Embed Size (px)
Citation preview

Espace L – JC Cassel
Conditionnement chez l’Homme et l’animal : principes et
domaines d’étude
Espace L – JC Cassel
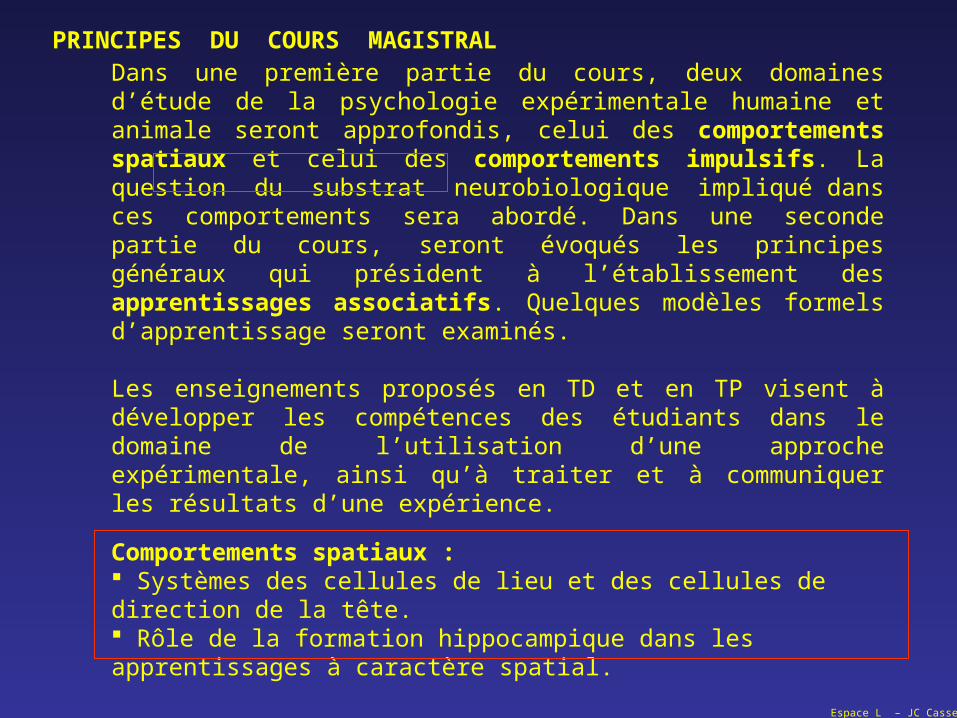
Dans une première partie du cours, deux domaines d’étude de la psychologie expérimentale humaine et animale seront approfondis, celui des comportements spatiaux et celui des comportements impulsifs. La question du substrat neurobiologique impliqué dans ces comportements sera abordé. Dans une seconde partie du cours, seront évoqués les principes généraux qui président à l’établissement des apprentissages associatifs. Quelques modèles formels d’apprentissage seront examinés. Les enseignements proposés en TD et en TP visent à développer les compétences des étudiants dans le domaine de l’utilisation d’une approche expérimentale, ainsi qu’à traiter et à communiquer les résultats d’une expérience.
PRINCIPES DU COURS MAGISTRAL
Comportements spatiaux : Systèmes des cellules de lieu et des cellules de direction de la tête. Rôle de la formation hippocampique dans les apprentissages à caractère spatial.
Espace L – JC Cassel
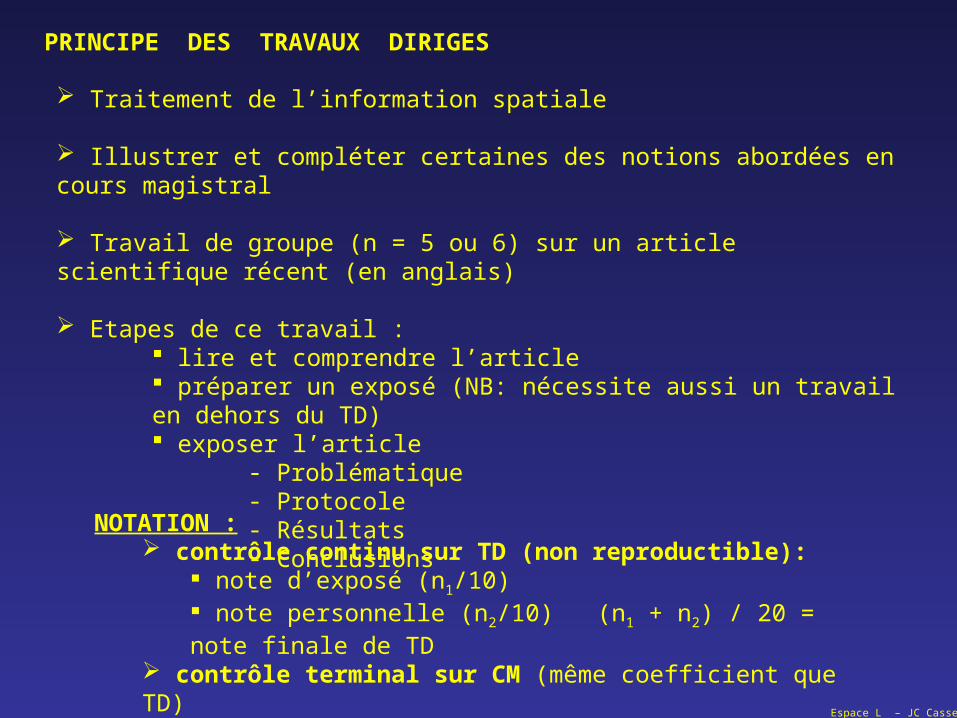
PRINCIPE DES TRAVAUX DIRIGES
Traitement de l’information spatiale
Illustrer et compléter certaines des notions abordées en cours magistral
Travail de groupe (n = 5 ou 6) sur un article scientifique récent (en anglais)
Etapes de ce travail : lire et comprendre l’article préparer un exposé (NB: nécessite aussi un travail en dehors du TD) exposer l’article
- Problématique - Protocole- Résultats- Conclusions
NOTATION : contrôle continu sur TD (non reproductible):
note d’exposé (n1/10) note personnelle (n2/10) (n1 + n2) / 20 = note finale de TD
contrôle terminal sur CM (même coefficient que TD) Note finale = TD + CM / 2 (coef. 1 pour chaque partie)

Espace L – JC Cassel
ORGANISATION DES TRAVAUX DIRIGES
10 octobre: Constitution des groupes. Qu’est-ce qu’un article scientifique ? Comment est-il construit ? Qu’est-ce qu’un
journal à comité de lecture? Comment ça marche? Présentation des articles choisis. Distribution des articles. Lecture des articles.
17 octobre: Lecture et compréhension des articles.
24 octobre: Compréhension des articles et préparation des exposés.
31 octobre: Annulation du TD.
7 novembre: Finalisation des exposés
14 novembre: Exposés (10 min d’exposé + 5 min de questions; prévoir 12h30)
21 novembre: Fin des exposés
28 novembre: Epreuve écrite (fin des TD)
Espace L – JC Cassel
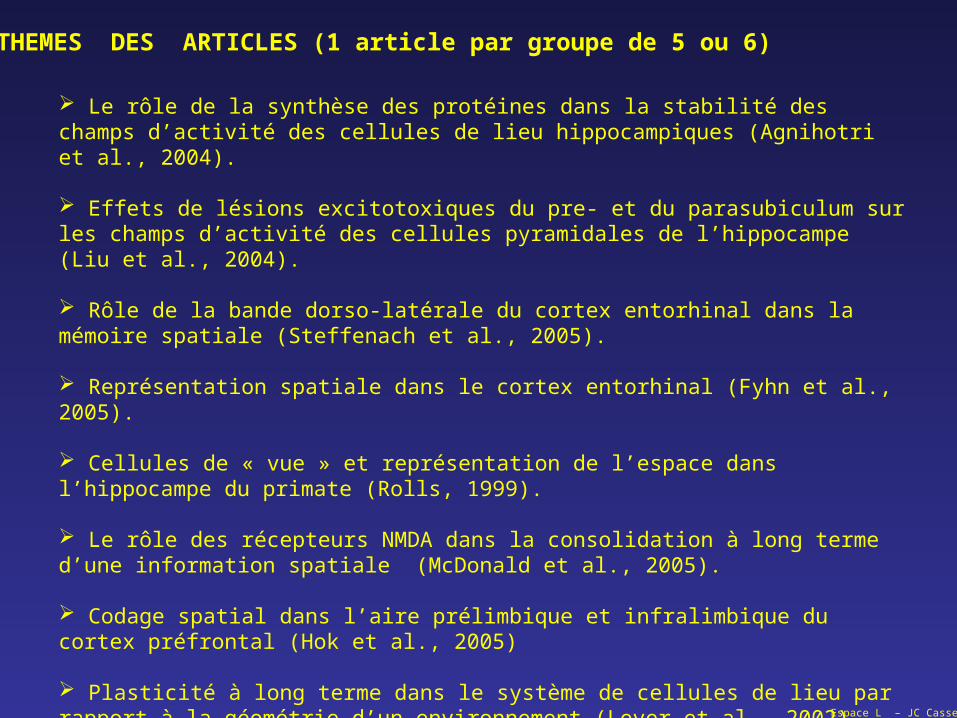
THEMES DES ARTICLES (1 article par groupe de 5 ou 6)
Le rôle de la synthèse des protéines dans la stabilité des champs d’activité des cellules de lieu hippocampiques (Agnihotri et al., 2004).
Effets de lésions excitotoxiques du pre- et du parasubiculum sur les champs d’activité des cellules pyramidales de l’hippocampe (Liu et al., 2004).
Rôle de la bande dorso-latérale du cortex entorhinal dans la mémoire spatiale (Steffenach et al., 2005).
Représentation spatiale dans le cortex entorhinal (Fyhn et al., 2005).
Cellules de « vue » et représentation de l’espace dans l’hippocampe du primate (Rolls, 1999).
Le rôle des récepteurs NMDA dans la consolidation à long terme d’une information spatiale (McDonald et al., 2005).
Codage spatial dans l’aire prélimbique et infralimbique du cortex préfrontal (Hok et al., 2005)
Plasticité à long terme dans le système de cellules de lieu par rapport à la géométrie d’un environnement (Lever et al., 2002).

Espace L – JC Cassel
LA REPRESENTATION DE L’ESPACE
D’après Robert Morris, Labyrinthe, 1999

Espace L – JC Cassel
Remarques introductives

Espace L – JC Cassel
Tout comportement, quel qu’il soit, a lieu à un endroit et un moment particuliers, et s’inscrit donc dans une double dimension définie par l’ESPACE et le TEMPS. Au cours de l’histoire de l’humanité, ces dimensions ont fait l’objet de tentatives de mesure et les unités ont tendu vers une certaine universalité, c’est dire leur importance. On trouve des traces de la mesure du temps et de l’espace, des représentations de l’organisation de ce dernier, en remontant assez loin dans l’histoire de l’Homme.
XVI° avant JCcadran solaire
3000 avant JC
clepsydre
sablier
2400 av JCgnomon
INTRODUCTION (1)
VI° avant JChorloge à encens

Espace L – JC Cassel
INTRODUCTION (2)
On trouve déjà des croquis dans certaines grottes préhistoriques, qui pourraient correspondre à des représentations du territoire occupé par leur(s) dessinateur(s).
Mésopotamie, 2500 avant JC ; carte en terre cuite (vallée d’un fleuve)
Carte « T dans l’O » du XIII° siècle (enluminure);convention =
Asie
Eur Afr
Image satellite
GPS embarqué
Carte de Cassini, Louis XV
Espace L – JC Cassel
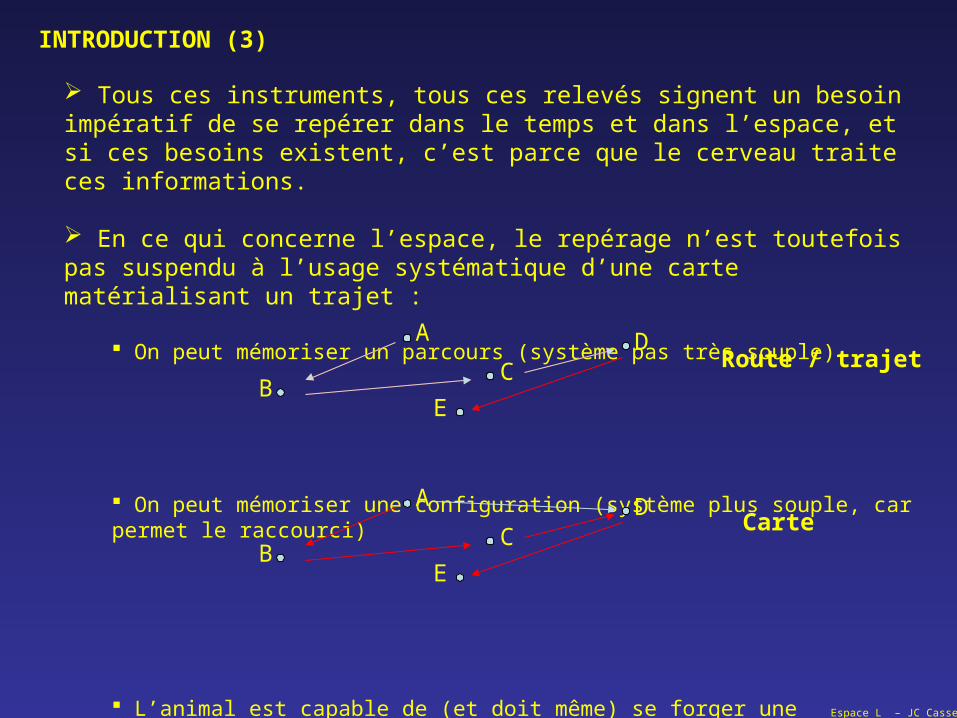
INTRODUCTION (3)
Tous ces instruments, tous ces relevés signent un besoin impératif de se repérer dans le temps et dans l’espace, et si ces besoins existent, c’est parce que le cerveau traite ces informations.
En ce qui concerne l’espace, le repérage n’est toutefois pas suspendu à l’usage systématique d’une carte matérialisant un trajet :
On peut mémoriser un parcours (système pas très souple)
On peut mémoriser une configuration (système plus souple, car permet le raccourci)
L’animal est capable de (et doit même) se forger une représentation de l’environnement dans lequel il vit (se nourrit, se reproduit, se repose) ; depuis Tolman, on parle de la « carte cognitive » (milieu XX°)
A
BC
D
E
A
BC
D
E
Route / trajet
Carte
Espace L – JC Cassel
INTRODUCTION (4)
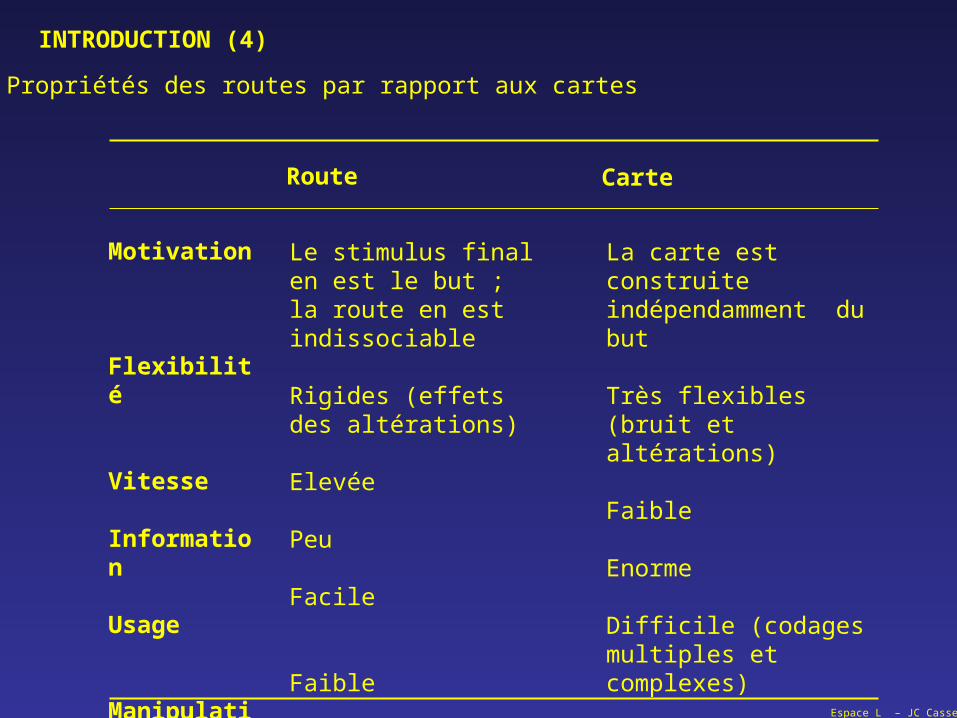
Propriétés des routes par rapport aux cartes
Route Carte
Le stimulus final en est le but ; la route en est indissociable
Rigides (effets des altérations)
Elevée
Peu
Facile
Faible
La carte est construite indépendamment du but
Très flexibles (bruit et altérations)
Faible
Enorme
Difficile (codages multiples et complexes)
On peut comparer des cartes, des lieus
Motivation
Flexibilité
Vitesse
Information
Usage
Manipulation
Espace L – JC Cassel
représentation mentale qu’un individu se fait de l’organisation de l’espace dans lequel il se trouve.
une représentation interne mentale qui implique que l’individu est capable d’inférer les relations, les distances ainsi que les directions qui lient différents points de l’espace sans avoir eu une expérience directe de chacun de ces points.
le concept de carte cognitive n’a pas 60 ans, et ne s’est imposé que vers la fin des années 70, en ce qui concerne l’animal (même si les recherches sur l’orientation dans l’espace sont plus anciennes / oiseaux migrateurs).
des arguments convergents ont fini par l’associer solidement (mais pas exclusivement) à une structure cérébrale particulière qu’on appelle l’hippocampe : l’hippocampe joue un rôle essentiel dans la représentation de l’espace, mais il n’est pas la seule structure impliquée.
cette association a été « un accouchement dans la douleur » puisqu’elle a dû s’imposer à contre-courant des conceptions behaviouristes.
INTRODUCTION (5) : LA NOTION DE « CARTE COGNITIVE »
Espace L – JC Cassel
ET POURTANT…
Watson et Lashley : recherches sur des oiseaux migrateurs (les sternes arctiques), 14 semaines en arctique, un peu plus en antarctique, le reste en voyage entre les deux (35000 km !): comment ces oiseaux s’orientent-ils pour leur migration? Comment font-ils, localement, pour retrouver leur nid?
En ce qui concerne le nid : arrivant de l’océan, ils suivent les indices les plus proéminents de l’île (les stimuli visuels) puis, à proximité du nid, ils rejoignent ce dernier sur la base d’habitudes indépendantes des stimuli visuels (notion de route); dans cette dernière partie de leur orientation, les oiseaux ne montraient rien qui indiquât la mise en œuvre de processus « idéatoires ».
En ce qui concerne la migration : les choses sont plus complexes (les hypothèses en vigueur : champs magnétiques, courants éoliens, position du soleil, perception visuelle du but, mémoire topographique héréditaire).Les chercheurs constatent que les oiseaux rejoignent leur objectif malgré une suture des paupières, puis abandonnent leurs recherches.
Hypothèse de la carte et du compas de Kramer (1953). En 1955, Griffin décrit trois processus: le pilotage (orientation sur la base d’indices familiers), le déplacement au compas (garder une direction constante) et la navigation (atteindre un but, quel que soit le point de départ et la direction).

Espace L – JC Cassel
Un peu d’histoire
L’hippocampe s’en va, la mémoire aussi, et les neurosciences commencent une longue croisière !
Espace L – JC Cassel
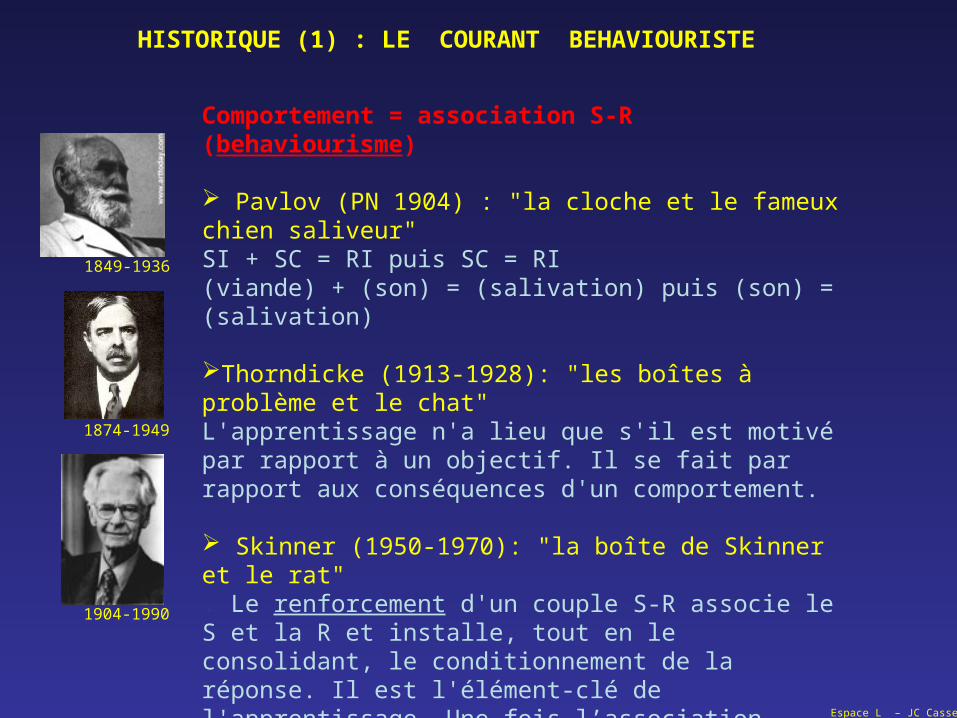
Comportement = association S-R (behaviourisme)
Pavlov (PN 1904) : "la cloche et le fameux chien saliveur"SI + SC = RI puis SC = RI(viande) + (son) = (salivation) puis (son) = (salivation)
Thorndicke (1913-1928): "les boîtes à problème et le chat"L'apprentissage n'a lieu que s'il est motivé par rapport à un objectif. Il se fait par rapport aux conséquences d'un comportement.
Skinner (1950-1970): "la boîte de Skinner et le rat". Le renforcement d'un couple S-R associe le S et la R et installe, tout en le consolidant, le conditionnement de la réponse. Il est l'élément-clé de l'apprentissage. Une fois l’association établie, une action appelle une conséquence quasi invariable qui est anticipée et attendue.Selon Skinner : Le fait de se croire libre d’agir n’est qu’une illusion, car le comportement est placé sous le contrôle de notre environnement et façonné au gré de nos expériences (sous-entendu: que nous ne contrôlons pas).
HISTORIQUE (1) : LE COURANT BEHAVIOURISTE
1849-1936
1874-1949
1904-1990
Espace L – JC Cassel
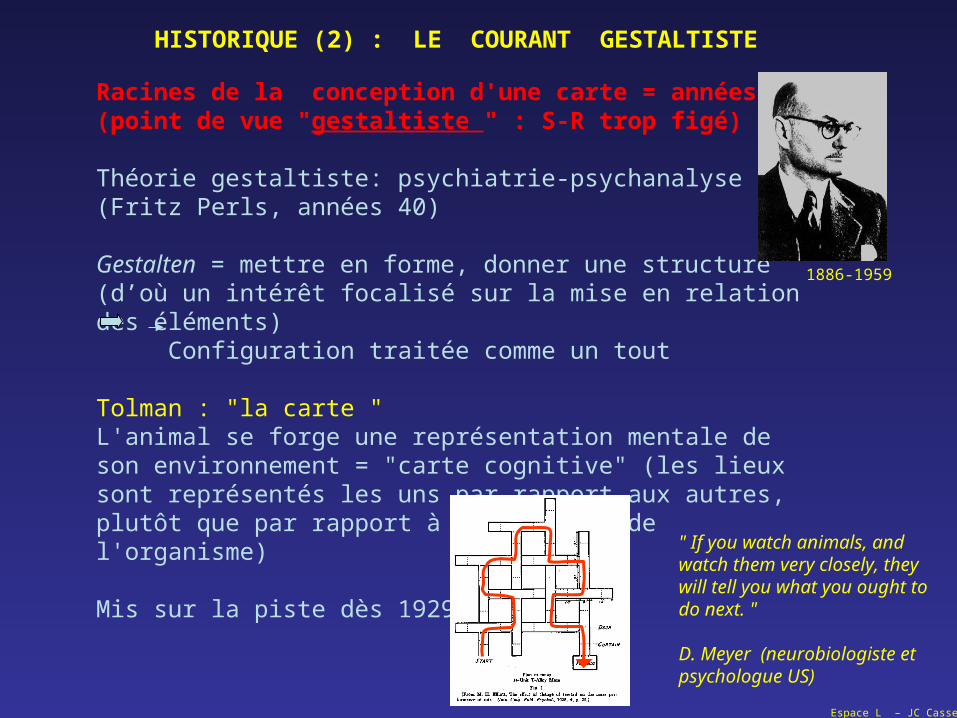
Racines de la conception d'une carte = années 30(point de vue "gestaltiste " : S-R trop figé)
Théorie gestaltiste: psychiatrie-psychanalyse(Fritz Perls, années 40)
Gestalten = mettre en forme, donner une structure(d’où un intérêt focalisé sur la mise en relation des éléments) Configuration traitée comme un tout
Tolman : "la carte " L'animal se forge une représentation mentale de son environnement = "carte cognitive" (les lieux sont représentés les uns par rapport aux autres, plutôt que par rapport à la position de l'organisme)
Mis sur la piste dès 1929…
HISTORIQUE (2) : LE COURANT GESTALTISTE
" If you watch animals, and watch them very closely, they will tell you what you ought to do next. "
D. Meyer (neurobiologiste et psychologue US)
1886-1959
Espace L – JC Cassel
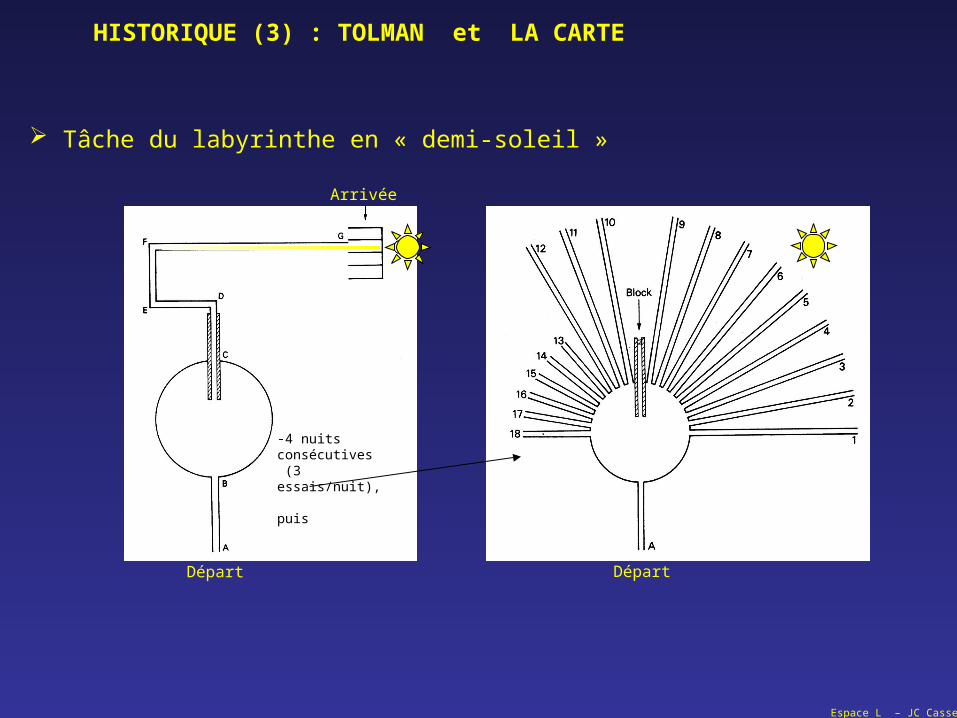
HISTORIQUE (3) : TOLMAN et LA CARTE
Tâche du labyrinthe en « demi-soleil »
Départ
Arrivée
-4 nuits consécutives (3 essais/nuit),
puis
Départ
Espace L – JC Cassel
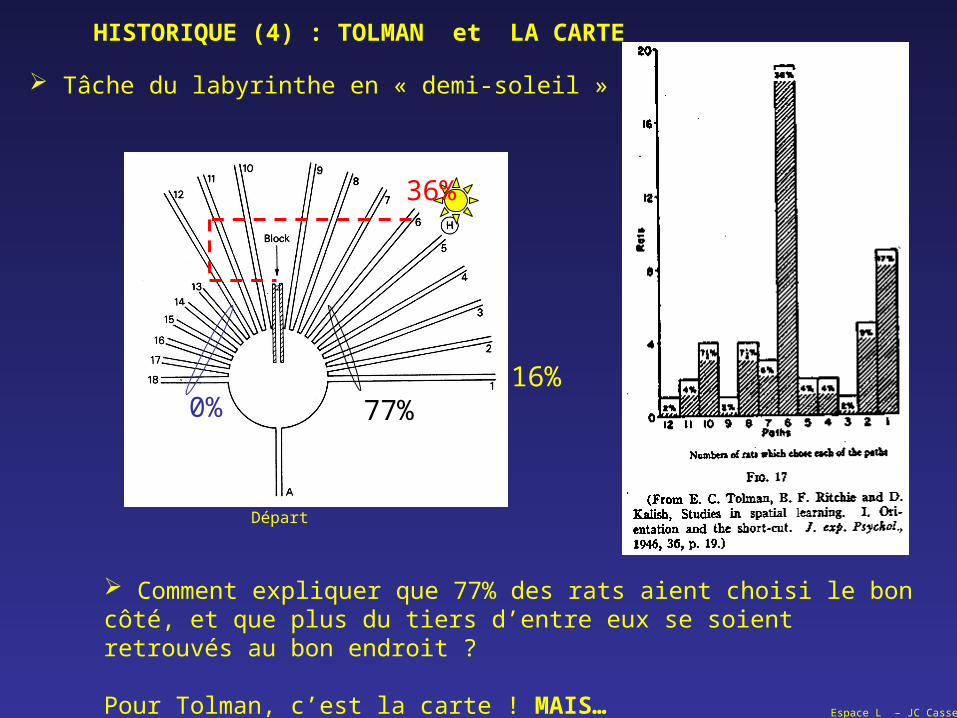
16%
Départ
Comment expliquer que 77% des rats aient choisi le bon côté, et que plus du tiers d’entre eux se soient retrouvés au bon endroit ?
Pour Tolman, c’est la carte ! MAIS…
HISTORIQUE (4) : TOLMAN et LA CARTE
Tâche du labyrinthe en « demi-soleil »
36%
0% 77%
Espace L – JC Cassel
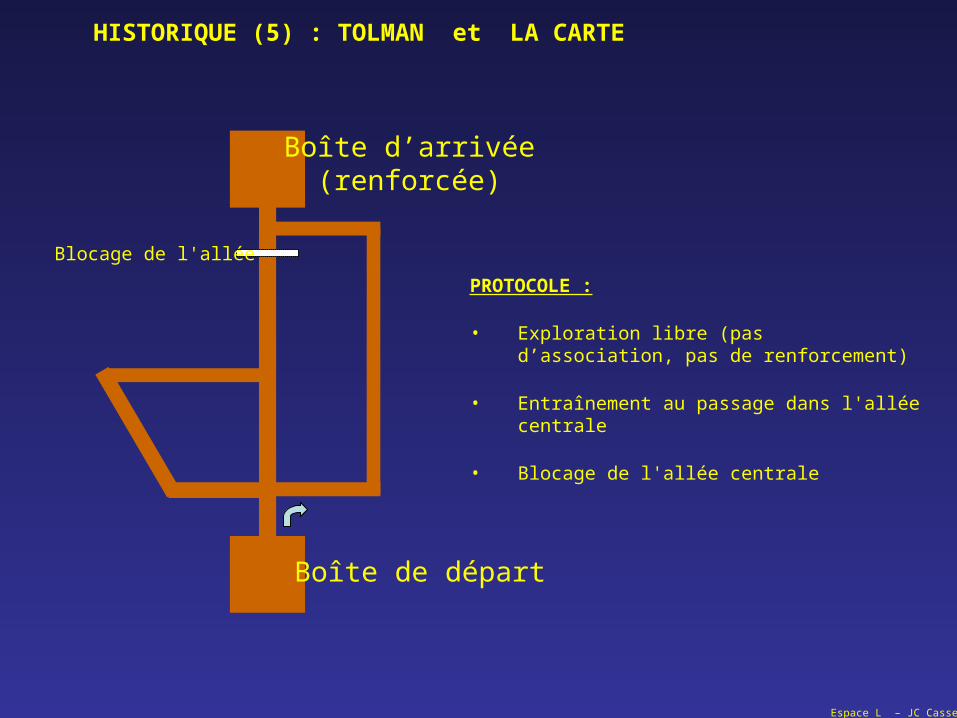
Boîte de départ
Boîte d’arrivée(renforcée)
PROTOCOLE :
• Exploration libre (pas d’association, pas de renforcement)
• Entraînement au passage dans l'allée centrale
• Blocage de l'allée centrale
Blocage de l'allée
HISTORIQUE (5) : TOLMAN et LA CARTE

Espace L – JC Cassel
HISTORIQUE (6) : TOLMAN et LA CARTE
La carte cognitive:
est le résultat d’un traitement central des informations spatiales et ce traitement est indépendant d’un renforcement (il s’agit d’un codage des aspects spatiaux d’une configuration 3D),
requiert une sélection et une organisation de l’information spatiale selon les relations spatiales qui relient les différents éléments constitutifs d’un environnement,
la représentation est celle d’un environnement vaste,
Ces points ont amené Gallistel (1990) à poser le principe d’un isomorphisme fonctionnel : correspondance formelle entre processus mentaux (ou neuronaux) et événements du monde extérieur (configuration = relations et opérations combinatoires sur le plan spatial)
Espace L – JC Cassel
Rep
aire
X
RAT Marcheur
Rep
aire
X
RAT
2 mètre
Mar
cheu
r
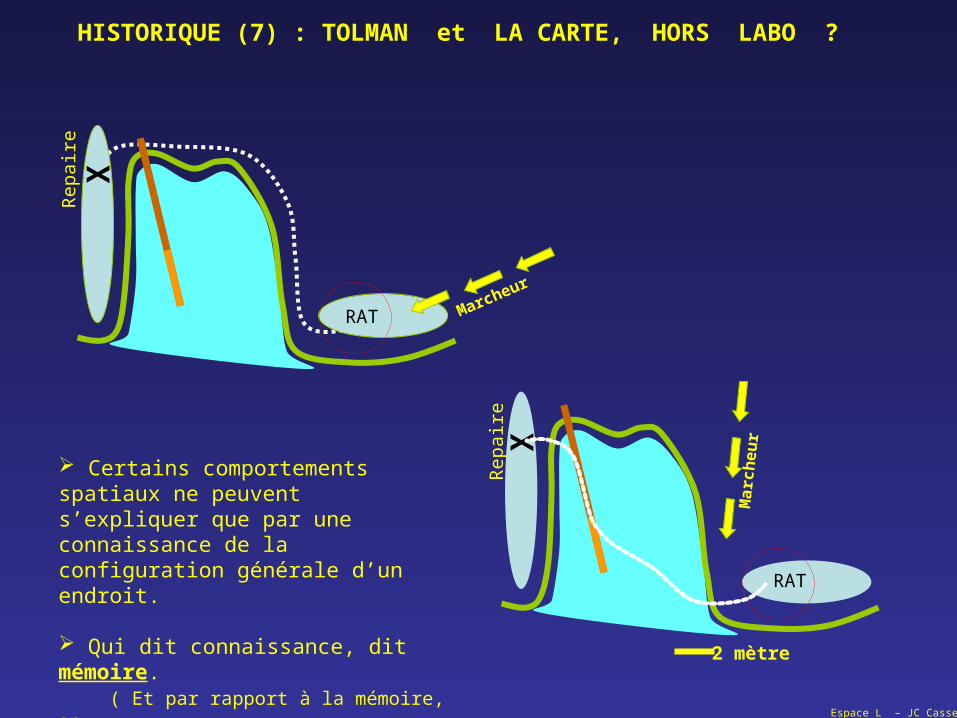
HISTORIQUE (7) : TOLMAN et LA CARTE, HORS LABO ?
Certains comportements spatiaux ne peuvent s’expliquer que par une connaissance de la configuration générale d’un endroit.
Qui dit connaissance, dit mémoire. ( Et par rapport à la mémoire, on remonte aussi au début du XX° )

Espace L – JC Cassel
HISTORIQUE (8) : LASHLEY ET LES CIRCUITS DE LA MEMOIRE
Dans les années 1920, Karl Lashley s’est intéressé aux circuits neuronaux impliqués dans la mémoire : quels sont ceux dont un rat ou un singe ne peut se passer pour procéder à un apprentissage ?
Il a retiré des fragments corticaux en divers endroits et à procédé à de multiples incisions sans jamais réussir à perturber significativement des rats ou des singes préalablement entraînés à une tâche impliquant des fonctions de perception et des fonctions motrices.
La conclusion au bout de 30 ans (donc dans les années 50): principes de l’action de masse et de l’équipotentialité (pas de localisation particulière)
[Erreur: mémoire implicite, focalisation sur le cortex]
Ironie du sort : 1953, Scoville opère H.M. alors âgé de 16 ans
1890-1958
Espace L – JC Cassel
HISTORIQUE (9) : H.M. ET SA MEMOIRE
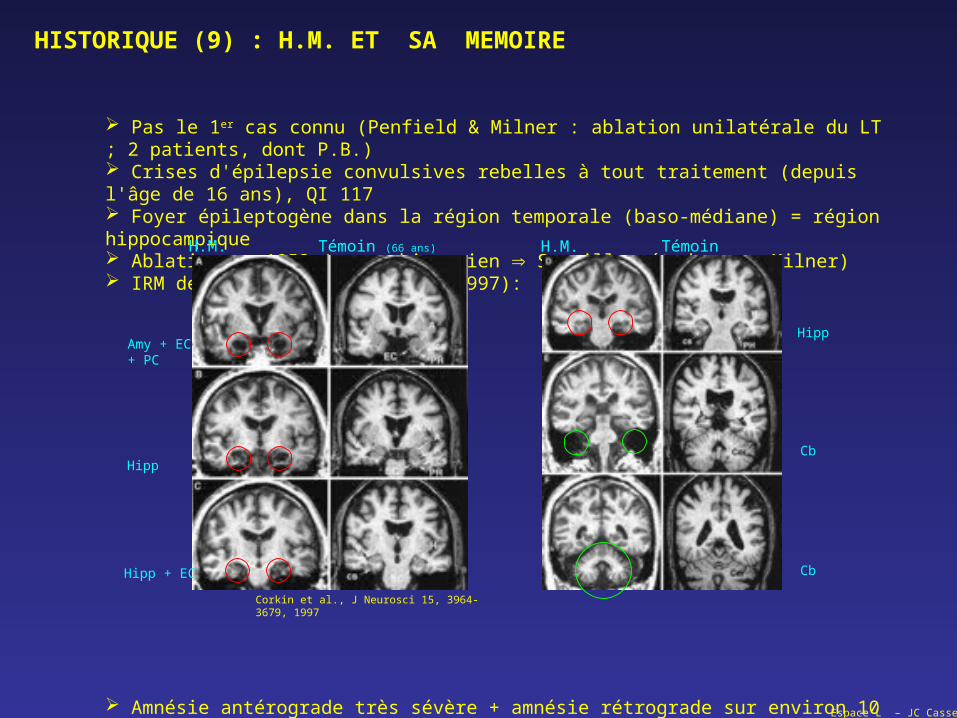
Pas le 1er cas connu (Penfield & Milner : ablation unilatérale du LT ; 2 patients, dont P.B.) Crises d'épilepsie convulsives rebelles à tout traitement (depuis l'âge de 16 ans), QI 117 Foyer épileptogène dans la région temporale (baso-médiane) = région hippocampique Ablation en 1953 (neurochirurgien Scoville, études par Milner) IRM de 1997 (Corkin et al., 1997):
Amnésie antérograde très sévère + amnésie rétrograde sur environ 10 ans, mais souvenirs les plus anciens relativement intacts
H.M. H.M.Témoin (66 ans) Témoin
Amy + EC+ PC
Hipp
Hipp + EC
Hipp
Cb
Cb
Corkin et al., J Neurosci 15, 3964-3679, 1997

Espace L – JC Cassel
H.M. ne connaît pas son âge, la date, le fait que ses parents soient décédés,… il est toutefois capable de réciter une liste de 7 items pendant quelques secondes à condition qu'il n'y ait aucune interférence, il ne fixe rien au delà de quelques secondes :
"Chaque jour est un jour singulier en soi, quelles que soient les joies ou les tristesses éprouvées… En ce moment, je me demande si j'ai pu dire quelque chose de travers. Voyez-vous, en ce moment tout me semble clair, mais que s'est-il passé il y a quelques instants? C'est ce qui me préoccupe. C'est comme si je me réveillais d'un rêve. Je suis incapable de me rappeler." (propos recueillis et rapportés par Brenda Milner, 1970 )
• Sa mémoire à long terme est atteinte, tout comme les processus de consolidation. Sa mémoire à court terme est aussi atteinte. • Rôle de la formation hippocampique dans les deux types de mémoire?
MAIS : Toutes les mémoires ne sont pas atteintes. Par ex., Sa mémoire à très court terme est quasi normale, de même que la mémoire procédurale. Autres structures impliquées?
DAYS
1 2 3 4 5 6 7 8 9 10
ER
RO
RS
5
10
15
20
25
30
35
DAYS
1 2 3 4 5 6 7 8 9 10
ER
RO
RS
0
5
10
15
20
25
30
DAYS
1 2 3 4 5 6 7 8 9 10
ER
RO
RS
0
5
10
15
20
25
30DAY 1 DAY 2 DAY 3
Dessin au miroir
Trials Trials Trials
a
a
HISTORIQUE (10) : H.M. ET SA MEMOIRE

Espace L – JC Cassel
H.M. : hippocampe, cortex entorhinal, cortex périrhinal, amygdale, cervelet
Question
Hippocampe / autres structures
Mémoire àcourt terme
Mémoire àlong terme? ?Acquisition, consolidation,rappel
HISTORIQUE (11) : H.M. OUVRE UNE NOUVELLE ERE !
Espace L – JC Cassel
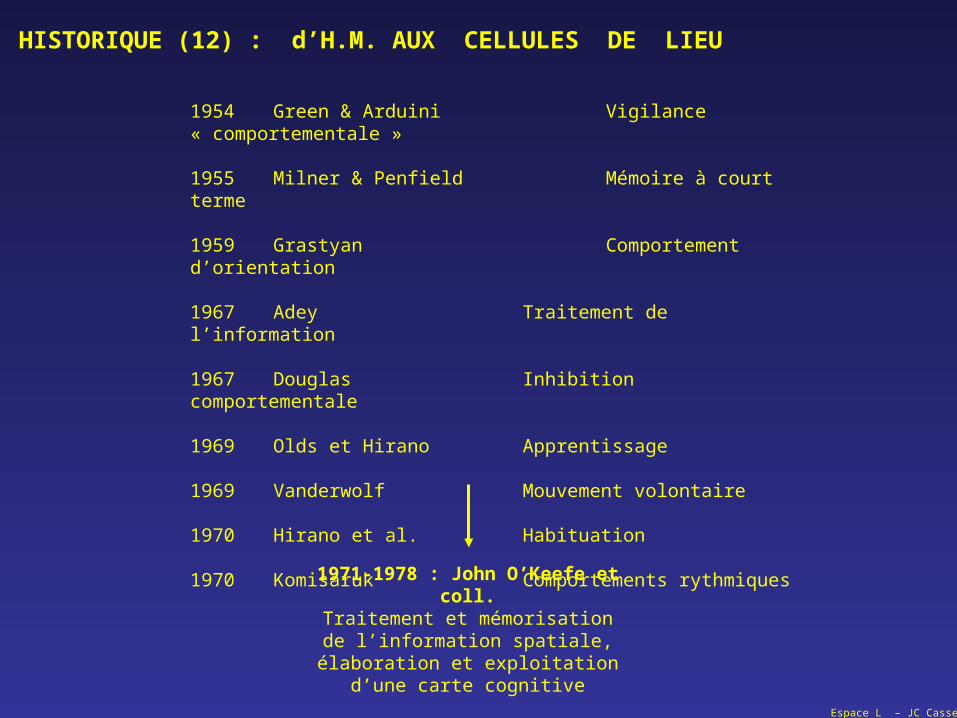
1954 Green & Arduini Vigilance « comportementale »
1955 Milner & Penfield Mémoire à court terme
1959 Grastyan Comportement d’orientation
1967 Adey Traitement de l’information
1967 Douglas Inhibition comportementale
1969 Olds et Hirano Apprentissage
1969 Vanderwolf Mouvement volontaire
1970 Hirano et al. Habituation
1970 Komisaruk Comportements rythmiques
1971-1978 : John O’Keefe et coll.Traitement et mémorisation
de l’information spatiale,élaboration et exploitation
d’une carte cognitive
HISTORIQUE (12) : d’H.M. AUX CELLULES DE LIEU

Espace L – JC Cassel
Les cellules de lieu
Crépitements et balbutiements…
Espace L – JC Cassel
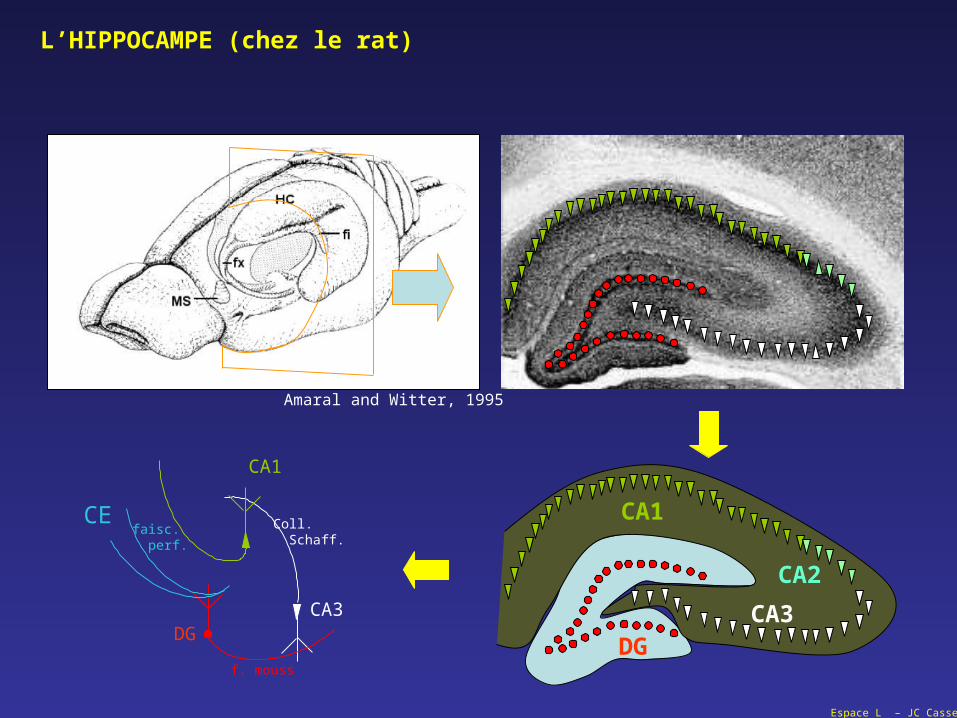
L’HIPPOCAMPE (chez le rat)
Amaral and Witter, 1995
f. mouss
Coll. Schaff.
CEfaisc. perf.
DGCA3
CA1
CA1
CA2
CA3DG
Espace L – JC Cassel
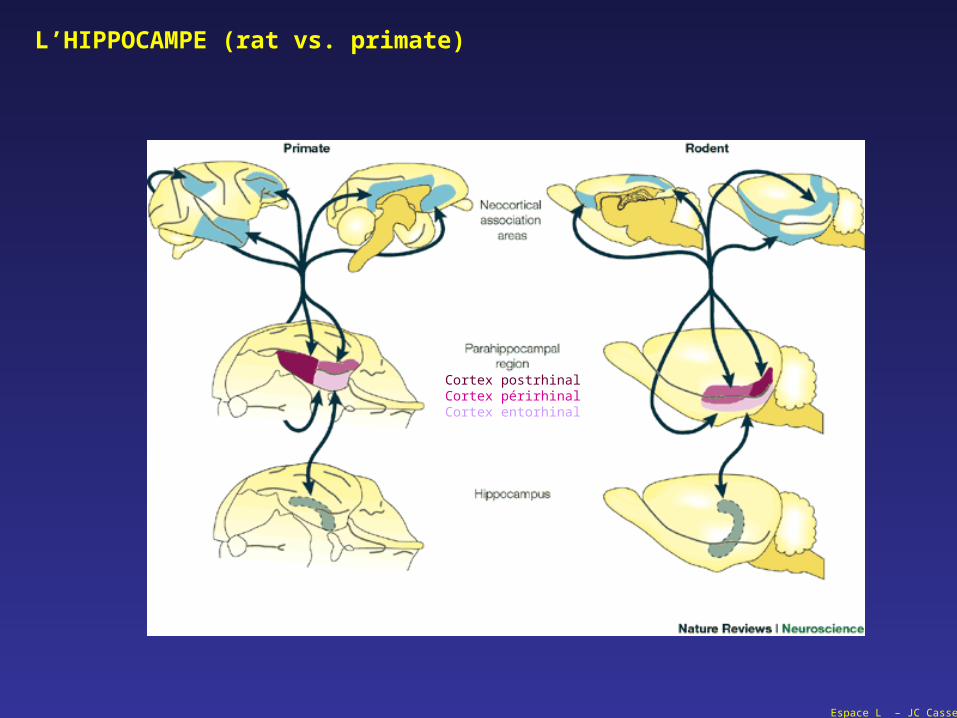
L’HIPPOCAMPE (rat vs. primate)
Cortex postrhinalCortex périrhinalCortex entorhinal
Espace L – JC Cassel
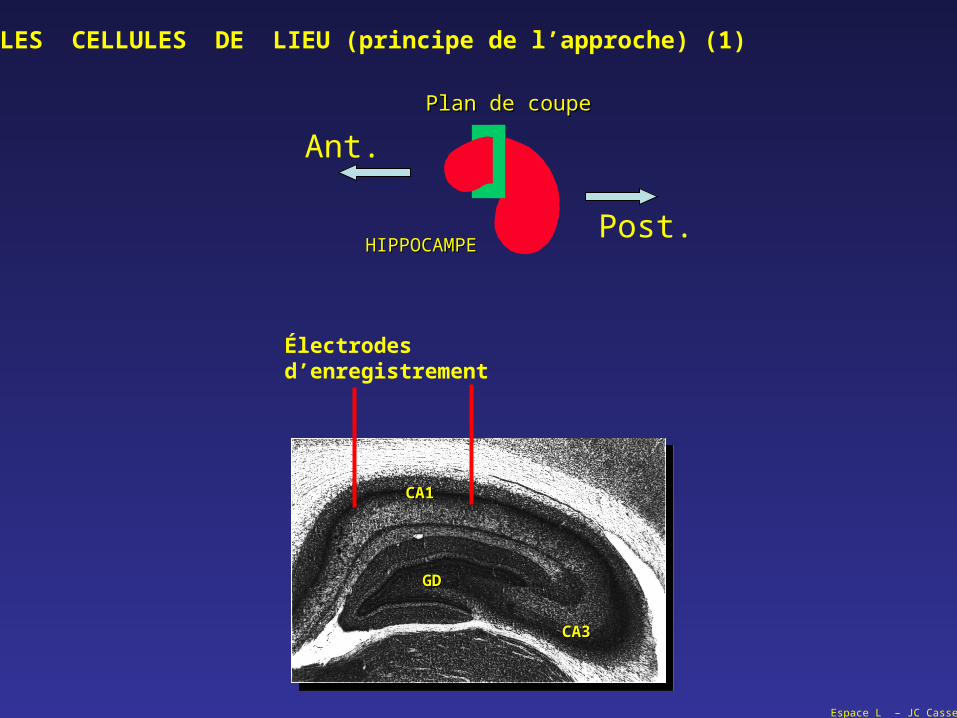
Plan de coupePlan de coupe
HIPPOCAMPEHIPPOCAMPE
Ant.
Post.
GDGD
CA1CA1
CA3CA3
Électrodesd’enregistrement
LES CELLULES DE LIEU (principe de l’approche) (1)
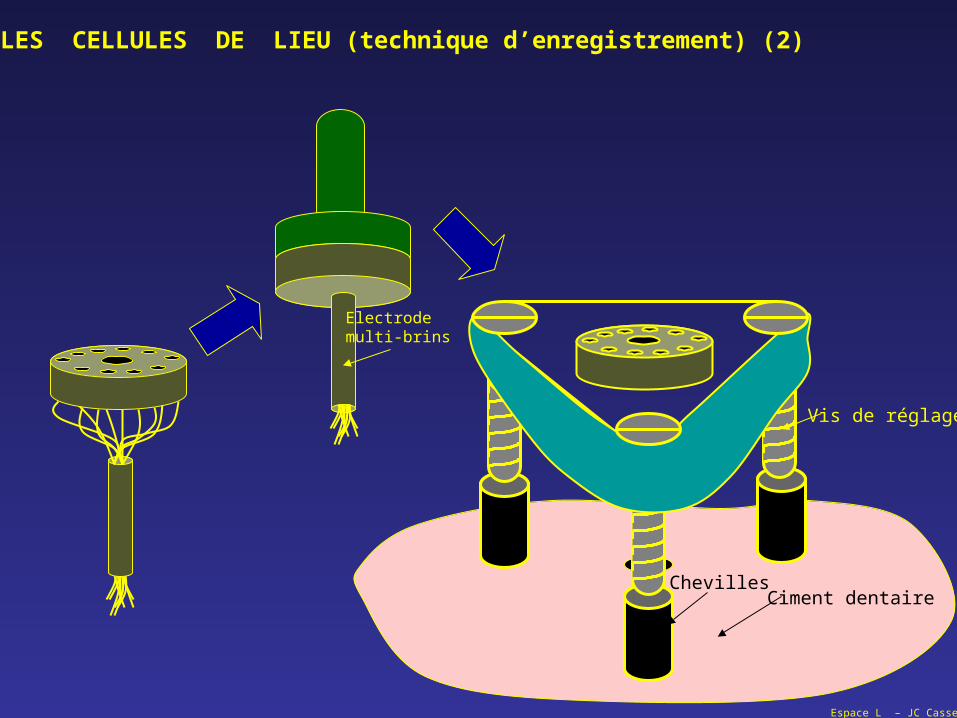
Espace L – JC Cassel
Vis de réglage
ChevillesCiment dentaire
Electrodemulti-brins
LES CELLULES DE LIEU (technique d’enregistrement) (2)
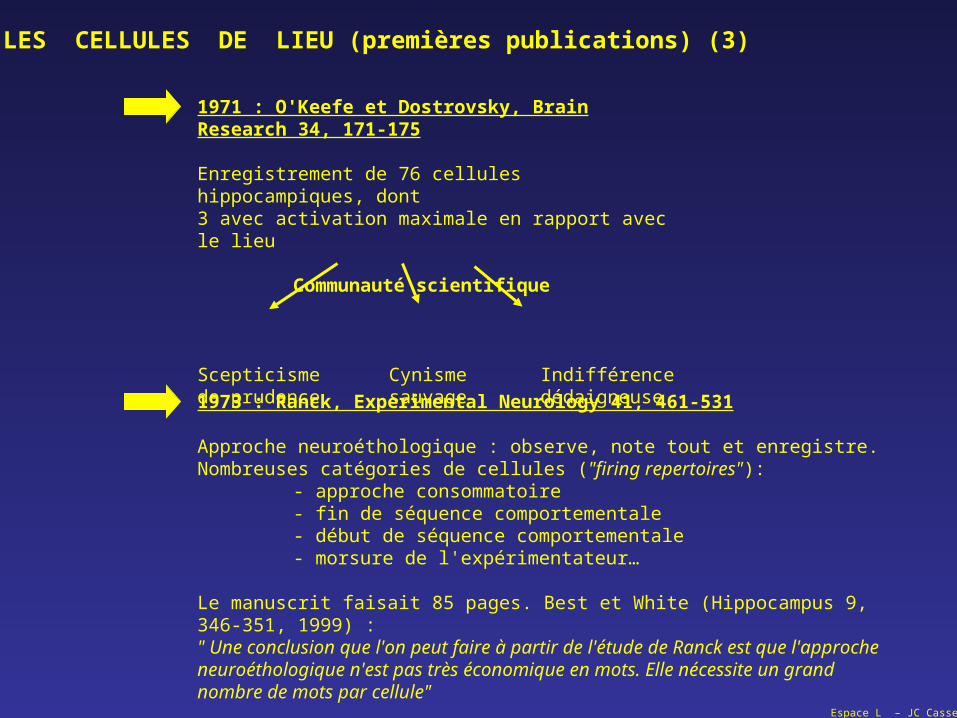
Espace L – JC Cassel
1971 : O'Keefe et Dostrovsky, Brain Research 34, 171-175
Enregistrement de 76 cellules hippocampiques, dont3 avec activation maximale en rapport avec le lieu
Communauté scientifique
Scepticisme Cynisme Indifférencede prudence sauvage dédaigneuse
LES CELLULES DE LIEU (premières publications) (3)
1973 : Ranck, Experimental Neurology 41, 461-531
Approche neuroéthologique : observe, note tout et enregistre.Nombreuses catégories de cellules ("firing repertoires"):
- approche consommatoire- fin de séquence comportementale- début de séquence comportementale- morsure de l'expérimentateur…
Le manuscrit faisait 85 pages. Best et White (Hippocampus 9, 346-351, 1999) :" Une conclusion que l'on peut faire à partir de l'étude de Ranck est que l'approche neuroéthologique n'est pas très économique en mots. Elle nécessite un grand nombre de mots par cellule"
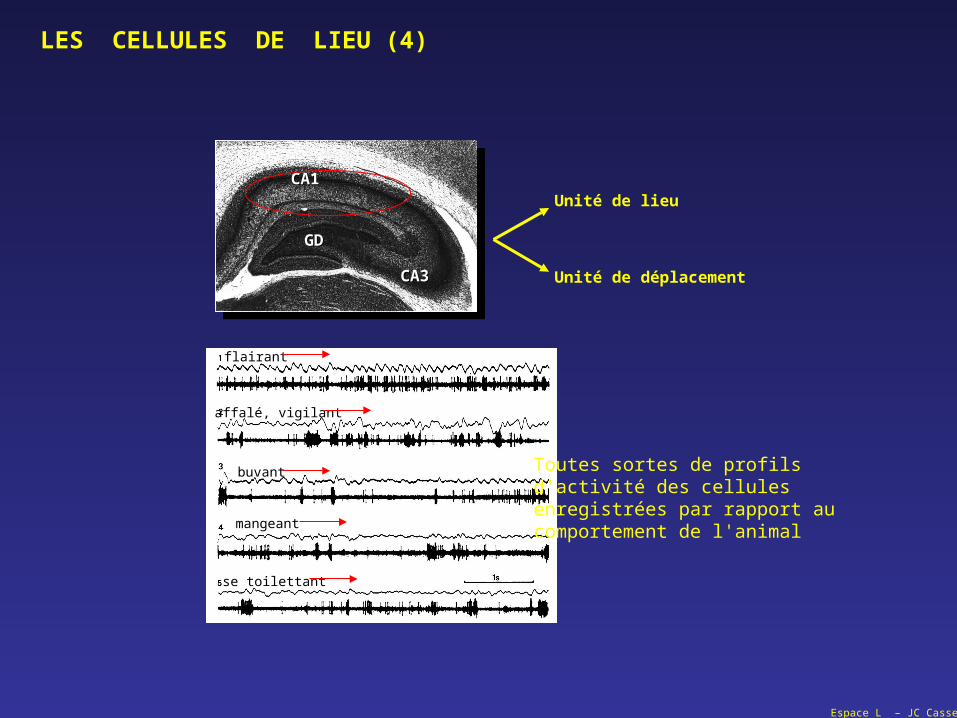
Espace L – JC Cassel
flairant
se toilettant
affalé, vigilant
buvant
mangeant
GDGD
CA1CA1
CA3CA3
Unité de lieu
Unité de déplacement
Toutes sortes de profilsd'activité des cellulesenregistrées par rapport aucomportement de l'animal
LES CELLULES DE LIEU (4)
Espace L – JC Cassel
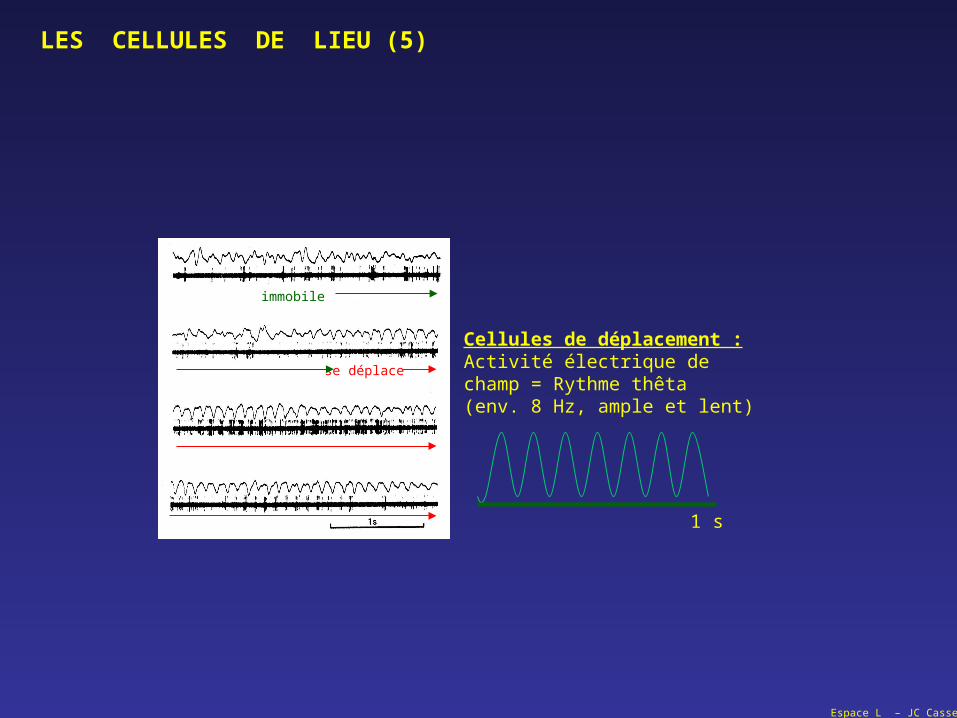
LES CELLULES DE LIEU (5)
immobile
se déplace
Cellules de déplacement :Activité électrique dechamp = Rythme thêta(env. 8 Hz, ample et lent)
1 s
Espace L – JC Cassel
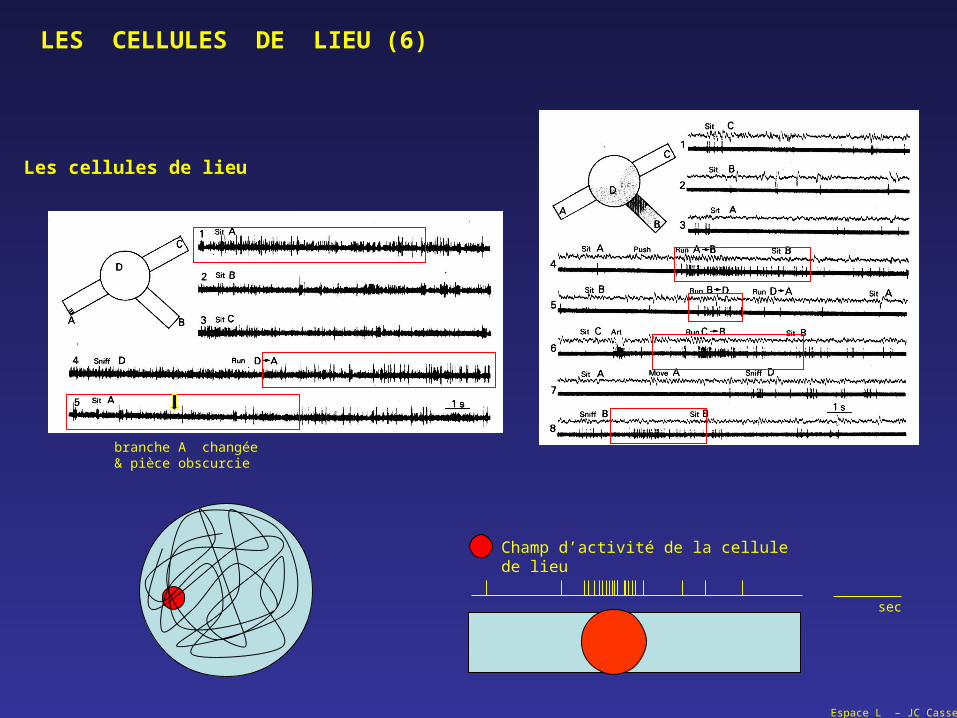
LES CELLULES DE LIEU (6)
branche A changée& pièce obscurcie
Les cellules de lieu
Champ d’activité de la cellule de lieu
sec

Espace L – JC Cassel
Les cellules de lieu.

Espace L – JC Cassel
ORGANISATION DES TRAVAUX DIRIGES
10 octobre: Constitution des groupes. Qu’est-ce qu’un article scientifique ? Comment est-il construit ? Qu’est-ce qu’un
journal à comité de lecture? Comment ça marche? Présentation des articles choisis. Distribution des articles. Lecture des articles.
17 octobre: Lecture et compréhension des articles.
24 octobre: Compréhension des articles et préparation des exposés.
31 octobre: Préparation et finalisation des exposés.
7 novembre: Exposés (10 min d’exposé + 5 min de questions; prévoir 12h30)
14 novembre: Fin des exposés
21 novembre: Epreuve écrite (fin des TD)
Espace L – JC Cassel
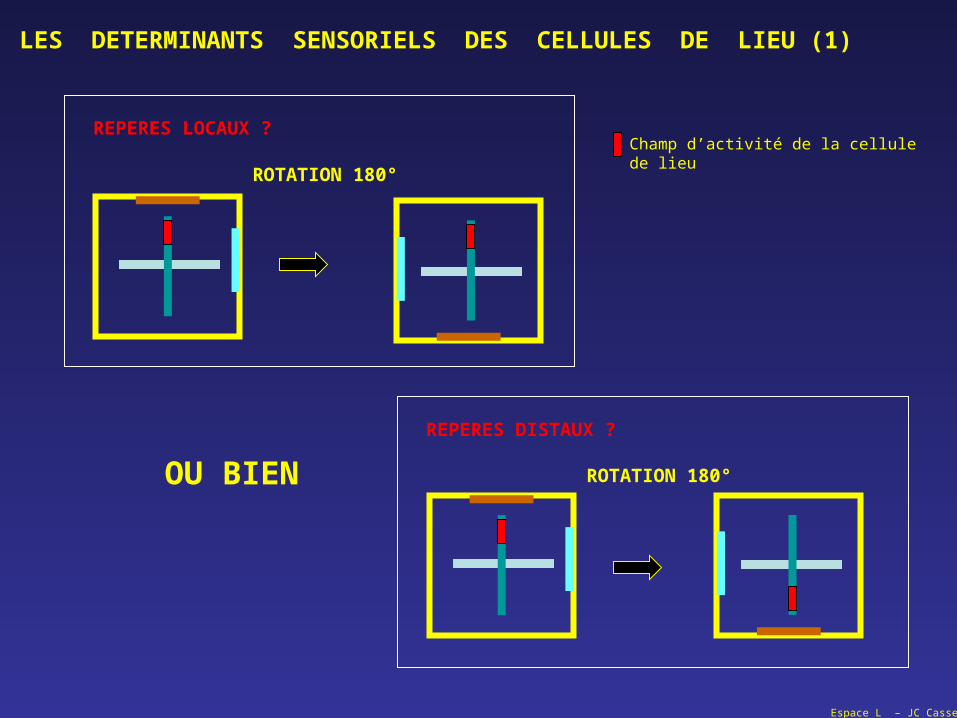
LES DETERMINANTS SENSORIELS DES CELLULES DE LIEU (1)
OU BIEN ROTATION 180°
REPERES DISTAUX ?
ROTATION 180°
REPERES LOCAUX ?Champ d’activité de la cellule de lieu
Espace L – JC Cassel
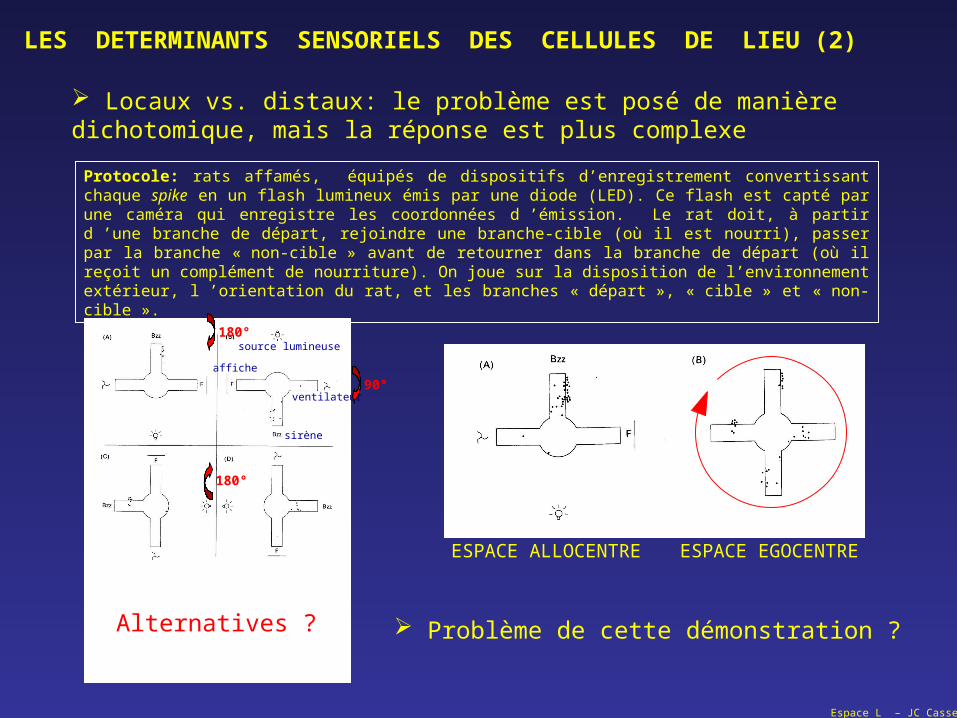
LES DETERMINANTS SENSORIELS DES CELLULES DE LIEU (2)
Locaux vs. distaux: le problème est posé de manière dichotomique, mais la réponse est plus complexe
Protocole: rats affamés, équipés de dispositifs d’enregistrement convertissant chaque spike en un flash lumineux émis par une diode (LED). Ce flash est capté par une caméra qui enregistre les coordonnées d ’émission. Le rat doit, à partir d ’une branche de départ, rejoindre une branche-cible (où il est nourri), passer par la branche « non-cible » avant de retourner dans la branche de départ (où il reçoit un complément de nourriture). On joue sur la disposition de l’environnement extérieur, l ’orientation du rat, et les branches « départ », « cible » et « non-cible ».
source lumineuse
ventilateur
sirène
affiche
180°
Alternatives ?
ESPACE ALLOCENTRE ESPACE EGOCENTRE
Problème de cette démonstration ?
180°
90°
Espace L – JC Cassel
LES DETERMINANTS SENSORIELS DES CELLULES DE LIEU (3)
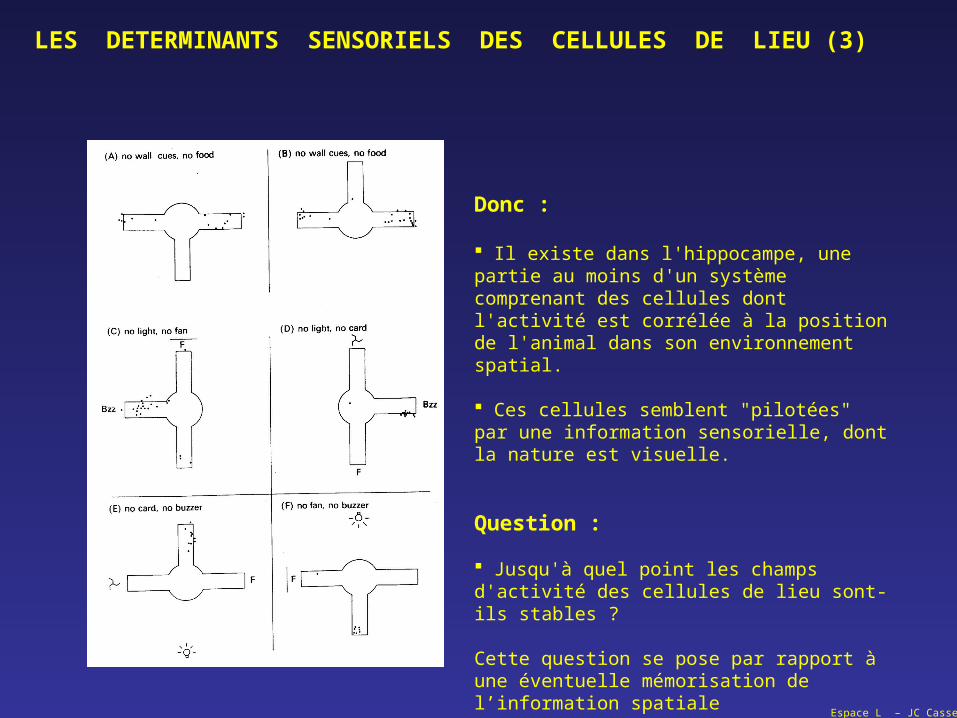
Donc :
Il existe dans l'hippocampe, une partie au moins d'un système comprenant des cellules dont l'activité est corrélée à la position de l'animal dans son environnement spatial.
Ces cellules semblent "pilotées" par une information sensorielle, dont la nature est visuelle.
Question :
Jusqu'à quel point les champs d'activité des cellules de lieu sont-ils stables ?
Cette question se pose par rapport à une éventuelle mémorisation de l’information spatiale
Espace L – JC Cassel
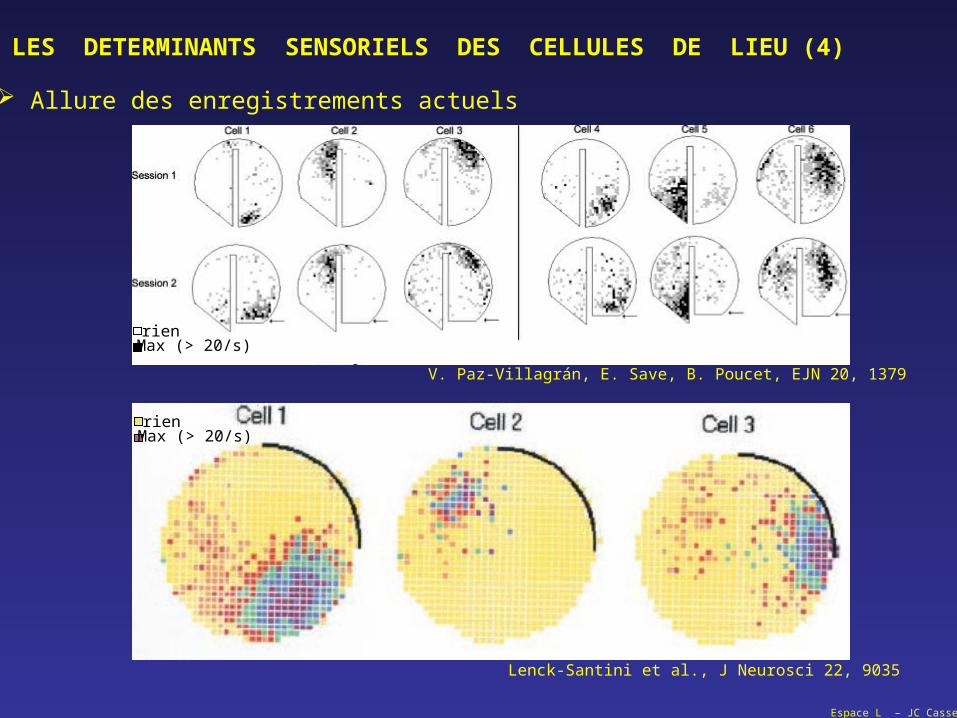
LES DETERMINANTS SENSORIELS DES CELLULES DE LIEU (4)
Allure des enregistrements actuels
Lenck-Santini et al., J Neurosci 22, 9035
rienMax (> 20/s)
V. Paz-Villagrán, E. Save, B. Poucet, EJN 20, 1379
rienMax (> 20/s)
Espace L – JC Cassel
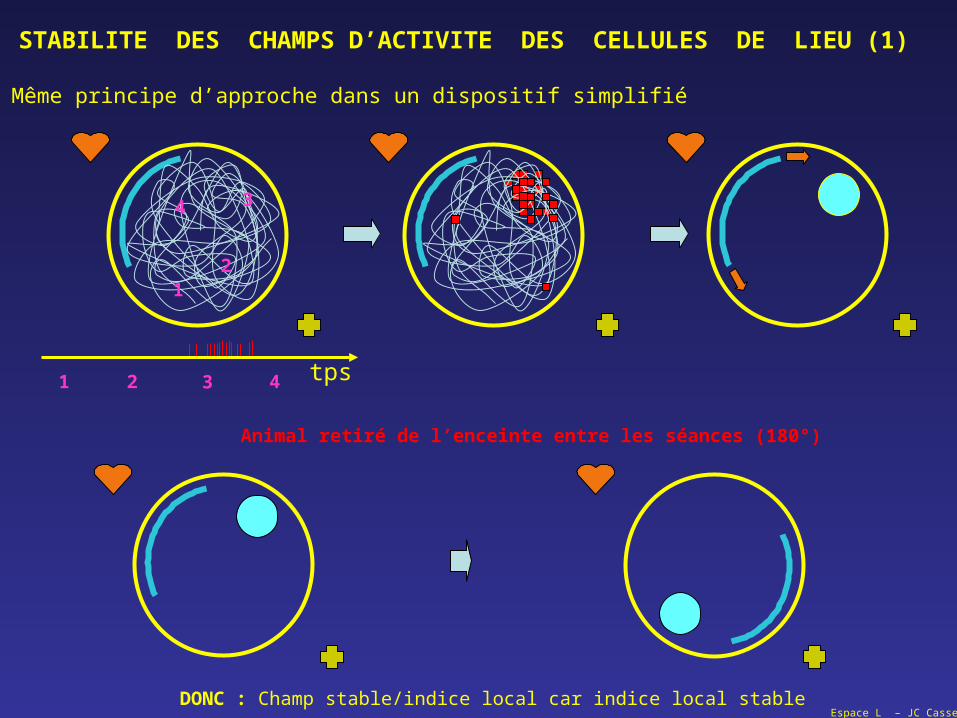
STABILITE DES CHAMPS D’ACTIVITE DES CELLULES DE LIEU (1)
Même principe d’approche dans un dispositif simplifié
tps
12
34
1 2 3 4
Animal retiré de l’enceinte entre les séances (180°)
DONC : Champ stable/indice local car indice local stable
Espace L – JC Cassel
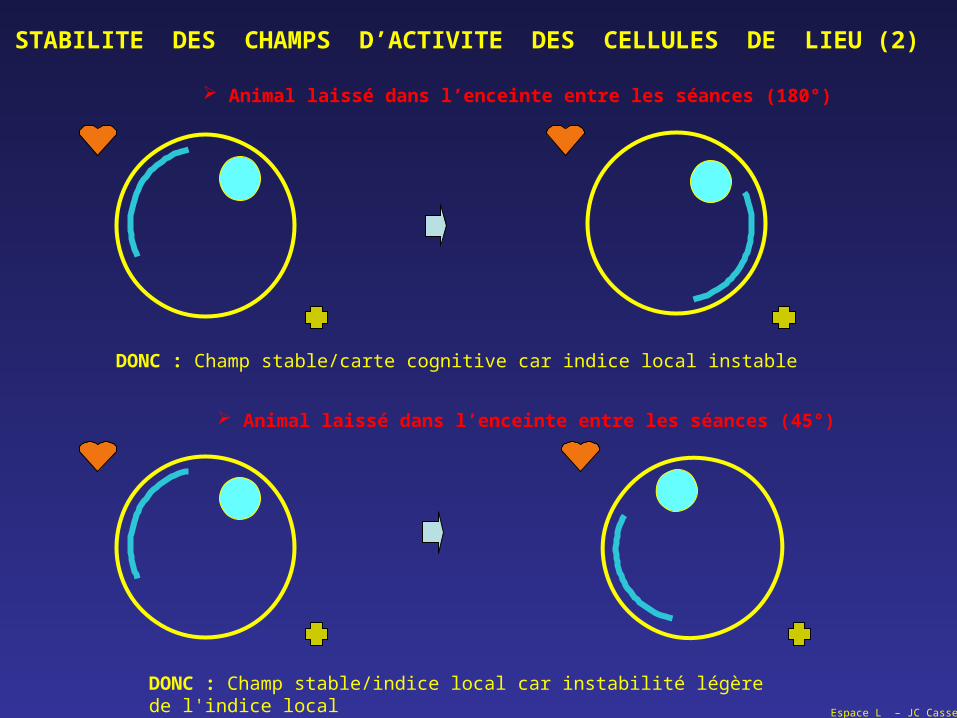
STABILITE DES CHAMPS D’ACTIVITE DES CELLULES DE LIEU (2)
Animal laissé dans l’enceinte entre les séances (180°)
DONC : Champ stable/carte cognitive car indice local instable
Animal laissé dans l’enceinte entre les séances (45°)
DONC : Champ stable/indice local car instabilité légère de l'indice local
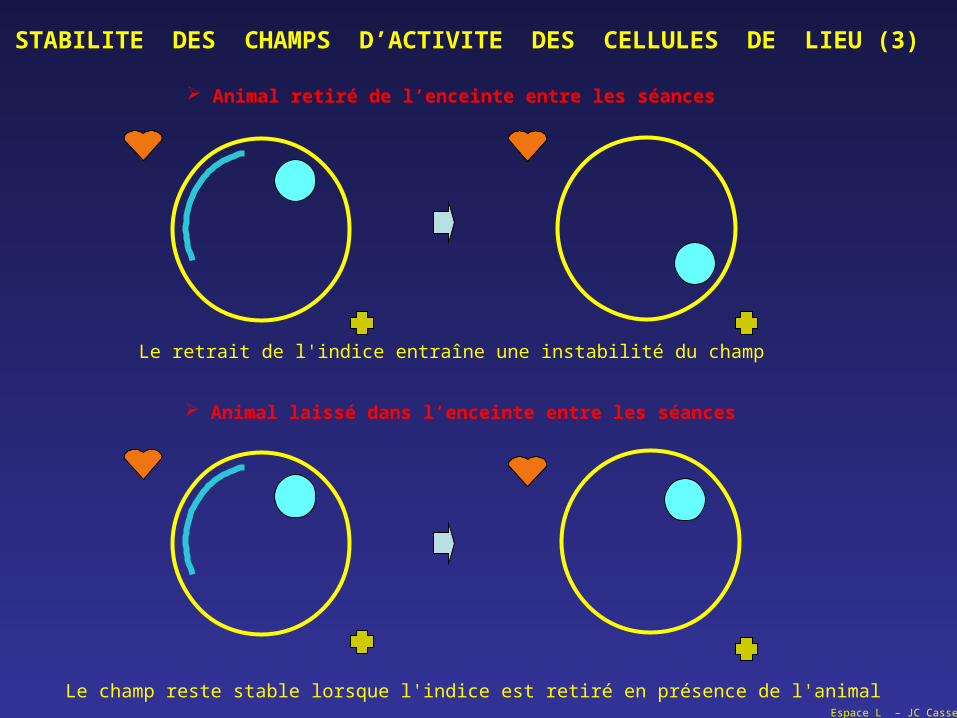
Espace L – JC Cassel
STABILITE DES CHAMPS D’ACTIVITE DES CELLULES DE LIEU (3)
Animal retiré de l’enceinte entre les séances
Animal laissé dans l’enceinte entre les séances
Le retrait de l'indice entraîne une instabilité du champ
Le champ reste stable lorsque l'indice est retiré en présence de l'animal

Espace L – JC Cassel
STABILITE DES CHAMPS D’ACTIVITE DES CELLULES DE LIEU (4)
Animal laissé dans l’enceinte entre les séances et les indices olfactifs du plancher sont brouillés.
Le retrait de l'indice conjugué au brouillage olfactif entraîne une instabilité du champ
Les cellules de lieu participent à un processus de traitement de l’information spatiale qui se base, dans la mesure du possible, sur les éléments qui, dans un environnement donné, sont susceptibles de permettre une orientation, et donc sur des éléments stables.
L’instabilité de ces éléments, lorsque flagrante, se traduit par une dérive/réorganisation des champs d’activité des cellules de lieu (déroute ou nouveau codage spatial?)

Espace L – JC Cassel
STABILITE DES CHAMPS D’ACTIVITE DES CELLULES DE LIEU (5)
Qu’en est-il de la stabilité dans le temps ? Epiphénomène transitoire ou reflet d’un véritable processus de traitement avec composante mnésique ?
2 jours
2 semaines
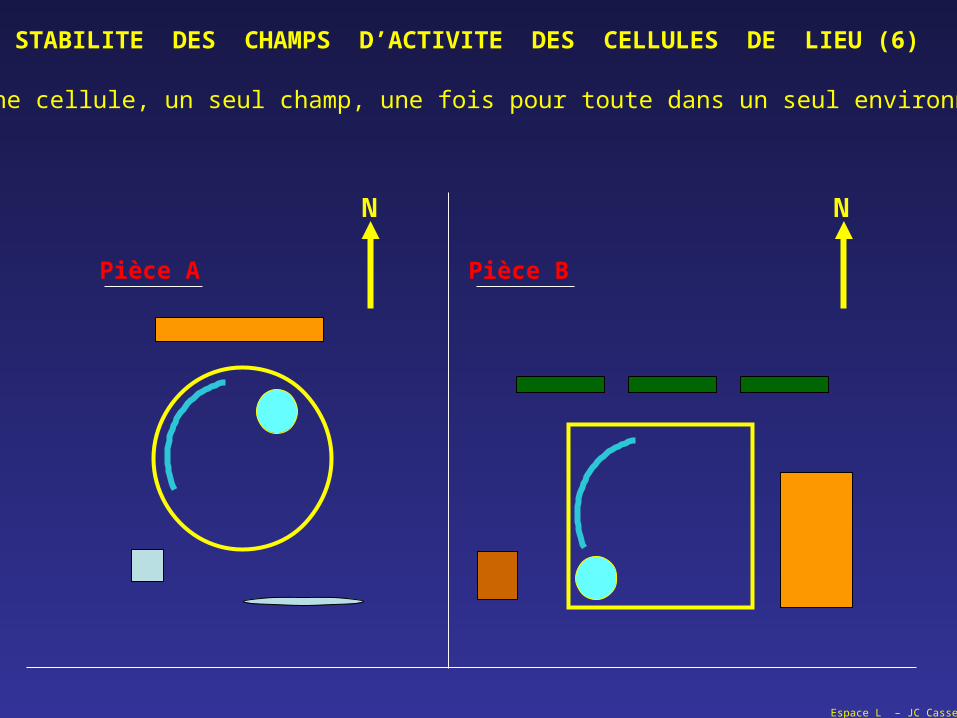
Espace L – JC Cassel
STABILITE DES CHAMPS D’ACTIVITE DES CELLULES DE LIEU (6)
N
Pièce A Pièce B
N
Une cellule, un seul champ, une fois pour toute dans un seul environnement ?

Espace L – JC Cassel
Expériences de manipulation de l'environnement sensoriel
pluralité de canaux sensoriels / champs
On retient
Prédominance visuelle Compensation olfactiveou autre possible
Champs d'activité d'une cellule donnée :
- différents dans plusieurs dispositifs- stables pendant une séance (plusieurs heures)- stables pendant des séances espacées par plusieurs semaines à condition que l'agencement de l'environnement reste stable- indépendants de la direction d'accès
EN RESUME
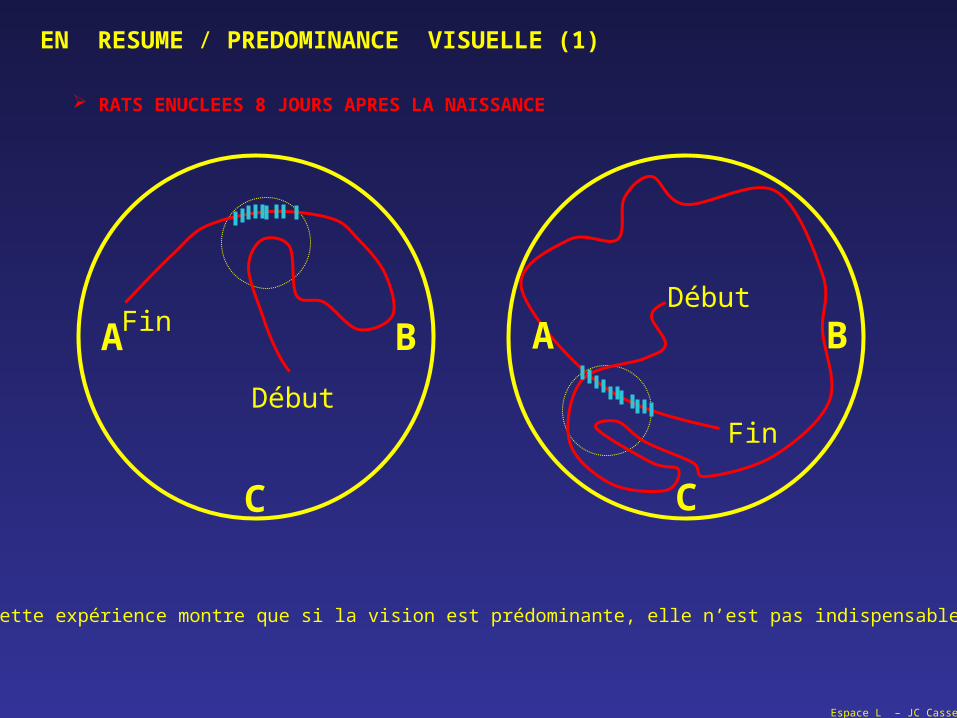
Espace L – JC Cassel
A B
C
Début
Fin
RATS ENUCLEES 8 JOURS APRES LA NAISSANCE
EN RESUME / PREDOMINANCE VISUELLE (1)
B
C
Fin
Début
A
Cette expérience montre que si la vision est prédominante, elle n’est pas indispensable
Espace L – JC Cassel
D'OU:
-Les informations visuelles ne sont pas indispensables à la navigation -La disposition des objets s'avère suffisante à assurer une navigation-Ce dernier système est toutefois moins fiable et nécessite un calibrage plus fréquent (NB : rats aveugles bien plus souvent au contact des objets); il est capable d'intégrer des informations propres aux mouvements effectués (informations kinesthésqiues), et probablement aussi des informations olfactives et tactiles.
DONC :
L'utilisation d'indices locaux non-visuels et d'indices kinesthésiquesest suffisante à assurer une navigation, à condition que l'animal aitpu interagir avec son environnement.
QUESTION :
Tous les indices sont-ils équivalents en matière de contrôle de l'activitédes cellules de lieu?
Jeu sur la configuration, le potentiel d'indication directionnelle d'une configuration d'objets et la proximité des objets par rapport au centre d'une enceinte.
EN RESUME / PREDOMINANCE VISUELLE (2)

Espace L – JC Cassel
Les cellules de lieu.
Farces et attrapes

Espace L – JC Cassel
Idée de jouer sur le niveau d’ambiguité du positionnement d’objets identiques dans un environnement circulaire
Triangle isocèle
Triangle équilatéral
En plus, on met en compétition les indices distaux et les indices proximaux !
INDICES PROXIMAUX vs. DISTAUX ET AMBIGUITE (1)
Espace L – JC Cassel
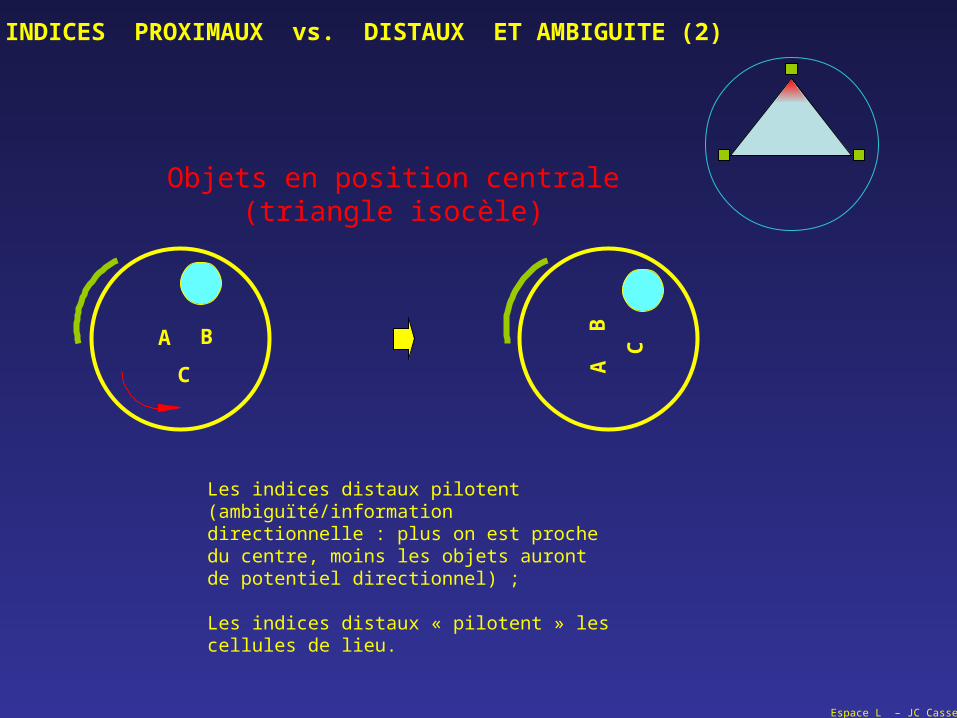
Objets en position centrale(triangle isocèle)
A B
C
Les indices distaux pilotent (ambiguïté/information directionnelle : plus on est proche du centre, moins les objets auront de potentiel directionnel) ;
Les indices distaux « pilotent » les cellules de lieu.
AB
C
INDICES PROXIMAUX vs. DISTAUX ET AMBIGUITE (2)
Espace L – JC Cassel
C
A B
INDICES PROXIMAUX vs. DISTAUX ET AMBIGUITE (3)
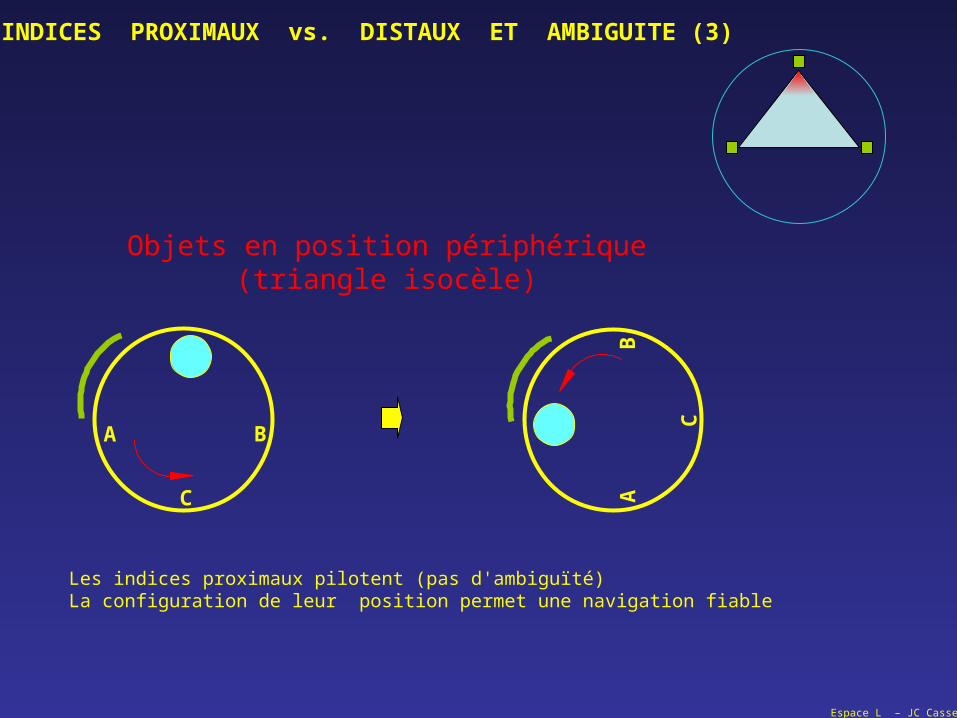
Objets en position périphérique(triangle isocèle)
C
AB
Les indices proximaux pilotent (pas d'ambiguïté)La configuration de leur position permet une navigation fiable
Espace L – JC Cassel
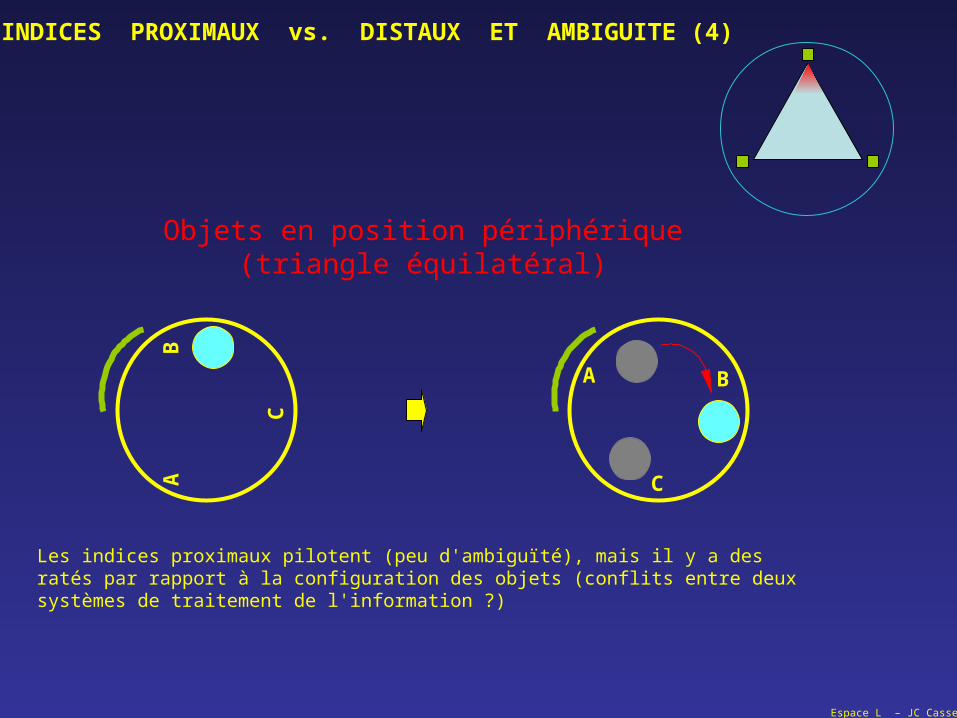
Objets en position périphérique(triangle équilatéral)C
AB
Les indices proximaux pilotent (peu d'ambiguïté), mais il y a desratés par rapport à la configuration des objets (conflits entre deuxsystèmes de traitement de l'information ?)
C
A B
INDICES PROXIMAUX vs. DISTAUX ET AMBIGUITE (4)
Espace L – JC Cassel
EN RESUME
Les cellules de lieu appartiennent à un système complexe de traitement et de codage de l'information spatiale. Ce traitement peut s’opérer sur la base d’informations allothétiques (locales ou distales) ou idiothétiques.
Ce système est très rapide et très souple en matière d’initialisation et de réinitialisation du traitement.
Ce système est alimenté de façon prédominante par l’information visuelle, mais il est capable d'intégrer une multitude d‘autres informations sensorielles (olfactives, auditives, kinesthésiques…). Cette intégration implique d’autres aires que l’hippocampe (cortex visuel, cortex pariétal).
Il sélectionne, voire trie les informations les plus pertinentes en termes de direction, d'organisation spatiale et de stabilité d’un environnement.
On peut le piéger en jouant sur le niveau de cohérence entre différentes sources d’informations (ce qui veut dire qu’on peut en anticiper la réponse et le décalage éventuel).
Espace L – JC Cassel
Hippocampe Subiculum
Cortex entorhinal
Cortex périrhinal Cortex rétrosplénial
Cortex parietal Cortex visuel
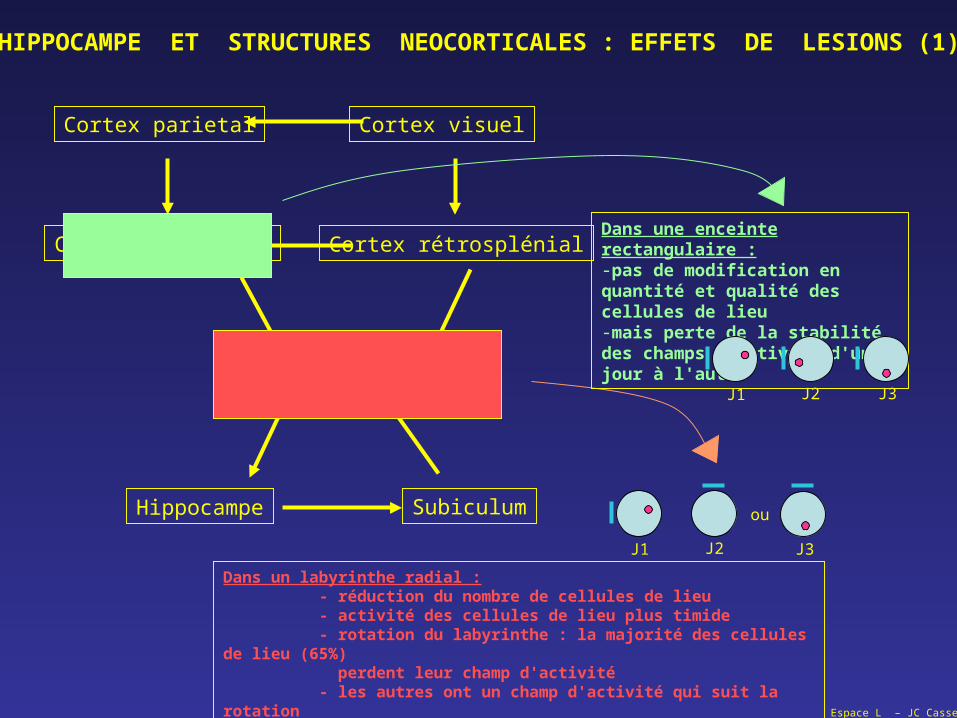
HIPPOCAMPE ET STRUCTURES NEOCORTICALES : EFFETS DE LESIONS (1)
Dans un labyrinthe radial :- réduction du nombre de cellules de lieu- activité des cellules de lieu plus timide- rotation du labyrinthe : la majorité des cellules de lieu (65%) perdent leur champ d'activité- les autres ont un champ d'activité qui suit la rotation
J1 J2
ou
J3
Dans une enceinte rectangulaire :-pas de modification en quantité et qualité des cellules de lieu-mais perte de la stabilité des champs d'activité d'un jour à l'autre
J1 J2 J3
Espace L – JC Cassel
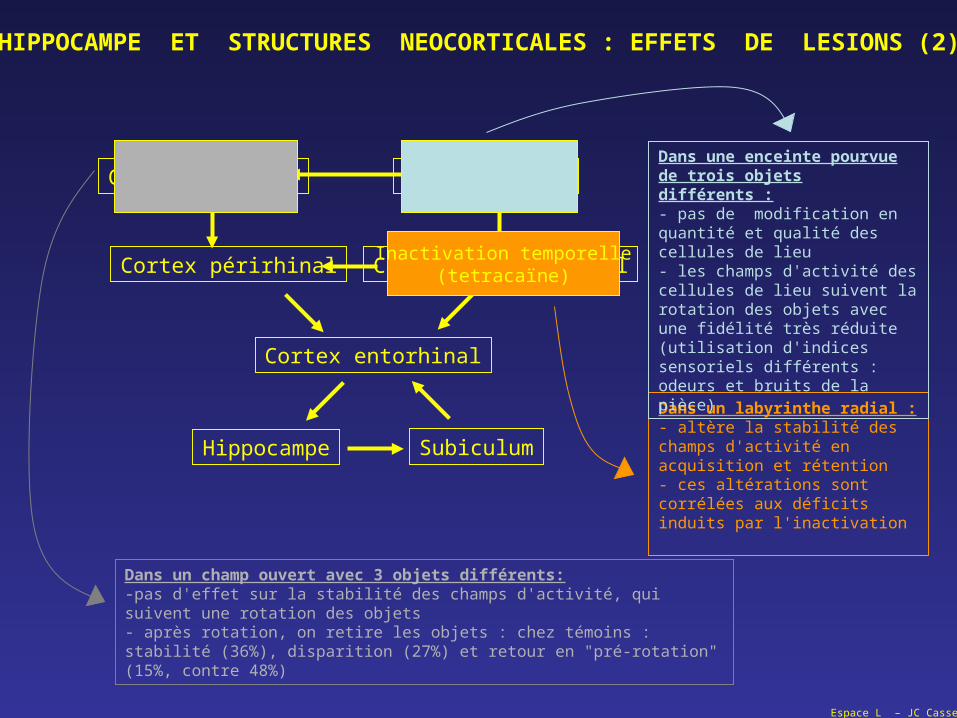
HIPPOCAMPE ET STRUCTURES NEOCORTICALES : EFFETS DE LESIONS (2)
Hippocampe Subiculum
Cortex entorhinal
Cortex périrhinal Cortex rétrosplénial
Cortex parietal Cortex visuel
Dans un champ ouvert avec 3 objets différents:-pas d'effet sur la stabilité des champs d'activité, qui suivent une rotation des objets- après rotation, on retire les objets : chez témoins : stabilité (36%), disparition (27%) et retour en "pré-rotation" (15%, contre 48%)
Inactivation temporelle(tetracaïne)
Dans un labyrinthe radial :- altère la stabilité des champs d'activité en acquisition et rétention- ces altérations sont corrélées aux déficits induits par l'inactivation
Dans une enceinte pourvue de trois objets différents :- pas de modification en quantité et qualité des cellules de lieu- les champs d'activité des cellules de lieu suivent la rotation des objets avec une fidélité très réduite (utilisation d'indices sensoriels différents : odeurs et bruits de la pièce)

Espace L – JC Cassel
HIPPOCAMPE ET STRUCTURES NEOCORTICALES : EFFETS DE LESIONS (3)
DONC :
Une atteinte de l'une ou l'autre des régions corticales connectées directement ou indirectement à l'hippocampe, et qui a (ou non) pour conséquence une altération des performances cognitives d'ordre spatial, peut aussi interférer avec l'une ou l'autre des caractéristiques fonctionnelles des cellules de lieu, mais sans oblitérer leur fonctionnalité.
Cette interférence ne se fait pas en « tout ou rien », ce qui suggère que le mécanisme d’intégration de l’information spatiale est distribué, plurimodal et plastique, au moins dans une certaine mesure.
Qu’entendre par distribution ? Il y a des structures qui acheminent de l’information spatiale, d’autres qui intègrent ces informations et certaines qui les stockent. Le traitement et l’intégration peuvent être étagés.
Les cellules de lieu ayant été décrites dans l’hippocampe, cette structure pourrait jouer un rôle central dans les processus d’intégration, éventuellement de stockage (stabilité des champs d’activité), de l’information spatiale.
Effets d’une atteinte ou d’une altération des fonctions hippocampiques ?

Espace L – JC Cassel
Les cellules de lieu
Mémoire de lieu ou lieu de mémoire ?
Espace L – JC Cassel
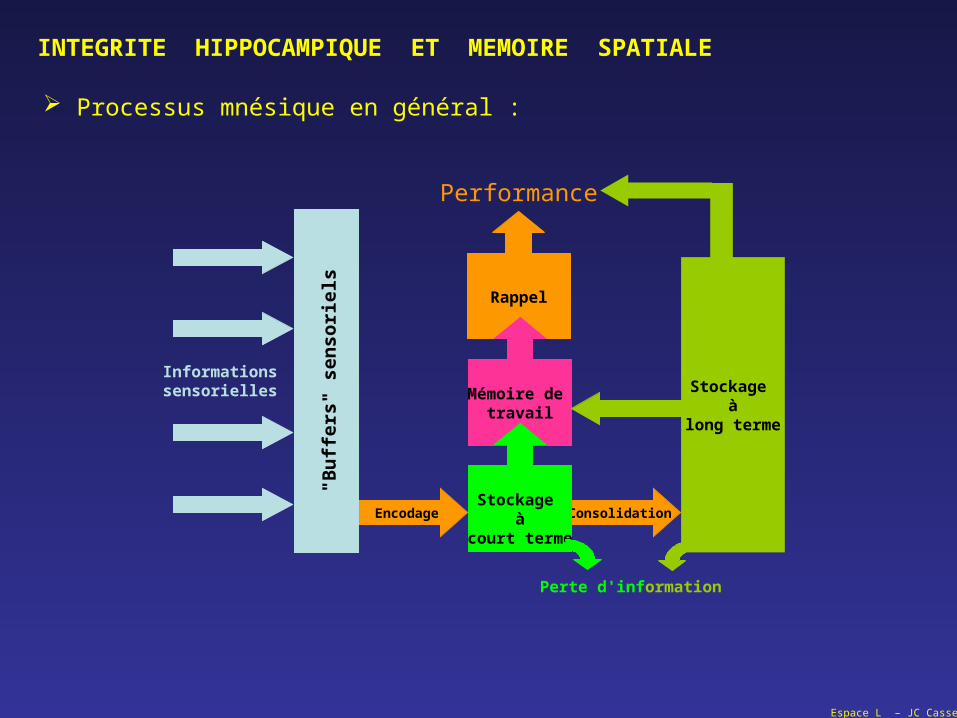
INTEGRITE HIPPOCAMPIQUE ET MEMOIRE SPATIALE
Processus mnésique en général :
Rappel
Stockage à
long terme
Encodage Consolidation
Mémoire de travail
Stockage à
court terme
Performance
Informationssensorielles
Perte d'information
"Bu
ffer
s" s
enso
riel
s
Espace L – JC Cassel
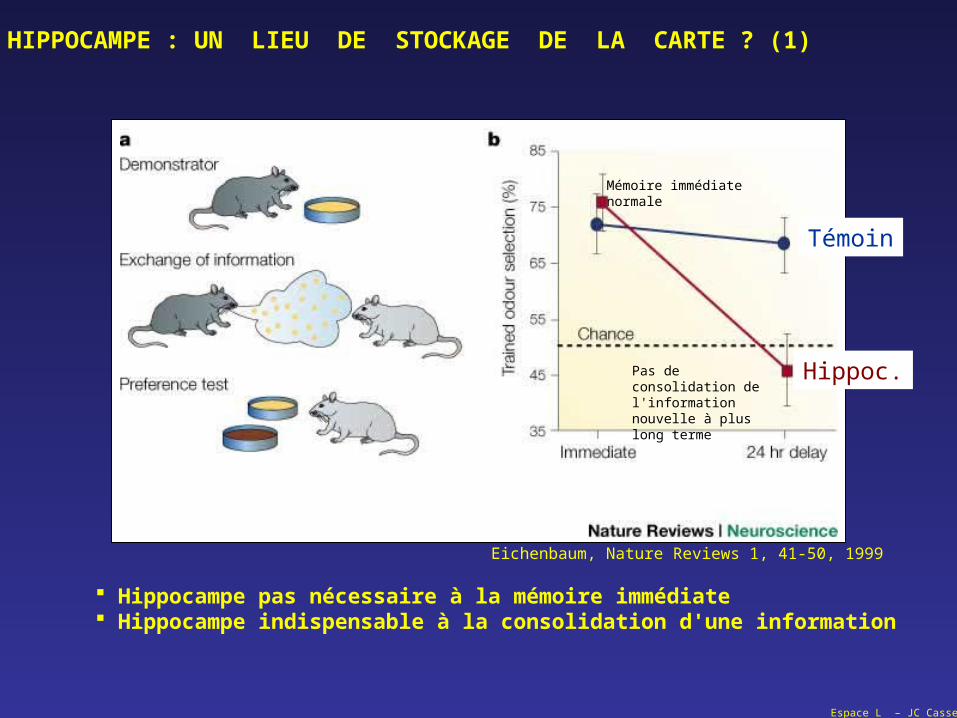
Hippocampe pas nécessaire à la mémoire immédiate Hippocampe indispensable à la consolidation d'une information
Mémoire immédiatenormale
Pas de consolidation de l'information nouvelle à plus long terme
Eichenbaum, Nature Reviews 1, 41-50, 1999
Témoin
Hippoc.
HIPPOCAMPE : UN LIEU DE STOCKAGE DE LA CARTE ? (1)
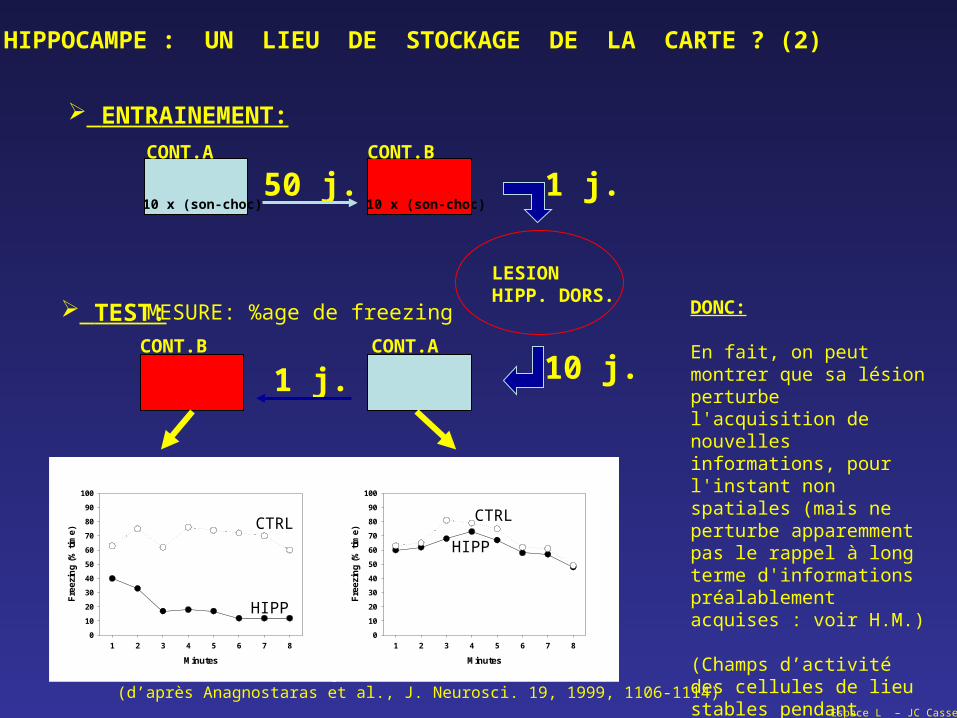
Espace L – JC Cassel
CONT.A CONT.B
LESIONHIPP. DORS.
50 j. 1 j.
ENTRAINEMENT:
10 x (son-choc) 10 x (son-choc)
CONT.ACONT.B
10 j.1 j.
TEST: MESURE: %age de freezing
Minutes
1 2 3 4 5 6 7 8
Fre
ezin
g (
% t
ime)
0
10
20
30
40
50
60
70
80
90
100
Minutes
1 2 3 4 5 6 7 8
Fre
ezin
g (
% t
ime)
0
10
20
30
40
50
60
70
80
90
100
HIPP
HIPP
CTRL CTRL
(d’après Anagnostaras et al., J. Neurosci. 19, 1999, 1106-1114)
HIPPOCAMPE : UN LIEU DE STOCKAGE DE LA CARTE ? (2)
DONC:
En fait, on peut montrer que sa lésion perturbe l'acquisition de nouvelles informations, pour l'instant non spatiales (mais ne perturbe apparemment pas le rappel à long terme d'informations préalablement acquises : voir H.M.)
(Champs d’activité des cellules de lieu stables pendant plusieurs semaines…)
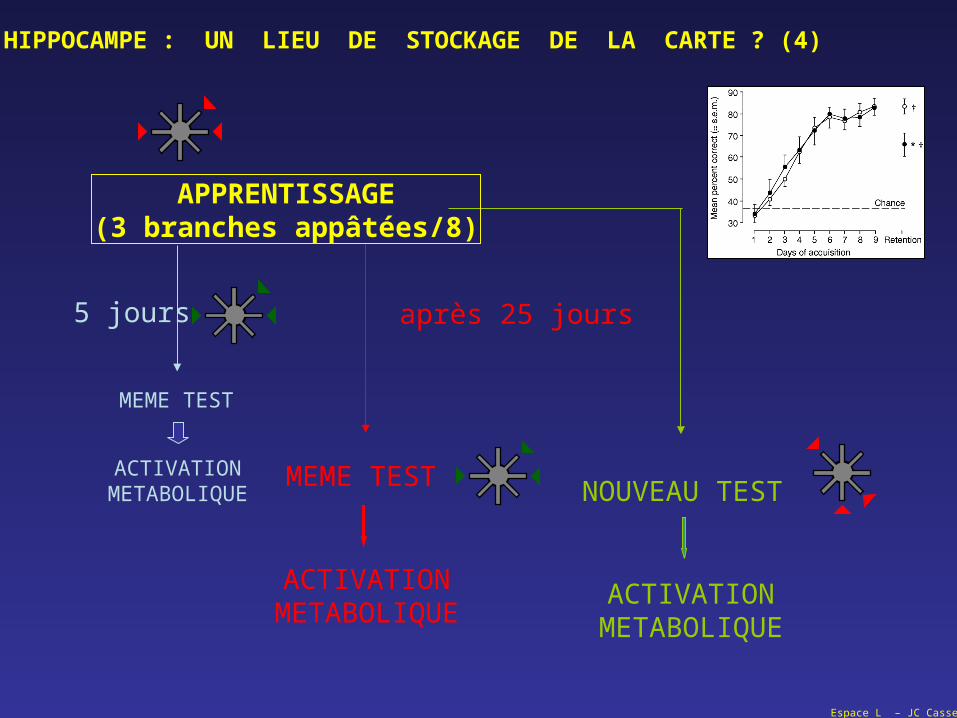
Espace L – JC Cassel
5 jours après 25 jours
MEME TEST
MEME TEST
ACTIVATIONMETABOLIQUE
NOUVEAU TESTACTIVATION
METABOLIQUE
ACTIVATIONMETABOLIQUE
HIPPOCAMPE : UN LIEU DE STOCKAGE DE LA CARTE ? (4)
APPRENTISSAGE(3 branches appâtées/8)
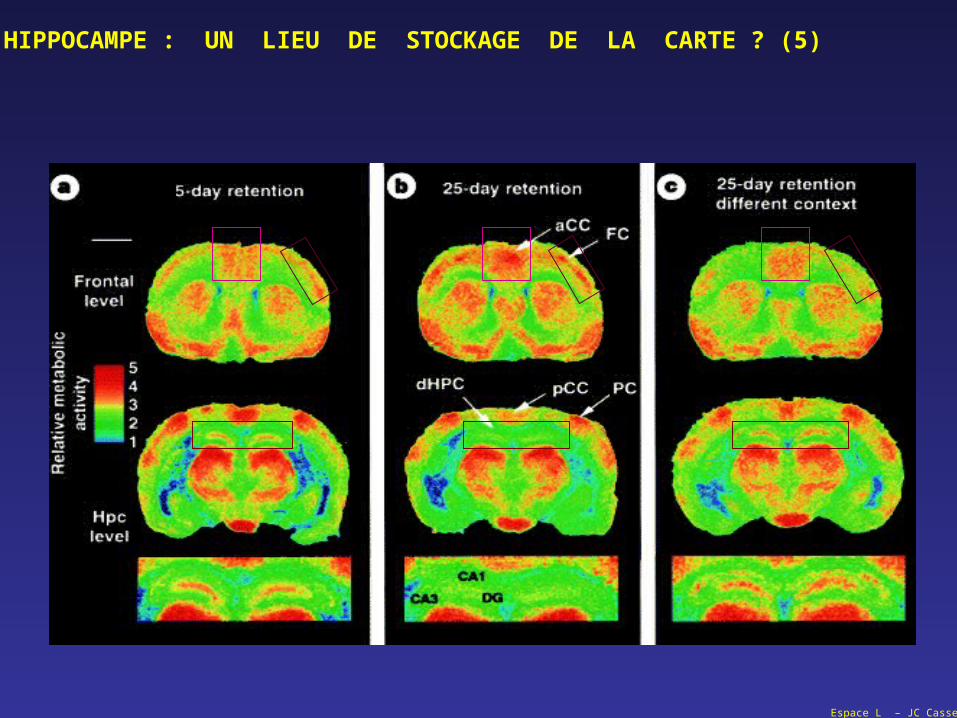
Espace L – JC Cassel
HIPPOCAMPE : UN LIEU DE STOCKAGE DE LA CARTE ? (5)

QUESTION :
Espace L – JC Cassel
EN RESUME
une lésion de l'hippocampe perturbe la mémoire épisodique (de travail) à caractère spatial,
une lésion de l'hippocampe ne perturbe pas le rappel d'une information spatiale mémorisée (et consolidée) depuis un certain temps (long terme),
un apprentissage spatial en cours ou le rappel d'un apprentissage récent se traduit par une activation de l'hippocampe (et l’inactivation de l’hippocampe empêche cette opération),
le rappel d'un apprentissage spatial établi depuis un certain temps s'accompagne d'une activation néocorticale, et non d'une activation de l'hippocampe (et l’inactivation du néocortex empêche cette opération)
la confrontation à une nouvelle tâche, y compris dans le même dispositif, provoque une nouvelle activation hippocampique.
Les cellules de lieu ont, en principe, un champ d’activité stable – Consolidation ?
Altération de la consolidation altération de cette stabilité ?Nous renseignerait sur leur éventuelle implication dans un processus mnésique !
Espace L – JC Cassel
CELLULES DE LIEU ET TRACE MNESIQUE :
Attributs d’une trace mnésique :
fonction de l’expérience(la formation de la trace est fonction d’une expérience)
spécifique à l’information acquise
durable(la disponibilité de la trace doit persister au-delà de l’exposition à l’information à mémoriser; nécessite une consolidation)
doit être réactivable(la présentation ultérieure d’une partie au moins des indices initiaux devrait réactiver la trace)

Espace L – JC Cassel
RECEPTEURS NMDA ET CONSOLIDATION
Le labyrinthe radial : test d’apprentissage et de mémoire spatiale
Espace L – JC Cassel
ROOM 2MK-801
4 8
Wo
rkin
g m
emo
ry e
rro
rs
0
1
2
3
4
5
1
2
3
4
5
1 2 3 4 5 6 7 8
ROOM 1
No Drug MK-801
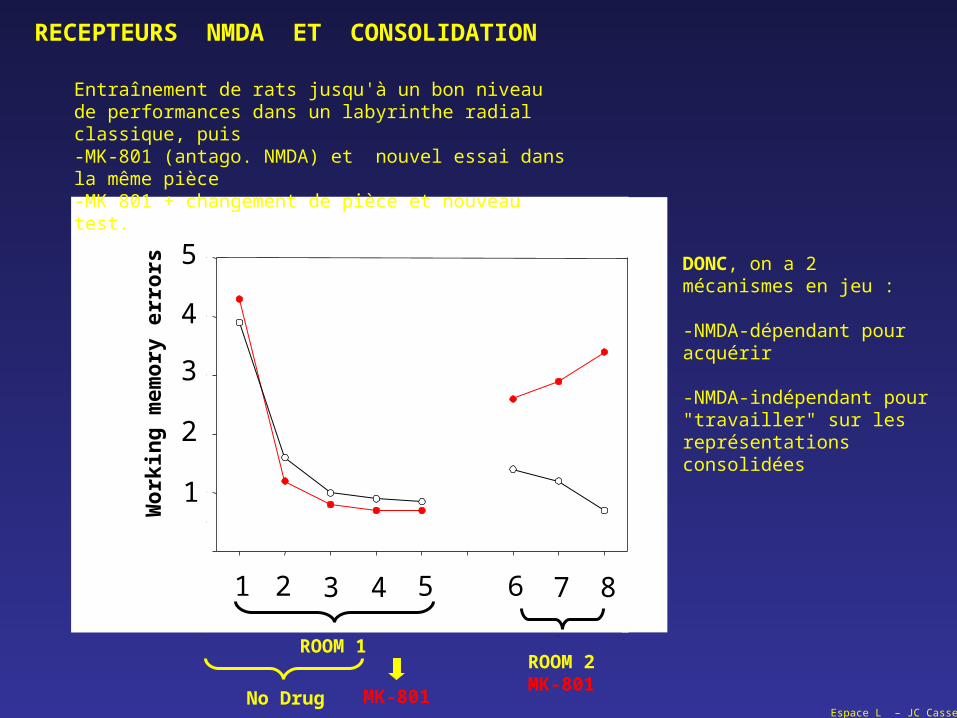
Entraînement de rats jusqu'à un bon niveau de performances dans un labyrinthe radial classique, puis-MK-801 (antago. NMDA) et nouvel essai dans la même pièce-MK 801 + changement de pièce et nouveau test.
DONC, on a 2 mécanismes en jeu :
-NMDA-dépendant pour acquérir
-NMDA-indépendant pour "travailler" sur les représentations consolidées
RECEPTEURS NMDA ET CONSOLIDATION
Espace L – JC Cassel
RECEPTEURS NMDA ET CONSOLIDATION
TRIAL
LA
TE
NC
Y
0
10
20
30
40
50
60
70
80
9015 sec 120 min20 min
CTRL
HIPP
1 2221 1
LESION HIPPOCAMPIQUE
TRIAL
1 2 1 2 1 2
LA
TE
NC
Y
0
20
40
60
80
D-AP5, i.c.v.
15 sec 120 min20 min
LCRa
INJECTION
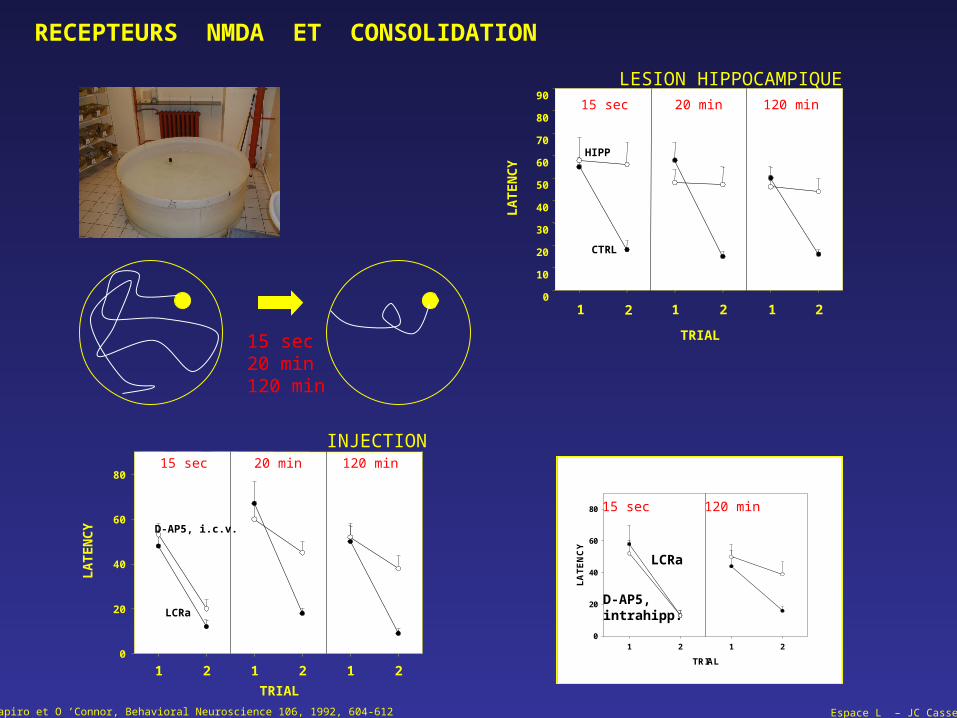
Shapiro et O ’Connor, Behavioral Neuroscience 106, 1992, 604-612
15 sec20 min120 min
TRIAL
1 2 1 2
LA
TE
NC
Y
0
20
40
60
80
D-AP5,intrahipp.
LCRa
15 sec 120 min
Espace L – JC Cassel
RECEPTEURS NMDA, CONSOLIDATION ET CELLULES DE LIEU
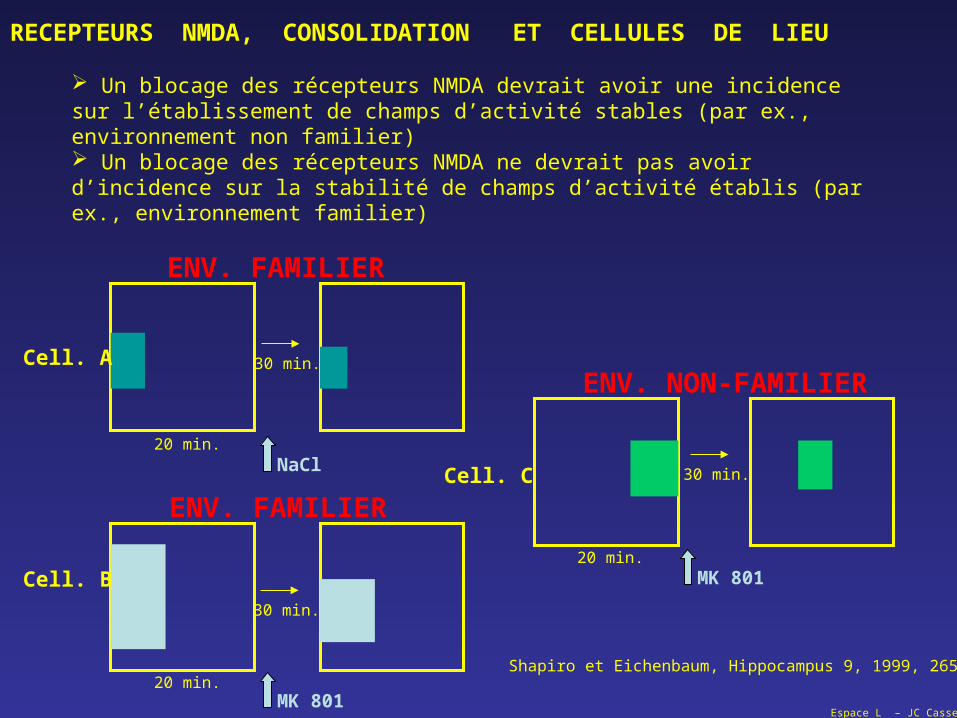
Un blocage des récepteurs NMDA devrait avoir une incidence sur l’établissement de champs d’activité stables (par ex., environnement non familier) Un blocage des récepteurs NMDA ne devrait pas avoir d’incidence sur la stabilité de champs d’activité établis (par ex., environnement familier)
Cell. A
Cell. B
ENV. FAMILIER
ENV. FAMILIER
30 min.
30 min.
20 min.
20 min.MK 801
NaCl Cell. C
ENV. NON-FAMILIER
30 min.
20 min.MK 801
Shapiro et Eichenbaum, Hippocampus 9, 1999, 265-384

Espace L – JC Cassel
RECEPTEURS NMDA , CONSOLIDATION ET CELLULES DE LIEU
Donc:
Un blocage de la consolidation (par blocage des récepteurs NMDA) bloque la stabilisation du champ d’activité de la cellule de lieu.
Par contre, le blocage des récepteurs NMDA ne modifie pas la stabilité des champs d’activité dans un environnement connu.
Espace L – JC Cassel
RECEPTEURS AMPA, CONSOLIDATION ET CELLULES DE LIEU
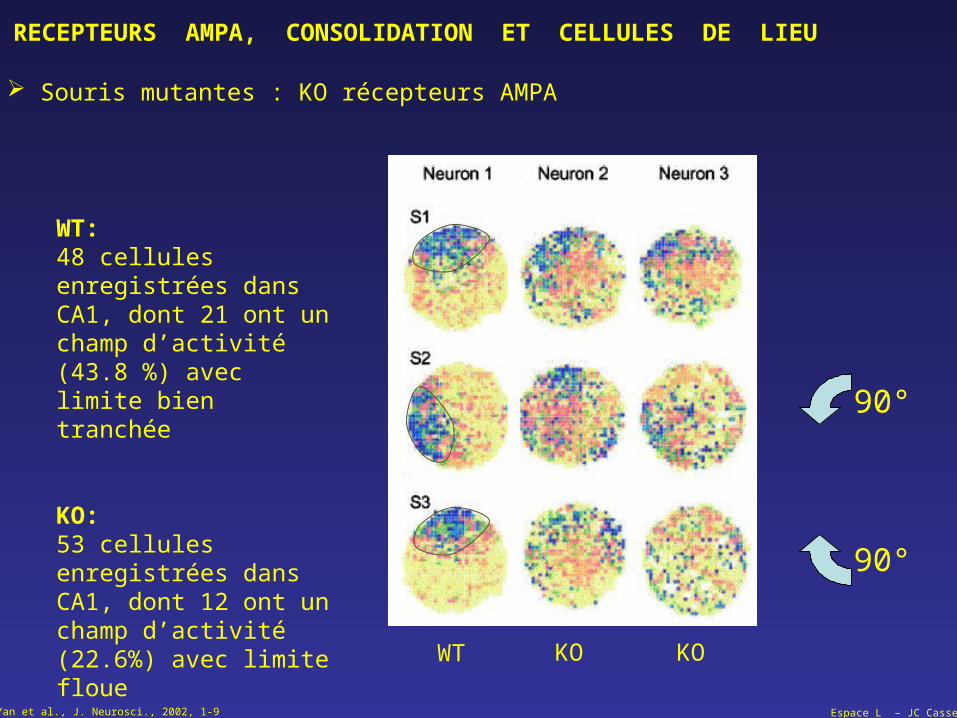
Souris mutantes : KO récepteurs AMPA
WT KO KO
90°
90°
WT:48 cellules enregistrées dans CA1, dont 21 ont un champ d’activité (43.8 %) avec limite bien tranchée
KO:53 cellules enregistrées dans CA1, dont 12 ont un champ d’activité (22.6%) avec limite floue
Yan et al., J. Neurosci., 2002, 1-9
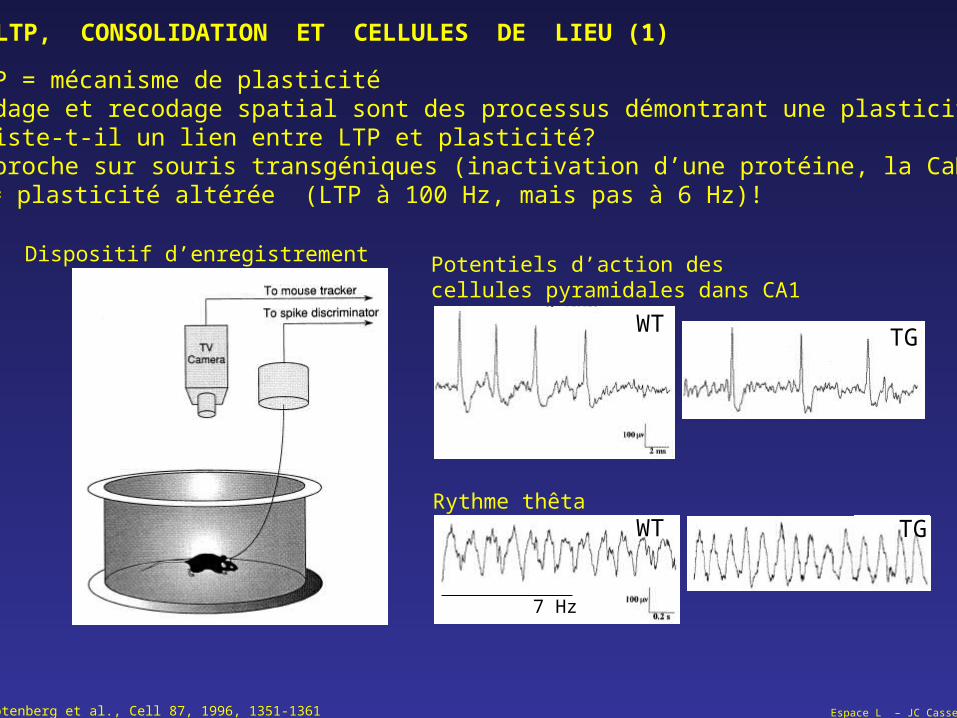
Espace L – JC CasselRotenberg et al., Cell 87, 1996, 1351-1361
LTP, CONSOLIDATION ET CELLULES DE LIEU (1)
LTP = mécanisme de plasticité Codage et recodage spatial sont des processus démontrant une plasticité Existe-t-il un lien entre LTP et plasticité? Approche sur souris transgéniques (inactivation d’une protéine, la CaMKII) = plasticité altérée (LTP à 100 Hz, mais pas à 6 Hz)!
Dispositif d’enregistrement Potentiels d’action des cellules pyramidales dans CA1
Rythme thêta
WTTG
WT TG
7 Hz
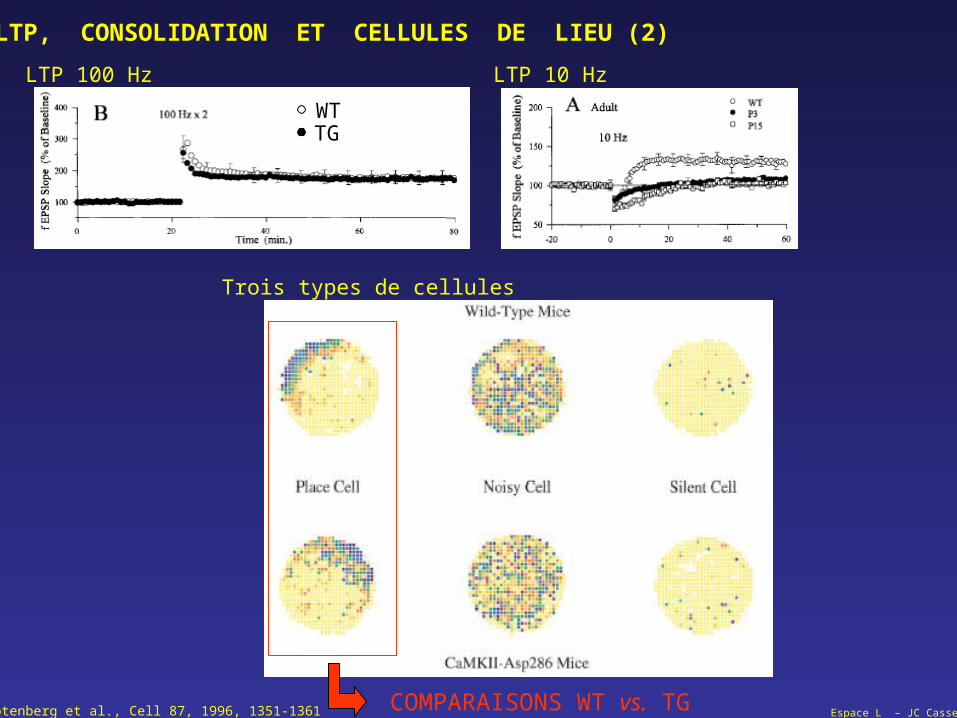
Espace L – JC CasselRotenberg et al., Cell 87, 1996, 1351-1361
LTP, CONSOLIDATION ET CELLULES DE LIEU (2)
Trois types de cellules
COMPARAISONS WT vs. TG
WTTG
LTP 100 Hz LTP 10 Hz
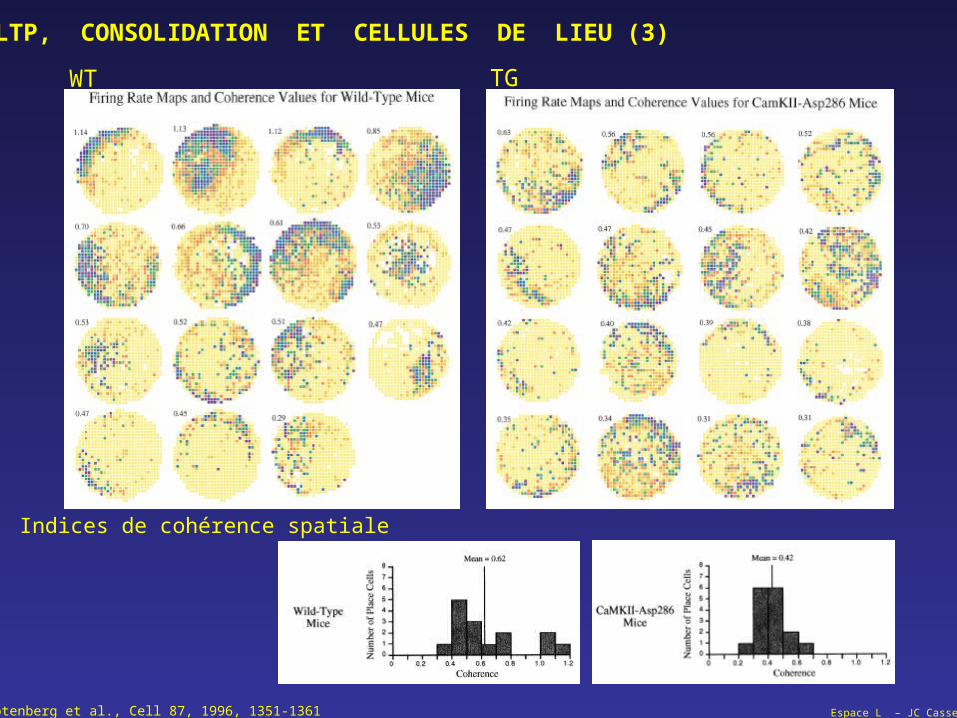
Espace L – JC CasselRotenberg et al., Cell 87, 1996, 1351-1361
LTP, CONSOLIDATION ET CELLULES DE LIEU (3)
WT TG
Indices de cohérence spatiale
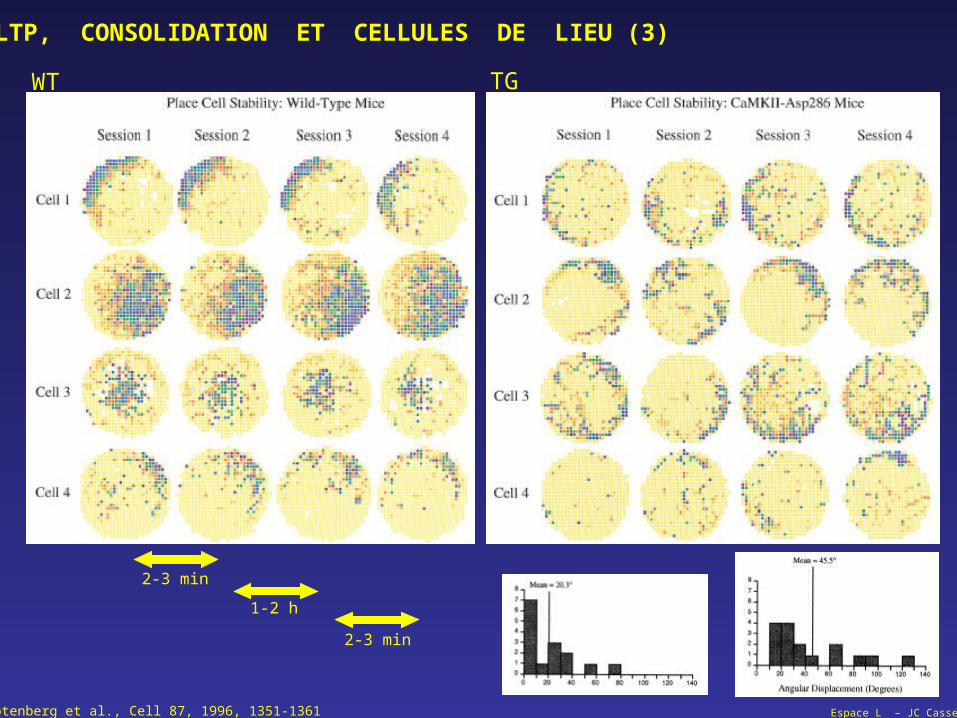
Espace L – JC CasselRotenberg et al., Cell 87, 1996, 1351-1361
LTP, CONSOLIDATION ET CELLULES DE LIEU (3)
WT TG
2-3 min
2-3 min
1-2 h
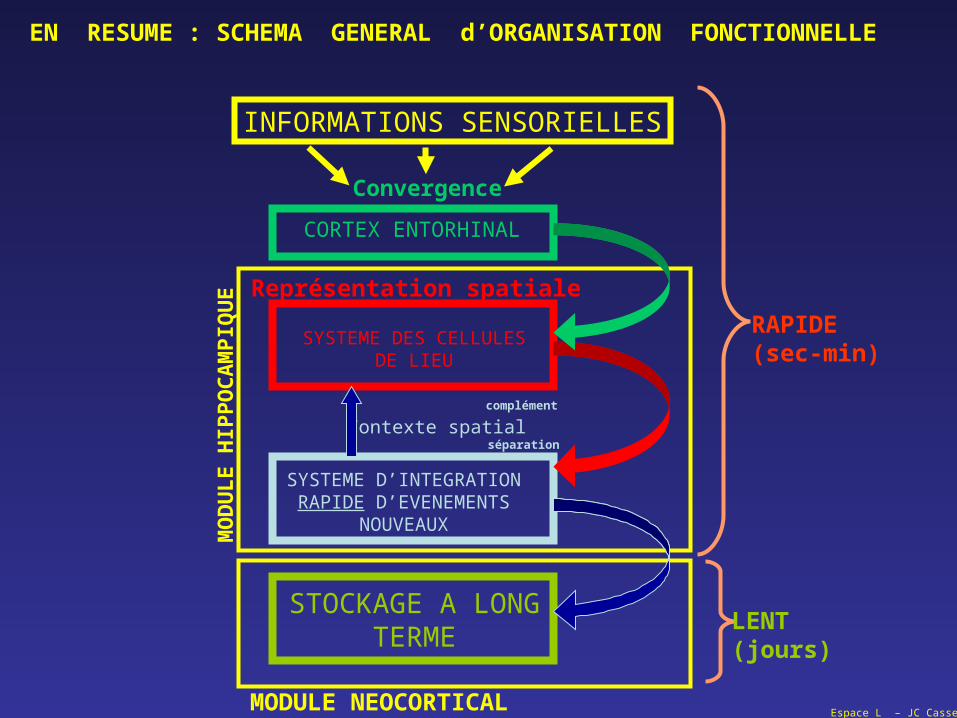
Espace L – JC Cassel
SYSTEME DES CELLULESDE LIEU
SYSTEME D’INTEGRATIONRAPIDE D’EVENEMENTS
NOUVEAUX
STOCKAGE A LONGTERME
CORTEX ENTORHINAL
INFORMATIONS SENSORIELLES
Contexte spatial
Représentation spatiale
MODULE NEOCORTICAL
MO
DU
LE
HIP
PO
CA
MP
IQU
E
Convergence
complément
séparation
LENT(jours)
RAPIDE(sec-min)
EN RESUME : SCHEMA GENERAL d’ORGANISATION FONCTIONNELLE
Espace L – JC Cassel
A partir de ce qui précède, on serait assez tenté de considérer l'hippocampe comme la structure cérébrale où sont traitées, encodées, rappelées… toute une série d'informations concernant les caractéristiques topographiques, proximales et distales, de l'environnement dans lequel évolue un animal comme le Rat.
Attention, un tel glissement apparaît assez rapidement comme décalé par rapport à une partie de la littérature, ce que Shapiro et Eichenbaum rappellent dans une revue de question de 1999.
En effet, la prise en compte d'un encodage de l'information "purement" spatiale ne suffit pas à expliquer tous les résultats expérimentaux publiés dans la littérature, et de loin pas (de la même façon, le codage spatial sollicite également d’autres cellules) !
Voyons quelques exemples :
REMARQUES :
Espace L – JC Cassel
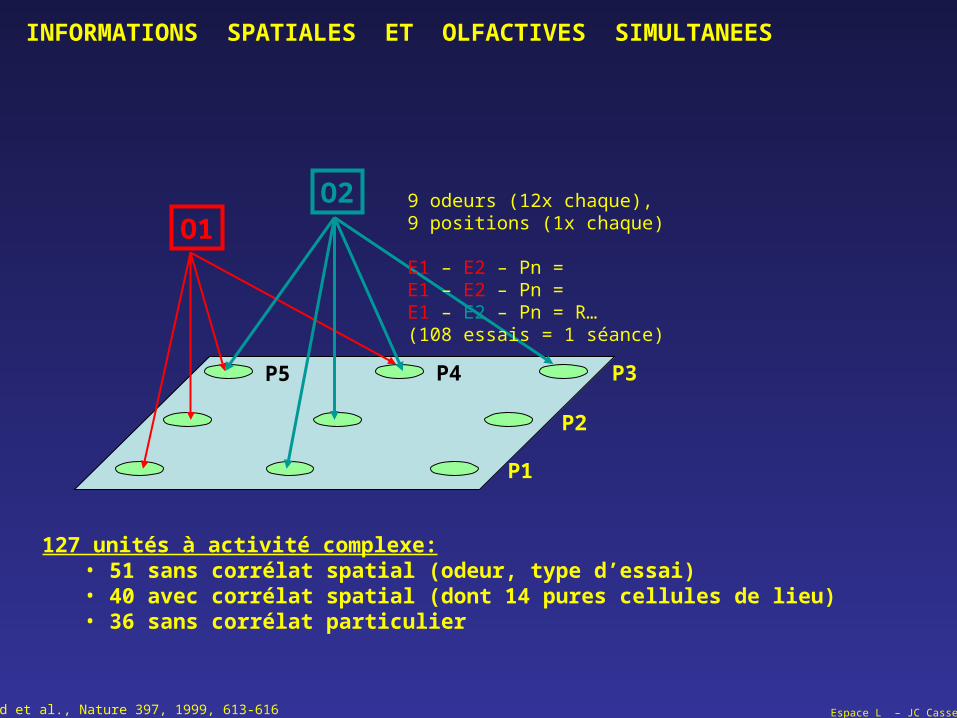
O1O2
P3
P2
P4
127 unités à activité complexe:• 51 sans corrélat spatial (odeur, type d’essai)• 40 avec corrélat spatial (dont 14 pures cellules de lieu)• 36 sans corrélat particulier
P1
P5
9 odeurs (12x chaque),9 positions (1x chaque)
E1 – E2 – Pn = E1 – E2 – Pn = E1 – E2 – Pn = R…(108 essais = 1 séance)
Wood et al., Nature 397, 1999, 613-616
INFORMATIONS SPATIALES ET OLFACTIVES SIMULTANEES
Espace L – JC Cassel
O1 O2 O3 O4 O5 O6 O7 O8 O9
FIR
ING
RA
TE
(H
z)
0
1
2
3
4
5
6
7
8
9
10
P1 P2 P3 P4 P5 P6 P7 P8 P9
FIR
ING
RA
TE
(H
z)
0
2
4
6
8
10
P1 P2 P3 P4 P5 P6 P7 P8 P9
FIR
ING
RA
TE
(H
z)
0
1
2
3
4
5
6
O1 O2 O3 O4 O5 O6 O7 O8 O9
FIR
ING
RA
TE
(H
z)
0
1
2
3
4
5
6
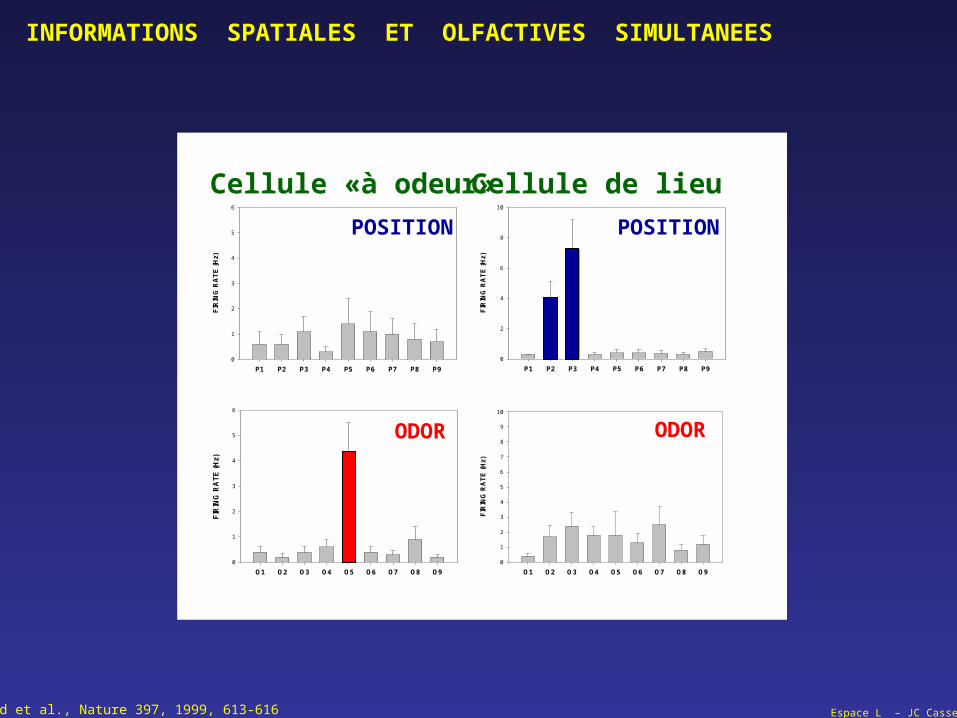
POSITION POSITION
ODOR ODOR
Cellule «à odeur» Cellule de lieu
Wood et al., Nature 397, 1999, 613-616
INFORMATIONS SPATIALES ET OLFACTIVES SIMULTANEES

Espace L – JC Cassel
Cellules de lieu et direction
Par où pour Caen ?
Espace L – JC Cassel
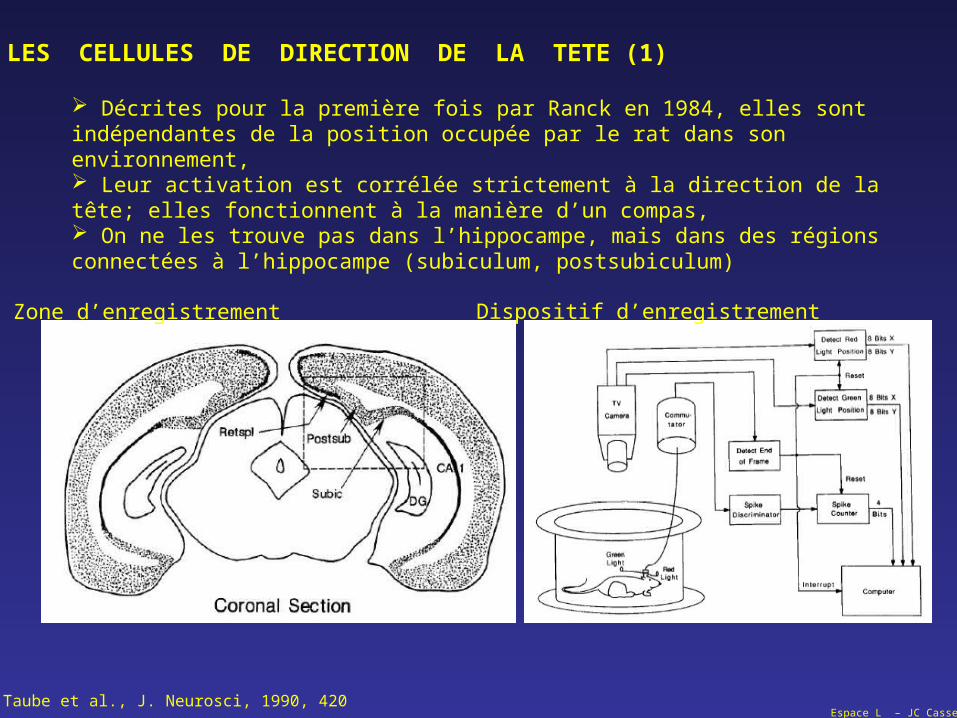
LES CELLULES DE DIRECTION DE LA TETE (1)
Décrites pour la première fois par Ranck en 1984, elles sont indépendantes de la position occupée par le rat dans son environnement, Leur activation est corrélée strictement à la direction de la tête; elles fonctionnent à la manière d’un compas, On ne les trouve pas dans l’hippocampe, mais dans des régions connectées à l’hippocampe (subiculum, postsubiculum)
Taube et al., J. Neurosci, 1990, 420
Zone d’enregistrement Dispositif d’enregistrement
Espace L – JC Cassel
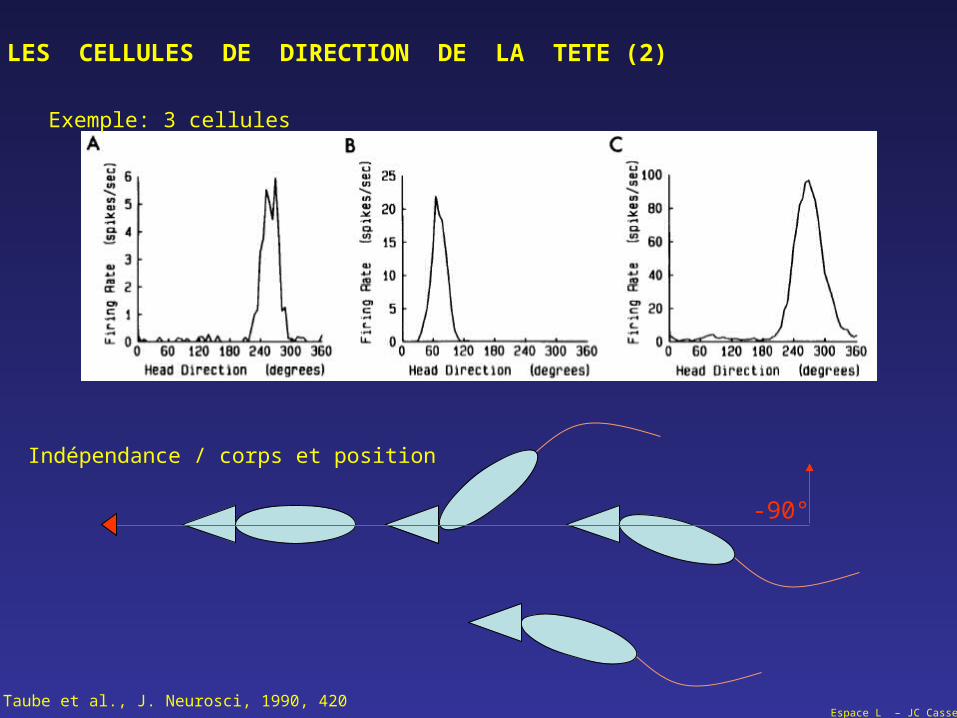
LES CELLULES DE DIRECTION DE LA TETE (2)
-90°
Taube et al., J. Neurosci, 1990, 420
Exemple: 3 cellules
Indépendance / corps et position
Espace L – JC Cassel
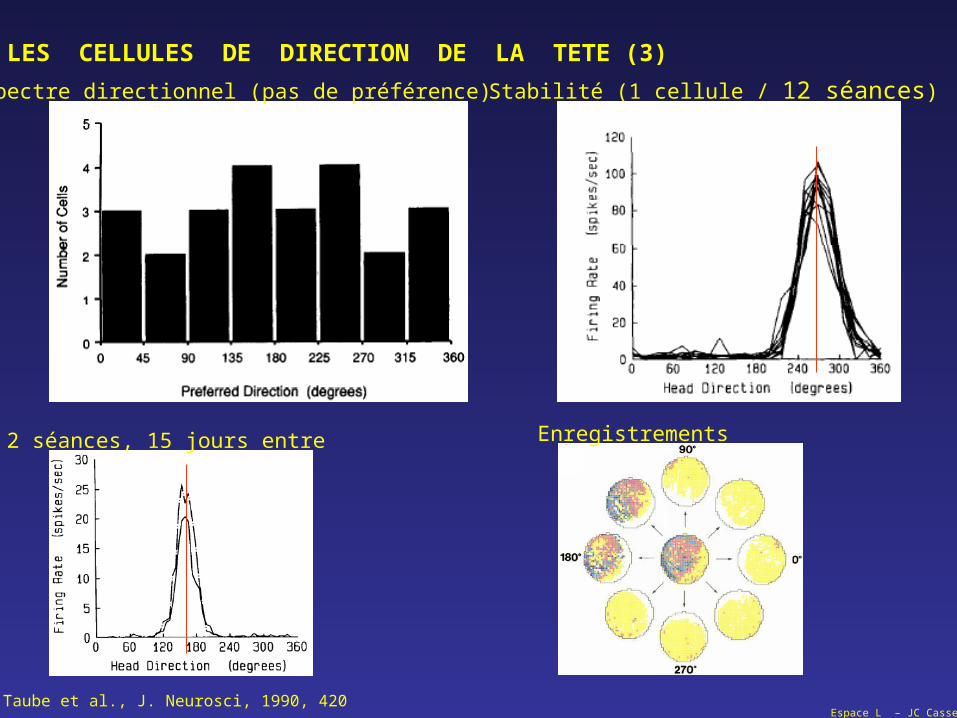
LES CELLULES DE DIRECTION DE LA TETE (3)
Taube et al., J. Neurosci, 1990, 420
Spectre directionnel (pas de préférence) Stabilité (1 cellule / 12 séances)
2 séances, 15 jours entre Enregistrements

Espace L – JC Cassel
LES CELLULES DE DIRECTION DE LA TETE (4)
Déterminants sensoriels : précautions méthodologiques (neutralisations des informations visuelles distales, olfactives et auditives; animal retiré avant tout changement) Plusieurs appareils sont utilisés :
ou ou carte à 3 h
Ordre des séances d’enregistrement :
standard (enceinte circulaire) – rotation de la carte – standard – retrait de la carte – standard – rectangle – standard – carré – standard
Effet de la rotation de la carte :
A: carte à 6 h (+90°)
B: carte à 9 h (180°)
La rotation n’est donc pas stricte-ment identique à celle de la carte, mais toujours elle s’en rapproche.
162°72°
Taube et al., J. Neurosci, 1990, 436
Espace L – JC Cassel
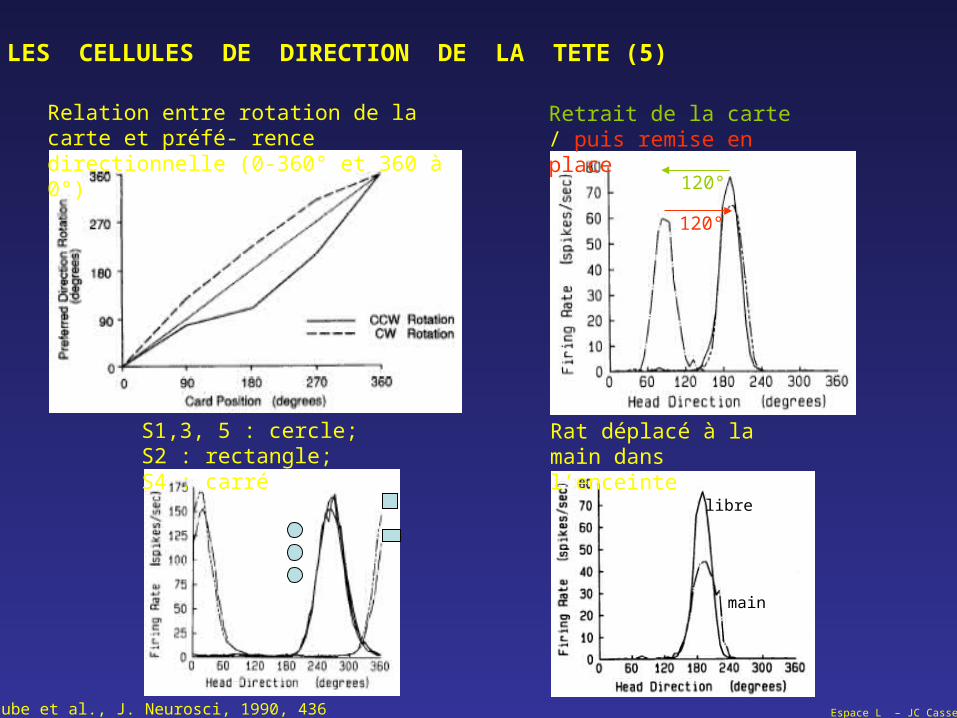
LES CELLULES DE DIRECTION DE LA TETE (5)
Relation entre rotation de la carte et préfé- rence directionnelle (0-360° et 360 à 0°)
Retrait de la carte / puis remise en place
120°
120°
S1,3, 5 : cercle; S2 : rectangle; S4 : carré
Rat déplacé à la main dans l’enceinte
libre
main
Taube et al., J. Neurosci, 1990, 436
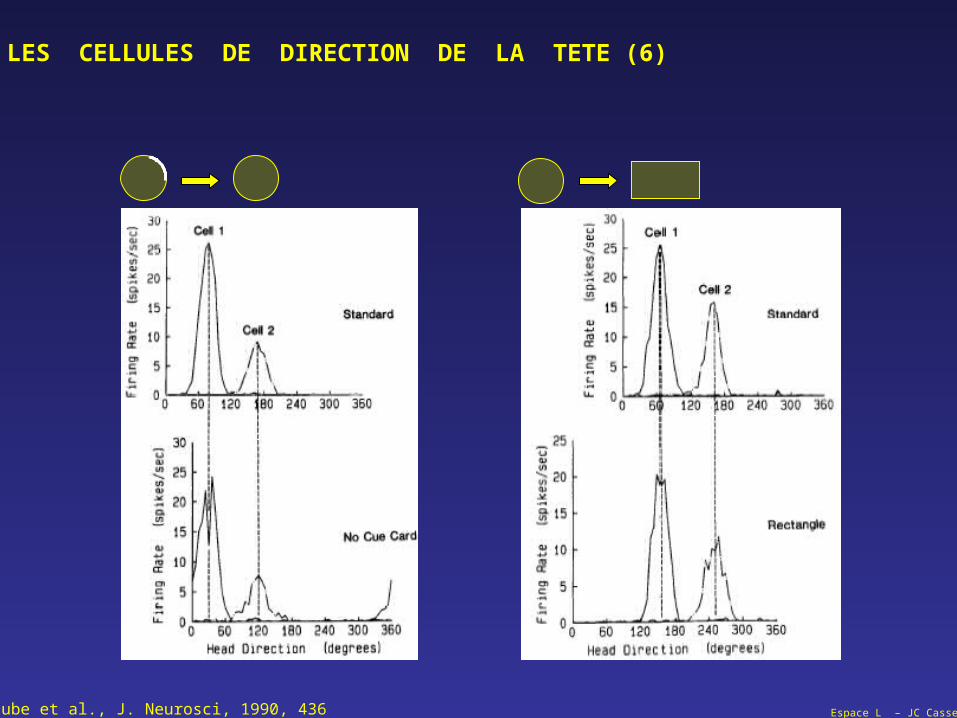
Espace L – JC Cassel
LES CELLULES DE DIRECTION DE LA TETE (6)
Taube et al., J. Neurosci, 1990, 436
Espace L – JC Cassel
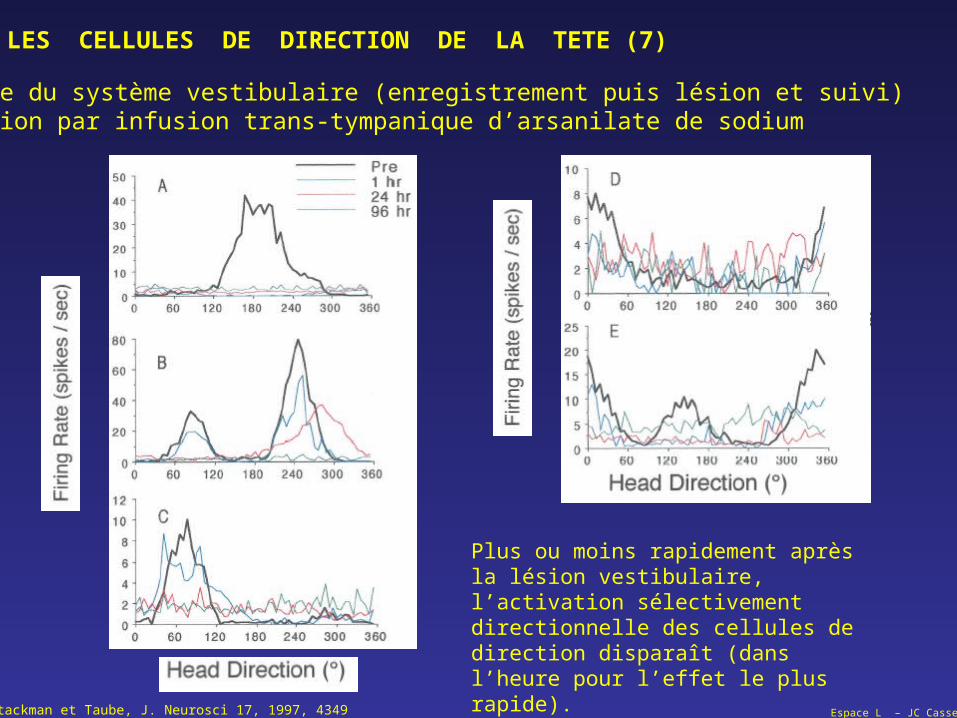
LES CELLULES DE DIRECTION DE LA TETE (7)
Rôle du système vestibulaire (enregistrement puis lésion et suivi) Lésion par infusion trans-tympanique d’arsanilate de sodium
Stackman et Taube, J. Neurosci 17, 1997, 4349
Plus ou moins rapidement après la lésion vestibulaire, l’activation sélectivement directionnelle des cellules de direction disparaît (dans l’heure pour l’effet le plus rapide).
Espace L – JC Cassel
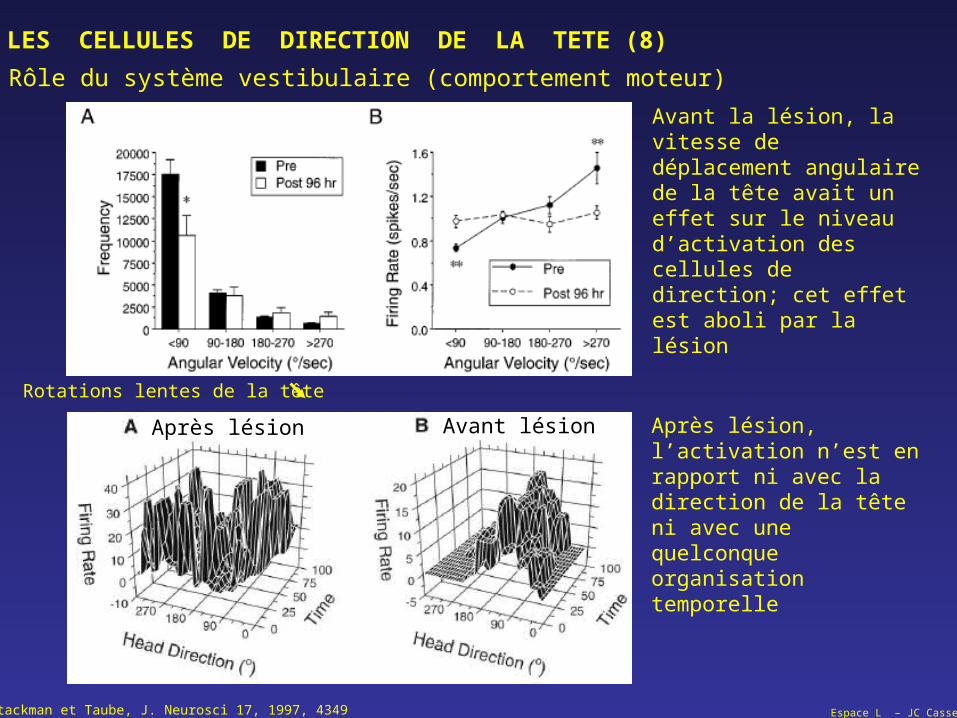
LES CELLULES DE DIRECTION DE LA TETE (8)
Rôle du système vestibulaire (comportement moteur)
Stackman et Taube, J. Neurosci 17, 1997, 4349
Rotations lentes de la tête
Avant la lésion, la vitesse de déplacement angulaire de la tête avait un effet sur le niveau d’activation des cellules de direction; cet effet est aboli par la lésion
Après lésion, l’activation n’est en rapport ni avec la direction de la tête ni avec une quelconque organisation temporelle
Après lésion Avant lésion
Espace L – JC Cassel
LES CELLULES DE DIRECTION DE LA TETE : EN RESUME
activation maximale lorsque l’axe de symétrie de la tête du rat pointe dans une direction particulière; l’activation décroît de part et d’autre de cet axe (base environ 90° et activation maximale entre 5 et 120 PA/s; 0-50 PA/s pour les cellules de lieu),
cette direction est indépendante de l’emplacement de l’animal dans un environnement donné, et il n’y a qu’une seule direction préférentielle par cellule (on peut avoir plus d’un champ d’activité pour les cellules de lieu),
leur activité est basée sur des indices visuels (comme les cellules de lieu) et lorsque ces derniers sont déplacés ou retirés, l’activité des cellules persiste, mais leur direction préférée est modifiée (lorsqu’ils sont modifiés en présence du rat, ce dernier peut se baser sur d’autres indices stables et ses cellules de direction de la tête peuvent conserver une direction préférentielle constante),
la modification des indices visuels altère la direction préférentielle, mais ne semble pas avoir d’impact sur l’écart entre les directions préférentielles différentes cellules de direction de la tête,
DONC: comme c’est le cas pour les cellules de lieu, leurs propriétés sont décrites au mieux dans leur lien avec les relations qu’un animal établit avec son environnement. Le système vestibulaire joue un rôle déterminant dans le fonctionnement de ces cellules de direction.
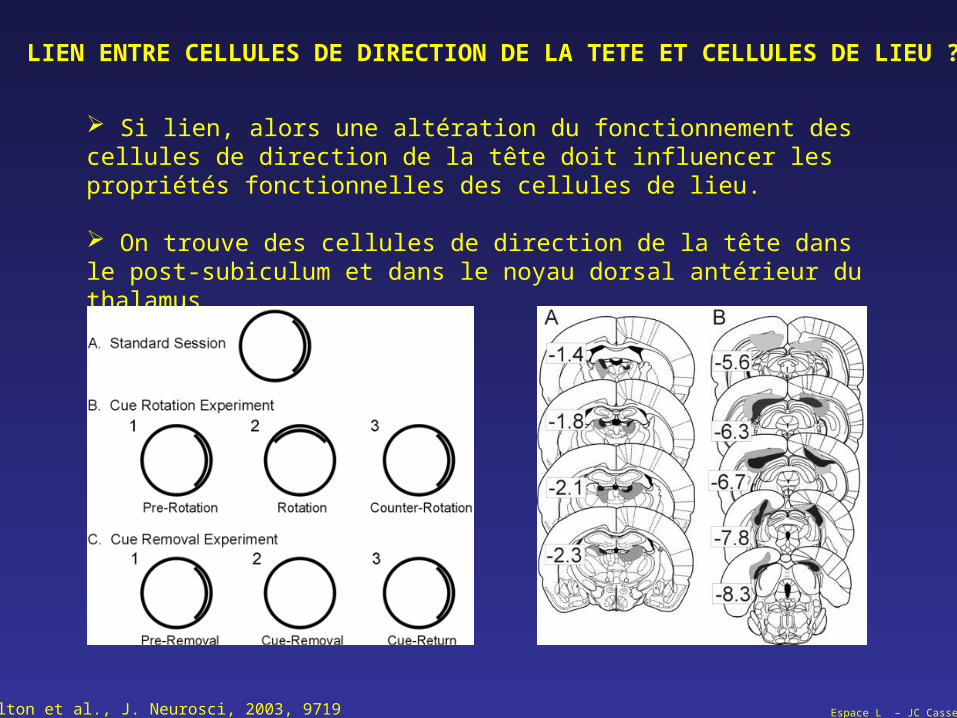
Espace L – JC Cassel
Si lien, alors une altération du fonctionnement des cellules de direction de la tête doit influencer les propriétés fonctionnelles des cellules de lieu.
On trouve des cellules de direction de la tête dans le post-subiculum et dans le noyau dorsal antérieur du thalamus
Calton et al., J. Neurosci, 2003, 9719
LIEN ENTRE CELLULES DE DIRECTION DE LA TETE ET CELLULES DE LIEU ?
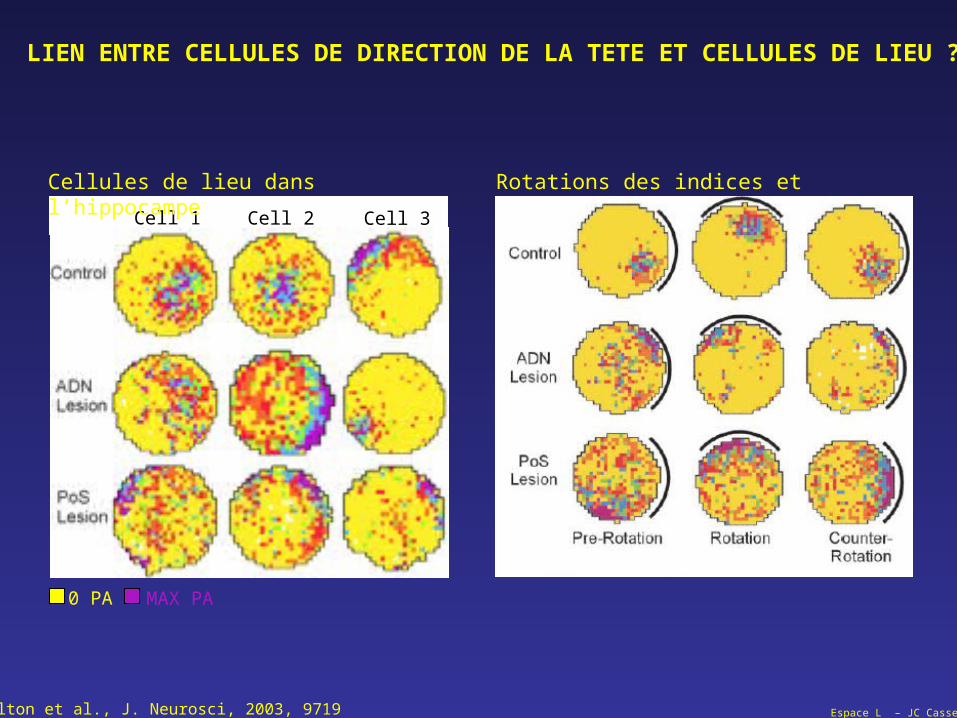
Espace L – JC Cassel
LIEN ENTRE CELLULES DE DIRECTION DE LA TETE ET CELLULES DE LIEU ?
Calton et al., J. Neurosci, 2003, 9719
0 PA MAX PA
Cell 1 Cell 2 Cell 3
Cellules de lieu dans l’hippocampe Rotations des indices et cellules de lieu
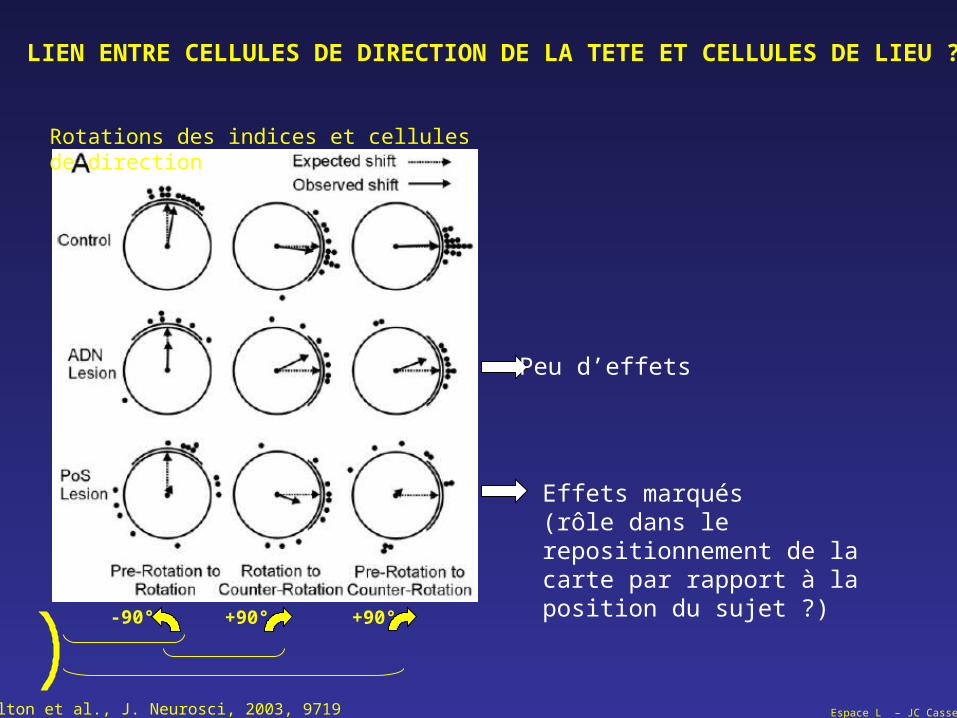
Espace L – JC CasselCalton et al., J. Neurosci, 2003, 9719
LIEN ENTRE CELLULES DE DIRECTION DE LA TETE ET CELLULES DE LIEU ?
Rotations des indices et cellules de direction
-90° +90° +90°
Peu d’effets
Effets marqués(rôle dans le repositionnement de lacarte par rapport à la position du sujet ?)
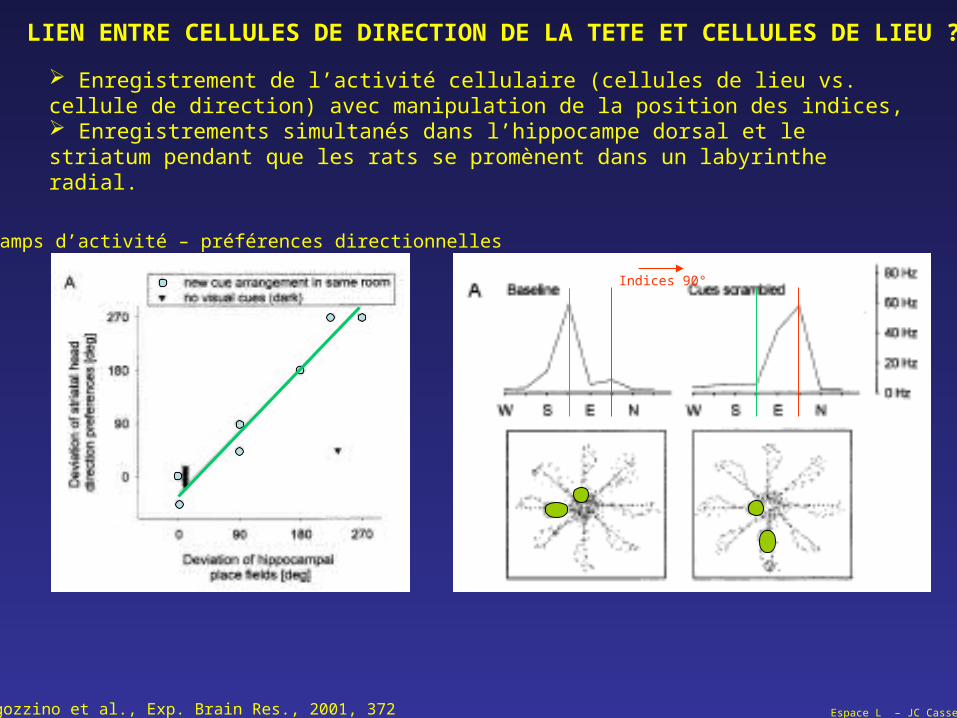
Espace L – JC Cassel
LIEN ENTRE CELLULES DE DIRECTION DE LA TETE ET CELLULES DE LIEU ?
Ragozzino et al., Exp. Brain Res., 2001, 372
Enregistrement de l’activité cellulaire (cellules de lieu vs. cellule de direction) avec manipulation de la position des indices, Enregistrements simultanés dans l’hippocampe dorsal et le striatum pendant que les rats se promènent dans un labyrinthe radial.
Champs d’activité – préférences directionnelles
Indices 90°

Espace L – JC Cassel
Et chez l’Homme ?
Espace L – JC Cassel
HIPPOCAMPE – MEMOIRE SPATIALE : ET CHEZ L’HOMME (1) ?air
e e
n m
m²
0
20
40
60
80
100
120
air
e e
n m
m²
0
20
40
60
80
100
120
témoinsconducteurs de taxi
ANT INT POST
0 350Expérience desconducteurs en mois
Ind
ice
de
vo
lum
eh
ipp
oc
am
piq
ue
(P
OS
T)
*
**
*
GAUCHE
DROITE
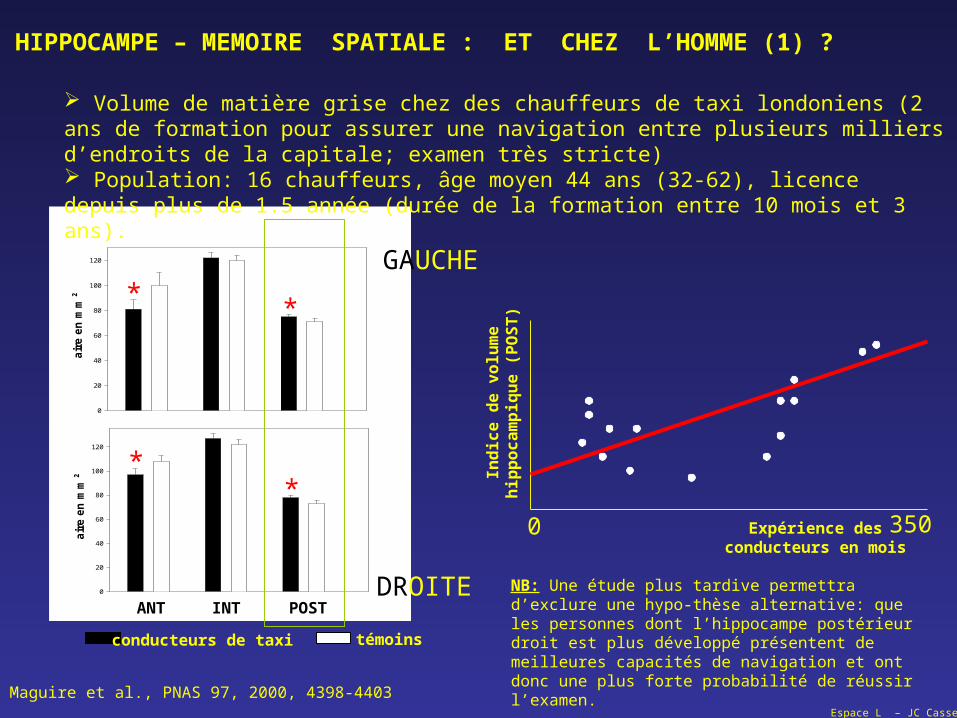
Maguire et al., PNAS 97, 2000, 4398-4403
Volume de matière grise chez des chauffeurs de taxi londoniens (2 ans de formation pour assurer une navigation entre plusieurs milliers d’endroits de la capitale; examen très stricte) Population: 16 chauffeurs, âge moyen 44 ans (32-62), licence depuis plus de 1.5 année (durée de la formation entre 10 mois et 3 ans).
NB: Une étude plus tardive permettra d’exclure une hypo-thèse alternative: que les personnes dont l’hippocampe postérieur droit est plus développé présentent de meilleures capacités de navigation et ont donc une plus forte probabilité de réussir l’examen.
Espace L – JC Cassel
HIPPOCAMPE – MEMOIRE SPATIALE : ET CHEZ L’HOMME (2) ?
Duke Nukem 3D
Correct, directCorrect, longErreur
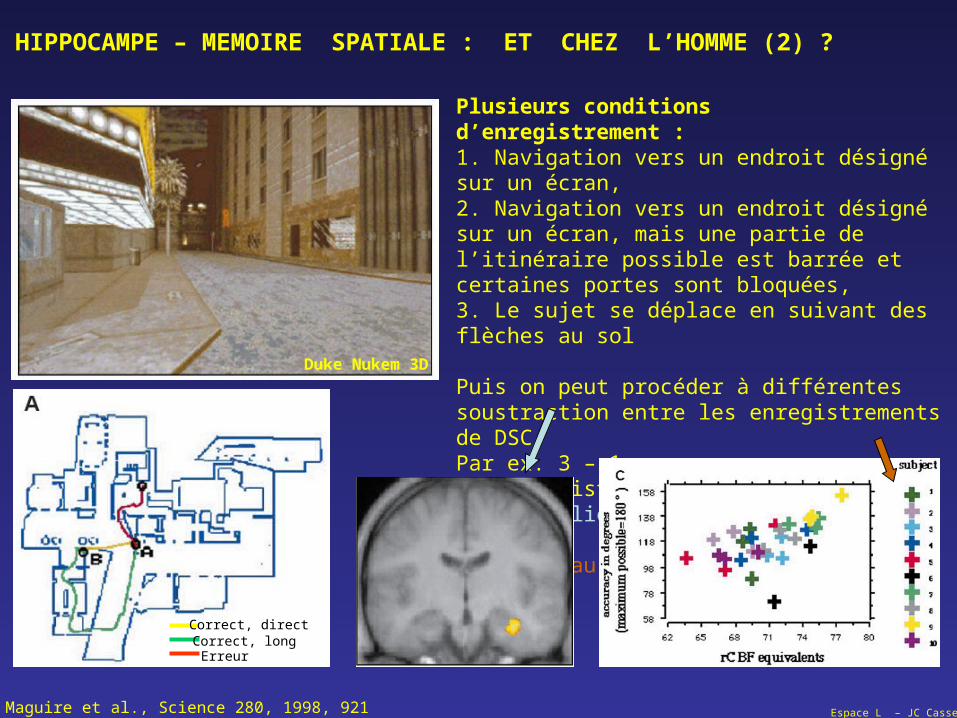
Plusieurs conditions d’enregistrement :1. Navigation vers un endroit désigné sur un écran,2. Navigation vers un endroit désigné sur un écran, mais une partie de l’itinéraire possible est barrée et certaines portes sont bloquées,3. Le sujet se déplace en suivant des flèches au sol
Puis on peut procéder à différentes soustraction entre les enregistrements de DSCPar ex. 3 – 1.On peut distinguer uneactivité liée à la navigation et relier cette dernière au
niveau de performance
Maguire et al., Science 280, 1998, 921
Espace L – JC Cassel
HIPPOCAMPE – MEMOIRE SPATIALE : ET CHEZ L’HOMME (3) ?
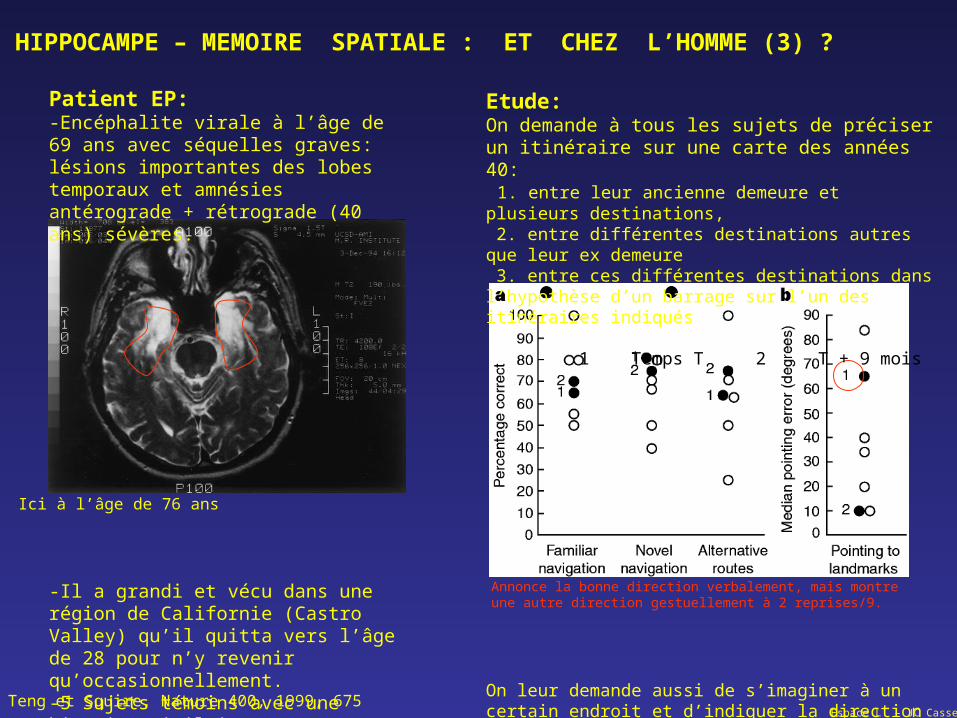
Patient EP:-Encéphalite virale à l’âge de 69 ans avec séquelles graves: lésions importantes des lobes temporaux et amnésies antérograde + rétrograde (40 ans) sévères.
-Il a grandi et vécu dans une région de Californie (Castro Valley) qu’il quitta vers l’âge de 28 pour n’y revenir qu’occasionnellement.-5 Sujets témoins avec une histoire similaire.
Teng et Squire, Nature 400, 1999, 675
Ici à l’âge de 76 ans
Annonce la bonne direction verbalement, mais montre une autre direction gestuellement à 2 reprises/9.
Etude:On demande à tous les sujets de préciser un itinéraire sur une carte des années 40: 1. entre leur ancienne demeure et plusieurs destinations, 2. entre différentes destinations autres que leur ex demeure 3. entre ces différentes destinations dans l’hypothèse d’un barrage sur l’un des itinéraires indiqués 1 Temps T 2 T + 9 mois
On leur demande aussi de s’imaginer à un certain endroit et d’indiquer la direction d’un autre endroit.

Espace L – JC Cassel
HIPPOCAMPE – MEMOIRE SPATIALE : ET CHEZ L’HOMME (4) ?
On dispose aussi d’enregistrements de l’activité des cellules au cours d’une tâche de navigation spatiale…- Enregistrements de l’activité cellulaire chez 7 patients épileptiques (on cherche le foyer)-Tâche: explorer un environnement virtuel (une cité) au volant d’un taxi et récupérer des clients- Enregistrement de 317 cellules (dont 67 dans hippocampe et 54 dans les régions parahippocampiques)
Dispositif Cellule à sélectivité de place
Ekstrom et al., Nature 425, 2003, 184
Espace L – JC Cassel
HIPPOCAMPE – MEMOIRE SPATIALE : ET CHEZ L’HOMME (5) ?
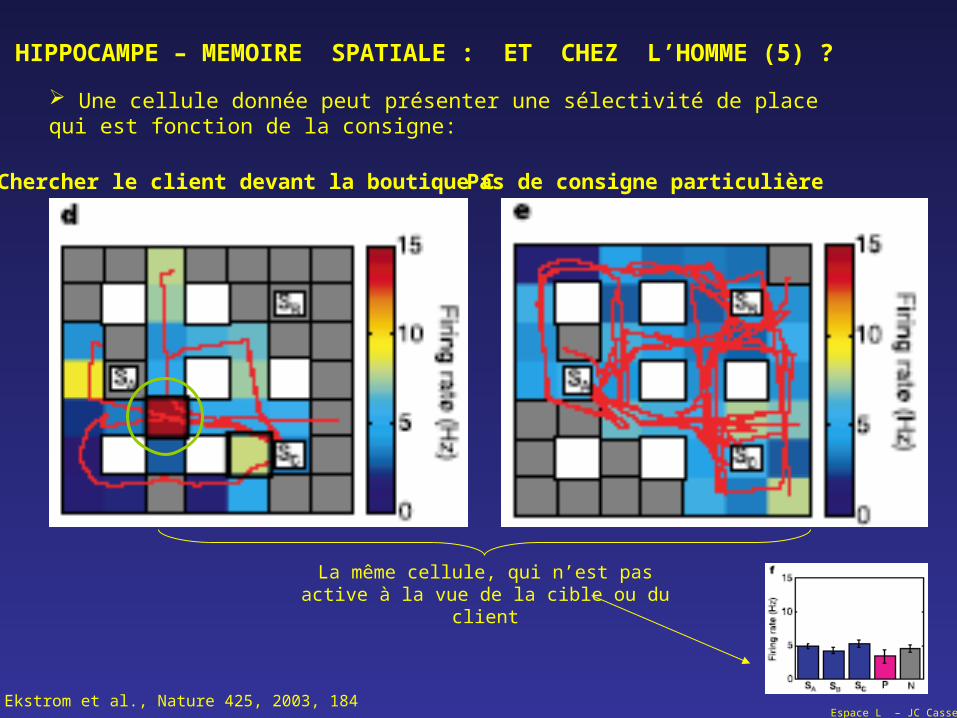
Une cellule donnée peut présenter une sélectivité de place qui est fonction de la consigne:
Chercher le client devant la boutique C Pas de consigne particulière
La même cellule, qui n’est pas active à la vue de la cible ou du client
Ekstrom et al., Nature 425, 2003, 184
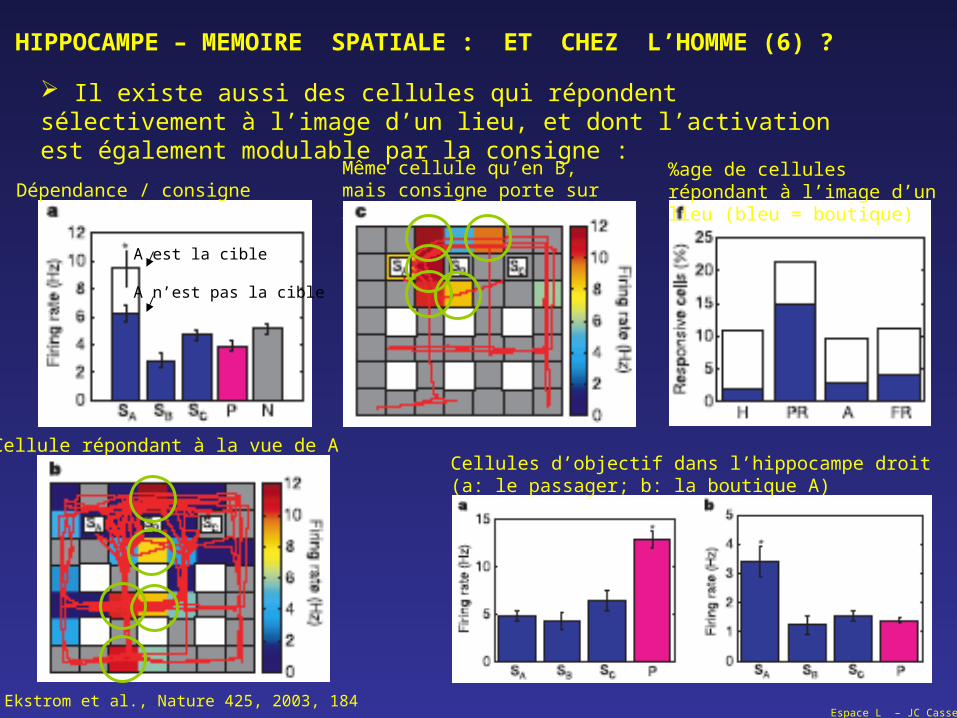
Espace L – JC Cassel
HIPPOCAMPE – MEMOIRE SPATIALE : ET CHEZ L’HOMME (6) ?
Il existe aussi des cellules qui répondent sélectivement à l’image d’un lieu, et dont l’activation est également modulable par la consigne :
A est la cible
A n’est pas la cible
Cellule répondant à la vue de A
Même cellule qu’en B, mais consigne porte sur ADépendance / consigne
%age de cellules répondant à l’image d’un lieu (bleu = boutique)
Cellules d’objectif dans l’hippocampe droit (a: le passager; b: la boutique A)
Ekstrom et al., Nature 425, 2003, 184

Espace L – JC Cassel
Résumé et conclusions générales
Espace L – JC Cassel
Le codage de l’information spatiale implique très fortement l’hippocampe (lésion, inactivation réversible, modèles transgéniques...) qui joue un rôle central (mais pas exclusif).
Ce codage repose sur un système complexe de neurones, principalement des régions CA1 et CA3, dont le niveau élémentaire d’organisation est la cellule de lieu.
Des cellules de lieu, ou des cellules aux propriétés comparables, ont été mises en évidence chez le Rat (première espèce), la Souris, le Singe et l’Homme (système commun à tous les vertébrés?).
Les cellules de lieu sont de cellules qui, après un certain temps, s’activent lorsque l’animal se trouve dans un endroit particulier de son environnement, quelle que soit la direction par laquelle il arrive à cet endroit (on appelle cet endroit le champ d’activité de la cellule de lieu).
Chaque cellule de lieu a un champ d’activité particulier qui restera stable des jours, des semaines et des mois, à la condition que l’environnement reste inchangé.
Lorsque cet environnement est modifié (et que l’animal n’assiste pas à la modification), le champ d’activité de la cellule de lieu est lui aussi modifié: on dit que la cellule procède à un recodage spatial (remapping).
CODAGE SPATIAL ET REPRESENTATION DE L’ESPACE (1)
Espace L – JC Cassel
CODAGE SPATIAL ET REPRESENTATION DE L’ESPACE (2)
Le champ d’activité d’une cellule de lieu donnée n’est pas piloté par un stimulus sensoriel particulier de l’environnement.
Les modalités sensorielles auxquelles les cellules de lieu sont sensibles sont multiples: quoique principalement visuelles, elles peuvent être olfactives, auditives, kinesthésiques… (allothétiques, idiothétiques).
En l’absence d’indices externes, qu’ils soient proximaux ou distaux, le codage de l’information spatiale peut aussi s’appuyer sur les déplacements de l’animal (plus grande marge d’erreurs, et même d’autant plus grande qu’il peut y avoir sommation de ces dernières).
Lorsqu’une partie de l’environnement est modifiée, il peut y avoir recodage de cette partie sans pour autant que la partie stable de l’environnement ne soit recodée.
Ces processus qui s’opèrent dans l’hippocampe sont des points de convergence de l’intégration d’une multiplicité d’informations impliquant une série d’aires corticales associatives (en amont).
La stabilité d’une représentation spatiale pourrait reposer sur des mécanismes de consolidation impliquant des processus cellulaires et moléculaires sous-tendant la plasticité synaptique (et donc les récepteurs NMDA et la synthèse protéique).
Le système des cellules de lieu opère de concert avec un système de codage directionnel indépendant de la position d’un animal (boussole/positionnement carte).
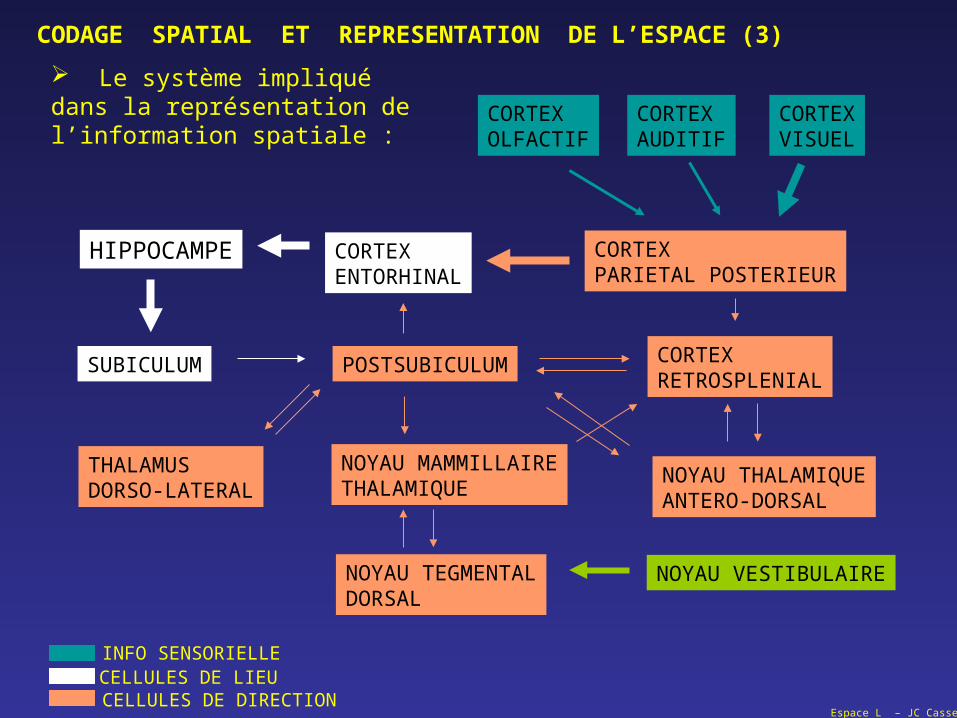
Espace L – JC Cassel
CODAGE SPATIAL ET REPRESENTATION DE L’ESPACE (3)
HIPPOCAMPE
SUBICULUM
CORTEXENTORHINAL
POSTSUBICULUM
NOYAU MAMMILLAIRETHALAMIQUE
NOYAU TEGMENTALDORSAL
THALAMUSDORSO-LATERAL
CORTEXPARIETAL POSTERIEUR
CORTEXRETROSPLENIAL
NOYAU THALAMIQUEANTERO-DORSAL
CORTEXOLFACTIF
CORTEXAUDITIF
CORTEXVISUEL
INFO SENSORIELLECELLULES DE LIEUCELLULES DE DIRECTION
NOYAU VESTIBULAIRE
Le système impliqué dans la représentation de l’information spatiale :