Embed Size (px)
Citation preview


2
L. Fortier
ArcticNet Annual Research Compendium (2011-12)
Arctic Cod
1.8 The Arctic cod (Boreogadus saida) ecosystem under the double pressure of climate change and industrialization (Arctic Cod)
Project LeaderLouis Fortier (Université Laval)
Project Team
Network InvestigatorsYves Gratton (Institut national de la recherche scientifi que - Eau, Terre et Environnement); Yvan Simard (Uni-versité du Québec à Rimouski); Jean-Eric Tremblay (Université Laval)
Collaborators and Research AssociatesShani Rousseau (ArcticNet Inc.); Dominique Robert (Québec-Océan); Alexandre Forest, Maxime Geoffroy, Makoto Sampei (Université Laval)
Postdoctoral FellowsKeita Suzuki (Université Laval)
PhD StudentsDelphine Benoit, Caroline Bouchard, Gerald Darnis, Jordan Grigor, Stephane Thanassekos (Université Laval)
MSc StudentsMarie-Claude Perreault (Université Laval)
Undergraduate StudentsSolenn Mordret (Université Laval)
Technical and Project Staff Hélen Cloutier, Louis Letourneau (Université Laval)

3
L. Fortier
ArcticNet Annual Research Compendium (2011-12)
Arctic Cod
ABSTRACT
The Arctic cod (Boreogadus saida), also known as the polar cod in Europe, is a key component of the Arctic Ocean pelagic ecosystem that effects up to 75% of the energy transfer between the plankton and the vertebrate fauna (fi sh, seals, whales and marine birds). Being an hyper-specialist adapted to life in ice-covered seas, Arc-tic cod is likely to be displaced by southern generalists such as the capelin and the sandlance as the ice regime becomes less severe. This project collaborates closely with “Hotspots”, “Moorings” and “Sea-ice” to map the distribution and migrations of Arctic cod populations in the Canadian Arctic, and to measure variations in hatching season and early growth in relation to annual changes in ice regime, surface temperature, and zoo-plankton prey abundance. In partnership with the Oil Exploration sector and the Department of Aboriginal Affairs and Northern Development (Beaufort Regional Environmental Assessment program), we assess the general distribution and reproduction of Arctic cod in the Beaufort Sea and the potential environmental risks of exploratory drilling on its ecology.
KEY MESSAGES
1. Seasonally migrating zooplankton assimilate car-bon in the surface layer and respire it at depth. A fi rst ever measurement of this vertical respiratory fl ux over an annual cycle shows that this process contributes signifi cantly to the sequestration of atmospheric carbon dioxide by the Arctic Ocean;
• Since 2002, reduced ice cover and increased up-welling in the Beaufort Sea have boosted the pop-ulation levels of the copepod Calanus glacialis, a major prey of the juvenile and adult stages of Arctic cod;
• The hatching season of Arctic cod extends from January to July in Arctic seas infl uenced by large rivers (Beaufort, Laptev, Hudson Bay) and is lim-ited to May-July in other regions;
• An Individual-Based Model (IBM) of Arctic cod bioenergetics successfully reproduced the ob-served growth and survival (0-45 d) of juveniles in the Northeast Water in 1993 and in the North Water in 1998, opening the way to modelling the response of Arctic cod to a decline in ice cover and the warming of the surface layer;
• Starting in 2010 and again in 2011, signifi cant numbers of Pacifi c sand lance (Ammodytes pa-cifi cus) juveniles were detected for the fi rst time in the offshore distribution area of Arctic cod juveniles, indicating that sand lance could have started displacing Arctic cod as the sea-ice regime becomes less severe;
• In the Amundsen Gulf, spawning Arctic cod start aggregating at depth precisely when the ice cover consolidate in early December, and disperse with the ice breakup in April. Aggregations move to deeper areas as the photoperiod lengthens.
• Contrary to expectations, Arctic cod does not seems to form dense schools in the surface layer of the offshore Beaufort Sea in late summer;
• The acoustic signatures (target strengths) of dif-ferent marine mammals in a 1.5 km radius around the Amundsen have been measured in August-September 2011 with a SX90 fi sheries sonar. These will help identify, monitor, and avoid ma-rine mammals during scientifi c and seismic op-erations.
OBJECTIVES
• Describe decadal variations (2002-2016) in the late-summer absolute and relative abundance of the main arctic copepods and their predators (Arc-tic cod, chaetognaths, and the hyperiid amphipod Themisto libellulla) in the Beaufort Sea;
• Track with unprecedented vertical resolution the seasonal migration at depth of the large calanoid copepods Calanus glacialis and C. hyperboreus and their lipid reserves using the new LOKI in-situ image profi ler;

4
L. Fortier
ArcticNet Annual Research Compendium (2011-12)
Arctic Cod
• Quantify with greater accuracy the role of cope-pod respiration and migration in the export of bio-genic carbon from the surface Arctic layer to the deep Atlantic layer;
• Assess regional and interannual (2002-2016) var-iations in the prey fi eld and food intake of larval and juvenile Arctic cod in the Beaufort Sea and North Water;
• Correlate decadal variations (2002-2016) in the hatch date frequency distribution of Arctic cod to variations in sea ice cover and surface tempera-ture in the Beaufort Sea;
• Apply our IBM of Arctic cod early growth to model and explain seasonal and interannual dif-ferences in the growth trajectory of juveniles captured in late summer in the Beaufort Sea from 2002 to 2012;
• Determine the summer distribution of adult and juvenile Arctic cod and their marine mammal predators in southeastern Beaufort Sea;
• Detect regional variations in the population ge-netics of Arctic cod using microsatellite markers;
• Document the vertical and horizontal distribution of Arctic cod and zooplankton under the ice cover of the Canadian Arctic Ocean using sonars carried by an Autonomous Underwater Vehicle (AUV).
INTRODUCTION
The pelagic ecosystem of the Arctic Ocean provides local communities with many goods and services that include waterways in summer and ice-ways in win-ter, traditional food (fi sh, marine mammals and birds), “traditional” health (omega-3, selenium, etc.), furs and leather, heating oil, substrates for sculpture (e.g. bones and ivory), inspiration for the arts, social cohesion, intergenerational bonding, spiritual comfort, and life fulfi lling. As the sea-ice cover of Arctic seas declines and their surface layer warms up, signs of the expected replacement of the unique Arctic pelagic ecosystem by North-Atlantic and North-Pacifi c ecosystem types are increasingly detected (e.g. Tynan and DeMaster 1997;
Gaston et al. 2003, Grebmeier et al. 2006). The atlan-tifi cation/pacifi cation of the Arctic pelagic ecosystem threatens the services rendered to northern communi-ties, and harbingers a shift to different ecosystem ser-vices that will benefi t primarily southern industries: for instance, access to oil and other mineral resources, new shipping lanes, fi sh stocks of commercial interest, and ecotourism.
The objective of ArcticNet Integrated Regional Impact Studies (IRISes) is to provide Inuit communities, the private sector, and governments with the relevant sci-entifi c information to formulate the policies, adaptation strategies, and decisions that will shape the response of Canada to climate change and modernization in the Arc-tic. As far as marine ecosystem services are concerned, anticipating the rate and timing of the expected trans-formation of the pelagic Arctic ecosystem is a crucial element for all four IRISes. The Arctic cod (Boreoga-dus saida) is a key component of the relatively simple Arctic Ocean pelagic ecosystem. This small forage fi sh channels up to 75% of the energy transfer between the plankton and the vertebrates such as fi sh, seals, whales and marine birds that supply many ecosystem services (Welch et al. 1992). A hyper-specialist adapted to life in ice-covered seas, Arctic cod is likely to be rapidly displaced by southern generalists as the ice regime be-comes less severe. In northern Hudson Bay where ice decline is intense, capelin (Mallotus villosus) and sand lance (Ammodytes spp.) have replaced Arctic cod as the main prey brought back to the nest to feed young thick-billed murres (Gaston et al. 2003). Similarly, in the Beaufort Sea where ice retreat is also severe, our own studies of the ichthyoplankton assemblage indicate the recent intrusion of the Pacifi c sand lance in the offshore distribution area of juvenile Arctic cod. We believe that changes in the ecology of Arctic cod, including its dis-placement by boreal forage fi shes, are among the most powerful indicators of the transition of the Arctic Ocean pelagic ecosystem to a new equilibrium.
The Arctic Cod project collaborates closely with the “Hotspots”, “Moorings” and “Sea-ice” projects to map the distribution of Arctic cod in the Canadian Arctic, to measure variations in its hatching season and early growth in relation to annual changes in ice regime, sur-

5
L. Fortier
ArcticNet Annual Research Compendium (2011-12)
Arctic Cod
face temperature, and zooplankton prey abundance, and to monitor changes in the ichthyoplankton assemblage to which it belongs. We also focus on numerical mod-els of the early growth and survival of the juveniles. In partnership with the Oil Exploration sector and the Department of Aboriginal Affairs and Northern Devel-opment (Beaufort Regional Environmental Assessment program), we contrast the abundance and reproduction of Arctic cod among different regions of the Beaufort Sea, including the edge of the continental shelf where oil exploration rights have been awarded, so as to assess the potential risks of exploration drilling on the Arctic cod ecosystem. Starting in 2011, our team has initiated collaborations with the Nunavut Department of Envi-ronment, Fisheries and Sealing, to coordinate opera-tions between the Amundsen and the research trawler Nuliajuk with the objective of documenting the inva-sion of Cumberland Sound by capelin.
ACTIVITIES
Time frame and study area. In 2011-2012, the team pursued the monitoring of zooplankton, larval fi sh, and adult Arctic cod based on the annual ArcticNet mission to the Canadian Arctic on board the CCGS Amundsen. Since 2002, we have collected zooplankton samples annually and measured the taxonomic composition of the zooplankton assemblage, species abundance, and developmental stages frequency of the main copepod species at the end of the summer in different regions of the Canadian Arctic with emphasis on the Beaufort Sea. The abundance and vertical distribution of juve-nile and adult fi sh (mainly Arctic cod) was assessed for the 15 m to bottom layer along the track of the ship using the EK60 fi sh fi nder. In 2011-2012, much effort was invested in the successful installation and deploy-ment of the SX90 fi sheries sonar for the detection of fi sh schools and marine mammals in a 1-2 km radius around the ship, including the insonifi cation of the sur-face layer (0-15 m) which until now we could not study.
Research activities in 2011-2012
• Installation on the Amundsen and preliminary test at sea of the new SX90 fi sheries sonar;
• Extensive test of the SX90 in the Gulf of St. Law-rence, the Labrador Sea, Baffi n Bay and Lancaster Sound with the successful detection of fi sh school and the acquisition of target strength for marine mammals including ringed seals, minke whales and right whales;
• Joint survey of fi sh distribution in Cumberland Sound combining SX90 and EK60 acoustic sur-veys by the Amundsen with experimental trawl-ing by the new Nunavut research trawler Nulia-juk;
• Sampling of zooplankton and ichthyoplankton at all basic stations along the ship track from Baffi n Bay to the Beaufort Sea and return via the North Water;
• Extensive acoustic surveys (EK60 and SX90) of fi sh and marine mammal distributions in the Beaufort Sea as part of the Beaufort Region Envi-ronmental Assessment (BREA);
• The completion and publication of our Indi-vidual Based Model of juvenile (0-45 d) Arctic cod growth and survival for the North Water and Northeast Water;
• First ever assessment of the fl ux of carbon linked to the annual cycle of vertical migration and respiration of zooplankton in the Arctic Ocean (Beaufort Sea);
• Analysis of interannual variations (2002-2009) in the abundance of copepod species in relation to sea-ice cover and upwelling in the Beaufort Sea;
• First acoustic description and publication of the offshore winter distribution and aggrega-tion behaviour of Arctic cod in the Arctic Ocean (Amundsen Gulf);
• Successful completion of major research partner-ships with IOL and BP on the ecological impor-tance of the offshore exploration claims relative to the overall Beaufort Sea;
• Successful completion of two doctoral theses, respectively on the IBM modeling of Arctic cod early bioenergetics (Thanassekos) and the acous-tic study of Arctic cod distribution (Benoit);
6
L. Fortier
ArcticNet Annual Research Compendium (2011-12)
Arctic Cod
• Analysis of zooplankton collections for the Nu-natsiavut Fjords project;
RESULTS
In the relatively simple ecosystem of the Arctic Ocean, copepods and the Arctic cod channel most of the car-bon and energy from ice algae and phytoplankton (the primary producers) to the vertebrate fauna (fi sh, marine mammals, marine birds) that provide services to com-munities. The magnitude and effi ciency of this trophic fl ux of carbon is strongly infl uenced by physical condi-tions such as sea-ice regime, temperature, wind mixing, and the upwelling of nutrients. The “Moorings” and “Hotspots” projects examine how ocean climate control primary production and the vertical fl ux of carbon (the fraction of primary production that ends up sequestered at depth). In close collaboration with these projects, the Arctic Cod project focuses in addition on the trophic fl ux of carbon that underpins ecosystem services.
Zooplankton respiration and the transfer at depth of atmospheric carbon (Darnis et al. in press)
In Arctic seas, lipids accumulated by zooplankton mi-grants in the surface layer in spring-summer are respired at depth during the winter. The resulting active down-ward transport of carbon by the 200-1000 and >1000 μm mesozooplankton fractions was quantifi ed based on
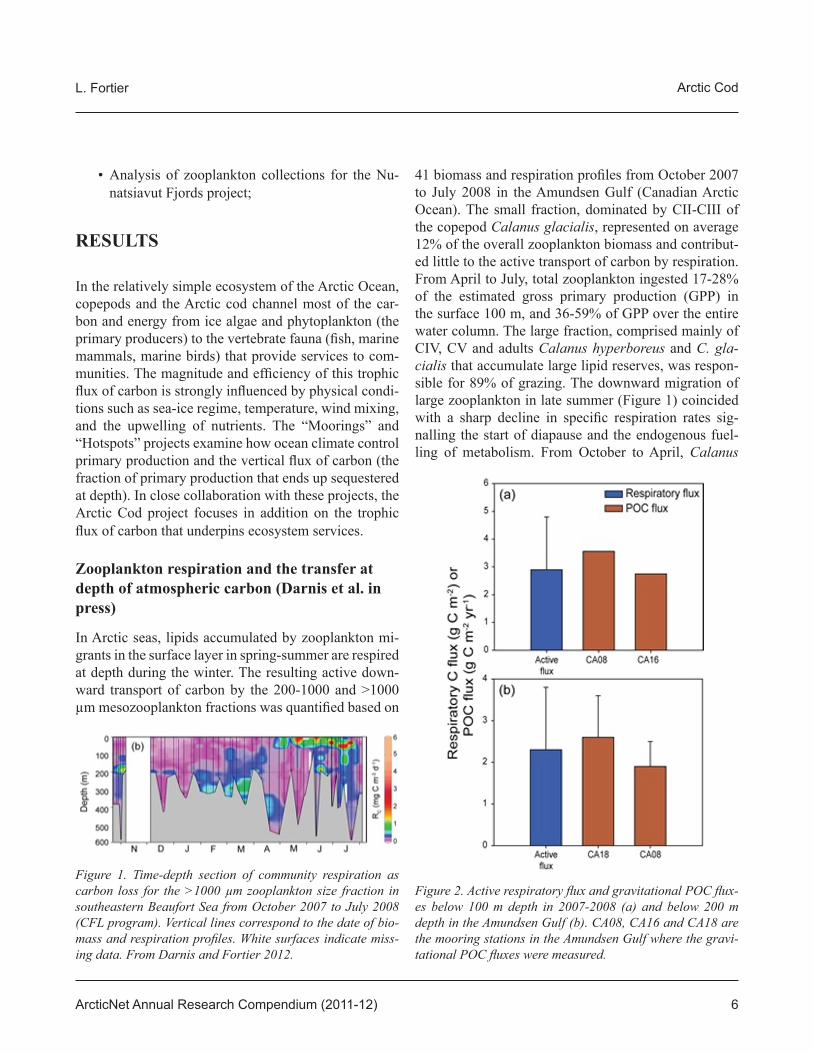
41 biomass and respiration profi les from October 2007 to July 2008 in the Amundsen Gulf (Canadian Arctic Ocean). The small fraction, dominated by CII-CIII of the copepod Calanus glacialis, represented on average 12% of the overall zooplankton biomass and contribut-ed little to the active transport of carbon by respiration. From April to July, total zooplankton ingested 17-28% of the estimated gross primary production (GPP) in the surface 100 m, and 36-59% of GPP over the entire water column. The large fraction, comprised mainly of CIV, CV and adults Calanus hyperboreus and C. gla-cialis that accumulate large lipid reserves, was respon-sible for 89% of grazing. The downward migration of large zooplankton in late summer (Figure 1) coincided with a sharp decline in specifi c respiration rates sig-nalling the start of diapause and the endogenous fuel-ling of metabolism. From October to April, Calanus
Figure 1. Time-depth section of community respiration as carbon loss for the >1000 μm zooplankton size fraction in southeastern Beaufort Sea from October 2007 to July 2008 (CFL program). Vertical lines correspond to the date of bio-mass and respiration profi les. White surfaces indicate miss-ing data. From Darnis and Fortier 2012.
Figure 2. Active respiratory fl ux and gravitational POC fl ux-es below 100 m depth in 2007-2008 (a) and below 200 m depth in the Amundsen Gulf (b). CA08, CA16 and CA18 are the mooring stations in the Amundsen Gulf where the gravi-tational POC fl uxes were measured.
7
L. Fortier
ArcticNet Annual Research Compendium (2011-12)
Arctic Cod
migration-respiration actively transported 3.1 g C m-2 beyond 100 m, a fl ux that represented 85 to 132% of the gravitational POC fl uxes at 100 m from October to July (Figure 2). Our results stress the importance of includ-ing active transport by large zooplankton migrants in carbon budgets of the Arctic Ocean.
Sea-ice decline, upwelling and copepod abundance in the Beaufort Sea (Suzuki et al. ms)
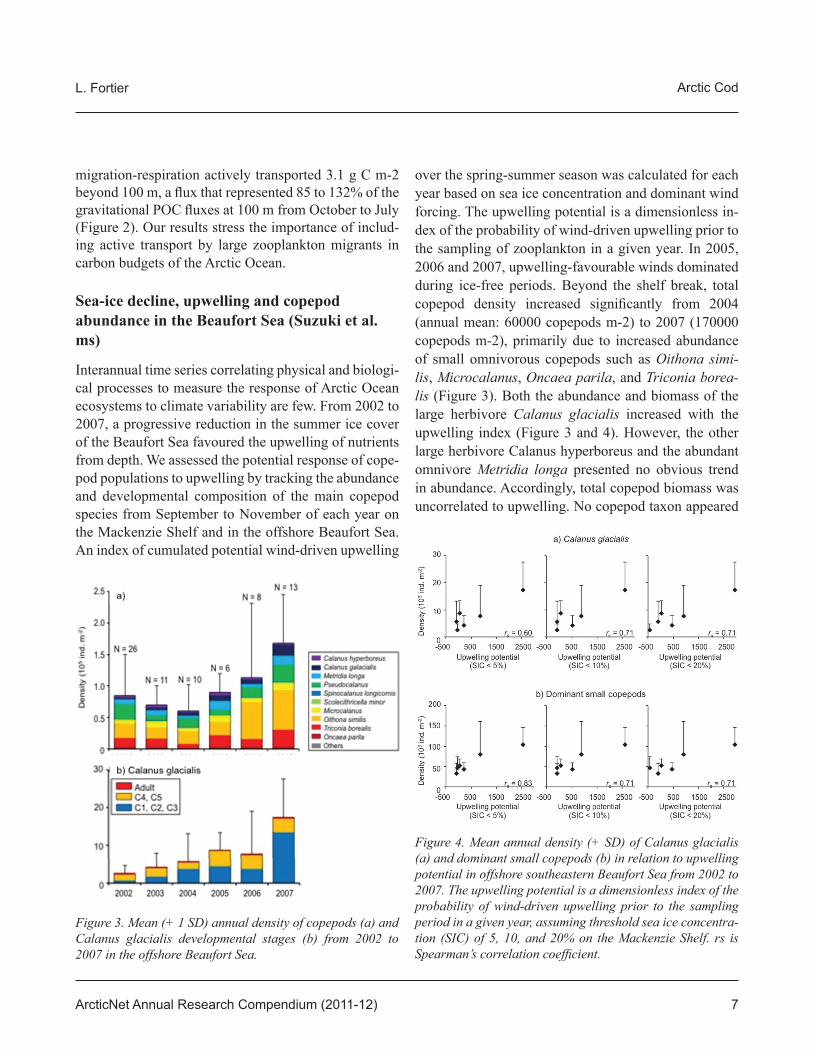
Interannual time series correlating physical and biologi-cal processes to measure the response of Arctic Ocean ecosystems to climate variability are few. From 2002 to 2007, a progressive reduction in the summer ice cover of the Beaufort Sea favoured the upwelling of nutrients from depth. We assessed the potential response of cope-pod populations to upwelling by tracking the abundance and developmental composition of the main copepod species from September to November of each year on the Mackenzie Shelf and in the offshore Beaufort Sea. An index of cumulated potential wind-driven upwelling
over the spring-summer season was calculated for each year based on sea ice concentration and dominant wind forcing. The upwelling potential is a dimensionless in-dex of the probability of wind-driven upwelling prior to the sampling of zooplankton in a given year. In 2005, 2006 and 2007, upwelling-favourable winds dominated during ice-free periods. Beyond the shelf break, total copepod density increased signifi cantly from 2004 (annual mean: 60000 copepods m-2) to 2007 (170000 copepods m-2), primarily due to increased abundance of small omnivorous copepods such as Oithona simi-lis, Microcalanus, Oncaea parila, and Triconia borea-lis (Figure 3). Both the abundance and biomass of the large herbivore Calanus glacialis increased with the upwelling index (Figure 3 and 4). However, the other large herbivore Calanus hyperboreus and the abundant omnivore Metridia longa presented no obvious trend in abundance. Accordingly, total copepod biomass was uncorrelated to upwelling. No copepod taxon appeared
Figure 3. Mean (+ 1 SD) annual density of copepods (a) and Calanus glacialis developmental stages (b) from 2002 to 2007 in the offshore Beaufort Sea.
Figure 4. Mean annual density (+ SD) of Calanus glacialis (a) and dominant small copepods (b) in relation to upwelling potential in offshore southeastern Beaufort Sea from 2002 to 2007. The upwelling potential is a dimensionless index of the probability of wind-driven upwelling prior to the sampling period in a given year, assuming threshold sea ice concentra-tion (SIC) of 5, 10, and 20% on the Mackenzie Shelf. rs is Spearman’s correlation coeffi cient.
8
L. Fortier
ArcticNet Annual Research Compendium (2011-12)
Arctic Cod
negatively impacted by increased upwelling. Our obser-vations may represent the fi rst signs of the general in-crease in secondary production predicted to result from a reduction of ice cover, a warming of the surface layer, and an intensifi cation of wind-induced upwelling in Arctic seas. Continued annual monitoring of zooplank-ton dynamics is crucial to anticipate the future services provided by pelagic ecosystems in the Arctic Ocean.
A pan-Arctic synthesis of the hatching season of Arctic cod (Bouchard and Fortier 2011)
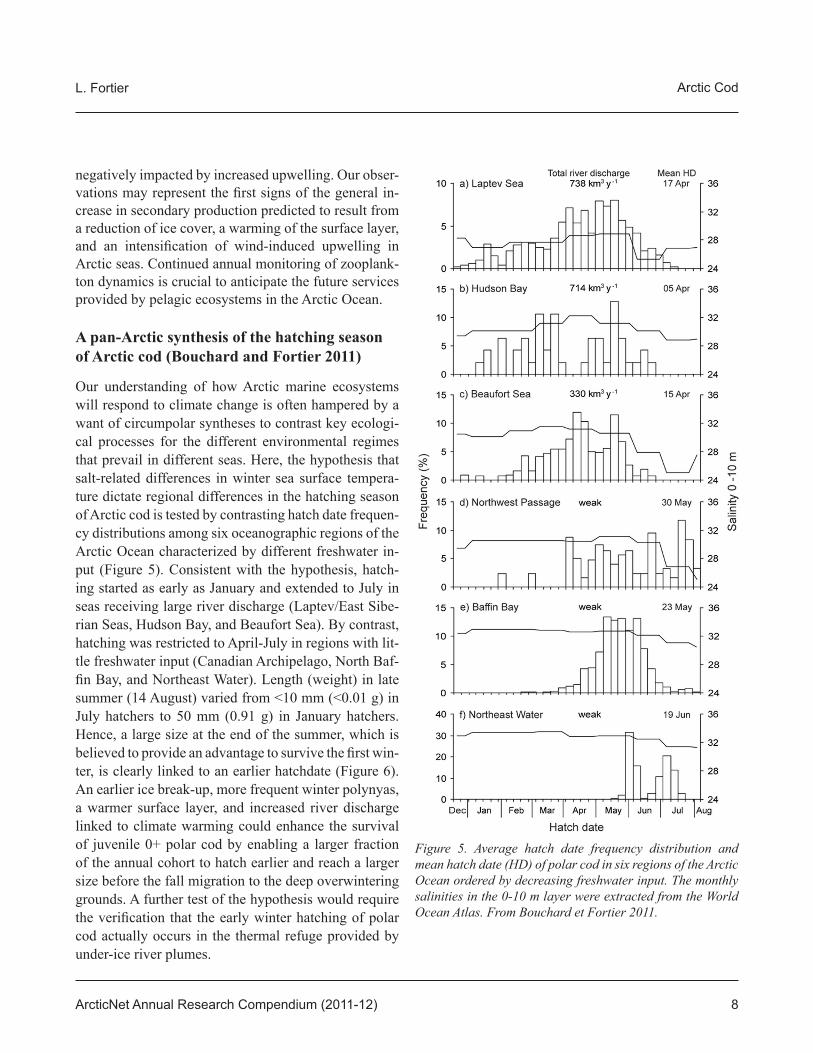
Our understanding of how Arctic marine ecosystems will respond to climate change is often hampered by a want of circumpolar syntheses to contrast key ecologi-cal processes for the different environmental regimes that prevail in different seas. Here, the hypothesis that salt-related differences in winter sea surface tempera-ture dictate regional differences in the hatching season of Arctic cod is tested by contrasting hatch date frequen-cy distributions among six oceanographic regions of the Arctic Ocean characterized by different freshwater in-put (Figure 5). Consistent with the hypothesis, hatch-ing started as early as January and extended to July in seas receiving large river discharge (Laptev/East Sibe-rian Seas, Hudson Bay, and Beaufort Sea). By contrast, hatching was restricted to April-July in regions with lit-tle freshwater input (Canadian Archipelago, North Baf-fi n Bay, and Northeast Water). Length (weight) in late summer (14 August) varied from <10 mm (<0.01 g) in July hatchers to 50 mm (0.91 g) in January hatchers. Hence, a large size at the end of the summer, which is believed to provide an advantage to survive the fi rst win-ter, is clearly linked to an earlier hatchdate (Figure 6). An earlier ice break-up, more frequent winter polynyas, a warmer surface layer, and increased river discharge linked to climate warming could enhance the survival of juvenile 0+ polar cod by enabling a larger fraction of the annual cohort to hatch earlier and reach a larger size before the fall migration to the deep overwintering grounds. A further test of the hypothesis would require the verifi cation that the early winter hatching of polar cod actually occurs in the thermal refuge provided by under-ice river plumes.
Figure 5. Average hatch date frequency distribution and mean hatch date (HD) of polar cod in six regions of the Arctic Ocean ordered by decreasing freshwater input. The monthly salinities in the 0-10 m layer were extracted from the World Ocean Atlas. From Bouchard et Fortier 2011.
9
L. Fortier
ArcticNet Annual Research Compendium (2011-12)
Arctic Cod
Modelling the early (0-45 d) growth and survival of Arctic cod larvae in the North Water and Northeast Water Polynyas (Thanassekos and Fortier 2012; Thanassekos et al. 2012).
Actual observations from a ship invariably provide a very limited picture of the population dynamics of planktonic organisms, a fortiori in ice-covered seas where sampling coverage is limited both in time and in space. Hence, studying and predicting the response of key components of the Arctic plankton community to climate variability and change will increasingly rely on numerical simulations. In such computer models, a virtual population of organisms is released in a virtual representation of its environment. The goal is to gen-erate a complete picture of its dynamics, over a given time interval, that reproduces and completes the partial picture provided by sampling at sea.
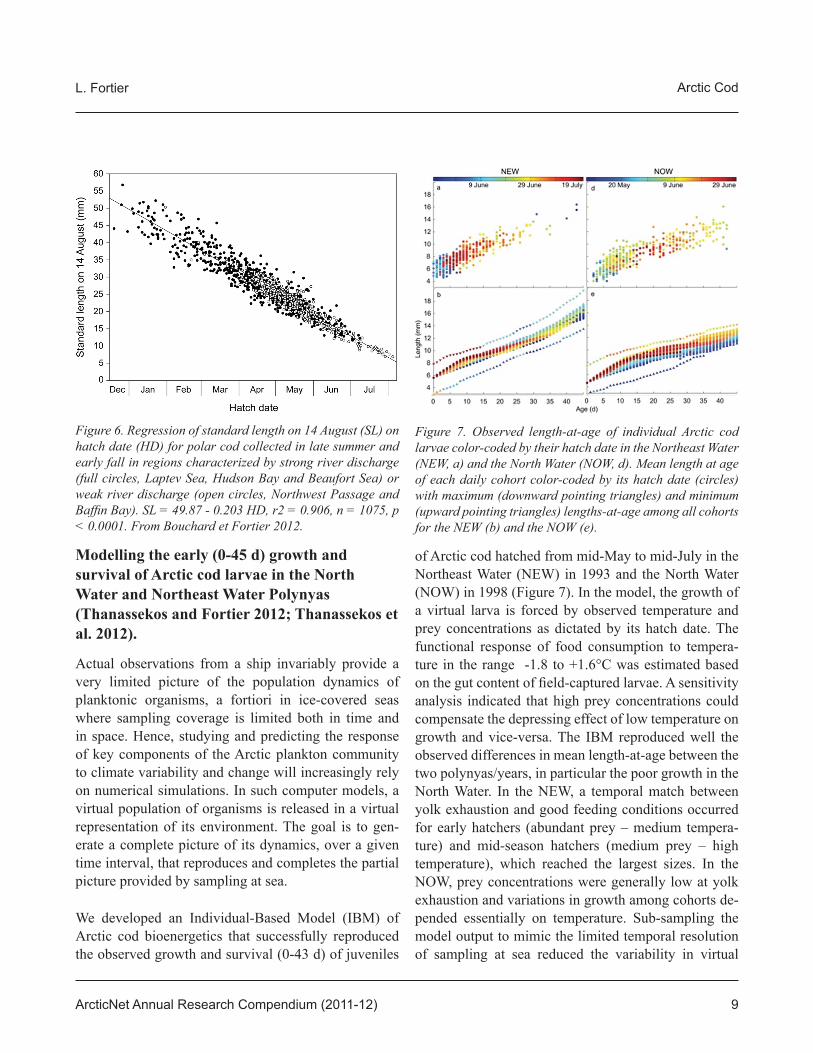
We developed an Individual-Based Model (IBM) of Arctic cod bioenergetics that successfully reproduced the observed growth and survival (0-43 d) of juveniles
of Arctic cod hatched from mid-May to mid-July in the Northeast Water (NEW) in 1993 and the North Water (NOW) in 1998 (Figure 7). In the model, the growth of a virtual larva is forced by observed temperature and prey concentrations as dictated by its hatch date. The functional response of food consumption to tempera-ture in the range -1.8 to +1.6°C was estimated based on the gut content of fi eld-captured larvae. A sensitivity analysis indicated that high prey concentrations could compensate the depressing effect of low temperature on growth and vice-versa. The IBM reproduced well the observed differences in mean length-at-age between the two polynyas/years, in particular the poor growth in the North Water. In the NEW, a temporal match between yolk exhaustion and good feeding conditions occurred for early hatchers (abundant prey – medium tempera-ture) and mid-season hatchers (medium prey – high temperature), which reached the largest sizes. In the NOW, prey concentrations were generally low at yolk exhaustion and variations in growth among cohorts de-pended essentially on temperature. Sub-sampling the model output to mimic the limited temporal resolution of sampling at sea reduced the variability in virtual
Figure 6. Regression of standard length on 14 August (SL) on hatch date (HD) for polar cod collected in late summer and early fall in regions characterized by strong river discharge (full circles, Laptev Sea, Hudson Bay and Beaufort Sea) or weak river discharge (open circles, Northwest Passage and Baffi n Bay). SL = 49.87 - 0.203 HD, r2 = 0.906, n = 1075, p < 0.0001. From Bouchard et Fortier 2012.
Figure 7. Observed length-at-age of individual Arctic cod larvae color-coded by their hatch date in the Northeast Water (NEW, a) and the North Water (NOW, d). Mean length at age of each daily cohort color-coded by its hatch date (circles) with maximum (downward pointing triangles) and minimum (upward pointing triangles) lengths-at-age among all cohorts for the NEW (b) and the NOW (e).

10
L. Fortier
ArcticNet Annual Research Compendium (2011-12)
Arctic Cod
growth and increased the match between simulated and observed variances in length-at-age. The IBM neverthe-less underestimated the observed exceptional growth during match events.
This fi rst effort opens the way to modelling the response of Arctic cod to a decline in ice cover and the warming of the surface layer.
Are we witnessing the fi rst signs of a displacement of Arctic cod by sand lance in the offshore Beaufort Sea?
Fish respond rapidly to changes in the geographical po-sition of isotherms in the Ocean. A general prediction of fi sheries ecology is that, with climate warming, the distribution of fi sh populations in the Northern Hemi-sphere will shift northward. Southern species of fi sh and their copepod prey have been observed moving to temperate areas, and temperate species have been seen expanding north in response to a shift in ocean climate (Beaugrand et al. 2002, Perry et al. 2005). Indications are that boreal species from the North Pacifi c and the North Atlantic are invading the Arctic Basin (Gaston et al. 2003, Babaluk et al. 2000). In particular, capelin (Mallotus villosus) and sand lance (Ammodytes spp.) are two small forage fi shes that could soon displace the Arctic cod on the continental shelves of the Arctic Ocean (Hunt and Megrey 2005, Barber et al. 2008, Ste-phenson and Hartwig 2010). Already, capelin and sand lance are increasingly replacing Arctic cod in the diet of thick-bill murres as the ice regresses in northern Hud-son Bay (Gaston et al. 2003). A replacement of Arctic cod by capelin and/or sand lance would harbinger a pro-found shift in the structure of the pelagic ecosystem and in the ecosystem services provided by the Arctic Ocean.
Starting in 2010, signifi cant numbers of Pacifi c sand lance (Ammodytes hexapterus) juveniles were detect-ed for the fi rst time in the offshore distribution area of Arctic cod juveniles. For example, in the exploration claim for oil at the edge of the continental shelf, sand lance juveniles were captured at none of 21 stations in 2009, 2 of 18 stations in 2010, and 9 of 13 stations in 2011 (Figure 8). Since the presence of juveniles neces-
sarily implies successful reproduction in the area, this progression suggests that sand lance could have started displacing Arctic cod as the sea-ice regime becomes less severe and the summer surface layer warms up in the Beaufort Sea. In addition to continued sampling in the coming years, the next step in verifying this hypoth-esis is to check for the presence of sand lance larvae and juveniles in our own ichthyoplankton collections in the area since 2002 and in inventory of fi sh larvae by other programs predating 2002.
Sea ice cover and the winter aggregation of Arctic cod in the offshore Amundsen Gulf (Beaufort Sea) (Geoffroy et al. 2011)
During the Circumpolar Flaw Lead System Study (CFL, 2007–2008), aggregations of polar cod were detected in winter in the Amundsen Gulf (Western Canadian Arctic) using the EK60 echosounder of the CCGS Amundsen
Figure 8. The distribution of Arctic cod (top panel) and Pa-cifi c sand lance (bottom panel) larvae and juveniles in the oil exploration claims of BP and IOL in 2009 (red circles), 2010 (orange) and 2011 (yellow).
11
L. Fortier
ArcticNet Annual Research Compendium (2011-12)
Arctic Cod
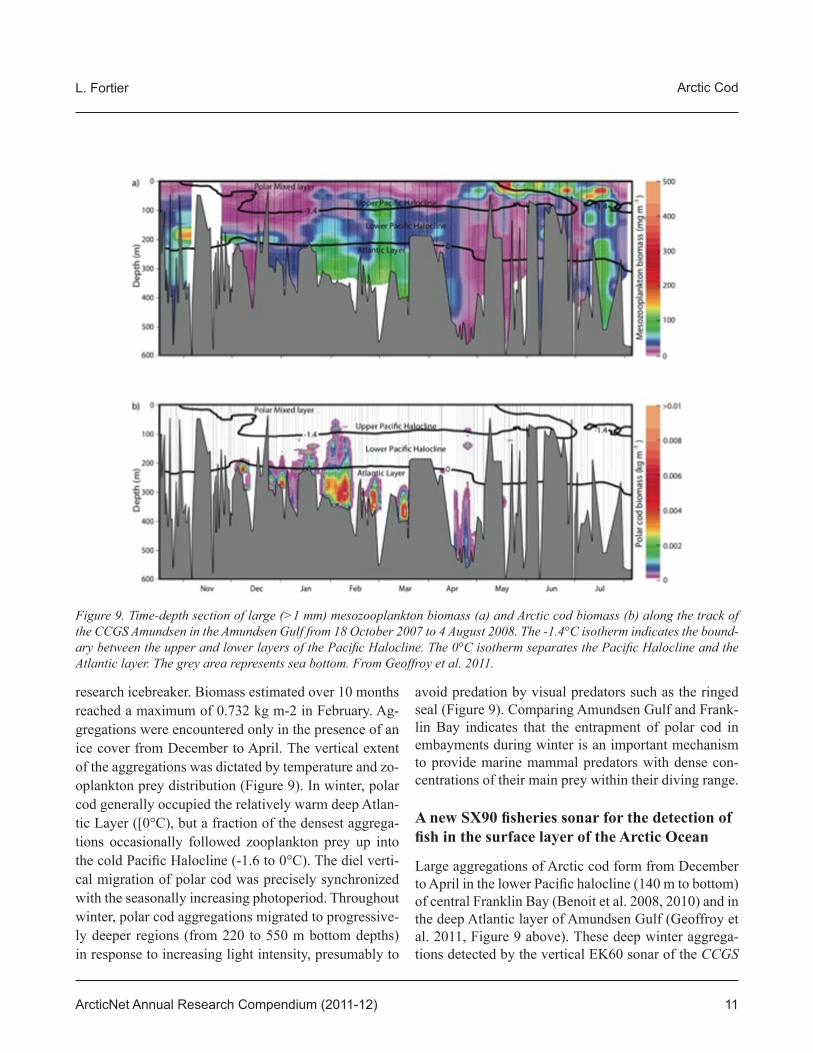
research icebreaker. Biomass estimated over 10 months reached a maximum of 0.732 kg m-2 in February. Ag-gregations were encountered only in the presence of an ice cover from December to April. The vertical extent of the aggregations was dictated by temperature and zo-oplankton prey distribution (Figure 9). In winter, polar cod generally occupied the relatively warm deep Atlan-tic Layer ([0°C), but a fraction of the densest aggrega-tions occasionally followed zooplankton prey up into the cold Pacifi c Halocline (-1.6 to 0°C). The diel verti-cal migration of polar cod was precisely synchronized with the seasonally increasing photoperiod. Throughout winter, polar cod aggregations migrated to progressive-ly deeper regions (from 220 to 550 m bottom depths) in response to increasing light intensity, presumably to
avoid predation by visual predators such as the ringed seal (Figure 9). Comparing Amundsen Gulf and Frank-lin Bay indicates that the entrapment of polar cod in embayments during winter is an important mechanism to provide marine mammal predators with dense con-centrations of their main prey within their diving range.
A new SX90 fi sheries sonar for the detection of fi sh in the surface layer of the Arctic Ocean
Large aggregations of Arctic cod form from December to April in the lower Pacifi c halocline (140 m to bottom) of central Franklin Bay (Benoit et al. 2008, 2010) and in the deep Atlantic layer of Amundsen Gulf (Geoffroy et al. 2011, Figure 9 above). These deep winter aggrega-tions detected by the vertical EK60 sonar of the CCGS
Figure 9. Time-depth section of large (>1 mm) mesozooplankton biomass (a) and Arctic cod biomass (b) along the track of the CCGS Amundsen in the Amundsen Gulf from 18 October 2007 to 4 August 2008. The -1.4°C isotherm indicates the bound-ary between the upper and lower layers of the Pacifi c Halocline. The 0°C isotherm separates the Pacifi c Halocline and the Atlantic layer. The grey area represents sea bottom. From Geoffroy et al. 2011.
12
L. Fortier
ArcticNet Annual Research Compendium (2011-12)
Arctic Cod
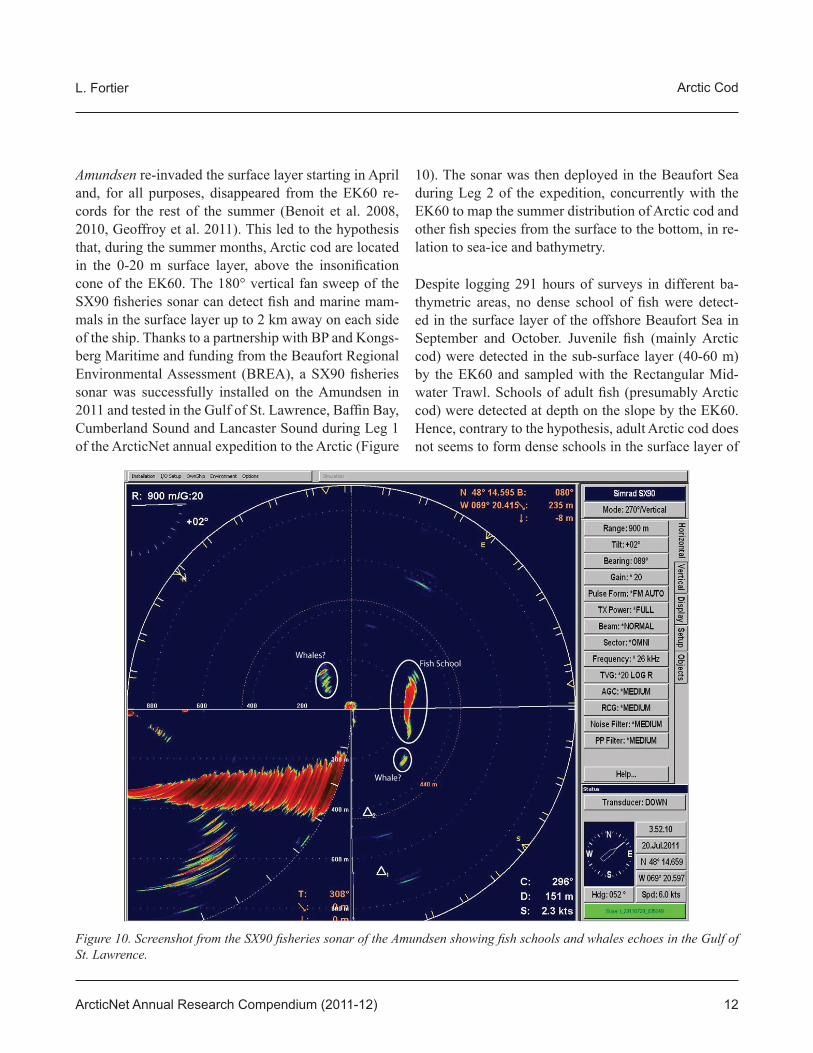
Amundsen re-invaded the surface layer starting in April and, for all purposes, disappeared from the EK60 re-cords for the rest of the summer (Benoit et al. 2008, 2010, Geoffroy et al. 2011). This led to the hypothesis that, during the summer months, Arctic cod are located in the 0-20 m surface layer, above the insonifi cation cone of the EK60. The 180° vertical fan sweep of the SX90 fi sheries sonar can detect fi sh and marine mam-mals in the surface layer up to 2 km away on each side of the ship. Thanks to a partnership with BP and Kongs-berg Maritime and funding from the Beaufort Regional Environmental Assessment (BREA), a SX90 fi sheries sonar was successfully installed on the Amundsen in 2011 and tested in the Gulf of St. Lawrence, Baffi n Bay, Cumberland Sound and Lancaster Sound during Leg 1 of the ArcticNet annual expedition to the Arctic (Figure
10). The sonar was then deployed in the Beaufort Sea during Leg 2 of the expedition, concurrently with the EK60 to map the summer distribution of Arctic cod and other fi sh species from the surface to the bottom, in re-lation to sea-ice and bathymetry.
Despite logging 291 hours of surveys in different ba-thymetric areas, no dense school of fi sh were detect-ed in the surface layer of the offshore Beaufort Sea in September and October. Juvenile fi sh (mainly Arctic cod) were detected in the sub-surface layer (40-60 m) by the EK60 and sampled with the Rectangular Mid-water Trawl. Schools of adult fi sh (presumably Arctic cod) were detected at depth on the slope by the EK60. Hence, contrary to the hypothesis, adult Arctic cod does not seems to form dense schools in the surface layer of
Figure 10. Screenshot from the SX90 fi sheries sonar of the Amundsen showing fi sh schools and whales echoes in the Gulf of St. Lawrence.

13
L. Fortier
ArcticNet Annual Research Compendium (2011-12)
Arctic Cod
the offshore Beaufort Sea in late summer. The acoustic signatures (target strengths) of different (visually iden-tifi ed) marine mammals in a 1.5 km radius around the Amundsen have been acquired with the SX90. These will help identify, monitor, and avoid marine mammals during scientifi c and seismic operations.
DISCUSSION
The long-term monitoring of the ecosystem of the Arctic cod is starting to reveal signifi cant trends in the abundance, reproduction, and population dynamics of the main actors (e.g. Arctic cod and copepods). Al-though still weak, these trends are generally coherent with predictions based on the ecology of the species in a context of progressive sea-ice reduction, warming of the surface layer, and increased upwelling of nutrient by wind. In the Beaufort Sea, they include primarily (1) a multi-year increase in the abundance of some key mem-bers of the low-diversity guild of copepods that makes up the bulk of the mesozooplankton of Arctic seas; (2) a potential shift in the hatching season of Arctic cod to-wards the preferential survival of early hatchers; and (3) the invasion of the offshore dispersion area of Arctic cod juveniles by sand lance juveniles. Such observa-tions are consistent with previous studies documenting higher nutrient availability, primary production in Arc-tic seas characterized by a longer ice-free season (e.g. Rysgaard et al. 1999, Tremblay et al. 2006, 2011); high-er copepod standing stocks in polynyas relative to adja-cent ice-covered regions (Ringuette et al. 2002); and a general (predicted and observed) northward expansion of the distribution of temperate/boreal zooplankton and fi sh into sub-arctic and arctic seas (Babaluk et al. 2000, Beaugrand et al. 2002, Gaston et al. 2003, Perry et al. 2005, Hunt and Megrey 2005, Barber et al. 2008, Ste-phenson and Hartwig 2010).
Although consistent with predictions, the observed trends remain statistically weak because of the short-ness of the time series. Also, while records have accu-mulated and been analysed for the Beaufort Sea where ice cover reduction and surface warming have been in-tense, observations for other regions of the Canadian
sector of the Arctic Ocean either have not been collected as extensively (e.g. Arctic Archipelago) and/or have not been analysed as completely (e.g. North Water, Hudson Bay). This regional imbalance in our monitoring effort can be attributed to the focus of several large initiatives on the Beaufort Sea (CASES, CFL, Malina, ArcticNet partnerships with the oil exploration sector, BREA, etc.), and to the relative success of the mooring program in this region compared to the North Water and Hudson Bay. Clearly, corroboration of the trends observed in the Beaufort Sea for other regions where climate warming has been intense (e.g. northern Hudson Bay), and their contradiction in other, more stable or cooling, regions (e.g. North Water) would be extremely informative. We conclude that, during Cycle II of ArcticNet, the success-ful monitoring of the Beaufort Sea must be expanded imperatively to Baffi n Bay and Hudson Bay, so as to fully capture the potential awakening and response of the Arctic cod ecosystem to climate change and to pro-vide baseline information for future generations of re-searchers.
CONCLUSION
Impacts of the proposed research. As with previous arctic work by my team, several components of the research help inform policy, decisions, and adaptation strategies of stakeholders (e.g. Inuit, Federal depart-ments, Oil Exploration sector) in the rapidly changing and developing Canadian Arctic. Examples of the sig-nifi cance of the research include:
• Our work on the present and future services pro-vided by pelagic arctic marine ecosystems is being incorporated into ArcticNet’s Integrated Regional Impact Assessments for the Canadian Western High Arctic, the Eastern High Arctic, and Hudson Bay;
• Our study of the summer offshore distribution of Arctic cod in southeastern Beaufort Sea is part of the Beaufort Regional Environmental Assessment (BREA) of Aboriginal Affairs and Northern De-velopment Canada (formerly Indian and Northern Affairs Canada);

14
L. Fortier
ArcticNet Annual Research Compendium (2011-12)
Arctic Cod
• The installation, adaptation and deployment of the new SX90 on the Amundsen is conducted in close collaboration with Simrad-Kongsberg and BP, two industrial partners that are interested in testing the capacity of the fi sheries sonar to de-tect marine mammals as a mitigation measure for seismic studies;
• In addition to its contribution to the BREA, the annual survey of the southeastern Beaufort Sea pelagic ecosystem provides the regional back-ground necessary to assess the ecological im-portance of the exploration claims acquired by Imperial Oil Limited and BP at the edge of the Mackenzie Shelf;
• As part of a building collaboration between my team and the Fisheries and Sealing Division of the Nunavut Department of Environment, the Amundsen and Nunavut’s scientifi c trawler MV Nuliajuk were deployed jointly in Cumberland Sound in August 2011 as a pilot attempt to assess pelagic fi sh stocks;
• All data sets generated by my arctic research pro-gram are integrated into the Polar Data Catalogue developed jointly by ArcticNet and the Canadian Cryospheric Information Network;
• The expertise of my team on arctic zooplankton and fi sh is increasingly in demand by other par-ties, for example the Oil Exploration sector (IOL, BP); the Nunatsiavut and Royal Military College joint program on the ecology of Labrador Fjords; the Institute of Marine Sciences at the University of Alaska, Fairbanks; and the Census of Marine Life;
Future Work
As discussed in the previous section, the long-term monitoring of the ecosystem of the Arctic cod is starting to yield signifi cant trends in the abundance, reproduc-tion and population dynamics of the main actors (e.g. Arctic cod and copepods), that are coherent with the general predictions based on the ecology of the species in a context of sea-ice reduction, warming of the surface
layer, and increased upwelling of nutrient by wind. Ac-cordingly, our priority in the coming years will be to maintain and expand the monitoring of the Arctic cod ecosystem in continued collaboration with the Hotspot and Mooring programs of ArcticNet, and in new part-nership with the Remote-Sensing program (CERC on the Remote-sensing of Canada’s new Arctic frontier at Université Laval).
This year we added successfully the SX90 fi sheries so-nar to our arsenal. As part of the renewal of Fortier’s Tier 2 CRC, the LOKI (Lightframe On-sight Keyspe-cies Investigation System), an in-situ taxonomist robot for the determination of the fi ne-scale vertical distribu-tion of mesozooplankton and fi sh larvae, will be devel-oped over the next year. Finally, due to the acoustic tan-trum generated when breaking ice, the many sonars of the Amundsen become useless in ice covers denser than 7/10th. As a result, the spatial distribution of fi sh and zooplankton under the ice remains poorly resolved. In partnership with Kongsberg Maritime and with fi nancial support from the Quebec Government we are planning the acquisition of a Hugin 1000 Autonomous Under-water Vehicle (AUV) with under-ice navigation capa-bility and a 80-km radius of operation to be deployed from the Amundsen. Carrying sonars like the EK60 and SX90, the ecological module of the AUV will enable us to document the vertical and horizontal distribution of Arctic cod and its zooplankton prey under the ice cover of the Canadian Arctic.
Six of the 8 engines and generators of the Amundsen must be replaced and the ship will not be available for science in 2012. With the agreement of AANDC, we are trying for 2012 to transfer our BREA program based on the SX90 to the trawler chartered by DFO (Jim Reist) before resuming operations on the Amundsen in 2013. This plan would allow a direct validation of the SX90 echoes with fi sh collections.
The non-availability of the Amundsen in 2012 will impact briefl y the monitoring of the transformation of ecosystems in the Canadian Arctic Ocean by the Arctic cod, Hotspot, Mooring, and Remote-Sensing programs of ArcticNet. However, these 4 programs will use this

15
L. Fortier
ArcticNet Annual Research Compendium (2011-12)
Arctic Cod
hiatus to regroup and refocus our highly successful and expanding efforts since 2002. We are proposing in 2012 a workshop gathering these and other ArcticNet pro-grams using the Amundsen to (1) discuss and advance the analysis and hybridization of existing data sets that can consolidate and complete the emerging picture of the changing ecosystem of Canadian Arctic seas; (2) to refocus the scientifi c program of the ArcticNet annual mission on the Amundsen as our objectives change in response to the results obtained; (3) to re-organize the Mooring program to offset the retirement of its leader; and (4) to re-build, consolidate and expand the tech-nical team in charge of the maintenance and deploy-ment of the growing pool of scientifi c equipment of the Amundsen.
ACKNOWLEDGEMENTS
In addition to Canada’s Network of Centres of Excel-lence (NCE) program, the Arctic cod (Boreogadus saida) ecosystem under the double pressure of climate change and industrialization (Arctic Cod) program of ArcticNet is supported by several collateral funding sources. We thank the Natural Sciences and Engineer-ing Research Council of Canada (NSERC), the Canada Research Chair program (CRC), the Canada Excel-lence Research Chair program (CERC), the Canada Foundation for Innovation (CFI), the Canadian Interna-tional Polar Year, the Beaufort Regional Environmen-tal Assessment (BREA) program of Aboriginal Affairs and Northern Development Canada, le Fonds québé-cois pour la recherche sur la nature et la technologie (FQRNT, volet Regroupements Stratégiques et volet Équipes), et le Comité national pour la recherche scien-tifi que français (CNRS). Major industrial partners such as BP, Imperial Oil Limited and Kongsberg Maritime contribute to our program. We acknowledge fruitful collaborations with several researchers in the Depart-ments of Fisheries and Oceans Canada, Environment Canada, and Natural Resources Canada. None of our achievements could be possible without the expertise and complicity of the personnel, offi cers and crew of the Canadian Coast Guard. We thank our numerous col-leagues in and outside ArcticNet for their expertise and
data, with emphasis on our friends at the Center for Ob-servation Studies (U. Manitoba), the Norwegian Polar Institute (U. Tromsø) and the National Institute of Polar Research in Tokyo. The research results presented here are contributions to the programs of ArcticNet, Québec-Océan at Université Laval, the Canada Research Chair on the response of marine arctic ecosystems to climate warming, and the Fisheries and Oceans Canada Re-search Chair in marine acoustics applied to resources and ecosystems.
REFERENCES
Babaluk, J.A., Reist, J.D., Johnson, J.D., Johnson, L. 2000. First records of sockeye (Oncorhynchus nerka) and pink salmon (O. gorbuscha) from Banks Island and other records of Pacifi c salmon in Northwest Territo-ries, Canada. Arctic 53(2): 161−164.
Barber, D, Lukovich, J.V., Keogak, J., Baryluk, S., For-tier, L., Henry, G. 2008. The changing climate of the Arctic. Arctic 61(suppl. 1): 7-26.
Beaugrand, G., Reid, P.C., Ibanez, F., Lindley, J.A., Edwards, M. 2002. Reorganization of north Atlantic marine copepod biodiversity and climate. Science 296 (5573): 1692−1694.
Benoit, D., Simard, Y., and Fortier, L. (2008). “Hydroa-coustic detection of large winter aggregations of Arctic cod (Boreogadus saida) at depth in ice-covered Franklin Bay (Beaufort Sea),” Journal of Geophysical Research-Oceans 113.
Benoit, D., Simard, Y., Gagne, J., Geoffroy, M., and Fortier, L. (2010). “From polar night to midnight sun: photoperiod, seal predation, and the diel vertical migra-tions of polar cod (Boreogadus saida) under landfast ice in the Arctic Ocean,” Polar Biol. 33, 1505-1520.
Gaston, A.J., Woo, K., Hipfner, J.M. 2003. Trends in forage fi sh populations in Northern Hudson Bay since 1981, as determined from the diet of nestling Thick-billed murres Uria lomvia. Arctic 56: 227-233.

16
L. Fortier
ArcticNet Annual Research Compendium (2011-12)
Arctic Cod
Geoffroy, M., Robert, D., Darnis, G., Fortier, L. 2011, The aggregation of polar cod (Boreogadus saida) in the deep Atlantic layer of ice-covered Amundsen Gulf (Beaufort Sea) in winter, Polar Biology 4 (12): 1959-1971.
Grebmeier, J.M., Overland, J.E., Moore, S.E., Farley, E.V., Carmack, E.C., Cooper, L.W., Frey, K.E., Helle, J.H., McLaughlin, F.A., McNutt, S.L. 2006. A major ecosystem shift in the northern Bering Sea. Science 311: 1461–1464.
Hunt, G.L., Megrey, B.A. 2005. Comparison of the bio- physical and trophic characteristics of the Bering and Barents Seas. Ices Journal of Marine Sciences 62(7): 1245 – 1255.
Perry, A. L., Low, P. J., Ellis, J. R., and Reynolds, J. D. (2005). “Climate Change and Distribution Shifts in Marine Fishes,” Science 308, 1912-1915.
Ringuette, M., Fortier, L., Fortier, M., Runge, J. A., Bé-langer, S., Larouche, P., Weslawski, J.-M., and Kwas-niewski, S. (2002). “Advanced recruitment and ac-celerated population development in Arctic calanoid copepods of the North Water,” Deep-Sea Research II 49 5081–5099.
Rysgaard, S., Nielsen, T.G., Hansen, B.W. 1999. Sea- sonal variation in nutrients, pelagic primary production and grazing in a high-Arctic coastal marine ecosystem, Young Sound, northeast Greenland. Marine Ecology Progress Series 179: 13–25.
Stephenson, S.A., Hartwig, L. 2010. The Arctic Marine Workshop. Canadian Manuscript Report of Fisheries and Aquatic Sciences 2934.
Thanassekos, S., Fortier, L. 2012. An individual based model of Arctic cod (Boreogadus saida) early life in Arctic polynyas: I. Simulated growth in relation to hatch date in the Northeast Water (Greenland Sea) and the North Water (Baffi n Bay). Journal of Marine Sys-tems 93: 25-38.
Thanassekos, S., Robert, D., Fortier, L. 2012. An indi-vidual based model of Arctic cod (Boreogadus saida) early life: II. Length-dependent and growth-dependent mortality. Journal of Marine Systems 93: 39-46.
Tremblay, J.E., Bélanger, S., Barber, D.G., Asplin, M., Martin, J., Darnis, G., Fortier, L., Gratton, Y., Link, H., Archambault, P., Sallon, A., Michel, C., Williams, W.G., Philippe, B., Gosselin, M. 2011. Climate forcing multiplies biological productivity in the coastal Arctic Ocean. Geophysical Research Letter 38, L18604. doi: 10.1029/2011GL048825.
Tremblay, J.E., Michel, C., Hobson, K.A., Gosselin, M., Price, N.M. 2006. Bloom dynamics in early open-ing waters of the Arctic Ocean. Limnology and Ocean-ography 51(2): 900–912.
Tynan, C.T., DeMaster, D.P. 1997. Observations and predictions of Arctic climatic change: Potential effects on marine mammals. Arctic 50: 308-322.
Welch, H.E., Bergmann, M.A., Siferd, T.D., Martin, K.A., Curtis, M.F., Crawford, R.E., Conover, R.J., Hop, H. 1992. Energy fl ow through the marine ecosystem of the Lancaster Sound Region, Arctic Canada. Arctic 45: 343–357.
2011-12 PUBLICATIONS
All ArcticNet refereed publications are available on the ASTIS website (http://www.aina.ucalgary.ca/arcticnet/).
Benoit, D., 2012, Océanographie, distribution et cycle annual de la more arctique (Boreogadus saida) en mer de Beaufort: une approche hydroacoustique, Ph.D. the-sis, Université Laval, Canada.
Bouchard, C., Fortier, L., 2011, Circum-arctic compari-son of the hatching season of polar cod (Boreogadus saida): a test of the freshwater winter refuge hypoth-esis, Progress in Oceanography, v.90, 105–116.

17
L. Fortier
ArcticNet Annual Research Compendium (2011-12)
Arctic Cod
Darnis, G., Fortier, L., 2012, Zooplankton respiration and the export of carbon at depth in the Amundsen Gulf (Arctic Ocean), Journal of Geophysical Research - Oceans, in press.
Darnis, G., Fortier, L., 2011, Respiration zooplancto-nique et exportation du carbone en profondeur dans le golfe d’Amundsen (Océan Arctique), Annual General Meeting of Québec-Océan, Lac Delage, Québec, 17 November.
Darnis, G., Fortier, L., 2011, Zooplankton respiration and the export of carbon at depth in the Amundsen Gulf (Arctic Ocean), High latitude pelagic and ice ecosys-tems meeting, University of Tromso, Tromso, 31 Oc-tober.
Darnis, G., Robert, D., Pomerleau, C., Link, H., Ar-chambault, P., Nelson, R.J., Geoffroy, M., Tremblay, J.-É., Lovejoy, C., Ferguson, S.H., Hunt, B.P.V., Fortier, L., 2012, Current state and changing trends in Canadian Arctic marine ecosystems: II. Secondary production and biodiversity, Climatic Change.
Deming J., Fortier, L., 2011, Introduction to the spe-cial issue on the biology of the circumpolar fl aw lead (CFL) in the Amundsen Gulf of the Beaufort Sea (Arc-tic Ocean)., Polar Biology, v.34, no.12, 1797-1801.
Forest, A., Babin, M., Stemmann, L., Picheral, M., Guidi, L., Robert, D., Sampei, M., Fortier, L., 2011, To-ward a size-structured ecosystem model for the Beau-fort Sea: data assimilation of zooplankton and vertical particle fl uxes, MEECE 2011 Summer School on ad-vanced ecosystem modelling, Ankara, 7-14 September.
Forest, A., Babin, M., Stemmann, L., Picheral, M., Sam-pei, M., Fortier, L., Sahlin, J.P., 2012, Physical and bio-logical determinants of particulate organic carbon and nitrogen across the Arctic shelf-basin system (Beaufort Sea)., Ocean Sciences Meeting 2012, Salt Lake City, 20-24 February.
Forest, A., Robert, D., Fortier, L., Stemmann, L., Picher-al, M., Burdorf, L., Sampei, M., Miquel, J.C., Babin, M., 2011, Zooplankton community structure during the Ajurak-Malina campaign 2009: a multi-methodological approach, Malina Plenary Meeting, Villefranche-sur-Mer, 10 May.
Forest, A., Stemmann, L., Picheral, M., Sampei, M., For-tier, L., Raimbault, P., Joux, F., Babin, M., 2011, Malina 2009: oceanographic context, carbon fl uxes and fi rst steps toward an integrative study, Malina Plenary Meet-ing, Villefranche-sur-Mer, 9 May.
Forest, A., Tremblay, J.E., Gratton, Y., Martin, J., Gag-non, J., Darnis, G., Sampei, M., Fortier, L., Ardyna, M., Gosselin, M., Hattori, H., Nguyen, D., Maranger, R., Vaqué, D., Marassé, C., Pedros-Alio, C., Sallon, A., Michel, C., Kellogg, C., Deming, J., Shadwick, E., Thomas, H., Link, H., Archambault, P., Piepenburg. D., 2011, Biogenic carbon fl ows through the plankton-ic food web of the Amundsen Gulf (Arctic Ocean): a synthesis of fi eld measurements and inverse modeling analyses, Progress in Oceanography, v.91, 410-436.
Fortier, L., Darnis, G., Robert, D., Link, H., Geoffroy, M., Forest, A., Wold, A., Archambault, P., Piepenburg, D., Hop, H., Falk-Petersen, S., 2012, Pelagic and Ben-thic Foodwebs, Circumpolar Flaw Lead Study, chapter 4.
Geoffroy, M., Robert, D., Darnis, G., Fortier, L., 2011, The aggregation of polar cod (Boreogadus saida) in the deep Atlantic layer of ice-covered Amund-sen Gulf (Beaufort Sea) in winter, Polar Biology, v.34, no.12, 1959-1971.
Geoffroy, M., Rousseau, S. Pyc, C., Fortier, L., 2011, Utilisation d’un sonar halieutique (Simrad SX90) pour l’étude des niveaux trophiques supérieurs de l’écosystème marin arctique, Assemblée générale annuelle de Québec-Océan, Lac Delage, 17-18 Novem-ber.
Geoffroy, M., Rousseau, S., Pyc, C., Fortier, L., 2011, Searching for the missing cod using hydroa-

18
L. Fortier
ArcticNet Annual Research Compendium (2011-12)
Arctic Cod
coustic technology, Beaufort Regional Environmental Assessment (BREA) Day, Inuvik, 5 December.
Polyakov, Y.V., Alexeev, V.A., Ashik, I.M., Bacon, S., Beszczynska-Möller, A., Carmack, E.C., Dmitrenko, I.A., Fortier, L., Gascard, J.-C., Hansen, E., Hölemann, J., Ivanov, V.V., Kikuchi, T., Kirillov, S., Lenn, Y.-D., McLaughlin, F.A., Piechura, J. Repina, I., Timokhov, L.A., Walczowski, W., Woodgate, R., 2011, Fate of early 2000s Arctic warm water pulse., Bulletin of the American Meteorological Society, v.92, no.5, 561-566.
Pyc, C., Geoffroy, M., Rousseau, S., Reier Knudsen, F., Gammelseter, O.B., Levesque, K., Fortier, L., 2011, Ca-nadian Beaufort Sea Active Acoustics Research pro-ject, BP external advisory panel meeting, Tampa, 24 November.
Robert, D., Falardeau, M., Suzuki, K., Fortier, L., 2011, Investigating trophic interactions between lar-val Arctic cod (Boreogadus saida) and their zooplank-ton prey in the rapidly warming Beaufort Sea, 35th An-nual Larval Fish Conference, Wilmington, 24 May.
Robert, D., Grant, C., Gagnon, J., 2011, Data from the environment and marine resources assessment of Im-perial Oil/BP Exploration License Areas 446, 449, 451 and 453 (Beaufort Sea), in the summers of 2009, 2010 and 2011, Québec, Canada, 74 pp.
Sampei, M., Sasaki, H., Forest, A., Fortier, L., 2012, A substantial export fl ux of particulate organic carbon linked to sinking dead copepods during winter 2007-2008 in the Amundsen Gulf (southeastern Beaufort Sea, Arctic Ocean), Limnology and Oceanography, v.57, no.1, 90-96.
Suzuki, K.W., Robert, D., Darnis, G., Sampei, M., Bé-langer, S., Fortier, L., 2012, Small copepods thrive as sea ice cover regresses in the southeastern Beaufort Sea, Canadian Arctic, Deep-Sea Research Part I: Oceano-graphic Research Papers.
Thanassekos S., Fortier, L., 2012, An individual based model of Arctic cod (Boreogadus saida) early life in Arctic polynyas: I. Simulated growth in relation to hatch date in the Northeast Water (Greenland Sea) and the North Water (Baffi n Bay), Journal of Marine Sys-tems, v.93, 25-38.
Thanassekos S., Robert, D., Fortier, L., 2012, An indi-vidual based model of Arctic cod (Boreogadus saida) early life: II. Length-dependent and growth-dependent mortality, Journal of Marine Systems, v.93, 39-46.
Thanassekos, S., 2011, Modélisation individu-centrée de la croissance et de la survie larvaire de la morue arctique (Boreogadus saida) dans deux polynies, Ph.D. thesis, Université Laval, Canada, 114 pp.
Tremblay J.-É., Bélanger, S., Barber, D.G., Asplin, M., Martin, J., Darnis, G., Fortier, L., Gratton, Y., Link, H., Archambault, P., Sallon, A., Michel, C., Williams, W.G., Philippe, B., Gosselin, M., 2011, Climate forcing multiplies biological productivity in the coastal Arctic Ocean, Geophysical Research Letter, v.38, L18604.
Wold, A., Darnis, G., Søreide, J.E., Leu, E., Philippe, B., Fortier, L., Poulin, M., Kattner, G., Graeve, M., Falk-Petersen, S., 2011, Life strategy and diet of Ca-lanus glacialis during the winter-spring transition in Amundsen Gulf, southeastern Beaufort Sea, Polar Biol-ogy, v.34, no.12, 1929-1946.
![MALLE NATALI FORTIER N° Inventaire [400]€¦ · Web view22 Des mains pleines de doigts François David. Natali Fortier Nathan Album. M F David 407 Mise à jour 03-07-2016 + DVD](https://img.pdfslide.fr/doc/110x75/60527a927a1d113bac1d5f99/malle-natali-fortier-n-inventaire-400-web-view-22-des-mains-pleines-de-doigts.jpg)




























