Embed Size (px)
Citation preview

Le flux membranaire
Le flux membranaire
I. Généralités1. Structures membranaires Données expérimental
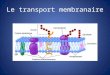
les données morphologiques microscopiques et biochimiques et l'étude de la membrane artificielle a permis d'établir un modèle appelé mosaïque fluide.
La mosaïque fluide :
la membrane et concilier par un film de lipides en il, l'intérieur du film contient les ans ne est trop forte, l'extérieur du film contient des lipides en Lille et on parle d'idyllique et protéines interagissent avec sept des couches envoie distinguer plusieurs cas :
protéine transmembranaire :
les protéines présentes des zones hydrophobes qui traversent là des couches lipidiques, ces protéine présente donc une partie cytosolique au lichen est parti transmembranaire une partie luminale ou extracellulaire.
Protéines ancrées par un lipide :
certains chaînes polypeptidiques sont liés de manière convaincante à des lipides qui retiennent sur une face de la membrane.
Protéines extrinsèques :
les protéines extrinsèques interagissent par des liaisons faibles avec la membrane se sent des interactions protéines - proteines au interaction protéines – lipides.
1

Le flux membranaire
Propriété générale des membranes :
dans une membrane des molécules peuvent bouger les unes par rapport aux autres dans le plan de la membrane par contre, les molécules ne peuvent pas spontanément changer d'orientation par rapport au plan de la membrane.
Malgré leur mobilité, la surface de la membrane n'est pas homogène.
Asymétrie membranaire une membrane sépare de compartiments sur une composition est une fonction différente, les deux faces de la membrane ne sont donc pas équivalentes les deux feuillets lipidiques , le feuillet lipidique externe n'a pas la même composition que feillet cytosolique.
Il était une transmembranaire présente une orientation donnée hérésie de lucidité. Uniquement du côté extracellulaire ou luminale glycoprotéine ou glycolipides.
La façon avec laquelle les protéines sont ancrées un lipide est différente sur la face du luminale et cytosolique.
Fonction générale des membranes :
à travers une lipidique le passage se fait par diffusion simple dans le sens de gradient électrochimique seules les petites molécules liposoluble peuvent passer .
Les autres passages en grâce à des protéines ces transports seront spécifiques régulés et saturable on atteint une vitesse maximale.
Flux membranaire :
mise en évidence :
par autoradiographie, pour les sécrétions, des vésicules partent du réticulum endoplasmique fusionne avec la face cis d'un appareil de Golgi, elles vont former de nouveau s'accumule se divisent en vésicules de sécrétions qui vont fusionner avec la membrane plasmique.
2

Le flux membranaire
Endocytoze :
les vésicules d'endocytoze qui se forment à partir des visécule…… Fusionne avec endosome, le endosome reçoit aussi les lysosomes primaires qui proviennent de la face trans de l'appareil de Golgi ,à partir de endosome se forment et visécule de recyclage qui retourne à la membrane plasmique… Et des lysosomes secondaires qui contiennent des Hydrolases acides et des éléments à dégrader.
Le flux rétro garde :
il permet le retour d'éléments vers le réticulum endoplasmique, visécule partent de l'endosome et fusionne avec la face transe de l'appareil de Golgi, puis des visécules vont retourner sacules trans vers sacule cis ….. Enfin de la face cis partent des vésicules qui retournent au réticulum endoplasmique.
C. mécanisme de transport :
3

Le flux membranaire
un certain point de la membrane vont s'organiser les ports transmembranaires ce qui permet la courbure de la membrane et la concentration de certaines protéines transmembranaires dans le bougre.
Par l'intermédiaire de protéine transmembranaire en 20 pouvoirs concentrés le bourdon des protéines et il se trouve dans la lumière de localité, en effet les protéines transmembranaires peut interagir avec les protéines solubles de la matière avec leur portion luminale.
Ce de manière générale les protéines transportées sont appelé carg.
Transports :
une fois la visécule formée la membrane se dissocie, ce transport peut faire intervenir le cytosquelette.
Fusion :
4

Le flux membranaire
La fusion entre une visécule et le compartiment accepteur est spécifique grâce un état des interactions entre une protéine transmembranaire et la visécule appelée SNARE.
Les SNARS peuvent être sous forme active ou inactive, elles sont des protéines donc régulées.
Contrôle de transport
les principales étapes de transport sont contrôlées par des protéines particulières qui X. les nucléotides ,elle sont appelés G-protéines il existe 2 grandes places de G protéines les protéines monomérique et les protéines trimérique .
Les petites G- protéines avec de nombreux nommé intercellulaire, il peut se trouver sous 2 formes :
une forme qui fixe le GTP est une forme qui fixe le GDP. En passe de la forme GTP à la forme GTP par une réaction d'échange,le GDP est libéré dans le cytosol et du GTP vient se fixer à sa place. Cette réaction d'échange et favorisé par un GEP et inhibé par un GDI .
5

Le flux membranaire
Les principales étapes de transport vésiculaire ne peuvent se faire que si une G proteine données est sous forme GTP , en fonction des activités GAP, GEF et GDI, le transport vésiculaire sera plus ou moins intense.
Léger protéines les plus importantes du transport vésiculaire sont les protéines de la famille RAB.
Membrane plasmique :
particularité structurale :
la face extracellulaire de la membrane d'un plasmique est recouverte de nombreux résidus des glucidiques qui forment des glycoprotéines ou des glycolipides et constitue le cellcoat ou glycocalix. Le rôle des glycocalix est un rôle de protection et de reconnaissance intercellulaire.
La membrane plasmique présente souvent des domaines de la composition biochimique est rende notamment dans les tissus qui couverte une cavité de l'organisme épithélium on distingue un domaine apical est un domaine baso-latéral
II. Transports à travers les membres :
transport passif :
généralités :
ne consomme pas d'énergie et se font donc en fonction du gradient électrochimique pour un
diffusion simple :
c’est le passage à travers les couches lipidiques, il concerne les molécules liposolubles et petites, ce sont les gaz respiratoires CO2 et O2 ET LE MONOXYDE D'AZOTE QUI EST UNE MOLÉCULE DE COMMUNICATION des hormones lipidiques et les médicaments.
Diffusion facilitée :
6

Le flux membranaire
elle s'est aussi dans le sens du gradient électrochimique mais met en jeu des protéines, ces transports seront donc spécifiques régulés et saturable.
Les cannaux :
ce sont des bouteilles de transmembranaire qui forme un petit port par lequel elle peut passer des ions minéraux ou de l’eau, le passage que des interactions entre les protéines et la molécule transportée, il est donc spécifique. Les cannaux peuvent être en ont fermé, on connaît des canaux régulés par la différence de potentiel transmembranaire, on connaît aussi des canaux régulés par la fixation de ligants, il existe encore d'autres types de régulation.
Les canons ont une capacité de transport extrêmement importante.
Les perméases forment un complexe avec la molécule transportée, elle présente 2 conformation, une ouverte d'un côté de la membrane, d'autre ouverte dans l'autre côté de la membrane le complexe se forme du côté de la plus forte concentration et se dissocie du côté de la plus faible concentration.
Transports actifs :
les transporteurs actifs consent de l'énergie mais peut le faire passer une molécule contre son gradient électrochimique en distingue deux grandes catégories.
Les pompes :
pompes à sodium potassium : savant couplé hydrolyse d’un nucléotide au transport d'une molécule . Hydrolyse du nucléotide entraîne des changements de conformation qui provoque le transport.
Les pompes de transport désirant minéraux :
cette pompe permet d’expulser trois cellules hors de la cellule et de faire rentrer 2 protons pour chaque ATP hydrolysé.
Pompe à calcium :
7

Le flux membranaire
présente sur la membrane plasmique toutes les cellules, elle expulse le calcium.
Pompe à protons H+ :
elle permet à certains cellules de sécréter des ions H+, on les ouvre aussi sur la membrane des lysosomes.
Banque transportant les molécules organiques :
les pompes appartiennent à la famille ABC et elles sont codées par des pompes MDR(multi drug resistance) ses pompes expulce hors de la cellule des molécules organiques avec une spécificité lâche, 50 permettent aux cellules d'expulser des toxiques, et peut aussi expliquer à résistance aux médicaments de certains cellules concernent ou de certains parasites.
Cotransporteur (transporteurs actifs secondaires) :
ce sont des molécules transmembranaires qui couplé le transport d'une molécule dans le sens de son gradient électrochimique au transport d'un autre type de molécules contre son gradient on appel symport un cotransporteur qui fait passer les deux molécules dans le même sens et l'antiport qui fait passer les molécules en sens inverse.
8

Le flux membranaire
perméabilité potentiellement membranaire :
dans toutes les cellules, la pompe à sodium potassium établi un gradient de sodium favorable à l'entrée de sodium dans la cellule et un gradient de potassium favorable à la sortie de potassium. La perméabilité de la membrane en sodium est très faible, on peut la négliger, par contre les membranes présentes des canaux potassiques ouverts en permanence, le potassium sort de la cellule est écrit un gradient de charges favorables à l'entrée dans la cellule excès de charges positives à l'extérieur de la cellule. Les ions de potassium sont donc soumis à 2 forces de sens opposé, il existe un gradient de concentration pour lequel les deux forces s'équilibrent, ce gradient correspond à un potentiel fixe pour une cellule donnée. Le potentiel de repos.
Potentiel d'action :
dans certains cas, des stimulus vont changer les propriétés de perméabilité de la membrane en aura donc un changement de la concentration des ions est donc un changement dans le potentiel transmembranaire, qui est appelé potentiel d'action.
Si un stimulus entraîne l'ouverture d'un canal sodique, il va y avoir entré du sodium sous l'effet du champ électrique et du gradient de concentration, ceci entraîne une dépolarisation, le changement du voltage transmembranaire entraîne la fermeture du canal sodique une ouverture de canaux
9

Le flux membranaire
potassiques de potassium va sortir sous l'effet du compagnon de charges et du gradient de la concentration, on va donc avoir repolarisation et fermeture du canal potassique la membrane a retrouvé ses propriétés, le fonctionnement de la pompe à sodium potassium va reconstituer le gradient de la cellule qui va revenir au potentiel de repos.
Certains membrane présente des canaux contrôlés par le page qui permette au potentiel d'action de se propager proche en proche sur la surface membranaire si la membrane contient des canaux sodiques sensibles au montage la dépolarisation initiale entraîne l'ouverture des canaux dansants voisinages l'ouverture de ces canaux provoque une dépolarisation qui entraîne la fermeture des canaux sodiques et l'ouverture de canaux potassiques, c'est-à-dire une repolarisation.
On a donc eu déplacement du potentiel d'action. La propagation s'effectue de la même façon de proches en proche, il ne peut pas y avoir de retour en arrière car après ouverture les canaux sodiques craignent une conformation ne permettant plus d'ouverture pendant quelques millisecondes, on parle de périodes de réfractaires.
Le récepteur transmembranaire est modifié de conformation de la partie cytosolique, ceci va modifier les propriétés biochimiques du récepteur est donc le fonctionnement cellulaire.
10

Le flux membranaire
Reconnaissance adhérence cellulaire :
généralités :
les cellules reconnaissance leur environnement direct grâce à des molécules transmembranaires appelées molécules d'adhérence pour qu'il y ait adhérence il faut que la partie cytosolique soit reliée au cytosquelette, ce lien n'est pas direct mais se fait par l'intermédiaire de protéine appelée protéines de la plaque, la partie extracellulaire voit reconnaître la manière spécifique et réversible une autre protéine de la cellule voisine ou sur la matrice extracellulaire, cette reconnaissance peut être réglée.
On peut avoir adhérence diffusée, molécules d'adhérence sur qu'une grande surface membranaire ou une adhérence régente charnelle et molécules d'adhérence sont regroupés en certains points de la membrane pour former une jonction.
11

Le flux membranaire
L'interaction d'une molécule d'adhérence avec son ligants peut entraîner une signalisation intracellulaire, la différence entre récepteur pour molécules de signalisation et molécule d'adhérence est définie expérimentalement par la force d'adhérence ou de cohésion, cette force est due au grand nombre de molécules exprimées, action est faible et c'est l'ensemble des interactions qui fait adhérence . Les sélectines sans des molécules d'adhérence intercellulaire qui reconnaisse des résidus glucidiques du glycocalix, et interviennent dans les deux interactions leucocyte –paroi-vasculaire.
Les immunoglobulines :
on les appelle souvent CAM (cell adhesion molecul) dans les protéines d'adhérence intercellulaire généralement homophile , elles interviennent notamment dans le système nerveux, parfois les CAM de la famille des immunoglobulines reconnaissent une integrine s'est notamment le cas dans le sujet immunitaire.
Cadherine :
les molécules cadherine sont des molécules intercellulaires homophile , en distingue deux sous-groupes : les cadherine desmosomales sont responsables d'une adhérence conjoncturelle classique peut intervenir dans une adhérence diffusée ou jonctionel .
On connaît environ une vingtaine de cadherine classique, un type cellulaire donné d'exprimer de manière spécifique à l'ensemble de quelques cadherine, les cadherine étant responsables d'interactions homophile, les cellules du même type vont se regrouper un ensemble pour former les tissus et des organes.
12

Le flux membranaire
Intégrine :
les intégrinesr sont formée de 2 chaînes transmembranaires associées de manière non covalentes qui forment un heterodimaire …… D'une manière générale, les intégrismes reconnaissent la nature extracellulaire.
Roles :
d'une manière générale les molécules d'adhérence informent la cellule sur son environnement direct immédiat par exemple : les cellules normales peuvent se diviser que celles sont en contact avec la matière extracellulaire, et ne se divise plus lorsque les contacts intercellulaires centraux importants inhibition par contact.
Les molécules d'adhérence permettent aux cellules de se regrouper pour former les tissus et les organes les molécules d'adhérence contribuent donc à la morphogenese pendant le développement embryonnaire, les tissus vont notamment se développer des structures stables appelaient jonction cellulaire qui relie les cellules entre elles ou à la MEC.
Jonction cellulaire :
d'ancrage :
13

Le flux membranaire
les jonctions d'ancrage quatre des jonctions intercellulaires ou des jonctions à la matrice, les molécules d'adhérence peuvent être reliées soit aux filaments intermédiaires soit aux filaments d’actines.
Communication :
Jonction de type GAP : les jonctions communicantes permettent à 2 cellule voisine d'échanger directement de petites molécules hydrosolubles sont passées vers le milieu extracellulaire.
Sur la membrane plasmique des cellules on trouve des protéines transmembranaires appelées connexines qui s'associent entre telle pour former un tunnel transmembranaire appelé connexion, 2 connexions de cellule voisine s'associe entre eux pour permettre le passage de molécules d'un cytosol à l'autre.
Les jonctions communicantes permettent le couplage métabolique et fonctionnel des cellules à l'intérieur des tissus. Ce couplage est régulé.
se à la limite du domaine apical et du domaines baso-latérale des cellules épithéliales au niveau de ses jonctions, les protéines transmembranaires des deux cellules voisines interagissent entre elles et forment l'espace intercellulaire, c'est pays rempart aura des petites parois qui laissent penser de
14

Le flux membranaire
manière spécifique une ou plusieurs molécules. Les jonctions contrôlent le passage par voie paracellulaire (transcellulaires).
Le réticulum endoplasmique constitue un réseau de citernes aplaties ou tubulaire, sa lumière communique avec l'espace péri nucléaire, le réticulum coûte plasmique peut fixer des ribosomes, les réticulums endoplasmiques rigoureux ou non , réticulum endoplasmique lisse n'aurait eu que l'on endoplasmiques représentent la majorité des surfaces membranaires de la cellule.
Adressage des protéines :
Protéine endoluminale :
dans ce cas la synthèse de la protéine débute par une séquence hydrophobe appelée peptide signale, type reconnaît un complexe macromoléculaire, la SRP (signale recognition particul) qui arrêtent l'élongation.
15

Le flux membranaire
La SRP va reconnaître un récepteur sur la membrane réticulum endoplasmique, le ribosomes va se fixer sur un petit canal protéique, translocon qui s'ouvre pour laisser passer le début de la chaîne polypeptidique le peptide signale se dissocie de la SRR et va interagir avec le translocon l'élongation reprend est la chaîne polypeptidique passe dans la lumière, au codant stop, l'extrémité c- terminale est libérée dans la lumière, le ribosomes se dissocie, le translocon se referme et le peptide signale est copé par un enzyme , la protéine est libérée dans la lumière, les peptides signalent repassent dans cytosol ou il est dégradé en acides aminés.
Protéine à passages transmembranaires :
c'est un peptide hydrophobe synthétisé par un ribosomes fixé sur un translocon , ce peptide interagit avec ce translocon est entré à la dissociation de la liaison ribosomes translocon, l'élongation se poursuit dans le cytosol, le translocon se referme et les petites et pour de passe dans la bicouche lipidique pour former un passage transmembranaire, si la chaîne polypeptidique débute par un peptide signale, on obtient une protéine à un passage transmembranaire a extrémité NH2 luminale.
Peptide d’ionisation de la translocation :
les peptides d'initiation à la translocation sont des peptides qui fonctionne comme un peptide signale qui sont à l'intérieur de la chaîne polypeptidique et qui ne sont pas clivé dans la protéine mature, il constitue donc un passage de transmembranaires.
16

Le flux membranaire
17

Le flux membranaire
Un pedigree d'initiation peut reconnaître la SRP avec 2 orientations, une orientation permet avoir des protéines à NH2 cytosolique, dans ce cas les peptides d'initiation se dissocientde laSRP , interagit avec le translocon, l'élongation reprend et permet le passage de l'extrémité COOH dans la lumière, d'orientation permet avoir des protéines à NH2 luminale, après fixation de laSRP sur son récepteur le peptides d'initiation interagit avec le translocon et permet le passage de l'extrémité NH2 dans la lumière, le ribosomes se dissocie du translocon et termine la synthèse dans le cytosol, l'extrémité COOH est donc cytosolique.
C. Peptides à plusieurs passages transmembranaires
les protéines à plusieurs passages transmembranaires présentes sur leur chaîne polypeptidique des successions des peptides hydrophobes qui se comportent soit comme un peptide initiation soit comme un peptide d'arrêt de la translocation.
3. Autre fonctiona. modification post-traductionnelle
les protéines prennent leur structure tridimensionnelle avec l'aide de chaperon et la disulfite isomèrase , en se trouve donc des ponts de disulfures que dans les partis luminale des protéines.La réaction de N glycolisation (fixation de glucides sur une fonction amine) débute dans le réticulum endoplasmiquetroisième réaction d'ancrage des protéines glycolipideils se fondent dans la lumière du réticulum d'endoplasmique
b. synthèse des lipides membranairesla synthèse de la plupart des lipides membranaires se fait sur la face cytosolique des membranes du réticulum endoplasmique, les lipides passent ensuite de manière spécifique dans le feuillet luminale, cette réaction appelée eliph-elop est catalysée par une flipase.
c. Ségrégation et les concentrationsla psychologue endoplasmique constituait un compartiment qui a une composition spécifique ,les ions calcium peuvent y être concentré grâce à une pompe à calcium.
d. Détoxication :au niveau du réticulum endoplasmique, les toxiques sont rendus hydrosolubles grâce à des réactions de conjugaison, il peut ensuite être expulsé par des pempes de la famille ABCi codé par le gène MPRi ils sont alors incapables de rentrer à nouveau dans la cellule, l'entrée se fait par diffusion simple.
III. L'appareil de GolgiUltrastructure l'appareil de Golgi est formé de plusieurs dictosomes, chaque dictosome est formé de sacule en forme de disque environ 10 sacules parallèle par dictosome, un dictosome présente une face convexe qui est en relation par les vésicules avec le réticulum endoplasmique, sur cette face se forme de nouveaux sacule , on parle de face cis ou de réseaux cis-golgien , sur la face concave les sacules se fragmentent en visécules, on parle de la vaste trans ou de raiseau trans-golgien.Les sacule progresse donc de la face cis à la face trans . en périphérie du dictosome, on trouve des visécule qui assure le transport vers les sacule précédents.
18

Le flux membranaire
2. Fonctiona. modification des protéines et des lipides
les molécules sans modifier de manière ordonnée, on a une succession coordonnée de réactions biochimiques dans les différents sacule, exemple : deuxième étape des N- glycolisation et O-glyciclisation.
b. Synthèse de polysaccharides :au niveau de l'appareil de Golgi vont être synthétisé des glucides complexes qui peuvent constituer des mucus ou participé à la matrice EC.
c. Ségrégation des vésicules
Au niveau du réseau cis-golgie il est visécule apporte des molécules synthétisées dans le réticulum endoplasmique, parmi celles-ci se trouvent des protéines qui présentent un signalent de rétention, ces protéines sont spécifiquement incorporées dans un bourgeon qui retourne au réticulum endoplasmique, ces protéines sont donc des protéines spécifiques du réticulum endoplasmique.Au niveau de la face transniveau de la face trans se forme à trois types de vésicules : les lysosomes, les visécule de ses créations contrôlées, les visécules de ses créations constitutives.Les protéines étaient présentes un residu mannose-6-phsophate sont spécifiquement incorporées dans les lysosomes. Les molécules qui forment des complexes avec la chromoganine sont spécifiquement incorporées dans les visécule de sécrétions contrôlées. Les molécules les gênant par de mannose-6-phosphates et qui ne forment pas se complexes avec la chromoganine sont incorporées dans des visécule de sécrétions constitutives.Le retour vers les sacules cis :
19

Le flux membranaire
les sacule se font en permanence sur la face cis,l faut que les visécules spécifiques vers le sacule qui le précèdent.
IV. Visécule de sécrétionsla sécrétion constitutive se produit en permanence et dans tous les types cellulaires.La sécrétion contrôlée se produit dans les cellules spécialisées, l'exorciste ne se produits qu'après un stimulus.1. Sécrétions contrôlées
le produit de sécrétions peut avoir été synthétisé dans le réticulum endoplasmique ou dans l'appareil de Golgi, il est incorporé spécifiquement dans l'est visécule de sécrétions contrôlées parce qu'ils forment des complexes avec le chromogranine , les protéines peuvent subir dans la vie séculaire des phénomènes de maturation(clivage proteio- lipidique) les visécules moléculaires sont parfois synthétisés dans le cytosol puis transporté spécifiquement à travers la membrane des visécules.
2. Sécrétions constitutivesce sont des visécule qu'ils fusionnent en permanence avec la membrane plasmique, elles permettent le renouvellement des membranes plasmiques et la synthèse de la MEC.Les régulations de cette voie se font au niveau des synthèses, ce sont des régulations à long terme (par heures).
V. Visécule d'endocytoze1. Phagocyte
la phagocyte permet l'absorption de particules, la membrane plasmique reconnaît la particule par des récepteurs plus ou moins spécifiques, ceci déclencherait une réorganisation du cytosquelette, ceci provoque également la formation d'expansion cellulaire appelée pseudopodes qui reconnut la particule
Il se forme une visécule appelée phagocyte. Cet phagocyte est particulièrement actif dans certains cellules spécialisées comme les macrophages ou polynucléaires neutrophiles. Le phagosome va fusionner avec l'endosome et se constituants et l’élément phagocyté sera généralement détruit.
2. Pinocytose :
20

Le flux membranaire
la pinocytose se produit en permanence dans tous les types cellulaires et permet l'absorption de molécules en solution. La pinocytose peut être spécifique grâce à des récepteurs membranaires
Là visécules brisure avec le endosome ce qui entraînait dissociation du complexe récepteur ligants, dans certains cas là visécules de pinocytose a simplement traversé la cellule, on aura passage du pôle apical au pôle baso-latérale ou vice versa, c'est le phénomène de tranxytose .
3. L’endosome :le endosome est un compartiment qui reçoit du visécule de phagocytose et de pinocytose ainsi que les lysosomes primaires le endosome va former dès visécule de recyclage qui renvoie à la membrane plasmique les molécules qui peuvent recevoir (exp récepteur après libération du ligants), de la même façon le endosome forme dès visécule qui rapporte des molécules vers la face trans de l'appareil de Golgi. À partir de l'endosom se forme dès visécule qui contiennent les hydrolase acides et les éléments à dégrader, ses visécule s'appelle des lysosomes secondaires.
VI. Lysosomes1. lysosomes primaires
ce sont visécule qui se forment à partir du réseau trans-golgien , leurs protéines transmembranaires sont fortement glycolisés ce qui les rend résistantes aux hydrolase acides, elle incorpore spécifiquement des hydrolase grâce à des récepteurs de mannose-6-phosphate et présente de nombreux transporteurs membranaires qui permettent l'acidification de la lumière et le transport de petites molécules vers le cytosol. Dans certaines cellules spécialisées, ces lysosomes peuvent fusionner avec la membrane plasmique, en générale île fusionne avec l'endosome.
2. Lysosomes secondaires
21

Le flux membranaire
Les lysosomes secondaires se forment à partir de l'endosome , les hydrolase et les transporteurs membranaire proviennent des lysosomes primaires, les concessions à dégrader proviennent de visécule de pinocytose, de phagosomes ou d’auto-phagosome.les auto-phagosomes sont des structures formées par deux membranes qui entourent les constituants cellulaires à dégrader,l’autophagosome fusionne avec l'endosom. Après dégradation, les petites molécules passent dans le cytosol où il se sent recycler, dans certains cas, il reste des éléments non dégradables qui forment des corps résiduels.
3. Lysosomes et les pathologies :dans certains cas, une ou plusieurs des enzymes de dégradation ne sont pas fonctionnelle à la suite de mutations, les cellules peuvent accumuler des corps résiduels, on parle de maladie de surcharge. L'accumulation de métabolites anormaux perturbe le fonctionnement cellulaire.Parfois la cellule phagocyte des cristaux minéraux non dégradables, les cristaux au niveau des lysosomes secondaires, perce la membrane et les hydrolase passe dans le cytosol, ceci provoque une inflammation chronique qui peut favoriser le développement de certains cancers.
22