Embed Size (px)
Citation preview

LES NAVETTES MITOCHONDRIALES
• Le NADH,H+ dans cytosol (produit lors réaction de déshydrogénation) ne franchit pas le membranemitochondriale Utilisation de navettes
• Navette Malate / Aspartate
• Navette Glycérol phosphate
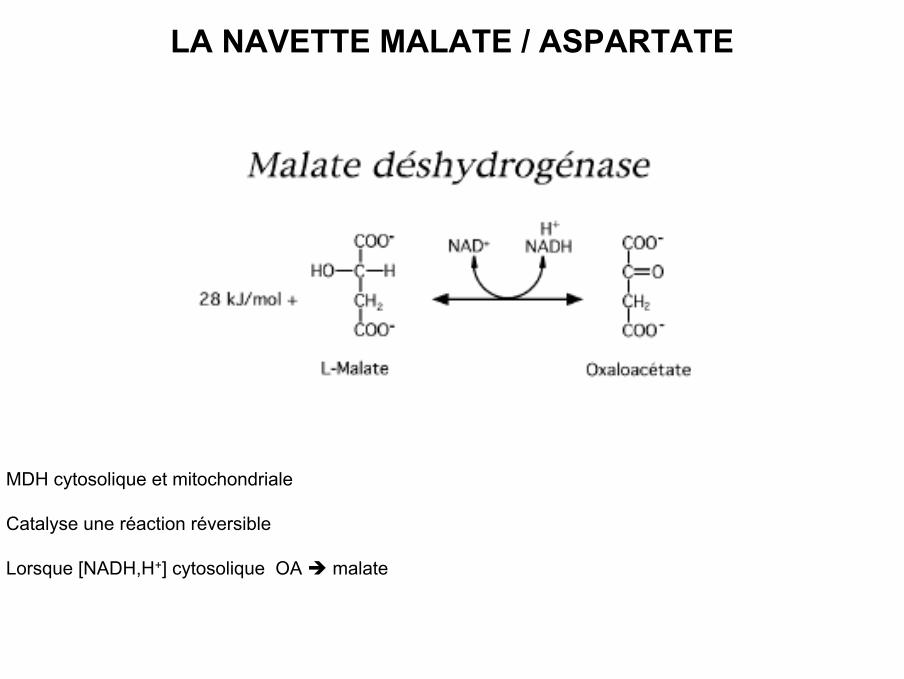
LA NAVETTE MALATE / ASPARTATE
• MDH cytosolique et mitochondriale
• Catalyse une réaction réversible
• Lorsque [NADH,H+] cytosolique OA malate
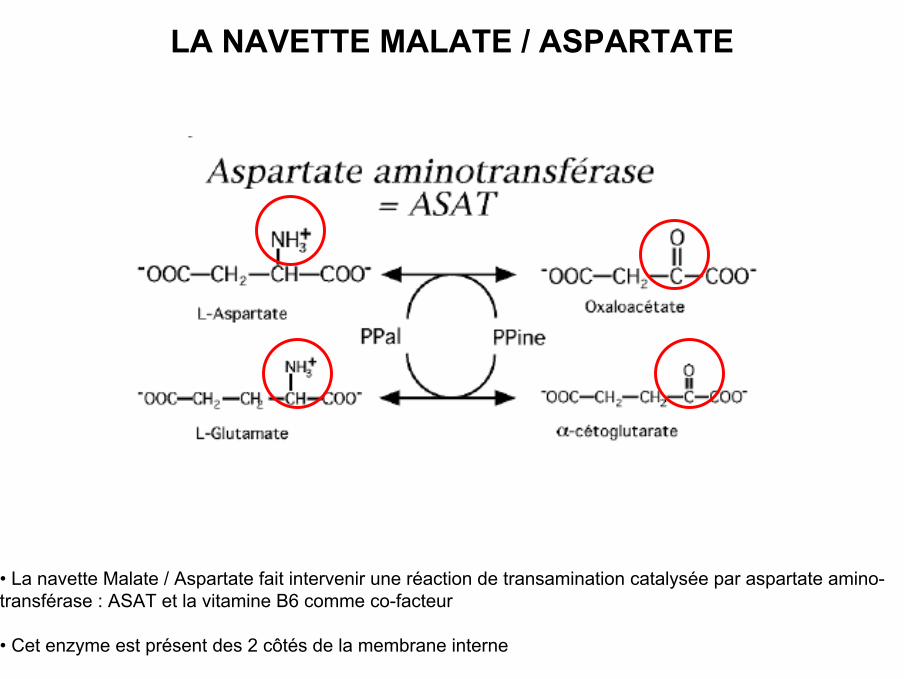
LA NAVETTE MALATE / ASPARTATE
• La navette Malate / Aspartate fait intervenir une réaction de transamination catalysée par aspartate amino-transférase : ASAT et la vitamine B6 comme co-facteur
• Cet enzyme est présent des 2 côtés de la membrane interne
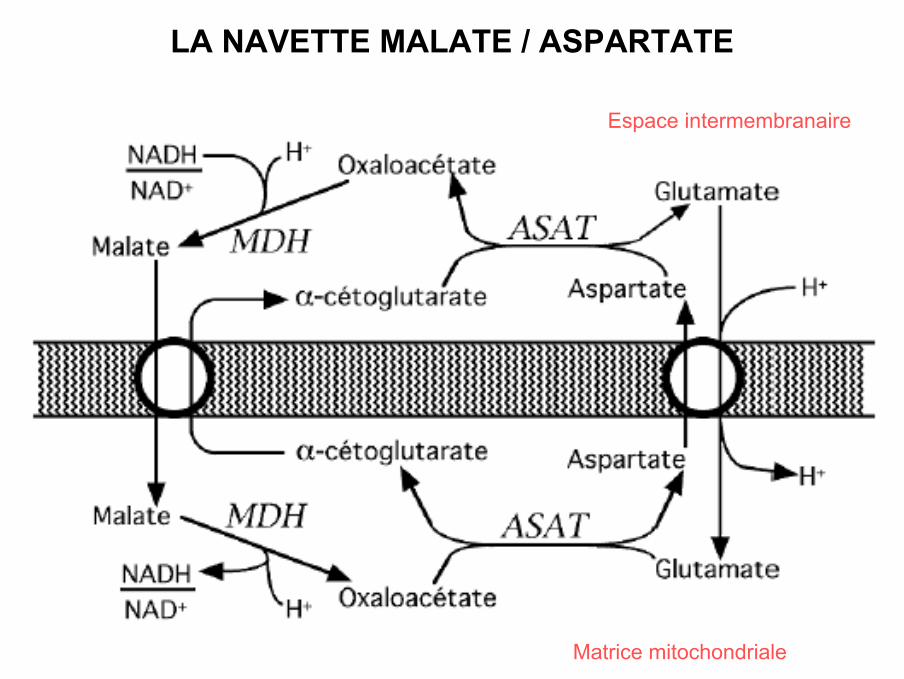
LA NAVETTE MALATE / ASPARTATE
Matrice mitochondriale
Espace intermembranaire

LA NAVETTE MALATE / ASPARTATE
Bilan
1 molécule de NADH,H+ cytosolique 1 molécule de NADH,H+ dans
mitochondrie
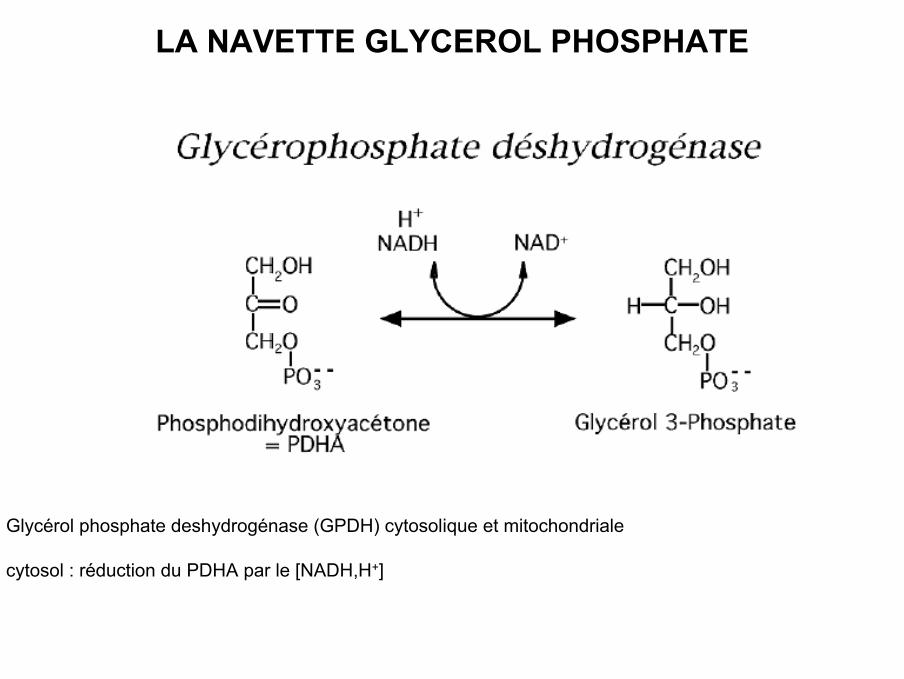
LA NAVETTE GLYCEROL PHOSPHATE
• Glycérol phosphate deshydrogénase (GPDH) cytosolique et mitochondriale
• cytosol : réduction du PDHA par le [NADH,H+]
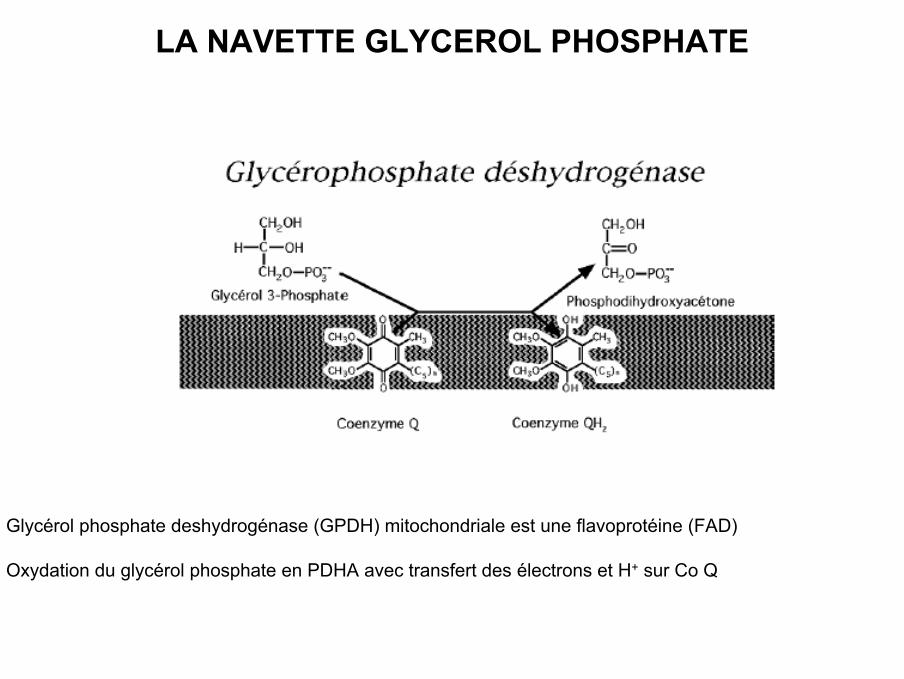
LA NAVETTE GLYCEROL PHOSPHATE
• Glycérol phosphate deshydrogénase (GPDH) mitochondriale est une flavoprotéine (FAD)
• Oxydation du glycérol phosphate en PDHA avec transfert des électrons et H+ sur Co Q
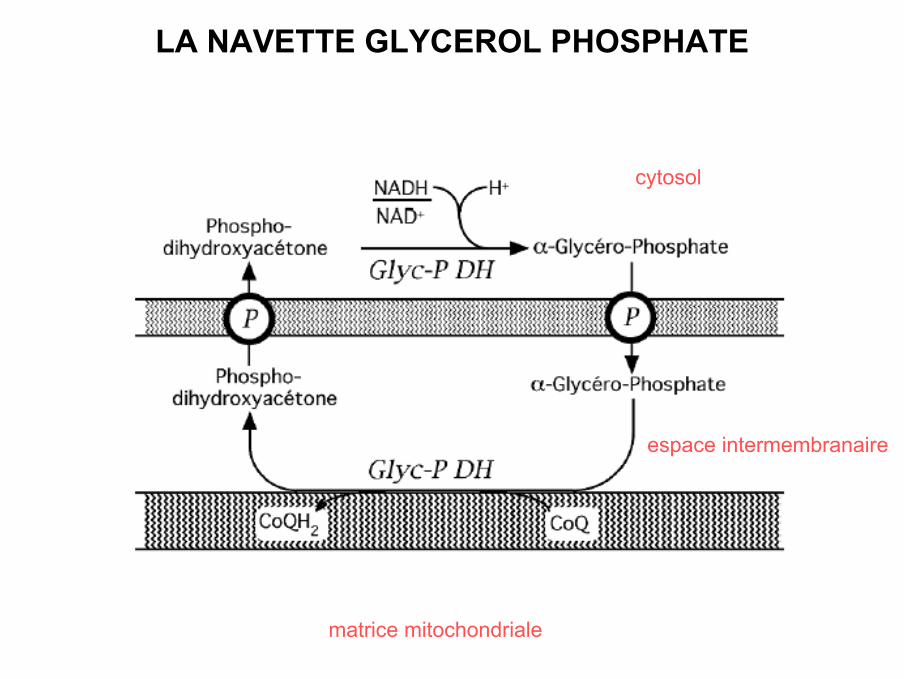
LA NAVETTE GLYCEROL PHOSPHATE
matrice mitochondriale
espace intermembranaire
cytosol

Bilan
1 molécule de NADH,H+ cytosolique 1 molécule de FADH2 dans
mitochondrie
LA NAVETTE GLYCEROL PHOSPHATE
Selon la navette utilisée : 1 molécule de glucose va générer36 ou 38 ATP
LA CHAINE RESPIRATOIRE :
TRANSFERT D’ELECTRONS ETPHOSPHORYLATION OXYDATIVE
Université de Caen – UFR de médecineS. Allouche - 2006/2007

• la présence d’oxygène est indispensable pour les eucaryotes aérobies (≠procaryotes anaérobies)
• L’oxygène est utilisé dans la respiration = processus le plus énergétique (ATP)
• La respiration cellulaire (certaines étapes) se déroule dans la mitochondrie
• La production d’énergie associe 2 processus :
• Le transport d’électrons à partir des cofacteurs réduits (NADH,H+, FADH2) jusqu’àl’oxygène
• La phosphorylation de l’ADP en ATP
PHOSPHORYLATION OXYDATIVE
INTRODUCTION
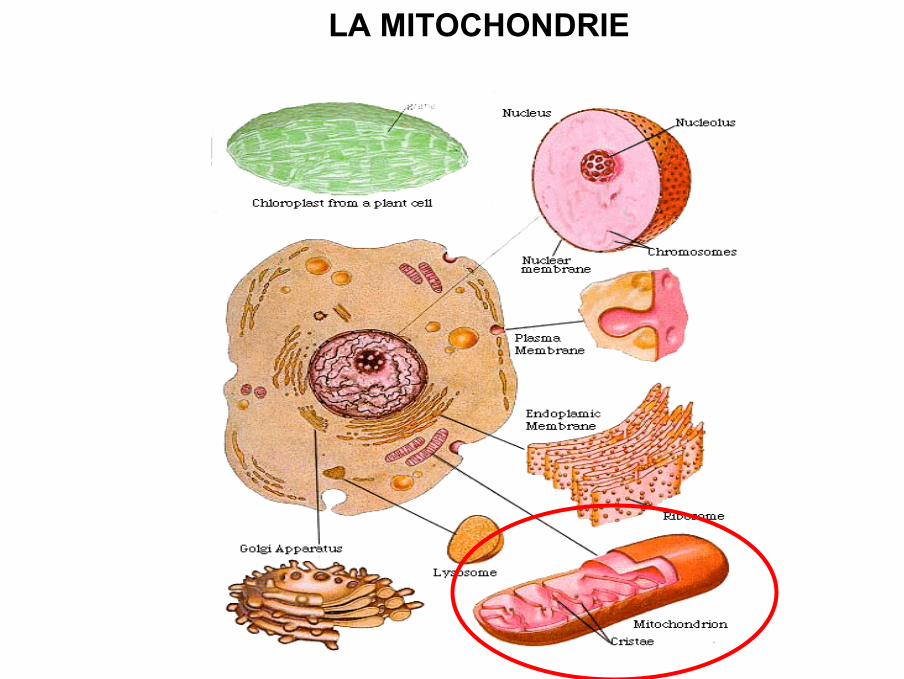
LA MITOCHONDRIE
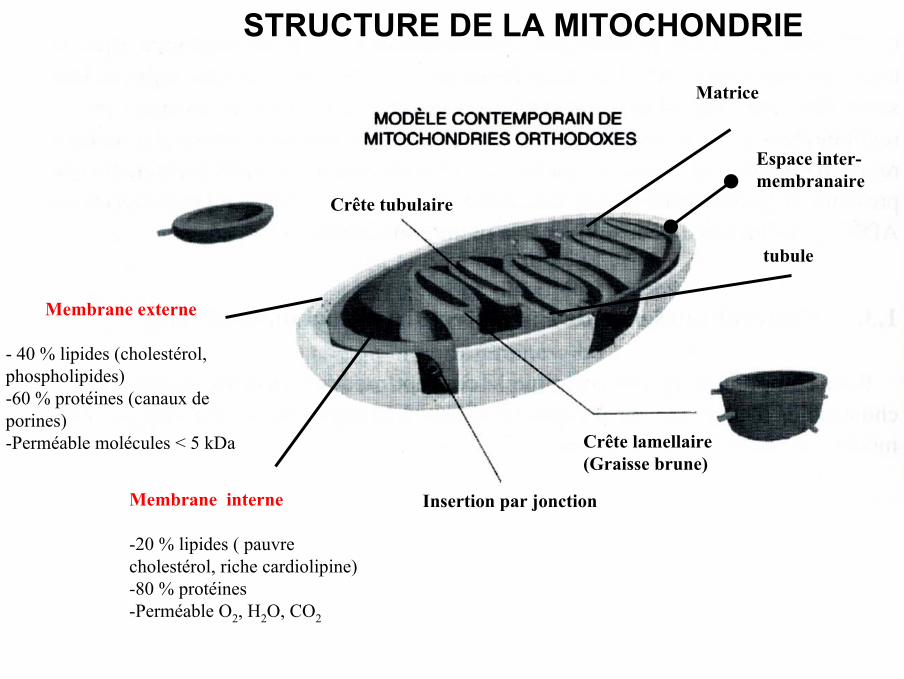
Crête tubulaire
Crête lamellaire(Graisse brune)
Insertion par jonction
tubule
Membrane externe
- 40 % lipides (cholestérol,phospholipides)-60 % protéines (canaux deporines)-Perméable molécules < 5 kDa
Matrice
Membrane interne
-20 % lipides ( pauvrecholestérol, riche cardiolipine)-80 % protéines-Perméable O2, H2O, CO2
STRUCTURE DE LA MITOCHONDRIE
Espace inter-membranaire
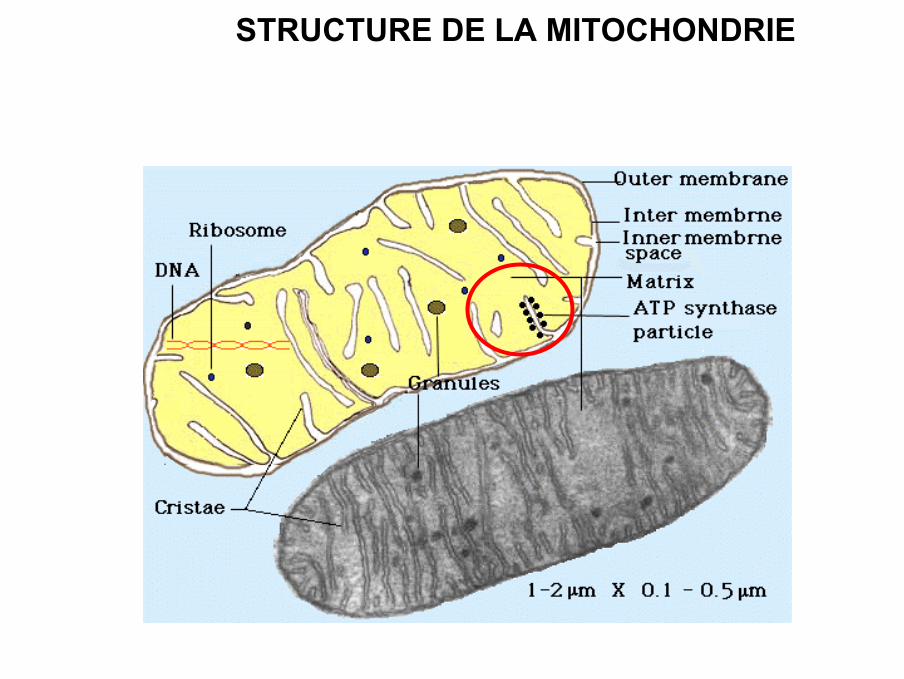
STRUCTURE DE LA MITOCHONDRIE
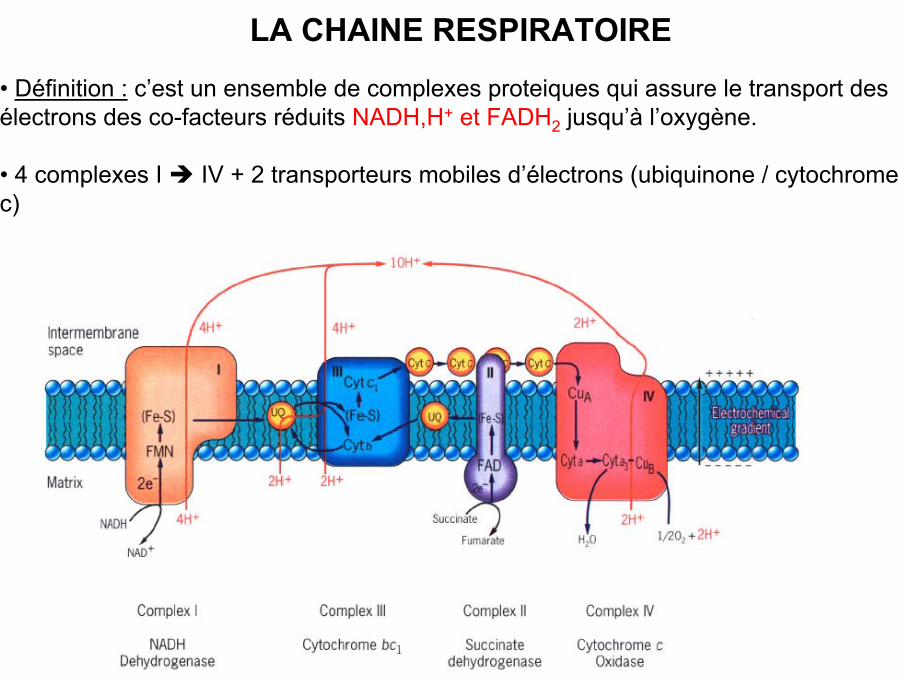
LA CHAINE RESPIRATOIRE
• Définition : c’est un ensemble de complexes proteiques qui assure le transport desélectrons des co-facteurs réduits NADH,H+ et FADH2 jusqu’à l’oxygène.
• 4 complexes I IV + 2 transporteurs mobiles d’électrons (ubiquinone / cytochromec)

• NAD (nicotinamide dinucléotide)
• FAD ou FMN (nucléotides flaviniques)
• Les électrons de ces cofacteurs proviennent des réactions de déshydrogénation:
-glycolyse
-Décarboxylation oxydative du pyruvate
-cycle du citrate
−β-oxydation
- Navettes Malate / Aspartate et Glycérol-phosphate
ORIGINE DES COFACTEURS REDUITS
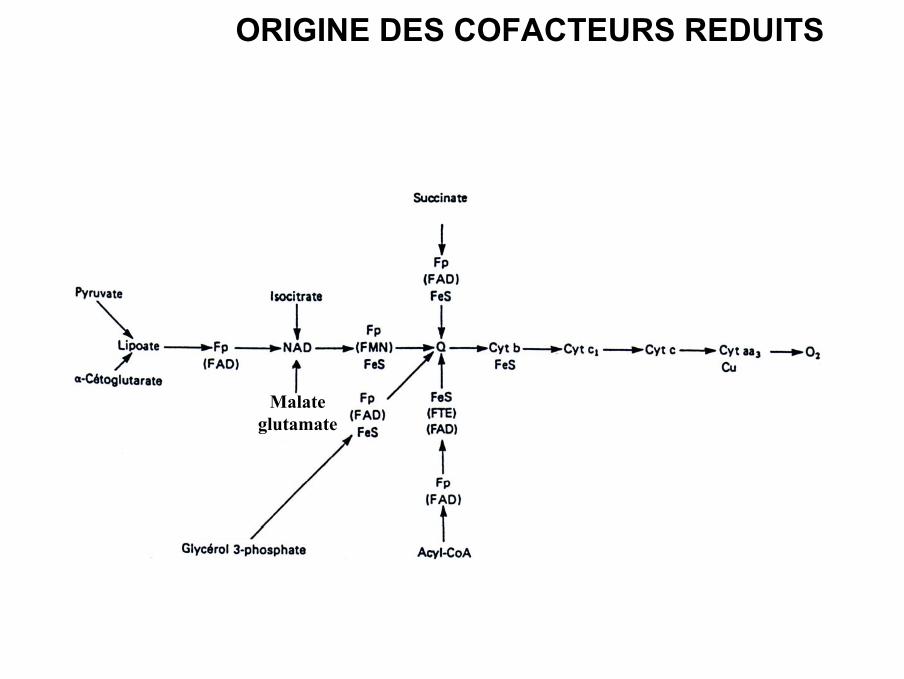
ORIGINE DES COFACTEURS REDUITS
Malateglutamate
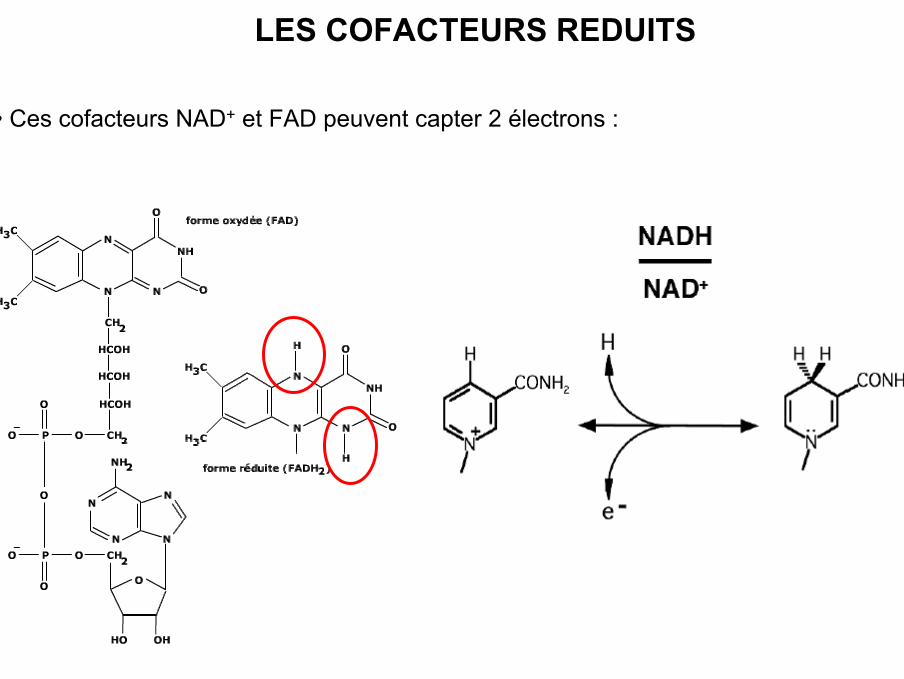
• Ces cofacteurs NAD+ et FAD peuvent capter 2 électrons :
LES COFACTEURS REDUITS

REACTIONS D’OXYDO-REDUCTION
• Les réactions au niveau de la chaîne respiratoire sont des oxydo-réductions
•définitions :
- toute molécule capable de gagner ou perdre un électron = couple d’oxydo-réduction ou rédox
-l’oxydation se définit comme une perte d’électron
-la réduction comme le gain d’électron.
Ces réactions sont toujours associées à une libérationd’énergie qui servira indirectement à la synthèse d’ATP

• La capacité à accepter ou à céder un électron est défini par le potentield’oxydoréduction ou rédox.
• Mesure du potentiel rédox (E0) au moyen d’une pile électrochimique : électroded’hydrogène de référence. Cette demi-pile est constituée d’une solution 1M en H+ (pH =0) et de H2 gazeux à une pression 1 atm en contact avec une électrode inerte de platine,et la F.E.M. (force électromotrice) est égale par convention à 0v.
• Lorsque le potentiel rédox d’un composé est négatif, cela signifie qu’il céderafacilement ses électrons à l’hydrogène (H2) : on qualifie alors le composé de réducteur.
•Inversement, lorsque le potentiel rédox d’un composé est positif , il possède uneaffinité plus importante pour les électrons comparée à l’hydrogène : on la qualified’oxydant.
REACTIONS D’OXYDO-REDUCTION
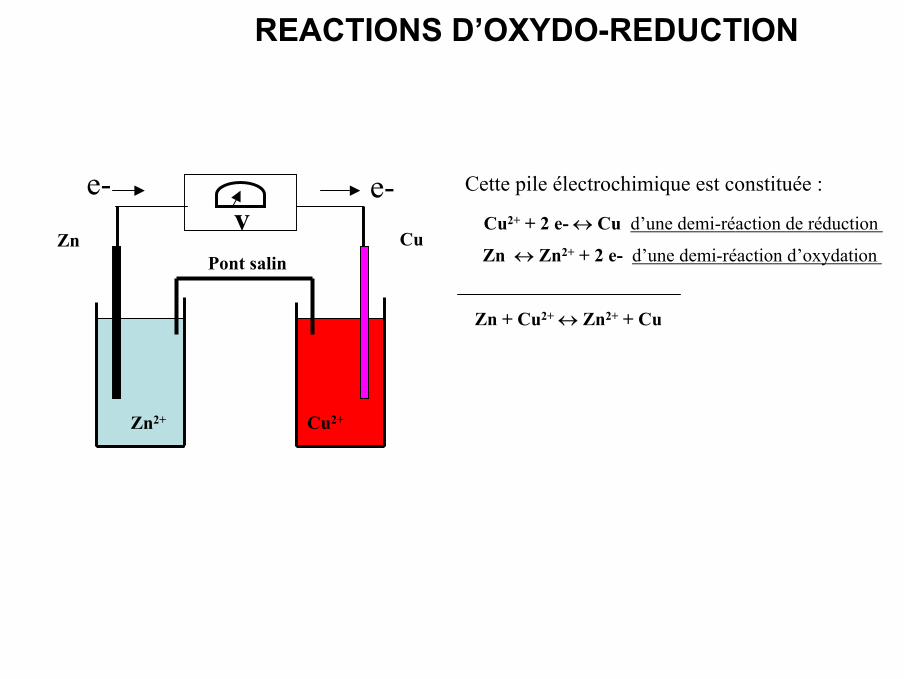
Pont salinZn Cu
Zn2+ Cu2+
ve- e-
Cu2+ + 2 e- ↔ Cu d’une demi-réaction de réduction
Zn ↔ Zn2+ + 2 e- d’une demi-réaction d’oxydation
Cette pile électrochimique est constituée :
Zn + Cu2+ ↔ Zn2+ + Cu
REACTIONS D’OXYDO-REDUCTION

• Réactions cellulaires dans des conditions standard à pH=7, 25°c, solutions desoxydants et réducteurs = 1 M F.E.M. de la demi-pile à hydrogène est alorségale à –0.42V.
• Pour les réactions biologiques, on parlera de potentiel rédox standard (Eo’)
• Différence de potentiel rédox (∆E0’) entre 2 couples rédox : ∆E0’= Eo’oxydant -Eo’ réducteur
• Ce transfert d’électron est étroitement associé à un transfert d’énergie par larelation suivante :∆G0’= - n X f X ∆E0’ = variation d’énergie libre standard
où n correspond au nombre d’électrons transférésf correspond à la constante de Faraday (23.06 kcal.V-1.mol-1 ou 96,406 kJ.V-1.mol-1)∆E0’ (en V)
REACTIONS D’OXYDO-REDUCTION

POTENTIELS REDOX

• Quelle est la différence de potentiel lorsque les 2 électrons passent du NADH,H+
jusqu’à l’oxygène ?
½ O2 + 2H+ + 2e- H2O E0’ = +0.82 V
NAD++2H++2e- NADH,H+ E0’ = -0.32 V
• Le NADH,H+ (réducteur) va céder ses 2 électrons qui vont aller réduire l’oxygène(oxydant) en H2O.• ∆E0’ = (+0.82)-(-0.32) = +1.14 V
• ∆G0’ = - 2 X 23.06 X 1.14 =-52.6kcal/mol ou 220 kJ/mol
• Pour la synthèse d’une molécule d’ATP ∆G0’ = + 8 kCal / mol
• Si rendement de 100 % : 52 / 8 = 6 ATP
• En pratique, seules 3 molécules d’ATP produites
TRANSFERTS D’ELECTRONS ET ENERGIE
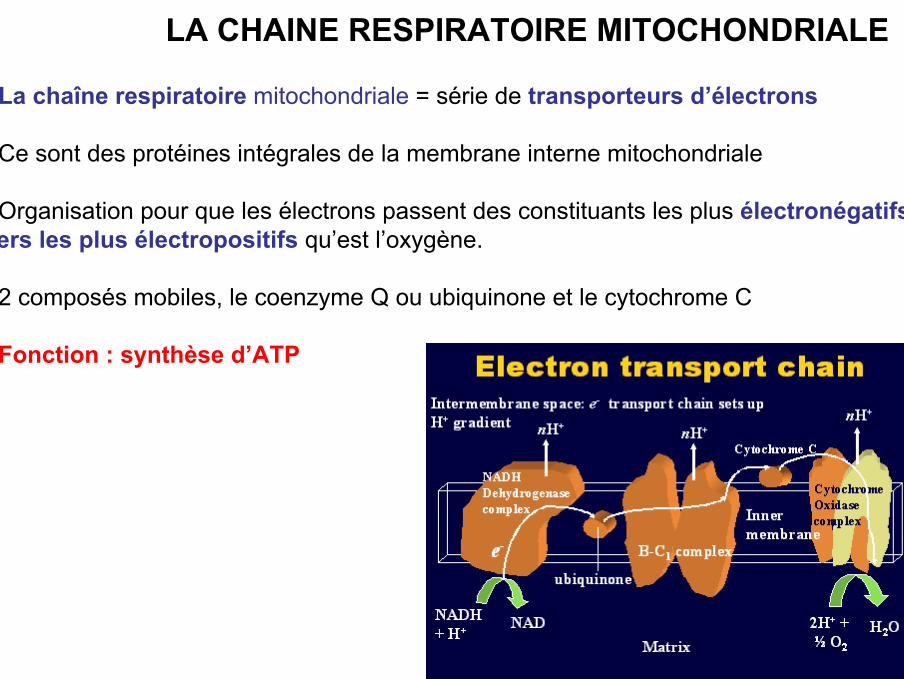
LA CHAINE RESPIRATOIRE MITOCHONDRIALE
• La chaîne respiratoire mitochondriale = série de transporteurs d’électrons
• Ce sont des protéines intégrales de la membrane interne mitochondriale
• Organisation pour que les électrons passent des constituants les plus électronégatifsvers les plus électropositifs qu’est l’oxygène.
• 2 composés mobiles, le coenzyme Q ou ubiquinone et le cytochrome C
• Fonction : synthèse d’ATP

COMPLEXE I
• Complexe I ou NADH-ubiquinone réductase ou NADH déshydrogénase
• Composition : 42 sous-unités dont 7 sont codées par l’ADN mitochondrial(ND1 7)
• Rôle : oxydation du NADH, H+ et transfert des électrons et protons sur lecoenzyme Q
• Transfert des électrons et protons assuré par 2 types de groupementsprostéthiques : FMN et 6 centres Fe / S
• Agencement des accepteurs d’électrons selon leur potentiel rédox (négatif positif)
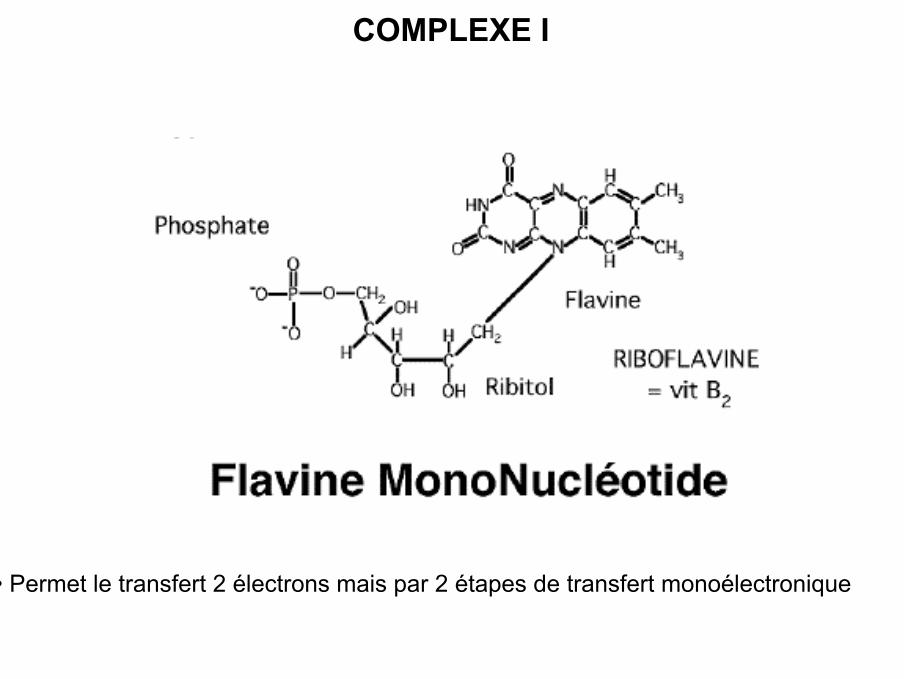
COMPLEXE I
• Permet le transfert 2 électrons mais par 2 étapes de transfert monoélectronique
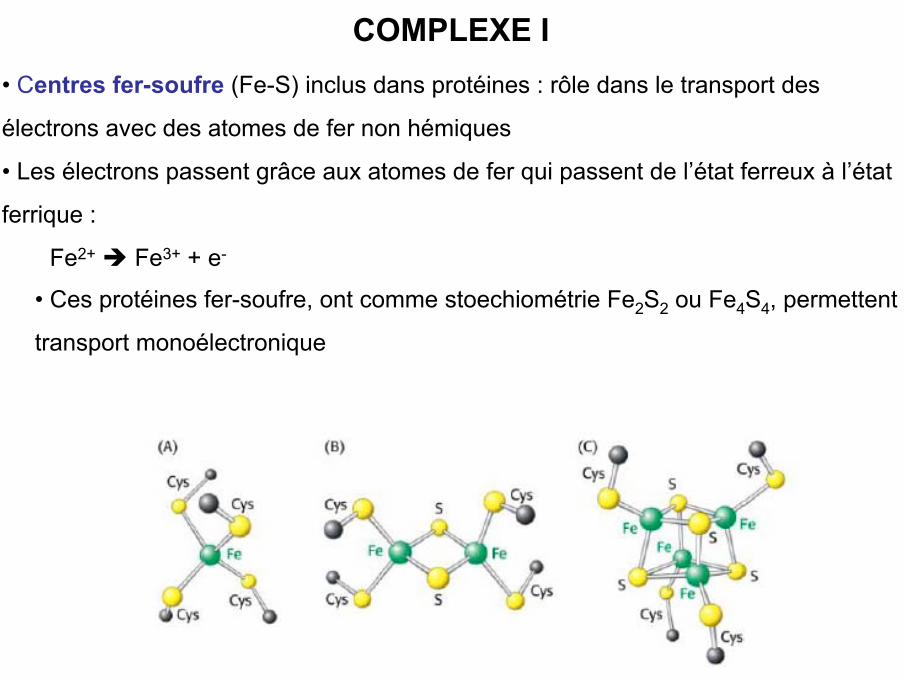
• Centres fer-soufre (Fe-S) inclus dans protéines : rôle dans le transport des
électrons avec des atomes de fer non hémiques
• Les électrons passent grâce aux atomes de fer qui passent de l’état ferreux à l’état
ferrique :
Fe2+ Fe3+ + e-
COMPLEXE I
• Ces protéines fer-soufre, ont comme stoechiométrie Fe2S2 ou Fe4S4, permettent le
transport monoélectronique
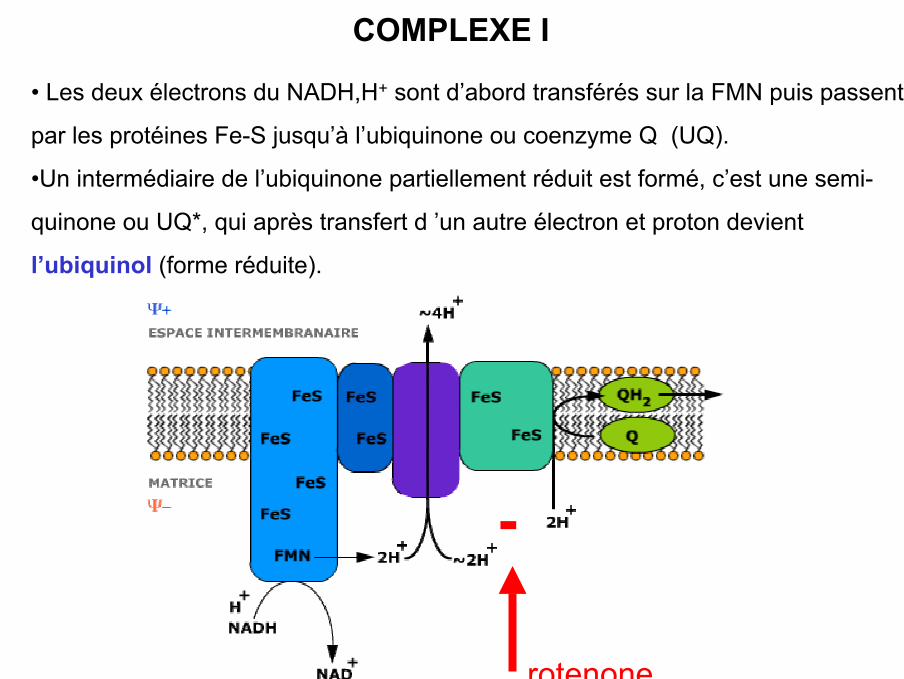
COMPLEXE I
• Les deux électrons du NADH,H+ sont d’abord transférés sur la FMN puis passent
par les protéines Fe-S jusqu’à l’ubiquinone ou coenzyme Q (UQ).
•Un intermédiaire de l’ubiquinone partiellement réduit est formé, c’est une semi-
quinone ou UQ*, qui après transfert d ’un autre électron et proton devient
l’ubiquinol (forme réduite).
-
rotenone
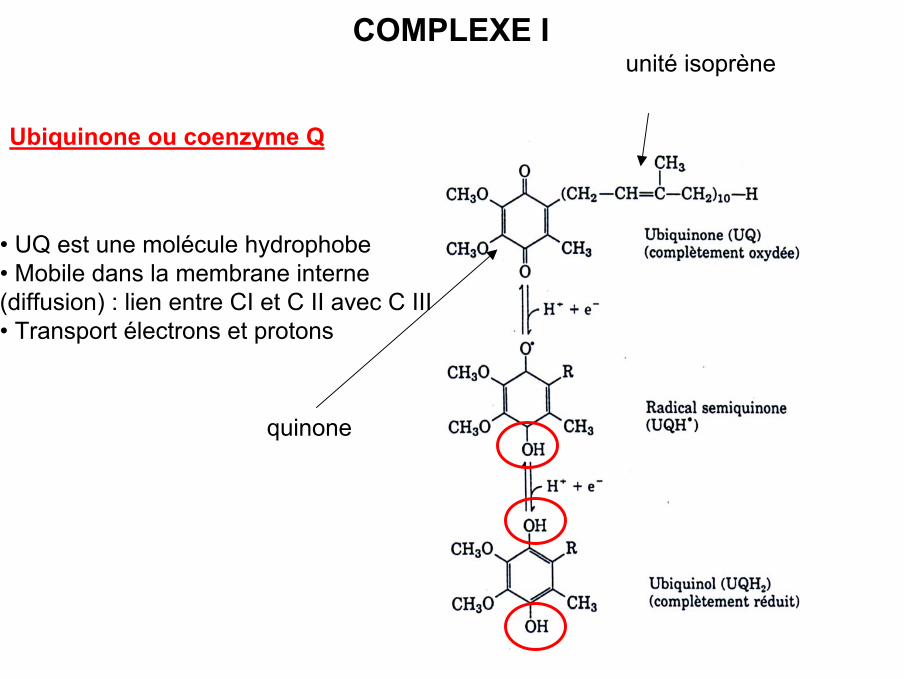
COMPLEXE I
Ubiquinone ou coenzyme Q
• UQ est une molécule hydrophobe • Mobile dans la membrane interne(diffusion) : lien entre CI et C II avec C III• Transport électrons et protons
unité isoprène
quinone
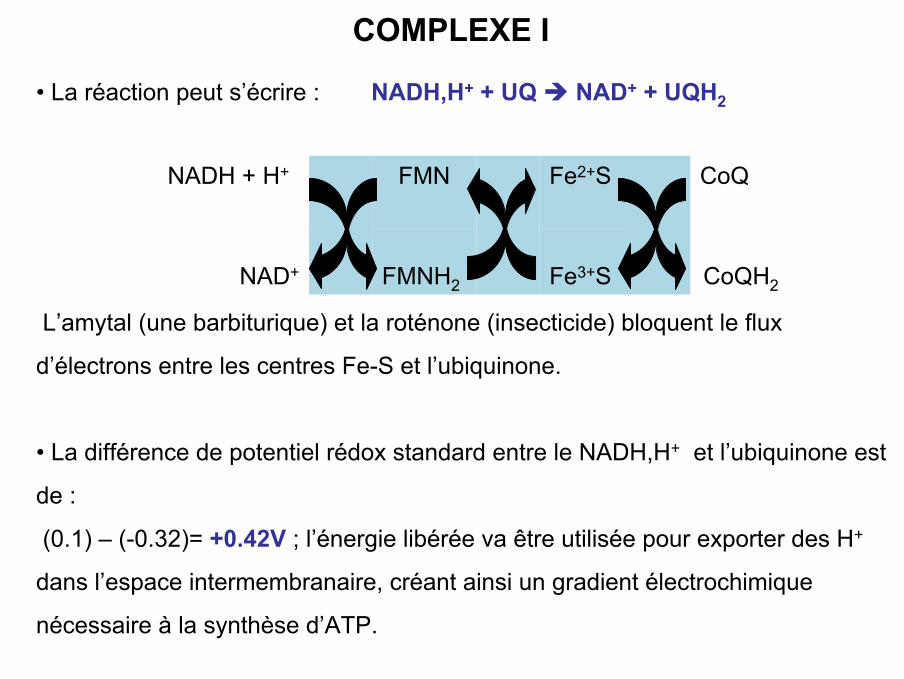
• La réaction peut s’écrire : NADH,H+ + UQ NAD+ + UQH2
L’amytal (une barbiturique) et la roténone (insecticide) bloquent le flux
d’électrons entre les centres Fe-S et l’ubiquinone.
• La différence de potentiel rédox standard entre le NADH,H+ et l’ubiquinone est
de :
(0.1) – (-0.32)= +0.42V ; l’énergie libérée va être utilisée pour exporter des H+
dans l’espace intermembranaire, créant ainsi un gradient électrochimique
nécessaire à la synthèse d’ATP.
COMPLEXE I
CoQH2Fe3+SFMNH2NAD+
CoQ
Fe2+S
FMN
NADH + H+

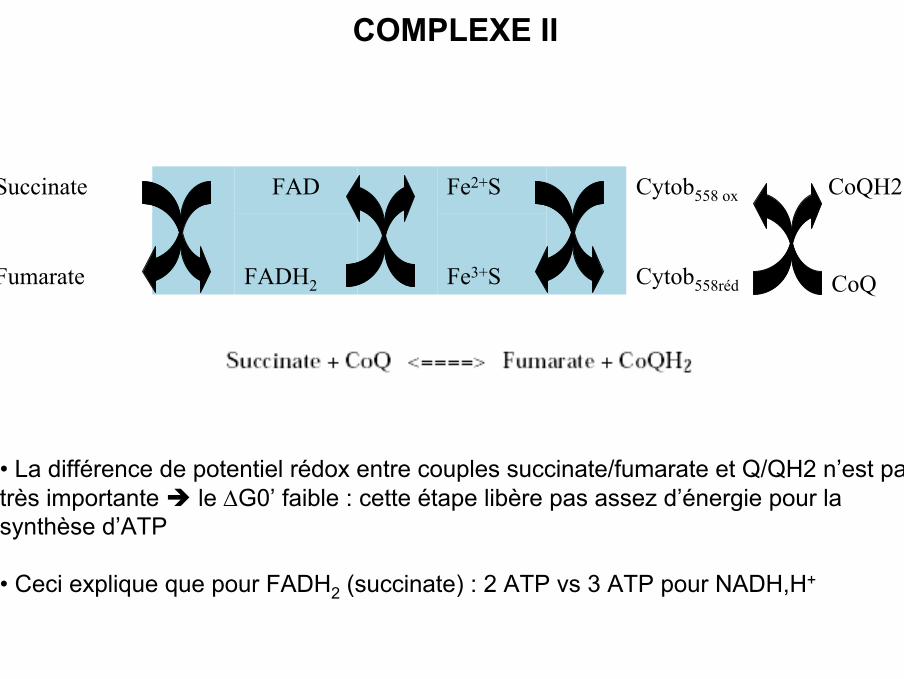
COMPLEXE II
• Complexe II ou succinate-ubiquinone réductase ou succinatedéshydrogénase
• Enzyme du cycle de KREBS
• Transfert d’électrons du succinate sur l’UQ.
• Composé de 8 polypeptides codés uniquement par l ’ADN nucléaire
• Ce complexe possède 4 centres Fe / S, un cytochrome b558 et l’une de ces sous-unités est une flavoprotéine (FAD) avec un centre Fe-S, qui va transférer lesélectrons sur une autre protéine Fe-S jusqu’à l’ubiquinone
• Le malonate est un inhibiteur compétitif de la succinate deshydrogénase.

COMPLEXE II
• Toutes les flavoprotéines n’ont pas le même potentiel rédox
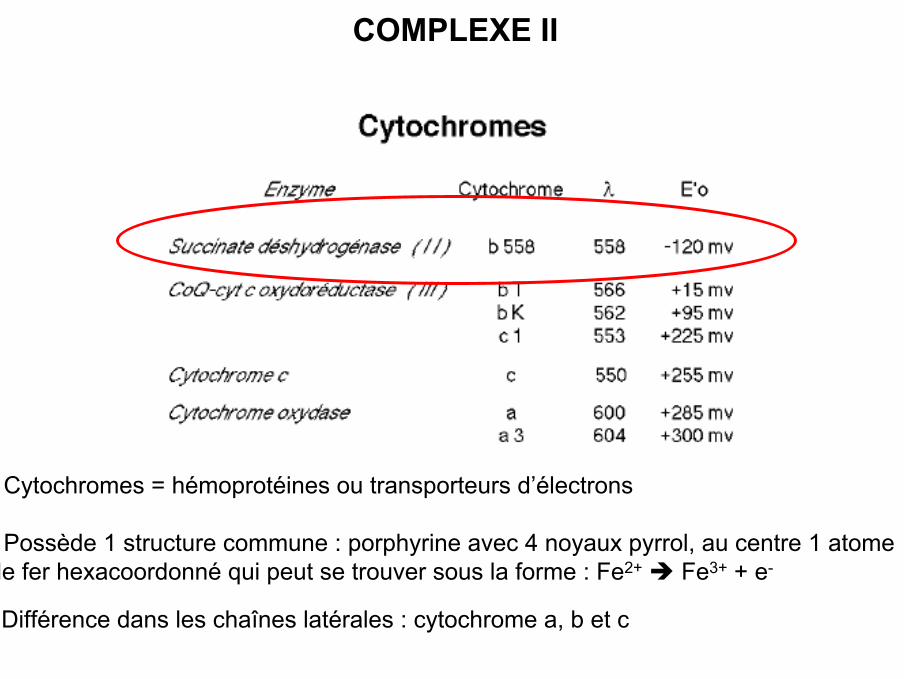
COMPLEXE II
• Cytochromes = hémoprotéines ou transporteurs d’électrons
• Possède 1 structure commune : porphyrine avec 4 noyaux pyrrol, au centre 1 atomede fer hexacoordonné qui peut se trouver sous la forme : Fe2+ Fe3+ + e-
• Différence dans les chaînes latérales : cytochrome a, b et c
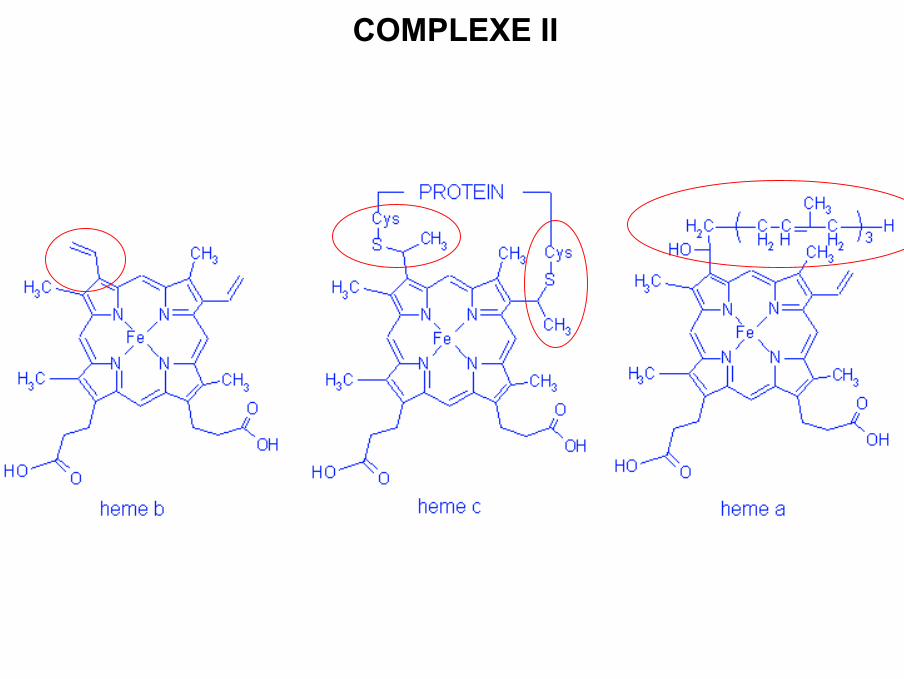
COMPLEXE II
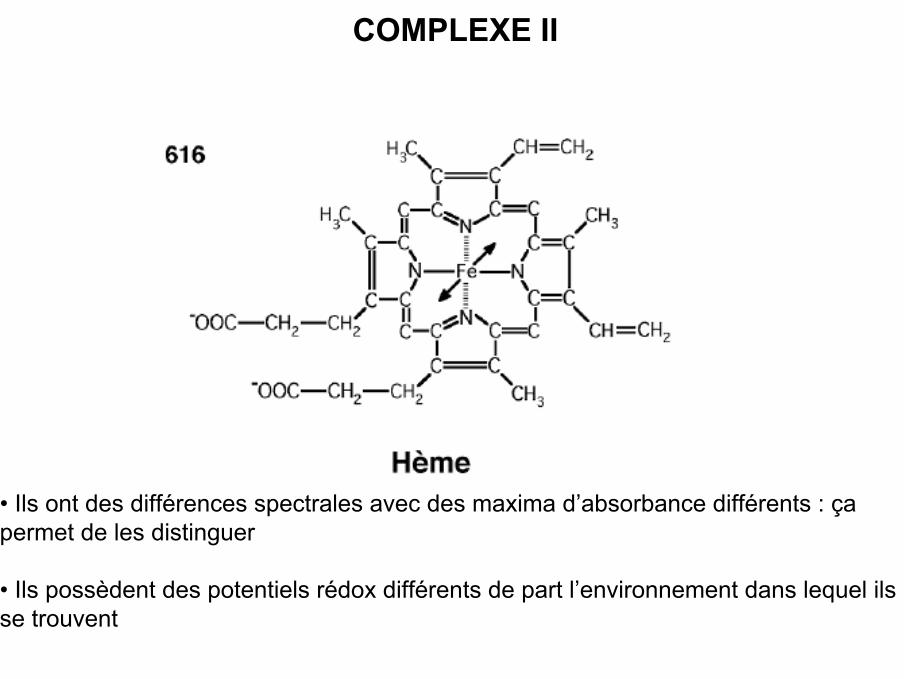
COMPLEXE II
• Ils ont des différences spectrales avec des maxima d’absorbance différents : çapermet de les distinguer
• Ils possèdent des potentiels rédox différents de part l’environnement dans lequel ilsse trouvent
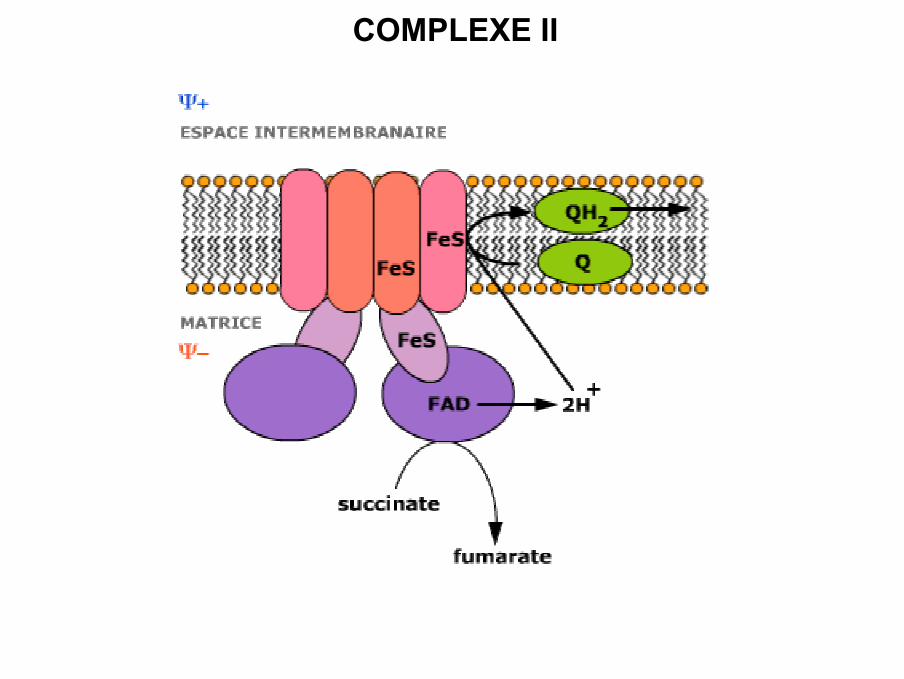
COMPLEXE II
COMPLEXE II
• La différence de potentiel rédox entre couples succinate/fumarate et Q/QH2 n’est pastrès importante le ∆G0’ faible : cette étape libère pas assez d’énergie pour lasynthèse d’ATP
• Ceci explique que pour FADH2 (succinate) : 2 ATP vs 3 ATP pour NADH,H+
Cytob558rédFe3+SFADH2Fumarate
Cytob558 ox
Fe2+S
FAD
Succinate CoQH2
CoQ

LES DESHYDROGENASES QUI TRANSMETTENTLES ELECTRONS SUR LE COENZYME Q
• Acyl-CoA déshydrogénase
• Glycérol phosphate déshydrogénase
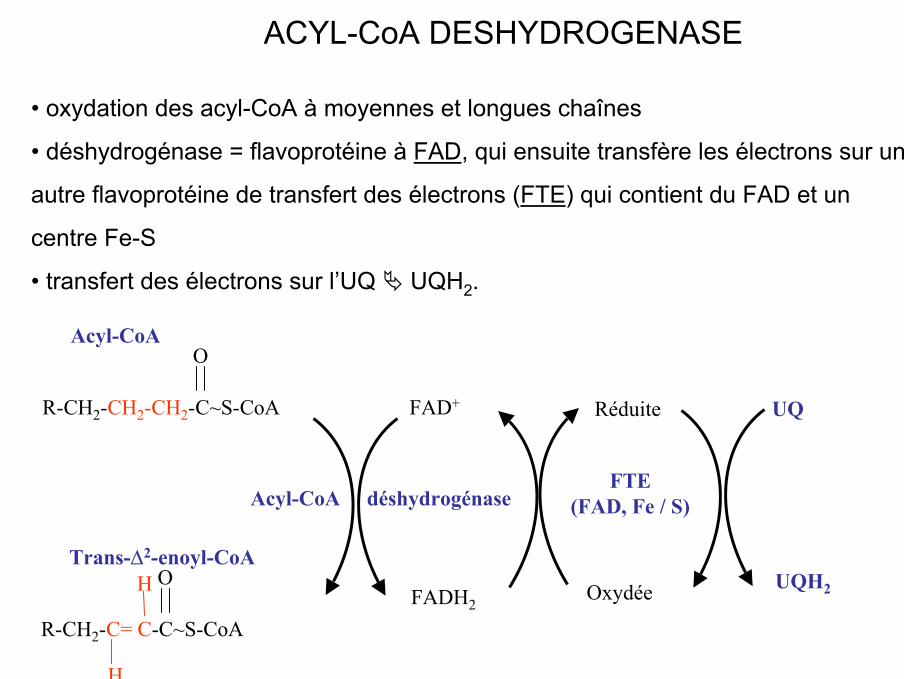
• oxydation des acyl-CoA à moyennes et longues chaînes
• déshydrogénase = flavoprotéine à FAD, qui ensuite transfère les électrons sur une
autre flavoprotéine de transfert des électrons (FTE) qui contient du FAD et un
centre Fe-S
• transfert des électrons sur l’UQ UQH2.
ACYL-CoA DESHYDROGENASE
R-CH2-CH2-CH2-C~S-CoA
OAcyl-CoA
R-CH2-C= C-C~S-CoA
OTrans-∆2-enoyl-CoA
H
H
Acyl-CoA déshydrogénase
FAD+
FADH2
FTE (FAD, Fe / S)
Oxydée
Réduite UQ
UQH2
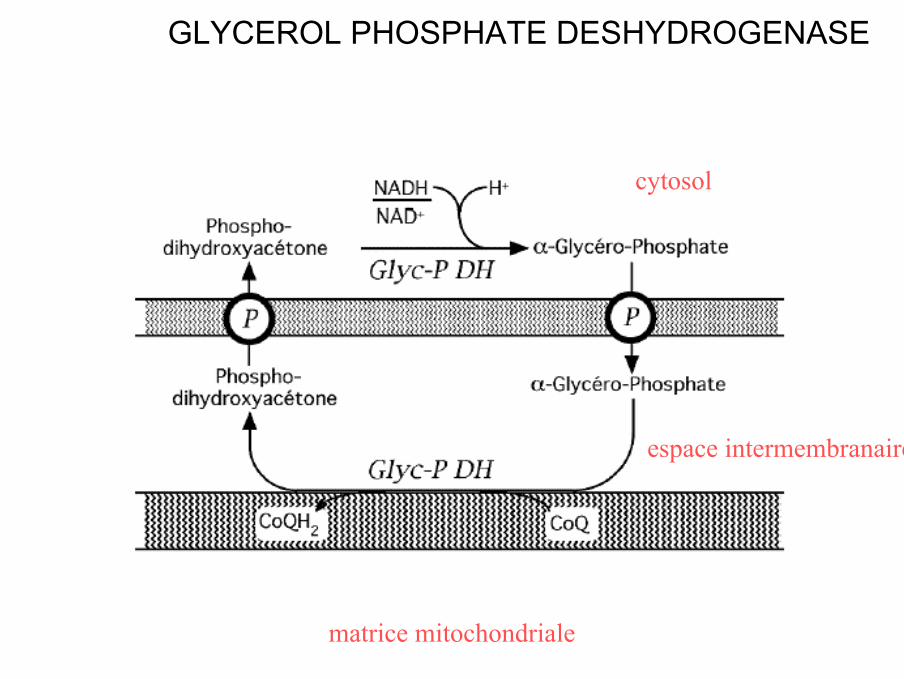
GLYCEROL PHOSPHATE DESHYDROGENASE
matrice mitochondriale
espace intermembranaire
cytosol
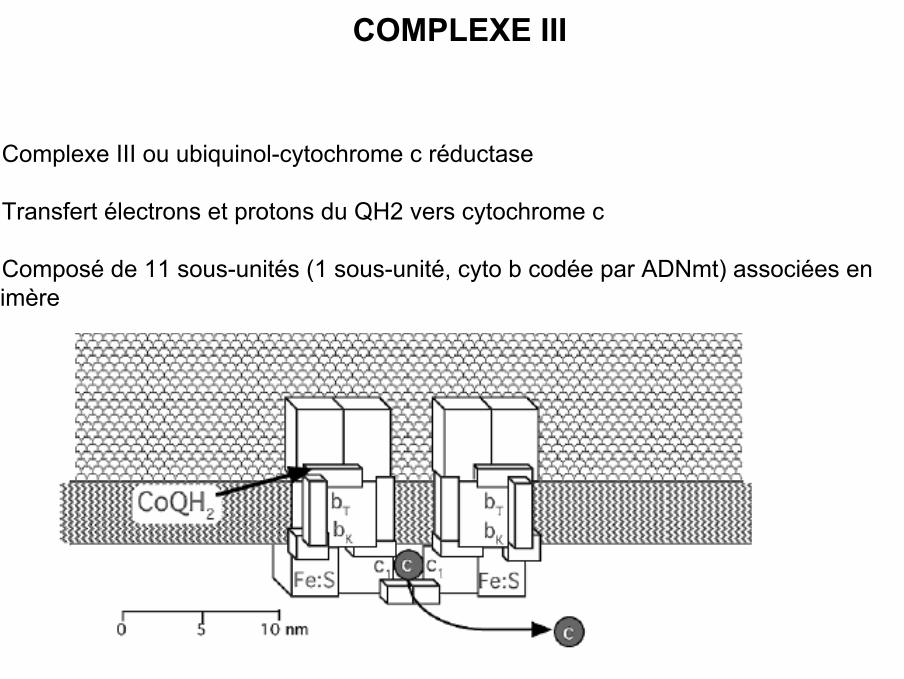
COMPLEXE III
• Complexe III ou ubiquinol-cytochrome c réductase
• Transfert électrons et protons du QH2 vers cytochrome c
• Composé de 11 sous-unités (1 sous-unité, cyto b codée par ADNmt) associées endimère

COMPLEXE III
• Chaque dimère contient :
- 2 cytochromes b
- 1 cytochrome c1
- 1 centre fer/soufre (centre Rieske 2 Fe / 2 S)
- 2 sites liaison pour Q : Qo (out) et Qi (inner, dans matrice)
• La réaction se déroule en plusieurs étapes avec transfert 1 électron à la fois :
- Fixation QH2 sur son site Q0- Transfert 1 électron sur centre fer / soufre- Transfert électron du centre fer / soufre sur cytochrome c1- Transfert cytochrome c1 sur cytochrome c oxydé (diffusion)- Deuxième électron du QH2 est transféré sur les cytochromes b puis sur Q oxydé
(site Qi)- Formation d’une semi-quinone sur Qi- 2ème molécule QH2 sur Qo et transfert idem sur cytochrome c- Sur Qi formation QH2 à partir semi-quinone et 2 protons de matrice
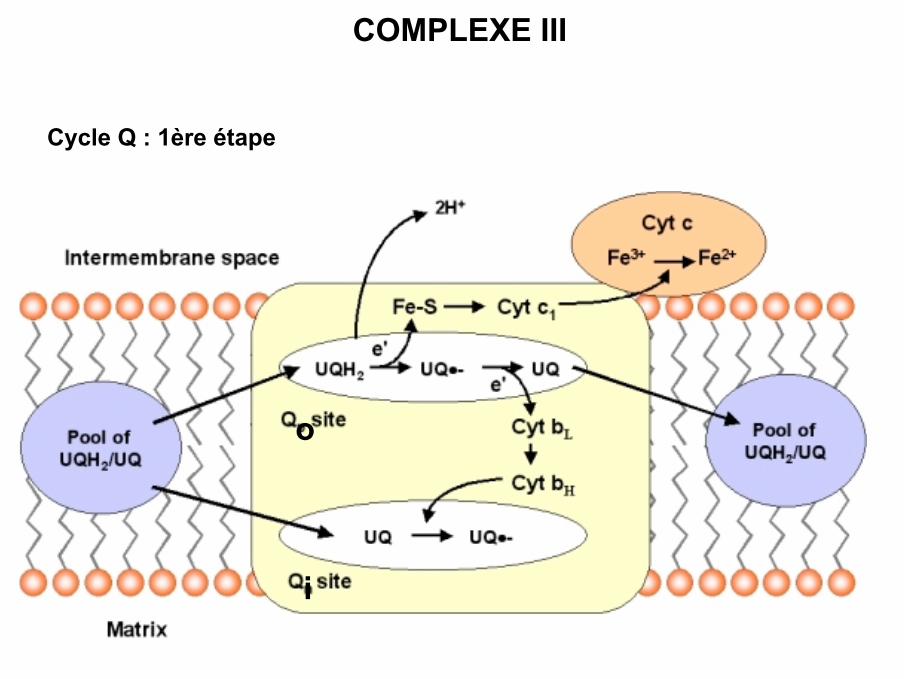
COMPLEXE III
Cycle Q : 1ère étape
i
o
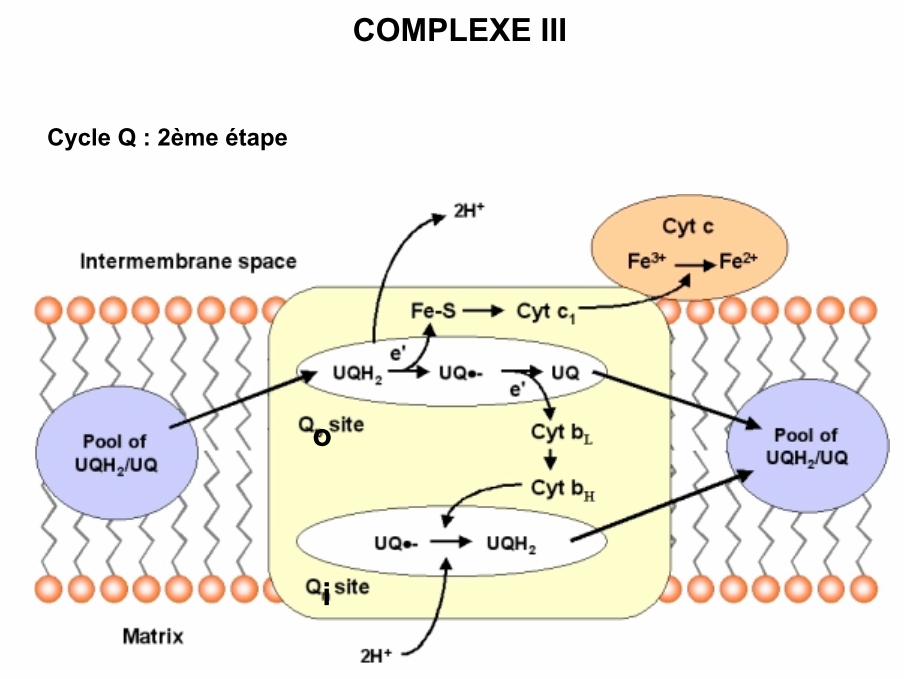
COMPLEXE III
Cycle Q : 2ème étape
i
o

COMPLEXE III

• UQH2 +2 cytochrome c (Fe3+) UQ + 2H+ + 2 cytochrome c (Fe2+)
• La différence de potentiel rédox standard entre le UQH2 et le cytochrome c est de :
+0.21 V libération d’énergie afin de contribuer à la formation du gradient de protons.
•L’antimycine A est un inhibiteur de ce complexe.
• Au total : 2 molécules QH2 oxydées, formation de Q et QH2, 2 cytochrome c réduit
COMPLEXE III
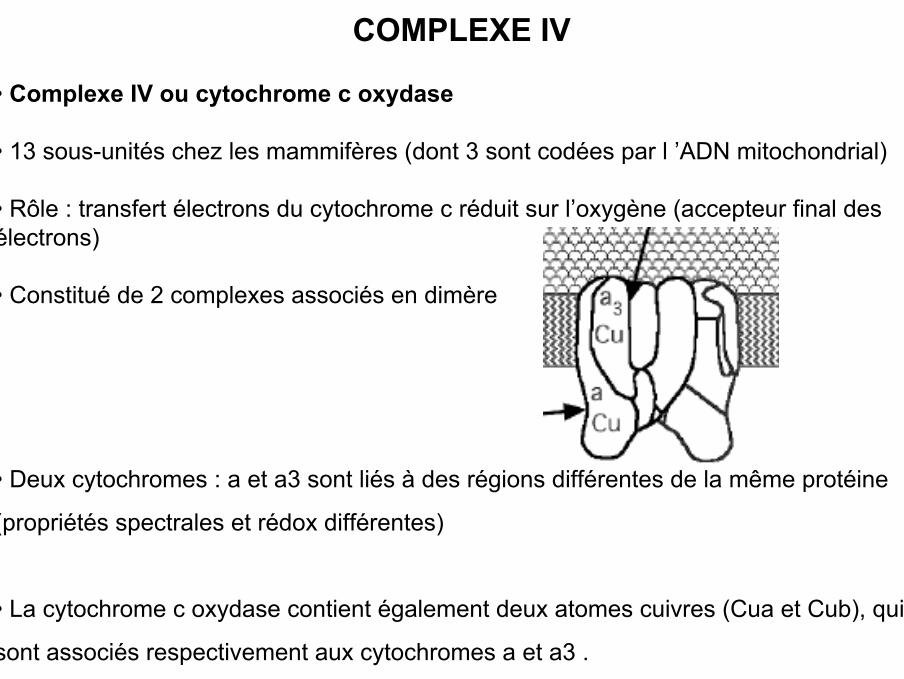
COMPLEXE IV
• Complexe IV ou cytochrome c oxydase
• 13 sous-unités chez les mammifères (dont 3 sont codées par l ’ADN mitochondrial)
• Rôle : transfert électrons du cytochrome c réduit sur l’oxygène (accepteur final desélectrons)
• Constitué de 2 complexes associés en dimère
• Deux cytochromes : a et a3 sont liés à des régions différentes de la même protéine
(propriétés spectrales et rédox différentes)
• La cytochrome c oxydase contient également deux atomes cuivres (Cua et Cub), qui
sont associés respectivement aux cytochromes a et a3 .
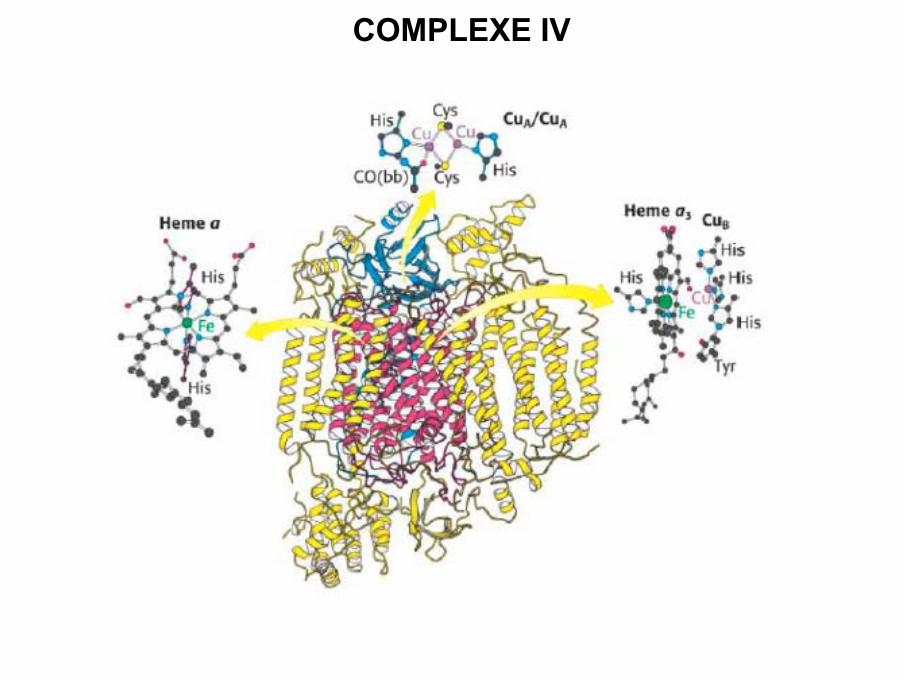
COMPLEXE IV

COMPLEXE IV
• Cytochrome c réduit par QH2(complexe III) se déplace jusqu’au complexe IV pour
céder son électron au premier atome de Cua
• Transfert de l’électron sur l’hème a
• L’électron est transféré sur le centre hème a3 / Cub
• Après capture de 2e- (Fe2+ / Cu+ ) transfert sur l ’oxygène (accepteur final des
électrons), et le centre devient (Fe3+- O22- / Cu2+ )
• 2 e- supplémentaires et 4 H+ vont permettre de produire 2 molécules d ’eau.
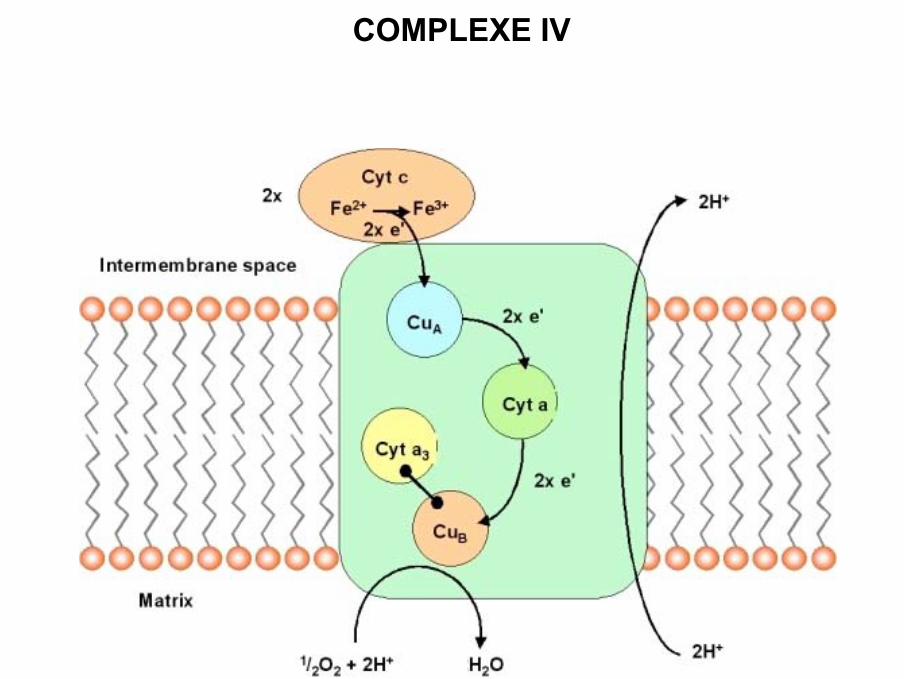
COMPLEXE IV

COMPLEXE IV

COMPLEXE IV
• La réaction peut s’écrire :
4 cytochrome c (Fe2+) + O2 + 4 H+ 4 cytochrome c (Fe3+) + 2 H2O
• La différence de potentiel rédox standard entre le cytochrome c et l’oxygène est de
+0.562 V
• Energie sert à pomper des protons de la matrice vers l’espace intermembranaire
• Le cyanure et le monoxyde de carbone sont des inhibiteurs du complexe IV en se
liant aux groupements hémiques.

• Synthèse d’ATP les travaux et les hypothèses introduits dans les années 60 parMitchell
• Transport d ’électrons et la synthèse d ’ATP sont couplés grâce à un gradient deprotons qui s ’établit de part et d’autre de la membrane interne mitochondriale
C’est la théorie chimio-osmotique.
SYNTHESE D’ATP
Entre NADH,H+ et O2 :• ∆G0’ = - 2 X 23.06 X 1.14 =-52.6kcal/mol ou 220 kJ/mol
• Pour la synthèse d’une molécule d’ATP ∆G0’ ~ + 8 kCal / mol
• Quel est le lien entre chaîne respiratoire et synthèse d’ATP ?

SYNTHESE D’ATP
La force proto-motrice
• Le transfert d’électrons le long de la chaîne respiratoire
libération d’énergie
passage de protons de la matrice vers l’espace intermembranaire contre gradient
accumulation dans l’espace intermembranaire car la membrane internemitochondriale est très imperméable
Attention : transfert de protons que si l’énergie libérée est assez importante (CI,III et IV)
Formation d’un gradient électrochimique (différence de potentiel membranaire ∆ψ+ différence de concentration en protons)= énergie potentielle pour la synthèsed’ATP grâce à l’ATP-synthase ou complexe V.
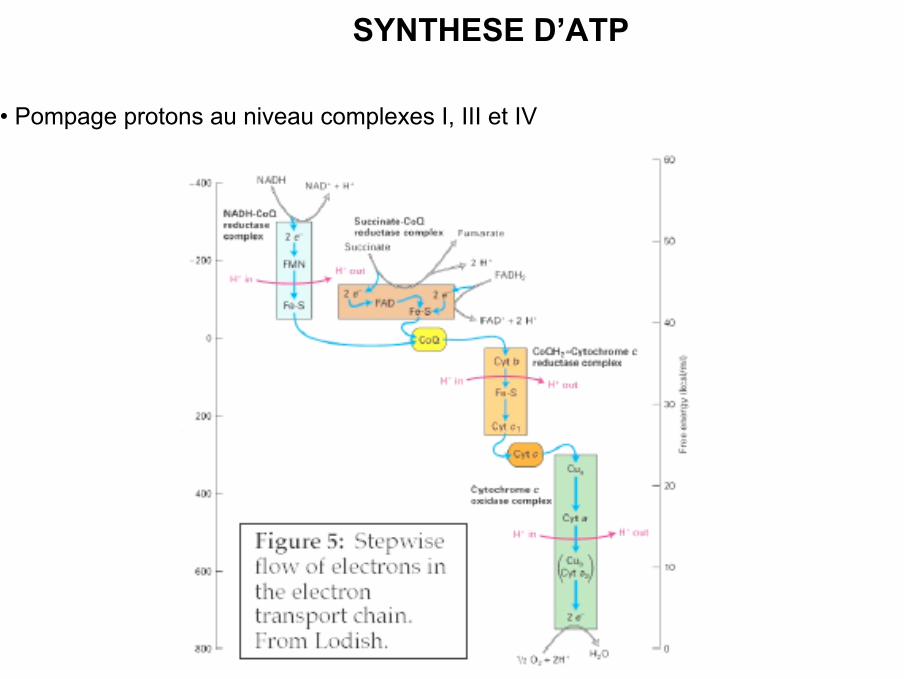
SYNTHESE D’ATP
• Pompage protons au niveau complexes I, III et IV
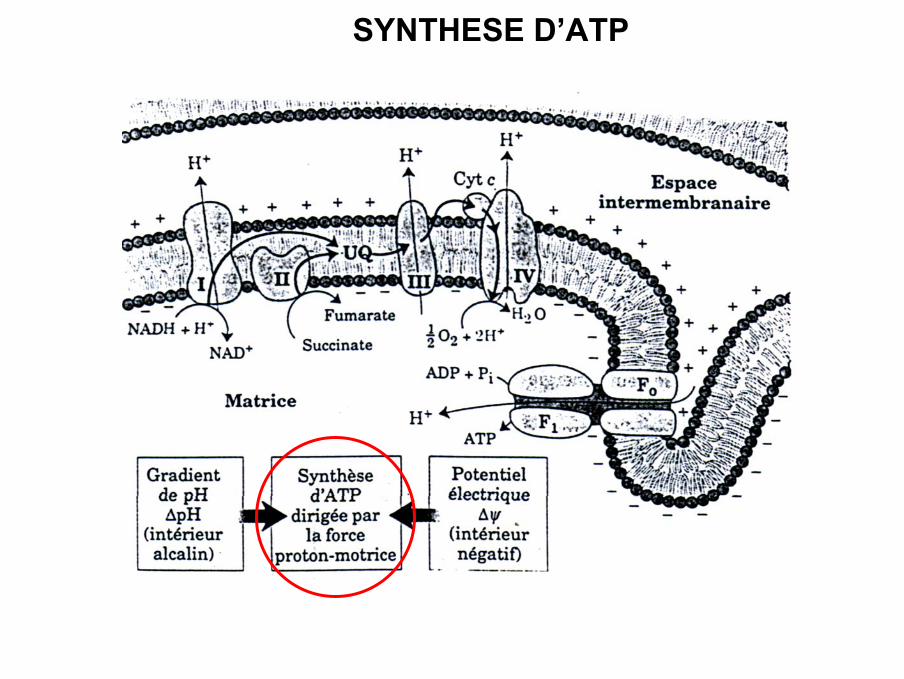
SYNTHESE D’ATP
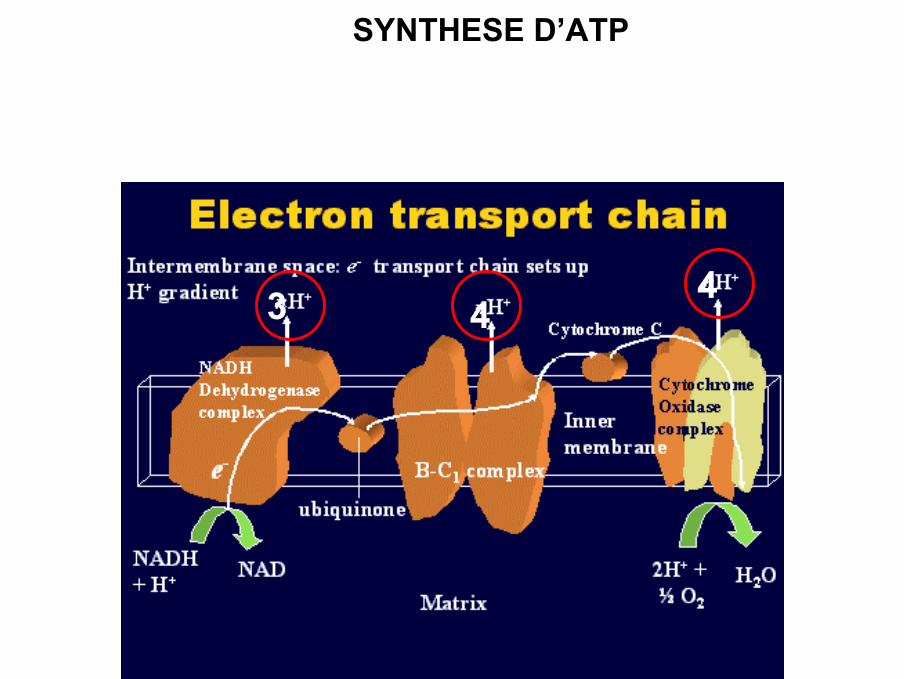
SYNTHESE D’ATP
3 44
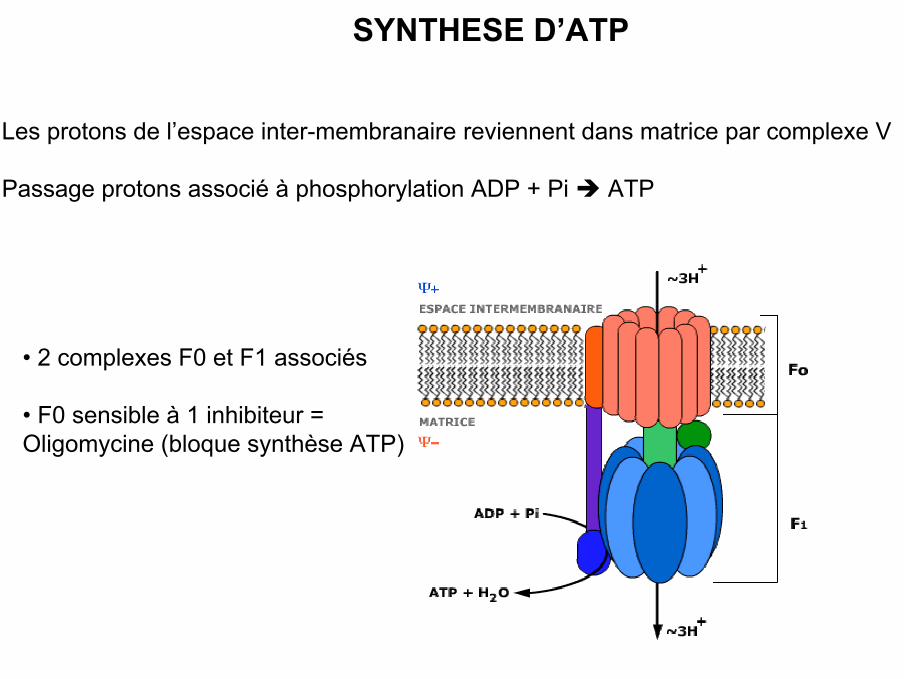
SYNTHESE D’ATP
• Les protons de l’espace inter-membranaire reviennent dans matrice par complexe V
• Passage protons associé à phosphorylation ADP + Pi ATP
• 2 complexes F0 et F1 associés
• F0 sensible à 1 inhibiteur =Oligomycine (bloque synthèse ATP)
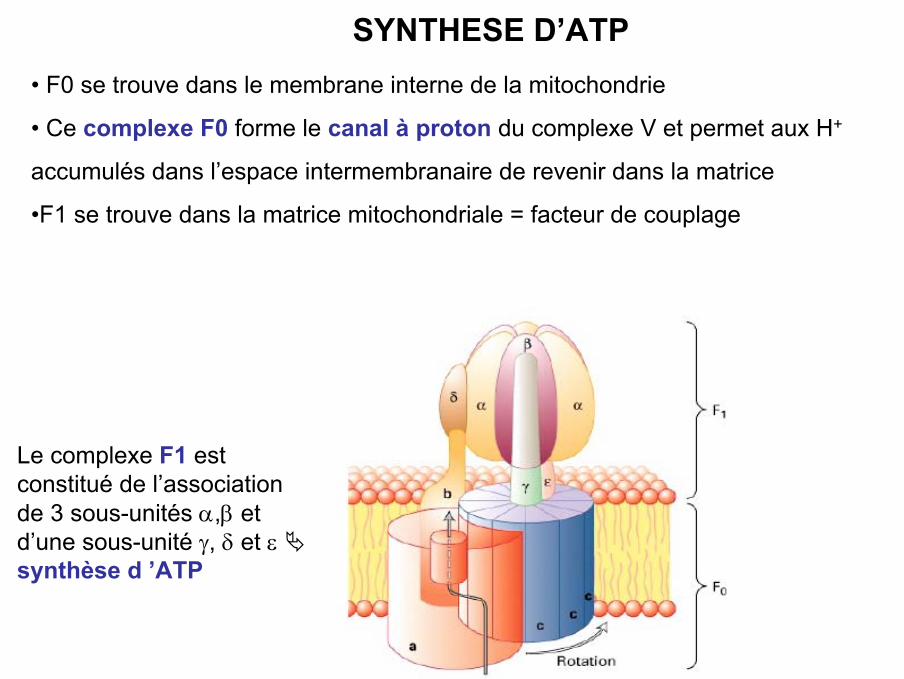
• F0 se trouve dans le membrane interne de la mitochondrie
• Ce complexe F0 forme le canal à proton du complexe V et permet aux H+
accumulés dans l’espace intermembranaire de revenir dans la matrice
•F1 se trouve dans la matrice mitochondriale = facteur de couplage
SYNTHESE D’ATP
Le complexe F1 estconstitué de l’associationde 3 sous-unités α,β etd’une sous-unité γ, δ et ε synthèse d ’ATP

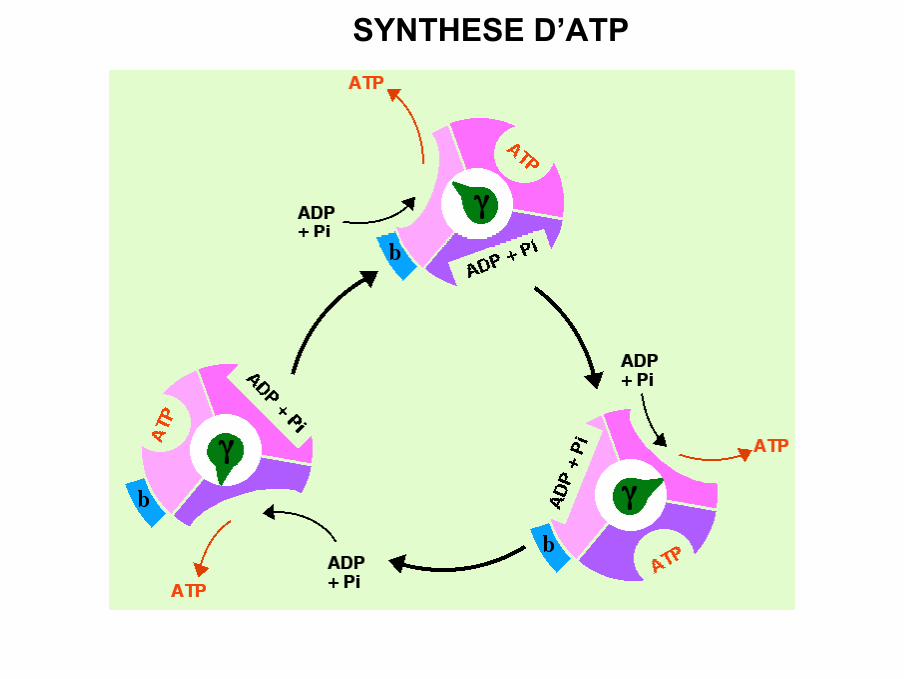
• Mécanisme de synthèse de l’ATP
L ’ATP synthase est capable de catalyser la formation d ’ATP à partir de l ’ADP et
du Pi même en absence de gradient de protons
Dans ces conditions, l ’ATP formé reste fixé au site actif et empêche toute nouvelle
réaction.
SYNTHESE D’ATP
• Modèle dit de changement de liaisonSur chaque paire de sous-unités αβ un site de liaison pour les nucléotides adényliques1 état de conformation pour chacun des 3 sites :- T = liaison forte- L = liaison faible- O = liaison très faible• Dans la conformation T: ATP est fortement lié à αβ, l’ADP et le Pi sont
faiblement liés à la conformation L.
La force protomotrice permettrait le départ de l’ATP, ce qui transformerait le
site T en site O, et le site L en site T.
SYNTHESE D’ATP

Synthase.mov
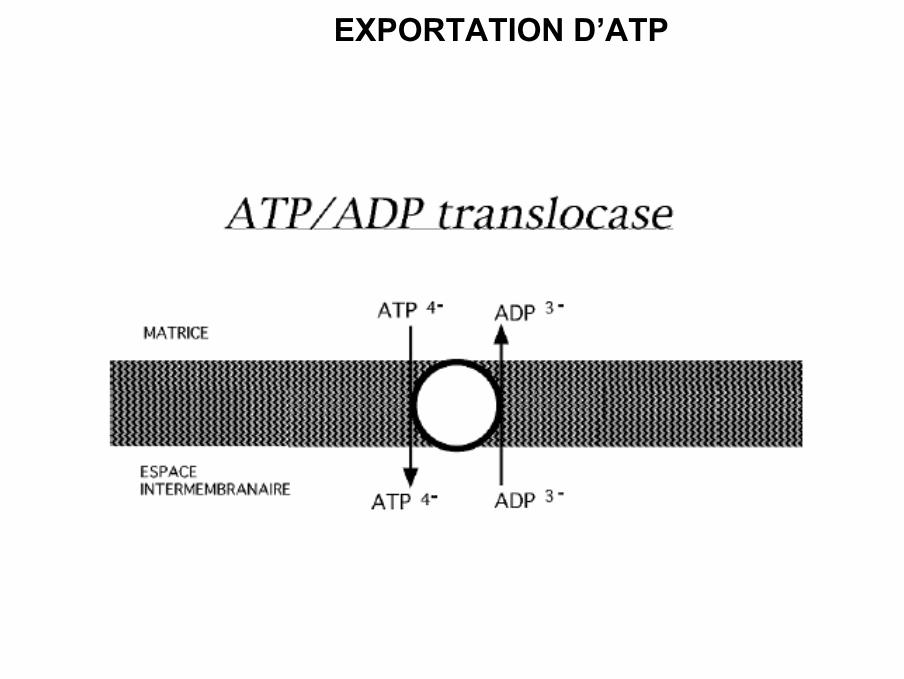
EXPORTATION D’ATP
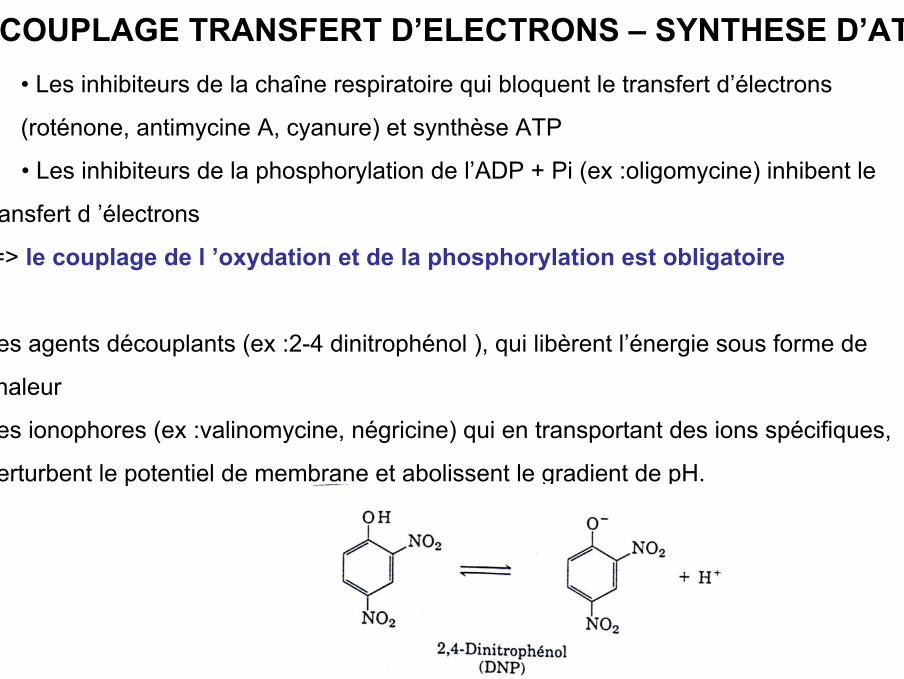
COUPLAGE TRANSFERT D’ELECTRONS – SYNTHESE D’ATP• Les inhibiteurs de la chaîne respiratoire qui bloquent le transfert d’électrons
(roténone, antimycine A, cyanure) et synthèse ATP
• Les inhibiteurs de la phosphorylation de l’ADP + Pi (ex :oligomycine) inhibent le
transfert d ’électrons
-=> le couplage de l ’oxydation et de la phosphorylation est obligatoire
-les agents découplants (ex :2-4 dinitrophénol ), qui libèrent l’énergie sous forme de
chaleur
-les ionophores (ex :valinomycine, négricine) qui en transportant des ions spécifiques,
perturbent le potentiel de membrane et abolissent le gradient de pH.
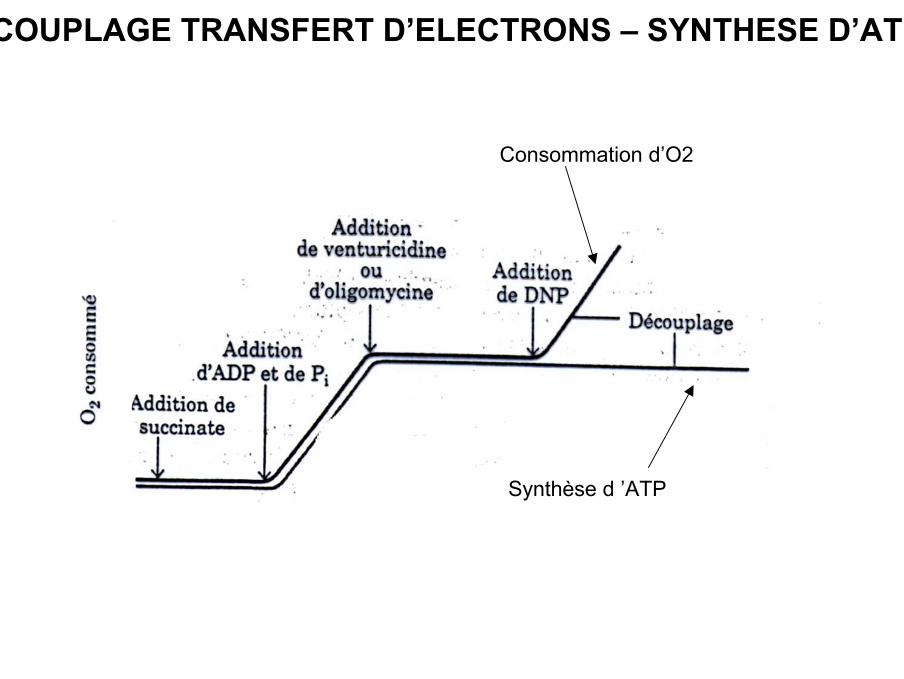
Synthèse d ’ATP
Consommation d’O2
COUPLAGE TRANSFERT D’ELECTRONS – SYNTHESE D’ATP

CONTROLE RESPIRATOIRE
• La respiration mitochondriale est soumise à une régulation étroite, et notamment par laconcentration en ADP (qui reflète l’état énergétique d’une cellule) substrat de laphosphorylation. Lorsque celle-ci augmente, elle augmente la respiration et entraîne lasynthèse d’ATP
• La respiration mitochondriale est soumise à une régulation étroite des concentrationsen cofacteurs réduits (NADH,H+ et FADH2)
• Un deuxième mécanisme de contrôle est réalisée sur la cytochrome c oxydase parl ’ATP et l ’ADP. Un rapport ATP/ADP élevé va inhiber de manière allostériquel ’enzyme
• Il existe dans la membrane interne de la mitochondrie un échangeur ADP-ATP ouadenyl nucléotide translocase (ANT). Il permet l’entrée de l’ADP pour être phosphoryléet la sortie de l’ATP dans le cytosol pour y être utilisé.

• A partir du NADH,H+: production de 3 molécules d’ATP pour ½ oxygène
consommé
Rapport P/O = 3
• A partir du succinate : production 2 molécules d’ATP pour ½ oxygène
consommé
Rapport P/O =2
• On peut calculer le rendement de la phosphorylation oxydative :
∆G0’ = - 220 kJ/mol pour le NADH,H+
∆G0’ = + 29.3 kJ/mol pour la synthèse d’ATP
rendement = (3 X 29.3 ) / 220 ~ 40 %
• Une partie de l’énergie est libérée sous forme de chaleur
• exemple : graisse brune du nouveau-né et UCP ou thermogenin
PHOSPHORYLATION OXYDATIVE
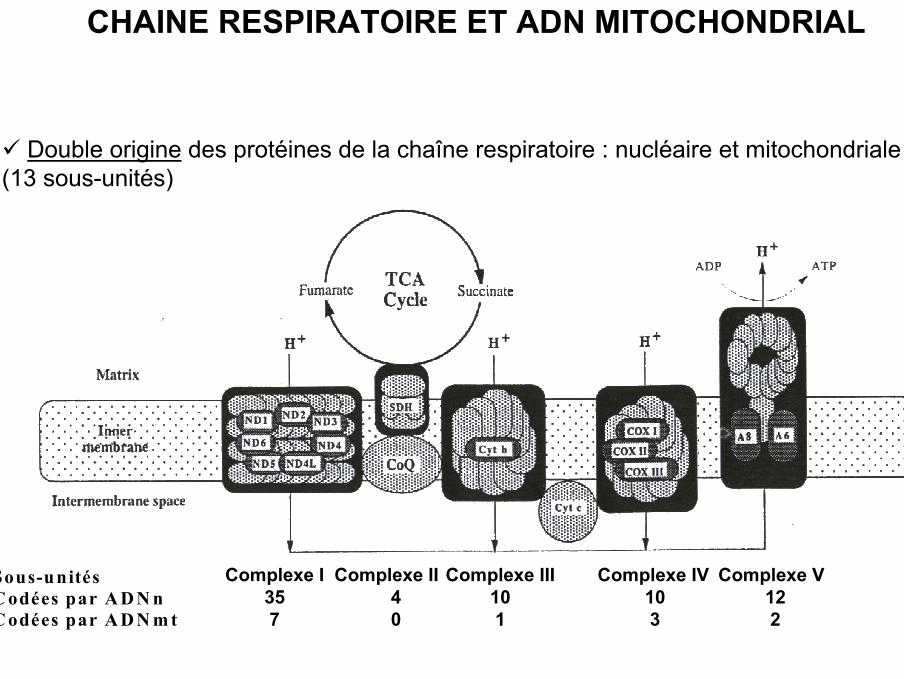
Sous-unitésC odées par A D N nC odées par A D N m t
Complexe I357
Complexe II40
Complexe III101
Complexe IV103
Complexe V122
Double origine des protéines de la chaîne respiratoire : nucléaire et mitochondriale(13 sous-unités)
CHAINE RESPIRATOIRE ET ADN MITOCHONDRIAL
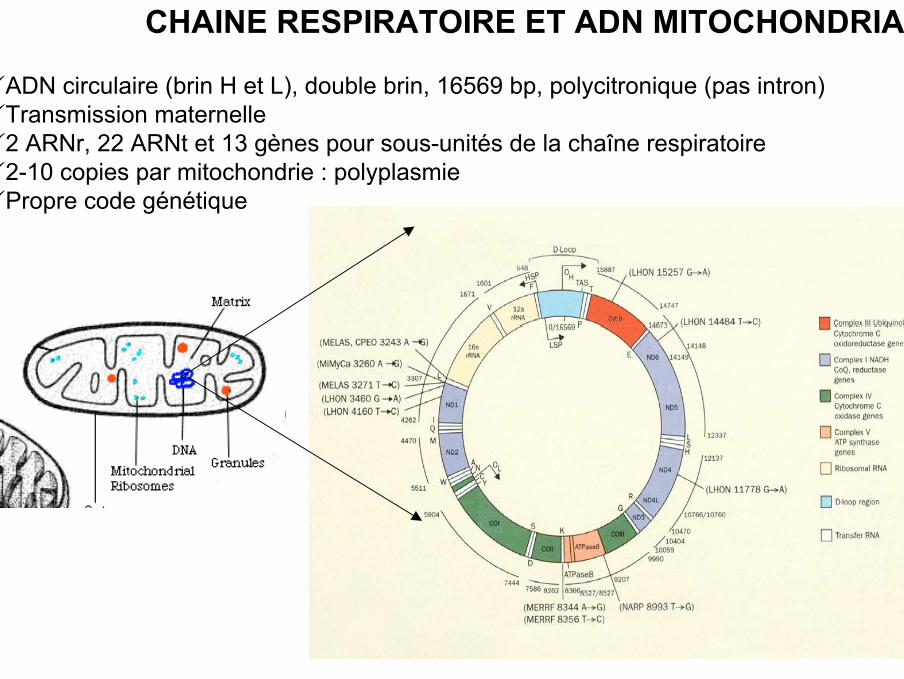
ADN circulaire (brin H et L), double brin, 16569 bp, polycitronique (pas intron)Transmission maternelle2 ARNr, 22 ARNt et 13 gènes pour sous-unités de la chaîne respiratoire 2-10 copies par mitochondrie : polyplasmiePropre code génétique
CHAINE RESPIRATOIRE ET ADN MITOCHONDRIAL

• On connaît plusieurs mutations ou délétions qui peuvent toucher cet ADNmitochondrial Ceci a pour conséquence une réduction du transfert d’électrons
• ces mitochondries ne peuvent subvenir aux besoins énergétiques des cellules. Il enrésulte également une accumulation des cofacteurs réduits NADH,H+, qui pour ne pasbloquer la glycolyse sont transformés en présence de pyruvate en acide lactique
• Ces pathologies ont comme caractéristiques d’être transmises uniquement par lesmères (les mitochondries proviennent de l’ovule) et se manifestent par des atteintesnerveuses (neuropathie optique de Leber, le syndrome MELAS) et musculaires.
CHAINE RESPIRATOIRE ET PATHOLOGIES

POINTS A RETENIR…..

POINTS A RETENIR…..
• Le rôle de chacun des complexes (ex: CIII va transférer électron du QH2 surcytochrome c)
• Les transporteurs mobiles d’électrons
• Théorie chimio-osmotique et relations transferts d’électrons et synthèsed’ATP





























