Embed Size (px)
Citation preview

Mécanisme d’action hépatique de la metformine dans le diabète de type 2Hepatic mechanism of metformin action in type 2 diabetes mellitus
M. Foretz 1,2, B. Viollet 1,21 Institut Cochin, Université Paris - Descartes, CNRS (UMR 8104), Département endocrinologie, métabolisme et cancer, Paris.2 Inserm, U567, Paris.
lisée avec succès depuis plus de 40 ans dans le traitement du diabète de type 2. La metformine est considérée comme le traitement de première intention chez la plupart des diabétiques de type 2, en particulier chez les patients en surpoids ou obèses, pour son efficacité et pour sa faible capacité à provoquer des hypogly-cémies. La metformine exerce des effets pléiotropiques bénéfiques sur le méta-bolisme énergétique et le système car-diovasculaire, ces deux actions pouvant être liées. Le foie joue un rôle central dans les effets métaboliques de la metformine. La principale action de la metformine est l’amélioration de l’hyperglycémie par inhi-bition de la gluconéogenèse hépatique. De manière secondaire, la metformine augmente la sensibilité à l’insuline dans le foie et dans les tissus périphériques. La metformine est également utilisée dans le traitement de la stéatose hépati-que non alcoolique, une conséquence de l’accumulation hépatique de lipides fré-quemment associée au diabète de type 2. Des travaux récents permettent de mieux comprendre les mécanismes moléculaires de la metformine dans le contrôle du méta-bolisme énergétique dans le foie au cours du traitement du diabète de type 2.
Le foie est l’organe cible de la metformine
Du fait de son administration orale, la metformine est absorbée par l’intestin et ses concentrations au niveau de la veine porte et au niveau hépatique sont élevées.
Correspondance :
Marc ForetzInstitut CochinDépartement endocrinologie, métabolisme et cancer24, rue du Faubourg Saint-Jacques75674 Paris cedex [email protected]
RésuméLa metformine est l’antidiabétique oral le plus utilisé pour réduire l’hyperglycémie au cours du diabète de type 2. La metformine agit principalement sur le foie en diminuant la synthèse de novo de glucose (gluconéogenèse). La metformine est également capa-ble d’améliorer la stéatose hépatique en inhibant la synthèse lipides et en augmentant l’oxydation des acides gras. Des observations récentes suggèrent que la metformine exercerait ses effets bénéfiques sur le métabolisme en diminuant la charge énergétique intracellulaire et en activant la protéine kinase activée par l’AMP (AMPK), un régulateur clé du métabolisme énergétique.
Mots-clés : AMPK – complexe I de la chaîne respiratoire – diabète de type 2 – gluconéogenèse hépatique – metformine – stéatose hépatique.
SummaryMetformin is the most frequently oral antidiabetic used to reduce hyperglycemia in type 2 diabetes. Metformin mainly acts on liver to reduce de novo glucose synthesis (gluconeogenesis). Metformin is also able to ameliorate hepatic steatosis by inhibiting lipid synthesis and increasing fatty acid oxidation. Recent observations suggest that metformin would exert its beneficial effects on metabolism through the reduction of intracellular energy charge and activation of AMP-activated protein kinase (AMPK), a key regulator of energy metabolism.
Key-words: AMPK – mitochondrial respiratory chain complex 1 – hepatic gluconeogenesis – metformin – hepatic steatosis – type 2 diabetes.
Introduction
La metformine (1,1-diméthylbiguanide hydrochloride) est un antidiabétique oral de la famille des biguanides. Les bigua-nides tirent leur origine du galéga officinal (Galega officinalis), une plante herbacée utilisée depuis le Moyen Âge pour réduire les symptômes du diabète. La metformine a été mise sur le marché pour la première fois en 1979 en France sous l’appellation de Glucophage®. La metformine est uti-© 2009 - Elsevier Masson SAS - Tous droits réservés.
Médecine des maladies Métaboliques - Janvier 2009 - Vol. 3 - N°1
48 Pour la science

Mécanisme d’action hépatique de la metformine dans le diabète de type 2 49
Médecine des maladies Métaboliques - Janvier 2009 - Vol. 3 - N°1
Des études récentes ont rapporté que le transport intracellulaire de la metformine est médié par le transporteur de cations organiques OCT1 (Organic cation trans-porter 1) [1]. Chez la souris, la délétion de OCT1 dans l’hépatocyte entraîne une diminution importante de l’accumulation de metformine dans le foie, ainsi qu’une altération de l’activation de l’AMPK (AMP-activated protein kinase) et de l’inhibition de la gluconéogenèse [2]. Chez l’homme, la présence de polymorphisme au niveau du gène OCT1 affecte l’effet de la met-formine au cours de tests de tolérance au glucose [2]. L’expression de OCT1 est élevée dans l’intestin, le foie et le rein, mais peu importante dans les autres tis-sus, en particulier le muscle, le cœur et le tissu adipeux. Dans le foie et le rein, OCT1 participe respectivement à l’ab-sorption et à l’excrétion de la metformine. L’expression importante du transporteur OCT1 dans l’intestin et le foie entraîne une accumulation plus marquée de la metformine dans ces deux organes [3]. L’accumulation de la metformine dans ces tissus est accentuée par le fait que la metformine n’est pas métabolisée par l’organisme, mais directement excrétée par la voie urinaire avec une demi-vie d’environ 6 heures. Il en résulte que le foie se trouve dans une situation idéale pour médier les effets de la metformine.
Cibles moléculaires de la metformine
Bien que la metformine soit largement utili-sée depuis des décennies, son mécanisme d’action cellulaire est resté longtemps obscur. Un certain nombre de mécanis-mes moléculaires ont été proposés pour expliquer les effets hépatiques de la met-formine dans l’inhibition de la gluconéo-genèse. Récemment, plusieurs études ont démontré, d’une part que la metformine inhibe modérément le complexe I (NADH : ubiquinone oxidoreductase) de la chaîne respiratoire et, d’autre part, active la pro-téine kinase activée par l’AMP (AMPK). L’AMPK est une sérine thréonine kinase extrêmement conservée chez les euca-ryotes [4]. L’AMPK est un hétérotrimère composé d’une sous-unité catalytique α et de deux sous-unités régulatrices β et γ. L’AMPK est activée de manière allostéri-
que par l’AMP et par phosphorylation de la thréonine 172 de la sous-unité catalytique α par une AMPK kinase, en particulier le gène suppresseur de tumeur LKB1 (ou serine/threonine kinase 11) [4] (figure 1). LKB1 est une protéine kinase dont des mutations ont été identifiées dans le syn-drome de Peutz-Jeghers, caractérisé par le développement de polypes intestinaux et d’adénocarcinomes pulmonaires. En association avec les protéines régulatri-ces STRAD et MO25, LKB1 joue le rôle d’un suppresseur de tumeur en contrôlant la prolifération et la polarité cellulaire [5]. L’AMPK est un senseur métabolique sen-sible au niveau énergétique de la cellule, qui permet d’ajuster en permanence, et au plus près, les besoins et disponibili-tés énergétiques de la cellule. L’AMPK est activée par une diminution du rap-port intracellulaire ATP/AMP suite à un
stress métabolique comme l’exercice, le jeûne ou l’hypoxie (figure 1). L’AMPK est également activée dans le foie par des adipokines sécrétées par le tissu adipeux, notamment l’adiponectine et la résistine [6]. Une fois activée, l’AMPK inhibe les voies métaboliques anaboliques consommatrices d’ATP et active les voies cataboliques productrices d’ATP afin de restaurer le rapport ATP/AMP intracellu-laires [4] (figure 1). Dans le foie, l’AMPK augmente l’oxydation des acides gras et inhibe la lipogenèse et la gluconéogenèse. En 2001, Zhou et al. ont montré, pour la première fois, que la metformine active l’AMPK dans des hépatocytes en culture et dans le muscle squelettique et ont pro-posé que l’activation de l’AMPK fournit une explication unifiée des effets pléiotro-piques bénéfiques de la metformine, sans toutefois expliquer le mécanisme de cette
Figure 1 : L’AMPK (AMP-activated protein kinase) est un senseur du statut énergétique intracellulaire.LKB1 : serine/threonine kinase 11.

Pour la science
Médecine des maladies Métaboliques - Janvier 2009 - Vol. 3 - N°1
50 Pour la science
activation [7]. L’AMPK n’est pas une cible directe de la metformine puisqu’elle est incapable de stimuler l’activité de l’AMPK ou d’augmenter sa phosphorylation par LKB1 in vitro [8]. Cependant, LKB1 est nécessaire à l’activation de l’AMPK par la metformine dans le foie [9]. Deux équi-pes ont montré indépendamment que la metformine induit une inhibition modérée et spécifique du complexe I de la chaîne respiratoire mitochondriale [10, 11]. Il semble maintenant clair que l’activa-tion de l’AMPK par la metformine est la conséquence de l’inhibition du complexe I (figure 2). Nous avons récemment mis en évidence que l’activation de l’AMPK par la metformine dans des hépatocytes en culture primaire est corrélée avec une diminution des niveaux intracellulaires en ATP [12, 13]. De plus, nous avons observé que l’inhibition du complexe I de la chaîne respiratoire par la metformine n’est pas altérée dans des hépatocytes de souris délétées des sous-unités catalytiques α1 et α2 [12]. Ces résultats démontrent que l’effet de la metformine sur la mitochondrie n’est pas une conséquence de l’activa-tion de l’AMPK et que le complexe I de la chaîne respiratoire est probablement la cible primaire de la metformine. Ainsi, ces observations suggèrent que l’activation de l’AMPK par la metformine résulte d’une modification du statut énergétique cellu-laire (figure 2). Comme nous le détaillerons ci-dessous, l’identification du complexe I et de l’AMPK comme cibles de la metfor-mine ont permis de mieux comprendre son mécanisme d’action dans le contrôle du métabolisme glucidique et lipidique dans le foie de sujets diabétiques de type 2.
Régulation de la production hépatique de glucose par la metformine
La metformine améliore l’hyperglycémie chez les diabétiques de type 2, princi-palement en diminuant la production de glucose hépatique et, dans une moindre mesure, en réduisant l’absorption gastro-intestinale du glucose et en augmentant l’utilisation du glucose par les tissus périphériques [14]. La diminution de la production hépatique de glucose par la metformine est attribuée à une inhibition de la gluconéogenèse [15]. Plusieurs
mécanismes ont été proposés pour expliquer cet effet métabolique, incluant l’amélioration du signal insuline [16], des changements d’activité d’enzymes clés [17-19], ou encore une diminution du transport hépatique des substrats glu-conéoformateurs [20]. Plus récemment, l’activation de l’AMPK par la metformine a été proposée pour expliquer l’inhibition de la production de glucose dans des hépatocytes via l’inhibition de l’expres-sion des gènes de la gluconéogenèse [7]. En effet, l’activation de l’AMPK est connue pour inhiber fortement la produc-tion hépatique de glucose et l’expression des gènes de la glucose 6-phosphatase (G6Pase) et de la phosphoénolpyruvate carboxykinase (PEPCK), deux enzy-mes clés de la gluconéogenèse [7, 21]. Récemment, le co-activateur trans-criptionnel TORC2 (Transcriptional co- activator transducer of regulated CREB activity 2) a été identifié comme un acteur central dans l’activation du programme gluconéogénique [22]. TORC2 interagit
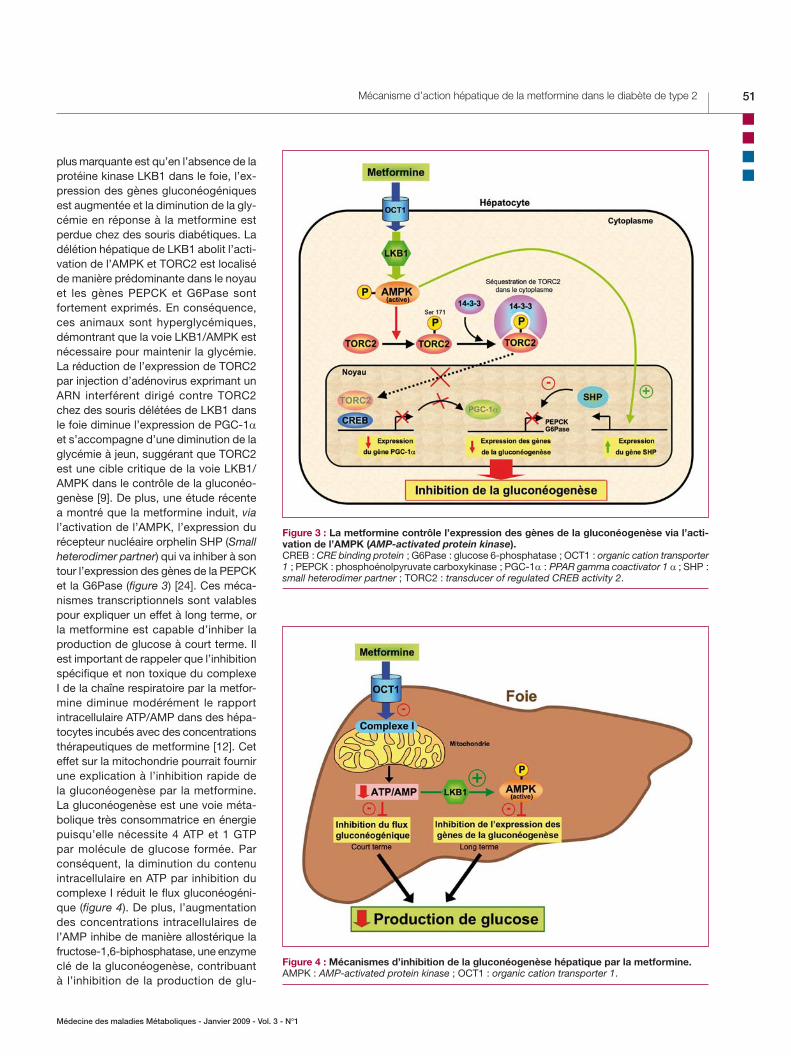
avec le facteur de transcription CREB (CRE binding protein) pour induire l’ex-pression d’un autre facteur de transcrip-tion PGC-1α (PPAR gamma coactivator 1 alpha) qui, à son tour, active l’expression des gènes PEPCK et G6Pase (figure 3). TORC2 est régulé par plusieurs voies de signalisation en réponse à des variations des niveaux d’insuline et du glucagon et au statut énergétique intracellulaire. La phosphorylation de TORC2 sur la sérine 171 lui permet de se lier à la protéine 14-3-3 et d’être séquestré à l’extérieur du noyau, entraînant une inhibition de l’expression des gènes de la gluconéo-genèse (figure 3). La kinase responsable de la phosphorylation de la serine 171 a été initialement identifiée comme SIK2 (Salt-inducible kinase 2), une protéine kinase phylogéniquement très proche de l’AMPK [23]. Récemment, l’AMPK a été montrée capable de phosphoryler TORC2 et de réguler sa translocation cytoplas-mique dans des hépatocytes en culture primaire (figure 3) [22]. La découverte la
Figure 2 : Cibles moléculaires de la metformine dans l’hépatocyte.AMPK : AMP-activated protein kinase ; OCT1 : organic cation transporter 1.
Mécanisme d’action hépatique de la metformine dans le diabète de type 2 51
Médecine des maladies Métaboliques - Janvier 2009 - Vol. 3 - N°1
plus marquante est qu’en l’absence de la protéine kinase LKB1 dans le foie, l’ex-pression des gènes gluconéogéniques est augmentée et la diminution de la gly-cémie en réponse à la metformine est perdue chez des souris diabétiques. La délétion hépatique de LKB1 abolit l’acti-vation de l’AMPK et TORC2 est localisé de manière prédominante dans le noyau et les gènes PEPCK et G6Pase sont fortement exprimés. En conséquence, ces animaux sont hyperglycémiques, démontrant que la voie LKB1/AMPK est nécessaire pour maintenir la glycémie. La réduction de l’expression de TORC2 par injection d’adénovirus exprimant un ARN interférent dirigé contre TORC2 chez des souris délétées de LKB1 dans le foie diminue l’expression de PGC-1α et s’accompagne d’une diminution de la glycémie à jeun, suggérant que TORC2 est une cible critique de la voie LKB1/AMPK dans le contrôle de la gluconéo-genèse [9]. De plus, une étude récente a montré que la metformine induit, via l’activation de l’AMPK, l’expression du récepteur nucléaire orphelin SHP (Small heterodimer partner) qui va inhiber à son tour l’expression des gènes de la PEPCK et la G6Pase (figure 3) [24]. Ces méca-nismes transcriptionnels sont valables pour expliquer un effet à long terme, or la metformine est capable d’inhiber la production de glucose à court terme. Il est important de rappeler que l’inhibition spécifique et non toxique du complexe I de la chaîne respiratoire par la metfor-mine diminue modérément le rapport intracellulaire ATP/AMP dans des hépa-tocytes incubés avec des concentrations thérapeutiques de metformine [12]. Cet effet sur la mitochondrie pourrait fournir une explication à l’inhibition rapide de la gluconéogenèse par la metformine. La gluconéogenèse est une voie méta-bolique très consommatrice en énergie puisqu’elle nécessite 4 ATP et 1 GTP par molécule de glucose formée. Par conséquent, la diminution du contenu intracellulaire en ATP par inhibition du complexe I réduit le flux gluconéogéni-que (figure 4). De plus, l’augmentation des concentrations intracellulaires de l’AMP inhibe de manière allostérique la fructose-1,6-biphosphatase, une enzyme clé de la gluconéogenèse, contribuant à l’inhibition de la production de glu-
Figure 3 : La metformine contrôle l’expression des gènes de la gluconéogenèse via l’acti-vation de l’AMPK (AMP-activated protein kinase).CREB : CRE binding protein ; G6Pase : glucose 6-phosphatase ; OCT1 : organic cation transporter 1 ; PEPCK : phosphoénolpyruvate carboxykinase ; PGC-1α : PPAR gamma coactivator 1 α ; SHP : small heterodimer partner ; TORC2 : transducer of regulated CREB activity 2.
Figure 4 : Mécanismes d’inhibition de la gluconéogenèse hépatique par la metformine.AMPK : AMP-activated protein kinase ; OCT1 : organic cation transporter 1.

Pour la science
Médecine des maladies Métaboliques - Janvier 2009 - Vol. 3 - N°1
52 Pour la science
cose. Nous avons pu montrer dans des hépatocytes issus de souris délétées de l’AMPK ou de LKB1, que la metformine est encore capable d’inhiber la produc-tion de glucose et que cette inhibition est corrélée avec une diminution du contenu intracellulaire en ATP, suggérant que la metformine est capable d’inhiber la pro-duction de glucose indépendamment de l’axe LKB1/AMPK par une action pure-ment énergétique [M. Foretz, résultats non publiés]. En résumé, la metformine inhibe la production de glucose hépati-que à long terme de manière transcrip-tionnelle via l’activation de l’AMPK, et à court terme en réduisant le flux gluco-néogénique par inhibition du complexe I de la chaîne respiratoire.
La metformine améliore la stéatose hépatique
Le diabète de type 2 est fréquemment associé à une stéatose hépatique non alcoolique. Il s’agit d’une pathologie chroni-que du foie dont l'incidence est croissante dans les pays occidentaux et peut évoluer vers une cirrhose. Par définition, la stéa-tose hépatique non alcoolique consiste en une accumulation de lipides au sein des hépatocytes, qui est une caractéristique importante du syndrome métabolique. En présence d'une inflammation, de nécrose ou de fibrose, on parle de stéatohépatite non alcoolique. Il existe une association forte entre stéatose ou stéatohépatite et insulinorésistance hépatique. Les seuls traitements ayant prouvé leur effica-cité dans cette pathologie sont la perte de poids - lorsque les patients sont en surpoids - et l’utilisation de médicaments améliorant la sensibilité à l’insuline. Ainsi, l’utilisation de la metformine chez l’homme améliore l'insulinosensibilité hépatique, réduit l’hépatomégalie de 20 % et dimi-nue significativement le taux de transami-nases [25]. L’efficacité de la metformine a été confirmée chez des souris obèses ob/ob déficientes en leptine qui présen-tent une insulinorésistance et une stéatose hépatique. Le traitement de ces animaux par la metformine améliore la stéatose hépatique, l'hépatomégalie et la cytolyse hépatique [26]. Chez le rat, la metformine prévient également l’apparition d’une insu-linorésistance dans le foie induite par la
perfusion de lipides en association avec une activation de l’AMPK [27]. Des études in vitro, réalisées sur des hépatocytes en culture primaire et des cellules d’hépa-tome HepG2, suggèrent que l’activation de l’AMPK par la metformine pourrait être en partie responsable de l’amélioration de la stéatose hépatique, en inhibant la lipo-genèse et en stimulant l’oxydation des acides gras [7, 28]. L’AMPK inhibe la syn-thèse d’acides gras par phosphorylation
et inactivation de l’acétyl-CoA carboxylase (ACC), une enzyme clé de la lipogenèse. L’inhibition de l’ACC diminue les concen-trations intracellulaires de malonyl-CoA et entraîne la levée de l’inhibition allostéri-que de la carnitine palmytoyl transférase I (CPTI), une enzyme limitante de l’oxyda-tion des acides gras qui permet l’entrée des acides gras dans la mitochondrie, entraînant la stimulation de la β-oxyda-tion et la production d’ATP (figure 5). De
Figure 5 : Mécanismes impliqués dans l’amélioration de la stéatose hépatique par la metformine.AMPK : AMP-activated protein kinase ; CoA : Coenzyme A ; ChREBP : carbohydrate response element-binding protein ; CPT1 : carnitine palmytoyl transférase I ; p-ACC : phosphorylated acetyl-Coenzyme A carboxylase ; SREBP-1 : sterol regulatory element-binding protein-1.
• Le foie est l’organe cible de la metformine. L’entrée de la metformine dans l’hépatocyte
est dépendante du transporteur OCT1.
• La metformine réduit l’hyperglycémie principalement en diminuant la production de
glucose hépatique.
• La metformine améliore la stéatose hépatique en inhibant la synthèse de novo de
lipides (lipogenèse) et en augmentant l’oxydation des acides gras.
• Le complexe I de la chaîne respiratoire est la cible primaire de la metformine.
• La metformine réduit la synthèse d’ATP en inhibant partiellement le complexe I de la
chaîne respiratoire conduisant à une déplétion énergétique relative.
• La réduction du rapport ATP/AMP générée par la metformine entraîne une diminution du
flux gluconéogénique et lipogénique, deux voies métaboliques consommatrices d’ATP.
• Cette réduction du rapport ATP/AMP entraîne également une activation de l’AMPK
qui va moduler à court terme et à long terme la gluconéogenèse et le métabolisme
lipidique.
Les points essentiels

Mécanisme d’action hépatique de la metformine dans le diabète de type 2 53
Médecine des maladies Métaboliques - Janvier 2009 - Vol. 3 - N°1
plus, l’HMG-CoA réductase (Hydroxy-3-méthylglutaryl-coenzyme A réductase), un des premiers substrats de l’AMPK identifié, est également phosphorylée et inactivée par l’AMPK, conduisant à une inhibition de la synthèse de cholestérol dans les hépatocytes. Comme la gluco-néogenèse, la lipogenèse nécessite une production soutenue d’ATP. Ainsi, l’inhi-bition de la synthèse d’ATP par la metfor-mine intervient également dans la dimi-nution du flux lipogénique. Parallèlement, l’activation de l’AMPK inhibe l’expression des facteurs de transcription SREBP-1c (sterol regulatory element-binding protein-1c) et ChREBP (carbohydrate response element-binding protein) et de leurs gènes cibles impliqués dans la lipogenèse (figure 5) [7, 21, 29, 30]. En conséquence, l’activation de l’AMPK par la metformine va réduire l’accumulation de lipides dans le foie, d’une part en inhi-bant la lipogenèse et, d’autre part, en activant l’oxydation des acides gras. De ce fait, la diminution du contenu lipidique dans le foie limite le phénomène de lipo-toxicité et contribue à améliorer la trans-duction du signal de l’insuline (figure 6).
Dans le cadre d’un diabète de type 2, cet effet sur la sensibilité à l’insuline pourra, de manière indirecte, participer à l’amé-lioration de l’hyperglycémie en restaurant l’inhibition de la gluconéogenèse hépati-que par l’insuline.
Conflits d’intérêt : Les auteurs déclarent ne pas
avoir de conflits d’intérêt relatif au contenu de
cet article.
Références[1] Wang DS, Jonker JW, Kato Y, et al. Involvement of organic cation transporter 1 in hepatic and intestinal distribution of metformin. J Pharmacol Exp Ther 2002;302:510-5.
[2] Shu Y, Sheardown SA, Brown C, et al. Effect of genetic variation in the organic cation trans-porter 1 (OCT1) on metformin action. J Clin Invest 2007;117:1422-31.
[3] Wilcock C, Bailey CJ. Accumulation of metfor-min by tissues of the normal and diabetic mouse. Xenobiotica 1994;24:49-57.
[4] Hardie DG. AMP-activated/SNF1 protein kina-ses: conserved guardians of cellular energy. Nat Rev Mol Cell Biol 2007;8:774-85.
[5] Baas AF, Smit L, Clevers H. LKB1 tumor sup-pressor protein: PARtaker in cell polarity. Trends Cell Biol 2004;14:312-9.
[6] Viollet B, Foretz M, Guigas B, et al. Activation of AMP-activated protein kinase in the liver: a new strategy for the management of metabolic hepatic disorders. J Physiol 2006;574:41-53.
[7] Zhou G, Myers R, Li Y, et al. Role of AMP-acti-vated protein kinase in mechanism of metformin action. J Clin Invest 2001;108:1167-74.
[8] Hardie DG. Neither LKB1 nor AMPK are the direct targets of metformin. Gastroenterology 2006;131:973; author reply 974-5.
[9] Shaw RJ, Lamia KA, Vasquez D, et al. The kinase LKB1 mediates glucose homeostasis in liver and therapeutic effects of metformin. Science 2005;310:1642-6.
[10] Owen MR, Doran E, Halestrap AP. Evidence that metformin exerts its anti-diabetic effects through inhibition of complex 1 of the mitochondrial respiratory chain. Biochem J 2000;348:607-14.
[11] El-Mir MY, Nogueira V, Fontaine E, et al. Dimethylbiguanide inhibits cell respiration via an indirect effect targeted on the respiratory chain complex I. J Biol Chem 2000;275:223-8.
[12] Guigas B, Bertrand L, Taleux N, et al. 5-Aminoimidazole-4-carboxamide-1-beta-D-ribofu-ranoside and metformin inhibit hepatic glucose phosphorylation by an AMP-Activated protein kinase-independent effect on glucokinase trans-location. Diabetes 2006;55:865-74.
Figure 6 : Mécanismes d’action de la metformine dans l’hépatocyte.ACC : acetyl-CoA carboxylase ; AMPK : AMP-activated protein kinase ; ChREBP : carbohydrate response element-binding protein ; OCT1 : organic cation transporter 1 ; SHP : small heterodimer partner ; SREBP-1 : sterol regulatory element-binding protein-1 ; TORC2 : transducer of regulated CREB activity 2.
Au niveau hépatique, la metformine induit de manière primaire une inhibition modérée
du complexe I de la chaîne respiratoire mitochondriale. Cette inhibition entraîne une
diminution pondérée du rapport ATP/AMP, mais suffisante pour réduire le flux de la
gluconéogenèse et de la lipogenèse (figure 6). Cette baisse de la charge énergétique
intracellulaire s’accompagne d’une activation de l’AMPK qui inhibe également, à court
terme et à long terme, ces deux dernières voies anaboliques et stimule l’oxydation
des acides gras (figure 6). Ces effets aboutissent à une réduction de la production
de glucose, une amélioration de la stéatose et de la sensibilité à l’insuline hépatique
(figure 6), et finalement à la diminution de l’hyperglycémie. Ce mécanisme impliquant
une diminution du niveau énergétique dans la cellule ne semble pas être propre à la
metformine. La berbérine, un antidiabétique issu de la pharmacopée asiatique, agit
également via l’inhibition du complexe I et l’activation de l’AMPK [31], suggérant que
le complexe I de la chaîne respiratoire, comme l’AMPK, est une cible thérapeutique
du diabète de type 2.
Conclusion

Pour la science
Médecine des maladies Métaboliques - Janvier 2009 - Vol. 3 - N°1
54 Pour la science
[13] Foretz M, Leclerc J, Hebrard S, Viollet B. Metformin inhibits hepatic gluconeogenesis through an AMPK-independent mechanism. Diabetes 2008;57(Suppl1):A423 [Abstract 1507-P].
[14] Caspary WF, Creutzfeldt W. Analysis of the inhibitory effect of biguanides on glucose absorption: inhibition of active sugar transport. Diabetologia 1971;7:379-85.
[15] Hundal RS, Krssak M, Dufour S, et al. Mechanism by which metformin reduces glu-cose production in type 2 diabetes. Diabetes 2000;49:2063-9.
[16] Gunton JE, Delhanty PJ, Takahashi S, Baxter RC. Metformin rapidly increases insulin receptor activation in human liver and signals preferenti-ally through insulin-receptor substrate-2. J Clin Endocrinol Metab 2003;88:1323-32.
[17] Argaud D, Roth H, Wiernsperger N, Leverve XM. Metformin decreases gluconeogenesis by enhancing the pyruvate kinase flux in isolated rat hepatocytes. Eur J Biochem 1993;213:1341-8.
[18] Large V, Beylot M. Modifications of citric acid cycle activity and gluconeogenesis in streptozo-tocin-induced diabetes and effects of metformin. Diabetes 1999;48:1251-7.
[19] Mithieux G, Guignot L, Bordet JC, Wiernsperger N. Intrahepatic mechanisms under-lying the effect of metformin in decreasing basal
glucose production in rats fed a high-fat diet. Diabetes 2002;51:139-43.
[20] Radziuk J, Zhang Z, Wiernsperger N, Pye S. Effects of metformin on lactate uptake and glu-coneogenesis in the perfused rat liver. Diabetes 1997;46:1406-13.
[21] Foretz M, Ancellin N, Andreelli F, et al. Short-term overexpression of a constitutively active form of AMP-activated protein kinase in the liver leads to mild hypoglycemia and fatty liver. Diabetes 2005;54:1331-9.
[22] Koo SH, Flechner L, Qi L, et al. The CREB coactivator TORC2 is a key regulator of fasting glucose metabolism. Nature 2005;437:1109-11.
[23] Screaton RA, Conkright MD, Katoh Y, et al. The CREB coactivator TORC2 functions as a cal-cium- and cAMP-sensitive coincidence detector. Cell 2004;119:61-74.
[24] Kim YD, Park KG, Lee YS, et al. Metformin inhibits hepatic gluconeogenesis through AMP-activated protein kinase-dependent regulation of the orphan nuclear receptor SHP. Diabetes 2008;57:306-14.
[25] Marchesini G, Brizi M, Bianchi G, et al. Metformin in non-alcoholic steatohepatitis. Lancet 2001;358:893-4.
[26] Lin HZ, Yang SQ, Chuckaree C, et al. Metformin reverses fatty liver disease in obese,
leptin-deficient mice. Nat Med 2000;6:998-1003.
[27] Cleasby ME, Dzamko N, Hegarty BD, et al. Metformin prevents the development of acute lipid-induced insulin resistance in the rat through altered hepatic signaling mechanisms. Diabetes 2004;53:3258-66.
[28] Zang M, Zuccollo A, Hou X, et al. AMP-acti-vated protein kinase is required for the lipid-lower-ing effect of metformin in insulin-resistant human HepG2 cells. J Biol Chem 2004;279:47898-905.
[29] Foretz M, Carling D, Guichard C, et al. AMP-activated protein kinase inhibits the glu-cose-activated expression of fatty acid syn-thase gene in rat hepatocytes. J Biol Chem 1998;273:14767-71.
[30] Kawaguchi T, Osatomi K, Yamashita H, et al. Mechanism for fatty acid "sparing" effect onMechanism for fatty acid "sparing" effect on glucose-induced transcription: regulation of car-bohydrate-responsive element-binding protein by AMP-activated protein kinase. J Biol Chem 2002;277:3829-35.
[31] Turner N, Li JY, Gosby A, et al. Berberine and its more biologically available derivative, dihydroberberine, inhibit mitochondrial respira-tory complex I: a mechanism for the action of berberine to activate AMP-activated protein kinase and improve insulin action. DiabetesDiabetes 2008;57:1414-8.







![IDIS SYNTHÈSE - ipubli.inserm.fr · croître (pour revue, voir [3]). A cet égard, il est important de noter que l'échange Na/Ca fut le premier mé canisme d'expulsion du Ca2+ à](https://img.pdfslide.fr/doc/110x75/5c83f14609d3f2b87d8c43b2/idis-synthese-croitre-pour-revue-voir-3-a-cet-egard-il-est-important.jpg)


![[Product Monograph Template - Schedule D] · Monographie d’OZEMPIC ® (sémaglutide injection) Page 4 de 64 tolérée de metformine) ne permettent pas de maîtriser adéquatement](https://img.pdfslide.fr/doc/110x75/5e1fb58418df9163b97aa3d8/product-monograph-template-schedule-d-monographie-daozempic-smaglutide.jpg)


















