Embed Size (px)
Citation preview
BIOCHIMIE, 1976, 58, 1273-1276. Br ves communications
Mise en dvidence d'un comportement anormal de diverses enzymes protdolytiques
en filtration sur gel de polyacrylamide-agarose.
G 6 r a r d PEAUCELLIER. Stat ion Biologique, 29211 Roscof f (France).
(19-2-1976).
L ' u t i l i s a t i o n c o m p a r a t i v e de d e u x gels e o m m e r c i a u x e n r u e de s 6 p a r e r p a r c h r o m a t o g r a p h i c d ' e x c l u s i o n les p r o t ~ a s e s d iges t i ve s d ' u n ann61ide, a f a i r a p p a r a i t r e des d i f f6 rences c o n s i d ~ r a b l e s e n t r e u n gel de d e x t r a n e s (Sephadex) c o u r a m m e n t u t i l i s6 et u n gel d ' a c r y l a m i d e - a g a r o s e (Ul t roge l ) n o u v e l l e m e n t ¢ o m m e r e i a l i s 6 . De n o m b r e u s e s 6 t u d e s o n t m o n t r 6 q u e le v o l u m e d ' 6 l u t i o n des m a e r o m o l ~ c u l e s en f i l t r a t i on s u r gel de d e x t r a n e s , de p o l y a c r y l a m i d e o u d ' a g a r o s e e s t f o n c t i o n de l e u r t a i l l e et avec u u e a s s e z b o n n e a p p r o x i m a t i o n de l eu r po ids m o l 6 e u l a i r e [11. Des prop~i6t6s s i m i l a i r e s a y a n t 6t6 d6cr i t es p o u r les gels d ' a c r y l a m i d e - a g a r o s e [2], l ' a n o m a l i e c o n s t a t 6 e a donc c o n d u i t h e x a m i n e r le c o m p o r t e m e n t c h r o m a t o g r a p h i q u e s u r ce gel de di- v e r s e s p r o t 6 a s e s pur i f i6es d u c o m m e r c e a i n s i que de q u e l q u e s p r o t 6 i n e s u t i l i s6es f r S q u e m m e n t c o m m e ~ta- lons .
M A T E R I E L ET METHODES.
Les exp6r i enees de c h r o m a t o g r a p h i e d ' e x e l u s i o n s u r e o l o n n e o u t ~t6 f a i t e s s e l o n la m 6 t h o d e d 'Andre~v [3], s u r u n e c o l o n n e de S e p h a d e x G 75 s u p e r f i n e ( P h a r - m a c i a lo t n ° 2165) 1,6 × 40 e m et u n e c o l o n n e d ' U I t r o - gel ACA 54 ( L . K . B . / I n d u s t r i e B i o l o g i q u e F r a n c a i s e ) 1,6 × 70 cm, ~lu6es p a r u n t a m p o n Tr is -HC1 p H 8, 0,05 M c o n t e n a n t 0,1 M de KC1 avec u n d6b i t de 7 m l / h d a n s u n e e n c e i n t e r6fr ig~r~e & 4°C. Les 5 e h a n - t i l l ons , e o n t e n a n t 2 h 8 m g de p r o t 6 i n e d i lnSe d a n s 0,5 m l de t a m p o n , o n t 6t~ a d d i t i o n n 6 s de 0,2 g de s a c c h a r o s e et de 0,25 m g de B leu D e x t r a n 2000. P o u r ~vi ter l ' a u t o l y s e , les p ro t6ases o n t 6t6 p r ~ a l a b l e m e n t i n a e t i v 6 e s p a r t r a i t e m e n t a u P h 6 n y l m 6 t h a n e S u ] f o n y l F l u o r u r e (P.M.S.F.). Les v o l u m e s d '61ut ion o n t ~t6 m e s u r 4 s d i r e e t e m e n t s u r l ' e n r e g i s t r e m e n t de la DO_~s,, de l ' e f f l u e n t de l a co lonne , s a u f p o u r i e s p r o t ~ a s e s de Sabellaria et la t r y p s i n e , qu i o n t 6t6 d~tect~es p a r m e - s u r e de l ' ae t iv i t~ e s t 6 r o l y t i q u e s c i o n le proc4d6 de H u m m e l [4], le t r a i t e m e n t a u P.M.S.F. a y a n t ~t6 n a t u - r e l l e m e n t o m i s d a n s ces eas- lh . L e s ~ l e c t r o p h o r 6 s e s s u r gel de p o l y a c r y l a m i d e (10 p. c e n t en a e r y l a m i d e ) en p r 6 s e n e e de S o d i u m Dod~cyl S u l f a t e (S.D.S.) e t de m e r c a p t o ~ t h a n o l o u t 6t6 ef fee tu~s s e l o n la m ~ t h o d e de W e b e r el al. [5]. L ' a c ~ t y l a t i o n des p r o t ~ a s e s p a r l ' a n h y d r i d e ac~ t ique effectu6e se lon le p r o c e s s u s de S h a r m a et al. [6], a 6t6 su iv ie d ' u n d e s s a l a g e s u r S e p h a d e x G 25 et d ' u u e c o n c e n t r a t i o n p a r u l t r a f i l t r a - t i o n (UM 2 A m i c o n ) et le cas 6ch~ant , les r 6 s i d u s t y ro - s ine on t 6t6 d~ac6tyl6s p a r l ' h y d r o x y l a m i n e 0,5 M [7].
R E S U L T A T S .
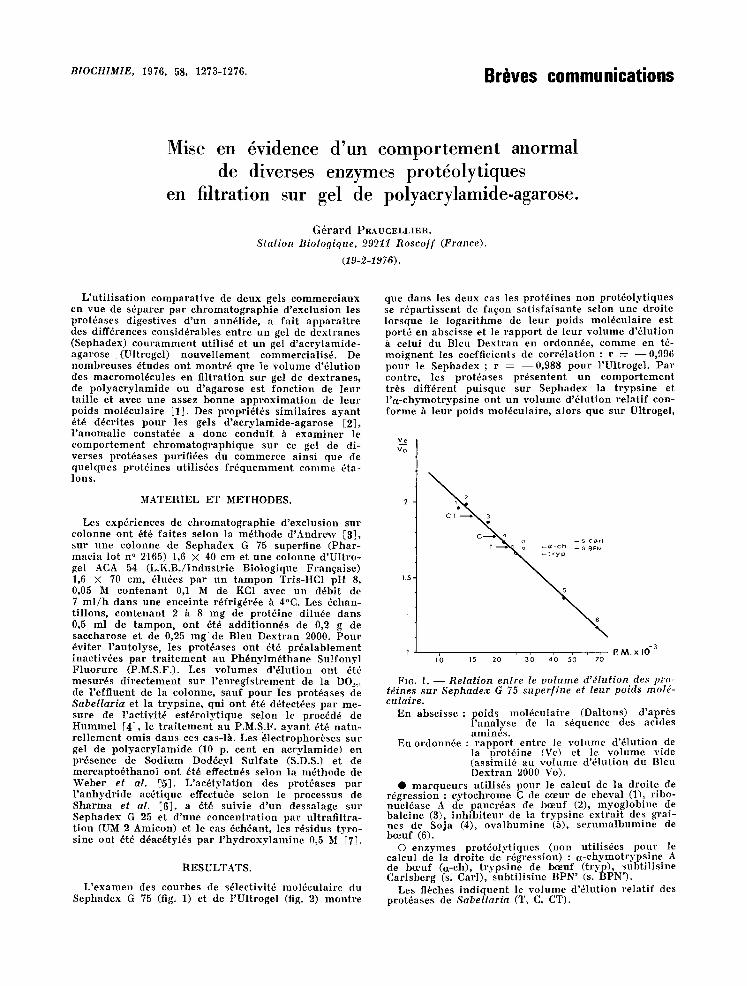
L ' e x a m e n des c o n r b e s de s61eetivit~ m o l ~ c u l a i r e du S e p h a d e x G 75 (fig. I) et de l ' U l t r o g e l (fig. 2) m o n t r e
que d a n s les d e u x cas les p r o t 6 i n e s n o n p r o t 6 o l y t i q u e s se r S p a r t i s s e n t de faqon s a t i s f a i s a n t e s e lon u n e d ro i t e l o r s q u e le l o g a r i t h m e de l eu r po id s m o l 6 c u l a i r e es t por t6 en a b s c i s s e et le r a p p o r t de l eu r v o l u m e d '61ut ion h ce lu i du Bleu D e x t r a n en o rdonn~e , c o m m e en t6- m o i g u e n t ies coef f ic ien ts de cor r61at ion : r : - - 0 , 9 9 6 p o u r le S e p h a d e x ; r : - - 0 , 9 8 8 p o u r l 'U l t rogeI . P a r eon t re , les p r o t 6 a s e s p r d s e n t e n t u n e o m p o r t e m e n t t r~s d i f f6rent p u i s q u e s u r S e p h a d e x la t r y p s i n e et l ' a - e h y m o t r y p s i n e on t u n v o l u m e d '61ut ion r e l a t i f con- f o r m e h l eu r po ids m o l 6 e u l a i r e , a lo r s que s n r Ul t roge l ,
V e
~o
2 2 I e
C T ~
C ~ 4 o - - s c a r l T ~ o ~ e { - c h - - S S P N
--tryp
L5
, , PM. xlO -3 1 I ~0 ] 5 2'0 ' 3 '0 4'0 .5'0 7 0
FIG. 1. - - Relat ion entre le vo lume d'dlution des pro- tdines sur Sephadex G 75 superf ine et leur poids mold- eulaire.
En a b s c i s s e : po ids m o l 6 c u l a i r e (Da l tons ) d ' a p r 6 s l ' a n a l y s e de la s6quence des ac ides a m i n 6 s .
En o r d o n n 6 e : r a p p o r t e n t r e le v o l u m e d '~ lu t i on de la o ro t~ ine (Ve) et le v o l u m e v ide ( a s s l m i l 6 au v o l u m e d '61ut ion du B leu D e x t r a n 2000 Vo).
• m a r q u e u r s u t i l i s6 s p o u r le ea lcul de l a d ro i t e de r d g r e s s i o n : e y t o e h r o m e C de coeur de cheva l ( l ) , r ibo- n u c l 6 a s e A de p a n e r 6 a s de ]meuf (2), m y o g l o b i n e de b a l e i n e (3), i n h i b i t e u r de la t r y p s i n e e x t r a i t des g ra i - ue s de Soja (4), o v a l b u m i n e (5), s e r u m a l b u m i n e de bceuf (6).
© e n z y m e s p r o t d o l y t i q u e s (non u t i l i s~es p o u r le ca lcu l de la d ro i t e de r6g re s s ion ) : a - c h y m o t r y p s i n e A de boeuf (ct-ch), t r y p s i n e de beeuf ( t ryp) , s u b t i l i s i n e C a r l s b e r g (s. Car l ) , s u b t i l i s i n e BPN ' (s. BPN') .
Les f lbches i n d i q u e n t le v o l u m e d '61ut ion r e l a t i f des p ro t~ases de Sabellaria (T, C, CT).
1 2 7 4 G. P e a u c e l l i e r .
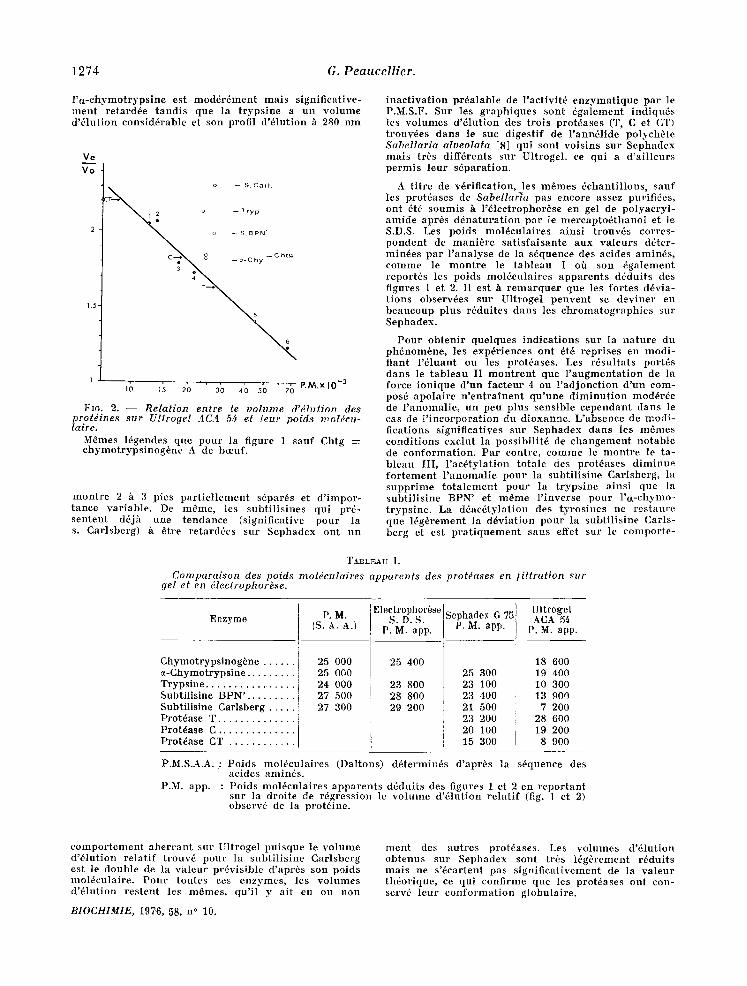
l ' t t - e h y m o t r y p s i n e es t m o d d r d m e n t m a t s s ign i f i ca t ive - m e n t r e t a rd~e t a n d i s que ]a t r y p s i n e a u n v o l u m e d '61ut ion e o n s i d d r a b l e et s o n profi l d ' d l u t i o n h 280 n m
V e
Vo o - - S . C a r l .
I 2 o - - T r y p
2 o - - S . B P N '
- - C h t g ~ " - - a - C h y . '
1 I; 1; 20 ' ' 30 4'0 5b ' 7; P ' M ' x l O ' 3
Fro. 2. - - R e l a t i o n en tre le v o l u m e d 'd la t ion des p ro td ines s u r Ul t roge l ACA 5~ et [eur po ids m o t dcu - laire.
Mdmes ldgendes qne p o u r la f igure 1 s a u f Ch tg -~ e h y m o t r y p s i n o g 6 n e A de bceuf.
m o n t r e 2 h 3 p ies p a r t i e l l e m e n t sdpards et d ' i m p o r - t a n c e v a r i a b l e . De m S m e , les s u b t i l i s i n e s qu i prd- s e n t e n t dd jh u n e t e n d a n c e ( s ign i f ica t ive p o u r la s. Ca r l sbe rg ) h a t re r e t a r d d e s s u r S e p h a d e x o n t u n
i n a c t i v a t i o n p r d a l a b l e de l ' ae t iv i td e n z y m a t i q u e p a r le P.M.S.F. Su r les g r a p h i q u e s s o n t 6 g a l e m e n t i n d i q u 6 s les v o l u m e s d ' d l u t i o n des t r o i s p r o t d a s e s (T, C et CT) t r o u v d e s d a n s le sue d ige s t i f de l ' a n n d l i d e po lyeh6 te S a b e l l a r i a a Iveo la ta [8] qu i s o n t v o i s i n s s u r S e p h a d e x m a i s t rbs d i f f~ren ts s u r Ul t roge l , ee qu i a d ' a i l l e u r s p e r m i s l eu r s d p a r a t i o n .
A t i t r e de vdr i f iea t ion , les m S m e s 6 e h a n t i l l o n s , s a u f les p r o t d a s e s de Sabe l lar ia pas encore a s sez pur i f i6es , o u t 6t6 s o u m i s h l '61ee t rophor~se en gel de p o l y a c r y l - a m i d e apr6s d d n a t u r a t i o n p a r le m e r e a p t o d t h a n o I et ]e S.D.S. Les po id s m o l ~ e u l a i r e s a i n s i t r o u v d s co r res - p o n d e n t de m a n i 6 r e s a t i s f a i s a n t e a u x v a l e u r s do te r - m i n d e s p a r l ' a n a l y s e de la s6quenee des ae ides a m i n d s , c o m m e le n m n t r e le t a b l e a u I off son d g a l e m e n t r epo r tds les po ids m o l d e u l a i r e s a p p a r e n t s d d d u i t s des f igures 1 et 2. I1 e s t h r e m a r q u e r que les f o r t e s ddv ia - t i o n s obse rvdes s u r Ul t roge l p e u v e n t se d e v i n e r en b e a u e o u p p l u s r d d u i t e s d a n s les e h r o m a t o g r a p h i e s su r S e p h a d e x .
P o u r o b t e n i r q u e l q u e s i n d i c a t i o n s s u r la n a t u r e du p h d n o m 6 n e , les expd r i enees o u t dtd r e p r i s e s en m o d i - f ian t l ' 61uant ou les p ro tdases . Les r d s u l t a t s por tds d a n s le t a b l e a u II m o n t r e n t que l ' a u g m e n t a t i o n de la force i o n i q u e d ' u n f a e t e u r 4 ou l ' a d j o n e t i o n d ' u n e o m - pos6 a p o l a i r e n ' e n t r a i n e n t q u ' u n e d i m i n u t i o n moddrde de l ' a n o m a l i e , u n peu p l u s s e n s i b l e e e p e n d a n t darts le cas de t ' i n e o r p o r a t i o n du d i o x a n n e . L ' a b s e n e e de m o d i - f i ca t ions s ign i f i ea t ives su r S e p h a d e x d a n s les m 8 l u e s c o n d i t i o n s e x e l u t la pos s ib i l i t d de e h a n g e m e n t n o t a b l e de c o n f o r m a t i o n . P a r eont re , e o m m e le m o n t r e le t a - b l e a u III, l ' a e d t y l a t i o n to t a l e des p ro tda se s d i m i n u e f o r t e m e n t l ' a n o m a l i e p o u r la s u b t i l i s i n e Ca r l sbe rg , la s u p p r i m e t o t a l e m e n t p o u r la t r y p s i n e a ins i que la s u b t i l i s i n e BPN ' et m 8 m e l ' i n v e r s e p o u r l ' a - e h y l n o - t r y p s i n e . La d d a e d t y l a t i o n des t y r o s i n e s ne r e s t a u r e que 16g6rement la d d v i a t i o n p o u r la s u b t i l i s i n e Ca r l s - be rg et es t p r a t i q u e m e n t s a n s effet su r le c o m p o r t e -
TABLEAU I.
C o m p a r a i s o n des po ids m o l d e u l a i r e s a p p a r e n t s des pro tdases en f i l t r a t i o n su r gel et en d lec trophorbse .
] P .M. Electrophor~se Sephadex G 75 ]- UltrogelACA 5~ Enzyme ] (S. A .A. ) S . D . S . P .M. app. ] P .M. app.
i P .M. app.
25 400 i
C h y m o t r y p s i n o g ~ n e . . . . . . i 25 000 a - C h y m o t r y p s i n e . . . . . . . . . i 25 000 T r y p s i n e . . . . . . . . . . . . . . . . . ,i 24 000 Subt i l i s ine B P N ' . . . . . . . . . I 27 500 Subt i l i s ine Car lsberg . . . . . :~ 27 300 Pro tdase T . . . . . . . . . . . . . . i P ro tdase C . . . . . . . . . . . . . i Pro t~as¢ CT . . . . . . . . . . .
23 800 28 800 29 200
25 300 23 100 23 400 21 500 23 200 20 100 15 300
18 600 19 400 10 300 13 900
7 200 28 600 19 200
8 900
P.M.S.A.A. ; P o i d s m o l d c u l a i r e s (Da l tous ) d d t e r m i n e s d ' a p r b s la sdquence des ae ides a m i n d s .
P.M. app . : P o i d s m o l d c u l a i r e s a p p a r e n t s d d d u i t s des f g u r e s 1 et 2 en r e p o r t a n t s u r la d ro i t e de r d g r e s s i o n le v o l u m e d ' d ]u t i on r e l a t i f (fig. 1 et 2) observd de la p ro t6 ine .
c o l n p o r t e m e n t a b e r r a n t s u r Ul t roge l p u i s q u e le v o l u m e d '61ut ion r e l a t i f t rouvO p o u r la s u b t i l i s i n e C a r l s b e r g es t le d o u b l e de la v a l e u r p r d v i s i b l e d ' a p r 6 s s o n po ids m o l d c u l a i r e . P o u r t o u t e s ces e n z y m e s , les v o l u m e s d ' d l u t i o n r e s t e n t les mOmes , qu ' i l y a i t eu o u n o n
BIOCHIMIE, 1976, 58, n ° 10.
mer i t des a u t r e s p ro tdases . Les v o l u m e s d '61ut ion o b t e n u s s u r S e p h a d e x s o n t t rbs l d g 6 r e m e n t r d d u i t s m a t s ne s ' d c a r t e n t pa s s i g n i l i e a t i v e m e n t de ]a v a l e u r t hdo r ique , ce qui con f i rme que les p ro t~ase s on t con- serv6 l e u r c o n f o r m a t i o n g lobu la i r e .
Filtration de protdases sur gel de polyacrylamide-agarose. 1275
TABLEAU II.
Ef fe t de l'~lnant snr l'dcart relatif du volume d'~lution des protdases & la valeur thdorique.
Ultr0gel ACA 54 Sepbadex G 75
I - -
+ 7 v + s z a-Chymotrypsine ". _ 2 . p. cem t p . c ¢ , t
- - 2 Wrypsine ! + • I 4 -. _ _ [ p. cent p. cent_
S u b t i l i s i n e BPN' ] + 20.4 + 20.7 _ _ _ _ _ I p. c e n t p. cen~
Subtillsine Carlsberg p-~. c3:n: p~.. 33nt8
p-~, 2.2 c e n t
c e n t
p~ 17.2 c e n t
cent
- - 0 . 3 - - 0.5 p. cent p. c e n t
1.3- ). cent p. cent
- +
). c en t p. c e n t
?. c e n t I p . c e n t
+ 0.3 p. cent
+ 5 . 2 p. cent
4 5 . 7 p. c e n t
+ 3 . 3 p. c e n t
Ve obs - - Ve th Les poureentages indiquds correspondent au rapport Ve th , diffd-
rence entre le volume d'6lution observd (Ve obs) et le volume d'dlution thdorique (Veth) (correspondant sur la droite de rdgression au P.M.S.A.A.) sur ee volume thdorique.
Pour chaque dluant les volumes d'~lution des six marqueurs ont dtd mesurds et utilisds pour le calcul des volumes d'dlution thdoriques des protdases par rdgression lindaire.
TABLEAU III.
Effe t de l'ac~tylation sur l'dcarl relatif du volume d'dlution des protdases h la valeur thdorique.
~-Chymotrypsine
Trypsine
Subtilisine BPN'
Subtilisine Carlsberg
Ultrogel ACA 54 Sephadex G 75
Acetylation hcdtylation Ae6tylation Aedtylation totale partielle totale partielle
, - - 2.5 p. cent + 4.0 p. c e n t
1.5 p. cent + 0.5 p. cent
8 . 3 p. cent
c e ° t
0--3p.p. cent
~ 7 . 1 p. c e - -
~ 0 . 1 p. cent
1.1 p. cent
0.5 p. cent
+ 2.8 p. c e n t
[ + 1.1 p. cent
La signification des pourcentages est la m~me clue dans le tableau II. La colonne << acdtylation totale >> donne les rdsultats obtenus avec les protdases
s implement acdtyldes, tandis que ccux des protdases traitdes par l 'hydroxylamine apr6s acdtylation sont portds dans la colonne << acdtylation part iel le >7.
DISCUSSION.
Les rdsultats prdcddents mont ren t c la i rement qu 'un facteur autre que le tamisage moldculaire in tervient lorsque les protdases sont filtrdes sur gel d 'acrylamide- agarose. Les retards d'dlution observds sugg6rent qu'il s 'dtablit entre ces enzymes et le gel des interact ions rdversibles qui re tardent la migration. Un certain hombre de comportements anorrnaux sur Sephadex ont dSjh dtd d6crits concernant des glycoprotdines telles l 'ovomucoide [8] dont le volume d'61ution est anorma- lement faible ou des enzymes glyco]ytiques tels le
BIOCHIMIE, 1976, 58, n ° 10.
lysozyme [9] et l 'amylase [10] dont l 'dlution est retardde, probablement par des interact ions de type enzyme-subs t ra t avec le gel, ce qui ne peut 6videm- merit ~tre invoqud dans le cas des protdases. Plus rdcemment cependant, le compor tement anormal d 'autres protdases sur Sephadex a dtd notd [11 et 12] sans qu'aucune explication nit dt6 avancde. En effet, on ne peut expliquer la rdtention par une simpIe inter- action dlectrostatique entre les quelques charges nd- gatives prdsentes dans le gel et la charge globale des prot6ines puisque selon cette hypoth6se la subti l is ine BPN' dont le point isodlectrique (Pi) est 7,8 ainsi que

1 2 7 6 G. P e a u c e l l i e r .
l ' (~-chymotrypsine (Pi = 8,1) ne devra ien t pas 8tre retard~es alors que la t ryps ine (Pi : 10,8) devra i t l 'Stre p lus que la subt i l i s ine Carlsberg (Pi ---- 9,4) et le cy tochrome C (Pi ---- 10) devrai t l '~tre au tan t . I1 f au t cependant noter que la charge globale peut 8tre responsable d 'une pat t ie de la rStent ion pu isque celle- ci d i m i n u e par a u g m e n t a t i o n de la force ioniqne dans le cas de la t ryps ine el de ]a subt i l i s ine Carlsberg.
Un aut re processus f r 6 q u e m m e n t invoqu~ dans les phdnom6nes d ' adsorp t ion est l ' in te rae t ion hydropbobe dont l ' i n te rven t ion a 6t6 d6montr~e duns l ' associa t ion de la t ryps ine avec l ' i nh ib i t eu r pancr6at ique [13] ou ut i l is6 pour la purif icat ion de l ' u - c h y m o t r y p s i n e su r Sepharose subs t i tu6 avec des rSsidus 2,4-dinitro- ph6nyl [14]. A la concent ra t ion de 10 p. cent le d ioxanne r6duit s ignif ica t ivement ]a r6 tent ion de routes les protdases alors que le degr6 d ' imbib i t ion du gel n 'es t pus modifi6 et q u ' a u c u n effet de par t i t ion notable n 'es t h craindre, comme Font mont r5 B u s h et al. [15] dans le cas du Sephadex. I1 semble done que des in te rac t ions hydrophobes in te rv iennent , au m o t h s pour pat t ie , dans la r6 tent ion des prot6ases test6es bien que duns le eas~:pr~sent l ' inac t iva t ion de la s6rine du site act i f ne modifie pas I ' interaet ion.
On pour ra i t 6galement penser h une act ion des acides amines a roma t iques puisque , outre leurs ten- dances hydrophobes , la r ichesse en dlectrons ~ de leur cycle a ~td reconnue responsable de len t adsorp t ion su r Sephadex [16]. En fait, ee type d ' in te rac t ion ne semble pas intervenir , sauf pour une fa ible par t dans la subt i l i s ine Carlsberg, car la ddac~tylat ion des tyro- s ines par l ' h y d r o x y l a m i n e ne res taure pas l ' anomaI ie .
Les charges posit ives des rdsidus lys inc superficieIs sont done les responsab les les plus v ra i semblab les de la r6 ten t ion puique leur neu t ra l i sa t ion par acfitylation la fair d ispara l t re . Ce r6sultat , assez s u r p r e n a n t ~tant donn6 ce qui a 6t6 dit plus haut , concorde p o u r t a n t avec ceux de S h a r m a et al. [6] qui ont mont rd qne l 'acStylat ion de l ' c t -chymotrypsine suppr ime 6galement une in terac t ion ind i spensab le "~ sa f ixation sur D-tryp- tophane m e t h y l ester-Sepharose. I1 f au t donc supposer commc ces au teurs , qu 'une r @ a r t i t i o n part icul i6rc des charges posi t ives des groupes e-aminds superficiels (donc sans r appor t direct avec le Pi) pe rmet des in ter- act ions dlectrosta t iques avec les quelques g roupemen t s
charges du gel. Toutefois , il se pour ra i t auss i que Fad- jonc t ion des g roupemen t s ac~tyl h la surface de la pro- t~ase crde u n emp~chement st6rique ou au t re h Fin- teract ion de rdsidus voisins.
Les anoma l i e s qui v i ennen t d 'etre d6crites ne de- v ra ien t cependant pas en t raver la g6n6ral isa t ion de l 'emploi des gels mix tes d ' ac ry lamide agarose qui sc just i f ie par leur rdsolu t ion tr6s sup6r ieure h celle du Sephadex. On peut mSme envisager l ' u t i l l sa t ion syst6- ma t iqne de l 'Ul trogel pour sdparer d iverses enzymes protdolyt iques d 'un m61ange comme cela a 6t6 fa i t avec succ6s pour les sucs digest ifs de Sabellaria alveolata.
Remerciements .
Je remercie v ivement le P rofesseur J. Pudles pour les prdcieux conseils qu ' i l m ' a a i m a b l e m e n t prodiguds.
BIBLIOGRAPHIE.
1. Rei land, . J . (1971) in Methods in Enzymology (Jakobi, W. B. ed.), vol. XXII, pp. 287-321, Aca- demic Press, New York and London.
2. Uriel, J., Berg6s, J., Boschetti , E. & Tixier, R. (1971) C. R. Acad. Sci. Paris, Ser. D, 273, 2358-2360.
3. Andre'ws, P. (1964) Biochem. J., 91, 222-2~33. 4. Hummel , B. C. W. (1959) Can. J. Biochem. Phgsiol. ,
3'7, 137-142. 5. Weber, K., Pringle, P. R. & Osborn, M. (1972) in
Methods in Enzymolo~y (Hirs, C. H. W, ,¢ Ti- mashe l l , S. ed.) vol. XXVI, pp. 3-27, Academic Press, Ne'w-Yo~k and London.
6. Sharma, S. K. & Hopkins, T. R. (1975) J. Chroma- tog., !1110, 321-326.
7. Labouesse, J..& Gerx ais, M. (1967) European J. Bio- chem., 2, 215-223.
8. Peaucell ier , G., m a n u s c r i t en prSparat ion. 9. Whi take r , J. R. (1963) Anal. Chem., 3'5, 1950-1953.
10. Gelotte, B. (1964) Acta Chem. Scan&, 18, 1283-1291. I1. Voordouv, G., Gaucher, G. M. & Roche, R. S. (I974)
Biochem. Bioph!ls. Res. Comm., 58, 8-12. 12. Starkcy, P. M. & Barret , A. J. (19'76) Biochem. J.,
.155, 255-263. I3. CIothia, C. a Jan in , J. (1975) Nature, '256, 705-708. 14. SteDanov, V. M., Lavrenova, G. I., Borovfl~ova, V.
P. & Balandina , G. N. (1975) J. Chromatog., 104, 373-377.
15. Bush, B., Jones, T. E. L. & T h o b u r n Burns , D, (1970) J. Chromatoq., 49, 448-454.
16. Janson , J. C. (1967) J. Chromatog., 28, 12-20.
BIOCH1M[E, 1976, 58, n ° 10.