Embed Size (px)
Citation preview

3
REPUBLIQUE ALGERIENNE DEMOCRATIQUE ET POPULAIRE MINISTERE
DE L’ENSEIGNEMENT SUPERIEUR ET DE LA RECHERCHE
SCIENTIFIQUE
UNIVERSITE D’ORAN ES-SENIA
FACULTE DES SCIENCES
DEPARTEMENT DE LA BIOTECHNOLOGIE
Mémoire de MAGISTER en Biotechnologie
Option: Intérêt des microorganismes en Agriculture et en Agro-alimentaire
Présenté par
SIDHOUM Warda
Thème :
Soutenu le / / 2011 devant la commission d’examination :
M. AOUES Aek . Professeur Université d’Oran Président
Mme KAÏD-HARCHE M. Professeur USTOMB (ex USTO), Oran Examinateur
M. KACEM M. Professeur Université d’Oran Examinateur
Mme FORTAS Z. Professeur Université d’Oran Rapporteur
Diversité des mycorhizes arbusculaires chez la variété « Sigoise » d’olivier (Olea europea L.) : étude
de leurs efficacités sur la croissance des plants.

4
Introduction :
L‟oléiculture à base d‟olivier (Olea europea L.), prend une place considérable dans l‟économie de
l‟Algérie. D‟ailleurs, une prise de conscience s‟est opérée récemment à ce sujet pour améliorer la
conduite du verger oléicole, réfléchir à son extension sur des terres où l‟intensification de la produc tion
est possible.
En effet, l‟État algérien a mis en place un Plan National Oléicole (PNO) en 2000. Ce plan avait
comme objectifs, l‟extension de la superficie des oliveraies à 500 000 ha, à l‟horizon 2010 (Argenson,
2008).
Cependant, ce regain d‟intérêt pour l‟arboriculture, d‟une manière générale et pour l‟oléiculture de
façon particulière, risque d‟être remis en cause par manque de plants sains, issus de variétés adaptées et
productives. Les nouvelles techniques de reproduction ou de multiplication, si elles sont maîtrisées
peuvent rendre de grands services dans ce domaine, le cas du microbouturage qui permet de multiplier
des espèces difficiles à reproduire naturellement, avoir des vitroplants facilement transportables sans
risques sanitaires et reboiser très rapidement avec une diminution du coût de production.
L‟olivier, comme la plupart des arbres fruitiers, est un partenaire symbiote potentiel des
champignons endomycorhiziens qui lui permettent de mieux résister à différents stress
environnementaux, la salinité, la sécheresse et certains microorganismes telluriques en zones aride et
semi aride de notre pays.
En Algérie, l'étude de la biodiversité des champignons mycorhiziens arbusculaires reste le parent
pauvre de l'écologie microbienne. La valeur agronomique relative des isolats indigènes est inconnue,
malgré l‟omniprésence des symbioses mycorhiziennes arbusculaires dans notre environnement naturel,
elles restent insuffisamment utilisées en agriculture moderne. C‟est dans ce cadre que nous avons
entrepris une étude sur les CMA indigènes des sols des vergers d‟oliviers et leur association
mycorhizienne avec la variété Sigoise d‟olivier in situ et en serre en conditions contrôlées avec comme
objectif d‟essayer de sélectionner des CMA capables de mycorhizer des boutures herbacées en pépinière.

5
Ainsi, notre étude consistera à :
- estimer le taux d‟infection endomycorhizienne des oliviers en conditions naturelles, de
déterminer le pouvoir endomycorhizogène du sol (PEM) et le nombre le plus probable de
propagules (NPP) ;
- isoler les spores des champignons mycorhiziennes arbusculaires indigènes du sol en essayant
de les caractériser pour les identifier;
- étudier la cinétique de la mycorhize à l‟intérieur des systèmes racinaires naturels ;
- réaliser des essais d‟endomycorhization contrôlée des boutures herbacées de la variété Sigoise
d‟olivier Olea europea L. avec des CMA sélectionnés et déterminer leur efficacité sur la
croissance et le développement de ces boutures.
- et enfin tenter la culture axénique in vitro des CMA sélectionnés dans le but de les multiplier,
d'évaluer la qualité de l'inoculum de production sans risque de contamination et d‟avoir une
collection locale de souches disponibles au laboratoire.
1. Généralités sur les CMA
1.1. Historique
Les premières études sur les racines auraient commencé vers le XVIIème siècle (1665).
En 1829, Meyen décrit d‟étranges formations dans le système racinaire de l‟hêtre qu‟il
supposa parasité. Fries (1829) par la suite, a rapporté la présence d‟hyphes à l‟extérieur des
racines de Monotropa hypopytis L.
Tulasne et Tulasne (1844) ont confirmé les observations faites par Théophraste (avant
2000 ans) sur l‟omniprésence des champignons autour des racines de quelques essences
forestières, ils ont également affirmé que les manchons d‟hyphes autour des racines de
quelques arbres provenaient de la truffe. Et depuis, Gasparini a signalé la présence du
mycélium de la truffe autour du pin, du noisetier et du châtaigner. Cependant, jusqu‟ici on
décrivait le phénomène comme étant du parasitisme. Le terme d‟association n‟est apparu au
grand jour qu‟après les études de Pfeffer (1877) et Kamienski (1881), qui a découvert
l‟association symbiotique entre les champignons et les racines de Monotropa, cette
association a été nommée Mycorhize en 1885 par Frank.

6
Les premières études sur l‟endomycorhize à arbuscules ont commencé durant la
période (1875- 1895), quant à l‟endomycorhize à pelotons des orchidées, elle a été
découverte par Bernard (1899) ; ce même auteur a fait les premiers isolements du
champignon.
Les plus importantes descriptions anatomiques des MA chez les plantes hôtes sont
celles rapportées par Schlicht (1889), Dangeard (1896), Janse (1897), Petri (1903), Gallaud
(1905), Peyronel (1923 et 1924), Jones (1924) et Lohman (1927).
Schlicht (1889) a signalé la présence de relations anatomiques entre les t issus de la
plante et le champignon et Janse (1897) a observé les structures intra-racinaires produites par
le symbiote fongique qu‟il a appelé vésicules et a mentionné la présence d‟autres structures
qu‟il n‟a pas pu définir ; celles- ci furent appelées arbuscules par Gallaud en 1905.
L‟appressorium a été décrit par Jones (1924) ; les spores et les sporocarpes à l‟origine de la
mycorhize arbusculaire ont été décrits par Peyronel (1924).
1.2. Distribution géographique
Le champignon mycorhizien étant peu spécifique, il n‟existe aucun site naturel où il y a
absence d‟endomycorhizes sauf dans des zones tourbeuses, des dunes récentes et des dépôts
morainiques (dépôts de sable, de cailloux et d'argile). La diversité et la distribution des CMA
résultent des processus écologiques temporaires agissant sur les communautés végétales et
fongiques, la température, le pH et le niveau de P dans la solution du sol ainsi que le génotype
des plantes constituent des facteurs limitant la distribution des espèces de CMA tels que
Glomus sp. et d’Acaulospora leavis (Johnson et al., 2004).
L‟intervention de l‟homme peut aussi aboutir à la création de zones dépourvues
d‟inoculum endomycorhizien, c‟est le cas d‟exploitations minières et des déblais de toutes
sortes. Il semblerait également que les landes à Callunes soient dépourvues de mycorhizes.
Les Éricacées forment des mycorhizes spécifiques, et lorsqu‟il y a monospécificité, la
disparition des plantes pouvant héberger des CMA provoque par conséquent la disparition du
symbiote fongique (Le Tacon et al., 1999).
Des espèces de CMA sont plus dominantes dans des régions plutôt que dans d‟autres, le
cas de Glomus intraradices , espèce à grande distribution géographique dans le monde

7
domine les sols agricoles (Mathimaran et al., 2005), tandis que Glomus constrictum est
spécifique du climat semi-aride (Azcon-Aguillar, 2003 ; Abbas et al., 2006) (tableau 1)
Tableau 1 : Habitats et plante hôtes des principales espèces de CMA.
Famille des CMA Espèce de CMA plante hôte Habitat Références
Acaulospora
bireticulata
Sassafrans albidum Ouest du Kentucky Rothwell et Trappe
(1979)
Les
Acaulosporacées
A. bireticulata Sentrosema
pubecensL.
Floride Schenck et Smith (1982)
A. bireticulata Malus domestica Michigan Miller et al. (1985)
A. bireticulata Allium sativum Sud de Chine Li et al. (2007)
A. bireticulata plantes diverses Rivière de Jansa,
Chine
Dandan et Zhiwei,
( 2007)
A.capsiculata plantes diverses Caroline du nord Schultz et al. (1999)
A. gerdemannii Araucaria
angustifolia
San Paulo, Brésil Moreira- Souza ( 2003)
A.gerdemannii Blutaparon
portulacoidesl
Santa Catarina sud
du Brésil
Cortoba et al. (2001)
A.gedanensis Festuca ovina Chalupy Bloszkowski (1988)
A. koskii Prosopis
glandulosa
Californie Sturmer (1998)
A. lacunosa Andropogon
viginicus
Virginie Sturmer (1998)
A.lacunosa Michelia champaca
L
Nord de l‟Inde Panna et Highland
(2010)
A.lacunosa les Meliacées Hainan Island Chine Shi et al. (2006)
A. mellea Ma. pumila Santa Catarina sud
du Brésil
Cavallazzi et al. (2007)
A. mellea Dioscorea spp Benin, ouest
d‟Afrique
Tchabi et al. (2008)
A. mellea plantes diverses Benjamin Brésil Stürmer et Siqueira
(2010)
A. paulinae plantes diverses Ouest de Poméranie Iwaniuk et Bloszkowski
(2004)

8
A. paulinae Poa annua Island Koske et al. (1997)
A.paulinae plantes diverses
Les dunes, Tel-Aviv.
Israel
Bloszkowski et al.(2001)
A. paulinae plantes diverses
Benjamin Brésil Stürmer et Siqueira
(2010)
Les
archaeosporacées
Archaeospora trappei plantes diverses USA Morton et Redecker
(2001)
Ar. trappei plantes diverses Iceland Greipsson et al. (2002)
Ar. trappei A. porrum et autre
plantes cultivées
Chine Gai et al. (2006)
Les
Entrophosporacé
es
Entrophospora
infrequens
plantes diverses Chine Tao et Zhiwei (2005)
E.infrequens plantes diverses Rivière de Jansa,
Chine
Dandan et Zhiwei
(2007)
Gigaspora albida Champ de tabac Chine Fang et al. (2000)
Les
Gigasporacées
G. albida Blutaparon
portulacoidesl
Sud du Brésil Cordoba et al. (2001)
G. albida Ma. pumila. Santa Catarina, sud
du Brésil
Cavallazzi et al. (2007)
G. albida Lawsonia inermis Goa, Inde Radhika et Rodrigues
(2010)
G. decipiens Dioscorea spp Benin, ouest
d‟Afrique
Tchabi et al. (2008)
G. gigantea plantes diverses Cotes de la mer
Baltique
Tadych et Bloszkowski
(2000)
G. gigantea
plantes diverses Cotes de l‟atlantique
du Québec jusqu‟au
nord d‟Amérique
Gemma et Koske,
(1989) ; Gemma et
al.(1989)
Tableau 1(suite)
G. gigantea plantes diverses Chine Wu et al. (2002)
G.gigantea plantes diverses Rivière de Jansa,
Chine
Dandan et Zhiwei,
(2007)

9
G. margarita Ar. angustifolia San Paulo, Brésil Moreira- Souza (2003)
G. margarita A. albida Sénégal Diop et al. (1994)
G.margarita P. glandulosa Sénégal Diallo et al, (1999)
G.margarita Les Meliacées Hainan Island, Chine Shi et al. (2006)
Scutellospora
calospora
Arachis hypogaea Sud de Chine Li et al. (2007)
S. calospora Ma. pumila. Santa Catarina, sud
du Brésil
Cavallazzi et al. (2007)
S. calospora Meliacées Hainan Island, China Shi et al. (2006)
S. heterogama A. sativum Sud de Chine Li et al. (2007)
S. heterogama Dioscorea spp. Benin, ouest
d‟Afrique
Tchabi et al. (2008)
S.heterogama plantes diverses Rivière de Jansa,
Chine,
Dandan et Zhiwei
(2007)
Les Gloméracées Glomus aggregatum Tetraclinis
articulata
Maroc Abbas et al. (2006)
Gl. aggregatum Ara. Araucaria a Sud de Chine Li et al. (2007)
Gl. aggregatum angustifolia
hypogae
San Paulo, Brésil Moreira- Souza (2003)
Gl. aggregatum Ara. hypogaea Benin, ouest
d‟Afrique
Tchabi et al. (2008)
Gl. aggregatum plantes diverses Rivière de Jansa,
Chine
Dandan et Zhiwei
(2007)
Gl. aggregatum Olea europea L. Murcia Azcon-Aguillar (2003)
Gl. aggregatum Azadirachta indica Goa, Inde Radhika et Rodrigues
(2010)
Gl. constrictum
Mi. champaca L. Nord de l‟Inde Panna et Highland
(2010)
Gl. constrictum T. articulata Maroc Abbas et al. (2006)
Gl. constrictum Di. spp Benin, ouest
d‟Afrique
Tchabi et al. (2008)
Gl. constrictum Blutaparon
portulacoidesl
Santa Catarina sud
du Brésil
Cordoba et al. (2001)

10
Glomus constrictum plantes diverses Rivière de Jansa,
Chine
Dandan et Zhiwei
(2007)
Gl. constrictum Mi.champaca L.
Nord de l‟Inde Panna et Highland
(2010)
Gl.constrictum Méliacées Hainan Island,
China
Shi et al. (2006)
Gl. etunicatum Am. arenaria ssp.
arundinacea
Comporta Portugal Rodríguez-Echeverría
et Freitas (2006)
Gl. etunicatum Malus pumila Santa Catarina, sud
du Brésil.
Cavallazzi et al. (2007
Gl. etunicatum Dioscorea spp Benin, ouest
d‟Afrique
Tchabi et al. (2008)
Glomus intraradices
Araucaria
angustifolia
San Paulo, Brésil Moreira et al. (2007)
Gl.intraradices Pistacia lentiscus Pianosa Island, Italie Turrini et al. (2010)
Gl.s intraradices plantes diverses chine Tao et Zhiwei (2005)
Gl. intraradices Plantes diverses Alicante (Espagne) Alguacil et al.(2009)
Gl. intraradices P. glandulosa Sénégal Diallo et al. (1999)
Gl. intraradices Andropogon
garardii
Kansas Sturmer et al. (1998)
Gl. intraradices Fragaria vesca L. Pologne Turneau et al. (2001)
Gl. intraradices plantes diverses Switzer land Jansa et al .(2002)
Gl. intraradices plantes diverses Israël Bloszkowski et
Czernawska (2006)
Tableau 1(fin)
Gl. intraradices plantes diverses Chine. Gai et al.(2006)
Gl. intraradices plantes diverses Afrique Oehl et al. (2005)
Gl. intraradices Dioscorea spp Benin, ouest
d‟Afrique
Tchabi et al. (2008)
Gl. intraradices Artemisia
campestris
Cotes nord de
Zélande, Danemark
Stukenbrock et
Rosendahl (2005)

11
Gl. intraradices Curcuma sp Goa, Inde Radhika et Rodrigues
(2010)
Gl. macrocarpum Ar.a angustifolia San Paulo, Brésil Moreira et al. (2007)
Gl.macrocarpum Gossypium
hirsutum
Benin, ouest
d‟Afrique
Tchabi et al. (2008)
Gl. macrocarpum Mi. champaca L Nord de l‟Inde Panna et Highland
(2010)
Gl. macrocarpum Méliacées Hainan island, China Shi et al. (2006)
Gl. macrocarpum Am. arenaria ssp.
arundinacea
Comporta Portugal Rodríguez-Echeverría
et Freitas (2006)
Gl. macrocarpum Dioscorea spp. Benin Ouest
d‟Afrique
Tchabi et al. (2009).
1.3. Écologie des CMA :
Par leur biotrophie obligatoire, les CMA acquièrent des sites écologiques très
contrastés ; ils influencent ainsi la composition et les successions des communautés végétales
dans les milieux peu ou pas perturbés (Vander Heijden et al., 1998et 2008 ; Kernaghan,
2005). Ils maintiennent la fertilité des sols (Cardoso et al., 2006) , la finesse des arbres et
l‟absorption des nutriments (Jeffries et al., 2007).
L‟absence de spécificité du partenaire végétal des CMA a été souvent démontrée
(Smith et Read, 1997), en dépit de la très forte association entre les deux partenaires. Des
études moléculaires ont confirmé qu‟aux champs les espèces de CMA sont spécifiques à leur
plante hôte (Sanders, 2002 et 2004) parce qu‟elles sont préférentiellement infectées par
certaines espèces de CMA du sol. Cela peut se traduire par la différence d‟infectivité, ou de la
quantité des propagules de chaque espèce dans le sol et /ou par la différence de susceptibilité
des racines à la colonisation de chaque champignon. Ce phénomène est appelé spécificité
écologique (McGonigle et Fitter, 1990). Par ailleurs, Van Tuinen et al. (1998) ont détecté
dans un fragment racinaire d‟un cm de longueur 4 espèces de CMA appartenant à 3 genres
différents.
La connaissance de l‟écologie des CMA est fondamentale pour comprendre la
coexistence des différentes espèces, l‟exploration par ces champignons des ressources
disponibles en nutriments et leur reproduction (Pringle et Bever, 2002; Hart et Klironomos,

12
2002). Ces connaissances ont permis de déterminer les différences dans les familles
d‟espèces.
Les saisons successives et les niches écologiques maintiennent la diversité au sein des
communautés des CMA (Pringle et Bever, 2002). En effet, la sporulation et le développement
ainsi que la mycorhization s‟effectuent pendant la saison froide avec Acaulospora colossica et
la saison chaude avec Gigaspora gigantea (Solaiman et Abbott, 2004).
Chez les Gigasporacées, le genre Scutellospora est plus diversifié que Gigaspora par la
morphologie des spores , la diversité génétique, le nombre d‟espèces décrites et sa présence
dans les écosystèmes naturels ou modifiés par l‟homme. Dans les sols des terrains agricoles
les espèces appartenant aux Gloméracées et aux Acaulosporacées sont plus abondantes dans
ces écosystèmes (Sieverding, 1991) que les Gigasporacées qui résistent moins aux pratiques
culturales (Daniell et al., 2001; Jansa et al., 2002; Vandenkoornhuyse et al., 2002; Oehl et al.,
2003; Johnson et al., 2004).
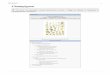
11..44.. TTaaxxoonnoommiiee ddeess CCMMAA
LLaa pprreemmiièèrree cc llaassss iiff iiccaatt iioonn aa éé ttéé éé ttaabb lliiee ppaarr TTaaxxttee rr ((11992222 )) pp uuiiss ee llllee aa ééttéé mmoodd iiffiiééee ppaarr ddee
nnoommbb rreeuuxx mmyyccoorrhhiizzoo lloo ggiisstteess ((ZZyycchhaa,, 11993355 ;; GGoodd ffeeyy,, 11995577 ;; MMoossss,, 11995599 eett 11997700 ;; MMoossss eett
BBoowweenn,, 11996688 ;; GGeerrddeemmaannnn eett TTrraappppee,,11997744 ;;WWaallkkeerr,, 11998855 ;; Walker et Sanders,, 11998866 ;; MMoorrttoo nn
eett BBeennnnyy,,11999900 ;; AAllmmaa iiddaa ee tt Schenck, 11999900 ;; WWaallkkee rr eett TTrraappppee,, 11999933 ;; RReeddeecckkeerr eett aall..,, 22000000 ;;
MMoorrttoo nn ee tt RReeddeecckkeerr,, 22000011 ;; SScchhuubb lleerr eett aall..,, 22000011 ))..
11..44..11.. CCllaassss iiffiiccaatt iioo nn ccllaassss iiqquuee
LLaa ttaaxxoonnoo mmiiee ddeess CCMMAA ééttaa iitt bbaassééee ssuurr llee mmoo rrpphhoottyyppaaggee ddeess ssppoorreess ;; ccee lllleess qquuii oo nntt uunnee
mmoorrpphhoo llooggiiee ss iimmiillaa iirree ee tt ffoorrmmeenntt uunn sseeuull ttyyppee ddee ppaarroo ii ssppoorraa llee ssoonntt aappppaarreennttééeess
pphhyylloo ggéénnéétt iiqquueemmeenntt ((MMoorrttoonn,, 22000022))..
MMoorrttoo nn eett BBeennnnyy ((11999900)) oonntt cc llaa sssséé lleess CCMMAA ddaannss ll‟‟oo rrddrree dd eess GGlloommaa lleess,, ssuubbdd iivviisséé eenn ddeeuuxx
ssoouuss--oorrddrreess GGlloo mmiinnééeess (( ffoorrmmaanntt ddeess vvééss iiccuulleess )) eett lleess GGiiggaassppoo rr iinnééeess..
-- LLeess GGlloommiinnééeess rreeggrroo uuppeenntt ddeeuuxx ggrraannddeess ffaammiill lleess,, lleess GGlloommaaccééeess rreepprréésseennttééeess ppaarr lleess
ggeennrreess GGlloommuuss eett SScclleerrooccyyssttyyss ((ggeennrree éé lliimmiinnéé ppaarr RReeddeecckkeerr eett aall.. ((22000000)) ee tt lleess
AAccaauulloossppoo rraaccééeess ppaarr AAccaauulloossppoorraa eett EEnnttrroopphhoossppoorraa..

13
-- LLeess GGiiggaassppoorr iinnééeess ccoommpprreennnneenntt tt rroo iiss ffaammiilllleess,, lleess GGiiggaassppoo rraaccééeess rreepp rréésseennttééeess ppaarr
GGiiggaassppoorraa eett SSccuutteelllloossppoorraa lleess AArrcchhaaeeoossppoorraaccééeess rreepprréésseennttééeess ppaarr llee ggeennrree AArrcchhaaeeoossppoorraa eett
lleess PPaarraagglloo mmaaccééeess ppaarr PPaarraagg lloommuuss ((MMoorr ttoonn eett RReeddeecckk eerr,, 22000011)),,
LL‟‟ iiddeenntt iiffiiccaatt iioo nn ddeess CCMMAA eess tt bbaassééee ssuurr llaa mmoorrpp hhoo lloo ggiiee ss ttrruuccttuurraa llee ddee llaa ssppoorree,,
cceeppeennddaanntt,, lleess ppooppuullaatt iioo nnss ddeess ssppoorreess iissoo llééeess eett iiddeenntt iiffiiééee àà ppaarr tt iirr ddeess ssoo llss rrhhiizzoosspp hhéérr iiqq uueess nnee
ccoonnss tt iittuueenntt ppaass ttoouujjoouurrss lleess ccoommmmuunnaauuttééss ddeess eessppèècceess ddee CCMMAA qquuii iinnffeecc tteenntt lleess rraacc iinneess
((SSaannddeerrss,, 22000044)).. LLeess ffaa iibb lleess vvaa rr iiaa tt iioonnss mmoorrpphhoo llooggiiqquueess eennttrree llee ss ssppoorreess rreenndd ssoouuvveenntt
ddiiffff iicc iillee lleeuurr iiddeenntt iiffiiccaa tt iioonn.. DDeess éé ttuuddeess ccoommpp lléémmeennttaa iirreess ddee ccaa rraaccttéé rr iissaatt iioonn mmoo llééccuullaa iirree
ddooiivveenntt êê ttrree ee ffffeeccttuuééeess ppoouurr aannaa llyyssee rr llaa vvaa rr iiaabb iilliittéé ggéénnéé tt iiqquuee ddeess CCMMAA..
LLeess CCMMAA aappppaarr tt iieennnneenntt aauu pphhyylluumm ddee GGlloommeerroommyyccoottaa,, ccee ddeerrnniieerr eess tt ccoonnsstt iittuuéé dd‟‟uunnee
sseeuullee cc llaassssee,, ccee llllee ddeess GGlloommeerroommyyccèètteess qq uuii eesstt ssuubbdd iivviissééee eenn qq uuaattrree oorrdd rreess,, dd iixx ffaammiilllleess eett
ttrree iizzee ggeennrreess (( ffiigg..99)).. LLaa ttaaxxoonnoo mmiiee bbaassééee ssuurr llee mmoorrpp hhoo ttyyppaaggee ddeess ssppoorreess ppeerrmmeett dd iisstt iinngguueerr
lleess qquuaattrree oorrddrreess..
11..22..11..11-- LLeess DDiivveerrss iissppoorraalleess
LLeess mmeemmbbrreess aappppaarrtteennaanntt àà ccee tt oorrddrree ffoorrmmeenntt uunnee mmyyccoorrhhiizzee eexxeemmpp ttee ddee vvééss iiccuulleess
((WWaallkkeerr ee tt Schüßler,, 22000044 )),, eenn rreevvaanncchhee llee mmyyccéé lliiuu mm eexxttrraammaattrr iicc iiee ll ffoo rrmmee ddeess ccee lllluu lleess
aauuxxiilliiaa iirreess eett ddeess ssppoorreess (( ffiigg..11))..
FFiigg..11 :: LLeess cceelllluulleess aauuxxiilliiaaiirreess ((aauuxx)) eett ss ppoorreess ((ss pp)) ffoorrmmééeess ppaarr llee mmyyccéé lliiuumm eexxttrraammaattrriicciiee ll ddee Scutellospora rubra (Sturmer et Morton, 1999).(flèches : mycélium extra-racinaire)
LLeess ssppoo rreess ssee ffoorrmmeenntt ssoo iitt àà ll‟‟ iinnttéérr iieeuurr dd ‟‟uunn ssaacc ssppoo rr iiffèèrree cchheezz KKuukk lloossppoorraa oouu de
manière intercalaire entre la saccule et l'hyphe suspenseur cchheezz EEnnttrroopphhoossppoorraa (( ffiigg..22))
((SS iieevveerrdd iinngg eett OOeehhll,, 22000066 )) oo uu llaa ttéérraa lleemmeenntt ssuurr llee ssaacc cchheezz AAccaauulloossppoo rraa (( ffiigg..33)) ee tt OOttoossppoorraa
((MMoorrttoonn ee tt BBeennnnyy,, 11999900)) oouu ssuurr uunn ppéédd iiccee llllee aattttaacchhééeess àà ll‟‟hhyypphhee ppoorr ttaanntt llee ssaacc cchheezz

14
AAmmbbiissppoorraa (( ffiigg..44 )) ((Walker et al., 2007b). EElllleess ssee ffoo rrmmeenntt aauussss ii aauu bboo uutt ddee ll‟‟hhyypphhee
ssppoorroo ggèènnee ssuurr uunnee ccee lllluullee ssppoo rrooggèènnee bb uullbbeeuussee cchheezz GGiiggaassppoo rraa eett SSccuutteell lloossppoorraa (( ffiigg.. 55 AA))
((BBeenntt iivveennggaa ee tt MMoorr ttoonn,, 11999955)) oouu ssuurr ll‟‟hhyypp hhee ssuussppeennsseeuurr cchheezz DDiivveerrcciissppoorraa eett PPaacc iissppoorraa
((Schübler eett aall.. ,, 22000011 ;; WWaallkkee rr eett Schübler,, 22000044 ))..
Fig. 2 : Spore et saccule sporifère d'Entrophospora infrequens ((BB llaass zzkkoowwsskkii,, 22000033))..
Fig. 4: Aspect d’une spore d’Ambispora fennica (Walker et al., 2007b).
LLaa ppaarroo ii ddeess ssppoorreess jjuuvvéénniilleess ddee GGiiggaassppoorraa eesstt ffoo rrmmééee ddee ddeeuuxx ccoouucchheess àà ééppaa iisssseeuurrss
ééggaa lleess,, lloorrssqquu‟‟ee lllleess ssoonntt ââggééeess,, ll‟‟aapppp aarr iitt iioonn ddee llaa ppaa rroo ii ggee rrmmiinnaatt iivvee ppeerrmmeett ll‟‟eexxttrruuss iioonn dd uu
ttuubbee ggeerrmmiinnaatt iiff..
LLeess ssppoorreess dd‟‟AAccaauulloossppoorraa oonntt uunnee ppaa rroo ii ffoorrmmééee dd‟‟uunnee sseeuullee ccoouucchhee ccoonntt iinnuuee aavveecc llaa
ppaarroo ii ddee ll‟‟hhyypphhee qquuii ppoo rrttee llee ssaacc ssppoo rr iiffèèrree..
Saccule Sporifère Spore
Pédicelle
Saccule
Sporifèr
e Spore
Fig. 3: Spore et saccule sporifère
d'Acaulospora rehmii ((BBllaasszzkkoowwsskkii,, 22000033))..

15
LLaa ggee rrmmiinnaatt iioo nn ddeess ssppoorreess ss ‟‟aaccccoommppaaggnnee ppaarr llaa ffoorrmmaatt iioo nn ddee «« SS hhiiee lldd »» ((Bouclier de
germination) cchheezz SSccuutteelllloossppoorraa (fig.5B) eett PPaacciissppoorraa oouu ddee «« OOrrbb »» cchheezz AAccaauu lloossppoorraa eenn
ggeerrmmiinnaa tt iioo nn (Sturmer et Morton, 1999 ; Jeffries et al., (2007) ..
Fig.5 :Caractéristiques du genre Scutellospora sp.
A : Aspect de la ccee lllluullee ssppoorrooggèè nnee (sc) et des différentes couches (iw1, iw2, 1, 2, sw (2) ,de la paroi d’une spore de Scutellospora rubra ((Sturmer et Morton, 1999) ;; BB :: AAss ppeecctt dduu bouclier de
germination ((BBGG)) «« SShhiiee lldd »» ddee Scutellospora heterogama (Jeffries et al., (2007).
11..22..11..22-- LLeess GGlloomméérraallee ss
LLeess cchhaammpp iiggnnoonnss aappppaarr tteennaanntt àà ll‟‟oorrddrree ddeess GGlloomméérraa lleess ssoo nntt hhyyppooggééss rraa rreemmeenntt éépp iiggééss,,
iillss ffoorrmmeenntt ddeess aa rrbbuussccuulleess,, ddeess vvééss iiccuulleess ee tt ddeess ssppoorreess iinntteerrccaa llaa iirreess oo uu tteerrmmiinnaa lleess àà llaa ffiinn dd‟‟uunn
hhyypphhee ssuussppeennsseeuurr ((DDeecc lleerrcckk eett aall..,, 22000000)) de 18-50 µm de diamètre chez Glomus mosseae,
(Sharma et al., 2008),, ll‟‟hhyypphhee ssppoorrooggèènnee ssee ddéévvee llooppppee àà ppaarr tt iirr dduu mmyyccéé lliiuu mm eexxttrraarraacc iinnaa iirree ddeess
rraacc iinneess mmyyccoorrhhiizzééeess..
LLeess ssppoo rreess ssoo nntt ssoo iitt ss iinngguull iièèrreess (( ffiigg..66 )),, eenn ggrraappppeess oouu eenn ssppoorroo ccaarrppeess (regroupement de
spores en une masse plus ou moins compacte),, mmuunniieess dd‟‟uunn ppéé rr iidd iiuumm (collerette
membraneuse protectrice enveloppant une masse de spores et permettant de différencier les
spores de quelques CMA) ((GGiioovvaanneett tt ii ee tt Salutini,, 11999911)).. EElllleess oo nntt uunnee sseeuullee paroi sporale (13
à 26 µm de diamètre) continue avec la paroi de l‟hyphe suspenseur et formée d‟une à deux
couches à structure laminaire (Blaszkowski et Czerniawska, 2008).
B
A
BG

16
FFiigg.. 66:: AAss ppeecctt ddee Glomus intraradices ((BB aaggoo eett CCaannoo ,, 22000055)).. FFllèècchhee :: ppaarrooii ss ppoorraallee..
11..22..11..33-- LLeess PPaarraagglloo mmaalleess
LLeess eessppèècceess aappppaa rrtteennaanntt àà ll‟‟oorrddrree ddeess PPaarraagglloo mmaa lleess ((MMoorrttoo nn ee tt RReeddeecckkeerr,, 22000011 )) ffoorrmmeenntt
ddeess aarrbbuussccuulleess,, rraa rreemmeenntt ddeess vvééss iiccuulleess eett ddeess ssppoorreess ss iinngguulliièè rreess ddaannss llee ssoo ll rreesssseemmbb llaanntt àà
ccee lllleess ddeess GGlloomméérraa lleess mmaa iiss ssoonntt iinnccoo lloorreess ((FF iigg..77)).. LLaa ppaa rroo ii ssppoo rraa llee eesstt ccoonnss tt iittuuééee ddee ddeeuuxx àà
ttrroo iiss ccoouucchheess ccoo nntt iinnuueess aavveecc ccee lllleess ddee ll‟‟hhyypphhee ssuussppeennsseeuurr,, llaa ccoouucchhee ssuuppeerr ffiicc iiee llllee ssee ddééttéé rr iioo rree
eett ssee ddééggrraaddee aavveecc ll‟‟ââggee..
FFiigg..77 :: AAssppeecctt dd’’uunnee ssppoorree ddee PPaarraagglloommuuss sspppp.. ((BBllaasszzkkoowwsskkii,, 22000033))..
11..22..11..44 LLeess AArrcchhaaeeoossppoorraa lleess
LLeess cchhaammpp iiggnnoonnss qquuii ffoorrmmeenntt ll‟‟oorrddrree ddeess AArrcchhaaeeoossppoo rraa lleess ((MMoorrttoonn eett RReeddeecckkeerr,, 22000011))
ffoo rrmmeenntt ddeess aarrbb uussccuulleess eett rraarreemmeenntt ddeess vvééss iiccuulleess ((SSppaa iinn eett aall..,, 22000066 ;; WWaallkkee rr eett aall..,, 22000077bb)) ..
LLeeuurr ssppoorree,, iiddeenntt iiqq uueess àà ccee lllleess dd‟‟AAccaauu lloossppoorraa eett ddee GGlloommuuss,, ssoo nntt iinnccoo lloorreess eett nnee rrééaaggiisssseenntt
ppaass aauu rrééaacctt iiff ddee MMeellzzeerr (( ffiigg.. 88)) ;; ee lllleess ssoo nntt ss iinngguulliièèrreess oouu eenn ggrraappppeess,, ee lllleess ppoossssèèddeenntt uunnee
ppaarroo ii ssppoorraa llee eett uunnee ppaarroo ii ggee rrmmiinnaatt iivvee ddoo nntt cchhaaccuunnee ccoo nntt iieenntt ddeeuuxx àà tt rroo iiss ccoouucchheess ((MMoorr ttoonn eett
RReeddeecckkeerr,, 22000011 ;; SSppaa iinn,, 22000033 ;; SSppaa iinn eett aall..,, 22000066 ;;SS iieevvee rrdd iinngg ee tt OOeehhll,, 22000066 ))..

17
FFiigg.. 88 :: AAss ppeecctt dd’’uunnee ss ppoorree dd’’AArrcchhaaeeoossppoorraa sspppp.. ((BB llaass zzkkoowwsskkii,, 22000033))..
11..44..22.. CCllaassss iiffiiccaatt iioo nn mmoollééccuullaaiirree
LLee pprr iinncc iippee ddee ll‟‟aapppprroocchhee mmoo llééccuullaa iirree eesstt bbaasséé ssuurr ll‟‟aammpp lliiff iiccaatt iioonn iinn vv iittrroo ddee ll‟‟AADDNN ddeess
rrééggiioonnss vvaa rr iiaabb lleess dd uu ggéénnoommee ddeess cchhaammpp iiggnnoonnss àà ppaarr tt iirr ddeess ssppoo rreess ((SScchhwwaarrzzootttt eett Schübler,,
22000011)) oouu àà ppaa rrtt iirr ddeess rraacc iinneess mmyyccoorrhhiizzééeess ((RReeddeecckkeerr eett aall..,, 22000000 ;; MMaarr tt iinn eett aall..,, 22000077 ;;
AAllgguuaacc iill,, 22000099))..
LLaa pp hhyyllooggéénniiee ddeess GGlloo mmeerroo mmyyccoottaa ((Schübler eett aall..,, 22000011)) eesstt eenntt iièè rreemmeenntt ccoonnsstt rruuiittee ssuurr llaa
bbaassee dd‟‟uunn sseeuull ggèènnee ((ggèènnee ccooddaanntt ppoouurr ll‟‟AARRNNrr 1188SS)).. LL‟‟aannaa llyyssee mmoo llééccuullaa iirree ddééttaa iill llééee dd uu
ggéénnoommee ddee GGlloommuuss eett ccee llllee dd uu ggèènnee SSSS UU ccooddaanntt ppoo uurr ll‟‟AARRNNrr.. aa ppeerrmmiiss ddee ccoonnnnaa îîttrree llaa
ttaaxxoonnoo mmiiee eett llaa pphhyyllooggéénniiee ddeess GGlloommeerroommyyccoottaa ((SScchhwwaarrzzoott tt eett aall.. ,,22000011)).. LL‟‟aammpp lliiffiiccaatt iioonn llaa
rrééggiioonn IITTSS ((IInnttee rrnnaa ll TTrraannssrr iippeedd SSppaaccee rr)) rreepprréésseennttaanntt lleess ppoo llyymmoorrpphhiissmmeess ddee ssééqquueenncceess ((DDiioopp
eett aall.. ,, 11999944)) aa ppeerrmmiiss ddee ddéétteecc tteerr llaa vvaarr iiaabb iilliittéé ggéénnéétt iiqquuee ddeess cchhaammpp iiggnnoonnss mmyyccoo rrhhiizziieennss
((HHeennrr iioonn eett aall..,, 11999944))..
LL‟‟aannaa llyyssee mmuulltt iiggéénniiqquuee aa éé ttéé uutt iill iissééee ppoouurr iiddeenntt iiffiiee rr lleess CCMMAA ((RReeddeecckkeerr eett RRaaaabb,,
22000066)) ppoouurr ccoo nnffiirrmmeerr llaa sstt rruuccttuurree pp hhyylloo ggéénnéé tt iiqquuee pp rrooppoossééee ppaarr Schübler eett aall.. ((22000011)).. LLaa
ddiivveerrss iittéé dd eess CCMMAA ddaannss lleess ssoo llss rrhhiizzoosspp hhéé rr iiqquueess aa éé ttéé éé ttuudd iiééee ppaa rr aannaa llyyssee mmoo llééccuullaa iirree
((AAllgguuaaccyyll eett aall..,, 22000099))..
PPaarr aa iilllleeuurrss,, ddeess aapppprroocchheess bb iioocchhiimmiiqquueess bbaassééeess ssuurr ll‟‟eemmpp lloo ii dd ‟‟aanntt iiccoorrppss ssppéécc iiffiiqquueess
((HHaahhnn eett aall..,, 11999933)),, ddeess iissooeennzzyymmeess oouu ddeess pprroo ffii llss lliipp iidd iiqq uueess (( MMaaddaann eett aall..,, 22000022 )),, ssoo nntt
ééggaa lleemmeenntt uutt iill iissééeess ppoouurr iiddeenntt iiffiieerr lleess CCMMAA..
Suspenseur

18
AAccttuuee lllleemmeenntt,, lleess CCMMAA aappppaarrtt iieennnneenntt aauu pphhyylluumm ddeess GGlloommeerroommyyccoottaa,, ccee ddeerrnniieerr eesstt
ccoonnss tt iittuuéé dd‟‟uunnee sseeuullee cc llaassssee,, ccee llllee ddeess GGlloomméérroommyyccèè tteess qquuii ee sstt ssuubbdd iivviissééee eenn qquuaa ttrree oorrdd rreess,,
ddiixx ffaammiillllee ss eett ttrree iizzee ggeennrreess (( ffiigg..99)).. CCeett ttee cc llaassss iiff iiccaatt iioo nn,, bbaassééee ssuurr ccee llllee ddee Schübler eett aall..
((22000011)),, aa éé ttéé mmoodd iiffiiééee ppaarr ddee nnoo mmbbrreeuuxx aauutteeuurrss ((Walker et Schübler ,2004 ; Oehl et
Sieverding,, 22000044 ;; Sieverding eett Oehl, 2006 ; Spain et al. 2006 ; Walker et aall..,, 22000077aa ee tt bb;;
PPaalleennzzuuee llaa eett aall..,, 22000088 ))..
LL‟‟eessppèèccee ffoonnggiiqquuee GGiioossiiffoonn ppyyrriiffoorrmmiiss,, ccaa rraaccttéé rr iissééee ppaarr Schübler et Wolf (2005), est
une eennddoossyymmbb iioo ttee ddeess CC yyaannoobbaaccttéé rr iiee ss ((NNoossttoocc sspp..)) ;; ee llllee aappppaarr tt iieenntt àà ll‟‟oo rrddrree GGlloommeerroommyyccoottaa
eett àà llaa ffaammiillllee ddeess GGeeoossiipphhoonnaaccééeess ((AArrcchheeoossppoorraalleess)) ppaarr sseess ss iimmiill iittuuddee mmoo llééccuullaa iirreess (Weber,
2009).

19
FFiigg.. 99 :: CCllaassss iiffiiccaattiioonn ddee SShhuubblleerr eett aall .. ((22000011)) mmooddiiff iiééee
((SS iieevveerrdd iinngg,, 22000044 ;; WWaallkkeerr ee tt SScchhuubb llee rr,, 22000044 ;;SS iieevveerrdd iinngg eett OOeehhll,, 22000066 ;; SSppaa iinn eett aall..,,22000066 ;;WWaallkkee rr eett
aall..,,22000077aaeettbb ;; PPaa lleennzzuuee llaa,, 22000088))
RRèèggnnee :: CC hhaammpp iiggnnoonn
PPhhyylluumm :: GGlloommeerroo mmyyccoottaa
CCllaassssee ::GGlloomméérroommyyccèètteess
OOrrddrree ::
AArrcchhaaeeoossppoorraalleess
PPaacciissppoorraaccééeess GGiiggaassppoorraaccééeess DDiivveerrcciissppoorraaccééeess AAccaauulloossppoorraaccééeess EEnnttrroopphhoossppoorraaccééeess
PPaarraagglloommaa lleess GGlloomméérraalleess
GGlloomméérraaccééeess PPaarraagglloommaaccééeess
DDiivveerrcciissppoorraa OOttoossppoorraa GGlloommuuss PPaarraagglloommuuss PPaacciissppoorraa
KKuukklloossppoorraa SSccuutteelllloossppoorraa GGiiggaassppoorraa AAccaauulloossppoorraa
AArrcchhaaeeoossppoorraa IInnttrraassppoorr
aa
OOttoossppoorraa
GGlloommuuss
AAmmbbiissppoorraaccééeess GGééoossiipphhoonnaaccééeess
GGee nnrree
EEnnttrroopphhoossppoorr
aa
AAmmbbiissppoorraa GGééoossiipphhoonn
AArrcchhaaeeoossppoorraaccééeess
DDiivveerrssiissppoorraallee
ss
FFaammiillllee

20
1.5. Cycle de vie des CMA et associations symbiotiques
LLee ccyyccllee ddee vviiee ddeess mmyyccoossyymmbbiiootteess aarrbbuussccuullaaiirreess nn‟‟eesstt ccoommppllééttéé qquu‟‟eenn pprréésseennccee ddee llaa
ppllaannttee hhôôttee,, dduurraanntt lleess pprroocceessssuuss ddee ffoorrmmaattiioonn ddee cceettttee ssyymmbbiioossee ((GGiioovvaannnneettttii eett aall..,, 22000000)) llaa
ppllaannttee «« aacccceeppttee »» llaa ccoolloonniissaattiioonn dduu cchhaammppiiggnnoonn ssaannss rrééaaccttiioonn ddee rreejjeett ((DDuummaass--GGaauuddoott eett aall..,,
22000000)) eett uunnee sséérriiee dd‟‟iinntteerraaccttiioonnss rraacciinneess--cchhaammppiiggnnoonn mmèènnee àà ll‟‟iinnttééggrraattiioonn ddeess ddeeuuxx
oorrggaanniissmmeess ..
LL‟‟ééttaabblliisssseemmeenntt ddee llaa ssyymmbbiioossee eesstt llee rrééssuullttaatt dd‟‟uunn «« ddiiaalloogguuee »» mmoollééccuullaaiirree ccoonnttiinnuuee
eennttrree llaa ppllaannttee eett llee cchhaammppiiggnnoonn àà ttrraavveerrss ddeess éécchhaannggeess ddee ssiiggnnaauuxx ddee rreeccoonnnnaaiissssaannccee eett
dd‟‟aacccceeppttaattiioonn ((VViieerrhheeiilliigg eett PPiicchhéé,, 22000022)) àà ll‟‟aaiiddee dd‟‟eexxssuuddaattss rraacciinnaaiirreess ((Akiyama,, 22000077;;
RReeiinnhhaarrddtt,, 22000077))..
EEnn rreevvaanncchhee,, llee mmooddèèllee dduu ccyyccllee ddee ccoolloonniissaattiioonn ddeess CCMMAA aa ééttéé bbaasséé ssuurr qquueellqquueess
eessppèècceess ddee GGlloommuuss ((DDoodddd eett aall..,, 22000000 ;; DDaallppéé eett aall..,, 22000055bb ;; HHaauussssee eett ffoosstteerr,, 22000055 ;; Bago et
Cano, 2005;; Balestrini et Lanfranco, 2006 ; Novero et al., 2008 ; BBoonnffaannttee eett aall..,,22000099 )),, eett lleess
GGiiggaassppoorraaccééeess ((ddee SSoouuzzaa eett aall..,, 22000055))..
1.5.1. Stade non symbiotique
1.5.1.1. Germination des propagules
Les propagules capables de germer en un mycélium et d‟initier une association
mycorhizienne sont les spores (intra-racinaires ou rhizosphériques), les vésicules et le
mycélium (intra-racinaire ou attaché à un péridium).
- La spore
La germination des spores des Gloméracées s‟effectue lorsque la paroi sporale interne
est forcée par le tube germinatif à partir de l‟hyphe suspenseur (de Souza et berbara, 1999) qui
traverse directement la paroi sporale (Tummerup et Kidby, 1980) ou la paroi de l‟hyphe
suspenseur (Giovanetti et al., 1991) et donne naissance à un ou plusieurs tubes germinatifs
droits à parois épaisses (fig. 10).

21
FFiigg.. 1100 :: GGee rrmmiinnaattiioonn dd’’uunnee ssppoorree ddee G. intraradices (Dalpé et al., 2005b).
TG : Tube germinatif sortant de la paroi de l‟hyphe suspenseur observé au MEB Susp : Suspenseur ;
Barre=7µm.
Dans le cas des Gigasporacées et des Acaulosporacées, les tubes germinatifs sortent
directement de la paroi sporale à l‟intérieur de laquelle se forme la paroi germinative et à
partir de celle-ci se différencient les différentes structures de germination de chaque espèce.
Les espèces appartenant au genre Scutellospora germent en formant un bouclier de
germination, localisé dans la couche interne de la paroi germinative de la spore, qui permet
la sortie des tubes germinatifs (Oehl et al., 2008), tandis que Chez Kuklospora et Acaulospora
la couche interne de la paroi germinative développe une forme sphérique appelé en anglais
« germination orb» (Spain, 1992) d‟où sortent les tubes germinatifs et qui persiste même
après la germination.
La germination des spores et des sporocarpes s‟effectue généralement entre 2 à 30 jours
en culture monoxénique, celle de Glomus sp. , après une phase de dormance d‟une durée de
quelques jours à 6 mois, le niveau de germination atteint 2 à 10%. La
germination de la spore nécessite de l‟humidité et l‟activation du métabolisme, durant laquelle
il y a une synthèse de l‟ADN et une réplication active du ou des noyaux multiples dans la
spore (Bianciotto et Bonfante, 1993), le contenu lipidique atteint 40-60% (Sancholle et al.,
2001), une synthèse des stérols, du diacylglycérol, des phospholipides et des acides gras
libres (Sancholle et al., 2001) et la consommation des triacylglycérides ( Gaspar et
al.,1994).
Généralement, il existe des facteurs qui activent la germination des spores, tels que le
pH neutre (Pons et al.,1984 ; Gunasekaran et al.,1987), la température optimale 20-30°C
(Sheikh et Sanders, 1988), les flavonoïdes, des faibles concentrations de phosphore, de
glucose et d‟acide galacturonique (Siquiera et Hubell, 1984), 50% de saccharose et de la
Susp TG

22
thiamine (Vilarino et Saiz, 1997). Cependant, d‟autres facteurs peuvent inhiber la
germination telle qu‟une extrême sécheresse ou un excès d‟humidité et l‟acidité du sol
(Siquiera et al., 1985).
De nombreux CMA sont incapables de produire des spores par contre, les formes intra-
racinaires dans les fragments de racines mycorhizées par ces champignons ont un potentiel de
germination non négligeable.
- Les vésicules
La germination des vésicules s‟effectue (Diop, 1994 ; Strullu et al., 1997 ; Dalpé et al.,
2005b) à travers l‟hyphe attaché à la vésicule (Declerck et al., 1998), le tube germinatif
produit génère des hyphes ramifiés similaires à ceux formés par la spore .
- Les hyphes extra et intra-racinaires
Quelques genres de Glomeromycota ne forment pas de vés icules, la germination
s‟effectue par les hyphes intra et extra racinaires qui germent et forment le pro-mycélium (
fig. 11) (de Souza et al., 2005) .
Fig. 11 : Développement du mycélium extra-racinaire (flèches) à partir d’un fragment racinaire
mycorhizé, en culture axénique (Bago et Cano, 2005).
- Croissance du pro-mycélium
Le tube germinatif se différencie en mycélium présymbiotique ou promycélium, sa
croissance dépend du contenu lipidique de la propagule (Sancholle et al., 2001) ; le
protoplasme de la spore contient toutes les organites requises pour assurer son développement
( Meier et Chavat,1992 ; Bananomi et al., 2009). Les grosses spores contiennent plus de

23
réserves nutritives (50% constituées par des lipides et plus particulièrement par des
triglycérides), elles peuvent tenter plusieurs germinations, et maintenir la croissance du
mycélium et son métabolisme lorsqu‟elles cherchent l‟hôte. Le mycélium généré se
différencie en des structures diverses les VLS (Vesicule like structure)( Budi et al., 1999) qui
ressemblent au vésicules et les BAS ( Branched Absorbing structures )(Bago et al., 1998).
Les VLS sont des formes globulaires qui ressemblent à des spores en miniatures, de 20
à 100 µm de diamètre, hyalines à paroi mince (fig.12) (Strullu et Romand, 1987), longtemps
considérées comme des spores juvéniles ; leur rôle est de permettre au mycélium néoformé de
survivre plus longtemps avant le contact avec la racine de l‟hôte (Dalpé et al., 2005b).
Fig. 12 : Aspect d’une VLS intercalaire (Dalpé et al., 2005b).
Les BAS sont des ramifications dites « absorbantes » à paroi mince, éphémères, leur
durée de vie et de 5 à 7 jours sauf celles qui sont liées aux spores, elles ont le rôle d‟absorber
les nutriments (Mosse, 1988) et confèrent au champignon le potentiel de croître en
saprophyte (Dalpé et al., 2005a).
Le promycélium se ramifie un grand nombre de fois et forme des hyphes « runner » qui
explorent le milieu (Declerck et al., 2000). La faible exposition à la lumière (Nagahashi et al.,
2000), le contenu sporale (Hepper, 1984), les exsudats et les substances volatiles racinaires
(Giovanetti et al., 1993), les flavonoïdes et les phénols Kaempferols, myricetine et quercetine
(Vierheilig et al., 1998 ; Buee et al., 2000), les acides aminés (Hepper et Jakobsen, 1983) et la
thiamine (Siquiera et al., 1982) ont un pouvoir de ramification de ces hyphes. Dans le cas
d‟absence de contact ou de détection de signaux provenant de la plante hôte, la croissance du
tube germinatif s‟arrête après quelques jours (Bécard et Piché, 1989).
1.5. 2. Stade pré-symbiotique

24
- Les signaux moléculaires
Les signaux racinaires préviennent le champignon de l‟existence d‟un hôte potentiel et
par conséquent d‟une source de carbone organique (Smith et al., 2006 ) et discriminent les
plantes hôtes des plantes non hôtes. D‟après Thomas et al. (2003), des éléments isolés à
partir d‟exsudats de racines transformées de carottes améliorent la respiration durant le stade
de pré-germination et stimulent la ramification des hyphes (Buee et al., 2000 ; Nagashi et
Douds, 2000) ( fig. 13).
Fig. 13 : Les différents signaux moléculaires synthétisés par les deux symbiotes dduurraanntt 44 pphhaasseess
((11,, 22,, 33,, 44)) ddee ll’’aassssoocciiaattiioonn ((BBoonnaannoommii eett aall..,, 22000099)).. 11 ::
GGeerrmmiinnaattiioonn ddee llaa ssppoorree eett ccrrooiissssaannccee ddeess hhyypphheess ;; 22 :: RRaammiiffiiccaatt iioonn ddeess hhyypphheess ;; 33 ::
DDéévveellooppppeemmeenntt ddeess aapppprreessssoorriiaa;; 44 :: ccrrooiissssaannccee iinnttrraa--rraacciinnaaiirree eett ffoorrmmaattiioonn ddeess aarrbbuussccuulleess ..
Les composés qui stimulent la ramification des hyphes et améliorent l‟activité
respiratoire dans les cellules sont désormais connus : ce sont les strigolactones
(phytohormones), qui ont été identifiés par Akiyama et al. (2005). Selon ces auteurs, la
production de 5- dioxy-strigolactone dans les racines de Lotus japonicum induit la
ramification des hyphes de Gigaspora margarita, ces molécules sont efficaces même à très
faible concentration (Besserer et al., 2006 ; Smith et al., 2006 ; Chen et al., 2009). D‟autres
strigolactones sont aussi efficaces à très faible concentration: les sorgolactones, les strigols,
les alectrils et les orobanchols, et leurs analogues GR 7et GR 24 ( Akiyama et Hayashi,
2006 ).
1
--CCOO22
--FFllaavvoonnoo iiddeess
--AAuuttrreess pphhéénnoo llss
3
-- SSiiggnnaauuxx
iinnccoonnnnuuss
4
--LLeess
pphhyyttooaa lleexxiinneess
2
- Les
strigolactones

25
Certains auteurs oo nntt ddéémmoonnttrréé qquuee llaa ccaa rreennccee eenn pphhoosspphhaattee aauuggmmeennttee llaa pprroodduucc tt iioonn
ddeess ssttrr iiggoo llaaccttoonneess ddaannss lleess eexxssuuddaa ttss rraacc iinnaa iirree ss eett ppaarr ccoonnssééqq uueenntt aamméé lliioorree llaa rraammiiffiiccaa tt iioonn ddeess
hhyypp hheess :: ccaass cchheezz llaa ttoommaattee ((LLóóppeezz--RRááeezz eett aall..,, 22000088)) eett llee mmee lloo nn ((Akiyama eett aall.. 22000022))..
DD‟‟aauuttrreess aauutteeuurrss oonntt confirmé que la présence de signaux moléculaires diffusibles dérivés du
champignon (Kosuta et al., 2003) auraient le pouvoir de stimuler la formation des racines
latérales chez la plante (Olah et al., 2005).
Selon Weidmann et al. (2004), des gènes de plantes sont régulés par les exsudats
hyphales lorsque les partenaires ne sont pas en contact physique direct. Des travaux sur la
tomate ont démontré que les CMA peuvent induire durant le stade pré-symbiotique
l‟expression des gènes de la plante impliqués dans la modification de la paroi cellulaire, la
réparation des tissus, les réponses aux attaques pathogènes et au stress oxydatif, 48h après
exposition de la plante à l‟inoculum fongique (Demartsev et al., 2005). Ainsi, le
champignon signale à la plante de préparer la voie de pénétration intracellulaire (Genre et al.,
2005).
1.5.3. Stade symbiotique
* Pénétration
Le contact entre les deux partenaires s‟effectue après la ramification des hyphes qui ssee
ddiifffféé rreenncc iieenntt eenn aapppprreessssoorr iiuumm (( ffiigg.. 1144 ))..
FFiigg.. 1144 :: AAss ppeecctt dd’’uunn aapppprreessssoorriiuumm ((AApppp)) àà ll’’iinnttéé rriiee uurr dd’’uunnee rraacciinnee mmyyccoorrhhiizzééee ((sseelloonn ((DDaallppéé eett
aall.. ,, 22000055bb))..
App

26
LL‟‟aapppp rreessssoorr iiuumm oouu hhyypp hhooppoodd iiuumm ::
CC‟‟eesstt uunn ggoonnfflleemmeenntt ddeess ccee lllluulleess àà ll‟‟eexxttrréémmiittéé dd‟‟uunn hhyypp hhee oouu dd‟‟uunn ttuubb ee ggeerrmmiinnaatt iiff
((BBllaasszzkkoowwsskk ii eett aall ..,, 22000033 )),, qquuii ssee ffoorrmmee eennvviirroonn 3366 hh aapprrèèss llee ccoonnttaacctt aavveecc lleess ccee lllluu lleess
éépp iiddeerrmmiiqquueess ((DDaa llppéé eett aall..,, 22000055bb)).. CCeett ttee sstt rruuccttuurree rreepprréésseennttee uunn ss iiggnnee ddee rreeccoonnnnaa iissssaannccee
eennttrree llee cchhaammpp iiggnnoonn eett lleess rraacc iinnee dd‟‟uunnee pp llaannttee ccoommppaa tt iibb llee ((RReeqq uueennaa eett aall..,, 22000077 )),, iill nnee ssee
ffoo rrmmee jjaammaa iiss ssuurr lleess rraacc iinneess dd‟‟uunnee pp llaannttee nnoo nn hhôôttee ((eexx :: BBrraassss iiccaaccééeess )) oouu ssuurr uunnee mmeemmbbrraannee
oouu uunn mmiill iieeuu ssyynntthhéétt iiqquuee eenn ll‟‟aabbsseennccee ddee rraacc iinneess (( HHaarr iissssoonn,, 11999977 ))..
Le processus de pénétration implique la participation active des cellules de la plante
hôte, la colonisation par les CMA provoque des modifications dans l‟organisation des cellules
de l‟hôte végétal (Bonfante, 2001) .En effet, la pénétration dans les cellules épidermiques
sous l‟effet des enzymes telles que les hydrolases et d‟autres molécules qui probablement
rendent la paroi cellulaire plus flexible (Garcia-Romera et al., 1991 ; Peretto et al. 1995),
provoque le changement de la position du noyau qui est positionné sous l‟appressorium, des
réarrangements des cytosquelettes et du réticulum endoplasmique, et la formation de
l‟appareil de pré-pénétration (PPA) 4 à 5h après la formation de l‟appressorium (Genre et al.,
2005). Cet appareil facilite le passage du champignon à travers les cellules épidermiques et
selon certains auteurs, il est responsable de la formation de l‟interface symbiotique et d‟un
nouveau compartiment apoplastique séparant les deux symbiotes (fig. 15).
RE
Champ
Surface
racinaire
C E
Champ
Ny
MT
MF
RE

27
FFiigg.. 1155 :: DDéévvee llooppppee mmee nntt ddee ll’’aappppaarree iill ddee pprréé --ppéénnéé ttrraattiioonn ((PPPPAA,, éé ttaappeess AA––EE)) ddaannss lleess cceelllluulleess
éé ppiiddee rrmmiiqquueess rraacciinnaaiirreess ee nn ccoonnttaacctt aavveecc uunn cchhaammppiiggnnoonn mmyyccoorrhhiizziiee nn aarrbbuussccuullaaiirree ((BBoonnffaannttee eett
aall.. 22000099))..
A : Cellule végétale; B: Adhésion de l‟hyphopodium; C: réorganisation des cytosquelettes, Repositionnement du noyau au site de contact et accumulation de réticulum endoplasmique (RE) ; D :
une seconde migration du noyau à travers la cellule est associée avec la formation de la colonne cytoplasmique ; E : Pénétration fongique par l‟hyphe à travers la voie du PPA . MT : Microtubules ;
MF : Micro filaments ; Ny : Noyau ; Champ : Champignon.
DDee nnoo mmbbrreeuuxx aauutteeuurrss oonntt ddéétteeccttéé uunnee ffaa iibb llee aacctt iivvaatt iioo nn ddeess ggèènneess ddee ddéé ffeennssee cchheezz llaa
ppllaannttee dd uurraanntt llaa ffoorrmmaatt iioo nn ddee ll‟‟aapppprreessssoorr iiuumm ((RRuuiizz-- LLoozzaannoo eett aall..,, 11999999 ;; BBllii lloouu eett aall..,, 22000000 ;;
BBoonnaannoo mmii eett aall..,, 22000099)).. LLaa ppéénnééttrraatt iioo nn ddeess hhyypphheess ddaannss llaa ppaarroo ii ccee lllluullaa iirree eess tt ppaarr ffoo iiss
aaccccoo mmppaaggnnééee ppaarr ll‟‟aa llttéérraa tt iioonn ddee ll‟‟hhyypphhee dduuee àà llaa rrééss iiss ttaannccee mmééccaanniiqq uuee ddeess ccee lllluu lleess
éépp iiddeerrmmiiqquueess ddee llaa pp llaannttee..
**LL’’iinntteerrffaaccee ssyymmbbiioo ttiiqquuee
LLoorrss ddee llaa ccoo lloonniissaatt iioo nn ppaarr lleess cchhaammpp iiggnnoo nnss mmyyccoorrhhiizziieennss,, llaa mmeemmbbrraannee pp llaassmmiiqquuee ddeess
ccee lllluulleess ccoorrtt iiccaa lleess nn‟‟eess tt jjaammaa iiss rroo mmpp uuee ((HHaarr iissssoonn,, 11999977)),, llaa ppaarroo ii eess tt dd iissssoocc iiééee ppuuiiss eennffoonnccééee,,
cceettttee mmeemmbbrraannee ss ‟‟ iinnvvaaggiinnee ee tt pprroo lliiffèè rree aauuttoo uurr
ddeess hhyypp hheess qq uuii ssee ddéévvee llooppppeenntt àà ll‟‟ iinnttéérr iieeuurr ddee
llaa ccee lllluullee,, ll‟‟ iinnttee rr ffaaccee eess tt ttoouujjoouurrss ddéé lliimmiittééee dd uu
ccôôttéé ddee ll‟‟hhôôttee ppaarr uunnee mmeemmbbrraannee ddee nnaa ttuurree
ppllaassmmiiqq uuee ((DDeexxhhee iimmeerr,,11999977 )) .. LLeess iinntteerr ffaacceess
ffoo rrmmééeess ddaannss lleess eessppaacceess iinnttrraa oo uu
iinntteerrccee lllluullaa iirreess ((FFeerrrroo ll eett aall..,, 22000022 )) oo nntt llaa
mmêêmmee ss ttrruucc ttuurree ddee bbaassee,, ee lllleess ssoonntt ccoommppoossééss
ddeess mmeemmbbrraanneess ddee ddeeuuxx ssyymmbb iiootteess ssééppaa rrééeess
ppaarr uunn eessppaaccee iinntteerr ffaacc iiaa ll éé ttrroo iitt (( ffiigg.. 1166))..
PPaarroo ii vvééggééttaa llee
MMeemmbbrraannee pp llaassmmiiqq uuee
IInntteerr ffaaccee
PPaarroo ii ffoonnggiiqquuee
cchhaammpp iiggnnoo nn

28
FFiigg..1166 :: SScchhéé mmaa rree pprréésseennttaanntt llaa ss ttrruuccttuurree ddee ll’’iinnttee rrffaaccee ss yymmbbiioottiiqquuee ooùù ssee pprroodduuiitt llee ttrraannss ffee rrtt ddee
nnuuttrriimmee nnttss ddaannss lleess rraacciinneess àà MMAA ((HHaauussee ee tt FFeessttee rr,, 22000055))..
LLaa ppéénnééttrraa tt iioo nn vv iiaa llee ss ccee lllluu lleess éépp iiddeerrmmiiqq uueess oouu lleess ppoo iillss aabbssoorrbbaannttss oouu mmêêmmee ppaarr lleess
rrhhiizzoommeess,, ddoonnnnee nnaa iissssaannccee àà uunn mmyyccéé lliiuu mm iinnttrraa-- rraacc iinnaa iirree sseeccoo nnddaa iirree qq uuii aa llaa mmêêmmee
mmoorrpphhoo llooggiiee ee tt ss ttrruuccttuurree qq uuee ccee llllee dduu mmyyccéé lliiuumm eexxttrraa-- rraacc iinnaa iirree ((TTiimmoonneenn eett aall..,, 22000011 ;;
Ligrone et al., 2007).. CCee mmyyccéé lliiuumm iinnttrraa-- rraacc iinnaa iirree ppoo uurrssuuiitt ssaa pprrooggrreessss iioonn ppaarr ddeess
rraammiiffiiccaa tt iioonnss eett ddeess aannaass ttoommoosseess ((fusion entre deux hyphes fongiques compatibles, dans le
but de former un réseau mycélien et de permettre des échanges de matériaux génétiques) ddaannss
ll‟‟eessppaaccee iinntteerrccee lllluu llaa iirree dduu ppaarreenncchhyymmee ccoo rrtt iiccaa ll,, eett ffoorrmmee dd eess ppee lloottoonnss iinnttrraaccee lllluullaa iirreess aauu
nniivveeaauu ddeess ccoouucchheess ssuubb--éépp iiddeerrmmiiqquueess ((DDoodddd eett aall..,, 22000000 ;; WWuubbee tt eett aall..,, 22000033)) eett ss‟‟éétteenndd
jjuussqq uu''aauuxx ccee lllluulleess ccoorrtt iiccaa lleess iinntteerrnneess ((PPaawwlloowwsskkaa eett aall..,, 11999999))..
- Les pelotons
Les pelotons mycéliens sont intra-racinaires, sont beaucoup plus abondants à coté du
point de pénétration lors de l‟infection par les CMA (fig.17), leur abondance varie selon le
type morphologique de la mycorhize arbusculaire (Dickson et al., 2007; Muthukumar et
PPrraakkaasshh ,2009 ) .
Fig.17 : Aspect des pelotons mycéliens (hc) (Muthukumar et PPrraakkaasshh, 2009).

29
-- LL‟‟aa rrbbuussccuullee ::
DDeess rraammiiff iiccaatt iioonnss dd iicchhoottoommiiqquueess ddeess hhyypphheess iinnttrraammaa ttrr iicc iiee llss ppeerr ffoo rreenntt llaa ppaarroo ii
ccee lllluullaa iirree eett ppéénnèèttrreenntt àà ll‟‟ iinnttéérr iieeuurr ddee llaa ccee lllluullee,, ll‟‟hhyypphhee eenn ppéénnééttrraatt iioonn ssee rraammiiff iiee uunn ggrraanndd
nnoommbbrree ddee ffoo iiss ppoouurr rrééaa lliissee rr uunnee ss ttrruucc ttuurree rraappppee llaanntt uunn ppee tt iitt aarrbbrree ((ddeess rraammiiff iiccaatt iioonnss ddee pp lluuss
eenn pp lluuss ffiinneess)),, uunn aarrbb uussccuullee ddee 2-6 µm ((DDeexxhhee iimmeerr,, 11999977)).. II ll rreepp rréésseennttee llaa ssttrruucc ttuurree cc lléé ddee llaa
ssyymmbb iioossee eett ssaa mmoorrpp hhoo lloo ggiiee ddééppeenndd dd uu ttyyppee ddee mmyyccoorrhhiizzee aarrbb uussccuullaa iirree ((DDiicckk ssoonn eett aall..,, 22000077))..
SSaa ffoorrmmaa tt iioonn eesstt ccoo nnttrrôô llééee ppaa rr llaa mmaacchhiinneerr iiee ggéénnéé tt iiqquuee ddee llaa pp llaannttee,, ee llllee eess tt aassssoo cc iiééee àà llaa
ssyynntthhèèssee ddee ll‟‟aacc iiddee jjaassmmoonniiqq uuee et àà ll‟‟aaccccuummuullaatt iioonn dduu HH22OO22 ((HHaauussssee eett aall..,, 22000077 ;; Herrera-
Medina et al., 2008)..Le nombre d‟arbuscules, différenciés en 4 jours à un mois après la
colonisation mycorhizienne (Simoneau et al., 1994), dépend de la plante, de la disponibilité
des nutriments et du partenaire fongique (Smith et Read ,1997; Harrison, 1999).
LL‟‟aa rrbbuussccuullee aauuggmmeennttee llaa ssuurr ffaaccee ddee ccoo nnttaacctt eennttrree lleess ssyymmbb iioo tteess,, cceett ttee iinnttee rr ffaaccee
aarrbbuussccuullaa iirree rreepp rréésseennttee llee ss iittee dd‟‟éécchhaannggee ddeess nnuuttrr iimmeennttss ((GGuueetthheerr eett aall..,, 22000099 )).. SSee lloo nn SS mmiitthh
eett RReeaadd ((11999977 )),, lleess ppee lloo ttoonnss ddaannss llaa mmyyccoorrhhiizzee ddee ttyyppee «« PPaa rr iiss »» eett llee mmyyccéé lliiuumm iinnttrraaccee lllluullaa iirree
ccoonnss tt iittuueenntt dd‟‟aauuttrreess ss iitteess dd‟‟éécchhaannggeess eett ddoonncc uunnee ssuurrffaaccee iimmppoorrttaannttee,, iillss oonntt uunnee dduurrééee ddee vviiee
pplluuss lloonngguuee,, eett ppaa rr ccoonnssééqquueenntt iillss ppeeuuvveenntt iinntteerrvveenniirr ddaannss llee ttrraannssffeerrtt ddee nnuuttrr iimmeennttss ((DDiicckkssoonn
eett KKoo llss iikk,, 11999999 ))..
LLeess aarrbbuussccuulleess nnééoo ffoorrmmééss dduurraanntt llaa ccoo lloonniissaatt iioonn,, ssoonntt ddeess ss ttrruucc ttuurreess éépphhéémmèèrreess eett oonntt
uunnee dduurrééee ddee vviiee ttrrèèss ccoo uurr ttee,, ddee 22 àà 1155 jjoouurrss ((HHaarr lleeyy,, 11998899)) oouu 77 àà 1111 jjoouurrss ((FFeerrrroo ll eett aall..,,
22000022)),, ppaarr ffoo iiss dd‟‟uunnee àà ttrroo iiss sseemmaa iinneess (Smith et Read 1997; Harrison, 1999). SSee lloonn ddee SSoouuzzaa eett
aall.. ((22000055)), la persistance de la totalité des arbuscules dans les racines mycorhizées des
plantes, en culture en pot, est plus longue pour les espèces de la famille des Gigasporacées
que celles des Gloméracées. LLeess aarrbb uussccuulleess qquuii ddééggéénnèèrreenntt ffoo rrmmeenntt ddeess aammaass àà ll‟‟ iinnttéérr iieeuurr
ddeess ccee lllluulleess,, mmaa iiss iillss ppeeuuvveenntt êêtt rree ssuubbsstt iittuuééss ppaarr dd‟‟aauuttrreess ((TTootthh ee tt MMiilllleerr,, 11998844 ;; AAlleexxaanndd eerr eett
aall..,, 11998888 eett 11998899))..
AApprrèèss llaa ffoo rrmmaa tt iioonn ddeess aarrbbuussccuulleess,, cceerrttaa iinneess eessppèècceess ddee GGlloommuuss eett dd‟‟AAccaauu lloossppoorraa,, ffoorrmmeenntt
ddeess vvééss iiccuulleess ((DDaa llppéé eett aall..,, 22000055bb))..
-- Les vésicules
Les vésicules sont des organes de réserve, constituées de lipides et de glycolipides
(Dodd et al., 2000) et des organites qui leur permettent d‟avo ir une croissance autonome

30
(Timonen et al., 2001). D‟ailleurs, elles sont des propagules infectives qui peuvent générer
un pro-mycélium (Declerck et al., 1998) et se former à partir du mycélium intra-racinaire
(fig., 18) (à l‟intérieur ou à l‟extérieur des couches du parenchyme cortical) (Paw lowska et
al.,1999).
Dans la mycorhize arbusculaire, les vésicules sont intra ou intercellulaires, terminales
ou intercalaires (Dalpé et al., 2005b).
Généralement, elles ont une forme ovoïde, globulaire ou ellipsoïde et contiennent une à
deux parois minces (1 à 2.5µm de diamètre). En effet, elles sont ellipsoïdes chez Glomus
spp. (Webet et al., 2003), tandis qu‟elles ont des formes variables chez Acaulospora spp.et
Kuklospora spp. (Blaszcowski et al., 2006), et ne se forment jamais chez Gigaspora spp. et
Scutellospora spp.
Après l‟établissement de la symbiose, le développement du mycélium extramatriciel
s‟effectue en quelques heures, et se différencie à nouveau en « BAS » et en « VLS »
(Declerck et al., 1998).
Fig.18 : Aspect des vésicules dans une coupe de racine mycorhizée (Dexheimer, 1997).
**LLee mmyyccéé lliiuumm eexxttrraa--mmaattrriicciiee ll
La structure de base de ce mycélium sont les hyphes « runner », hyphes coenocétiques
droits, larges, à paroi épaisse ; ils sont composés d‟une seule paroi chez Gl. versiforme (Carr
et al., 1985) et d‟une double paroi chez Gl. fasciculatum (Bonfante-Fasolo et Grippiolo,
1982).
V

31
Leur appellation est due à leur rapidité d‟extension et de colonisation des substrats (sol
ou gélose en culture monoxénique) et de leur association avec la racine ; leur vitesse
d‟élongation atteint 750µm h-1 ( Dalpé et al., 2005b). CCeess hhyypphheess ffoorrmmeenntt uunn rréésseeaauu ddaannss llee
ssoo ll,, iillss aaggiisssseenntt ccoommmmee uunnee eexxtteennss iioo nn ddeess rraacc iinneess ddeess pp llaannttee ss hhôô tteess,, eett aauuggmmeenntteenntt llee vvoo lluummee
dduu ssoo ll aacccceessss iibb llee àà llaa pp llaannttee,, aabboouutt iissssaanntt aa iinnss ii àà uunnee mmee iilllleeuurree mmoobb iilliissaatt iioo nn ddee ll‟‟eeaauu eett ddeess
nnuuttrr iimmeennttss dd uu ssoo ll ((NN,, PP,, KK,, MMgg)).. LLee rréésseeaauu dd‟‟hhyypphheess jjoo uuee uunn rrôô llee ffoonnddaammeennttaa ll ddaannss llaa
ffoo rrmmaa tt iioo nn,, llaa ssttaabb iill iissaatt iioo nn ddeess aaggrrééggaattss dd uu ssoo ll ppaarr llaa pp rroodd uucctt iioo nn ddee llaa gglloommaa lliinnee ((DDoodddd,, 22000000 ;;
CChhoott ttee,, 22000011 ;; MMuurruullaannddaa eett aall..,, 22000033 ;; PPoorrccee ll eett aall..,, 22000033 )) eett ll‟‟ iinniitt iiaatt iioonn dd‟‟uunnee nnoouuvvee llllee
iinnffeecctt iioo nn..
LLee mmyyccéé lliiuumm ddeess CCMMAA aauuggmmeennttee llaa ssuurrffaaccee ddee ccoonnttaacctt aavveecc lleess aauuttrreess mmiiccrroooo rrggaanniissmmeess
dduu ssoo ll ee tt llaa vvoo iiee ddee ttrraannss llaatt iioonn ddee ll‟‟éénneerrggiiee ddeess pprroodduuiittss aassss iimmiillééss ppaarr llaa pp llaannttee vveerrss llee ssoo ll
((JJoohhnnssoo nn eett aall..,, 22000044 )) ..
- Les cellules auxiliaires :
Chez les genres Gigaspora, Pacispora et Scutellospora (Blaszkowski et al., 2006), le
mycélium extramatriciel forme des cellules auxiliaires (AC) (formes globulaires regroupées
en grappe, attachées aux hyphes extramatriciels et souvent ornementées) ( fig. 19). En
culture monoxénique, les jeunes cellules contiennent des gouttelettes lipidiques tandis que les
vieilles apparaissent vides (de Souza et Declerck, 2004).
Les cellules auxiliaires sont formées lors des ramifications d‟hyphes extramatriciels ; chaque
ramification génère plusieurs branches qui forment à leur tour des structures sphériques
regroupées en masses dont chacune contient 2 à 20 sphères mesurant 12 à 39µm de diamètre.
Les cellules auxilliaires sont riches en noyaux, en organites et en lipides (Bonfante et Perotto,
1995) ; leur surface est lisse chez Scutellospora sp. et rugueuse chez Gigaspora sp.
Declerck et al. (2004) ont observé en culture monoxénique, une croissance des hyphes à
partir des cellules auxiliaires chez S. reticulata, ils ont suggéré que les longs fragments de
mycélium intacts portant plusieurs cellules auxiliaires peuvent induire une symbiose.
LLee rréésseeaauu dd‟‟hhyypphheess eexxttrraammaattrr iicc iiee llss ppeeuutt êêtt rree àà ll‟‟oo rr iiggiinnee ddeess ssppoorreess,, cceess ddeerrnniièèrreess àà
mmaattuurraa tt iioonn,, ssoonntt lliibbéérrééeess ddaannss llee ssoo ll eett lleeuurr ggeerrmmiinnaa tt iioonn ddoonnnnee rraa llee pp rroo--mmyyccéé lliiuumm ddee ddééppaa rrtt..

32
Fig. 19 : Aspect des cellules auxiliaires (CA) » (Bago et Cano, 2005).
A : morphologie des cellules auxiliaires attachées aux hyphes extra-racinaires (HyER) ;
B : Développement des cellules auxiliaires de Gi. margarita sur le tronc des « BAS ».
- Les spores
Elles sont unicellulaires, multinuclées, leur diamètre est compris entre 50 et
500 μm ; il est de 314 µm chez les Gigasporacées, 158 µm chez les Acaulosporacées et
127μm chez les Gloméracées.
Les spores sont produites à l‟extrémité d‟un hyphe sporogène ou suspenseur (structure
reliant la spore aux hyphes du mycélium dont la morphologie est utilisée pour identifier
certains genres de CMA) ou à l‟intérieur des racines, dans le sol ou encore elles adhèrent
aux racines du végétal.
La sporulation intra-racinaire a été observée au champ et en pot chez Gigaspora et
Scutellospora (Schenck et Perez, 1990; de Souza et al., 2005), Gl. aggregatum et
Gl.intraradices (Schenck et Smith, 1982). Gadkar et Adholeya (2000) ont rapporté qu‟une
culture monoxénique d‟un isolat de Gigaspora margarita produit 10–15% de spores intra-
racinaires. Pour certains auteurs (de Souza et al., 2005) , les spores intra-racinaires sont les
propagules infectives dans les racines mycorhizées des Gigasporacées.
Cependant, le nombre de spores produites dépend de l‟espèce fongique, de l‟espèce
végétale, de la fertilité du sol, de l‟application des fertilisants et de l‟intensité de la lumière
(Blaszcowski, 1993).
La reproduction des CMA est clonale, les spores ont le rôle de séquestrer l‟information
génétique de l‟espèce fongique et de la disperser dans un nouvel habitat.
Ces spores se différencient à nouveau en VLS en 2 à 8 jours (fig.20). LLoorrss ddee llaa
sséénneesscceennccee ddee llaa rraacc iinnee,, oouu ddeess tt iissssuueess ccoorr tt iiccaauuxx,, lleess vvééss iiccuulleess ssoo nntt lliibbéé rrééeess ddaannss llee ssoo ll,, ee lllleess
pprroodduuiisseenntt ddeess hhyypp hheess aapp rrèèss ggeerrmmiinnaa tt iioo nn qq uuii ssee ddéévvee llooppppeenntt eenn hhyypphheess sseeccoonnddaa iirreess pp lluuss
ffiinnss,, ee tt rréé iinnffeecctteenntt lleess rraacc iinneess..
CA
HyER
A B
BAS
MyER CA

33
FFiigg.. 2200 :: AAss ppeecctt dd’’uunnee ss ppoorree mmaattuurree pprroodduuiittee ppaarr llee mmyyccéélliiuumm eexxttrraammaattrriicciiee ll eett ddeess VVLLSS ((DDaallppéé
eett aall.. ,, 22000055bb))..
11..66.. NNuuttrriittiioo nn ddeess CCMMAA
11..66..11.. TTrraannssffeerrtt dduu ccaarrbboonnee vveerrss llee cchhaammppiiggnnoo nn
LLeess CCMMAA ssoo nntt ddeess ssyymmbb iiootteess oobb lliiggaa ttoo iirreess,, iillss ddééppeennddeenntt ppoo uurr lleeuurr ccrroo iissssaannccee eett lleeuurr
aacctt iivviittéé ddee ll‟‟aapppprroovviiss iioonnnneemmeenntt eenn ccoommppoossééss ccaarrbboo nnééss ssoouuss ffoo rrmmee ddee CCOO22 ((BBééccaarrdd eett aall..,,
11999922)) ,, eett ddee mméé ttaabboo lliitteess ddee ttyyppee CC33 eett CC44 ((LLuuggoo eett CCaabbee lllloo,, 22000022)) àà ppaarr tt iirr dd uu ppaa rrtteennaa iirree
pphhoottoossyynntthhéé tt iiqquuee ((DDoodddd,, 22000000 ;; DDaa llppéé,, 22000011)).. LLeess pp hhoottoo ssyynntthhéé ttaattss ssoonntt dd iirr iiggééss vvee rrss llaa rraacc iinnee
eett llee ttaauuxx ddee pphhoottoossyynntthhèèssee ddee ll‟‟hhôôttee aauuggmmeennttee ((WWrriigghhtt eett Upadhyaya,, 11999988)).. PP lluuss ddee 2200%%
ddeess pprroodduuiittss ddee pphhoo ttoossyynntthhèèssee aassss iimmiillééss ppaarr llaa pp llaannttee ssoonntt ttrraannssfféérrééss aauu cchhaammpp iiggnnoonn
((MMeerrrryywweeaa tthheerr ,, 22000011))..
LLee tt rraannss ffee rrtt ddeess ccoo mmppoossééss ccaarrbboonnééss iimmpp lliiqq uuee ddeess ee fffflluuxx ppaassss iiffss àà ttrraavvee rrss llaa mmeemmbbrraannee
ppllaassmmiiqq uuee ddee llaa pp llaannttee,, ssuuiivviiss ppaa rr uunn ppaassssaaggee aacc tt iiff vveerrss llaa mmeemmbbrraannee dduu cchhaammpp iiggnnoonn ppaarr
ll‟‟aappoopp llaa ssttee iinnttee rr ffaacc iiaa ll ((FFeerrrroo ll eett aall..,, 22000022)) (( ffiigg..2211))..
11..66..22.. TTrraannssffeerrtt dduu pphhooss pphhaa ttee vveerrss llaa ppllaannttee ::
LLaa ccoo lloonniissaatt iioo nn ddeess rraacc iinneess ppaarr llee cchhaammpp iiggnnoonn eesstt aaccccoo mmppaaggnnééee ppaarr llee ddéévvee llooppppeemmeenntt
dduu rréésseeaauu eexxttrraa-- mmaattrr iicc iiee ll,, ppaarr ccoo nnssééqq uueenntt ll‟‟aauuggmmeennttaa tt iioonn dduu ttaauuxx dd‟‟aabbssoorrpp tt iioonn ddeess mmiinnéérraauuxx
S VLS

34
lleess mmoo iinnss mmoobb iilleess ddaannss llee ssoo ll,, ssuurr ttoouutt llee PP ((CCaarrddoossoo eett aall..,, 22000066)).. DDuurraanntt llaa ssyymmbb iioossee,, llee
mmoouuvveemmeenntt dduu PP ccoommmmeennccee ppaarr ssoonn aabbssoo rrpptt iioonn àà ppaarr tt iirr ddee llaa ssoo lluutt iioonn dduu ssoo ll ppaarr ddeess
ttrraannssppoorr tteeuurrss ss iittuuééss ddaannss llee mmyyccéé lliiuumm eexxttrraa-- rraacc iinnaa iirree ((GGuueetthhee rr eett aall..,, 22000099 ;;SS mmiitthh eett aall..,,
22001100)),, iill eesstt ssuuiivvii ppaarr ssaa ttrraannss llooccaa tt iioonn vvee rrss lleess ss ttrruuccttuurreess iinnttrraa-- rraacc iinnaa iirreess dd uu cchhaammpp iiggnnoo nn eett
ffiinnaa lleemmeenntt vveerrss lleess rraacc iinneess ccoo lloo nniissééeess vv iiaa lleess iinnttee rr ffaacceess aarrbbuussccuullaa iirreess ((FFee rrrroo ll eett aall..,, 22000022 ))..
LLee ttrraannssffeerr tt dduu PP àà ttrraavveerrss ll‟‟ iinntteerr ffaaccee ssyymmbb iioott iiqq uuee ddéébbuuttee ppaarr ddeess ee fffflluuxx àà ppaarr tt iirr dduu
cchhaammpp iiggnnoonn vveerrss ll‟‟aappoopp llaassttee iinntteerr ffaacc iiaa ll.. LLeess pp rroocceessssuuss ddee ttrraannssppoo rrtt mmeemmbbrraannaa iirree ffaavvoorr iisseenntt llaa
ppeerrttee dduu PP ddee ll‟‟aarrbb uussccuullee ((CC llaarrkkssoonn,, 11998855 ;; GGuuee tthheerr eett aall..,, 22000099)),, ssoonn ttrraannssppoorrtt àà ttrraavvee rrss llaa
mmeemmbbrraannee aarrbbuussccuullaa iirree iinndduuiitt ddeess ttrraannssppoorrtteeuurrss oo uu ddeess cchhaa iinneess dd‟‟ee fffflluuxx ssppéécc iiffiiqquueess ((FFeerrrroo ll eett
aall..,, 22000022))..
FFiigg..2211 :: SScchhéé mmaa rreepprréésseennttaanntt lleess éécchhaannggeess ddee nnuuttrriimmeennttss ddaannss lleess rraacciinneess àà MMAA àà
ttrraavvee rrss ll’’iinnttee rrffaaccee ssyymmbbiioott iiqquuee ((HHaauussssee ee tt FFeesstteerr,, 22000055))..
Cellule du champignon
Apoplaste
Cellule de la
plante

35
2. Données sur les mycorhizes
2.1. Définition :
Le terme « mycorhize » désigne une symbiose mutualistique bénéfique entre les
champignons et les racines de la majorité des plantes vasculaires qui leur permet de survivre,
croître plus efficacement (Smith et Read, 1997).
Les plantes autotrophiques capables de transformer l‟énergie en éléments nutritifs et les
champignons hétérotrophiques spécialisés dans l‟absorption de l‟eau et des éléments
minéraux à partir du substrat de sol associent leurs fonctions et leurs rôles compléme ntaires
(Dalpé, 2001).
Les symbioses mycorhiziennes se rencontrent chez 90% des plantes terrestres (Dalpé,
2001;Smith et Read, 2008) et chez les plantes aquatiques et marécageuses (Beck- Nielsen et
Vindaek, 2001 ; Radhika et Rodriguez, 2006), distribuées dans tous les climats et les
écosystèmes , indépendamment du type de sol (Khade et Adoleya, 2009 ; Alguacil et al.,
2009 ;Smith et al., 2010 ), de la végétation (Wubet et al., 2003 ; Brundrett, 2009) et des
conditions environnementales (Tao et Zhiwei, 2005 ; Cho et al., 2006) ; elles jouent un rôle
actif dans l‟amélioration de la diversité et la productivité des plantes, dans la diversité de la
microflore , de la microfaune et de la qualité du sol.
D‟ailleurs, la colonisation des terres émergées a été rendue possible à l‟échelle des
temps géologiques par ce phénomène (Simon et al., 1993 ; Redecker et al., 2000), cela
suggère que les plantes vasculaires et les champignons ont co-évolué (Schwarzott et al.,
2001). En effet, les champignons ont été détectés dans les racines de plantes fossiles
d‟Aglaophyton (plante pré-vasculaire, ancêtre des plantes vasculaires) datant de la période
dévonienne (période géologique entre -420 et -367 millions d‟années) (Taylor et al., 1995).
Cette période géologique est caractérisée par la colonisation du milieu terrestre par des
organismes aquatiques et le développement des plantes en milieu terrestre. Il a été observé
que de nombreuses espèces de mousses pouvaient être colonisées dans leur partie
chlorophyllienne par des champignons MA des genres Glomus, Paraglomus, Acaulospora et
Gigaspora (Zhang et Guo, 2007).
Quelques familles de plantes vasculaires sont qualifiées de non mycotrophes, elles
n‟établissent pas de symbiose mycorhizienne : les Crucifères, les Chénopodiacées, les
Amaranthacées les Cypéracées, les Urticacées, les Juncacées, et quelques espèces de

36
Caryophyllacées comme Ceratium tatrae et Dianthus plumarius L. Subsp. praecox (Barea et
al., 2005 ; Zubek et al., 2008 ; Vosatka et Albrechtova, 2009).
Il est connu que les plantes dont le système racinaire est très ramifié (ex. les Graminées)
sont moins mycotrophiques, elles ne dépendent pas du champignon dans l‟exploration du sol
et l‟absorption du P pour une croissance optimale. Tandis que d‟autres plantes mycotrophes
(poireau, tomate, oignon, carotte…) ont un système racinaire grossier qui détermine leur
dépendance de la symbiose (Dodd, 2000 ; Merryweather, 2001 ; Janos, 2007 ; Fortin et al.,
2008).
Il existe plusieurs formes d‟associations mycorhiziennes (fig.22) (Fortin et al., 2008).
Les champignons qui forment les divers types de mycorhizes appartiennent à des groupes
fongiques différents. Les morphotypes adoptés par les symbiotes fongiques lorsqu‟ils
colonisent les systèmes racinaires des végétaux, sont utilisés pour classer et distinguer les
associations mycorhiziennes (tableau 2) (Brundrett, 2004 ; Smith et Read, 2008).
D‟ailleurs, au sein d‟un même écosystème et parfois autour d‟un seul et même arbre, la
succession et la concurrence peuvent intervenir entre les différents types de mycorhizes, cela
a été mis en évidence chez les aulnes (Beddiar, 1984), les hélianthèmes (Read et al., 1977 ;
Ian et al., 2004), les Eucalyptus (Pagano et al., 2009), les peupliers et les saules (Khasa et al.,
2002).
Le type biologique d‟une espèce fongique mycorhiziene donnée n‟est pas figé. Le
champignon peut commencer son existence en temps que saprophyte (Strullu et al., 1997 ;
Davey, 2006) avant de rencontrer son partenaire spécifique avec lequel il établira une relation
mycorhizienne, cette dernière pouvant laisser place, dans certaines conditions, à un mode de
vie parasitaire (Merryweather, 2001).

37
Fig. 22 : Les principaux types de mycorhize représentés sur une coupe transversale de
racine (Le Tacon, 1985) .
sporocyste rempli
de spores
Endomycorhizes à pelotons
des Orchidées
Endomycorhizes à pelotons
des Ericacées et hélianthèmes
Endomycorhizes
à arbuscules
à arbuscules
Cèpe Truffe
Ectomycorhizes
Ectendomycorhizes

38
Tableau 2. Les caractéristiques des différents types de mycorhizes d‟après Smith et Read
(1997) in Siddiqui et Pichtel (2008). Type
mycorhizien
Mycorhize
Arbusculair
e
Ectomycorhiz
e
Ectendo-
mycorhize
arbutoïd
e
Monotro-
poïde
Ericoïd
e
Orchi-
doïde
Taxonomie
du
champignon
Glomeromycot
a
Basidiomycètes
Ascomycètes
Zycomycètes
Basidio-
mycètes
Ascomycète
s
Basidio-
mycètes
Basidio-
mycètes
Asco-
mycètes
Basidio-
mycètes
Taxonomie
de la Plante
Bryophytes
Ptéridophyte
s
Gymnosperm
es
Angiosperme
s
Gymnosperme
s
Angiospermes
Gymno-
spermes
Angiosperm
es
Éricales
Monotro
-poïdées
Éricales
Orchi-
dacées
Colonisation
intra-
cellulaire
+
-
+ +
+ + +
Manteau
fongique
- + +/- +/- +
- -
Réseau de
Hartig
-
+
+ +
+
-
-
Vésicule + /- -
- - - - -
Achlorophyllie - -
- - + - +

39
2.2. Les différents types de mycorhizes :
2.2.1. Les ectomycorhizes :
Fig. 23: Ectomycorhize de Hebeloma mesophaeum sur racine de l’épicéa (Egli et Brunner, 2002).
a : Aspect du champignon macroscopique. b : Manteau fongique épais entourant les radicelles de la
plante. Rad : Radicelle ; M : Manteau fongique.
Il existe environ 6000 espèces de plantes à ectomycorhizes (Rinaldi et al., 2008;
Brundrett, 2009) ; ces plantes appartiennent aux Angiospermes (environ 5600 espèces),
Gymnospermes (285 espèces), aux Ericacées , Pinacées, Bétulacées et Fagacées (Smith et
Read, 2008;Van der Heijden et al., 2008 ; Matsuda et al., 2009 ; Brundrett, 2009).
Les plantes ectomycorhizées sont généralement des espèces forestières, principalement des
arbres et arbustes, avec lesquelles le partenaire fongique joue un rôle important dans la
croissance des arbres dans divers habitats à travers le globe terrestre (Le Tacon, 1985;
Dexheimer ,1997 ; Kernaghan, 2005; Fortin et al., 2008 ; Brundrett, 2009).
Les champignons et les plantes mycorhizées ont une distribution taxonomique large
dans tous les continents (sauf l‟Antarctique) ce qui suggère une évolution ancienne de cette
symbiose. Les Pinacées sont certainement la famille de plantes la plus ancienne ayant établi
une symbiose avec les champignons ectomycorhiziens (Hibbett et Matheny, 2009).
Les champignons qui forment des ectomycorhizes (fig. 23 a) appartiennent aux
Ascomycota (27 lignées), aux Basidiomycota (37 lignées) et aux Zygomycota (deux lignées)
(Bruns et Shefferson, 2004 ; Hibbett et al., 2000 et 2007; Matheny et al., 2007; Tedersoo et
al., 2010).
Ces mycosymbiotes peuvent appartenir aux ordres des Pézizales (y compris les genres
Tuber , Terfezia et Tirmania (Fortas et Chevalier, 1992a)) , Agaricales, Bolétales, Hélotiales,
Endogonales, Sordariales, Athéliales, Russulales, Théléphorales, Hyménochaetales,
Gomphales, Hystérangiales, Cantharellales, Sebacinales, Eurotiales, (Tedersoo et al., 2010).
a b
Rad M

40
Chez les ectomycorhizes, la racine mycorhizée est transformée, elle présente la
morphologie d‟une racine courte, renflée, ramifiée, ou même peu prendre la forme d‟un
racème (grappe) (Dexheimer, 1997 ; Fortin et al., 2008). Le mycélium se développe à
l‟extérieur des cellules corticales, les hyphes forment un manchon ou manteau fongique
autour des radicelles (fig.23 b) puis ils s‟insinuent entre les cellules pour former le réseau de
Hartig. Suivant les partenaires, le réseau s‟étend jusqu‟au cylindre central ou reste aux
premières assises cellulaires (Dexheimer, 1997). Dans la plupart des ectomycorhizes, les
cellules corticales de l‟hôte dégénèrent rapidement ou se transforment en cellules à tannins
alors que le réseau contient encore des hyphes vivants ( Dexheimer, 1997).
Contrairement aux champignons qui forment des mycorhizes arbusculaires et éricoïdes,
la plupart des champignons ectomycorhiziens se reproduisent par voie sexuée lorsque les
conditions sont favorables, les hyphes dicaryotiques donnent naissance à des corps fructifères
(carpophores épigés ou hypogés) (Kernaghan, 2005).
22..22..22.. LLeess eecctteennddoommyyccoo rrhhiizzeess
LLeess eecctteennddoommyyccoorrhhiizzeess pprréésseenntteenntt àà llaa ffoo iiss ddeess ss ttrruucc ttuurreess ccaarraacc ttéérr iisstt iiqquueess ddee llaa
ssyymmbb iioossee ectomycorhizienne eett eennddoo mmyyccoorr iizziieennnnee.. EElllleess ssoonntt ccoonnnnuueess cchheezz lleess EErr iiccaaccééeess,, lleess
PPiinnaaccééeess eett MMoonnoottrrooppaaccééeess (Brudrett et al., 1996 ; Barea et al., 2005) eett lleeuurrss ccaarraacc ttéérr iisstt iiqquueess
uullttrraa ssttrruucc ttuurraa lleess dd iiffffèè rreenntt ssee lloonn qq uuee ll‟‟hhôôttee vvééggééttaa ll ssoo iitt aauuttoo ttrroopphhee oouu hhéé ttéérroo ttrroopphhee..
LLaa mmyyccoo rrhhiizzee mmoo nnoottrrooppoo ïïddee ssee ffoorrmmee eennttrree uunn cchhaammpp iiggnnoonn eecc ttoommyyccoo rrhhiizziieenn eett uunn
mmoonnoo ttrrooppee ((eexx.. MMoonnoottrrooppaa hhyyppooppiittyyss)) qquuii eess tt uunnee pp llaannttee nnoo nn cchhlloo rroopphhyylllliieennnnee ppaa rraass iittee
dd‟‟aarrbbrreess aavveecc lleessqquuee llss ee llllee pp aarrttaaggee uunn mmêêmmee cchhaammpp iiggnnoonn mmyyccoo rrhhiizziieenn ((FFoo rrtt iinn eett aall..,, 22000088 )),, ccee
ddeerrnniieerr ffoorrmmee uunn mmaanntteeaauu ,, uunn rréésseeaauu ddee HHaarr tt iigg ,, àà pp aarrtt iirr dduuqquuee ll uunnee rraammiiffiiccaa tt iioo nn ppee rr ffoorree llaa
ppaarroo ii ccee lllluullaa iirree eett ffoo rrmmee llaa«« cchheevviillllee »» (( ffoorrmmaatt iioonn eennddoommyyccoorrhhiizziieennnnee)) ((DDeexxhhee iimmeerr,, 11999977)) ..
LLaa mmyyccoorrhhiizzee aarrbb uuttoo ïïddee ccoonnnnuuee cchheezz AArrbbuuttuuss uunneeddoo eett lleess EErr iiccaaccééeess,, llee cchhaammpp iiggnnoonn
ffoo rrmmee llee mmaanntteeaauu ffoo nnggiiqq uuee eett llee rréésseeaauu HHaarrtt iigg,, àà ppaarr tt iirr ddee ccee ddeerrnniieerr,, lleess hhyypphheess ppéénnèèttrreenntt àà
ll‟‟ iinnttéérr iieeuurr ddeess ccee lllluulleess ccoo rrtt iiccaa lleess eett ffoo rrmmeenntt ddeess ppee lloottoonnss mmyyccéé lliieennss ((DDeexxhhee iimmeerr,, 11999977))..
2.2.3. Les endomycorhizes

41
2.2.3.1. Les endomycorhizes à pelotons
2.2.3.1.1. Les endomycorhizes éricoïdes
Cette symbiose est présente chez toutes les espèces d‟Ericacées appartenant aux genres
Calluna sp , Erica sp., Gaulteria sp., Vaccinum myrtillus, Rhododendrous sp., Centauria
cyanus (Dexheimer , 1997) mieux connus sous le nom de bruyère de callune , bruyère,
gaulthérie ou palommier, myrtilles , Azalées (rhodo) et bleuet (Fortin et al., 2008). Elle est
prédominante dans les sols acides riches en matières organiques récalcitrantes mais pauvres
en phosphores et en azote tels que les landes à callunes ou les milieux sujets à l‟entourbement
(Cairney et Burke, 1998).
Le partenaire fongique (Martin et al., 2007) est un Ascomycète ( Smith et Read, 1997)
tels que : Hymenoscyphus ericae (ordre des Hélotiales), Scytalidium vaccinii Dalpé (ordre
des Hélotiales) (Egger et Sigler, 1993) et Oidiodendron spp. (ordre des Onygénales)
(Monreal et al., 1999 ; Sharples et al., 2000 ; Johansson, 2001).
Le mycélium extra-racinaire forme un réseau d‟hyphes vivants dans le sol et le
mycélium intra-racinaire colonise l‟épiderme et forme des pelotons mycéliens intracellulaires,
cette association reste active pendant une courte période de temps, après quoi, les cellules
corticales dégénèrent et le champignon colonise en saprophyte les cellules mortes de la plante.
2.2.3.1.2. Les endomycorhizes Orchidoïdes
CCeettttee mmyyccoorrhhiizzee eesstt ssppéécciiffiiqquuee ddeess OOrrcchhiiddééeess,, aappppaarrtteennaanntt aauuxx ggeennrreess OOrrcchhiiss,,
DDaaccttyyllaarrcchhiiss,, VVaanniillllee ……eettcc.. IIllss ss‟‟aassssoocciieenntt aavveecc ddeess BBaassiiddiioommyyccèètteess aappppaarrtteennaanntt aauu ggeennrree
RRhhiizzooccttoonniiaa.. LLee cchhaammppiiggnnoonn ppéénnèèttrree ddaannss lleess cceelllluulleess rraacciinnaaiirreess,, mmaaiiss nnee ffoorrmmee qquuee ttrrèèss
rraarreemmeenntt uunn rréésseeaauu iinntteerrcceelllluullaaiirree,, lleess hhyypphheess ss‟‟eennrroouulleenntt ssuurr eeuuxx mmêêmmeess eett ffoorrmmeenntt ddeess
ppeelloottoonnss iinnttrraacceelllluullaaiirreess,, ll‟‟iinnffeeccttiioonn ssee pprrooppaaggee ddiirreecctteemmeenntt dd‟‟uunnee cceelllluullee àà uunnee aauuttrree,, ppuuiiss lleess
ppeelloottoonnss dd‟‟hhyypphheess ddééggéénnèèrreenntt eett ssoonntt ddiiggéérrééss ppaarr lleess cceelllluulleess pplluuss iinntteerrnneess ddiitteess
pphhaaggooccyyttaanntteess,, dd‟‟aaiilllleeuurrss àà ll‟‟ââggee aadduullttee,, cceerrttaaiinnss OOrrcchhiiddééeess éélliimmiinneenntt ccoommppllèètteemmeenntt llee
cchhaammppiiggnnoonn aalloorrss qquuee dd‟‟aauuttrreess,, llee ggaarrddeerroonntt ttoouuttee lleeuurr vviiee ((FFoorrttiinn eett aall..,, 22000088))..
22..22..33..22.. LLaa mmyyccoorrhhiizzee aarrbbuussccuullaaiirree

42
LL‟‟aassssoocciiaattiioonn mmyyccoorrhhiizziieennnnee ddee ttyyppee aarrbbuussccuullaaiirree eesstt uunnee aassssoocciiaattiioonn ssyymmbbiioottiiqquuee
mmuuttuuaalliissttee oobblliiggaattooiirree ((FFeerrrrooll eett aall..,,22000022 ;; VVeessttbbeerrgg eett ccaasssseellllss..,, 22000099 )) qquuii ss‟‟eeffffeeccttuuee eennttrree
ddeess cchhaammppiiggnnoonnss aayyaanntt lleeuurr pprroopprree pphhyylluumm mmoonnoopphhyyllééttiiqquuee ((RReeddeecckkeerr eett aall..,, 22000000 ;;
SScchhwwaarrzzootttt eett aall..,, 22000011)),, GGlloommeerroommyyccoottaa ((SScchhuubblleerr eett aall..,, 22000011 )) eett 8800%% ddee ttoouutteess lleess
eessppèècceess ddee ppllaanntteess ((SSmmiitthh eett RReeaadd,, 22000088 ;; KKhhaaddee eett AAddhhoolleeyyaa,, 22000099 )) ;; eellllee eesstt llaa ssyymmbbiioossee llaa
pplluuss rrééppaanndduuee ddaannss ll‟‟ééccoossyyssttèèmmee tteerrrreessttrree (( SSmmiitthh eett RReeaadd,, 11999977 ;; HHaarriissssoonn,, 11999977 ,, SSttuurrmmeerr,,
11999988 ,, WWuubbeett eett aall..,, 22000033 ;; GGuueetthheerr eett aall..,, 22000099)).. EEnn eeffffeett,, eellllee ccoonncceerrnnee 220000 000000 eessppèècceess ddee
ppllaanntteess ((BBoonnffaannttee eett aall..,, 22000099)) lleess AAnnggiioossppeerrmmeess,, lleess GGyymmnnoossppeerrmmeess,, lleess PPttéérriiddoopphhyytteess,, lleess
RRuuttaaccééeess eett lleess BBrryyoopphhyytteess (( WWuubbeett eett aall..,, 22000033 ;; DDiicckkssoonn eett aall..,, 22000077 ;; AAppppeellhhaannss eett aall..,,
22000088 ;; MMuuttkkuummuurr eett Prakash,, 22000099 ))..
LLeess cchhaammpp iiggnnoonnss mmyyccoorrhhiizziieennss aarrbbuussccuullaa iirreess ((CCMMAA)),, tt iieennnneenntt lleeuurr nnoomm ddee lleeuurr
ssttrruucc ttuurree ccaarraaccttéé rr iiss tt iiqquuee,, ll‟‟aarrbb uussccuullee oo ffffrree uunnee ssuurrffaaccee ddee ccoonnttaacctt iimmppoorrttaannttee,, iill ee sstt
iinnttrraaccee lllluullaa iirree ee tt ccoonnsstt iittuuee llee ss iittee dd‟‟éécchhaannggee ddee nnuuttrr iimmeennttss eennttrree lleess ddeeuuxx ppaarr tteennaa iirreess ((SS mmiitthh eett
RReeaadd,, 11999977 ;; CC hhiifffflloo tt,, 22000088 ;; GGuuee tthheerr eett aall..,, 22000099 )) .. LL‟‟aarrbb uussccuullee éé ttaanntt llaa ssttrruucc ttuurree llaa pp lluuss
iimmppoorr ttaannttee ddee llaa mmyyccoo rrhhiizzee,, dd‟‟aauuttrreess ss ttrruucc ttuurreess mmoorrpp hhoo lloo ggiiqq uueess ccaarraacc ttéérr iisseenntt ccee ttttee
mmyyccoorrhhiizzeess ttee llss qquuee :: llee mmyyccéé lliiuu mm eexxttrraa mmaa ttrr iicc iiee ll,, lleess aapppprreessssoorr iiaa,, lleess vvééss iiccuulleess,, lleess ssppoo rreess,, eett
lleess ccee lllluulleess aauuxxiilliiaa iirreess..
LLeess ttyyppeess mmoorrpphhoolloogg iiqquueess ddeess mmyyccoorrhhiizzeess aarrbbuussccuullaa iirreess ((MMAA))
LLaa ccaappaacc iittéé ddeess CCMMAA àà ccoo lloonniisseerr uunnee pp llaannttee hhôôttee ee tt àà lluuii ffoouurrnniirr ddeess nnuuttrr iimmeennttss dd iiffffèèrree
ssee lloo nn llee ttyyppee ddee ccoo lloo nniissaatt iioonn aa rrbb uussccuullaa iirree ((MMuuttkk uummaarr ee tt PPrraakkaasshh,, 22000099 )).. LL‟‟eexxpp rreessss iioonn dduu ttyyppee
mmoorrpphhoo llooggiiqq uuee ddee llaa ssyymmbb iioo ssee eesstt ccoo nnttrrôô llééee ppaarr llee ggéénnoommee ddee llaa pp llaannttee hhôô ttee ((SS mmiitthh eett SS mmiitthh,,
11999977)) eett llee ggéénnoo mmee ffoonnggiiqquuee ((CCaavvaaggnnaarroo eett aall.. ,,22000011))..
GGaa llllaauudd ((11990055)) aa rrééppaarr tt ii lleess mmyyccoorrhhiizzeess aarrbb uussccuullaa iirreess ((MMAA)) eenn ddeeuuxx cc llaasssseess mmoorrpphhoo llooggiiqquueess
mmaajjeeuurreess,, llee ttyyppee «« AArruumm »» eett llee ttyyppee «« PPaarr iiss »»..
-- TTyyppee «« AArruumm »»
CCee ttyyppee ee sstt ddoommiinnaanntt cchheezz lleess pp llaanntteess ccuulltt iivvééeess ((SS mmiitthh eett SS mmiitthh,, 11999977 ;; YYaammaattoo,,
22000044 ;;DDiicckkssoonn eett aall..,,22000077 )) ee tt ddaannss lleess ééccoossyyss ttèèmmeess nnaa ttuurree llss ((OO‟‟ccoo nnnnoo rr eett aall..,, 22000011 ;;
MMuutthhuukk uummaarr eett aall..,, 22000033 ee tt 22000066 ;; FF iisshhee rr ee tt JJaayyaacchhaannddrraann,, 22000055 ;; YYaammaattoo,, 22000055)).. CCee
mmoorrpphhoottyyppee eesstt ccaa rraaccttéé rr iisséé ppaarr uunnee eexxtteennss iioonn rraapp iidd ee dd uu cchhaammpp iiggnnoonn àà tt rraavveerrss lleess eessppaacceess
aappoopp llaasstt iiqquueess,, eennttrree lleess ccee lllluu lleess ccoo rrtt iiccaa lleess dduu ppaarreenncchhyymmee rraacc iinnaa iirree,, lleess vvééss iiccuulleess,, ss ii pprréésseenntteess,,

43
ssoonntt iinntt rraa oo uu iinntteerrccee lllluullaa iirreess,, lleess aa rrbb uussccuulleess ssoonntt tteerrmmiinnaauuxx ssuurr ddeess bb rraanncchheess iinnttrraaccee lllluullaa iirreess
(( ffiigg..2244)) ((DDiicckkssoo nn eett aall..,, 22000077)).. LLeess ppee lloo ttoo nnss ssoonntt ffaaccuullttaatt iiffss,, ee tt nnee ssoo nntt ppaass uunn ccoo mmppoo ssaanntt
mmaajjeeuurr dd uu mmyyccéé lliiuumm iinnttrraarraacc iinnaa iirree ((WWuubbee tt eett aall..,, 22000033)) ..
DDaannss llaa MMAA ddee ttyyppee «« AArruumm »»,, llee mmyyccéé lliiuumm iinnttee rrccee lllluullaa iirree ss ‟‟éétteenndd dd aannss ll‟‟eessppaaccee
iinntteerrccee lllluullaa iirree dd uu ccoo rrtteexx rraacc iinnaa iirree,, ccee qq uuii ffaacc iill iittee ssaa pprrooppaaggaa tt iioo nn rraapp iidd ee eett llee ttrraannss ffeerr tt ddeess
nnuuttrr iimmeennttss ((DDiicckkssoonn eett aall..,, 22000077)),, eenn ee ffffeett,, llee ddéévvee llooppppeemmeenntt ddee llaa MMAA dduu ttyyppee «« AArruumm »» eesstt
pplluuss rraapp iiddee qq uuee ccee lluuii ddee ttyyppee «« PPaarriiss »» ((BBrruunnddrree tttt ee tt KKuunnddrr iicckk,,11999900))
FFiigg.. 2244 :: AAssppeecctt dd’’uunn aa rrbbuussccuullee ddaa nnss llaa MMAA ddee ttyyppee «« AArruumm »» ((SS mmiitthh eett aall ..,, 22000066))..
-- TTyyppee «« PPaarriiss »» CCee ttyyppee ddee MMAA eesstt ddoommiinnaanntt cchheezz lleess lliiggnneeuuxx ((YYaammaa ttoo eett IIwwaazzaakk ii,,
22000022)),, cchheezz GGeenntt iiaannaa lluutteeaa ((Gianinazzi -Pearson eett GGiiaanniinnaazzzzii,, 11998833)),, iill eess tt ccaarraacc ttéérr iisséé
ppaarr llaa pprréésseennccee ddee ppee lloottoo nnss iinnttrraaccee lllluullaa iirreess,, dd eess ppee lloo ttoonnss aarrbbuussccuullaa iirreess ((CCaavvaaggnnaarroo eett aall..,,
22000011)) (( ffiigg..,, 2255 )) ,, ddeess vvééss iiccuulleess iinnttrraaccee lllluu llaa iirreess ((GGuutt tteennbbeerrggee rr,, 22000000 ;; WWuubbee tt eett aall..,, 22000033 ;;
MMuutthhuukk uummaarr eett PPrraakk aasshh,, 22000099)),, ee tt ppaarr ll‟‟aabbsseennccee ddee mmyyccéé lliiuumm iinntteerrccee lllluu llaa iirree ((WWuubbee tt eett
aall..,, 22000033))..
FFiigg.. 2255 :: AAss ppeecctt dd’’uunn ppeelloottoonn aarrbbuussccuullaaiirree ((aacc)) ddaannss llaa mmyyccoorrhhiizzee MMAA ddee ttyyppee «« PPaarriiss »»
((MMuutthhuukkuummaarr ee tt PPrraakkaass hh,, 22000099))..

44
LLee cchhaammpp iiggnnoo nn ssee pprrooppaaggee dd iirreecc tteemmeenntt dd‟‟uunnee ccee lllluullee àà uunnee aauuttrree àà ll‟‟ iinnttéé rr iieeuurr dduu ccoo rrtteexx
rraacc iinnaa iirree ee tt ffoo rrmmee pp lluuss iieeuurrss ppee lloottoonnss,, àà ppaarrtt iirr ddeessqquuee llss,, lleess aarrbb uussccuulleess ssoonntt ssoouuss ffoorrmmee ddee
ssttrruucc ttuurreess iinnttee rrccaa llaa iirreess (( ffiigg.. 2266))..
FFiigg.. 2266 :: MM yyccéé lliiuumm iinnttrraaccee lllluullaaiirree ssee pprrooppaaggee aanntt ee nnttrree ddee uuxx ccee lllluulleess ccoorrttiiccaalleess ee tt ffoorrmmaanntt ddeess
aarrbbuuss ccuulleess iinnttee rrccaallaaiirreess ((MM uutthhuukkuummaarr ee tt PPrraakkaass hh,, 22000099)).. aa :: AArrbbuuss ccuullee ;; iihh :: hhyypphhee
iinnttrraaccee lllluullaaiirree ;; ccww :: ppaarrooii ccee lllluullaaiirree ..
PPlluuss iieeuurrss eessppèècceess vvééggéé ttaa lleess ffoorrmmeenntt lleess ddeeuuxx ttyyppeess mmoorrpphhoo lloo ggiiqquueess ddee MMAA ((AArruumm eett
PPaarr iiss)) :: llaa ccaarroo ttttee eenn ccuullttuurree mmoonnooxxéénniiqquuee pprréésseennttee ddeess MMAA dd ee ttyyppee «« AArruumm »» bb iieenn dd iifffféérreenncc iiéé
oouu uunn ttyyppee iinntteerrmméédd iiaa iirree aavveecc GGll.. eettuunniiccaattuumm oouu eennccoorree lleess ddeeuuxx mmoorrpp hhoo ttyyppeess eenn ccoommmmuunn
ddaannss lleess ccee lllluulleess ccoorrtt iiccaa lleess ((SS mmiitthh eett RReeaadd,, 11999977)).. Le type intermédiaire de MA a aussi été
observé chez quatre espèces de palmiers infectés par Glomus mosseae (Nicol. & Gerd.)
Gerdemann & Trappe (Dreyer et al., 2009).
33.. RRôôlleess ee tt iimmppoo rrttaa nnccee ddeess MMAA ppoouurr llee ss ppllaa nntteess
33..11.. Effet sur la croissance et le développement ddeess lliiggnneeuuxx
LLaa pprréésseennccee ddeess CCMMAA ppeerrmmeett uunnee mmee iill lleeuurree ccrroo iissssaannccee ddeess lliiggnneeuuxx,, ll‟‟uunnee ddeess ffoonncc tt iioonnss
mmaajjeeuurreess ddee cceett ttee aassssoocc iiaa tt iioo nn eess tt ddee ppeerrmmeett ttrree aauuxx pp llaannttee ss ddee ss‟‟aa jjuusstteerr aauuxx ccoonndd iitt iioonnss llooccaa lleess
dduu ssoo ll eett ddee ss‟‟aacccc lliimmaa tteerr ((RRaa ii,, 22000011 ;; PPoo rrrraass Soriano eett aall..,, 22000022 )).. CCeett ttee ssyymmbb iioossee ppeeuutt
éévvoo lluueerr,, ssoo uuss ddeess ccoonndd iitt iioo nnss lliimmiittaanntteess vveerrss ddeess ccoommbb iinnaa iissoo nnss aa rrbbrree //cchhaammpp iiggnnoonn pp lluuss
rroobbuusstteess ee tt ee ffffiiccaacceess,, ee llllee ppeeuutt mmêêmmee jjoouueerr llee rrôô llee ddee mméédd iiaa tteeuurr ddeess iinntteerraacctt iioo nnss eennttrree lleess
aarrbbrreess ((HHaarr tt eett aall..,, 22000033 ;; EEnnkkhhttuuyyaa eett VVooss ttaakkaa,, 22000055))..

45
LLeess CCMMAA ccoonnttrr iibbuueenntt àà llaa ssuurrvviiee,, llaa cc rroo iissssaannccee ee tt llaa pp rroodduucctt iivviittéé ddeess aa rrbbrreess ffrruuiitt iieerrss ppaarr
llee rrôô llee bbéénnéé ffiiqquuee qquuee jjoouuee ccee ttttee aassssoocc iiaa tt iioo nn ddaannss llaa nnuuttrr iitt iioonn mmiinnéé rraa llee ((BBaa eett aall..,, 22000011 ;;
CCaammpprruubb ii eett aall..,, 22000000)) eett hhyyddrr iiqquuee ((AAuuggéé eett aall..,, 22000044)),, llaa fflloorraa iissoo nn eett llaa ffrruucctt iiffiiccaa tt iioo nn ddee
ll‟‟aarrbb rree (Azcon-Aguilar et Barea, 1997), llaa ttoo lléé rraannccee aauuxx ss ttrreessss bb iioo tt iiqq uueess ((LLiinnddeerrmmaannnn,, 22000000))
eett aabb iioo tt iiqquueess ((CChhoo eett aall..,,22000066 )) eett ll‟‟aaggrrééggaatt iioo nn dduu ssoo ll ((RRiill lliigg ee tt MMuummmmyy,, 22000066))..
LLeeuurr ee ffffeett ssuurr llee ddéévvee llooppppeemmeenntt ee tt llaa ccrroo iissssaannccee dd uu ssyymmbb iioottee vvééggéé ttaa ll aa éé ttéé ééttuudd iiéé ppaarr
pplluuss iieeuurrss aauutteeuurrss,, ((DDuuccoo uussssoo eett PPoo lloo nnaa,, 11999944 ;; JJaa iizzmmee –– VVeeggaa ee tt AAzzccoonn,, 11999955 ;; RRaavviikk uummaarr eett
aall..,, 11999977 ;; BBaa eett aall..,, 22000011 ;; JJaa iizzmmee --VVeeggaa eett aall..,, 22000033 ;; BBiinneett eett aall..,, 22000077 ;; EEcchhaa iirr ii eett aall..,,
22000088 ;; SSaaaadd,, 22000099)) ;; iill ssee ttrraadd uuiitt ppaarr ll‟‟aauuggmmeennttaatt iioonn ddee llaa hhaauutteeuurr eett dduu ppoo iiddss ddeess pp llaannttss
mmyyccoorrhhiizzééss ppaarr rraappppoorrtt àà ddeess pp llaannttss nnoonn mmyyccoorrhhiizzééss eett ll‟‟ iimmppoo rrttaannccee dd uu ssyyssttèèmmee rraacc iinnaa iirree..
DDee nnoo mmbbrreeuuxx eexxeemmpp lleess ddee ttrraavvaauuxx oonntt mmoo nnttrréé qquuee lleess ssyymmbb iioosseess mmyyccoo rrhhiizziieennnneess
aamméé lliioorreenntt llaa ccrroo iissssaannccee :: ccaass ddeess pp llaannttss dd‟‟aarrggaanniieerr mmyyccoo rrhhiizzééss ppaa rr uunnee ssoouucchhee ccoommmmeerrcc iiaa llee
ddee GGll.. IInnttrraarraadd iicceess ((EEcchhaa iirr ii eett aall.. 22000088)) oo ùù llaa hhaauutteeuurr mmooyyeennnnee ddeess pp llaannttss ââggééss ddee 66 mmoo iiss
eesstt 33 àà 44 ffoo iiss ssuuppéérr iieeuurree àà ccee llllee ddeess ttéémmoo iinnss ((NNoouuaa iimm ee tt Chaussod, 1996 )) ;; ccaass des variétés
Picual et Arbequina d‟olivier mycorhizés par Gl. intraradices (MYCOSYM TRI-TON,
souche commercialisée) (de Rougemont, 2008). DDeess rrééssuullttaattss sseemmbb llaabb lleess oo nntt aauussss ii ééttéé
oobbtteennuuss cchheezz 4 arbres fruitiers (Balanites aegyptiaca L. Del., Parkia biglobosa (Jacq.) Benth.,
Tamarindus indica L. et Zizyphus mauritiana Lam) des systèmes agroforestiers des zones
arides et semi-arides mycorhizés par Glomus aggregatum Schenck & Smith emend. Koske
(Guissou et al., 2001), et chez des pp llaannttss ddee bbaammbboouu iinnffeecc ttééss ppaarr GGll.. aaggggrreeggaattuumm eett GGll..
FFaasscciiccuullaattuumm ((RRaavviikk uummaarr eett aall..,, 11999977))..
Un effet positif sur la croissance, la fixation de l‟azote et la nodulation de Casuarina
equisetifolia, d’Acacia cyanophylla et de Faiderbia albida associé Gl. mosseae a été rapporté
par certains auteurs (Cornet et Diem, 1982 ; Nasr et Diem, 1988 ; Ducousso et Polona, 1993).
DDeess ccaass ppaarrtt iiccuulliieerrss dd‟‟aabbsseennccee dd‟‟amélioration de la croissance de plants infectés par un
seul champignon ou deux ensembles oonntt ééttéé ss iiggnnaa llééss ttee ll eess tt llee ccaass ddee Tripleurospermum
inodorum et Calamagrostis epigejos mycorhizées associées à Glomus intraradices et Gl.
claroideum (Janoušková et al., 2009).
LLeess CCMMAA ccoonnttrriibbuueenntt aauussssii àà llaa ssuurrvviiee ddeess vviittrrooppllaannttss lloorrss ddee lleeuurr ttrraannssppllaannttaattiioonn eett ddee
lleeuurr aacccclliimmaattaattiioonn ddaannss lleeuurr nnoouuvveeaauu mmiilliieeuu ((GGiilllleemmiinn eett GGiiaanniinnaazzzzii,, 11999944 ;; JJaaiizzmmee--VVeeggaa eett
aall..,, 22000022 ;; BBiinneett eett aall..,, 22000077))..

46
33..22.. EEffffee tt ssuurr llaa nnuuttrriittiioo nn mmiinnéérraallee ddeess ppllaa nntteess
LLeess aassssoocc iiaatt iioo nnss mmyyccoo rrhhiizziieennnneess nnaattuurree lllleess ssoonntt pp lluuss ffrrééqquueenntteess ddaannss lleess ssoo llss ooùù llaa
ddiissppoonniibb iilliittéé ddeess éé lléémmeennttss mmiinnéérraauuxx eesstt ffaa iibb llee ((BBaa eett aall..,, 22000011 )),, eenn ee ffffeett,, lleess aarrbbrreess aamméé lliioo rreenntt
lleeuurr nnuuttrr iitt iioo nn aazzoo ttééee eett pphhoosspphhoorrééee àà ttrraavveerrss llee ppaarrtteennaa iirree ffoonnggiiqquuee ((GGoovviinnddaarraajjuulluu eett aall..,,
22000055 ;; SS mmiitthh eett RReeaadd,, 22000088 ))..
LLee cchhaammpp iiggnnoo nn àà ll‟‟aa iiddee dd uu rréé sseeaauu eexxttrraammaattrr iicc iiee ll eexxpp lloo rree uunn vvoo lluummee pp lluuss iimmppoo rrttaanntt ddee
ssoo ll eett ppeerrmmee tt àà llaa pp llaannttee ddee ss‟‟aapppprroo vviiss iioonnnneerr eenn éé lléémmeennttss mmiinnéérraauuxx ccoo mmmmee llee SS ((AAlllleenn eett
SShhaacchhaarr,, 22000099)),, llee CCuu,, ZZnn,, MMnn,, FFee eett llee KK ((SSoorreennsseenn eett aall..,, 22000055 ;; CCaarrddoossoo eett aall..,, 22000066 ;;
GGuueetthhee rr eett aall,, 22000099)) ..
LLaa mmoobb iilliissaatt iioo nn dduu pphhoo sspphhoorree àà ppaa rrtt iirr dd uu pp hhoosspp hhaa ttee nnaa ttuurree ll ((PPNN)) ((CCaarrddoossoo eett aall..,, 22000066 ;;
BBaabbaannaa eett AAnnttoo uunn,, 22000066)) eett ll‟‟aabbssoorrpp tt iioonn dduu pphhoosspphhaa ttee iinnoorrggaanniiqq uuee ddee llaa ssoo lluutt iioo nn dd uu ssoo ll ppaarr
lleess CCMMAA ssoo nntt lleess pprroocceessssuuss cc llééss qquuii ppeerrmmeett tteenntt dd ‟‟aamméé lliioo rreerr llaa ccrroo iissssaannccee ddeess vvééggééttaauuxx
((BBuusshhee rr,, 22000077))..
33..33.. EEffffee tt ssuurr llee ss ffaacctteeuurrss bbiioo ttiiqquueess ee tt aabbiioott iiqquueess
33..33..11.. LLeess ffaacctteeuurrss aabbiioottiiqquueess
LLeess CCMMAA ccoonnttrr iibb uueenntt aauu mmaa iinntt iieenn ee tt aauu ffoonncc tt iioonnnneemmeenntt ddee ll‟‟ééccoossyyssttèèmmee eenn ee ffffeett,, llaa
mmyyccoorrhhiizzee aarrbbuussccuullaa iirree aauuggmmeennttee llaa pprroodd uucctt iioonn ee tt llaa ssuurrvviiee ddeess pp llaanntteess ddaannss lleess ccoonndd iitt iioonnss ddee
ssttrreessss aabb iioott iiqq uuee ((RRuuiizz-- LLoozzaannoo,, 22000033 ;;AAuuggéé,, 22000011 ,, CChhoo eett aall..,, 22000066 ,,CCoorrnnee tt eett DDiieemm 11998822 ;;
SSttuurrmmeerr,, 11999988 ;; TTaaoo eett ZZhhiiwwee ii..,, 22000055))..
LLee ss ttrreessss aabb iioo tt iiqquuee ttee ll qquuee llaa sséécchheerreessssee,, lleess eexxttrrêêmmeess tteemmppéérraa ttuurreess,, llaa ssaa lliinniittéé ppaarr ttaaggeenntt
uunn ccoommppoossaanntt oo ssmmoott iiqquuee ccoo mmmmuunn llee ddéé ffiicc iitt eenn eeaauu ppoouurr lleess pp llaanntteess ((WWaanngg eett aall..,, 22000033 eett
22000044)) .. II ll rreepprréésseennttee uunnee sséé rr iieeuussee mmeennaaccee ppoo uurr ll‟‟aaggrr iiccuullttuurree ((PPoorrccee ll eett aall..,, 22000066 )) ..
LLaa ttoo lléérraannccee ddee llaa pp llaannttee mmyyccoorrhhiizzééee àà llaa sséécchhee rreessssee (( ffaa iibb llee hhyyggrroo mmééttrr iiee ddee ll‟‟aa iirr))
ss‟‟ee ffffeeccttuuee àà ddeess nniivveeaauuxx ccee lllluu llaa iirreess,, pphhyyss iioo lloo ggiiqquueess nnuuttrr iitt iioonnnnee lllleess eett pphhyyss iiqquueess ((RRuuiizz--
LLoozzaannoo,, 22000033))
DD‟‟aauuttrreess mmééccaanniissmmeess iinntteerrvviieennnneenntt ddaannss llaa ttoo lléérraannccee ddeess pp llaannttee ss mmyyccoorrhhiizzééeess àà llaa
sséécchheerreessssee ttee llss qq uuee ll‟‟aabb ssoorrpptt iioonn dd iirreeccttee ee tt llee tt rraannssffee rrtt dd‟‟eeaauu àà ttrraavvee rrss llee mmyyccéé lliiuumm ffoonnggiiqquuee
cchheezz lleess pp llaanntteess hhôôtteess (( MMaarruullaannddaa eett aall..,,22000033 )),, uunn mmee iilllleeuurr aajjuusstteemmeenntt oo ssmmoott iiqquuee ddeess pp llaanntteess

47
((KKuubb iikkoo vvaa eett aall..,, 22000011)),, uunnee aamméé lliioorraa tt iioonn ddeess éécchhaannggeess ggaazzeeuuxx ddee llaa pp llaannttee ((GGrreeeenn eett
aall..,,11999988)),, ddeess cchhaannggeemmeennttss ddaannss llaa rréé tteenntt iioo nn dd‟‟eeaauu ((AAuuggéé,, 22000011 ;; AAuuggéé eett aall..,, 22000044)) eett llaa
pprrootteecctt iioo nn ccoonnttrree lleess ddoo mmmmaaggeess ooxxyyddaatt iiffss ggéénnéérrééss ppaarr llaa sséécchheerreessssee ((PPoorrccee ll eett aall..,,22000033 ;;
PPoorrccee ll eett RRuuiizz-- LLoozzaannoo,, 22000044))..
SSeelloonn PPoo rrccee ll eett aall.. ((22000066)),, llaa pprroottéé iinnee ((1144--33--33)) pprréésseennttee cchheezz GGlloommuuss iinnttrraarraadd iicceess eess tt
iimmpp lliiqquuééee ddaannss llaa pprroo tteecctt iioonn // ll‟‟aamméé lliioorraatt iioonn ddee llaa ttoo lléérraannccee aauu ssttrreessss hhyyddrr iiqquuee..
Fig. 27 : Mécanisme d’absorption des minéraux et des métaux lourds par les CMA et leur
translocation vers la plante (Giasson et al., 2008).
MyIR : Mycélium intra-racinaire ; MyER : Mycélium extra-racinaire
LLeess MMAA aamméé lliioorreenntt llaa ccrroo iissssaannccee ee tt llaa ssuurrvviiee ddeess pp llaanntteess ddaannss lleess ss iitteess ppeerr ttuurrbbééss
iinncc lluuaanntt cceeuuxx ccoo nnttaammiinnééss ppaarr lleess mméé ttaauuxx lloo uurrddss,, ppaarr ffoo iiss pp rréésseennttss eenn eexxccèèss ddaannss ccee rrttaa iinnss ssoo llss
aacc iiddeess ((AAll,, CCdd,, ZZnn,, AArr,, CCrr)) ((BBii eett aall..,,22000033 ;; VViivvaass eett aa ll ..,,22000055 ;; WWaanngg eett aall..,,22000055 ;; AAhhmmeedd eett
aall..,,22000066 ;; PPooppee eett aall..,, 22000077 ;; XXiiaa eett aall..,, 22000077 ;; UUllttaa eett aall ..,,22000077;; KKhhaaddee eett AAddoo lleeyyaa,, 22000077 eett
22000099 ;; SS mmiitthh eett aall ..,, 22001100 )).. EElllleess aamméé lliioorreenntt ééggaa lleemmeenntt llaa nnuuttrr iitt iioonn mmiinnéérraa llee ddeess pp llaanntteess
ccuulltt iivvééeess ssuurr ddeess ssoo llss ccoonnttaammiinnééss ppaa rr lleess mméé ttaauuxx lloouurrdd (( VViivvaass eett aall..,, 22000033 ;; WWaanngg eett
aall..,,22000055 )) eett rréédd uuiisseenntt lleeuurr tt rraannssppoorr tt vveerrss llaa ppaarr tt iiee aaéé rr iieennnnee ddee llaa pp llaannttee ((VViivvaass eett aall..,, 22000055))
cceess ddeerrnniiee rrss ssoonntt mmiieeuuxx ttoo lléérrééss (( GGaauurr eett AAddoo lleeyyaa,, 22000044))..
LLeess mmééccaanniissmmeess iimmpp lliiqquuééss ddaannss ll‟‟aaccccuummuullaa tt iioonn ddeess mméé ttaauuxx ssoo nntt :: llaa dd iilluutt iioonn ddeess
éé lléémmeennttss ttoo xxiiqquueess ddaannss llaa pp llaannttee,, lleeuurr ssééqquueess ttrraa tt iioonn ddaannss lleess ss ttrruucc ttuurreess ffoonnggiiqquueess eett
My ER
Spores Racines mycorhizées
Arbuscule My IR
My ER
Absorption des mmiiccrrooéélléé mmeennttss
eett ddeess mméétt aauu xx ll oouurr ddss
Accumulation des
métaux dans les racines

48
ll‟‟aamméé lliioo rraatt iioo nn ddee lleeuurr ttoo lléérraannccee ppaarr lleess CCMMAA.. LLeess mmyyccéé lliiuummss iinnttrraa ee tt eexxttrraa mmaattrr iicc iiee ll,,
ccoonnttrr iibbuueenntt dd iirreecctteemmeenntt àà ll‟‟aabbssoo rrpptt iioo nn ee tt llaa tt rraannss llooccaatt iioonn ddeess mmiiccrrooéé lléémmeennttss ((CC uu,, ZZnn)),, eett ddeess
éé lléémmeennttss ttooxxiiqquueess ((CCdd,, AArr,, PPbb)) vvee rrss lleess rraacc iinneess ddee ll‟‟hhôôttee ((LLeeyyvvaa ll eett aall..,, 22000022 ))(( ffiigg.. 2277))..
CCeeppeennddaanntt,, lleess ccoo nncceennttrraa tt iioonnss éé lleevvééeess eenn CCrr,, CCdd ee tt PPbb ppeeuuvveenntt aavvoo iirr uunn ee ffffeett nnééggaatt iiff
ssuurr lleess CCMMAA ddaannss lleess ss iitteess ccoonnttaammiinnééss,, eenn rree ttaarrddaanntt,, rréédduuiissaanntt eett mmêêmmee eenn éé lliimmiinnaanntt
ccoommpp llèè tteemmeenntt llaa ccoo lloonniissaatt iioo nn mmyyccoorrhhiizziieennnnee ee tt llaa ggeerrmmiinnaa tt iioonn ddeess ssppoorreess dduu cchhaammpp iiggnnoonn
((KKhhaaddee ee tt AAddoo lleeyyaa,, 22000099))..
33..33..22.. IInnttee rraacctt iioonn ddeess MMAA--mmiiccrroooorrggaanniiss mmeess
33..33..22..11.. MMiiccrrooff lloorree ttee lllluurriiqquuee ee tt mmyyccoorrhhiizzoosspphhèèrree
LLaa ffoorrmmaatt iioonn ddee llaa MMAA cchhaannggee qq uuaanntt iittaatt iivveemmeenntt ee tt qquuaa lliittaatt iivveemmeenntt,, lleess ppoopp uullaa tt iioonnss
mmiiccrroobb iieennnneess ddee llaa rrhhiizzoosspp hhèè rree ((BBaarreeaa eett aall..,, 22000022 ;; JJoo hhaannssssoonn eett aall..,, 22000044 )),, eenn ee ffffeett,, lleess
hhyypp hheess eexxttrraa rraacc iinnaa iirreess ddeess MMAA ccoonnss tt iittuueenntt jjuussqquu''àà 8800 %% ddee llaa mmaassssee mmiiccrroobb iieennnnee,, aavveecc pp rrèèss ddee
115500 ccmm//ccmm33
ddee ssoo ll ((KKaabb iirr eett aall..,,11999977)).. LLeess CCMMAA iinntteerraaggiisssseenntt aavveecc lleess bbaaccttéé rr iieess,, lleess
cchhaammpp iiggnnoonnss ee tt dd‟‟aauuttrreess mmiiccrroooorrggaanniissmmeess aassssoocc iiééss àà llaa pp llaannttee ee tt iinnfflluueenncceenntt lleess iinnttee rraacctt iioonnss
eennttrree ll‟‟hhôôttee ee tt lleeuurr ssyymmbb iioottee mmiicc rroobb iieenn ((AAzzccoo nn--AAgguuiill llaarr ee tt BBaarreeaa,, 11999966))..
CCeerrttaa iinneess iinntteerraacctt iioonnss eennttrree lleess CCMMAA eett lleess mmiicc rroooorrggaanniissmmeess dduu ssoo ll aa ffffeecc ttaanntt
ppooss iitt iivveemmeenntt llaa ccrroo iissssaannccee ddee llaa pp llaannttee ttee ll eess tt llee ccaass ddeess RRhhiizzoobb iiuummss ((bbaaccttéé rr iieess ffiixxaa ttrr iicceess
dd‟‟aazzootteess )) qquuii ccoo lloonniisseenntt ss iimmuu llttaannéémmeenntt aavveecc lleess CCMMAA lleess rraacc iinneess ddee LLéégguummiinneeuusseess,, ssaannss
ccoommppéétt iitt iioonnss ppoo uurr lleess ss iitteess dd ‟‟ iinnffeecctt iioonn ((XXaavviiee rr eett GGeerrmmiiddaa,, 22000022 ;; BBaarreeaa eett aall..,, 22000055)).. CCeess
bbaaccttéérr iieess aadd hhèèrreenntt àà ll‟‟hhyypp hhee dduu cchhaammpp iiggnnoonn eett aauuxx ppoo llyyssaacccchhaarr iiddeess eexxttrraaccee lllluu llaa iirreess eett
ll‟‟uutt iill iisseenntt ppoouurr ccoolloonniissee rr lleess rraacc iinneess ((BBiiaanncc iioo ttttoo eett aall..,, 22000011)).. CCeettttee iinntteerraacctt iioonn aamméé lliioorree llaa
ccrroo iissssaannccee ddeess pp llaanntteess ssppéécc iiaa lleemmeenntt ddaannss lleess ssoo llss ppeeuu ffeerr tt iilleess ;; cchheezz llee ss pp llaanntteess hhôôtteess,, ee llllee
aamméé lliioorree llaa ffiixxaatt iioonn bb iioo llooggiiqquuee ddee ll‟‟aazzoottee eenn aauuggmmeennttaanntt llaa nnoodd uullaa tt iioonn,, lleeuurr iinnffeecc tt iioonn
mmyyccoorrhhiizziieennnnee,, lleeuurr bb iioo mmaassssee vvééggéé ttaa llee eett llaa qquuaanntt iittéé ddee NN ee tt PP,, ((XXaavviieerr eett GGeerrmmiiddaa,, 22000022))..
EEllllee iinndduuiitt llee mmééccaanniissmmee ddee lleeuurr ttoo lléé rraannccee aauuxx ppaatthhoo ggèènneess ((DDaarr eett aall..,, 11999977)).. DD‟‟aauuttrreess
mmiiccrroooo rrggaanniissmmeess iinnttee rraaggiisssseenntt ppooss iitt iivveemmeenntt aavveecc lleess CCMMAA ccoo mmmmee ::
-- lleess bbaacc ttéérr iieess ee tt cchhaammpp iiggnnoonnss ssoo lluubb iilliissaatteeuurrss ddee pp hhoosspphhaa ttee ddaannss llee ssoo ll (( PPsseeuuddoommoonnaass sspp..
PPeenniicciill lliiuumm cchhrryyssoogg eennuumm,, AAssppeerrggiill lluuss aawwaannoorrii )) ((BBaabbaannaa ee tt AAnnttoouunn ,, 22000066))..
-- lleess PPGGPP RR ((eenn aannggllaa iiss :: PP llaanntt GGrroowwtthh PPrroommoott iinngg RRhhiizzoobbaaccttee rr iiaa)) ssoonntt ddeess bbaaccttéérr iieess lliibb rreess
ddaannss llee ssoo ll ((PPsseeuuddoommoonnaass,, BBaacc iilllluuss)) ((RRaammaammoooo rrtthhyy eett aall..,, 22000011 ;; KK llooeeppppeerr eett aall..,, 22000044))
ddoonntt llaa pprréésseennccee aamméé lliioorree llaa ccoo lloonniissaatt iioonn ddeess rraacc iinneess ppaarr lleess CCMMAA ((BBaarreeaa eett aall..,,22000022)),,

49
sstt iimmuullee llaa ggeerrmmiinnaa tt iioonn ddeess ssppoorreess eett llee ddéévvee llooppppeemmeenntt dd uu mmyyccéé lliiuumm ;; eelllleess ssoonntt aappppee llééeess
lleess MMHHBB ((MMyyccoorrrrhhiizzaa ll HHee llppeerr BBaacc tteerr iiaa)) ((VViivvaass eett aall..,, 22000033 eett 22000055 )) ..
LL‟‟aapppp lliiccaa tt iioonn ccoommbb iinnééee ddeess CCMMAA eett ddeess MMHHBB,, dd ‟‟aa iilllleeuurrss ttrrèèss uutt iilliissééee ddaannss llaa lluuttttee
bbiioo llooggiiqq uuee,, aauuggmmeennttee llaa ccoo lloonniissaa tt iioo nn ddeess rraacc iinneess,, ll‟‟ iinnhhiibb iitt iioo nn ddeess ppaatthhoo ggèènneess,, ((BBuudd ii eett aall..,,
11999999 ;; SS iidddd iikk ii eett AAkkhhttaarr,, 22000088 ;; VVeessttbb eerrgg ee tt CCaassssee llllss,, 22000099 )),, aamméé lliioo rree llaa pprroodduucc tt iioonn ddeess
rraacc iinneess cchheezz lleess pp llaanntteess aa iinnss ii qquuee lleeuurr cc rroo iissssaannccee ee tt lleeuurr nnuuttrr iitt iioonn ((AAzzccoonn--AAgguuiill llaarr eett
BBaarreeaa,,11999966 ))..
LLeess BTPs ((pp llaanntteess ttrraannssggéénniiqquueess Bt mmoodd iiff iiééeess ppaa rr Bacillus thuringiensis) ont par contre
des effets négatifs sur la croissance de l‟hyphe pré-symbiotique des CMA et sur l‟infection
racinaire par l‟arrêt du développement des appressoriums (Liu, 2010). Cet effet a été observé
chez le maïs (Turrini et al. 2004 ; Castaldini et al. 2005) et le riz (Yang et al. 2002 ; Ren,
2006).
33..33..22..22.. MMiiccrroooo rrggaanniiss mmeess ppaatthhooggèènneess
LLeess CCMMAA pprroo ttèèggeenntt llee ss pp llaanntteess ccoo nnttrree llee sstt rreessss bb iioo tt iiqquuee ccaauusséé ppaarr lleess ppaatthhoo ggèènneess dd uu ssoo ll
ffoonnggiiqquueess ((PPhhyyttoopphhttoorraa,, RRhhiizzooccttoonniiaa,, VVeerrtt iicc iill lliiuumm,, GGaaeeuummaannoonnyycceess,, FFuussaarr iiuumm,, TThhiiee llaavv iiooppssiiss,,
PPyytthhiiuumm,, AApphhaannoonnyycceess)) eett cceerrttaa iinnss nnéémmaa ttooddeess ((RRhhoott iilleenncchhuuss,, PPrraa lliilleenncchhuusseett MMeelloo iiddooggyynnee))
((LLiinnddeerrmmaannnn,, 11999944 ;; SS iinngghh eett aall..,, 22000000 ;; AAzzccoo nn--AAgguuiill llaarr eett aa ll..,,22000022 ;; PPoo zzoo eett aall..,, 22000022 ;;
XXaavviiee rr eett BBoo yyee llcchh,, 22000022 ;; EEllsseenn eett aall..,, 22000022 eett 22000033 ;; WWhhiippppss,, 22000044 ))..
DDeess éé ttuuddeess ssuurr llee bbaannaanniieerr oonntt mmoo nnttrréé qquuee ssoo nn iinnooccuullaa tt iioonn aavveecc GGll.. ccaa lleeddoonniiuumm ee tt GGll..
mmaaccrrooccaarrppuumm rréédduuiitt ss iiggnniiffiiccaa tt iivveemmeenntt llaa pp rroodd uucctt iioonn ddee ggaa llllee ssuurr sseess rraacc iinneess ((EEllsseenn eett aall..,,
22000022)) eett aavveecc GGll.. iinnttrraarraaddiicceess llaa rreepprroodd uucc tt iioo nn ddee RRaaddoopphhoo lluuss ssiimmii lliiss (( nnéémmaa ttooddee ll‟‟oo rrddrree ddeess
TTyylleenncchhiiddaa )) eesstt rréédd uuiittee ddee 5500%% ((EEllsseenn eett aall..,, 22000011)),, cceerrttaa iinnss nnéémmaattooddeess ppeeuuvveenntt ppaarr ccoonnttrree
aavvoo iirr uunn ee ffffeett nnééggaatt iiff ssuurr llee ddéévvee llooppppeemmeenntt iinnttrraarraadd iiccuullaa iirree ddeess CCMMAA ((EEllsseenn eett aall..,, 22000022))..
CCeeppeennddaanntt,, llaa ttoo lléé rraannccee aauuxx aatt ttaaqquueess ddeess ppaatthhooggèènneess ddeess pp llaanntteess mmyyccoorrhhiizzééeess vvaa rr iiee ssee lloo nn lleess
eessppèècceess ddee CCMMAA,, llee ss ppaatthhooggèènneess,, lleess ssuubbss ttrraattss eett lleess ccoo nndd iitt iioonnss eennvviirroonnnneemmeennttaa lleess ((BBaarreeaa eett
aall..,, 22000055))..
PPaarr aa iilllleeuurrss,, dd‟‟aauuttrreess ttrraavvaauuxx oonntt mmoonnttrréé qq uuee ll‟‟ iinnooccuullaatt iioonn dduu ccoonnccoommbbrree ppaarr GGll..
eettuunniiccaattuumm ddiimmiinnuuee ffoo rrtteemmeenntt ll‟‟ iinncc iiddeennccee ddee llaa ffuussaa rr iioossee,, aauuggmmeennttee llaa ccoo nncceennttrraatt iioonn dd uu PP
ddaannss lleess tt iiggeess ddee llaa pp llaannttee,, ccee llllee ddee llaa pprroo lliinnee ee tt ll‟‟aacc tt iivviittéé ddee llaa pp rroo lliinnee ooxxyyddaassee,, iinndd iiqq uuaanntt
ll‟‟ee ffffeett dd uu CCMMAA ssuurr ll‟‟aacctt iivvaatt iioonn dduu mmééccaanniissmmee ddee rrééss iissttaannccee cchheezz llaa pp llaannttee ((HHaaoo eett aall..,, 22000055))..

50
LL‟‟eexxttrraa iitt dduu mmyyccéé lliiuumm eexxttrraarraacc iinnaa iirree ddee GGll.. iinnttrraarraaddiicceess rréédd uuiitt llaa ggee rrmmiinnaatt iioonn ddee FFss..
qqxxyyssppoorruumm ((FFiilliioonn eett aall..,, 11999999))..
LLee ppootteenntt iiee ll ddeess CCMMAA ccoommmmee aaggeennttss ddee lluutt ttee bb iioo lloo ggiiqquuee ccoonnttrree llee ss iinnffeecc tt iioonnss
ppaarraass iittaa iirreess eesstt rree lliiéé àà cc iinnqq mmééccaanniissmmeess dd‟‟aacctt iioo nn ((AAzzccoonn--AAgguuiillllaa rr eett aall..,,22000022 ;; PPoozzoo eett aall..,,
22000022 ;; DDaa llppéé,, 22000055 )) ssuusscceepptt iibb lleess dd‟‟êê ttrree iinnff lluueennccééss pp aarr ddeess ffaacc tteeuurrss aabb iioo tt iiqq uueess (( sséécchheerreessssee,,
tteemmppéérraattuurree oouu ttaauuxx ddee ffeerr tt iilliissaa tt iioonn)) oouu bb iioo tt iiqq uueess ((ppaarraass iittee ,, mmiiccrroo fflloorree dd uu ssoo ll ,, ggéénnoottyyppee ddee llaa
ppllaannttee mmyyccoorrhhiizzééee ((DDaa llppéé,, 22000055 ;; VVeessttbbee rrgg eett CCaassssee llllss ,, 22000099)).. CCeess mmééccaanniissmmeess ccoo mmpprreennnneenntt ::
-- llaa ss tt iimmuullaatt iioonn ddee llaa ccrroo iissssaannccee ddee llaa pp llaannttee ppaarr llee bb iiaa iiss dd ‟‟uunn aappppoorr tt aaccccrruuee eenn nnuuttrr iimmeennttss ;;
-- llaa ccoommppéé tt iitt iioonn ppoouurr lleess pp rroodduuiittss ddee pphhoottoossyynntthhèèssee ddee ll‟‟hhôô ttee ee tt ppoouurr llee ss iittee ddee ccoo lloo nniissaa tt iioonn..
EEnn ee ffffeett,, ddaannss lleess rraacc iinneess ffoorr tteemmeenntt ccoo lloo nniissééeess ppaa rr lleess ssoo uucchheess àà ppoo tteenntt iiee ll mmyyccoo rrhhiizziieenn éé lleevvéé,,
lleess hhyypphheess ssaattuurreenntt ddaavvaannttaaggee lleess ss iitteess dd‟‟ iinnffeecctt iioo nn dd iissppoonniibb lleess ppoo uurr lliimmiitteerr llaa ppéénnéé ttrraa tt iioonn ddeess
hhyypp hheess dd ‟‟uunn ppaarraass iittee ddaannss llee ss rraacc iinneess..
-- ll‟‟ iinndduucctt iioo nn dduu cchhaannggeemmeenntt ddee ll‟‟aannaa ttoommiiee eett ddee ll‟‟aarrcchhiitteeccttuurree dd uu ssyyss ttèèmmee rraacc iinnaa iirree.. LLaa
mmyyccoorrhhiizzaatt iioo nn eennttrraa îînnee llaa rraammiiffiiccaa tt iioo nn eett llaa lliiggnniiff iiccaatt iioonn dduu ssyyss ttèèmmee rraacc iinnaa iirree,, ccoo nnsstt iittuuaanntt uunnee
bbaarrrr iièè rree ddee pprroo tteecctt iioo nn ccoo nnttrree llaa pp éénnééttrraatt iioo nn dd uu ppaarraass iittee ss ‟‟aaccccoo mmppaaggnnaanntt dd ‟‟uunnee aaccccuummuullaa tt iioo nn ddee
ccoommppoossééss pp hhéénnoo lliiqquueess eett ssuuiivviiee dd‟‟uunnee aacctt iivviittéé cchhiitt iinnoo llyytt iiqquuee aa llttéérraanntt lleess ppaa rroo iiss dduu ppaa rraass iittee
ffoonnggiiqquuee ((SS yyllvviiaa ee tt CChhee lllleemmii,, 22000011 ))..
-- llaa mmoodd iiffiiccaa tt iioo nn ddee llaa mmiicc rroo fflloo rree rrhhiizzoosspphhéérr iiqquuee,, ssoouuss ll‟‟ee ffffee tt ddeess eexxssuuddaa ttss rraacc iinnaa iirree iinndd uuiittss
ppaarr llaa rraammiiffiiccaa tt iioo nn iinntteennssee ddeess rraacc iinneess..
-- eett eennffiinn ll‟‟aacctt iivvaatt iioo nn dduu mmééccaanniissmmee ddee ddéé ffeennssee cchheezz lleess pp llaanntteess..
44.. EEffffee ttss ddeess ddiifffféérreennttss ffaacctteeuurrss ssuurr llee ss CCMMAA
44..11.. EEffffee ttss ddeess ffaacctteeuurrss lliiééss àà ll’’eennvviirroonnnneemmeenntt
LLeess CCMMAA ppeeuuvveenntt êê ttrree iinnfflluueennccééss ppaarr ddeess mmoodd iiffiiccaa tt iioonnss cchhiimmiiqq uueess eett pp hhyyss iiqquueess ddee
cceerrttaa iinnss ffaacctteeuurrss aauuttoo uurr ddee llaa rraacc iinnee ttee llss qq uuee llee ttyyppee ddee ssoo ll,, llee ss pprraa tt iiqq uueess ccuullttuurraa lleess eett dd‟‟aauuttrreess
ffaacctteeuurrss eennvviirroonnnneemmeennttaauuxx ((ppHH,, tteemmppéérraa ttuurree ee tt hhuummiidd iittéé)) ((CCaarrddoo ssoo eett KK uuyyppeerr,, 22000066 ))..
LLaa ssppoo rruullaa tt iioonn eett llaa ddeennss iittéé ddeess ssppoorreess aassssoo cc iiééeess àà llaa pp llaannttee hhôôttee ssppéécc iiffiiqquuee vvaarr iieenntt aauussss ii
ssee lloo nn lleess ssaa iissoonnss,, lleess ffaacctteeuurrss ééddaapp hhiiqquueess ((ppHH,, ttaauuxx ddee PP eett hhuummiidd iittéé dduu ssoo ll)),, ll‟‟oo mmbbrree,, llaa
ddééppeennddaannccee ddee ll‟‟hhôôttee ,, ll‟‟ââggee ddee llaa pp llaannttee hhôôttee,, llaa ccaappaacc iittéé ddee ssppoorruullaatt iioonn ddeess CCMMAA,, llaa ppéé rr iiooddee
ddee ddoorrmmaannccee,, llaa dd iiss ttrr iibb uutt iioo nn ddeess CCMMAA ddaannss llee ssoo ll ((LLuuggoo eett aall..,, 22000088)).. SSee lloonn ccee tt aauutteeuurr,,

51
ll‟‟aa lltt iittuuddee eesstt uunn ffaacc tteeuurr iimmppoorr ttaanntt qq uuii aaggiitt ssuurr llaa dd iivvee rrss iittéé ddeess CCMMAA,, ccaa rr ll‟‟aa lltt iittuuddee
aaccccoo mmppaaggnnééee dd‟‟uunnee dd iimmiinnuutt iioonn ddee llaa tteemmppéérraattuurree aa ffffeeccttee nnééggaatt iivveemmeenntt cceess cchhaammpp iiggnnoonnss..
LLaa lluummiièè rree eess tt ééggaa lleemmeenntt uunn ffaacctteeuurr lliimmiittaanntt,, eenn ee ffffeett,, ee llllee ss tt iimmuullee llee ddéévvee llooppppeemmeenntt ddeess
CCMMAA ((NNaaggaasshhii ee tt DDoouuddss,, 22000044)).. LL‟‟oommbbrree,, ppaarr ccoonnttrree,, rréédduuiitt nnoonn sseeuulleemmeenntt llaa ccoo lloo nniissaa tt iioonn
ddeess rraacc iinneess ppaarr lleess CCMMAA eett lleeuurr ssppoo rruullaa tt iioonn mmaa iiss aauussss ii llaa rrééppoonnssee ddee llaa pp llaannttee àà llaa mmyyccoorrhhiizzee,,
cceecc ii eesstt pp rroobbaabb lleemmeenntt dd ûû àà uunnee mmaauuvvaa iissee ccrroo iissssaannccee ddeess hhyypp hheess iinntt rraarraacc iinnaa iirreess ee tt dduu rréésseeaauu
dd‟‟hhyypphheess eexxttrraammaa ttrr iicc iiee llss ddaannss llee ssoo ll.. QQuuaanntt àà llaa tteemmppéérraa ttuurree oopptt iimmaa llee,, ee llllee vvaa rr iiee ssee lloonn
ll‟‟eessppèèccee ffoo nnggiiqq uuee ((SScchheenncckk eett aall..,, 11997755)).. SSeess vvaa rr iiaa tt iioonnss iinnff lluueenncceenntt tt rroo iiss ssttaaddeess ddee
ddéévvee llooppppeenntt ddee llaa mmyyccoorrhhiizzee :: llaa ggee rrmmiinnaatt iioo nn ddeess ssppoorreess,, llaa ppéénnéé ttrraa tt iioo nn ddee ll‟‟hhyypphhee ddaannss llaa
rraacc iinnee ee tt ssaa pprroo llii fféérraa tt iioonn àà ll‟‟ iinnttéérr iieeuurr ddeess ccee lllluulleess ((Goltapeh ,, 22000088 ))..
LL‟‟ee ffffiiccaacc iittéé ddeess eennddoopp hhyyttee ss eess tt aauussss ii lliiééee aauu ppHH dduu ssoo ll ((ccoo mmpprr iiss ggéénnéérraa lleemmeenntt eennttrree 55,,66
eett 77)),, ccee ddeerrnniieerr iinnfflluuee ssuurr llaa ggeerrmmiinnaa tt iioonn ddeess ssppoorreess ((DDaanniiee llss eett TTrraappppee ,,11998800)).. PPoouurr lleess
eessppèècceess ddee Glomus , le pH adéquat est compris entre 5 .5 et 6.5 (Porter et al. 1987 ; Wang et
al. 1993), les Acaulosporacées, par contre, sporulent abondamment dans des sols plus acides
(Gai et Liu, 2003).
LLaa rree llaatt iioonn eennttrree llee ppHH dd uu ssoo ll ee tt ll‟‟ee ffffee tt ddee llaa mmyyccoorrhhiizzee ddééppeenndd ddee ll‟‟eesspp èèccee vvééggééttaa llee,, dduu ttyyppee
dduu ssoo ll,, ddeess ffoo rrmmeess dd uu PP eett ddeess eessppèècceess ffoo nnggiiqq uueess ((Goltapeh,, 22000088 ))..
LLeess CCMMAA pp rroottèèggeenntt ggéénnéé rraa lleemmeenntt lleess pp llaanntteess ccoonnttrree llee sstt rreessss ssaa lliinn ;; ddaannss lleess mmiill iieeuuxx àà
ffoo rrttee ssaa lliinniittéé,, llaa pprroodd uucc tt iioonn ddeess pprrooppaagguulleess ddee CCMMAA dd iimmiinnuueenntt ((PP ffee iiffffee rr eett BBlloo ssss ,, 11998888 ))..
44..22.. EEffffee ttss ddeess pprraa ttiiqquueess ccuullttuurraalleess ssuurr lleess CCMMAA
LLaa ssyymmbb iioossee mmyyccoorrhhiizziieennnnee ppoo uurrrraa iitt aavvoo iirr uunn ppootteenntt iiee ll aaggrr iiccoo llee,, ccaa rr ee llllee aaggiirraa iitt ccoo mmmmee
uunn bb iioo-- ffee rrtt iilliissaanntt ((PP lleenncchhee ttttee eett aall..,, 22000055 ;; HHiijjrr ii eett aall..,,22000066))..
LLaa dd iivvee rrss iittéé ddeess ccoo mmmmuunnaauuttééss ddee CCMMAA eess tt pp lluuss ffaa iibb llee ddaannss lleess ssoo llss ccuulltt iivvééss qquuee ddaannss lleess
ssoo llss ddee pp rraa iirr iieess oo uu dd ee ffoorrêê ttss ((DDaanniiee llll eett aall..,, 22000011 ;;OOeehhll eett aall..,, 22000033,, 22000044 ee tt 22000055)) ;; cceett ttee ppee rrttee
ddee dd iivveerrss iittéé aa éé ttéé aa tttt rr iibbuuééee àà uunnee sséé lleecctt iioonn ccoommpp lleexxee ddeess CCMMAA ppaarr ddeess pp rraatt iiqquueess aaggrr iiccoo lleess
ccoommmmee llaa ffeerrtt iill iissaa tt iioo nn mmiinnéérraa llee,, llee tt rraavvaa iill dd uu ssoo ll,, ll‟‟eemmpp lloo ii ddeess ppeesstt iicc iiddeess eett llaa mmoonnooccuullttuurree..
LLee ttyyppee ddee ccuullttuurree pprraatt iiqq uuééee eett llaa nnaattuurree dduu ssoo ll iinnfflluueenntt aauussss ii ssuurr llaa dd iivvee rrss iittéé ddeess CCMMAA ((HHiijj rr ii
eett aall..,, 22000066))..
44..22..11.. EEffffee tt ddee llaa ffeerrtt iilliissaattiioo nn ss uurr llee ss CCMMAA ::

52
LL‟‟aappppoo rrtt ddee ffee rrtt iilliissaanntt mmiinnéérraa ll ssoo lluubb llee aauuggmmeennttee ssaa dd iissppoonniibb iilliittéé ppoo uurr llee ss pp llaanntteess mmaa iiss iill
rréédduuiitt ffoo rrtteemmeenntt eett mmêêmmee ssuupppprr iimmee llaa ssyymmbb iioossee mmyyccoorrhhiizziieennnnee ((BBaa eett aall..,, 22000011)).. DDeess
ccoonncceennttrraa tt iioo nnss éé lleevvééeess eenn aazzoottee eett eenn pphhoosspphhoorree ddaannss llee ssoo ll aauuggmmeenntteenntt llee ss ssyynntthhèèsseess
pprroottéé iiqquueess eett lleess ccoo mmppoossééss pphhoosspphhoorryyllééss cchheezz lleess pp llaanntteess,, dd iimmiinnuueenntt llaa tteenneeuurr eenn ssuucc rreess
ssoo lluubb lleess ddaannss lleess rraacc iinneess,, ee tt ppaarr ccoonnssééqquueenntt llee ttaauuxx ddee ccoo lloonniissaa tt iioo nn mmyyccoorrhhiizziieennnnee ((LLee TTaaccoonn
,,11999999)).. LLeess eennggrraa iiss oorrggaanniiqq uueess (( ffuummiieerr,, ccoommppoosstteess,, rrééss iidd uuee llss ddee ccuullttuurreess,, bboo iiss,, rraammeeaauuxx
ffrraaggmmeennttééss)),, aauu ccoonnttrraa iirree nnee ssoo nntt ppaass nnéé ffaass tteess ppoouurr lleess CCMMAA,, eett ppeeuuvveenntt mmêêmmee lleess sstt iimmuu lleerr
((GGooss lliigg eett aall..,, 22000066 ))..
44..22..22.. TTrraavvaaii ll dduu ssooll ::
LLee ttrraavvaa iill dduu ssoo ll,, ccoo mmmmee llee tt iillllaaggee pprroovvooqquuee llaa ddeesstt rruucctt iioonn dduu rréésseeaauu dd ‟‟hhyypphheess eett
ll‟‟eennffoouuiisssseemmeenntt ddeess pprrooppaagguulleess eenn pprroo ffoo nnddeeuurr ((OOeehhll eett aall ..,, 22000033,, 22000044,, 22000055)),, iill rraa lleenntt iitt llaa
ccoo lloo nniissaa tt iioonn ddeess pp llaanntteess ee tt ll‟‟eexxpp lloorraatt iioonn dduu ssoo ll ppaarr lleess CCMMAA ((GGoossss eett ddee VVaarreennnneess ,,22000022 ;;
KKaabbiirr,, 22000055 )),, eett ppaa rr ccoo nnssééqq uueenntt aa ffffeeccttee llaa nnuuttrr iitt iioo nn ee tt llaa cc rroo iissssaanncceess ddeess pp llaanntteess .. II ll ppeeuutt
ééggaa lleemmeenntt mmoodd iiffiiee rr llaa ss ttrruucc ttuurree ddeess ccoommmmuunnaauuttééss ddee CCMMAA ((JJaannssaa eett aall..,, 22000022 eett 22000033)),, eenn
éé lliimmiinnaanntt cceerr ttaa iinneess ffaammiilllleess dd‟‟AAccaauu lloossppoorraaccééeess ee tt GGiiggaassppoorraaccééeess eett eenn aauuggmmeennttaanntt llaa ddeennss iittéé
dd‟‟aauuttrreess eessppèècceess ddee GGlloo mméérraa lleess ((GGlloommuuss dd uu ggrroouuppee AA )) ((DDoodddd ,, 22000000 )).. CCeess ddeerrnniieerrss ssoonntt
ccaappaabb lleess ddee ddéévvee llooppppeerr uunn rréésseeaauu dd ‟‟hhyypp hheess oo uu ssppoorruulleerr rraapp iiddeemmeenntt aapprrèèss uunnee ppeerr ttuurrbbaa tt iioo nn ddee
ll‟‟ééccoossyyss ttèèmmee ppaarr llee llaabboo uurr ((JJaannssaa,, 22000033 ;; LLeeaakkee eett aall..,, 22000044 ;; OOeehhll,, 22000055 ;; MMaatthhiimmaarraann eett aall..,,
22000055))..
44..22..33.. RRoottaattiioonn ddeess ccuullttuurreess ::
LLeess ffaa iibb lleess dd iivveerrss iittééss ddeess ccuullttuurreess aa iinnss ii qquuee lleess ssyyssttèèmmeess dd ee mmoonnooccuullttuurreess mmoodd iiffiieenntt llaa
ddiivveerrss iittéé ddeess CCMMAA ddaannss lleess ssoo llss ccuulltt iivvééss ee tt aauuggmmeenntteenntt llaa ddeennss iittéé ddee ccee rrttaa iinneess eessppèècceess
ffoonnggiiqquueess pp lluuttôôtt qquuee dd‟‟aauuttrreess ((BBuurrrroowwss eett PP fflleeggeerr,, 22000022 ;; OOeehhll eett aall ..,,22000033)).. II ll eexxiissttee eenn ee ffffeett
uunnee cceerr ttaa iinnee ssppéécc iiffiicc iittééss ddeess CCMMAA ppoouurr lleeuurr hhôô ttee ((HHee llggaassoo nn eett aall..,, 22000022 ;; MMuummmmeeyy eett aall..,,
22000055)) eett llaa ssuucccceessss iioonn aannnnééee aapprrèèss aannnnééee ddee llaa mmêêmmee ccuullttuurree ppeerrmmeett ddee sséé lleecc tt iioo nnnnee rr uunn
nnoommbbrree rréédduuiitt dd ‟‟eessppèècceess ssppéécc iiffiiqq uueess àà cceett ttee pp llaannttee ..
LLeess BBrraassss iiccaaccééeess ee tt lleess CChhéénnooppoodd iiaaccééeess nnee ffoorrmmeenntt ppaass dd‟‟aassssoocc iiaatt iioo nn mmyyccoorrhhiizziieennnnee,, eett
ll‟‟ iinnttrroodduucc tt iioo nn dd ‟‟uunnee ccuullttuurree ddee ccoo llzzaa oouu ddee bbeett tteerraavvee ppaarr eexxeemmpp llee,, nnee ppeerrmmeett ppaass ddee mmaa iinntteenniirr
oouu dd ‟‟aamméé lliioo rreerr ll‟‟ iinnooccuulluumm ffoonnggiiqquuee ddaannss llee ssoo ll,, aauu ccoo nnttrraa iirree,, ee llllee dd iimmiinnuuee llee nnoo mmbbrree ddee
pprrooppaagguulleess vviiaabb lleess eett ddee ssppoorreess qq uuii ddééggéénnèè rreenntt,, eett ppaarr ccoo nnssééqq uueenntt,, ee llllee ppeeuutt rreettaarrdd eerr oouu mmêêmmee
ddiimmiinnuuee rr llaa ccoo lloo nniissaa tt iioonn ddee llaa ccuullttuurree ssuuiivvaannttee ((KKaarraassaawwaa eett aall..,, 22000022 ;; SSoorreennsseenn eett aall..,, 22000055))..

53
PPaarr ccoonnttrree,, ll‟‟ iinnttrroodduucctt iioo nn dd‟‟uunnee pp llaannttee hhaauutteemmeenntt mmyyccoottrroopphhee eenn ttêê ttee ddee rroottaa tt iioonn eess tt bbéénnéé ffiiqquuee
ppoouurr llee ppoo tteenntt iiee ll iinnffeecc tt iieeuuxx mmyyccoorrhhiizzoo ggèènnee dduu ssoo ll,, ccaarr ll‟‟ iinnooccuulluumm llaa iisssséé ppaarr llaa ccuullttuurree
pprrééccééddeennttee iinnfflluueennccee ppooss iitt iivveemmeenntt llaa ccoo lloonniissaatt iioonn ddee llaa ccuullttuurree ssuuiivvaannttee ((PP lleenncchhee ttttee eett aall..,,
11998833 ;; AArr iihhaarraa ee tt KKaarraassaawwaa,, 22000000 ;; SSoo rreennsseenn eett aall..,, 22000055 ))..
44..22..44.. LLeess ppeess ttiicciiddeess
LLeess ppeesstt iicc iiddeess ppeeuuvveenntt ssuupppprr iimmeerr lleess cchhaammpp iiggnnoo nnss ((CCMMAA eett aauuttrreess ppaa tthhooggèènneess)),, lleess
nnéémmaattooddeess eett lleess aaddvveenntt iicceess ppaa rr ccoonnssééqquueenntt,, ll‟‟ee ffffee tt gglloobbaa ll ppeeuutt êêttrree ppoo ss iitt iiff ((ttaabb lleeaauu 33))..
LLeess ffoo nnggiicc iiddeess oonntt uunn ee ffffeett ttoo xxiiqquuee ssuurr llee ss CCMMAA eett lleess hhee rrbb iicc iiddeess ppeeuuvveenntt aauussss ii aavvoo iirr
uunn ee ffffeett nnééggaa tt iiff dd iirreecctt ssuurr lleess ppooppuullaatt iioo nnss ddeess CCMMAA,, eenn éé lliimmiinnaanntt uunn hhôôttee ppoo tteenntt iiee ll ppoo uurr llaa
ssuurrvviiee ddeess cchhaammpp iiggnnoonnss ((GGooss lliigg eett aall..,, 22000066 ))..
PPaarr aa iilllleeuurrss,, lleess mmiiccrroooo rrggaanniissmmeess uutt iilliissééss eenn lluutt ttee bb iioo lloo ggiiqq uuee ppeeuuvveenntt êêtt rree iinnttéérreessssaannttss,, lleess
PPGGPPRR qq uuii iinntteerraaggiisssseenntt aavveecc lleess CCMMAA,, aauuggmmeenntteenntt llee nnoo mmbbrree ddee pprrooppaagguulleess ddaannss llee ssoo ll..
TTaabblleeaauu 33 :: EEffffeett ddeess dd iifffféé rreenntteess tteecchhnniiqquueess ccuullttuurraa lleess ssuurr lleess CCMMAA ((dd ‟‟aapprrèèss GGooss lliigg eett aall..
((22000066))..
PPrraattiiqquueess EEffffeettss
LLeess pprraattiiqquueess bbéé nnéé ffiiqquueess
--UUttiilliissaattiioonn ddee ffeerrttiilliissaannttss àà bbaassssee ssoolluubbiilliittéé --LLeess bbaasssseess ccoonncceennttrraattiioonnss ddee nnuuttrr iimmeennttss
uuttiilliissaabblleess ddaannss llee ssooll ffaavvoorr iisseenntt llaa
mmyyccoorrhhiizzaatt iioonn..
--LL‟‟eexxcclluussiioonn ddeess bbiioocciiddeess --QQuueellqquueess bbiioocc iiddeess oonntt uunn eeffffeett ttooxxiiqquuee ssuurr lleess
CCMMAA..
--RRoottaattiioonnss ddiivveerrsseess --FFaavvoorriisseenntt llaa ddiivveerrssiittéé ddeess CCMMAA..
LLeess pprraattiiqquueess nnuuiiss iibblleess
--LLee ttiillllaaggee mmééccaanniiqquuee ddaannss llaa lluuttttee ccoonnttrree lleess
mmaauuvvaaiisseess hheerrbbeess..
--DDeessttrruuccttiioonn dduu rréésseeaauu dd‟‟hhyypphheess eexxttrraa--mmaattrriicciieellss..
--CCuulltt uurreess nnoonn mmyyccoorrhhiizzooggèènneess --DDiimmiinnuutt iioonn ddeess pprrooppaagguulleess..
--UUttiilliissaattiioonn ddeess ffoonnggiicciiddeess àà bbaassee ddee ccuuiivvrree --TTooxxiiqquuee ppoouurr lleess CCMMAA..
55.. LLaa mmyyccoorrhhiizzaattiioo nn ccoo nnttrrôôllééee

54
LLaa ssyyllvviiccuullttuurree eett ll‟‟aarrbboo rr iiccuullttuurree ffrruuiitt iièè rree ccoonnsstt iittuueenntt uunn ddoo mmaa iinnee dd‟‟aapppp lliiccaa tt iioonn
pprr iivviillééggiiéé ddeess ssyymmbb iioosseess ppoo uurr ll‟‟aamméé lliioo rraatt iioo nn ddee llaa pprroodd uucc tt iioo nn vvééggéé ttaa llee,, eenn rraa iissoo nn ddeess pprroojjee ttss
ddee rreebboo iisseemmeenntt ddaannss llee mmoonnddee,, dd uu ppeeuu ddee rreennttaabb iill iittéé ddeess aarrbb rreess àà lloonngg ttee rrmmee ccee qq uuii jjuusstt iiff iiee llaa
rreecchheerrcchhee ddee tteecchhnniiqquueess ééccoonnoo mmiissaanntt lleess ddééppeennsseess dd ‟‟éénneerrggiiee eett ddee mmaa tt iièè rreess pprreemmiièè rreess
ccooûûtteeuusseess ee tt ppoo lllluuaanntteess ((eennggrraa iiss,, ppeesstt iicc iiddeess )),, ssaacchhaanntt qquuee llaa ssyymmbb iioossee ppee rrmmee tt aauuxx aarrbb rreess ddee
mmiieeuuxx rréé ss iiss tteerr aauuxx mmaa llaadd iieess rraacc iinnaa iirreess eett ddee mmiieeuuxx eexxpp lloo iitteerr lleess rreessssoo uurrcceess dduu ssoo ll..
LLaa mmyyccoo rrhhiizzaa tt iioonn ccoo nnttrrôô llééee ee sstt uunn eennsseemmbb llee ddee tteecchhnniiqq uueess qquuii ccoo nnss iiss tteenntt àà iissoo lleerr,,
ccuulltt iivveerr,, sséé lleecc tt iioo nnnnee rr,, mmuulltt iipp lliieerr,, iinnccoorrppoorreerr ee tt ssuuiivvrree llee cchhaammpp iiggnnoonn aa ffiinn ddee pp rroodd uuiirree ddeess
ppllaannttss bb iioo llooggiiqquueemmeenntt aamméé lliioo rrééeess ppaarr ll‟‟oopp tt iimmiissaa tt iioonn ddee llaa ssyymmbb iioossee ((BBaa eett aall..,, 22000011))..
LLeess pp rraatt iiqq uueess ddee llaa mmyyccoorrhhiizzaatt iioonn ccoonnttrrôô llééee oonntt ééttéé ddééccrr iitteess ppaa rr pp lluuss iieeuurrss aauutteeuurrss ((BBaa eett
aall..,, 22000011 ;; EEcchhaa iirr ii eett aall..,, 22000088)) ;; cceess tteecchhnniiqq uueess dd iiffffèèrreenntt ppaarr llee cchhoo iixx ddee llaa pp llaannttee hhôô ttee eett ppaarr
lleess ccoonndd iitt iioonnss ddee ccuullttuurree :: ccuullttuurree eenn ppoott,, ccuullttuurree hhyyddrrooppoonniiqquuee oouu aaéérrooppoonniiqquuee,, eenn aaxxéénniiqquuee
oouu nnoonn..
EEnn pprraa tt iiqq uuee,, ll‟‟ iinnooccuullaa tt iioo nn ccoo nnss iiss ttee àà iinnccoorrppoorree rr aauu ssoo ll ddééss iinnffeeccttéé ddee ppéépp iinniièèrree
ll‟‟ iinnooccuulluumm ccoonntteennaanntt llee cchhaammpp iiggnnoonn mmyyccoorrhhiizziieenn ddee ssoonn cchhoo iixx..
55..11.. PPrroo dduucctt iioonn dd’’iinnooccuulluumm ddee CCMMAA
LLaa pp rroodduucctt iioonn dd ‟‟ iinnooccuulluumm ccoo nnss iiss ttee àà mmuulltt iipp lliiee rr llee cchhaammpp iiggnnoonn,, iissoo lléé àà ppaarr tt iirr dduu ssoo ll
ssoouuss ffoo rrmmee ddee ssppoo rreess,, eenn uutt iilliissaanntt uunnee pp llaannttee mmyyccoottrroopphhee ,, ttee llss qquuee llee ppoo iirreeaauu ((FFoorr tt iinn eett aall..,,
22000088)),, llee mmiill ((BBaa eett aall..,, 22000011)),, llee ttrrèè ffllee oouu PPlleeccttrraanntthhuuss aauussttrraalliiss ((BBoouuddaarrggaa eett DDeexxhhee iimmeerr,,
11998899 ;; EEcchhaa iirr ii eett aall..,, 22000088 )),, llaa ttoommaa ttee .. II ll ffaauutt éévviitteerr ttoo uuttee ffoorrmmee ddee ccoo nnttaammiinnaa tt iioonn ppaarr ddeess
aaggeennttss ppaatthhoo ggèènneess eett ttoo uutt rr iissqquuee ddee mméé llaannggee eennttrree lleess ssoo uucchheess,, lleess ccuullttuurreess ssee ffoo nntt ssuurr dd uu ssoo ll
ppaauuvvrree eenn pphhoosspp hhoorree pp rrééaa llaabb lleemmeenntt ddééss iinnffeecc ttéé..
PPoouurr mmiinn iimmiisseerr lleess vvaarr iiaatt iioonnss dduueess aauu ssoo ll,, iill eess tt pprréé fféérraabb llee dd‟‟uutt iilliisseerr ddee ll‟‟aa rrggiillee ccaa llcc iinnéé
oouu dd‟‟aauuttrreess ssuubbss ttrraa ttss àà bbaassee ddee ttoo uurrbbee,, ddee ppee rr lliittee ee tt ddee vveerrmmiiccuulliittee qquuii ffaacc iilliitteenntt llaa rrééccuuppéérraa tt iioonn
ddeess ssppoorreess,, ddeess ssppoorrooccaa rrppeess ee tt ddeess mmyyccoorrhhiizzeess ppaarr llee ttaammiissaaggee hhuummiiddee..
55..22.. SSyynntthhèèsseess mmyyccoorrhhiizziieennnneess
55..22..11.. SSyynntthhèèssee mmyyccoorrhhiizziieennnnee eenn ccoonnddiittiioo nnss ggnnoottooxxéénniiqquueess
LLaa ssyynntthhèèssee mmyyccoo rrhhiizziieennnnee eenn ccoo nndd iitt iioonnss ggnnoo ttooxxéénniiqq uueess eess tt rrééaa lliissééee eenn cchhaammbbrree ddee
ccuullttuurree cc lliimmaa tt iissééee,, eenn ssee rrrree oo uu aauu cchhaammpp ;; ee llllee uutt iilliissee ccoo mmmmee iinnooccuulluumm ddeess rraacc iinneess iinnffeecc ttééeess,,
dduu ssuubbsstt rraatt ccoonntteennaanntt ddeess ssppoorreess ee tt dd uu mmyyccéé lliiuumm qquuee ll‟‟oo nn pp llaaccee aauu ccoonnttaacctt ddeess rraacc iinneess ddee llaa
ppllaannttee hhôôttee.. ((BBaarreeaa,, 22000000 ;; FFoo rrtt iinn eett aall..,, 22000088)).. LL„„ iinnoo ccuulluumm ppeeuutt êê ttrree ccoonndd iitt iioonnnnéé ssoouuss

55
ddiifffféé rreenntteess ffoorrmmeess,, ppaarr llee mméé llaannggee ddeess mmyyccoorrhhiizzeess aavveecc ddee ll‟‟aa rrggiillee ccoonnffeecc tt iioo nnnnééeess eenn ppeett iittee
bboouulleetttteess,, eenn ssuussppeennss iioo nn ddaannss ll‟‟eeaauu oouu eennccaappssuullééss ddaannss ddeess bb iilllleess dd‟‟aa llggiinnaa tteess ppoouurr ffaacc iilliitteerr
lleeuurr iinnccoo rrppoorraa tt iioo nn aauu ssoo ll oouu lleeuurr pp llaacceemmeenntt aauu vvoo iiss iinnaaggee ddeess sseemmeenncceess..
CCeettttee tteecchhnniiqquuee aa éé ttéé uutt iilliissééee ddaannss llaa mmyyccoorrhhiizzaatt iioonn ddee pp lluuss iieeuurrss eesspp èècceess lliiggnneeuusseess eett
ffrruuiitt iièèrreess,, eenn rraa iissoonn ddeess aavvaannttaaggeess qquu‟‟oo ffffrree ll‟‟aassssoocc iiaatt iioo nn àà llaa pp llaannttee hhôô ttee,, ssuurr ttoouutt ppoouurr lleess
bboouuttuurreess sseemmii lliiggnneeuusseess,, eett lleess vviitt rroo--ppllaannttss,, ppoo uurr lleeuurr éévviitteerr llee cchhoocc ddeess tt rraannsspp llaannttaatt iioonnss
dduurraanntt llaa ppéérr iiooddee dd ‟‟aacccc lliimmaa ttaa tt iioonn.
LLaa mmyyccoorrhhiizzaattiioonn ccoonnttrrôôllééee ddee ll‟‟oolliivviieerr aa ééttéé uuttiilliissééee ppaarr ddee nnoommbbrreeuuxx cchheerrcchheeuurrss qquuii
oonntt mmyyccoorrhhiizzéé ddeess mmiiccrroobboouuttuurreess sseemmii lliiggnneeuusseess dd‟‟oolliivviieerr ddee llaa vvaarriiééttéé «« CCoorrnniiccaabbrraa »» iissssuueess
ddee nnéébbuulliissaattiioonn aavveecc ttrrooiiss eessppèècceess ddee GGlloommuuss ((GGll.. iinnttrraarraaddiicceess,, GGll.. mmoosssseeaaee eett GGll..
CCllaarrooiiddiiuumm)) ((PPoorrrraass SSoorrooaannoo eett aall..,, 22000022 ;; PPoorrrraass PPiieeddrraa eett aall..,, 22000055)),, eett cceelllleess ddeess vvaarriiééttééss
«« AArrbbeeqquuiinnaa eett LLeecccciinnoo »» aavveecc lleess ddeeuuxx pprreemmiièèrreess eessppèècceess ddee GGlloommuuss eett GGll.. VViissccoossuumm
((CCaallvveennttee eett aall..,, 22000044)).. SSaaaadd ((22000099)) aa aauussssii iinnooccuulléé ddeess bboouuttuurreess hheerrbbaaccééeess ddee llaa vvaarriiééttéé
«« SSiiggooiissee »» dd‟‟oolliivviieerr dd‟‟AAllggéérr iiee ppaarr ddeess eessppèècceess iinnddiiggèènneess ddee CCMMAA..
LLaa mmyyccoorrhhiizzaattiioonn ccoonnttrrôôllééee eesstt aapppplliiqquuééee àà dd‟‟aauuttrreess aarrbbrreess ffrruuiittiieerrss ccoommmmee llee jjuujjuubbiieerr
(( BBaa eett aall..,, 22000011)),, llee kkiiwwii (( CCaallvveett eett aall..,, 11998899)),, llee ffrraaiissiieerr eett llee ppoommmmiieerr (Cavallazzi et al.,
2007 ; Derkowska et al..,, 22000088)),, llee ppiissttaacchhiieerr (( GGrreeeenn eett aall..,, 22000055)) eett llee ppaallmmiieerr ddaattttiieerr ((DDrreeyyeerr
eett aall..,, 22000099))..
Les plants micropropagés peuvent être aussi mycorhizés durant la phase
d‟enracinement, au début de la phase d‟acclimatation ou après la phase d‟acclimatation mais
avant la période post-acclimatation en chambre de culture (Azcon-Aguilar et Barea, 1997;
Taylor et Harrier, 2001).. DDiivveerrss ttrraavvaauuxx oonntt mmoonnttrréé qquuee llaa nnuuttrriittiioonn mmiinnéérraallee eett llaa ccrrooiissssaannccee
aaiinnssii qquuee lleeuurr ssuurrvviiee ddeess ppllaannttss mmiiccrroopprrooppaaggééss ssoonntt aamméélliioorrééss ggrrââccee àà llaa mmyyccoorrhhiizzaattiioonn :: ccaass
ddeess vviittrrooppllaannttss dd‟‟aannaannaass iinnooccuullééss aavveecc GGlloommuuss sspp.. ((GGuuiilllleemmiinn eett GGiiaanniinnaazzzzii,, 11999944)),, ddee cceeuuxx
dd‟‟oolliivviieerr et de deux cultivars de bananier mycorhizés par Gl. intraradices et Gl. manihotis
((BBiinneett eett aall..,, 22000077))..
55..22..22.. SSyynntthhèèssee mmyyccoorrhhiizziieennnnee eenn ccoonnddiittiioonnss aaxxéénniiqquueess ((ccuullttuurree mmoonnooxxéénniiqquuee ))
OOnn qquuaalliiffiiee ddee ccuullttuurree mmoonnooxxéénniiqquuee,, uunnee ccuullttuurree ffoonnggiiqquuee aavveecc uunn oorrggaanniissmmee ééttrraannggeerr,,
llaa rraacciinnee ((SSttrruulllluu eett aall..,, 11999977)) ((ffiigg.. 2288))..

56
LLeess CCMMAA nnee ssee ccuullttiivveenntt ppaass oouu ssee ccuullttiivveenntt ddiiffffiicciilleemmeenntt eenn ccuullttuurree ppuurree,, iillss oonntt bbeessooiinn
dd‟‟uunn ppaarrtteennaaiirree vvééggééttaall ppoouurr aaccccoommpplliirr lleeuurr ccyyccllee vviittaall ((DDeecclleerrcckk eett aall..,, 22000055)) ;; eenn eeffffeett,, lleess
ssyymmbbiioosseess mmyyccoorrhhiizziieennnneess ssoonntt ééttaabblliieess eenn ssuuccccèèss,, eenn
mmiilliieeuu ggéélloosséé ((FFoorrttiinn eett aall..,, 22000022))..
FFiigg.. 2288 :: CCuullttuurree mmoo nnooxxéénniiqquuee ssuurr bbooîîtteess ddee PPeettrrii ((CCaannoo eett aall ..,, 22000088))..
LLaa mmyyccoorrhhiizzaattiioonn ccoonnttrrôôllééee eenn aaxxéénniiqquuee ss‟‟eeffffeeccttuuee ppaarr llaa sséélleeccttiioonn ddee ll‟‟iinnooccuulluumm qquuii
ppeeuutt êêttrree ccoonnssttiittuuéé ddee ssppoorreess ((BBoouuddaarrggaa eett DDeexxhheeiimmeerr,, 11998899 )),, dd‟‟ hhyypphheess oouu ddee ffoorrmmeess
iinnttrraarraacciinnaaiirreess,, lleess vvééssiiccuulleess (( SSttrruulllluu eett RRoommaanndd,, 11998866 eett 11998877 ;; DDiioopp eett aall..,, 11999944;; SSiimmoonneeaauu
eett aall..,, 11999944 ;; DDeecclleerrcckk eett aall.. ,, 11999966 ;; SSttrruulllluu eett aall..,, 11999977 ;; FFoorrttiinn eett aall..,, 22000022)).. LLeess vvééssiiccuulleess eett
lleess hhyypphheess iinnttrraa-- rraacciinnaaiirreess ssoonntt iissoollééss àà ppaarrttiirr ddeess mmyyccoorrhhiizzeess ppaarr ddeess ttrraaiitteemmeennttss
eennzzyymmaatt iiqquueess..
LL‟‟iinnooccuulluumm ddooiitt êêttrree ddééssiinnffeeccttéé,, ccaarr llaa ddééssiinnffeeccttiioonn eesstt ll‟‟ééttaappee llaa pplluuss iimmppoorrttaannttee ppoouurr llaa
rrééuussssiittee ddee llaa tteecchhnniiqquuee,, iill eenn eexxiissttee pplluussiieeuurrss pprrooccééddééss sseelloonn llee ttyyppee dd‟‟iinnooccuulluumm ((SSttrruulllluu eett
RRoommaanndd,, 11998866 ;; BBoouuddaarrggaa eett DDeexxhheeiimmeerr,, 11998899)),, llaa pplluuss uuttiilliissééee eesstt cceellllee ddee CCrraanneennbbrroouucckk eett
aall.. ((22000055)) ((TTaabblleeaauu 44))..
TTaabblleeaauu 44 :: MMéétthhooddeess ddee ddééssiinnffeecctt iioonn ddee ll‟‟iinnooccuulluumm ddeess CCMMAA ((CCrraanneennbbrroouucckk eett aall.. 22000055))..
TTyyppee dd’’eess ppèèccee IInnoocc uulluu mm DDééss iinnffeeccttiioonn
eenn uu ttii ll ii ss aanntt
PPrrééttrraaii ttee mmee nntt
11
PPrrééttrraaii ttee mmee nntt
22
EEtt aappee 11
cchhlloorraammii nn
ee TT
EEtt aappee 22
AAnnttii bbii oo--
ttii qquueess

57
CCeettttee tteecchhnniiqquuee ffoo uurrnniitt ééggaa lleemmeenntt uunnee eexxccee lllleennttee eennccee iinnttee ppoouurr llaa ccoonnsseerrvvaa tt iioonn ddeess
ssoouucchheess,, ppoo uurr ll‟‟ééttuuddee ddee lleeuurr ccyycc llee ddee vviiee ((SS ttrruulllluu eett aall..,, 11999977 ;; DDiioopp eett aall..,, 11999988)),, ddee
ll‟‟oonnttooggéénniiee ssppoo rraa llee aa iinnss ii qquuee ppoo uurr ll‟‟éévvaa lluuaatt iioonn ddeess ccoonndd iitt iioonnss ffaavvoorraabb lleess àà llaa ggeerrmmiinnaa tt iioonn ee tt àà
llaa ssppoorruullaatt iioonn ;; ee llllee eess tt ccoommppaatt iibb llee àà ll‟‟ iinnooccuullaatt iioonn ddeess pp llaannttss ddee lliiggnneeuuxx iissssuuss ddee
mmiiccrroopp rrooppaaggaatt iioo nn (DDeessjjaarrdd iinnss eett aall..,, 22000055))..
CCeess ccuullttuurreess ccoonnsstt iittuueenntt llee ss RROOCC ((rroooo tt oorrggaann ccuullttuurree )),, ee lllleess ppeerrmmeett tteenntt llaa pprroodduucc tt iioonn
dd‟‟ iinnooccuulluumm,, eenn ee ffffee tt,, oonn oobbtt iieenntt eenn qq uuee llqq uueess sseemmaa iinneess eenn bboo îîtteess ddee PPeettrr ii ddeess mmiill lliieerrss ddee ssppoo rreess
eexxeemmpptteess ddee ttoouuttee ccoo nnttaammiinnaa tt iioonn ((DDeecc lleerrcckk eett aall..,, 22000022 ))..
LLeess mmiilliieeuuxx ddee ccuullttuurree uutt iill iissééss ssoonntt llee mmiilliieeuu MMSS RR ((SS ttrruulllluu ee tt RRoommaanndd mmoodd iiffiiéé )) (( SSttrruulllluu eett
aall..,, 11999977)) oouu llee mmiilliieeuu CCrruusshh eett HHaayy ((11998811))..
LLaa mmyyccoorrhhiizzaatt iioonn ccoonnttrrôô llééee eenn ccoonndd iitt iioonnss aaxxéénniiqquueess aa ééttéé rrééaa lliissééee ssuurr ddeess pp llaannttss
mmiiccrroopp rrooppaaggééss ddee bbaannaanniiee rr ((DDeecc lleerrcckk eett aall..,, 11999955 ;; JJaa iizzmmee --VVeeggaa ee tt AAzzccoonn,, 11999955 ;; JJaa iizzmmee --
VVeeggaa eett aall..,, 22000022 eett 22000033)),, dd‟‟aannaannaass ,, ddee ppaappaayyee eett dd ‟‟aavvooccaatt ((JJaa iizzmmee --VVeeggaa eett AAzzccoonn,,11999911 eett
11999955)) eett dd‟‟oo lliivv iieerr ((BBiinneett eett aall..,, 22000077))..
LL‟‟ iinnooccuullaa tt iioonn ssee ffaa iitt aauu mmoo mmeenntt ddee llaa tt rraannsspp llaannttaatt iioo nn ddeess pp llaannttss (( ffiigg.. 2299)).. DDeess pp llaannttss ddee
bbaannaanniieerr mmiiccrroopprrooppaaggééss oonntt ééttéé iinnooccuullééss ppaarr ttrroo iiss eessppèècceess ddee GGlloomméérraaccééeess iissssuueess dd‟‟uunnee
ccuullttuurree rraacc iinnaa iirree ((RROOCC)),, eennccaappssuullééeess ddaannss ddeess bb iilllleess ddee CCaa--aa llggiinnaa ttee ((JJaa iizzmmee --VVeeggaa eett aall..,,
22000033)).. LL‟‟eennccaappssuullaatt iioonn ddeess CCMMAA ddaannss lleess bb iillllee ss dd‟‟aa llggiinnaa ttee oo ffffrree llaa ppoossss iibb iilliittéé ddee dd iivveerrss iiff iieerr
lleess pprroocceessssuuss dd ‟‟ iinnooccuullaatt iioonn,, iill ssee rraa iitt iinnttéérreessssaanntt dd‟‟ iinnccoorrppoorree rr ddaannss lleess ccaappssuullee ss ddeess ffllaavvoonnoo ïïddeess
oouu ddeess mmiicc rroooorrggaanniissmmeess ssyynneerrggéé tt iiqquueess..
OOnn ppeeuutt aauussss ii iinnooccuullee rr lleess rraacc iinneess dd‟‟uunn ssyymmbb iioottee vvééggéé ttaa ll ccoommmmee llaa ccaarroo ttttee ((DDaauuccuuss
ccaarrrroottaa LL..)) oouu llaa ttoommaattee ttrraannss ffoorrmmééee oouu nnoonn ppaa rr AAggrroobbaacctteerriiuumm rrhhiizzooggeenneess ((MMuuggnniiee rr eett
MMoossss,, 11998877))..
EEssppèè ccee ss
pprroo dduuii ssaanntt ddee ss
ssppoorree ss
GGiiggaass ppoorraaccééeess
GGlloo mméé rraaccééeess
EEssppèè ccee ss
pprroo dduuii ssaanntt ddee ss
ssppoorrooccaarr ppee ss
GGlloo mméé rraaccééeess
EEssppèè ccee ss aavvee cc
ii mmppoorrttaannttee ss
ii nnttee nnssii ttéé
mmyyccoorrhhii zzii eennnnee ss
GGlloo mméé rraaccééeess
AAccaauu lloossppoorraaccééeess
SS ppoorree
SS ppoorroocc aarr ppee
MMyycc oorrhhiizzee
AAppppaarreeii ll àà
ffii ll ttrraattii oonn
AAppppaarreeii ll àà
ffii ll ttrraattii oonn
BBaaiinnss
dd’’uull ttrr aass oonnss
AAuuccuu nn
UUll ttrraass oonnss ,,
11mmiinn
(( EEtthh aann ooll 9988%%
1100ss ))22
AAuuccuu nn
AAuuccuu nn
HHyyppoocchhll oorrii ttee
ddee CC aa 66%% ,,
22mmiinn
22--1100 mmiinn
22--1100 mmiinn
22--1100 mmiinn
22--1100 mmiinn
22--1100 mmiinn
22--1100 mmiinn

58
QQuueellqquueess eessppèècceess ppeerrddeenntt lleeuurr iinnffeeccttiivviittéé aapprrèèss pplluussiieeuurrss rréépplliiccaattiioonnss iinn vv iittrroo
((PPlleenncchheettttee eett aall..,, 11999966 ;; DDeecclleerrcckk eett aall..,, 22000055)) cc‟‟eesstt llaa rraaiissoonn ppoouurr llaaqquueellllee,, iill eesstt nnéécceessssaaiirree
dd‟‟éévvaalluueerr llee ppootteennttiieell ddee ll‟‟iinnooccuulluumm ddeess ddiifffféérreenntteess ggéénnéérraattiioonnss ddee CCMMAA eenn ccuullttuurree
mmoonnooxxéénniiqquuee ccoonnttiinnuuee.. PPoouurr pprréésseerrvveerr llaa bbiiooddiivveerrssiittéé ffoonnggiiqquuee ppeerrmmaanneennttee,, ddeess ccoolllleeccttiioonnss
ddooiivveenntt êêttrree mmaaiinntteennuueess iinn vv iittrroo eett iinn vv iivvoo..
FFiigg.. 2299 :: SScchhéé mmaa dduu pprrooccééddéé ddee ll’’iinnooccuullaattiioonn ddeess ppllaannttss mmiiccrroopprroo ppaaggééss
(DDeessjjaa rrddiinnss eett aall ..,, 22000055)).. AA :: IInnooccuullaattiioonn ddeess CCMMAA ssuurr rraacciinneess ddee ccaarroottttee ttrraannssffoorrmmééeess ((iinnooccuullaattiioonn pprr iimmaa iirree)) ;; BB ::
MMiiccrroopprrooppaaggaattiioonn ddeess ppllaannttss ;; CC :: TTrraannssffeerrtt ddeess ppllaannttss ssuurr mmiilliieeuu dd‟‟eennrraacciinneemmeenntt ;; DD :: IInnooccuullaatt iioonn ddeess
ppllaannttss ppaarr lleess rraacciinneess mmyyccoorrhhiizzééeess ((iinnooccuullaattiioonn sseeccoonnddaaiirree))..
55..44.. SSéé lleeccttiioonn ddeess CCMMAA
LL‟‟iinnooccuullaattiioonn ddee llaa ppllaannttee hhôôttee eesstt ssuuiivviiee ppaarr llaa sséélleeccttiioonn ddee ll‟‟eessppèèccee llaa pplluuss
ppeerrffoorrmmaannttee oorr,, lleess eessppèècceess mmoonnttrraanntt uunn ppootteennttiieell iimmppoorrttaanntt ppoouurr ll‟‟aabbssoorrppttiioonn ddeess nnuuttrriimmeennttss
Chambre de
culture
Incubation5 semaines à 27°C

59
ddooiivveenntt êêttrree sséélleeccttiioonnnnééeess aavvaanntt ll‟‟iinnooccuullaattiioonn ddeess ppllaannttss ((AAzzccoonn--AAgguuiillaarr eett BBaarreeaa,, 11999977;;
PPuutthhuurr eett aall..,, 11999988 )).. DDee nnoommbbrreeuusseess oobbsseerrvvaattiioonnss oonntt mmoonnttrréé qquuee lleess eessppèècceess ddee CCMMAA nnee
pprroodduuiisseenntt ppaass llee mmêêmmee eeffffeett ssuurr lleess aarrbbrreess dd‟‟ooùù llaa nnéécceessssiittéé ddee bbiieenn lleess sséélleeccttiioonnnneerr ((CCiitteerrnneessii
eett aall..,, 11999988 ;; BBaarreeaa eett aall..,, 11999999)) ppoouurr oobbtteenniirr uunn iissoollaatt eeffffiiccaaccee ssuurr pplluussiieeuurrss eessppèècceess ddee
ppllaanntteess lliiggnneeuusseess ((SSiieevveerrddiinngg ,,11998899)).. CCeeppeennddaanntt,, mmaallggrréé lleeuurr iimmppoorrttaannttee iinnffeeccttiivviittéé,, lleess CCMMAA
ppeeuuvveenntt ppoossssééddeerr ddeess ddeeggrrééss ddee ssppéécciiffiicciittéé ffoonnccttiioonnnneellllee vviiss--àà--vviiss dduu ggéénnoottyyppee ddee lleeuurrss
ppllaanntteess hhôôtteess ((VVeessttbbeerrgg eett CCaasssseellll,, 22000099))..
LLeess ccrriittèèrreess ddee sséélleeccttiioonn ddeess CCMMAA ssoonntt :: ll‟‟iinntteennssiittéé dd‟‟iinnffeeccttiioonn rraacciinnaaiirree ddee llaa ppllaannttee
hhôôttee,, llaa rraappiiddiittéé dd‟‟iinnffeeccttiioonn eett llee ddéévveellooppppeemmeenntt dduu rréésseeaauu eexxttrraammaattrriicciieell,, ll‟‟aamméélliioorraattiioonn ddee
llaa nnuuttrriittiioonn mmiinnéérraallee ddeess ppllaannttss qquuii ppeeuutt êêttrree eessttiimmééee ppaarr llee ccaallccuull dduu fflluuxx ddee pphhoosspphhoorree
ppéénnééttrraanntt lleess hhyypphheess.. PPaarr eexxeemmppllee,, ..AAccaauulloossppoorraa llaaeevv iiss eesstt uunn bboonn ccaannddiiddaatt ppoouurr ll‟‟iinnooccuullaattiioonn
ppaarrccee qquuee llee fflluuxx ddee pphhoosspphhaattee eesstt ddee 22..88 nnmmooll//mm22ss eett ssaa vviitteessssee ddee ccrrooiissssaannccee ddaannss llee ssooll eesstt
ddee 33..11 mmmm//jjoouurr.. IIll yy aa dd‟‟aauuttrreess ccrriittèèrreess ccoommmmee llaa ttoolléérraannccee aauuxx ffoorrtteess ccoonncceennttrraattiioonnss eenn
pphhoosspphhoorree qquuii ppeerrmmeetttteenntt aauuxx CCMMAA ddee vviivvrree ddaannss lleess ssoollss ffeerrttiilleess eenn ssyynneerrggiiee aavveecc dd‟‟aauuttrreess
mmiiccrroooorrggaanniissmmeess tteelllluurr iiqq uueess..
6. L’olivier
6.1. Situation de l’olivier dans le monde et en Algérie
L‟oléoculture est une des cultures la plus ancienne ; l‟olivier (Olea europea L.) est
cultivé depuis 4000 ans (Besnard et al., 2001). Il est originaire d‟Asie et sa culture s‟étend sur
le pourtour du Bassin méditerranéen (Blazquez, 1996), on le retrouve dans les décorations des
stèles funéraires et des pyramides dès la IVème dynastie (-4505 à -4475ans) (Breton et al.
,2006).
L‟olivier est rattaché aux paysages méditerranéens, il est cité dans les textes classiques
grecs, la Bible et le Coran, comme symbole de force, de longévité et de paix. Cet arbre est une
des plantes les plus cultivées, il occupe la 24éme place parmi les 35 espèces les plus répandues
dans le monde. Sa plantation est intéressante pour des raisons économiques et culturales, du
fait de l‟adaptation de l‟huile d‟olive et des olives de table dans plusieurs régimes ; le bois, en
revanche, a une importance non négligeable, il est apprécié spécialement dans les travaux de
construction et dans les œuvres artistiques et artisanales (Rhizopoulou, 2007). Le fruit n‟est
pas seulement utilisé dans la production de l‟huile ou dans les préparations culinaires mais il
est aussi utilisé dans la fabrication des savons, parfums et dans l‟éclairage.

60
La culture de l‟olivier est localisée préférentiellement en zones littorales et sub-littorales
du Bassin méditerranéen (95% à 98% du patrimoine oléicole) (Rugini et al., 2005) , dans les
régions où les conditions de croissance sont relativement difficiles (Zohary et Spiegel, 1975)
telles que la chaleur, la sécheresse et les basses températures et sur les sols peu reproductifs
secs et pierreux (Omrani- Sabbaghi et al., 2007). Cependant, l‟olivier est sensible au gel et à
l‟humidité, en revanche, il a une large distribution dans le monde, puisqu‟il est cultivé au
Nord et au Sud d‟Afrique (Égypte, Tunisie, Maroc et Algérie, Afrique du sud) dans l‟Est et le
Sud de l‟Asie (Liban, Israël Malaisie, Hawaï, Maui et même en Chine), (Rony et al., 2009),
en Europe (France, Espagne, Italie, Portugal) (Cordeiro et al., 2008), dans l‟est de l‟Australie,
dans le Nord et le Sud de l‟Amérique (Argentine, Brésil, Chili, Pérou, Mexique et Californie),
et dans la nouvelle Calédonie (Starr et al., 1999 ; Rhizopoulou, 2007).
L‟olivier constitue en Algérie, une des principales essences fruitières. Le verger oléicole
occupe 200 000 ha soit 45% de la surface arboricole nationale à peine 2,3% de la superficie
agricole utile totale et produit près de 20.103 tonnes d‟huile par an (Sahli, 2009).
Le potentiel oléicole est concentré dans les régions montagneuses (Sahli, 2009). La
Kabylie occupe à elle même 54.3% de la superficie oléicole nationale dans le centre Nord
(Istambouli, 1976 ; Mazliak, 1982 ; Hamlat, 1995), le reste est distribué dans l‟est (28,3 %) et
l‟Ouest (17 %) (Yakoub-Bougdal et al., 2007).
Le profil variétal est constitué essentiellement de deux variétés très répandues (les plus
dominantes) Chemlal et Sigoise. Cette faible diversité variétale contraste avec celle des
milieux de production. Il existe des variétés populaires « Aberkane », « Aidel » ,
« Bouchouk », « Agraraz », « Aime », « Azeradj », « Bouchouk », « Rugette »,
« Blanquette », « Hamma », et « Limli » (Sahli, 2009) très rustiques et très adaptées aux
conditions pédoclimatiques de leur milieu d‟implantation mais qui ne sont pas multipliées
(fig.30).
La Sous espèce Olea Laperrini (Batt. et Tr. In Benarar et Bouguedoura, 2003) se
rencontre à l‟état sauvage jusqu‟à 2700m d‟altitude dans les zones arides (de l‟Atlas marocain
au massif du Hoggar et du Tassili des Adjers en Algérie) où les conditions d‟humidité et de
température sont favorables.

61
N
Témouchent
Tlemcen
Aïn-
Oran
Mostaganem
Sidi bel-Abbès
Naâma
M
A
R
O
C
Mascara
Tiaret
Djerlfa
El-ByadhLaghouat
Relizane
Chlef Aïn-Defla
TipazaALGER
Blida
Médea
M‟Sila
Biskra
Batna
Khenchla
Mila
Sétif
Béjaïa JijelTizi-Ouzou
Boumerdès
Bouira
TissemssiltOum-el-Bouagui
AnnabaSkikda
Cons-tantine
B. Bou-Arriridj
El-Oued
Ghardaïa
Guelma
El-Tarf
Tebessa
T U
N I S
I E
Saïda
SigoiseBourichaSouidiLimliBlanquette de GalmaRougette de MitidjaHamra, RougetteChamlalGrosse de HammaFerkaniAzeradjlangue de MilianaChef-lieu de wilaya
Douk-AhrasM e
r M
é d
i t
e r r a n
é e
n n
e
Légende
Fig. 30 : Les différentes variétés d’olivier produites en Algérie (d’après Saad, 2009).
6.2. Ethymologie de l’olivier :
La première appellation de l‟olivier date du 13ème siècle avant notre aire en Grèce
(trouvé sur une table en argile) (Chadwick, 1958). D‟ailleurs le terme « olive » et les autres
noms communs, dérivent des termes grecs « elaa » et « elam » (Hoad, 1991), dont :
Ar : Zeitoun (olivier cultivé), زيتون et zeboudje (olivier sauvage) ; Berbère : Ezemour ; Fr :
Olivier ; Angl. : olive tree, Ital.: Olivo ; Latin: oliva, oleum, olivum ; vieux Scandinavie :
Olia ; Portugais : Oliveira, zambugeiro ; Türk: Zeytin ; Touaregs : aleo (olivier sauvage)
(Hoad, 1991 ; Liddell et Scott, 1991; Rhizopoulou, 2007).
6.3. Systématique et Biologie de l’olivier
6.3.1. Classification de l’olivier
La famille des Oléacées comprend 29 genres et près de 600 espèces, sous forme
d‟arbres et arbustes, entre autre des plantes ornementales, les lilas (Siringa), les troènes
(Ligustrum), les frênes (Fraximus), les jasmins (Jasmin) et l‟olivier (Olea) (Ozgenturk et al.,
2010) .

62
L‟Olea europea est la seule espèce d‟olivier portant des fruits comestibles, elle inclue
l‟olivier sous sa forme cultivée Olea europaea sativa et sous sa forme sauvage ou l‟oléastre
Olea europaea sylvestris, à partir desquels sont issues 35 espèces, 6 sous-espèces (Heywood,
1978 ; de Casas et al., 2006 ; Breton et al. ,2006) : Olea europaea subsp. europaea (Europe),
Olea europaea subsp. Guanchica (Iles de Canarie), Olea europaea subsp. Cerasiformis
(Madeira archipelago), Olea europaea subsp. maroccana (Maroc), Olea europaea subsp.
laperrinei (montagnes du Sahara) et Olea europaea subsp. cuspidata (du Sud au Nord-est
Africain et du Sud-ouest de l‟Asie jusqu‟en Chine) ; et plus de 2500 cultivars (Rugini et al.,
2005 ; Cordeiro et al., 2008 ; Ozgenturk et al., 2010) .
Selon les systématiques moléculaires de Claros et al. (2000), Rugini et Baldoni (2004),
Cardoso et al. (2005) et de Strikic et al. (2010), la classification de l‟olivier est la suivante :
Règne : Plantae
Sous règne : Tracheobionates
Division : Magnoliophytes
Embranchement : Spermaphytes (Phanérogames)
Sous-embranchement : Angiospermes
Classe : Dicotylédones (ou Thérébinthales)
Sous-classe : Astéridées (ou Gamopétales)
Ordre : Lamiales
Famille : Oléacées.
Genre : Olea
espèce : europea
6.3.2. Description de l’arbre
L‟olivier est un arbre rustique, il se distingue des autres arbres fruitiers par sa grande
longévité, il conserve ses caractéristiques des milliers d‟années, toutes les espèces du genre
Olea ont un nombre chromosomique de base 2n =46 (x = 23) (Rugini et al., 2005).
Arbrisseau ou arbre de 2 à 10 mètres, il peut même atteindre 15 à 2m de diamètre, son
hauteur dépend du type de climat ; dans les climats froids ils sont généralement petits.
Généralement très rameux d'un blanc grisâtre, les rameaux sont spinescents (couvert
d‟épines) à l'état sauvage, les feuilles sont persistantes et disposées de façon opposée sur les
rameaux, oblongues ou ovales lancéolées, entières, coriaces, glabres, d'un vert cendré en
dessus, blanches-soyeuses en dessous, atténuées en court pétiole, à nervure médiane seule
saillante (Loussert et Brousse, 1978).

63
Les fleurs sont blanchâtres, regroupées en petites grappes axillaires dressées, calice en
coupe, à 4 dents très courtes.
La drupe charnue, ellipsoïde ou arrondie, verte, à la fin noire, à noyau osseux renfermant 1 à 2
graines, suivant les variétés (Loussert et Brousse, 1978).
6. 3.3. Description de la variété Sigoise d’olivier
La Sigoise (olive du Tell ou olive de Tlemcen) est originaire de la plaine de Sig, elle
occupe 25% du verger oléicole algérien, elle est utilisée pour la production de l‟olive de table
et de l‟huile avec un rendement moyen de 18 à 22% (Mendil et Sebri, 2006).
Fig. 31 : Aspect d’un olivier de la variété Sigoise d’un verger de Djdiouia (W. de Relizane).
C‟est un arbre à vigueur moyenne (fig.31), les feuilles sont longues, à largeur moyenne,
elliptiques lancéolées (fig.32), peu d‟inflorescences de longueur moyenne. Le fruit est
ovoïde, légèrement asymétrique, à sommet pointu, et à base tronquée de couleur noire en
pleine maturation, l‟endocarpe est elliptique à sommet pointu avec mucron à son extrémité et
à base arrondie (Mendil et Sebri, 2006).

64
Fig. 32 : Aspect des fruits et des feuilles d’olivier de la variété Sigoise de la région de
Djdiouia (W. de Relizane).
6.3.4. Voies de propagations de l’olivier
Pour conserver les caractéristiques organoleptiques de l‟olive et de l‟huile, l‟olivier est
multiplié par voie végétative, en utilisant la propagation par l‟ovule (sphaéroblaste) ou le
souquet, le bouturage ou le greffage (Rugini, 2002 ; Fabbri et al., 2004 ). Le microbouturage
permet de multiplier des espèces difficiles à reproduire naturellement, avoir des vitroplants
facilement transportables sans risques sanitaires et reboiser très rapidement avec une
diminution du coût de production (Benderradji et al., 2007).
La micropropagation des cultivars de l‟olivier est aussi utilisée avec succès (Rugini,
1984; Sayhan et Ozzambak, 1994 ; Rugini et al.,1999 ; Rugini et al., 2005 ; Binet et al., 2007,
Brhadda et al., 2008) pour sélectionner des espèces résistantes aux stress biotique et
abiotique (Sasanelli et al.,2000 ; Bartolozzi et al., 2001) dans l‟amélioration génétique
(Rugini et al., 1999 ; Rugini et Pesce, 2006).
En Algérie, la propagation de la variété Sigoise d‟olivier s‟effectue généralement par
voie végétative (ex : propagation par l‟ovule) ou par greffage (Fantanazzo et Baldoni, 1999) ;
cependant ces deux modes de multiplication ne sont pas très pratiques pour l‟obtention de
plantations intensives d‟olivier (Abdul Hussain et Abdul Hussain, 2004) ; pour y remédier on
utilise récemment le micro bouturage (Benderradji et al., 2007).
6.4. Les maladies de l’olivier
Fruits
Feuilles

65
L‟olivier est sensible à des attaques parasitaires, fongiques et bactériennes (Rugini,
2002 ; Phillips et al., 2005 ; Vidović et Petanović, 2008) (Tableau 5) et à certains virus
(Pasquini et al., 2002) susceptibles de causer de sérieux dommages économiques concernant
le rendement de sa production ; en revanche, les conditions météorologiques tels que le froid
(gel, neige…) , l‟humidité et la sécheresse sont considérés comme ennemis N°1 de l‟olivier
(Loussert et Brousse, 1978).
Tableau 5: Les différentes maladies de l‟olivier

66
Agent causal La maladie Les symptômes Références
Cyclconium oleaginum
Œil de paon
-Défoliation, chute des
fruits, il affecte
considérablement la
récolte.
Coulomb et al.
(2004) ;
Guechi et Girre
(2002).
Spilocea oleagina
Œil de paon
-Parasite des feuilles,
des olives et de leurs
pédoncules, il
provoque la
défoliation.
Régis (2007)
Verticillium dahliae
Verticilliose
-Dépérissement lent ou
aigu de l‟arbre,
défoliation, peut
provoquer la mort de
l‟arbre
Brickel et Zuk,
(1997);
Bellahcene
(2004) et
Bellahcene et
al. (2005a et b)
Bactrocera oleae
Mouche de l’olive.
-Chute prématurée du
fruit, altération de la
pulpe de l‟olive, il
abime la qualité de
l‟huile.
Avensenq et
al.(2005)
Prays oleae Teigne de l’olivier
-Chute prématurée du
fruit.
Avensenq et al.
(2005)
Saissetia oleae
Cochenille noire de
l’olivier
-Défoliation et perte de
récolte significative
Avensenq et al.,
2005
Dacus oleae Gmel.
Mouche de l’olivier
-Chute prématurée du
fruit, perte de poids, il
abime la qualité de
l‟huile.
Gaouar-
Benyelles
(1996)

67
Pseudomonas syringae
pv.savastanoi Smith.
Tuberculose
Développement de
tumeurs sur les organes
de la plante, cause la
mort des branches et
des bourgeons
Gaouar-
Benyelles
(1996)
1. Présentation des stations d’étude L‟étude sur l‟olivier Olea europea L.var. Sigoise, a été menée dans deux stations
oléicoles, dont l‟une est située en zone semi aride, dans la région de Sig (Wilaya de
Mascara) et l‟autre en zone aride, dans la région de Béchar.
1.1. Localisation des stations d’étude
1.1.1. Localisation de la station de Sig
La ville de Sig (ex wilaya d'Oran en 1975) se situe à l'ouest de l‟Algérie (0°11′29″W
35°31′44,2″N), à 50 km au sud-est d'Oran (fig.33) au débouché de la vallée de la
Mekerra. Elle regroupe plus de 65 800 habitants (2008) (regroupant les daïras de Sig et
d‟Oggaz). Son ancien nom français est Saint-Denis-du Sig [1].
Dans la région oléicole de Sig, l‟étude a été effectuée dans deux parcelles d‟oliviers.
-Parcelle 1 : située à 8 Km de la route n°4, à une altitude de 38m (fig.34), dans laquelle
les oliviers sont âgés de 5 ans (fig.35) et de 8 ans.
Fig.33 : situation géographique de la ville de Sig[2]
Sig
0
N
80 Km

68
-Parcelle 2 : située à 2 Km au nord de la Daïra de Sig, à une altitude de 49m (fig.34),
comporte des oliviers âgés de 17 ans (fig.36) et d‟autres centenaires (fig.37).
[1] http://fr.wikipedia.org/wiki/Sig_(Alg%C3%A9rie) [2]. www.Google earth.fr
Fig. 34 : Localisation des deux parcelles d’oliviers (A, B) dans la station de Sig
[3].
2 Km0
Bou Henni
SIGDouar el
Mkhalif
Ouled Nair
Douar ez
Zm ela
Koubbet
Sid i Lakhdar
MezraetBenkadda
Belkheir
DjebelBou Setta
Hamm arHaouadh
el Kelkh
W5N4
N4
N6
W30W21
W41
W5Douar’Atba
Djemmâla
Oggaz
O u
e
d
S
i g
N
Stations d’étude A et B.
AB

69
Fig. 35 : Olivier âgé de 5 ans
[3].http://maps.google.fr//
Fig.36 : Olivier âgé de 17 ans
Fig. 37 : Aspect d’oliviers centenaires. a : Verger d’oliviers de la variété Sigoise âgé de 100
ans; b : Morphologie d’un olivier du verger
1.1.2. Localisation de la Station de Béchar
a b

70
La wilaya de Béchar se situe au sud –ouest du territoire national ( 31°37′N 2°13′E),
elle est limitée à l‟est par la Wilaya d‟Adrar, à l‟ouest par le Maroc (600 Km de la
frontière marocaine), au nord par les Wilayates de Naama et d‟El Bayadh , au sud-ouest
par Adrar et au sud-est par Tindouf (fig.38). Elle occupe une superficie de161.400 Km2,
soit 6,77% du territoire national et compte une population de 272.436 habitants
équivalente à une densité moyenne de 169/ Km². Béchar est une région agricole où les
vergers d‟oliviers regroupent 3200 arbres [4].
Fig. 39: Localisation de la station
d’étude (A) par rapport à la ville de
Béchar [2].
N
Fig.38 : Situation géographique de la
Wilaya de Béchar [4].
N
A
0 50

71
La station oléicole se localise à environ 15 Km du sud de la ville de Béchar (fig.39).
1.2. Caractéristiques climatiques des deux stations d’étude
Les données climatiques proviennent de l‟Unité Météorologique Régionale
d‟Oran (UMRO).
La station de Sig est située dans la plaine de la Habra de Sig ; elle se trouve dans l‟étage
bioclimatique semi-aride, dû à l‟écran de la péninsule ibérique et du Rif qui la met en
position d‟abri pluviométrique (Bouchetata, 2006).
[4].http://fr.wikipedia.org/wiki/Béchar.
1.2.1. La température
1.2.1.1. Températures annuelles
Les données thermiques des stations d‟étude s‟étalant de 1999 jusqu‟à 2009 (fig. 40).
Dans la station de Sig, l‟année 1999 constitue l‟année la plus chaude (21.3°C), l‟année
qui suit (2000) s‟avère la plus froide (14.8°C). Quant à la station de Béchar, les années les
plus tempérées (1999 et 2002) ont des températures moyennes respectivement de 22.3 et
22.8 ° C, pour les autres années la température moyenne annuelle est de 20.9-21.9°C.
1.2.1.2. Températures mensuelles
Les deux stations d‟étude sont caractérisées par des températures moyennes
extrêmement élevées lors de la période estivale et des températures assez clémentes
durant la période hivernale. La température moyenne des mois les plus chauds (juillet et
Août) pour la station de Sig est de 28° C alors que celle du mois le plus froid (décembre)
est de 7.9° C (Fig.41). Pour la station de Béchar, les températures moyennes des mois
(juillet et Août) les plus chauds sont respectivement 35.4 et 34.5° C, alors que celles des
mois les plus froids (décembre et janvier) sont respectivement 8.8 et 10.3°C.

72
Fig. 40 : Répartition des températures annuelles moyennes des deux stations d’étude
(Unité météorologique régionale d’Oran).
Fig.41 : Températures moyennes annuelles (1999-2009) des deux stations d’étude
(Unité météorologique régionale d’Oran).
0
5
10
15
20
25
30
1999 2000 2001 2002 2003 2004 2005 2006 2007 2008 2009
T(°C)
Années
Températures annuelles moyennes
Sig
Béchar
0
5
10
15
20
25
30
35
40
T(°C)
Mois
Températures mensuelles moyennes de
l'année 2009
Sig
Béchar

73
1.2.2. La pluviométrie
1.2.2.1. Pluies annuelles
Les données pluviométriques des stations d‟étude s‟étalant de 1999 jusqu‟à 2009
(Fig.42). Pour la station de Sig, l‟année 2007 constitue l‟année la plus pluvieuse (419
mm), précédée des années 2003 et 2004 (385 mm). L‟année 2001 est considérée comme
la plus sèche avec des précipitations de l‟ordre de 196 mm. En revanche, la station de
Béchar est caractérisée par un climat plutôt sec, les années relativement pluvieuses sont
2008 et 2009 (241 mm), et l‟année la plus sèche est l‟année 2001, où la pluviométrie
moyenne annuelle atteint 13 mm.
1.2.2.2. Pluies mensuelles
Pour la station de Sig, le mois le plus pluvieux est celui de décembre, avec une
moyenne de l‟ordre de 82 mm suivi par le mois de novembre avec une pluviométrie
moyenne de 64 mm (Fig. 43). Le mois le plus sec est celui d‟août avec une précipitation
mensuelle moyenne de 2mm. Pour celle de Béchar, le mois le plus pluvieux est celui
d‟octobre caractérisé d‟une pluviométrie mensuelle moyenne de 153mm suivi des mois
de février et de septembre avec des pluviométries moyennes de 23 et 21 mm
respectivement. Le reste de l‟année est relativement sec, caractérisé par des précipitations
moyennes maximales de 10 mm et minimales de 0 à 3mm.

74
Fig.42 : Répartition des pluies annuelles des deux stations d’étude
(Unité météorologique régionale d’Oran).
0
50
100
150
200
250
300
350
400
450
1999 2000 2001 2002 2003 2004 2005 2006 2007 2008 2009
Pré
cip
itati
on
(m
m)
Années
Répartition des pluies annuelles
Sig
Béchar
0
20
40
60
80
100
120
140
160
180
Pré
cip
ita
tio
n (m
m)
Mois
Répartition des pluies mensuelles
Sig
Béchar

75
Fig. 43: Histogramme des pluies mensuelles moyennes des deux stations d’étude
(Unité météorologique régionale d’Oran).
Les données des précipitations mensuelles (Fig.43) montrent que dans la station
de Sig, la période la plus humide se situe entre les mois d‟octobre jusqu‟en juin, cette
station est caractérisée par une longue période de sécheresse estivale qui s‟étale du mois
de juillet jusqu‟à septembre. En revanche, la station de Béchar se caractérise par une
sécheresse qui dure presque toute l‟année, les précipitations se situent pendant les mois
de février, septembre et octobre.
2. Caractérisation des MA indigènes des sols des oliveraies de Sig et de Béchar
2.1. Prélèvement des échantillons de sol et des racines
Les échantillons de sol rhizosphérique et des racines d‟olivier ont été collectés
dans les deux parcelles (1 et 2) de Sig en février 2009 et dans la station de Béchar au
mois de mai de la même année. Les échantillons ont été prélevés au niveau de trois points différents au pied de 15 arbres
de chaque parcelle choisis aléatoirement.
La collecte de sol est effectuée à des profondeurs variant de 10 à 20cm (Calvente et
al., 2004), ensuite mélangés pour obtenir un échantillon représentatif. Après une semaine
de séchage à l‟air libre, le sol recueilli est passé au tamis (à mailles 5mm) pour éliminer
les cailloux et les gros débris de matière organique puis sur tamis à mailles 2mm.
Les racines secondaires très fines blanchâtres et légèrement gonflées, fraichement
récoltées, sont nettoyées à l‟eau de robinet puis conservées dans du FAA (Annexe 1).
2.2. Analyse physico-chimique du sol Les échantillons de sol rhizosphérique de l‟olivier ont été analysés au Laboratoire
Régional des analyses du sol (INSID) d‟El Matmar (Wilaya de Relizane) (Saad, 2009).
2.3. Mise en évidence de l’infection endomycorhizienne Les mycorhizes naturelles de l‟olivier ont été observées au microscope photonique
après leur coloration par la méthode de Phillips et Hayman (1970) et l‟estimation de
l‟infection mycorhizienne des racines est réalisée selon la méthode de Trouvelot et al.
(1986).

76
2.3.1. Coloration de Phillips et Hayman (1970)
Des fragments racinaires de 1cm de longueur sont soigneusement lavés à l‟eau de
robinet pour éliminer les traces de FAA et immergés dans une solution de KOH à 10%
pendant 45min à 90°C ; cette étape permet de vider le contenu cellulaire des racines. Ils
sont ensuite soigneusement lavés à l‟eau distillée, blanchis et neutralisés dans de l‟acide
lactique à 10% puis colorés avec le bleu de trypan à 0.05% ou la fuscine acide à 0.1%
(annexe 1) pendant 1h à 90°C. Ces colorants réagissent avec la chitine, composante
essentielle de la paroi fongique.
2.3.2. Technique d’évaluation de l’infection endomycorhizienne
Après la coloration, 50 fragments d‟environ 1 cm prélevés au hasard, sont montés
sur entre lame et lamelle, à raison de 10 fragments par lame, dans une goute de
lactoglycérol (V/V) puis examinés au microscope photonique.
Cette technique calcule 4 paramètres d‟infection mycorhizienne arbusculaire. Le calcul
de ces 4 paramètres d‟infection est réalisée en utilisant les formules de calcul définies par
(Trouvelot et al., 1986):
1. La fréquence de mycorhization (F) qui reflète le degré d‟infection du système
racinaire.
F(%) = 100 (N-n0) /N
N : nombre de fragment observés.
n0 : nombre de fragment sans trace de mycorhization.
2. L’intensité de mycorhization (M : intensité de mycorhization relative et m :
intensité de mycorhization absolue) qui exprime la portion du cortex colonisée par
rapport à l‟ensemble du système racinaire.
m(%) = (95n5 + 70n4 + 30n3 + 5 n2 +n1)/ (N-n0)
M(%) = (95n5 + 70n4 + 30n3 + 5 n2 +n1)/ N
n5, n4, n3, n2 et n1sont les nombres de fragments respectivement notés dans les cinq
classes d‟infection marquant l‟importance de la mycorhization à savoir :
5= plus de 95%, 4= de 50à 95%, 3= 30à 50%, 2= 1à 30% et 1= 1% du cortex (fig.44).

77
3. La teneur en arbuscules (a) de la partie mycorhizée.
a%= (100 mA3 + 50 mA2+ 10mA1)/ 100
mA3, mA2 et mA1 sont les pourcentages de mycorhization de qualité
arbusculaire données, calculés suivant le modèle :
mA1= (95n5Ai + 70n4 Ai+ 30n3 Ai+ 5 n2 Ai+n1 Ai) F/M (N-n0)
n5Ai, n4 Ai, n3 Ai, n2 Ai et n1 Ai sont respectivement les nombres de fragments notés
5Ai, 4Ai, 3Ai, 2Ai et 1Ai (fig. 45).
4. la teneur en arbuscules (A) du système radiculaire qui reflète les potentialités
d‟échange symbiotique de l‟association, son efficacité.
A(%) = a M/100
2.3.3. Prospection des espèces de CMA indigènes aux sols des oliveraies
- Extraction des spores à partir des sols rhizosphériques de l’olivier :
L‟extraction des spores et des sporocarpes de CMA est effectuée par la technique de
tamisage humide de Gerdemann et Nicolson (1963) suivie d‟une centrifugation sur
gradient de saccharose selon la méthode décrite par Giovanetti et al. (1991).
A partir de chaque échantillon de sol, 100g sont prélevés et mis dans 1000 ml d‟eau. La
suspension de terre est ensuite vigoureusement agitée puis filtrée dans une série de tamis
Fig. 44: les classes d’infection (0à5) d’un fragment
racinaire endomycorhizé
Fig.45 : Estimation de l’abondance des arbuscules dans un fragment
racinaire mycorhizé. A1 : quelques arbuscules ; A2 : arbuscules
fréquents ; A3 : Arbuscules abondants.

78
à mailles décroissantes (500, 350, 250, 80 et 40µm) sous un jet d‟eau. Les tamisâts
contenant les spores des CMA sont mélangés et mis en suspension dans de l‟eau distillée.
La suspension sporale est centrifugée une première fois à 2000rpm (tours par
minute) pendant 5 min pour réduire les particules de sol et les fragments racinaires, le
surnageant est ensuite filtré sur un tamis de 40µm. Le dernier tamisât est centrifugé de
nouveau sur un gradient de saccharose à (20 /60%) (p : v) ( Sturmer et al., 1998) afin
d‟obtenir un grand nombre des spores, le surnageant est filtré sur un tamis de 40µm et le
tamisât est lavé pour éliminer l‟excès de saccharose. Les spores et les sporocarpes sont
recueillis dans des boîtes de Petri , examinés sous la loupe stéréoscopique puis récoltés à
l‟aide d‟une micropipette et montés entre lame et lamelle dans une goutte de polyvinyl-
lactoglycérol (PVLG) (Koske and Testier, 1983) ou dans une goutte de PVLG
additionnée du réactif de Melzer(V :V) (Morton, 1991) (Annexe 1).
Les lames ainsi préparées sont laissées pendant 4 à 5 min à la température
ambiante, avant de les recouvrir par une lamelle puis pendant une nuit pour la
polymérisation des produits de montage des préparations.
Les préparations sont ensuite examinées au microscope photonique après une légère
pression exercée sur la lamelle pour briser la paroi sporale et observer ses différentes
couches.
Les spores et les sporocarpes sont caractérisés sur la base de leurs critères
morphologiques :
- la dimension des spores, leur couleur et ornementation, l‟épaisseur de la
paroi sporale et le nombre de couches.
- et le diamètre des hyphes suspenseurs, l‟épaisseur de leur paroi et leur
mode d‟attachement à la spore.
Nous avons essayé d‟identifier les spores en se basant sur les travaux de Sturmer
et Morton (1997 et 1999), BBllaasszzkkoowwsskk ii ((22000033)),, WWaallkkeerr eett SS hhuubb lleerr ((22000044)),, SS iieevveerrdd iinngg ee tt
OOeehhll ((22000066 )),, SSppaa iinn eett aall..(( 22000066 )),, Walker et al. ((2007b), Jeffries et al. (2007), Sharma et
al. (2008), Blaszkowski et Czerniawska (2008).
La couleur des couches de la paroi sporale est déterminée d‟après la carte de
couleur INVAM des spores (INVAM color chart for spores) indiquée en Annexe
5((BBllaasszzkkoowwsskk ii,, 22000033))

79
NB: le réactif de Melzer est un réactif chimique à base d‟iode qui permet de révéler la
présence d‟amidon (coloration bleu) et de sucres de type dextrose (coloration brun roux).
Il est utilisé pour mettre en évidence la coloration dextrinoïde de certaines parois des
spores des espèces de Glomeromycota.
2.3.4. Étude du pouvoir endomycorhizogène du sol (PEM)
Selon Plenchette et al. ( 1989), le potentiel infectieux mycorhizogène (PEM) d‟un
sol représente sa capacité à initier la formation d‟une association mycorhizienne à partir
d‟une quantité d‟inoculum présente dans ce sol sous forme de spores, de mycélium et des
débris racinaires portant des vésicules.
Le test biologique est basé sur une relation de type dose (quantité de sol non
stérilisé)-réponse (statut mycorhizien des plates tests) et le potentiel mycorhizien des
échantillons du sol est mesuré par le calcul du nombre le plus probable de propagules
(NPP) mycorhizogènes viables et capables d‟infecter des racines d‟une plante
mycotrophe (dite plante test) par Kg de sol.
2.3.4.1. Préparation des dilutions et repiquage des plantules de poireau
La technique consiste à réaliser cinq séries de dilution de sol allant de 10-1 jusqu‟à
10-5 Ainsi, 20g de sol non désinfecté sont ajoutés à 180g de sol désinfecté (autoclavage
à 120°C pendant 1h ) (Fortas, 1990) pour obtenir la dilution 10-1 puis 20g de cette
dilution sont ajoutés à 180g de sol désinfecté pour obtenir la dilution 10-2 ensuite on
procède de la même manière pour les autres dilutions 10-3, 10-4 et 10-5 (Tableau 6).
Les différentes dilutions de sol ainsi préparées (200g) sont mises dans des pots de 250ml.
Tableau 6 : Obtention des dilutions de sol.
Sol
non
stérile
10-0 10-1 10-2 10-3 10-4 Sol
non
stérile
Quantité
de sol à
ajouter(g)
200 20 20 20 20 20 0
Quantité
de sol
stérile(g)
0 180 180 180 180 180 200
Dilution 10-0 10-1 10-2 10-3 10-4 10-5 Témoin
stérile.

80
2.3.4.2. Production de plantes tests
Les essais sont réalisés sur une plante-hôte mycotrophe, herbacée, bisannuelle de la
famille des Alliacées qui est le poireau (Allium porrum L.).
Les graines de poireau sont d‟abord immergées dans de l‟eau distillée pendant
15min , désinfectées dans une solution d‟hypochlorite de sodium à 10% (préparée à partir
d‟hypochlorite de sodium à 12 ° chlorométriques) additionnée de 300µl de Tween 20
pendant 15min avec agitation puis rincées soigneusement trois fois à l‟eau distillé stérile.
Après désinfection, dix graines sont mises à germer directement dans chaque pot de
culture.
2.3.4.3. Conditions de culture :
Les cultures de poireau sont placées dans une serre non climatisée, à température
ambiante et arrosées 2 fois par semaine à l‟eau de robinet.
2.3.4.4. Méthode de calcul du NPP :
La durée de la culture du poireau était de 16 semaines pour le sol provenant de la
parcelle 1 et 20 semaines pour la parcelle 2. Le système racinaire de chaque plant est
prélevé et traité par la coloration de Phillips et Hayman (1970) ; chaque système
racinaire présentant un point d‟infection est considéré mycorhizé et on lui attribue la
valeur (1). La valeur (0) est attribuée à ceux exempts de toute infection mycorhizienne.
Par ailleurs, on dénombre pour chaque dilution le nombre de systèmes racinaires
infectés et on détermine la dernière dilution (P1) qui représente celle où le nombre de
plants mycorhizés est supérieur à 5, P2 et P3 désignent le nombre de plants infectés dans
les deux dilutions successives.
Le nombre le plus probable (NPP) est calculé par la méthode d‟Alexander (1965)
suivant la table de Cochrane (1950) puis exprimé par Kg de sol. Les valeurs sont
obtenues dans un intervalle de confiance de 95% (NPP÷3.30< NPP<NPP× 3.30).
Lorsque NPP>1500, le PEM est élevé, il reflète un bon état biologique du sol, il est jugé
acceptable lors que NPP= 1500 et faible lors que MPP<1500.

81
3. Effet des CMA indigènes du sol rhizosphérique de la station oléicole de Sig sur la
croissance des plants de poireau.
Le poireau (Allium porrum) est utilisé pour piéger les complexes mycorhiziens
contenus dans les échantillons de sol provenant des deux parcelles (1 et 2) de la station
de Sig.
3.1. Technique de piégeage avec le poireau:
Les graines de poireau désinfectées comme précédemment, sont semées dans des
pots de 250 ml contenant 150g de sol rhizosphér ique de l‟olivier non désinfecté
provenant des parcelles (1 et 2) de Sig. Après leur germination (une semaine), les
cultures sont mises dans une serre non climatisée à température ambiante et arrosées
régulièrement une fois par semaine.
Le dispositif comprend quatre niveaux d‟inoculation :
- Niveau 1 : les graines de poireau sont semées dans le sol rhizosphérique
désinfecté de la parcelle 1.
- Niveau 2 : les graines de poireau sont semées dans le sol rhizosphérique
désinfecté de la parcelle 2.
- Niveau 3 : les graines de poireau sont semées sur le sol rhizosphérique non
désinfecté de la parcelle 1.
- Niveau 4 : les graines de poireau sont semées sur le sol rhizosphérique non
désinfecté de la parcelle 2.
Dans chaque niveau, nous avons effectué 50 répétitions. Après chaque semaine de
culture, environ 15 plants sont prélevés pour suivre l‟état de mycorhization (estimation
des paramètres d‟infection) des systèmes racinaires et cela durant dix semaines.
3.2. Mesure des paramètres de croissance des plants de poireau
La réponse des plants de poireau à la mycorhization est estimée par la biomasse
des parties aériennes, exprimée en matière sèche (MS) après leur séchage à l‟étuve à
105°C pendant 24 heures (Oihabi et Meddich, 1996).
L‟effet de la mycorhization est estimé par le degré de dépendance mycorhizienne
correspondant à la nécessité d‟une plante à porter une mycorhize pour pouvoir atteindre
une croissance maximale ou fournir un rendement donné en fonction d‟un certain niveau
de fertilité du sol (Jaizme-Vega et al., 2002), ce paramètre est calculé selon la formule :

82
DM = [(PS plante M - PS plante NM)/ PS plante NM] × 100
D‟où PS plante M et PS plante NM représentent respectivement le poids sec des parties
aériennes des plantes mycorhizés et non mycorhizés.
4. Effet des CMA indigènes du sol des oliveraies sur la croissance de l’olivier
La mise en évidence de l‟efficacité de l‟infection, par les champignons
endomycorhiziens à arbuscules et vésicules des sols, nécessite l‟utilisation de souches de
champignons endomycorhiziens connues pour être infectives et performantes. Mais par
manque de ces souches fongiques, nous avons entrepris notre essai avec un inoculum
contenant un mélange de champignons endomycorhiziens indigènes des sols étudiés.
4-1: Matériel végétal: Obtention des boutures herbacées d’olivier var. Sigoise.
Nous avons utilisé des boutures herbacées de la variété Sigoise d‟olivier
enracinées, âgées de deux mois. Leur phase de multiplication s‟est déroulée dans une
serre à nébulisation (Fig. 46) appartenant à la ferme expérimentale d‟El Matmer (Wilaya
de Relizane).
Fig.46: Disposition des boutures herbacées d’olivier sur les tables de multiplication
(Saad, 2009).
4-2: Matériel fongique
L‟inoculum fongique utilisé est constitué d‟un mélange de fragments de racines de
poireau, infectés par les espèces endomycorhizogènes indigènes du sol des oliveraies et

83
d‟un mélange de substrat rhizosphérique provenant de culture de poireau en pots et
contenant des spores et du mycélium.
4.2.1. Obtention des plantules de poireau
Les graines de poireau sont désinfectées puis mises à germer dans des pots
contenant 200g de sol préalablement désinfecté (à l‟autoclave à 120° C, pendant 1 h).
Une dizaine de jours après la mise en germination des graines, les jeunes plantules de
poireau atteignent environ 4 cm de hauteur.
4.2.2. Production d’inoculum
Nous avons effectué quatre séries d‟expérience :
- La première série (nommée CMS : complexe mycorhizien de la station de
Sig) consiste à semer des graines de poireau désinfectées dans des pots contenant du sol
non désinfecté provenant de la station des Sig.
- La seconde série (nommée CMB : complexe mycorhizien de la station de
Béchar) consiste à semer des graines de poireau désinfectées dans des pots contenant du
sol non désinfecté provenant de la station d e Béchar.
- La troisième et (nommée I1) consiste à inoculer les plantules de poireau par des
spores de CMA morphologiquement semblables isolées à partir du sol rhizosphérique de
l‟olivier de la station de Sig. Une suspension sporale dans de l‟eau distillée stérile est
mise directement au contact du système racinaire des plantules; ces spores constituent
l‟isolat n° 1.
- La quatrième (nommée I2) consiste à inoculer les plantules de poireau par des
spores de CMA morphologiquement semblables isolées à partir du sol rhizosphérique de
l‟olivier de la station de Béchar ; ces spores constituent l‟isolat n° 2.
Les pots de chaque série d‟expérience sont placés dans une serre non climatisée et
les jeunes plantules sont arrosées périodiquement à l‟eau de robinet.
Après trois mois de culture, les plants de poireau inoculés sont soigneusement
déterrés de leurs substrats, leurs racines sont délicatement et abondamment rincées à l‟eau
distillée stérile. A partir de chaque série d‟expérience, des plants de poireau sont choisis
au hasard et partagés en 2 lots. Un lot de racines est traité et coloré selon la méthode modifiée de Phillips et
Hayman (1970) et les racines sont examinées au microscope photonique afin de contrôler
la présence de l‟infection des champignons endomycorhizogènes indigènes.
Les racines du second lot de plants de poireau mycorhizés, issues de chaque série
d‟expérience, sont découpées en petits fragments de 0,5 à 1mm de longueur. Ces
fragments constituent l‟inoculum qui sera utilisé pour inoculer d‟autres plants de poireau.

84
Les fragments de racines issus de ces derniers plants âgés de quatre mois constitueront
l‟inoculum pour la mycorhization des bouture herbacées d‟olivier.
4.3. Technique d’inoculation des boutures herbacées d’oliviers
L‟inoculation des boutures herbacées d‟olivier a été initiée après l‟enracinement,
nous avons utilisé 50 boutures herbacées de la variété Sigoise enracinées et âgées
d‟environ 2 mois (Fig. 47).
Fig. 47: Boutures herbacées enracinées de la variété Sigoise d’olivier, âgées de 2
mois.
Les plants d‟oliviers ont été transférés séparément dans des pots de 400 ml
remplis à mi-hauteur de substrat stérile composé d‟un mélange de sable et de tourbe
blonde (V/V) préalablement désinfecté . L es plants sont ensuite répartis en quatre
groupes:
- le premier groupe: les plants sont inoculés avec l‟inoculum provenant de l‟expérience
de la série 1;
- le deuxième groupe: les plants sont inoculés avec l‟inoculum provenant de
l‟expérience de la série 2;
- le troisième groupe est inoculé avec l‟inoculum provenant de l‟expérience de la série
3.
- le quatrième groupe est inoculé avec l‟inoculum provenant de l‟expérience de la série
4. L‟inoculation des boutures herbacées consiste à mettre à proximité de chaque
système racinaire environ 50 g d‟inoculum utilisé sous forme de sol renfermant à la fois
des propagules de champignons endomycorhizogènes indigènes et des racines d‟oignon
infectées. Après l‟inoculation, le reste du substrat de culture désinfecté est ajouté jusqu‟à

85
ce que les pots en soient remplis. Les boutures de chaque série sont placées séparément
dans des plateaux (fig.48). Les plants sont élevés en serre non climatisée et sont
régulièrement arrosés avec l‟eau de robinet.
4.4. Mise en évidence de l’infection après inoculation des boutures
Après trois mois de culture, l‟infection et la colonisation des racines par les
champignons endomycorhizogènes indigènes est décelée au microscope photonique sur
des segments de racines traités et colorés selon la méthode modifiée de Phillips et
Hayman (1970).
4.5. Mesure des paramètres de croissance
Pour mettre en évidence les effets de la mycorhization, la croissance des plants
est évaluée par la hauteur de la partie aérienne, le nombre de tiges et de feuilles formées
par plante, la longueur des racines ainsi que les poids frais et sec de la partie aérienne des
plants.
Analyse statistiques
Les résultats des mesures de la croissance des plants sont traitées par des analyses de
variance(ANOVA) et des comparaisons de moyennes à l‟aide du logiciel STATISTICA
(6.0), les différences entres les traitements sont confirmées par le test de Newman-Keuls
au seuil de 5% (p<0.05)(Guissou, 2009).
4.6. Évaluation du degré de dépendance mycorhizienne des plants d’olivier
L‟effet de la mycorhization est aussi estimé par le degré de dépendance mycorhizienne
calculé à partir des matières sèches des parties aériennes des plants d‟olivier.
DM = [(PS plante M - PS plante NM)/ PS plante NM] × 100
Fig. 48 : Boutures herbacées d’olivier inoculés.

86
D‟où PS plante M et PS plante NM représentent respectivement le poids sec des parties
aériennes des plantes mycorhizés et non mycorhizés.
4.7. Ré-isolement et identification des spores à partir du substrats de culture en pot
Ce test de ré- isolement des spores permet de contrôler la présence de propagules
viables dans les substrats de culture des boutures d‟olivier inoculées. La présence des
espèces fongiques endomycorhiziennes à arbuscules est confirmée après leur isolement
par les techniques précédemment décrites (Brundrett et al., 1996) et leur observation
microscopique.
5. Culture axénique mycorhizienne des CMA indigènes de la rhizosphère de
l’olivier.
5.1. Isolement de spores
L‟isolement des espèces de CMA est effectué à partir de la culture de poireau
utilisée pour l‟étude du PEM (dilution 10-1) et à partir du sol rhizosphérique d‟olivier de
la station de Béchar.
Les spores sont isolées par tamisage humide (tamis 500, 250 à 50µm) et récupérées
selon les techniques décrites précédemment.
5.2. Production d’inoculum
Les spores ayant le même morphotype généralement en forme de grappes sont
prélevées sous la loupe stéréoscopique et mises à proximité du système racinaire des
plants d’Allium porrum, âgés de 15 jours et cultivés sur du sol désinfecté.
Après 12 semaines de culture en serre, des racines sont prélevées, lavées et des
fragments racinaires de 1 à 2mm de longueur servent d‟inoculum pour une seconde
culture monospécifique du poireau permettant d‟obtenir un nombre plus important de
spores.
Les plants de poireau inoculés par chaque isolat sont placés séparément dans des
plateaux et élevés en serre non climatisée. Après 12 semaines de culture, les plants sont
déterrés et leur système racinaire ainsi que le sol sont utilisés pour initier une infection
mycorhizienne en conditions axéniques.
5.3. Désinfection des spores et des racines mycorhizées
5.3.1. Désinfection des spores
Les spores recueillies sous la loupe stéréoscopique, après tamisage humide
du sol, sont désinfectées pendant 20 min avec la solution « A » composée
d‟hypochlorite de sodium à 7% (à partir d‟une solution à 12° chlorométriques) ,
de 300µl de Tween 20 et de 40 mg de streptomycine. Après filtration sur
membrane millipore (0.45µm) suivie de plusieurs rinçages à l‟eau distillée stérile,

87
le filtre est aseptiquement retiré à l‟aide d‟une pince stérile et les spores retenues
sur le filtre sont mises en suspension dans 2 ml d‟eau distillée stérile.
Cette suspension sporale est soit étalée sur de l‟eau gélosée à 1% (Annexe 5) ou
utilisée pour inoculer des racines secondaires des plantules de tomate cultivées sur
le milieu d‟association (annexe 5).
5.3.2. Désinfection des mycorhizes
Les racines mycorhizées de poireau prélevées sont lavées avec l‟eau pour
éliminer les particules de sol, désinfectées par immersion dans une solution « A
selon la méthode de Diop (2003), soigneusement rincées avec de l‟eau distillée
stérile et déposées aseptiquement sur de l‟eau gélosé à 1 % pour obtenir le
développement d‟un mycélium aseptique.
5.4. Production des plantules de tomates
Désinfection et mise en germination des graines de tomate
Nous avons utilisé la tomate (Solanum lycopersicum L. , famille des Solanacées),
espèce herbacée qui présente un système racinaire très dense et ramifié.
Les graines de tomate sont désinfectées comme celles du poireau et mises à germer dans
des boîtes de Petri contenant de l‟eau gélosée (1%) à raison de 7 à 8 graines par boîte
(Bouderga et Dexheimer, 1989).
5.5. Reconstitution de l’association en conditions contrôlées
L‟association est réalisée en boîtes de Petri de 90 mm de diamètre, sur
milieu Crush et Hay (1981 in Bouderga et Dexheimer, 1989) (annexe 5) ou sur
milieu MSR (Strullu et al., 1997), ces milieux sont appropriés pour la croissance
des racines car ils leur fournissent les sels minéraux essentiels à leur
développement. Leur composition est indiquée en annexe 5
Le système racinaire de tomate (prélevé et découpé aseptiquement) (fig.
49B) et l‟inoculum (fig. 49A) (racines mycorhizées en culture sur eau gélosé,
spores germées ou mises directement sans germination préalable) sont transférés
sur le milieu de symbiose.
L‟inoculum est placé à proximité des racines secondaires émises par le pivot des
jeunes plantes (fig. 49 C).
Les boîtes de Pétri sont fermées par un ruban adhésif et disposées en
position inclinée à 30° sous une rampe lumineuse réglée en jour de 12h, le suivi
de l‟expérimentation se fait quotidiennement sous la loupe stéréoscopique à
travers le couvercle des boîtes de Petri maintenues fermées.

88
1.
Co
mposition physico-chimique du sol des oliveraies de la station de Sig
Les résultats des analyses physico-chimiques du sol des oliviers des deux parcelles 1
et 2 de la station de Sig sont mentionnés dans le tableau 7 (Saad, 2009).
Le sol a une structure grumeleuse ; il est sablo- limoneux avec une large prédominance
de sable par rapport aux autres éléments tels que les limons et l‟argile. Sa teneur en eau est de
18,3 à 21,8% ; son pH est alcalin ( 8,03< pH <8,43). C‟est un sol non salé (0,704< Ce< 1,266
ms/c ) moyennement pauvre en calcaire (calcaire total 17,83< taux < 18,25% et calcaire actif (
5,37 <CaCO3<5,56% ), en azote total (0,09 à 0,14%) , en phosphore (0.008 à 0.02%) et en
matière organique (1,69 et 2,00%).
Les caractéristiques du sol des deux parcelles indiquent que les sols des parcelles1 et 2
sont favorables à la prolifération des CMA et à l‟établissement de la MA avec la variété
Sigoise d‟olivier. Des caractéristiques analogues des sols à CMA ont été rapportés par divers
auteurs (Djili et al. 1999 ; Le Tacon et al., 1999 ;Giri et al., 2005 ; Cortoba et al., 2001 ;
Cantrell et Linderman, 2001 ; Cavalazzi et al., 2007).
Tableau 7 : Analyses physico-chimiques du sol des deux parcelles de la station de Sig (Saad, 2009).
Caractéristiques Parcelle 1 Parcelle 2
Structure Grumeleuse Grumeleuse
Perméabilité Suffisante Bonne
Sables grossiers (%) 4% 5%
Sables fins(%) 50% 60%
Limons (%) 30 – 35% 30- 32%
A C B
Fig.49 : Technique de la culture monoxénique à partir d’une spore germée
(Cranenbrouck et al. 2005).
A : Extraction du fragment de gélose contenant la spore germée ; B : Prélèvement du
système racinaire de la plante hôte ; C : Association de l’inoculum à la racine sur le
milieu de symbiose.

89
Argile (%) 9 - 11% 5-3%
Poids total humide (g) 961 954
Poids total sec (g) 862 860
Poids de la tare (g)
407
347
Poids de l‟eau (g)
99
94
Poids du sol sec (g)
455
513
Teneur en eau (%) 21.8 18.3
pH 8,43 8,03
Conductivité électrique
(Ce) (l/g ms/cm)
0,704 1,266
Calcaire total% 18,25 17,83
Calcaire actif % 5,56 5,37
Matière organique % 2,00 1,69
Azote total ‰ 0,962
(0,09%)
1,40
(0,14%)
P2O5 assimilable ppm 83,3
(0,008%)
224,99
(0,02%)
2. Mise en évidence de l’infection endomycorhizienne chez l’olivier
2.1. Estimation des paramètres d’infection mycorhizienne .
Les examens microscopiques effectués sur des racines d‟oliviers de la région de Sig
(âgés de 5, 17 et 100 ans) prélevées en hiver et sur de jeunes oliviers de la région de Béchar
(âgés d‟environ 5 ans) prélevées au printemps de la même année, ont montré que la fréquence
de mycorhization des arbres est importante quels que soit l‟âge des arbres, les saisons de
prélèvement et les régions prospectées (aride ou semi aride) (fig. 50).
L‟olivier s‟est montré hautement mycotrophique, les pourcentages d‟infection sont
importants pendant les deux saisons de prélèvement, le taux d‟infection est supérieur à 80% ,
il est de 82% pour les oliviers âgés de 5 ans et de 100% pour les autres oliviers indiquant ainsi
la richesse du sol en propagules viables et son bon état biologique. Ces résultats rejoignent

90
ceux de Wubet et al. (2003), d‟Azcón-Aguilar et al. (2003), de Caravaca et al.(2005) et de
Saad (2009).
L‟intensité de mycorhization est plus élevée (70.5%) chez les jeunes oliviers de Béchar
que celle des jeunes oliviers de Sig. Le taux d‟arbuscules est par contre très faible, il est de 12
% chez les jeunes oliviers et de 33,67 % chez les oliviers âgés de 15 ans, ceci est
probablement dû à la courte durée de vie des arbuscules (une à trois semaines) qui dégénèrent
ensuite pour être remplacés par des hyphes et des vésicules qui persistent jusqu‟à la mort de
la cellule. Nos résultats sont analogues à ceux de Saad (2009) et selon l‟estimation de
Kormanick et McGraw (1982), on peut déduire que la mycorhize de la variété Sigoise
d‟olivier correspondrait à la classe 5 (Wubet et al. 2003).
Fig. 50: Estimation des paramètres d’infection mycorhizienne de l’olivier. F : fréquence de mycorhization ; M et m : intensité de mycorhization relative et absolue respectivement, du cortex colonisée par rapport à l‟ensemble du système racinaire ; A: teneur en arbuscules du
système radiculaire et (a) de la partie mycorhizée
2.2. Mise en évidence des structures de l’association endomycorhizienne
Les examens microscopiques des racines de tous les oliviers révèlent la présence des
mycorhizes arbusculaires. Des résultats analogues ont été rapportés pa r d‟autres auteurs
(Porras Soriano et al., 2002; Wubet et al., 2003 ; Porras Piedra et al., 2005 ; Calvente et al.,
2004 ; de Rougement, 2007 ; Saad, 2009). Ce type de mycorhize est commun dans les
systèmes de culture naturels ( Roldan-Fajardo et Barea, 1986).
0
20
40
60
80
100
120
F % m % M % a % A %
(%)
5 ans
17ans
100ans
Béchar

91
Les différentes structures que nous avons observées sont : les hyphes extra-racinaires,
les appressoriums, les hyphes intra-racinaires, les enroulements, les arbuscules, les vésicules
et les spores intra-racinaires.
2.2.1. Les hyphes extramatriciels
Les hyphes extra-racinaires (fig.51) mesurent environ 8 à 9,6 µm de diamètre et plus
d‟une centaine de µm de longueur et portent parfois des spores.
2.2.2. Les appressoriums
Les CMA pénètrent dans les cellules corticales de l‟hôte à l‟aide d‟appressoriums ou
par les poils absorbants (fig. 52), comme le décrit Ligrone et al. (2007).
Nous avons constaté que la morphologie des appressoriums est variable (fig. 53) ;
certains sont lobés tandis que d‟autres portent des ramifications, le champignon forme
généralement une boucle dans la première cellule épithéliale qu‟il pénètre puis progresse entre
les cellules adjacentes pour coloniser le cortex racinaire grâces aux hyphes intra-racinaires.
2.2.3. Les hyphes intra-racinaires
Les hyphes intra-racinaires sont coenocytiques, d‟aspect et de diamètre différents, fins
(mesurant 3 à 5 µm) ou épais (8 à 12 µm). Ces résultats rejoignent ceux de Zubek et al.
(2008) qui ont mentionné que le mycélium intra-racinaire de la majorité des plantes
prospectées a 2 µm de diamètre ; il est de 20µm selon Plenchette (2005) et de 5–10 µm
selon Giovannetti et al. (2010).
Nous avons observé trois types de mycélium bien distincts:
Type 1 : le mycélium de diamètre moyen (6 à 7 µm) qui se colore mal avec la
coloration au bleu de trypan et à la fuschine acide. Il est difficilement discernable au
microscope photonique, par contre, on peut observer la présence de gouttelettes lipidiques
dans les hyphes lorsqu‟ils envahissent les cellules racinaires et prennent la forme
irrégulière d‟un « H » ; on observe des enroulements en forme de boucles (fig. 54) et des

92
vésicules de forme irrégulière. Selon Dodd et al. (2000), le mycélium présentant cette
description appartiendrait au genre Acaulospora.
Type 2. Le mycélium est composé d‟hyphes droits qui s‟étendent entre les cellules en
formant un « Y », et des liaisons en « H » qui se colorent intensément ; ce type de mycélium
est présent dans tous les systèmes racinaires (Fig. 55). Selon Dodd et al. (2000) et
Blascowski, (2003), il correspondrait au mycélium du genre Glomus.
Type 3. Mycélium constitué d‟hyphes épais (8 à 9 µm) qui se colorent intensément à
la fuschine acide et au bleu de trypan et progressent en boucles à l‟intérieur de la racine ; ce
type est caractérisé par l‟absence de vésicules. D‟après Morton (2000), Dodd et al. (2000) et
Blascowski (2003), ce mycélium caractérise les espèces appartenant aux Gigasporacées
(genres Gigaspora et Scutellospora ).
2.2.4. Les enroulements
Peu d‟enroulements sont formés par les complexes mycorhiziens des oliveraies
prospectées, ils sont localisés dans les couches corticales externes des systèmes racinaires, généralement liés aux points de pénétration, ils sont intracellulaires (fig. 56a, b, et c).
La présence des pelotons mycéliens a été détectée dans le système racinaire des oliviers
âgés de 15 ans, ils sont intracellulaires (fig. 57, a et b).
2.2.5. Les vésicules
Les vésicules ont une forme soit ovalaire (fig. 58, a), rectangulaires (fig. 58, b) ou
parfois sphériques (fig. 58, c) et intracellulaires (fig.58, d).
Il est connu que les vésicules se forment seulement chez les genres appartenant aux
Acaulosporacées, Gloméracées et Pacisporacées (Zubek et al., 2008).
La présence importante de vésicules ovalaires et allongées dans les sols des régions
semi arides indique la prédominance d‟espèces appartenant surtout au genre Glomus ; celles
d‟Acaulospora sont irrégulières et lobées et généralement peu colorées avec le bleu de
trypan. La dominance des Gloméracées dans les zones aride et semi aride a été rapportée par
de nombreux travaux (Sieverding, 1991 ; de Souza et al., 1999; Bever et al.,2001; Franke-
Snyder et al., 2001; Lovelock et al., 2003).
2.2.6. Les arbuscules
Les arbuscules (fig. 59) sont généralement plus petits que les vésicules (23 ×12.8 µm),
ils sont éphémères (une à trois semaines).

93
Nous avons observé dans le système racinaire des oliviers âgés de 15 ans, le type 3 qui
est caractérisée par des hyphes épais (8 à 9 µm), qui se colorent intensément avec le bleu de
trypan, les arbuscules sont composés d‟un tronc gonflé et de ramifications fines, les vésicules
sont absentes (fig. 60) ; ce morphotype correspond aux espèces appartenant aux
Gigasporacées (Dodd et al., 2000 ; Blaszcowski, 2003).
Nous avons observé quasiment dans tous les systèmes racinaires d‟oliviers examinés
le type 2 caractérisé par des arbuscules à tronc cylindrique (fig. 61) et des hyphes
intercellulaires plus au moins épais (6 à 8 µm), intensément colorés avec la coloration de
Phillips et Hayman, (1970), les vésicules sont présentes ; ce morphotype est dominant il
correspond aux espèces appartenant aux Gloméracées. (Dodd et al., 2000 ; Blaszcowski,
2003).
Nos observations microscopiques des racines des oliveraies prospectées révèlent que
la mycorhize arbusculaire formée par les CMA indigènes des sols est de type
« Arum » caractérisé ppaarr ll‟‟eexxtteennss iioonn iinnttee rrccee lllluullaa iirree dd uu cchhaammpp iiggnnoo nn ddaannss llaa rraacc iinnee,, lleess hhyypphheess
ffoo rrmmeenntt ddeess ttrroonnccss llaa ttéérraauuxx qquuii ppéénnèèttrreenntt àà ll‟‟ iinnttéé rr iieeuurr ddeess ccee lllluulleess ccoo rrtt iiccaa llee ss eett ddéévvee llooppppeenntt
ddeess rraammiiff iiccaatt iioonnss dd iicchhoottoommiiqquueess ttrrèèss ffiinneess ffoorrmmaanntt ddeess aarrbb uussccuulleess tteerrmmiinnaauuxx (( ffiigg.. 6622)), les
enroulements se localisent généralement près des points de pénétrations.
Nos résultats rejoignent ceux de nombreux auteurs qui ont rapporté que les Oléacées
forment des MA de type « Arum » (Smith et Smith ,1997 ; Wubet et al. 2003 ; Azcon-
Aguillar et al. 2003 ; Ahulu et al. 2005 ; Dickson et al. 2007). Ils sont également présents
chez d‟autres ligneux (Yamato 2004 et 2005 ; Muthukumar et Prakach, 2009) et chez
d‟autres familles de plantes telles que les Astéracées, les Campanulacées, les Fabacées, les
Lamiacées et les Rosacées (Zubek et al., 2008). Ce type morphologique de MA serait contrôlé
par la plante hôte au niveau des espèces, c'est-à-dire que la morphologie de la MA est
influencée par l‟identité de la plante (Smith et Smith, 1997).
2.2.7. Les spores intra-racinaires
Les spores intra-racinaires sont observées dans le système racinaire des oliviers âgés de
15 ans et de ceux de Béchar (fig. 63). La plupart de ces spores sont très petites (≤25µm)
globulaires ou sphériques, en grappe de 10 à 15 spores, attachées à un hyphe suspenseur de 2
à 4 µm de diamètre ; ces spores appartiennent au genre Glomus . Des résultats analogues ont
été rapportés par Tao et Zhiwei (2005) et Wubet et al., (2003). Selon ces auteurs, quelques

94
espèces de Gloméracées comme Gl. intraradices, Gl. microaggregatum et Gl. clarum,
adaptées à un environnement chaud et aride, forment des spores intra-racinaires (Tao et
Zhiwei , 2005). Gl. tenui forme aussi des spores intra-racinaires très petites (diamètre≤
10µm) sous forme de renflements intra ou intercellulaires généralement appelés endophytes
fins ; elles sont communes dans les climats tempérés et absentes dans les climats tropicaux
(Dodd et al., 2000).
Nous avons aussi retrouvé dans le système racinaire des oliviers provenant de Sig un
mycélium cloisonné (fig. 64) mais à très faible fréquence (≤5%), il s‟agit de champignons
appartenant aux EMS.
LLaa pprréésseennccee ddeess eennddoopphhyytteess àà mmyyccéélliiuumm sseeppttéé ((EEMMSS)) ccoolloonniissaanntt ssiimmuullttaannéémmeenntt lleess
ééccoossyyssttèèmmeess aavveecc lleess CCMMAA aa ééttéé ccoonnssttaattééee ssuurrttoouutt ddaannss lleess hhaabbiittaattss aallppiinnss eett aarrccttiiqquueess
((HHaasseellwwaannddtteerr eett RReeaadd,, 11998800 ;; RReeaadd eett HHaasseellwwaannddtteerr,, 11998811 ;; JJuummppoonneenn,, 22000011)).. EEllllee aa aauussssii
ééttéé ddéétteeccttééee ddaannss lleess ssyyssttèèmmeess rraacciinnaaiirreess ddee nnoommbbrreeuusseess ppllaanntteess mmyyccoorrhhiizzééeess tteellss qquuee
LLeeuuccaanntthheemmuumm wwaallddsstteeiinniiii ((AAsstteerraaccééeess)),, CCaammppaannuullaa ppoollyymmoorrpphhaa ((CCaammppaannuullaaccééeess)) eett
FFeessttuuccaa vveerrssiiccoolloorr ((PPooaaccééeess)) eett ddaannss cceeuuxx ddeess ppllaanntteess nnoonn mmyyccoorrhhiizzooggèènneess aappppaarrtteennaanntt aauuxx
BBrraassssiiccaaccééeess,, CCaarryyoopphhyyllllaaccééeess eett SSccrrooffuullaarriiaaccééeess ((ZZuubbeekk eett aall..,, 22000088)) ;; sseelloonn cceess aauutteeuurrss,, iillss
ssoonntt iimmppoorrttaannttss ppoouurr llaa ccrrooiissssaannccee ddee cceess ppllaanntteess.. CCeess eennddoopphhyytteess ssoonntt ccoonnssiiddéérrééss ccoommmmee
mmyyccoorrhhiizziieennss àà nnee ppaass nnéégglliiggeerr ddaannss lleess ééttuuddeess ééccoollooggiiqquueess ((JJuummppoonneenn,, 22000011))..
3. Diversité des CMA du sol rhizosphérique de l’olivier
La méthode par tamisage humide du sol rhizosphérique d‟olivier nous a permis d‟isoler
diverses spores qui se différencient par leurs caractéristiques morphologiques et leur
dimension (diamètre généralement ≤80µm parfois de 40 et 500 µm).
La dominance des spores de petite taille dans les sols rhizosphériques a été rapportée
par de nombreux auteurs (Mangan et al., 2004 ;Tao et Zhiwei, 2005 ; Abbas et al., 2006) ; elle
est probablement liée à leur adaptation sélective au stress hydrique (Brundrett et al., 1999 ;
Boddington et Dodd, 2000).
3.1 Diversité des CMA du sol rhizosphérique de l’olivier de Sig
Nous avons constaté la prédominance des spores globulaires ou ovoïdes reliées ou non à
un hyphe suspenseur, volumineuses (≤200 µm de diamètre) qui caractérisent le genre
Glomus (fig.65A et B).

95
D‟autres spores sont aussi présentes, sessiles, sphériques de couleur jaune doré à orange
(fig. 66, F) parfois sombres (figs. 66 D et J), le saccule sporifère est souvent absent en raison
de son détachement lors du tamisage humide ; la morphologie de ces spores est
caractéristique d‟Acaulospora.
Ces résultats sont en accord avec ceux de Abbas et al. (2006) puisque les spores
d‟Acaulospora sont lisses (fig.66A, B ; I) ou ornementées (fig.66 C et E), sessiles (fig.66 A à
H) (sans pédicelle) et leur position est latérale sur l‟hyphe du saccule sporifère. Les spores
juvéniles ont une paroi formée d‟une seule couche qui est continue avec la paroi de l‟hyphe
du saccule et qui se différencie ensuite en une couche laminaire constituée de trois couches
dont la troisième couche est difficile à distinguer. La spore mure développe une à deux
parois germinatives flexibles ou semi flexibles parfois ornementées entre lesquelles se
présente l‟orb de germination (fig.66 A) d‟où sort le tube germinatif.
Nous avons constaté la présence de spores à l‟intérieur d‟autres spores plus
volumineuses (fig.67 A et B). Selon Gerdemann et Nicolson (1963), ces spores mortes
appartenant au type 6 sont parfois présentes chez les Glomus.
3.2. Diversité des CMA du sol rhizosphérique de l’olivier de Béchar.
Les observations sous la loupe stéréoscopique des spores du sol rhizosphérique de
l‟olivier de Béchar ont révélé la prédominance des Gloméracées et la présence de quelques
espèces appartenant aux genres Acaulospora et de Scutellospora.
La dominance des Gloméracées dans les sols rhizosphériques des régions arides et
semi-arides a été rapportée par certains auteurs (Azcon-Aguilar et al. ,2003 ; Wang et
al.,2008). Les espèces les plus dominantes dans le sol rhizosphérique de l‟olivier en Espagne
sont : Glomus coronatum, Gl. constrictum, Entrophospora sp. et Scutellospora sp.
Les spores peuvent former un sporocarpe (fig. 68). La couleur des spores peut être
transparente ou sombre mais la plupart sont jaunes dorés, marrons ou oranges.
SP

96
Nous avons remarqué que la densité des spores isolées du sol rhizosphérique de
l‟olivier en zone aride est plus importante que celle en zone semi aride. Selon certains auteurs
la densité des spores de CMA augmente dans les climats secs (Uhlmann et al., 2006) ; elle est
liée à leur adaptation aux écosystèmes chauds secs et arides (Mangan et al., 2004 ; Tao et
Zhiwei, 2005) .
3.2.1. Les morphotypes d’Acaulospora
Quatre morphotypes appartenant au genre Acaulospora ont été isolés. Les spores sont
sessiles généralement sans le saccule sporifère (fig. 69 E) de couleur jaune doré à orange
(fig.69 A ; B ; C et D).
3.2.2. Les morphotypes de Glomus
Nous avons isolé six morphotypes de Glomus
3.2.2.1. Fiche descriptive de Glomus sp.1 (Planche 1)
Spore : de forme globuleuse, de 115.2 µm de diamètre, de couleur brun orange (0/60/50/10)
issue d‟un hyphe suspenseur non cloisonné (Planche 2, figs. A et B).
Paroi de la spore : elle est composée de trois couches (Planche 2, fig. C).
Couche 1 : très fine, elle se colore en brun foncé dans le réactif de Meltzer.
Couche 2 : c‟est la couche la plus épaisse (8.96 µm), unitaire et lisse, en continuité avec la
paroi de l‟hyphe suspenseur.
Couche 3 : très fine (1.28 µm), transparente, de type membranaire.
Le suspenseur : de forme cylindrique, plus au moins droit, de couleur jaune pale, mesurant
12.8 µm de diamètre au point d‟attachement et de 9 µm plus bas, attaché à son extrémité en
position perpendiculaire à un hyphe très fin (3,84 µm) de même couleur que le suspenseur.
(Planche 2, fig. E)
La paroi de l’hyphe suspenseur : composée d‟une couche de couleur hyaline, de 5.12 µm
d‟épaisseur au point d‟attachement qui se rétrécit jusqu‟à atteindre 2.56 µm de diamètre au
centre et au bas de l‟hyphe

97
Pore : ouvert de 2.56µm de diamètre (Planche 2, fig. D).
Le contenu de la spore : la spore contient des globules ressemblant à des spores miniatures
avec attachement, de 30,7 à 32 µm de diamètre.
3.2.2.2. Fiche descriptive de Glomus sp.2 (Planche 2)
Spore : de forme globuleuse à subglobuleuse parfois ovoïdes, isolée ou en grappe portant 6
à 8 spores de 64 à 89µm de diamètre à l‟état juvénile et 148.48µm à maturité . Elles sont
de couleur brun orange (0/60/50/10) et ont une réaction positive dans le Meltzer (Planche 2,
figs. A et C).
Paroi de la spore : Elle est composée de deux groupes (I et II) (Planche 2, fig. B).
Groupe(I) formé de deux couches
Couche 1 : unitaire, externe, singulière, clairement distinguée des autres couches, de 5.12 µm
chez les jeunes spores et 7.68µm chez les matures, de couleur plus foncée (20/80/80/0) que
celle de la spore mais non continue avec celle du suspenseur ;
Couche 2 : c‟est la couche la plus épaisse 19.2µm, elle adhère étroitement à la surface interne
de la couche 1 et a la même couleur que la spore.
Groupe (II) comprenant deux autres couches :
Couche 3 : de type membranaire, hyaline, elle est de 1.28µm d‟épaisseur.
Couche 4 : de type membranaire (1.28µm), hyaline, entoure l‟intérieur de la spore qui mesure
89.6µm de diamètre, le contenu des jeunes spores se colore en plus foncé, au contact du
réactif de Meltzer et apparait sous forme d‟une couche de 7.68 à 19.2µm mais le fond reste
clair.
L’hyphe suspenseur : cylindrique, courbé et flexible, de 19.2 µm de diamètre au point
d‟attachement (6.4µm chez les jeunes spores) mais de 12.8 µm plus loin, de couleur marron
pale (0/20/20/0) (Planche2 , fig. D).
La paroi du suspenseur : composée d‟une seule couche hyaline de 3.84µm d‟épaisseur
(Planche 2, fig. E).

98
3.2.2.3. Fiche descriptive de Glomus sp. 3 (Planche 3)
Spore : de forme globuleuse, 117,76 µm de diamètre, de couleur jaune, singulière dans le sol,
issue d‟un hyphe suspenseur (Planche 3, fig. A).
Paroi de la spore : elle est composée de trois couches (Planche 3, fig. B).
Couche 1 : très fine, à contours irréguliers ; elle se colore en brun foncé dans le réactif de
Meltzer,
Couche 2 : c‟est la couche la plus épaisse (7,68 µm), unitaire et lisse.
Couche 3 : très fine (1.28 µm), transparente, de type membranaire.
Le suspenseur : de forme cylindrique (Planche 3, fig. C), flexible, de 19,2 µm de diamètre à
son attachement à la spore et 12,8 µm plus loin, de couleur jaune pale.
La paroi de l’hyphe suspenseur : elle est composée d‟une seule couche (Planche 3, fig. C).
Couche 1 : de même couleur que le suspenseur, de 4 µm d‟épaisseur au point d‟attachement
puis se rétrécit jusqu‟à atteindre 1 µm d‟épaisseur au centre et à la partie inférieure de
l‟hyphe
Pore : fermé par un septum.
Le contenu de la spore : riche en globules lipidiques (Planche 3, fig. B)
3.2.2.4. Fiche descriptive de Glomus sp. 4 (Planche 4).
Spore : de forme sub-globuleuse, (64 × 66.56µm) de diamètre, de couleur jaune, singulière
dans le sol, issue d‟un long hyphe suspenseur (Planche 4, fig. A).
Paroi de la spore : elle est composée de deux couches (Planche 4, fig. B).
Couche 1 : de type laminaire, de 6.4 µm d‟épaisseur, elle est en continuité avec la paroi de
l‟hyphe suspenseur, elle se colore en brun orange dans le réactif de Meltzer.
Couche 2 : de type membranaire, elle est transparente, adhère étroitement à la surface interne
de la couche 1.
Le suspenseur : cylindrique, transparent, un peu courbé au niveau de l‟attachement puis
devenant droit, de 7, 68 µm de diamètre au point d‟attachement à la spore.
La paroi de l’hyphe suspenseur : est composée d‟une seule couche de même couleur que
le suspenseur, de 1.28µm d‟épaisseur.

99
3.2.2.5. Fiche descriptive de Glomus sp. 5
Spore : de forme sub-globuleuse, de 65 ×70 µm de diamètre, de couleur jaune orangé,
singulière dans le sol, issue d‟un hyphe suspenseur.
Paroi de la spore : elle est composée de trois couches (figure ci-dessous).
Couche 1 : de type unitaire, de 7 µm d‟épaisseur, lisse ;
Couche 2 : de type membranaire, de 3.83µm d‟épaisseur, translucide plus au moins large,
elle est continue avec la paroi de l‟hyphe suspenseur ;
Couche 3 : de type unitaire, de 5.12 µm d‟épaisseur.
Le suspenseur : cylindrique mais irrégulier, de 10 µm de diamètre à son attachement à la
spore et 7 µm plus loin, de couleur jaune doré.
La paroi de l’hyphe suspenseur : est composée d‟une seule couche de même couleur que le
suspenseur, de 2 µm d‟épaisseur au point d‟attachement à la spore puis se rétrécit jusqu‟à
atteindre 1 µm d‟épaisseur au centre et à la partie inférieure de l‟hyphe.
Pore : ouvert.
3.2.2.6. Fiche descriptive de Glomus sp. 6 (Planche 5)
Spore : de forme globuleuse à sub-globuleuse en grappe constituée de 7 spores de 32×34,56
µm de diamètre, de couleur jaune d‟or (0/20/80/0), terminales portées par un hyphe
suspenseur (Planche 5, figs. A et B).
Paroi de la spore : elle est composée de trois couches (Planche 5, fig. C).
Couche 1 : de type unitaire, qui se colore en marron (20/80/80/0) au contact du Meltzer, de
4.8µm d‟épaisseur ;
Couche 2 : hyaline et plus épaisse (9.6 µm) ;
Le suspenseur : droit reliant toutes les spores, de couleur jaune (0/0/20/0), de 4.8 µm de
diamètre .
La paroi de l’hyphe suspenseur : composée d‟une seule couche de même couleur que le
suspenseur, de 2 µm d‟épaisseur (Planche 5, fig. D).
Pore : fermé par la paroi sporale.

100
3.2.3. Morphotype de Scutellospora (Planche 6)
Spore : de forme globuleuse à subglobuleuse, de diamètre compris entre 38,4 à 51.2×51.2 à
54.4 µm, singulière (planche 6, fig.A ) ou en deux unités dans le sol (planche 6 , fig.B ), de
couleur rouge écrevisse.
Paroi de la spore : elle est composée de deux couches (Planche 6, fig.C )
Couche 1 : ornementée (1.28 µm), se colore en brun dans le réactif de Melzer ;
Couche 2 : c‟est la couche la plus épaisse, de type unitaire, de 4 µm d‟épaisseur
Le suspenseur : bulbeux parfois très large ou ellipsoïde au point d‟attachement (19,2 à 23,04
× 25,6 µm), de même couleur que la spore, épineux (Planche 6, fig.D), l‟hyphe qui porte le
suspenseur est cloisonné et bulbeux, de 6,4 à 12.8 µm de diamètre (Planche 6, fig..E ).
La paroi du suspenseur : est composée d‟une couche de couleur hyaline, de 1.28 µm
d‟épaisseur
Essais d’identification des CMA isolés
A l‟aide des observations microscopiques et des clefs de détermination des CMA
(Walker ,1983 ; Stürmer et Morton ,1997 ; Blaszkowski et Czerniawska, 2008), nous avons
pu décrire six morphotypes du genre Glomus. L‟espèce Glomus sp. 3 présente de
nombreuses caractéristiques qui la rapprochent de Glomus macrocarpum ( Gerdemann et
Trappe, 1974 ; Berch et Fortin ,1983 ; Blaszkowski, 2003 ; Sharma et al. , 2008) :
morphologie et dimension de la spore, singulière dans le sol ou en sporocarpe, épaisseur de la
2ème couche de la paroi sporale, présence d‟un pore sporal fermé par un septum et de globules
lipidiques dans le contenu sporal, dimension de l‟hyphe suspenseur présentant une seule
couche.
D‟autres morphotypes du genre Glomus (05), Acaulospora (04) et Scutellospora (01)
n‟ont pas pu être identifiés à cause de l‟incertitude de la caractérisation morphologique.
4. Le pouvoir endomycorhizogène du sol (PEM)
Les plants de poireau âgés de 4 mois présentent une meilleure croissance surtout avec
les dilutions du sol 10-2, 10-3 et 10-4 (planche 1). Les résultats du calcul du nombre le plus
probable (NPP) de propagules viables révèlent que :
- dans le sol de la station d‟étude 1, comprenant les vergers âgés de 5 ans, NPP= 5500
propagules par Kg de sol avec un intervalle de confiance à 95 % de 0.33 et 4.4.

101
- dans le sol de la station d‟étude 2, comprenant les vergers âgés de 17 ans, NPP= 7000
propagules par Kg de sol avec un intervalle de confiance à 95 % de 0.42 et 4.62.
Nos résultats rejoignent ceux de Saad (2009) qui a montré que ces sols sont riches en
propagules viables indiquant leur bon état biologique. Ils sont aussi en accord avec ceux de
Azcon-Aguilar et al. (2003) qui ont constaté que la rhizosphère de l‟oliv ier Olea europea
sub.sp.sylvestris d‟une région semi aride d‟Espagne (province d‟Alicante-sud d‟Espagne) est
plus riche en propagules infectieuses (NPP=2970 propagules par Kg de sol) que celle
d‟autres plantes.
Certains travaux ont mentionné un PEM de 33000 (propagule)/Kg dans 50 terrains
agricoles (Wang et al., 2008) et de 36000 p/Kg dans la rhizosphère de l‟oignon (Sow et al. ,
2008), dans d‟autres, il est supérieur à 100000 propagules par Kg de sol (Scheltema et al.
1987; Land et al. 1993).
Par ailleurs, les analyses de sols des oliveraies, des stations d‟étude, ont montré que ces
sols sont pauvres en éléments minéraux (Saad, 2009) surtout en phosphore (0.008% et 0.02%
pour les stations 1 et 2 respectivement). La pauvreté du sol en phosphore assimilable explique
souvent la richesse en propagules de CMA (Plenchette et al., 1983 ; Scheltema et al., 1987;
Chelius et Triplett, 1999; Troech et Loynachan, 2003) . EEnn ee ffffee tt,, lleess MMAA ssoonntt pp lluuss ffrrééqquueenntteess
ddaannss lleess ssoo llss ppaauuvvrreess éé lléémmeennttss mmiinnéérraauuxx eesstt ffaa iibb llee ((Bâ eett aall..,, 22000011)) eett l‟importance de la
colonisation racinaire et la sporulation dépend du niveau de P assimilable dans le sol (Douds
et Millner, 1999; Kahiluoto et al. 2001).
DD‟‟aauuttrreess ffaacc tteeuurrss ppeeuuvveenntt affecter le potentiel infectieux ddeess CCMMAA ddaannss llee ssoo ll ttee llss qquuee
ll‟‟eennvviirroo nnnneemmeenntt,, llee ttyyppee ddee ssoo ll eett llaa ccuullttuurree ((AAddee llmmaann eett MMoorrttoonn,, 11998866)),, le pH (Daniels et
Trappe, 1980) et le nombre des spores dans le sol (Harinikumar et Bagyaraj, 1989; Arihara et
Karasawa ,2000; Vestberg et al. ,2005).
Il faut noter que le PEM permet d‟estimer seulement le potentiel de colonisation relatif
des CMA dans les conditions expérimentales utilisées et des situations écologiques qui sont
souvent très différentes et les intervalles de confiance de la méthode PEM sont le reflet du
facteur de dilution et du nombre de réplications (Cochran, 1950).
D‟autres facteurs influent aussi sur les résultats du PEM (plante hôte, température, période
de récolte, dimensions des pots, état du sol (sec ou humide) et son statut nutritionnel, la
densité des racines) (Wilson et Trinick, 1982).

102
5. Cinétique de la mycorhize
Les résultats représentés par l‟histogramme (Fig. 70) montrent que le poireau est
hautement mycotrophe par son piégeage des CMA contenus dans les sols des deux
parcelles, de la région semi aride. Après dix semaines de sa culture, les fréquences de
mycorhization (F) atteignent des niveaux élevés : 85% avec les CMA de la parcelle1 et
94.59% et avec ceux de la parcelle 2. Quant aux intensités de colonisation du cortex
racinaire (M), elles sont de 44.25% et de 60.08 % pour les deux parcelles (1 et2
respectivement) ; les taux d‟arbuscules du système radiculaire de la partie mycorhizée (a)
sont de 79.86% et 89.46% respectivement dans les parcelles 1 et 2.
Fig 70: Estimation des paramètres d’infection mycorhizienne des plants de poireau
F :Fréquence de mycorhization ; M et m : intensité de mycorhization, A et a : Teneur en
arbuscules du système radiculaire et de la partie mycorhizée respectivement ; P1 : Parcelle 1 ;
P2 :Parcelle2.
5.1. Estimation des paramètres d’infection mycorhizienne des plants de poireau au cours
de dix semaines de culture
5.1.1. Fréquence de mycorhization :
On constate que les fréquences d‟infection des systèmes racinaires sont pratiquement
nulles lors de la première semaine qui suit les semis des graines de poireau ; au 10ème jour de
86
58,15
50,4
79,86
40,24
94,89
63,5160,04
89,46
53,74
0
10
20
30
40
50
60
70
80
90
100
F % m % M % a % A %
(%)
P1
P2

103
culture, elles sont < 20%. Au cours de cette période, la plante produit son système racinaire
tandis que les propagules fongiques germent pour produire du mycélium pré-symbiotique.
Après la phase de reconnaissance entre les deux partenaires par leurs échanges
moléculaires, l‟interaction entre les deux symbiotes débute. Ainsi, on constate une
augmentation progressive du taux d‟infection qui atteint 93% et 94.59% respectivement à la
neuvième et la dixième semaine avec les CMA de la parcelle 2, par contre, il ne dépasse pas
86 % avec ceux de la parcelle 1 même à la fin de l‟expérience (fig. 71).
5.1.2. L’intensité de mycorhization :
Les hyphes des espèces fongiques indigènes provenant de la parcelle 2 envahissent les
cellules racinaires à la troisième semaine et celles la parcelle 1 à la 5ème semaine. Aux 5ème et
6ème semaines, les complexes fongiques de la parcelle 2 progressent à l‟intérieur des cellules
corticales avec une intensité (M) de 23 % et 28 % qui augmente jusqu‟à 60% à la fin de
l‟expérience.
Au cours des premières semaines de culture des plants de po ireau issus de semis, sur le
sol provenant de la parcelle (1), l‟intensité de mycorhization reste très faible mais constante
(0.44, 0.70 %) jusqu‟à la sixième semaine. Selon Trouvelot et al. (1986), la colonisation est
de niveau 1 dès la 7ème semaine, ensuite, l‟intensité de mycorhization augmente jusqu‟à 50 %
à la fin de l‟expérience (fig. 72).
5.1.3. Le taux d’arbuscules :
Les examens microscopiques des systèmes racinaires montrent que la production des
arbuscules débute dès la 5ème semaine avec les CMA de la parcelle 2 et à la 7ème semaine avec
ceux de la parcelle 1, c'est-à-dire 3 à 5 semaines après la colonisation des cellules par le
champignon. Nos résultats se rapprochent de ceux de Simoneau et al. (1994) qui ont montré
que les arbuscules se différentient en 4 jours à un mois après l‟infection mycorhizienne ;
cette période est probablement nécessaire pour l‟extension mycélienne intra-racinaire
jjuussqq uu''aauuxx ccee lllluulleess ccoorr tt iiccaa lleess iinntteerrnneess où se forment les arbuscules. Les arbuscules se trouvent
dans les racines fines pendant les stades précoces de l‟infection racinaire (Smith et Read,
1997; Harrison, 1999).

104
Nous avons constaté que le taux d‟arbuscules augmente progressivement jusqu‟à
atteindre 50% et 60 % du système racinaire du poireau cultivé sur les sols provenant des
parcelles 1 et 2 (fig.73).
En revanche, la formation des vésicules par les complexes fongiques indigènes des deux
parcelles s‟effectue une semaine après la formation des arbuscules et à des taux très faibles,
sachant que les vésicules sont abondantes dans les colonisations anciennes (Brundrett et al.,
1999).
Ces résultats rejoignent ceux de DDoodddd eett aall.. ((22000000 )) eett DDaa llppéé eett aall.. ((22000055)) qui affirment que la
formation des arbuscules précède celle des vésicules..
Dans tous les cas, les cinétiques de mycorhization se présentent sous forme de courbes
sigmoïdes comme l‟ont décrits Gianinazzi- Pearson et al. (1985) et Oihabi et Meddich (1996)
chez le trèfle, Blal et al. (1990) et Oihabi (1991) chez le palmier dattier.
L‟infection endomycorhizienne du poireau est plus élevée avec les CMA du sol
rhizosphérique des oliviers âgés de 15 ans que celle du sol des jeunes oliviers de 4 ans ; cette
différence proviendrait du nombre de propagules viables qui diffèrent dans les deux sols,
puisque nos résultats comme ceux de Saad (2009) ont montré que le sol de la parcelle 2 est
plus riche en propagules que celui de la parcelle 1. Ces résultats pourraient s‟interpréter par
les niveaux de fertilité des deux sols en matière organique ou en minéraux.
P 1
P 2
0
10
20
30
40
50
60
70
80
90
100
1 2 3 4 5 6 7 8 9 10
pourcentage (%)
temps (semaine)
F%
F%

105
Fig.71 : Variation des taux d’infection des CMA indigènes des parcelles 1 (P1) et 2 (P2)
pendant 10 semaines de culture des plants de poireau.
Fig.72: Variation de l’intensité mycorhizienne des CMA indigènes des parcelle s 1 (P1) et
2 (P2) pendant 10 semaines de culture des plants de poireau.
P 1
P 2
0
10
20
30
40
50
60
70
1 2 3 4 5 6 7 8 9 10
po
urc
en
tage
(%
)
temps (semaine)
M %
M %

106
Fig.73: Variation du taux arbusculaire des CMA indigènes des parcelles 1 (P1) et 2 (P2)
pendant 10 semaines de culture des plants de poireau.
5.2. Étude du rôle de la mycorhize sur la production de biomasse des plants de poireau
Les figures 74 et 75 représentent l‟évolution dans le temps de la croissance des plants de
poireau élevés sur le sol rhizosphérique des oliviers de la parcelle 1 ou 2. Après dix
semaines de culture, l‟effet de la mycorhization est nettement visible, les CMA indigènes de
la parcelle 2 semblent mieux favoriser la croissance des plants que ceux de la parcelle 1.
La biomasse des parties aériennes des plants mycorhizés (en matière sèche) est deux fois
supérieure à celle des plants témoins ; elle est de 152.6 % avec une dépendance
mycorhizienne de 52% à la fin de l‟expérience.
Nos résultats rejoignent ceux de Fortin et al. (2008) qui ont montré que la dépendance
mycorhizienne du poireau peut atteindre 97% dans un sol pauvre en phosphore parce que
cette plante, comme d‟autres plantes (tels que la tomate, l‟oignon et la carotte), possède un
système racinaire grossier qui dépend complètement du champignon dans l‟exploration du
sol et l‟absorption du P pour une croissance optimale contrairement aux plantes qui ont un
système racinaire très ramifié leur permettant de s‟alimenter sans l‟association mycorhizienne
(ex. les graminées) (Dodd, 2000 ; Merryweather, 2001 ; Janos, 2007 ; Fortin et al., 2008).
D‟autres travaux ont mentionné que le ppoo iirreeaauu nnoo nn mmyyccoorrhhiizzéé aa uunnee ccrroo iissssaannccee
oopptt iimmaa llee ssuurr uunn ssoo ll ooùù llaa ccoo nncceennttrraa tt iioonn ddee PP aassss iimmii llaabb llee eesstt ddee 8800 àà 110000mmgg//KKgg (Jakobsen,
P 2P 1
0
10
20
30
40
50
60
1 2 3 4 5 6 7 8 9 10
pou
rcen
tage
(%)
temps (semaine)
A %
A %

107
1995), cceeppeennddaanntt,, iill ppeeuutt aavvoo iirr llee mmêêmmee nniivveeaauu ddee cc rroo iissssaannccee ss ‟‟ iill eess tt mmyyccoorrhhiizzéé ssuurr uunn ssoo ll
ccoonntteennaanntt 1155 àà 2255 mmgg //KK gg ddee PP aassss iimmii llaabb llee ((SSoorreennsseenn eett aall..,, 22000055)).. DD‟‟aauuttrreess pp llaanntteess ssoonntt
ddééppeennddaanntteess ddee llaa mmyyccoorrhhiizzee ccoo mmmmee llee JJuujjuubb iieerr ((>> 7700%%)) ((BBaa eett aall..,, 22000011)),, ll‟‟aarrggaanniieerr ((48 %)
(Echairi et al., 2008) et le bananier (35 à 50%) (JJaa iizzmmee --VVeeggaa eett aall..,,22000022).
Nous avons constaté que l‟effet de l‟endomycorhization sur la croissance des plants
varie selon les complexes fongiques des deux parcelles ; les plants âgés de cinq semaines
mycorhizés avec les CMA de la parcelle 2 sont plus développés que les témoins, tandis que
chez ceux de la parcelle 1, cet effet apparaît après 6 à 7 semaines de culture. Il y a aussi une
augmentation de l‟intensité de l‟infection mycorhizienne et la production d‟arbuscules qui
représentent des sites d‟échange de nutriments chez les deux partenaires de la symbiose. En
effet, le gain de croissance observé pourrait s‟expliquer soit par l‟amélioration de
l‟absorption de l‟eau et des minéraux présents dans le sol ou par une meilleure assimilation
du phosphate et d‟autres éléments peu mobiles tels que le cuivre et le zinc.
Pendant les premiers stades du développement des plants, la croissance des plants
mycorhizés était analogue à celle des témoins ; c‟est à partir de la 6ème semaine qu‟on
observe leurs différences de croissance. La croissance la plus importante des plants serait liée
aux CMA qui ont donné le meilleur taux d‟infection mycorhizienne et qui sont aptes à
coloniser plus rapidement les racines (meilleure intensité mycorhizienne).
Des résultats analogues ,montrant la relation entre la stimulation de la croissance des
plants et le niveau élevé d‟infection mycorhizienne par les CMA, ont été obtenus dans les
cas de la vigne (Olivier et al., 1983) et du palmier dattier (Oihabi, 1991).
La mycorhization des plants améliore aussi la croissance de la partie aérienne des plants (fig.
76). Après 10 semaines de culture, la hauteur des plants mycorhizés est encore plus
importante que celle des témoins , elle est de 110% (parcelle 1) et de 130% ( parcelle 2).

108
Fig. 74 : Influence des CMA de la parcelle 2 sur la biomasse des plants de poireau.
Psm2 : Poids sec des plants mycorhizées. Pst2 : Poids sec des plants témoins;
Ltm2 : Longueur des tiges des plants mycorhizés ; Ltt2 : Longueur des tiges des plants témoins.
Fig. 75 : Influence des CMA de la parcelle 1 sur la biomasse des plants de poireau.
Psm1 : Poids sec des plants mycorhizés; Pst1 : Poids sec des plants témoins ;
Ltm1 : Longueur des tiges des plants mycorhizés ; Ltt1 : Longueur des tiges des plants
témoins
1,31 1,42,3
2,78 33,8
4,5
6,23
8,7
45
6,37,1
9,7
12,34
14,7
17,318,2
0 0 01 1,33
2,333
4,665,7
1
3
5
6,27
8
11
13,6 14
0
2
4
6
8
10
12
14
16
18
20
2 3 4 5 6 7 8 9 10
Psm2
Ltm2
Pst2
Ltt2
2
3
4,48
5,62
7
9,6
11,212
13
0 0 0
11,5
2
3
4
5
0 0
1
3
4,5
6
7,5
9
10,5
1,1 1,181,8 1,9
2,33
3,7
4,57
5,6
0
2
4
6
8
10
12
14
2 3 4 5 6 7 8 9 10
Ltm1
Pst1
Ltt1
Psm1

109
5.3.Étapes de l’infection racinaire des plants de poireau inoculés par les CMA indigènes
5.3.1. La pénétration
Nous avons constaté que les premiers signes d‟infection débutent par l‟apparition des
appressoriums après 7 à14 jours de culture (fig. 77 B) c'est-à-dire 4 à 5 jours après la
germination des graines et la formation du système racinaire (fig. 77 A). Ces résultats
concordent avec ceux de DDaa llppéé eett aall.. ((22000055)) qquuii oonntt montré que l‟infection s‟effectue en 3 à
12 jours ; llaa ffoorrmmaatt iioonn ddee ll‟‟aapppp rreessssoorr iiuumm ssee ffaa iitt eenn 3366 hh aapprrèèss llee ccoonnttaacc tt dduu cchhaammpp iiggnnoonn
aavveecc lleess ccee lllluu lleess éépp iiddeerrmmiiqquueess eett ll‟‟aappppaa rree iill ddee pprréé--ppéénnééttrraatt iioonn ssee pprrééppaa rree pour la ppéénnéé ttrraa tt iioonn
dduu cchhaammpp iiggnnoonn àà ll‟‟ iinnttéérr iieeuurr ddee llaa ccee lllluullee ..
DDiifffféérreenntt aassppeecc ttss dd‟‟aapppp rreessssoo rr iiuumm oonntt ééttéé oobbsseerrvvééss ddaannss lleess ssyyss ttèèmmeess rraacc iinnaa iirreess (( ffiigg.. 7788))..
55..33..22.. DDéévvee llooppppeemmee nntt ddeess hhyypphheess ddaannss llee ccoorrtteexx rraa cciinnaaiirree
55..33..22..11.. LLee mmyyccéé lliiuumm iinnttrraa --rraacciinnaa iirree
LLaa ppéénnééttrraa tt iioonn ddeess hhyypphheess ddaannss lleess ccee lllluulleess ccoo rrtt iiccaa lleess ss‟‟ee ffffeecc ttuuee ppaarr lleess ccee lllluulleess
éépp iiddeerrmmiiqquueess pp uuiiss iillss ss‟‟eennrroo uulleenntt ddaannss lleess pp rreemmiièèrreess ccee lllluulleess dduu pp aarreenncchhyymmee ccoo rrtt iiccaa ll
(( ffiigg.. 7799))..
LLeess hhyypphheess ssee pprrooppaaggeenntt ddaannss lleess eessppaacceess iinntteerrccee lllluu llaa iirreess dd uu ppaarreenncchhyymmee ccoorrtt iiccaa ll ddee llaa
rraacc iinnee eenn ssee rraammiiffiiaanntt eett eenn ffoo rrmmaanntt ddeess aannaass ttoommoosseess eenn «« HH »» ee tt eenn «« YY »»(( ffiigg..8800 )) .. DD‟‟aapp rrèèss
lleess mmoorrpp hhoo ttyyppeess ddeess hhyypphheess iinnttrraa-- rraacc iinnaa iirreess,, oo nn ppeeuutt ssuuppppoosseerr llaa pprrééddoo mmiinnaannccee ddeess ggeennrreess
GGlloommuuss eett AAccaauulloossppoorraa nnééaannmmoo iinnss,, oonn oobbsseerrvvee aauussss ii ddeess hhyypphheess ééppaa iiss dduu ttyyppee 33,, qquuii ssoonntt
ffoo rrtteemmeenntt ccoo lloorrééss aauu bb lleeuu ddee ttrryyppaann..
55..33..22..22.. LLeess ppee lloottoonnss
La troisième et la quatrième semaine après le semis des graines, les hyphes
progressent dans le cortex racinaire (fig. 81) ; ils se différencient en pelotons mycéliens intra
ou intercellulaires qui sont généralement peu abondants et souvent localisés dans les cellules
les plus externes du cortex.
CC
E
C

110
5.3.2. 3. Les arbuscules
Les arbuscules apparaissent à partir de la 5ème semaine de culture (fig. 82); ils sont de
type « Arum » (type 1). Nos résultats rejoignent ceux de Simoneau et al. (1994) qui ont
montré que ces structures se différencient un mois après la colonisation du cortex. Les
arbuscules de type « Arum » que nous avons observés se rencontrent fréquemment chez les
plantes cultivées (Fisher et Jayachandran, 2005 ; Muthukumar et al., 2006 ;Muthukumar et
PPrraakkaasshh ,2009). Ce morphotype serait approprié aux cultures annuelles en raison de la
rapidité de la croissance racinaire des plantes qui est en harmonie avec l‟extension rraapp iiddee de
ce type de MA (Yamato, 2004). D‟autres plantes cultivées forment aussi ce type de MA
comme le maïs (Greny, 1973).
Les arbuscules de type 1 observés sont caractérisés par un tronc cylindrique, des branches ou
ramifications fines et lâches (fig. 83), leur coloration au bleu de trypan est peu intense, ils
sont caractéristiques du genre Acaulospora.
Dans les systèmes racinaires, nous avons aussi observé d‟autres types morphologiques
d‟arbuscules :
des arbuscules de type 2 sont les plus fréquents (fig.84 A et B); ils sont à tronc
cylindrique avec des ramifications dichotomiques, ils prennent une coloration modérée
avec le bleu de trypan et sont caractéristiques du genre Glomus.
des arbuscules de type 3 sont très peu répandus, ils sont à tronc bulbeux ou gonflé
(fig. 85) et sont souvent formés par les genres Gigaspora et Scutellospora.
La fréquence des arbuscules des types 1 et 2 reflète la dominance des CMA
appartenant des genres Acaulospora et Glomus et explique le nombre de morphotypes de ces
deux genres que nous avons isolés et décrits précédemment.
La présence relative des arbuscules de type 3 caractéristiques des Gigasporacées
confirme nos résultats puisque le nombre de morphotypes de Scutellospora isolés est très
faible. Nos résultats rejoignent ceux de nombreux travaux qui ont montré que les
Gigasporacées cohabitent souvent avec d‟autres CMA dans les écosystèmes naturels, mais
elles sont peu abondantes aussi bien dans les écosystèmes tropicaux (Lovelock et al., 2003 ;
Siqueira et al., 1989 ; Zhao et al., 2001 et 2003 ; Khade et Rodrigues, 2003) que tempérés
A B

111
(Helgason et al., 1998). Elles sont relativement présentes ou absentes dans les cultures
annuelles, (Sieverding, 1991; Helgason et al., 1998; Daniell et al.,2001; Jansa et
al.,2002;Vandenkoornhuyse et al., 2002; Oehl et al., 2003; Johanson et al., 2004).
3.2.4. Les vésicules
Après la 7ème semaine de culture, nous avons observé la présence de quelques vésicules
dans les tissus de la plante (fig. 86) de forme globulaire ou ellipsoïde (fig. 90) qui
confirment encore la prédominance des Glomus. Ces Gloméracées ssppoo rruulleenntt eett ddéévvee llooppppeenntt
uunn rréésseeaauu dd‟‟hhyypp hheess pp lluuss rraapp iiddeemmeenntt qquuee lleess aauuttrreess CCMMAA.. CCeess rrééssuullttaa ttss rree jjoo iiggnneenntt cceeuuxx ddee
Souza, (2005) et de Wubet et al. (2003) qui ont rapporté que les Gloméracées infectent et
colonisent plus rapidement les racines que les Gigasporacées et les Acaulosporacées.
Dans les systèmes racinaires, on observe aussi des vésicules qui sont lobées irrégulières
et faiblement colorés au bleu de trypan (fig.88); elles sont caractéristiques d’Acaulospora.
L‟ensemble de nos observations indiquent la présence des structures intra-racinaires
caractéristiques des genres Glomus, Acaulospora et Scutellospora ou Gigaspora. Des
résultats analogues ont été obtenus avec la mycorhize naturelle de l‟olivier de la région de
Sig , donc nous pouvons déduire que les plants de poireau ont été infecté par des espèces
de CMA de la rhizosphère de l‟olivier appartenant à ces genres.
Lors des examens microscopiques des racines des plants de poireau mycorhizées, nous
avons remarqué dans 40 % des systèmes racinaires en plus des CMA, la présence d‟autres
espèces fongiques bien colorées au bleu de trypan. Ces espèces sont des endophytes, leurs
filaments mycéliens sont cloisonnés (fig. 89) et ils forment des pelotons intracellulaires (fig.
90).
DDaannss lleess rraacc iinneess nnaattuurree lllleess ddee ll‟‟oo lliivviieerr ddee llaa rrééggiioonn ddee SS iigg,, nnoouuss aavvoo nnss aauussss ii oobbsseerrvvéé uunn
mmyyccéé lliiuumm sseeppttéé ddeess EEMMSS ddoonncc lleeuurr pp rréésseennccee ddaannss llaa rrhhiizzoosspp hhèèrree ddee ll‟‟oo lliivv iieerr eesstt bb iieenn àà
ll‟‟oorr iiggiinnee ddee ll‟‟ iinnffeecc tt iioonn oobbsseerrvvééee ddaannss llee ss rraacc iinneess ddeess pp llaannttss ddee ppoo iirreeaauu.. CCeeppeennddaanntt,, nnoouuss nnee
ppoouuvvoonnss ppaass aa ffffiirrmmeerr ll‟‟ee ffffee tt ddee cceess eennddoopphhyytteess ssuurr llee ddéévvee llooppppeemmeenntt ddeess pp llaannttss ddee ppoo iirreeaauu
ccaarr iill ffaauuddrraa iitt lleess iissoo llee rr eett lleess iiddeenntt iiffiiee rr aavvaanntt ddee mmeett ttrree eenn éévviiddeennccee lleeuurr rrôô llee..
6. Effet des CMA indigènes au sol des oliveraies sur la croissance des boutures
enracinées de la variété Sigoise d’olivier.

112
6.1. Estimation des paramètres d’infection des plants d’olivier par les CMA
Les observations microscopiques des tissus racinaires des plants d‟olivier inoculés par
les fragments racinaires et par du substrat de culture du poireau infecté par les isolats de
CMA, ont révélé la présence de mycorhizes arbusculaires, après trois mois de culture. Les
résultats représentés par l‟histogramme (fig. 91) montrent que l‟olivier est mycotrophe.
Après trois mois de sa culture, les fréquences de mycorhization (F) atteignent des
niveaux moyens dans les deux premières séries d‟inoculation (CMS et CMB) 30 et 58 %
respectivement et très faibles (< 10%) pour les deux autres séries d‟inoculation (I1 et I2).
Quant aux intensités de colonisation du cortex racinaire (M), elles sont de 11.14% et de
13.04 % pour les deux séries CMS et CMB, et sont quasiment nulles pour les deux autres
séries d‟inoculation (0.74% et 1% pour I1 et I2).
Les taux d‟arbuscules du système radiculaire de la partie mycorhizée (a) sont de
81.7% et 59.13% respectivement dans les séries d‟inoculation CMS et CMB et
pratiquement nuls pour les deux autres séries d‟inoculation.
Fig. 91: Estimation des paramètres d’infection mycorhizienne des plants d’olivier
F : Fréquence de mycorhization M et m : intensité de mycorhization relative et absolue
respectivement, du cortex colonisée par rapport à l‟ensemble du système rac inaire ; Teneur en arbuscules du système radiculaire (A) et de la partie mycorhizée (a).
Les examens microscopiques des racines montrent que la présence d‟appressorium,
des hyphes intra et extra-racinaires (planche 8, fig. c; planche 9, fig. c), des enroulements
30
37,13
11,14
81,7
9,1
58
22,48
13,04
59,13
7,718 9,25
0,74 0 0
6
0,06 1 0 00
10
20
30
40
50
60
70
80
90
F % m % M% a% A %
(%)
CMS
CMB
I1
I2

113
(planche 8, fig. d ; planche 9, fig. d) des arbuscules (planche 8, fig. e ; planche. 9, fig. e) et
quelques vésicules de formes ovoïdes (planche 8, fig. f) ou allongées (planche 9,fig. f)
caractéristiques des Gloméracées. La présence de quelques vésicules dans les racines des
plants d‟olivier indique que les MA sont au stade jeune. La morphologie des mycorhizes
arbusculaires est analogue à celle des MA naturelles.
Nous avons aussi observé la présence des eennddoopphhyytteess àà mmyyccéé lliiuumm sseeppttéé ((EMS) dans les
séries d‟inoculation CMS (planche 8, fig. g), CMB (planche 9, fig. g) et I1 (planche 10, fig. c)
mais à des taux très faibles entre 4 et 8 %. En revanche, les racines des plants témoins sont
exemptes d‟infection (fig. 92).
6-2: Estimation de la croissance des plants d’olivier mycorhizés, en conditions
contrôlées
Six paramètres de croissance ont été estimés : la longueur des racines, la hauteur, le
nombre de tiges et de feuilles et les poids frais et secs de la partie aérienne.
Les résultats des mesures de paramètres de croissance des plants d‟olivier ( longueur des
racines ; hauteur, nombre de tiges et de feuilles, poids frais et secs de la partie aérienne) sont
présentés en Annexe 3.
Les résultats des analyses statistiques sont présentés en Annexe 4.
Les résultats montrent que la mycorhization stimule la croissance des plants inoculés par
rapport aux témoins ; son effet se manifeste par :
L‟augmentation des paramètres de croissance des plants mycorhizés par rapport aux
témoins (fig.93, A).
et l‟importance du chevelu racinaire chez les plants inoculés (Fig.93, B).
Les résultats obtenus montrent une différence de croissance entre les plants d‟olivier
mycorhizés et les témoins dans les 4 niveaux d‟inoculation CMS (P lanche 8, figs. a et b),
CMB (Planche 9, figs. a et b), I1 (Planche 10, figs. a et b) et I2 (Planche 10 figs. d et e).

114
Hauteur de la partie aérienne des plants
Les analyses de variances montrent des effets significatifs marqués de la mycorhization
sur la hauteur des plants inoculés (Annexe 4).
Le test de Newman-Keuls confirme ces effets (Annexe 4) et souligne que la différence
se situe entre la hauteur des plants inoculés par I2 et les témoins. Le rapport hauteur des plants
inoculés/hauteur des plants témoins varie de 1.18 à 1.47 ce qui explique l‟importance des
plants inoculés par rapports aux témoins (fig. 94).
Longueur des racines
Les analyses de variance montrent des effets significatifs marqués de la mycorhization
sur la longueur des racines des plants inoculés (Annexe 4) le test de Newman-Keuls confirme
ces effets (Annexe 4 ).
Les plants inoculés par les CMA ont une longueur des racines plus importante que celle
des plants non mycorhizés (fig. 95)
Nombre de feuilles
Les analyses de variance montrent des effets significatifs marqués de la mycorhization
sur le nombre de feuilles des plants inoculés (Annexe 4). Le test de Newman-Keuls confirme
ces effets (Annexe 4).
Les plants inoculés par les CMA ont un nombre de feuilles plus important que celui des
plants non mycorhizés (fig. 96)
Nombre de tiges

115
Les analyses de variance montrent des effets significatifs marqués de la mycorhization
sur le nombre de tiges des plants inoculés (Annexe 4). Le test de Newman-Keuls confirme ces
effets (Annexe 4 ) Les plants inoculés par les CMA ont un nombre de tiges plus important que
celui des plants non mycorhizés (fig. 97).
Poids frais
Les analyses de variance montrent des effets significatifs de la mycorhization sur le
poids frais de la partie aérienne des plants inoculés (Annexe 4). Le test de Newman-Keuls
confirme cet effet (Annexe 4 ). Le rapport poids frais des parties aérienne des plants inoculés/
poids frais des parties aérienne des plants témoins varie de 1.1 à 1.78, ces résultats montrent
que la croissance des plants inoculés est importante que celle des témoins (fig.98).
Poids sec de la partie aérienne
Les analyses de variance montrent des effets significatifs marqués de la mycorhization
sur le poids sec de la partie aérienne des plants inoculés (Annexe 4). Le test de Newman-Keuls
confirme cet effet (Annexe 4). Le rapport poids sec des parties aérienne des plants inoculés/
poids sec des parties aérienne des plants témoins varie de 1.341 à 1.729 sauf pour l‟isolat I1 qui
est inférieur à 1 (0.95), ces résultats montrent que la croissance des plants inoculés (sauf celle
des plants inoculés par I1) est importante que celle des témoins (fig. 99).La différence entre le
poids frais et le poids sec des plants inoculés et témoins nous renseignent sur la teneur en eau
qui est plus élevée chez les plants inoculés. L‟analyse de tous ces paramètres de croissance
montre que l‟inoculation des plants d‟olivier par les isolats I2, CMB et CMS améliorent
significativement la croissance en hauteur et en biomasse des plants d‟olivier.
La meilleure croissance des plants inoculés est obtenue dans la série CMB où les rapports
de la hauteur des plants inoculés /hauteur des plants témoins est de 1.35 ; ceux des poids frais
de la partie aérienne des plants mycorhizés/ poids frais de la partie aérienne des plants témoins
est de 1.78 ; ceux des poids sec de la partie aérienne des plants mycorhizés/ poids sec de la
partie aérienne des témoins est de 1.729. D‟ailleurs, les plants d‟oliviers appartenant à cette
série ont un nombre plus important de tiges et de feuilles ; leurs racines sont plus longues.

116
Ces résultats rejoignent ceux de nombreux auteurs qui ont montré les avantages de la
mycorhization sur des boutures semi ligneuses de la variété « Sigoise » d‟olivier inoculées par
des CMA indigènes des sols d‟oliviers de Sig (Saad, 2009 ; Saad et al. 2010), et sur des
variétés « Cornicabra, Arbequina, Leccino et Picual » d‟oliviers inoculés par des Glomus
(Gl. mosseae, Gl. intraradices, Gl. claroideum et Gl. viscuosum) (Calvente et al. , 2004 ; Porras
Piedra et al., 2005 ; de Rougemont , 2008). Des résultats analogues ont été rapportés par Jasper
et al. (1989) qui ont remarqué que les associations symbiotiques avec les CMA améliorent la
croissance précoce de certains arbres et leur maintien dans les sols des zones arides et semi
arides déficients en nutriments. D‟autres travaux ont aussi montré que l‟inoculation par des
CMA des espèces fruitières, en particulier le pommier et le fraisier Derkowska et al., 2008),
et des vitroplants de pommier améliore leur croissance et leur nutrition surtout pendant la
période d‟acclimatation des pommiers micropropagés (Cavalazzi et al. 2007).
En ce qui concerne, les systèmes racinaires des boutures herbacées mycorhizées, nos
résultats ont montré que la mycorhization augmente la ramification des racines avec un
chevelu racinaire important CMS (Planche 8, fig. b), CMB (Planche 9, fig.b), I1 (Planche 10,
fig. b) et I2 (Planche 10 fig. e). Ces résultats rejoignent ceux décrits chez certaines espèces
fruitières mycorhizées comme Olea europea L. (Citernesi et al., 1998), Prunus cerasifera L.
(Berta et al., 1995) , Vitis vinifera L (famille des Vitacées) (Schellenbaum et al., 1991), le
palmier dattier (Dreyer et al. 2009) et Fragaria ananassa Duch. (famille des Rosacées)
(Norman et al., 1996).
Cette stimulation de la croissance racinaire (modification de la morphologie des racines,
augmentation de leur nombre et leurs ramifications, jaunissement et absence totale des poils
absorbants) améliore, selon certains auteurs, l‟absorption de l‟eau et la nutrition minérale
(Daniels-Hetrick et al., 1988 ; Kaldorf et Ludwig-Müller , 2000 ; Fidelibus et al., 2000 ;
Fester et al. 2002 ; Derkowska et al. , 2008).
6.3. Estimation de la dépendance mycorhizienne des plants d’olivier
Les résultats obtenus montrent que les plants d‟oliviers sont différemment dépendants
de la mycorhization (fig. 100).
Avec un degré de dépendance mycorhizienne de 72.9%, les plants d‟oliviers expriment
le plus haut degré de dépendance vis-à-vis du complexe mycorhizien de Béchar (CMB), ils

117
répondent moyennement à la mycorhization par I2 et CMS (avec des DM de 50.57% et
34.18% respectivement), et faiblement vis-à-vis de I1 (-4.32 %).
Les importants degrés de dépendance mycorhizienne des plants d‟oliviers inoculés par
CMB et CMS sont dus à leurs taux de mycorhizations qui sont relativement élevés (58% et
30 % respectivement), en revanche, le faible degré de dépendance des plants inoculés par
I1 est du à sont taux de mycorhization qui est presque nul (F= 8%).
La dépendance mycorhizienne dans notre expérience est déterminée par l‟inoculum
fongique utilisé et non par la teneur du sol en P (Ba et al., 2001 ; Fortin et al., 2008) puisque
le sol ainsi que l‟espèce végétale utilisés sont homogènes pour les quatre séries
d‟inoculation. Nos résultats confirment donc que le complexe mycorhizien CMB assure une
meilleure alimentation minérale (surtout le P) pour une bonne croissance des plants.
Ces résultats rejoignent ceux de Jaizme-Vega et al. (2002) qui ont rapporté que le
bananier montre une plus grande dépendance vis-à-vis de Glomus intraradices (50%) que Gl.
manihotis ((3355%%))..
Fig. 94 : Estimation de la hauteur des boutures d’olivier mycorhizées
0
5
10
15
20
25(cm) Hauteur de la partie aérienne
témoin
I1
I2
CMS
CMB

118
Fig.95: Estimation de la longueur des racines des boutures d’olivier mycorhizées
Fig. 96 : Estimation du nombre de tiges des boutures d’olivier mycorhizées
0
5
10
15
20
25
(cm)Longueur moyenne des racines des plants
d'olivier
Témoin
I1
I2
CMB
CMS
0
0,5
1
1,5
2
2,5
Nombre moyen des tiges
Témoin
I1
I2
CMS
CMB

119
Fig. 97 : Dénombrement des feuilles des boutures d’olivier mycorhizeés.
Fig. 98 : estimation des poids frais des parties aériennes des boutures d’olivier
mycorhizeés.
0
5
10
15
20
25
30Nombre moyen des feuilles
Témoin
I1
I2
CMS
CMB
0
1
2
3
4
5
6
7
8
9(g)
Poid frais moyen des plants d'olivier
Témoin
I1
I2
CMS
CMB

120
Fig. 99 : Estimation des poids sec des parties aériennes des boutures d’olivier
mycorhizeés
Fig. 100: degré de dépendance mycorhizienne des plants d’olivier dans les quatre
séries d’inoculation
0
1
2
3
4
5
6
(g)Poids sec moyen des plants d'olivier
Témoin
I1
I2
CMS
CMB
-4,32
50,57
34,18
72,9
-10
0
10
20
30
40
50
60
70
80
I1 I2 CMS CMB
(%)

121
6.4. Test de ré-isolement des spores
L‟examen microscopique du tamisat du substrat de culture des séries d‟inoculation des
boutures d‟oliviers, monté entre lame et lamelle dans une goutte de polyvinyl- lactoglycérol, a
permis de certifier d‟une part, la pureté des isolats I1 (fig. 101, A) et I2 (fig. 101, B), et
d‟autre part, une diversité de spores (fig. 101, C et D) et des sporocarpes (fig. 101, E) qui ont
une morphologie analogue à ceux isolés du sol des deux stations d‟étude.
7. La culture axénique des CMA :
Les essais de culture des CMA in vitro ont été réalisés à partir de fragments de racines
de poireau mycorhizées ou des spores.
7.1. Culture des CMA à partir de racines de poireau mycorhizées .
Des fragments racinaires de poireau mycorhizés et désinfectés sont aseptiquement
déposés sur de l‟eau gélosée puis incubés à 27°C pendant plusieurs mois.
Ces fragments ont été obtenus à partir de cultures « monosporales » de poireau dont les
plants ont été inoculés par des spores morphologiquement identiques, isolées de la
rhizosphère de l‟olivier.
Après 24 h d‟incubation, des hyphes se développent autour des fragments racinaires du
poireau (fig.102) montrant ainsi que des formes intra-racinaires (vésicules, hyphes ou
probablement des spores intra-racinaires) sont à l‟origine de ce développement mycélien. Ces
résultats montrent qu‟un CMA peut se développer en l‟absence de l‟hôte comme l‟ont
rapportés Giovannetti et al., (1993) pour Gl. mosseae dont la vitesse de croissance est de 1.65
et 2.7 µm /min.
Des résultats analogues ont été obtenus par plusieurs auteurs (Diop, et al., 1994; Strullu
et al.,1997 ; Diop, 2003) expliquant que ce développement serait dû à la richesse des formes
intra-racinaires en contenu lipidique qui supporterait cette croissance sans tenir compte de la
diminution de l‟activité métabolique par les processus de désinfection et d‟excision des
racines. Au cours de l‟incubation, les cellules racinaires se dégradent progressivement, les
hyphes extra-matriciels croissent, leur densité augmente pendant plusieurs semaines et les
filaments se ramifient et tapissent les boîtes.
Après 48h de culture des fragments racinaires, le mycélium extra-racinaire s‟étend sur
l‟ensemble du milieu ; il ressemble à celui décrit par Friese et Allen (1991), Bago et al
(1998), Bago et Cano (2005) et Cano et al. (2008) puisqu‟il forme un réseau d‟hyphes très

122
ramifiés dans les cultures ; des hyphes plus fins et très ramifiés se différencient
probablement en « BAS » puis produisent les VLS (fig. 103 A) et plus tard des spores.
Les VLS sont hyalines, globulaires de 6.4 à 19.2µm de diamètre , terminales ou intercalaires, portées par un thalle (fig. 103 B). Des structures similaires de 20 à 100 µm de
diamètre ont été décrites chez les Gloméracées (Dalpé et al. 2005b).
Après 10 jours d‟incubation, les fragments racinaires produisent des spores brunes
d‟abord à l‟intérieur ( fig. 104 A) puis elles sont ensuite émises à l‟extérieur des racines dans
le milieu de culture ( fig. 104B). Les spores produites sont isolées, globulaires à
subglobulaires, de 153.6 à 217.6 (en moyenne 179.2 µm) de diamètre, de couleur marron
claire à un stade jeune à brun foncé (40/80/100/10 ) à stade adulte, elles sont issues d‟un
hyphe sporogène.
Des résultats analogues ont été obtenus par Diop et al., (1994) qui ont montré que les
racines excisées inoculées par Gl. versiforme et Gl. intraradices développent sur l‟eau gélosée
des structures extra-racinaires extensives jusqu‟à la sporulation sans la présence d‟un hôte
vivant pendant trois mois. La présence des signaux de l‟hôte végétal ne sont pas nécessaires
lors de cette phase (Solaiman et Abbott, 2004), ce qui confirme que la qualité ainsi que la
quantité des exsudats racinaires sont impliqués dans l‟élongation des hyphes (Carr et al.,
1985; Elias et Safir,1987).
Pour mettre en évidence les structures produites par le mycélium extra-racinaire et non
discernables par examen microscopique direct de la culture en boîte de Pétri, nous avons
inondé pendant une nuit la surface du milieu avec une solution de bleu de trypan à 0.1%
dans de l‟acide lactique puis éliminé l‟excès de colorant par plusieurs lavages à l‟eau
courante.
Cette coloration, souvent utilisée pour mettre en évidence le mycélium extraracinaire
et les BAS (Bago et al. 1999 ; Cano et al., 2008), nous a permis d‟observer différentes
structures (Fig.105) : des hyphes extraracinaires non septés (fig.107), cylindriques de 2.5 à
3.8 µm de diamètre à partir duquel sortent des pédicelles portant des VLS ; les spores
apparaissent de couleur plus foncée. Le diamètre des hyphes extra-racinaires se rapprochent
de celui de G. intraradices (3.45 µm) en co-culture cependant, il peut être plus important
chez d‟autres CMA comme Gi margarita (6.02 µm) (Tiwari et Adholeya , 2002).

123
Nous avons aussi constaté la présence de sporocarpe de 230.4 µm de diamètre
(fig.107).
La formation des sporocarpes est caractéristique des Gloméracées et leur présence de
chez plusieurs espèces de CMA indique que la spore appartient au genre Glomus (Bi et al.,
2004).
Après un mois de culture, les spores perdent à leur tour leur forme initiale, elles
doublent de volume, germent à nouveau en émettant plusieurs tubes germinatifs (fig.108) qui
se différencient en pro-mycélium grâce aux réserves contenues dans les spores. Cependant, à
un certain stade de leur développement, ils se rétractent en raison de l‟absence de l‟hôte
végétal. L‟isolement des CMA à partir de racines mycorhizées permet d‟obtenir du
mycélium extra-racinaire mais pas une culture pure du champignon.
Selon certains auteurs, le nombre de tubes germinatifs émis à partir de l‟hyphe
suspenseur est variable chez les espèces de Glomus, il peut être un seul (Gl. mosseae et Gl.
caledonium) ou plusieurs ( Gl. clarum ) ou seulement un gonflement à l‟extrémité de l‟hyphe
suspenseur(Gl. viscosum ) (Walker et al. 1995).
Le pro-mycélium explore le milieu de culture et se différencie en « BAS » (fig. 109A
et B). Ces résultats confirment ceux de Dalpé (2005b), Bago et al. (1998) et Cano et al.(
2008).
Les BAS ne sont pas formés seulement par les Gloméracées, mais aussi par les
membres de la famille des Gigasporasporacées (de Souza et Declerck, 2004 ; Bago et Cano,
2005) et des Acaulosporacées (Dalpé et Declerck, 2002) ; ces structures se forment en milieu
axénique mais aussi dans le sol. Bago et al. (1998) a supposé qu‟en plus de leur rôle
physiologique dans l‟absorption des nutriments, ces structures seraient impliquées dans le
maintien de la structure du sol et la formation des agrégats.
Ces résultats affirment que les racines mycorhizées accomplissent leur cycle par le
mycélium extra matriciel et par la formation des spores qui à leur tour germent pour initier
une nouvelle association avec des racines vivantes.
Synthèse mycorhizienne entre mycorhizes en culture et le système racinaire de la
tomate
Nous avons réalisé une culture monoxénique en associant in vitro des fragments
racinaires de poireau mycorhizés, issus d‟une culture sur eau gélosée, avec le système
VLS

124
racinaire de plantules de tomate (issus de la germination de graines en conditions axéniques).
L‟association est réalisée en boîtes de Petri sur milieu d‟association Crush et Hay (1981).
Après environ une semaine de culture, les fragments racinaires de poireau mycorhizés
produisent des hyphes qui se développent et se dirigent vers des racines secondaires de la
tomate, ils se ramifient énormément et forment les VLS (planche 11, fig. A).
Les VLS sont similaires à celles produites sur l‟eau gélosée (planche 11, fig. B), ces
hyphes se développent sur la surface du milieu en boîte de Pétri (planche11, fig. C) et entrent
en contact avec les racines de la tomate (planche 11, fig. D) et forment des appressoriums qui
apparaissent sous forme de très petites taches brunes ou noires (planche 11, fig. E). Ces
résultats concordent avec ceux de Dalpé et al. (2005b) qui ont montré que la présence de la
racine d‟une plante hôte stimule le développement du mycélium extra-racinaire. Diverses
cultures des Gloméracées, incluant Gl. caledonium (Karandashov et al. 1999), Gl. clarum (de
Souza et Berbara, 1999), Gl. fistulosum (Nuutila et al., 1995) et Gl. intraradices (Chabot et
al.,1992), produisent les VLS, elles se différencient en 2 à 8 jours après le contact avec les
racines.
Des spores sont aussi produites par le mycélium extraracinaire sur le milieu
d‟association (planche 11, fig. F) ; leur couleur diffère de celles produites en eau gélosée.
Elles apparaissent globulaires et crémeuses (planche 11, fig. G).
Des observations analogues ont été décrites par de nombreux auteurs qui montré qu‟en
culture monoxénique, les spores produites sont moins pigmentées et plus petites et que celle
produites dans le sol (Pawlowska et al., 1999 ; Dalpé, 2001).
Elles subissent une réduction importante de l‟épaisseur de leur paroi, néanmoins, elles
sont nettement plus infectieuses même plus efficaces dans la colonisation et la croissance des
plants micropropagés cultivés en serre (Vimard et al., 1999; Filion et al., 2001; Declerck et
al., 2002 ; Jaizme-Vega et al., 2003). D‟ailleurs, des plants inoculés avec ces spores et
cultivées sur du sol donneront naissance à une première génération de spores à paroi épaisse.
Un tel résultat indique une grande plasticité des CMA qui s‟adaptent rapidement aux
changements de l‟environnement tout en préservant l‟intégrité de leurs colonies fongiques
(Bago et al. 2004 ; Bago et Cano, 2005).
Les synthèses in vitro entre mycorhizes en culture et le système racinaire des plantules
de tomate réalisées nous ont permis de suivre trois stades de développement d‟une espèce de
CMA comme l‟ont décrits Bago et al. (1998): la prolifération des hyphes, le développement

125
des BAS le long des hyphes et la formation des spores dans les espaces colonisés par les
hyphes et les BAS.
7.2. Essais de cultures axéniques de CMA à partir de spores
Les spores utilisées dans cette expérimentation ont été isolées de la rhizosphère de
l‟olivier de la région de Béchar puis piégées dans une culture d’Allium porrum, les spores
ainsi multipliées ont été ré- isolées.
7.2.1. Caractéristiques des spores
Ces spores sont globulaires à sub-globulaires (96 µm), isolées ou en deux unités, de
couleur marron foncé, attachées à un hyphe suspenseur (fig.110A). Leur paroi est constituée
de trois couches, la couche supérieure de 0.7µm, la deuxième unitaire de 4 µm d‟épaisseur et
de couleur marron foncé, suivie d‟une couche laminaire de 2.56 µm.
L‟hyphe suspenseur est jaune pale, droit, cylindrique (11.5 µm), sa paroi est composée
d‟une seule couche de la même couleur de celle du suspenseur (fig.110B).
Les caractéristiques morphologiques des spores correspondent à celles du genre
Glomus décrit par certains auteurs (Walker ,1983 ; Stürmer et Morton, 1997 ; Błaszkowski et
Czerniawska, 2008) et plus particulièrement à l‟espèce Gl . multicaulis décrite par Panna et
Highland (2010).
Essai de culture axénique de CMA à partir de spores en germination
Les spores isolées sont mises à germer dans de l‟eau gélosée. Après 4 jours de culture
elles augmentent trois fois leur volume et émettent plusieurs tubes germinatifs (fig.112).
Le taux de leur germination est de 60%. Les 40% de spores qui n‟ont pas germé, sont
probablement mortes ou en phase de dormance. Ce phénomène de dormance est connue chez
diverses espèces des genres Glomus (Juge et al., 2002) et Acaulospora (Tommerup, 1983;
Douds et Schenck, 1991 ; Gazey et al. 1993).
Les tubes germinatifs, émis par les spores, s‟arrêtent de croître après 15 jours culture.
Nos résultats rejoignent ceux de certains travaux qui ont montré que cet arrêt de croissance a
lieu après 8 à 20 jours de culture (Giovanetti et al. 1993 ; Logi et al. 1998). Selon ces mêmes
auteurs, cet arrêt est dû au rétrécissement du protoplasme de l‟apex hyphal qui est séquestré
par des cloisons répétées dans l‟hyphe vide. Cette septation s‟accompagne d‟une

126
vacuolisation intense et d‟une rétraction du cytosol, des noyaux et des mitochondries vers la
spore. La rétraction des septa sépare les segments des hyphes viables des autres vides (non
viables) (Logi et al., 1998). Pour d‟autres auteurs, la croissance s‟arrête en l‟absence d‟un
signal ou d‟un contact de l‟hôte (Bécard et Piché, 1989 ; Giovannetti et al., 2000 ;Giovannetti
et al., 2010).
Pour établir l‟association mycorhizienne, plusieurs essais de germination de spores
doivent être tentés (Dalpé et al., 2005b).
7.2.1.2. Essai d’association mycorhizienne entre une spore de CMA en germination et le
système racinaire des plantules de tomate.
L‟association in vitro entre une spore de CMA en germination et le système racinaire
des plantules de tomate est réalisée sur le milieu de Crush et Hay (1981).
Nous avons constaté qu‟une spore en germination émet plusieurs tubes germinatifs
dans toutes les directions (fig. 112 A et B). L‟élongation des tubes germinatifs donne
naissance à un réseau mycélien lâche dont la croissance est très faible et variable selon les
espèces appartenant ou non au même genre. (Louis et Lim, 1988 ; Bécard et Piché, 1989 ;
Gianinazzi-Pearson et al., 1989 ; Douds et al., 1996 ; Logi et al. 1998).
Les tubes germinatifs issus de la spore se différencient ensuite en un mycélium pré-
symbiotique ; ce dernier croît et explore le milieu dans la direction de la racine comme l‟ont
montré les travaux de Declerck et al., (2000).
7.2.2. Synthèse mycorhizienne entre une spore et les racines de plantules de tomate
Après trois mois de culture, nous avons observé la formation de nombreuses spores
jeunes et translucides devenant noires à maturité (fig.113 A et B). Leur isolement et leur
réassociation avec des plantules de tomate, sur milieu MRS a donné naissance à un mycélium
extra-racinaire (fig. 114) qui produit des spores se développant sur les systèmes racinaires de
la tomate (fig.115), certaines sont soit isolées sur le milieu de culture ou regroupées en
sporocarpe (116).
Pour étudier leurs caractéristiques, les spores sont prélevées à partir du milieu de
culture et montées entre lame et lamelle dans une goutte de PVLG ou de PVLG+ Melzer.
S

127
Les spores issues du mycélium extra-racinaire possèdent des caractéristiques
morphologiquement similaires à celles dont est issu le mycélium extra-racinaire. Elles sont
sub-globulaires (fig.117 E), aux contours irréguliers, translucides à l‟état jeune (32µm de
diamètre) et noires à maturité (217.6×240 µm); elles sont reliées à la racine (fig.117) par le
suspenseur (6.4 µm). Leur surface est rugueuse, cependant leur dimension réduite et leur
couleur sombre ne nous ont pas permis de distinguer les couches de la paroi sporale.
Quant aux sporocarpes, ils apparaissent après 4 mois de culture, ils sont sans péridium,
moins volumineux (140 µm) et portent plusieurs spores (de 57.6 µm de diamètre). (fig. 116).
Les spores semblent contenir des petites structures intra-sporales ovalaires (7.68×5.12
µm de diamètre) qu‟elles excrètent après la rupture de la paroi (fig.118).
caractéristiques morphologiques de la spore (forme, le type et mode du suspenseur )
correspondent au genre Glomus. Elles rejoignent d‟ailleurs les descriptions de ce genre
rapportés par de nombreux auteurs (Berch, 1988; Schenck et Pérez, 1990 ; Blaszkowski et
Czerniawska ,2008)
Mise en évidence de l’infection du système racinaire des plantules de tomate
Les examens microscopiques des racines de plantules de tomate en culture axénique et
colorées par le bleu trypan (Phillips et Hayman, 1970) révèlent l‟omniprésence du mycélium
extra-racinaire sur la surface des racines (fig. 119) et des spores (figs. 120). On observe à
l‟intérieur des cellules corticales des VLS attachées au mycélium intra-racinaire (fig.121).
Les subcultures des CMA sur le milieu MSR modifié, permettent donc d'avoir une
production régulière d'inoculum et surtout de maintenir in vitro des souches de CMA
indigènes. Il restera à vérifier si une souche conserve ou non son potentiel infectieux au cours
de multiples repiquages.
Conclusion :
Dans ce travail, nous nous sommes proposé d‟étudier les CMA des sols d‟oliviers et
leur association mycorhizienne avec la variété Sigoise d‟olivier dans le but de sélectionner des
CMA pour la mycorhization des boutures herbacées produites en pépinière.
RT
A
S

128
Cette étude a porté sur la diversité des champignons mycorhiziens à arbuscules (CMA)
associés à la variété Sigoise d‟olivier (Olea europea L.), cultivée dans la région de Sig et à
Béchar.
Les données climatiques (température et pluviométrie) des deux stations d‟étude ont
révélé qu‟elles sont caractérisées par des températures moyennes extrêmement élevées lors de
la période estivale (28° C et 35.4 pour les stations Sig et Béchar respectivement) et assez
clémentes durant la période hivernale (7.9° C et 8.8° C pour les stations Sig et Béchar
respectivement), des pluviométries moyenne de l‟ordre de 2 à 82 mm pour la station de Sig
(durant les mois d‟août et de décembre respectivement), de 23 mm (février) et de 0 à 3mm le
reste de l‟année pour la station de Béchar.
Les données pédologiques ont montré que les sols des vergers de Sig sont sablo-
limoneux, calcaires, alcalins, pauvres en sel, en azote, en phosphore et en matière organique.
Les examens microscopiques des fragments racinaires naturels d‟oliviers ont révélé la
présence de mycorhizes arbusculaires de type « Arum » à des taux très élevés (82% pour les
oliviers âgés de 5 ans, et de 100% pour les autres oliviers) indépendamment de l‟âge et de la
saison de prélèvement des échantillons confirmant ainsi le caractère mycotrophique de cette
variété d‟olivier. Ces champignons forment des structures caractéristiques des MA :
arbuscules, vésicules, enroulements et spores intra-racinaires. Un mycélium septé a été
observée (≤5%).
Les caractéristiques morphologiques des spores du sol rhizosphérique de la variété
Sigoise de l‟olivier ont révélé la prédominance des Gloméracées et la présence de quelques
espèces appartenant aux genres Acaulospora et Scutellospora. Deux espèces de CMA
appartenant au genre Glomus se rapprochant par leurs caractéristiques morphologiques de
Glomus macrocarpum et Gl. muticaulis.
L‟étude du pouvoir endomycorhizogène (PEM) du sol des deux parcelles d‟olivier de
Sig révèle la richesse de ce sol en propagules infectieuses des CMA. Le nombre la plus
probable de propagules (parcelle 1 : NPP= 5500 propagules par Kg de sol et 7000 pour la
parcelle 2).
L‟utilisation des plants de poireau, comme plante-test pour piéger les CMA des sols
rhizosphériques des oliviers de Sig, a montré que cette plante est hautement mycotrophe.
Nous avons constaté que l‟infection apparait dès la deuxième semaine de culture, la

129
production des arbuscules débute à partir de la 5ème semaine et celle des vésicules (à des taux
très faibles) s‟effectue une semaine après la formation des arbuscules. La présence du
mycélium septé a aussi été observée, à une fréquence de 40 %. Les résultats révèlent que les
complexes mycorhiziens indigènes des sols favorisent la croissance aérienne des plants de
poireau mycorhizés (parcelle 1 : 110% parcelle 2 ; 130%), avec une dépendance
mycorhizienne des plants de 52%.
L‟inoculation des boutures herbacées d‟olivier par des isolats de CMA (isolat I1 ou
I2 préalablement sélectionné et multiplié sur culture de poireau) ou des complexes
mycorhiziens indigènes du sol des vergers de Sig (CMS et CMB) a montré ses effets
favorables sur la croissance des plants qui forment des MA analogues aux mycorhizes
naturelles.
L‟examen microscopique du tamisat du substrat de culture des boutures herbacées
d‟oliviers mycorhizéés a permis de contrôler que les inoculums utilisés étaient bien à l‟origine
de ces MA. Des inoculations régulières de nouveaux plants de poireau avec un inoculum
sélectionné et leur culture en pots ouverts sur un substrat de culture permettront de conserver
les isolats de CMA les plus efficaces.
Les essais de culture axénique des CMA , après association in vitro des fragments
racinaires mycorhizés de poireau ou de spores de CMA à des racines de tomate, nous ont
permis de suivre l‟infection racinaire, le développement du mycélium extra-racinaire et des
structures qu‟il produit : spores, sporocarpes, VLS et BAS. Des subcultures des CMA, sur
un milieu d‟association bien défini, permettent donc de produire régulièrement l'inoculum et
surtout de maintenir in vitro des souches de CMA sélectionnées. Cependant, il est toujours
indispensable vérifier si la souche conserve ou non son potentiel infectieux au cours de
multiples repiquages.
Ainsi, notre travail a permis de compléter le travail du magister de SAAD (2009)
par : la description des principaux morphotypes de CMA de la rhizosphère des sols de la
variété sigoise d‟olivier, l‟identification des deux espèces de Glomus, la caractérisation de
deux isolats de CMA et leur efficacité sur la croissance des boutures herbacées d‟olivier, la
conservation des CMA sélectionnés sur culture de poireau ou en culture axénique pour leur
utilisation ultérieure en pépinière.

130
En perspectives, il serait également intéressant de poursuivre ce travail :
- en complétant la caractérisation les deux espèces de Glomus isolées par des
techniques de Biologie moléculaire ;
- en isolant et caractérisant d‟autres espèces de CMA de la rhizosphère des vergers
d‟oliviers des régions arides et semi-arides ;
- et enfin, en inoculant des boutures herbacées d‟oliviers ou des vitroplants
d‟oliviers avec les CMA sélectionnées et suivre leur développement en pépinière
et au champ.
Références bibliographiques :
Abbas Y., Ducousso M., Abourouh M., Azcon R., Duponois R., 2006. Diversity of arbuscular mycorrhizal fungi in Tetraclinis articulate (Vahl) Masters woodlands in Morocco. Ann. For.Sci., 63: 285-291. Abbott L. K., 1982. Comparative anatomy of vesicular-arbuscular mycorrhizas formed on subterranean cover. Aust. J. Bot., 30:485–490.
Abbott L. K., Robson A.D., Gazey C., 1992. Selection of inoculants vesicular arbuscular mycorrhizal fungi. In: Norris JR, Read DJ, Varma AK (eds) Methods in microbiology: experiments with mycorrhizas (vol. 24). Academic, London, pp. 1–121. Abdul Hussain K. H., Abdul Hussain M. S., 2004. Influence of the gibberelic acid on the germination of the seeds of olive-tree Olea europea L. J. Eur.Agr., 5 (1):1-4 Adelman M. J., Morton J. B., 1986. Infectivity of vesicular-arbuscular mycorrhizal fungi: influence of host–soil diluent combinations on MPN estimates and percentage colonization. Soil Biol. Biochem., 18:77–83 Adholeya A., Tiwari P., Singh R., 2005 .Large-Scale Inoculum Production of Arbuscular Mycorrhizal Fungi on Root Organs and Inoculation Strategies. In Vitro Culture of Mycorrhizas (ed. by S. Declerck, D.-G. Strullu, and A. Fortin). Soil Biol., 4: pp.315-338.
Afek U., Rinaldelli E., Menge J.A., Johson E.L.V., Pont E., 1990 . Mycorrhizal species, root age, and position of mycorrhizal inoculum influence colonization of cotton, onion, and pepper seedlings. Amer. J. Soc. Hort. Sci., 115(6): 938-942. Ahmed F.R.S., Killham K., Alexander I., 2006 Influences of arbuscular mycorrhizal fungus Glomus mosseae on growth and nutrition of lentil irrigated with arsenic contaminated water. Plant Soil, 258:33–41. Ahulu E. M., Nakata M., Nonaka M., 2005. Arum- and Paris-type arbuscular mycorrhizas in a mixed pineforest on sand dune soil in Niigata Prefecture, central Honshu. Japan Mycorrhiza,15: 129–136.

131
Akiyama, K., 2007. Chemical identification and functional analysis of apocarotenoids involved in the development of arbuscular mycorrhizal symbiosis. Bioscience Biotec. Biochem.,71(6): 1405-1414.
Akiyama K., Hayashi H., 2006. Strigolactones: Chemical signals for fungal symbionts and parasitic weeds in plant roots. Ann. Bot. Lond., 97: 925-931.
Akiyama K., Matsuoka H., Hayashi H., 2002. Isolation and identification of a phosphate deficiency-induced C-glycosylflavonoid that stimulates arbuscular mycorrhiza formation in melon roots. Mol. Plant Microbe Inter., 15(4): 334-40.
Akiyama K., Matsuzaki K., Hayashi H., 2005. Plant sesquiterpenes induce hyphal branching in arbuscular mycorrhizal fungi. Nature, 435: 824-827.
Alexander M., 1965. Most- probable-number method for microbial populations. In: Methods of soil analysis part 2, chemical and microbiological properties. (Ed.) Black C.A. Amer. Soc. Agron. Madison, Wis. Part 2, pp. 1467-1472.
Alexander T., Meier R., Toth R., Weber, H.C., 1988. Dynamics of arbuscule development and degeneration in mycorrhizas of Triticum aestivum L. and Avena sativa L. with reference to Zea mays L. New Phytol. 110: 363–370.
Alexander T., Toth R., Meier R., Weber H.C., 1989. Dynamics of arbuscule development and degeneration in onion, bean and tomato with reference to vesicular-arbuscular mycorrhizae in grasses. Can. J. Bot. 67, 2505–2513.
Alguacil M. M., Roldan A., Torres M. P., 2009. Complexity of semiarid Gypsophilous Shrub Communities Mediates the AMF Biodiversity at the Plant Species Level. Microbiol. Ecol., 57: 718-727.
Al-Karaki G., 2004. Field response of wheat to arbuscular mycorrhizal fungi and drought stress. Mycorrhiza, 14: 263-269.
Allen J.W., Shachar-Hill Y., 2009. Sulfur transfer through an arbuscular mycorrhiza. Plant Physiol., 149: 549-560.
Almeida R. T., Schenck N. C., 1990. A revision of the genus Sclerocystis (Glomaceae, Glomales). Mycologia, 82: 703-714.
Amijee F., Tinker P.B., Stribley D.P., 1989. The development of endomycorrhizal root systems. 7. A detailed study of effects of soil-phosphorus on colonization. New Phytol., 111: 435-445.
Appelhans M., Weber C. H.. Imhof S., 2008. Rutaceae sampled from Germany, Malta, and Mallorca (Spain) are associated with AMF clustering with Glomus hoi Berch & Trappe Mycorrhiza, 18:263–268.
Argenson C., 2008. La culture de l‟olivier dans le monde, ses productions, les tendances. Le
Nouvel Olivier, 61: 8-11.
Arihara J., Karasawa T., 2000. Effect of previous crops on arbuscular mycorrhizal formation and growth of succeeding maize. Soil Sci. Plant Nutr., 46:43–51.
Augé R., 2001. Water relations, drought and vesicular arbuscular mycorrhizal symbiosis .Mycorrhiza, 11:3-42.

132
Augé R. M., Sylvia D. M., Park S., Buttery B.R., Saxton A., M., Moore J. L., Cho K., 2004. Partitioning mycorrhizal influence on water relations of Phaseolus vulgaris into soil and plant components. Can. J. Bot., 82:503-514.
Aversenq S., Gratraud C., Pinatel C., 2005. Ravageurs et auxiliaires des oliviers. Phytoma : 32-36.
Azcón-Aguilar C., Barea J. M., 1996. Arbuscular mycorrhizas and biological control of soil-borne plant pathogens - An overview of the mechanisms involved. Mycorrhiza, 6: 457-464. Azcón-Aguilar C., Barea J. M., 1997. Applying mycorrhiza biotechnology to horticulture, significance and potentials. Scientia Horticult., 68: 1-24.
Azcón-Aguilar, C., Jaizme-Vega, M.C., Calvet, C., 2002. The contribution of arbuscular mycorrhizal fungi to the control of soil-borne plant pathogens. In Mycorrhizal Technology in Agriculture: From Genes to Bioproducts., S. Gianinazzi, H. Schuepp, K. Haselwandter, and J.M. Barea, eds (Basel: ALS Birkhauser Verlag, Germany), pp. 187-197.
Azcón-Aguilar C., Palenzuela J., Roldán A., Bautista S., Vallejo R., Barea J.M., 2003. Analysis of the mycorrhizal potential in the rhizosphere of representative plant species from desertification-threatened Mediterranean shrublands. Appl. Soil Ecol., 22: 29-37.
Bâ A., Guisson T., Duponnois R., Plenchette C., Sacko O., Sidibé D., Sylla K., Windou B., 2001. Mycorhization contrôlée et fertilisation phosphatée: Applications à la domestication du jujubier. Fruits, 56(4): 261-269
Babana A. H., Antoun H., 2006. Effect of Tilemsi phosphate rock-solubilizing microorganisms on phosphorus uptake and yield of field-grown wheat (Triticum aestivum L.) in Mali. Plant Soil, 287:51–58. Bago B., Cano C., 2005. Breaking Myths on Arbuscular Mycorrhizas in Vitro Biology. In Vitro Culture of Mycorrhizas.(ed. by S. Declerck, D.-G. Strullu, and A. Fortin). Soil Biol., 4:111-138.
Bago B., Zipfel W., Williams R. M., Piché Y., 1999. Nuclei of symbiotic arbuscular mycorrhizal fungi as revealed by in vivo two-photon microscopy. Protoplasma, 209:77-89.
Bago B., Cano C., Samson J., Coughlan A. P., Piché Y., Azcón-Aguilar C., 2004. Differential morphogenesis of the extraradical mycelium of an arbuscular mycorrhizal fungus grown monoxenically on spatially heterogeneous culture media. Mycologia, 96:452–462.
Bago B., Zipfel W., Williams R. M., Chamberland H., Lafontaine J.G., Webb W.W., Piché Y., 1998. In vivo studies on the nuclear behavior of the arbuscular mycorrhizal fungus Gigaspora rosea grown under axenic conditions. Protoplasma, 203:1–15.
Balestrini R., Lanfranco L., 2006. Fungal and plant gene expression in arbuscular mycorrhizal symbiosis. Rev. Mycorrhiza,16: 509-524.
Barea, J. M., 2000. Rhizosphere and mycorrhiza of field crops. In J. P. Toutant, E. Balazs, E. Galante, J.M. Lynch, J. S. Schepers, D. Werner and P. A. Werry (Eds.), Biological Resource Management, Connecting Science and Policy (OECD) Heidelberg, Germany: INRA, Editions and Springer, pp. 110-125. Barea J. M., Azcón R., Azcón-Aguilar C., 2002. Mycorrhizosphere interactions to improve plant fitness and soil quality. Antoine van Leeuwenhoek, 81:343-351.

133
BB aarree aa JJ..MM..,, PPéé rree zz SS.. EE..,, DDeell VVaall CC..,, AAzzccóónn--AAgguuiillaarr CC..,, 11999999.. IImmppoorrttaanncciiaa ddee llaass mmiiccoorrrriizzaass eenn
eelleessttaabblleecciimmiieennttoo yy pprrootteecccciióónn ddee llaass ppllaannttaass eenn ssuueellooss ddee--ggrraaddaaddooss.. PPhhyyttoommaa..,, 111111::1188--3300
Barea J. M., Werner D., Azcon-Guilar C., Azcon R., 2005. Interactions of arbuscular mycorrhiza and nitrogen-fixing symbiosis in sutainable agriculture. In: Nitrogen Fixation in Agriculture. Eds. Werner D., Newton W. E. Forestry, Eco. Environ.: 199-222.
Bartolozzi F., Mencuccini M., Fontanazza G., 2001. Induction of frost tolerance in olive plants in vitro by cold acclimation and sucrose increase. Plant Cell., Tissue and Organ Culture, 67 (3): 299-302.
Beck-Nielson D., Vindaek M., 2001. Occurrence of vesicular arbuscular mycorrhiza in aquatic miccrophytes from lakes and streams. Aqua. Bot., 71:141-148.
Bécard G., Douds D. D., Pfeffer P. E., 1992. Extensive invitro hyphal growth of vesicular-arbuscular mycorrhizal fungiin the presence of CO2 and flavonols. Appl. Environ. Microbiol., 58: 821-825. Bécard, G., Kosuta S., Tamasloukht M., Séjalon-Delmas N., Roux C., 2004. Partner communication in the arbuscular mycorrhizal interaction. Can. J. Bot., 82 (8): 1186-1197.
Bécard G., Piché Y., 1989. Fungal growth stimulation by CO2 and root exudates in vesiculararbuscular mycorrhizal symbiosis. Appl. Environ. Microbiol., 55:2320–2325.
Bécard G., Roux C., Sejalon-Delmas N., 2005. Modulators of the development of mycorrhizal fungi with arbuscules and uses thereof, Patent W, Editor. , Berlin, pp. 355-370.
Becker W., Gerdemann J., 1977. Colorimetric quantification of vesicular–arbuscular mycorrhizal infection in onion. New Phytol 78:289–295
Beddiar A., 1984. Les possibilités d‟associations symbiotiques de l‟Aulne glutineux dans divers sols de l‟Est de France. Thèse de doctorat, Université de Nancy, France. 200p. Bellahcene M., 2004. La verticilliose de l‟olivier: étude épidémiologique et diversité génétique de Verticillium dahliae Kleb., agent de la verticilliose. Thèse de doctorat d‟état es Science, Université d‟Oran, Algérie, 145 p.
Bellahcene M., Fortas Z., Fernandez D., Nicole M., 2005b. Vegetative compatibility of Verticillium dahlia isolated from olive trees ( Olea europea L.) in Algeria. Afric. J. Biotechn., 4(9): 963-967. Bellahcene M., Assigbetsé K., Fortas Z., Geiger J.P., Nicole M., Fernandez D., 2005a. Genetic diversity of Verticillium dahliae isolates from olive trees in Algeria. Phytopathol. Mediterr., 44: 566-274.
Benarar D., Bouguedoura N., 2003. Essai de germination de l‟olivier de Laperrine (Olea Laperrini
Batt. et Tr.). La forêt Algérienne, 5: 16-18.
Benderradji L., Bouzerzour H., Ykhle f N., Djekoun A., Kellou K., 2007. Réponse à la culture in vitro de trois variétés de l‟olivier (Olea europaea L.). Sci. Technol., 26 :27-32
Berch S., 1988. Compilation of the Endogonacae.Mycologue, Waterloo, Canada, 228p.
Berch S. M., Fortin J.A., 1983. Lectotypification of Glomus macrocarpum and proposal of new combination: Glomus austral, Glomus versiforme, and Glomus tenebrum (Endogonaceae). Can. J. Bot., 61: 2608-2617.

134
Berta, G., Trotta, A., Fusconi, A., Hooker, J.E., Munro, M., Atkinson, D., Giovannetti, M., Morini, S., Fortuna, P., Tisserant, B., Gianinazzi-Pearson, V., Gianinazzi, S., 1995. Arbuscular mycorrhizal induced changes to plant growth and root system morphology in Prunus cerasifera. Tree Physiol., 15: 281–293.
Bernard N., 1899. Sur la germination du Neotia nidus avis. Compte rendu Hebdomadaire des Séances de l‟Académie des sciences Paris, 128 : 1253-1255
Besnard G., Baradat P., Chevalier D., Tagmount A., Bervillé A., 2001. Genetic differentiation in the olive complex (Olea europaea L.) revealed by RAPDs and RFLPs in the rRNA genes. Gen. Res. Crop. Evol., 48 : 165-82.
Besserer A., Puech-Pages V., Kiefer P., Gomez-Roldan V,Jauneau A, Roy S, Portais JC, Roux C, Bécard G, Séjalon-Delmas N., 2006. Strigolactones stimulate arbuscular mycorrhizal fungi by activating mitochondria. PLoS Biol., 4: 1239–1247. Bever J.D., Schultz P.A., Pringle A., Morton J.B., 2001. Arbuscular mycorrhizal fungi: more diverse than meets the age, and the ecological tale of why. Bioscience, 51: 923- 931.
Bi Y. L., Li X. L., Christie P., Hu Z. Q., Wong M. H., 2003. Growth and nutrient uptake of arbuscular mycorrhizal maize in different depth of soil overlying coal fly ash. Chemosphere, 50:863-869. Bi Y., Li X., Wang H., Christie P., 2004. Establishment of monoxenic culture between the arbuscular mycorrhizal fungus Glomus sinuosum and Ri T-DNA-transformed carrot roots. Plant Soil, 261: 239-244.
Bianciotto V., Andreotti S., Balestrini R., Bonfante P., Perotto S., 2001 . Extracellular polysaccharides are involved in the attachment of Azospirillum brasilense and Rhizobium leguminosarum to arbuscular mycorrhizal structures. Eur. J. Histochem., 45: 39-49. Bianciotto V., Bonfante P., 1993. Evidence of DNA replication in an arbuscular mycorrhizal fungus in the absence of the host plant. Protoplasma, 176:100-105.
Binet M.N., Lemoine M.C., Martin C., Chambon C., Gianinazzi S., 2007. Micropropagation of olive (Olea europaea L.) and application of mycorrhiza to improve plantlet establishment. Dev. Biol. Plant, 43:473-478. Biryukova O. N., Orlov D. S., Biryukov M. V., 2008. Genesis and geographyof soils Humus Status of Buried Soils in Loess Deposits of the Minusinsk Intermontane Trough. Soil Science, 41 (5): 471–480 Blal B., Gianinazzi-Pearson V., Fardeau J.C., Gianinazzi S., 1990. Influence of vesicular–arbuscular mycorrhizae on phosphate fertilizer efficiency in two tropical acid soils planted with micropropagated oil palm (Elaeis guineensis Jacq.). Biol. Fert. Soil, 9:43–48
Blaszkowski J., 1988. Three new vesicular-arbuscular mycorrhizal fungi (Endogonaceae) from Poland. Bull. Pol. Ac. Sci. Biol. Sci., 36 : 10-12. Blaszkowski J., 1993. Scutellospora armeniaca, a new species in glomales (zygomycetes) from poland. Mycologia, 84(6):939-944.

135
Blaszkowski J. 2003. Arbuscular mycorrhizal fungi (Glomeromycota), Endogone and Complexipes species deposited in the Department of Plant Pathology, University of Agr iculture in Szczecin, Poland, 230p.
Blaszkowski J., Czerniawska B., 2006. The occurrence of arbuscular mycorrhizal fungi of the phylum Glomeromycota in Israeli soils. Acta Soc. Bot. Pol., 75: 339-350.
Blaszkowski J., Czerniawska B., 2008. Glomus eburneum and Scutellospora fulgida, species of arbuscular mycorrhizal fungi (Glomeromycota) new for Europe. Acta Mycol. 43 (1): 57–65. Blaszkowski J., Kowalczyk S., Czerniawska B., 2006 . Acaulospora rehmii and Gigaspora margarita, arbuscular mycorrhizal fungi (Glomeromycota) new for Europe and Poland, respectively. Acta Mycol., 41: 41-48.
Blaszkowski J., Blanke V., Renker C., Buscot F. 2004 . Glomus aurantium and G. xanthium, new species in Glomeromycota. Mycotaxon, 90: 447-467.
Blaszkowski J., Tadych M., Madej T., Adamska I., Iwaniuk A. 2001. Arbuscular mycorrhizal fungi (Glomales, Zygomycota) of Israeli soils. Mat. II Polsko-Izraelskiej Konf. Nauk. nt. „Gospodarowanie zasobami wodnymi i nawadnianie roslin uprawnych”. Przeglad naukowy Wydz. Inz. Ksztalt. Srod., 22: 8-27. Blazquez J. M., 1996. The origin and expansion of olive cultivation. In: International Olive Oil Council (ed) World Olive Encyclopedia. IOOC, Madrid, pp.19-20.
Blilou I., Bueno P., Ocampo J. A., Garcia-Garrido J., 2000. Induction of catalase and ascorbate peroxidase activities in tobacco roots inoculated with the arbuscular mycorrhizal Glomus mosseae. Mycol. Res., 104:722–725. Boddington C.L., Dodd J.C., 2000. The effect of agricultural practices on the development of indigenous arbuscular mycorrhizal fungi. I. Field studies in an Indonesian ultisol. Plant Soil, 218: 137-144.
Bonanomi G., Vinale F., Scala F., 2009. The Role of Natural Products in Plant-Microbe Interactions. A.E. Osbourn and V. Lanzotti (eds.), Plant-derived Natural Products, pp. 301-320.
Bonfante-Fasolo P., Grippioli R., 1982. Données actuelles sur la cytologie des mycorhizes. In: Les mycorhizes, partie intégrante de la plante: biologie et perspectives d‟utilisation. Coll. I.N.R.A., Dijon, France, 13: 25-36
Bonfante P., 2001. At the interface between mycorrhizal fungi and plants: the structural organization of cell wall, plasma membrane and cytoskeleton. In: Hock B (ed) The Mycota IX: fungal associations. Springer, Berlin Heidelberg . New York, pp. 45 – 61.
Bonfante P., Balestrini R., Genre A., Lanfranco L., 2009 . Establishment and Functioning of Arbuscular Mycorrhizas. The Mycota VH. Deising (Ed.) 2nd Edition. Plant Relationships , , pp. 259-274.
Bonfante P., Perotto S., 1995. Strategies of arbuscular mycorrhizal fungi when infecting host plants. New Phytol., 130: 3-21.

136
Bouchetata T.B., 2006. Analyse des agro-systèmes en zone tellienne et conception d‟une base de
données, Mascara, Algérie.-. Thèse Master of Science, IAMM. Montpellier, France, 170 p.
Boudarga K., Dexheimer J., 1989. Sur la mycorhization contrôlée de semis d‟Eucalyptus camaldulensis Dehnardt par Gigaspora margarita Becker et Hall. Ann. Sci. For., 46: 131-139. Bouwmeester H. J., 2008. Tomato strigolactones are derived from carotenoids and their biosynthesis is promoted by phosphate starvation . New Phytol., 178; 863 – 874.
Breton C., Médail F., Pinatel C., Bervillé A., 2006. De l‟olivier à l‟oléastre : origine et domestication de l‟Olea europaea L. dans le Bassin méditerranéen. Cahiers Agricult. , 15(4) :329-336.
Brhadda N., Loudyi D. E. W., Abousalim A., 2008. Effet du sucre sur l‟embryogenèse somatique de l‟olivier (Olea europea L.) cv. „Picholine marocaine‟. Biotechnol. Agron. Soc. Environ., 12(3) : 245-250. Brundrett M. C., 2004. Diversity and classification of mycorrhizal associations. Biol. Rev. 79: 473–495.
Brundrett M.C., 2009. Mycorrhizal associations and other means of nutrition of vascular plants: understnding global diversity of hostplants by resolving conflicting information and developingreliable means of diagnosis. Plant Soil, 320:37–77.
Brundrett M. C., Kendrick B., 1990. The roots and mycorrhizas of herbaceous woodland plants I. Quantitative aspects of morphology. New Phytol., 114:457–468.
Brundrett M.C., Piche Y., Peterson R.L., 1984. A new method for observing the morphology of vesicular-arbuscular mycorrhizae. Can. J. Bot., 62:2128–2134.
Brundrett M.C., Ashwath N., Jasper D.A., 1996. Mycorrhizas in the Kakadu region of tropical Australia. II. Propagules of mycorrhizal fungi in disturbed habitats. Plant Soil, 184: 173-184. Brundrett M.C., Abbott L.K., Jasper D.A., 1999. Glomalean fungi from tropical Australia. I. Comparison of the effectiveness of isolation procedures. Mycorrhiza, 8: 305-314.
Bruns T. D., Shefferson R. P., 2004. Evolutionary studies of ectomycorrhizal fungi: recent advances and future directions. Can. J. Bot., 82:1122–1132.
Bucher M., 2007. Functional biology of plant, Phosphate uptake at root and mycorrhiza interfaces. New Phytol., 173: 11-26.
Budi S.W., Blal B., Gianinazzi S., 1999. Surface-sterilization of Glomus mosseae sporocarps for studying endomycorrhization in vitro. Mycorrhiza, 9:65–68.
Buee , M. , Rossignol , M. , Jauneau , A. , Ranjeva , R., Becard G., 2000. The pre-symbiotic growth of arbuscular mycorrhizal fungi is induced by a branching factor partially purified from plant root exudates . Mol. Plant-Microbe Inter., 13 : 693 – 698.
BB uurrrroowwss RR.. LL..,, PPffllee ggee rr FF.. LL..,, 22000022.. AArrbbuussccuullaarr mmyyccoorrhhiizzaall ffuunnggii rreessppoonndd ttoo iinnccrreeaassiinngg ppllaanntt
ddiivveerrssiittyy.. CCaann.. JJ.. BBoott.. ,, 8800:: 112200-- 113300..
Cairney J. W. G., Burke R. M., 1998. Extracellular enzyme activities of the ericoid mycorrhizal endophyte Hymenoscyphus ericae (Read) Korf & Kernan: their likely roles in decomposition of dead plant tissue in soil. Plant Soil, 205: 181–1923.

137
Calvente R., Cano C., Ferrol N., Azcón-Aguilar C. Barea J.M., 2004. Analysing natural diversity of Arbuscular mycorrhizal fungi in olive tree (Olea europaea L.) plantations and assessment of the effectiveness of native fungal isolates as inoculants for commercial cultivars of olive plantlets. Appl. Soil Ecol., 26: 11-19.
Calvet C., Pera J., Estaun V., Camprub A., 1989. Vesicular-arbuscular mycorrhizae of kiwi fruits in an agricultural soil: inoculation of seedlings and hardwood cutting with Glomus mosseae. Agron., 9: 181-185 Camprubi A., Calvet C., Estaun V., 2000. Micorrizas arbusculares en produccion agricola. Horticult., 144 : 38-41
Cano C., Dickson S., González-Guerrero M., Bago A., 2008. In vitro Cultures Open New Prospects for Basic Research in Arbuscular Mycorrhizas. A. Varma (ed.). Mycorrhiza, pp. 627-654.
Cantrell I. C., Linderman R. G., 2001. Preinoculation of lettuce and onion with VA mycorrhizal fungi reduces deleterious effects of soil salinity. Plant Soil, 233: 269–281. Caravaca F., Alguacil M.M., Barea J.M., Roldán A., 2005. Survival of inocula and native AM
fungi species associated with shrubs in a degraded Mediterranean ecosystem. Soil Biology &
Biochemistry, 37: 227-233.
Cardoso I.M., Kuyper T.W., 2006. Mycorrhizas and tropical soil fertility. Agri. Ecosy. Envir., 116: 72-84.
Cardoso I., M., Boddington C., Janssen B. H., OenemaO., Kuyper T.W., 2006. Differential access to phosphorus pools of anOxisol by mycorrhizal and non-mycorrhizal maize. Soil Sci. plant Anal., 37: 1-15. Cardoso J. M. S., Felix M. R., Clara M. I. E., Oliveira S., 2005.The complete genome sequence of a new necrovirus isolated from Olea europaea L. Arch. Virol., 150: 815–823.
Carr G.R., Hinkley M.A., LeTacon F., Hepper C.M., Jones M.G.K, Thomas E., 1985. Improved hyphal growth of two species of vesicular arbuscular mycorrhizal fungi in the presence of suspension cultured plant cells. New Phytol., 101:417–426.
Castaldini M., Turrini A., Sbrana C., Benedetti A., Marchionni M., Fabiani A., Landi S., Santomassimo F., Pietrangeli R., Nuti M. P., Miclaus N., Giovannetti M., 2005. Impact of Bt corn on rhizospheric and soil eubacterial communities and on beneficial mycorrhizal symbiosis in experimental microcosms. Appl. Environ. Microbiol., 71:6719–6729.
Cavagnaro T. R., Gao L-L., Smith F. A., Smith S. E., 2001. Morphology of arbuscular mycorrhizas is influenced by fungal identity. New Phytol., 151:469–475.
Cavallazzi J.R. P., Filho O. K., Sturmer S. L., Rygiewicz P. T., de Mendonc M.M., 2007.Screening and selecting arbuscular mycorrhizal fungi for inoculating micropropagated apple rootstocks in acid soils. Plant Cell. Tiss. Organ Cult., 90:117–129. Chabot S., Bécard G., Piché Y., 1992. Life cycle of Glomus intraradix in root organ culture. Mycologia, 84:315–321. Chadwick J., 1958. The Decipherment of Linear B. Cambridge University Press, pp. 29-40.

138
Chelius M. K., Triplett E. W., 1999. Rapid detection of arbuscular mycorrhizae in roots and soil of an intensivelymanaged turfgrass systemby PCR amplification of small subunit rDNA. Mycorrhiza, 9:61–64. Chen C.Y., Zou J., Zhang S.Y., Zaitlin D., Zhu L. H., 2009. Strigolactones are a new-defined class of plant hormones which inhibit shoot branching and mediate the interaction of plant-AM fungi and plant-parasitic weeds. Review Sci.China Series C.Life Sci., 52(8): 693-700.
Chifflot V., 2008. Étude moléculaire des CMA dans le système agri sylvicole. Mémoire, Université Laval. Québec, 63p.
Cho K., Toler H., Lee J., Ownley B., Stutz J. C., Moore J. L., Augé R. M., 2006. Mycorrhizal symbioses and response of sorghum plants to combined drought and salinity stresses. J. Plant Physiol., 163: 517-528 .
Chotte J .L., 2005. Importance of microorganisms for soil aggregation. In : microorganisms in soil; roles in genesis on functions (eds . Buscot F., Varma A., Berlin ) ,Soil Biol., 3: 107–122. Citernesi, A.S., Vitagliano, C., Giovannetti, M., 1998. Plant growth and root system morphology of Olea europea L. rooted cuttings as influenced byarbuscular mycorrhizas. J. Horticult. Sci. Biotechnol.,73: 647–654.
Clark R. B., 1997. Arbuscular mycorrhizal adaptation, spore germination, root colonization, and host plant growth and mineral acquisition at low pH. Plant Soil, 192:15–22.
Clarkson DT. 1985. Factors affecting mineral nutrient acquisition by plants. Annual Review of Plant Physiology, 36: 77–115.
Claros M. G., Crespillo R., Aguilar M.L., Canovas F. M., 2000. DNA fingerprinting and classification of geographically related genotypes of olive-tree (Olea europaea L.). Euphytica, 116: 131–142, Cochran W.G., 1950. Estimation of bacterial densities by means of the „most probable number‟ method. Biometrica, 6: 105-116.
Cordeiro A. I., Sanchez-Sevilla J.F., Alvarez-Tinaut M.C., Gomez-Jimenez M.C., 2008. Genetic diversity assessment in Portugal accessions of Olea europea by RAPD markers. Biol. Plant., 52 (4): 642-647. Cornet F., Diem H. G., 1982. Etude comparative de l‟efficacité des souches de rhizobium d‟Acacia isolées de sols du Sénégal et effet de la double symbiose Rhizobium-Glomus mosseaesur la croissance de Acacia holosericea et A. raddiana. Revue Bois et Forêts desTropiques, 198 :3-15.
Cordoba A. S., de Mendonca M. M., Sturmer S. L., Rygiewicz P. T., 2001. Diversity of arbuscular mycorrhizal fungi along a sand dune stabilization gradient: A case study at Praia da Joaquina, Ilha de Santa Catarina, South Brazil. Mycoscience, 42: 379-387.
21 and Paul T. Rygiewicz 3) Coulomb P. J., El Maataoui M., Coulomb P. O., 2004. L‟œil de Paon (Cyclconium oleaginum). Le nouvel olivier, pp. 25-26. Cranenbrouck S., Voets L., Bivort C., Renard L., Strullu D.G., Declerck S., 2005. Methodologies for in Vitro Cultivation of Arbuscular Mycorrhizal Fungi with Root Organs. In Vitro Culture of Mycorrhizas (ed. by S. Declerck, D.-G. Strullu and A. Fortin). Soil Biol., 4: 341-375.

139
Crush J.R., Hay M.J.M., 1981. A technique for growing mycorrhizal clover in solution culture. N.Z.J. Agric. Res., 24: 371-372.
Dalpé Y., 2001. In vitro monoxenic culture of arbuscular mycorrhizal fungi: a major tool for taxonomical studies. In: Proc 3rd Natl. Symp. Mycorrhizal Symbioses, Guanajuato, Mexico, pp.321-325.
Dalpé Y., 2004. Mycorrhizal fungi biodiversity in Canadian soils. Can. J. Soil. Sci., 83: 321-330.
Dalpé Y., 2005. Les mycorhizes: un outil de protection des plantes mais pas une panacée. Phytoprotect., 86 : 53-59 .
Dalpé Y., Declerck S., 2002. Development of Acaulospora rehmii spore and hyphal swellings under root-organ culture. Mycologia, 94: 850-855.
Dalpé Y., de Souza F. A., Declerck S., 2005a. Life Cycle of Glomus Species in Monoxenic Culture. In : In Vitro Culture of Mycorrhizas. Eds. Declerck S., Strullu D.-G., Fortin A. Soil Biol., 4: 49-71
Dalpé Y., Cranenbrouck S., Séguin S., Declerck S., 2005b.The Monoxenic Culture of Arbuscular Mycorrhizal Fungi as a Tool for Systematics and Biodiversity. In: In Vitro Culture of Mycorrhizas . eds: Declerck S., Strullu D.-G., Fortin A. Soil Biol., 4: 31-48.
Dandan Z., Zhiwei Z., 2007. Biodiversity of arbuscular mycorrhizal fungi in the hot-dry valley of the Jinsha River, southwest China. Appl. Soil Ecol., 37 : 118 – 128. Dangeard P.A., 1896. Une maladie du peuplier dans l‟ouest de la France. Botaniste, 58:38–43.
DDaanniiee llll TT.. JJ..,, HHuuss bbuunndd RR..,, FFiittttee rr AA.. HH..,, YYoouunngg JJ..PP..WW..,, 22000011 .. MMoolleeccuullaarr ddiivveerrssiitt yy ooff aarrbbuussccuullaarr
mmyyccoorrrrhhiizzaa ll ffuunnggii ccoolloonniizziinngg aarraabbllee ccrrooppss.. FFEEMMSS MMiiccrroobbiiooll.. EEccooll.. ,, 3366:: 220033--220099.. Daniels B., Trappe J. M., 1980. Factors affecting spore germination of the vesicular-arbuscular fungus, Glomus epigaeus. Mycologia, 72:457–471.
Daniels-Hetrick B.A., Leslie J.F., Wilson G.T., Kitt D.G., 1988. Physical and topological assessment of effects of a vesicular-arbuscular mycorrhizal fungus on root architecture of big bluestem. New Phytol., 110: 85–96. Dar G. H., Zargar M.Y., Beigh G.M. 1997. Biocontrol of Fusarium Root Rot in the Common Bean (Phaseolus vulgaris L.) by using Symbiotic Glomus mosseae and Rhizobium leguminosarum. Microb. Ecol., 34:74–80
Davey, M. C. R.S, 2006. Interactions between mosses (Bryophyta) and fungi. Can. J. Bot.,. 84: 1509-1519.
de Casas R. R., Besnard G., Schönswetter P. , Balaguer L., Vargas P., 2006. Extensive gene flow blurs phylogeographic but not phylogenetic signal in Olea europaea L. Theor. Appl. Genet., 113:575–583
de Rougement M., 2007. Les mycorhizes et l‟olivier: Effet sur le développement des plants en pépinière et en verger. Journées méditerranéennes de l‟olivier, Meknès, 22-26 octobre 2007, pp. 2-7. de Souza F. A., 2005. Biology, ecology and evolution of the family of Gigasporaceae, arbuscular mycorrhizal fungi (Glomeromycota). Thèse de doctorat. Leiden University, The Netherlands, 55p.

140
de Souza F. A., Berbara R. L. L., 1999. Ontogeny of Glomus clarum in Ri T-DNA transformed roots. Mycologia, 91:343–350.
de Souza F. A, Declerck S., 2004. Mycelium development and architecture, and spore production of Scutellospora reticulata in monoxenic culture with Ri T-DNA transformed carrot roots. Mycologia, 95:1004–1012.
de Souza F.A., Trufem S.F.B.T., Almeida D.L., Silva E.M.R., Guerra J.G.M., 1999. Efeito de pré-cultivos sobre o potencial de inóculo de fungos micorrízicos
arbusculares e produção de mandioca. Pesquisa Agropecuária Brasileira, 34(10): 1913-1923
de Souza F. A ., DalpéY., Declerck S., de la Providencia. E., Séjalon-Delmas N., 2005. Life History Strategies in Gigasporaceae: Insight from Monoxenic Culture. In: In Vitro Culture of Mycorrhizas. Eds. Declerck S., Strullu D.-G., Fortin A. Soil Biol., 4: 73-91.
Declerck S., Plenchette C., Strullu D.G., 1995. Mycorrhizal dependency of banana (Musa acuminata ,AAA group ) cultivar. Plant soil, 176: 183-187.
Declerck S., Strullu D. G., Plenchette C., 1998 . Monoxenic culture of the intraradical forms of Glomus sp. isolated froma tropical ecosystem: a proposed methodology for germplasm collection. Mycologia, 90:579–585.
Declerck S., Risede J.M., Delvaux B., 2002. Greenhouse response of micropropagated bananas inoculated with invitro monoxenically produced arbuscular mycorrhizal fungi. Mycol. Res., 100:1237–1242.
Declerck S., Séguin S., Dalpé Y., 2005. The Monoxenic Culture of Arbuscular Mycorrhizal Fungi as a Tool for Germplasm Collections. In: In Vitro Culture of Mycorrhizas. Eds. Declerck S., Strullu D.-G., Fortin A. Soil Biology, 4: 17-30. Declerck S., D’or D., Cranenbrouck S., Le Boulengé E., 2001. Modelling the sporulation dynamics of arbuscular mycorrhizal fungi in monoxenic culture. Mycorrhiza, 11:225– 230.
Declerck S., D’Or D., Bivort C., de Souza F. A., 2004. Development of extraradical mycelium of Scutellospora reticulata under root-organ culture: spore production and function of auxiliary cells. Mycol. Res., 108:84–92. Declerck S., Strullu D.G., Plenchette C., Guillemette T., 1996. Entrappement of invitro produced spores of Glomus versiforme in alginate beads: invitro and invivo inoculum potentials. J. Biot., 48: 51-57
Declerck S., Cranenbrouck S., Dalpé Y., Seguin S., Grandmougin-Ferjani A., Fontaine J., Sancholle M., 2000. Glomus proliferum sp. Nov.: a description based on morphological, biochemical, molecular and monoxenic cultivation data. Mycologia, 92 : 1178-1187.
Demartsev V., Aussenberg E. R., Gadkar V., Koltai H., Zilberstein A., Kapulnik Y. 2005. Alteration in tomato (Lycopersicon esculentum) gene expression during early stages of the interaction with Glomus intraradices. In: Management committee and final meeting on “Achievements and Future Landscape for Arbuscular Mycorrhiza Research”, Dijon, France, pp.369-377.
Derkowska E., Sas - Paszt L., Sumorok, B., 2008. The influence of mycorrhization and organic mulches on mycorrhizal frequency in apple and strawberry roots. J. Fr. Orn. Plant Res., 16: 227-242.

141
Desjardins Y., Hernández-Sebastià C., Piché Y., 2005. Monoxenic Culture as a Tool to Study the Effect of the Arbuscular Mycorrhizal Symbiosis on the Physiology of Micropropagated Plantlets in Vitro and ex Vitro. In : In Vitro Culture of Mycorrhizas. Eds. Declerck S., Strullu D.-G., Fortin A. Soil Biol., 4: 181-199.
Dexheimer J. ,1997. Étude structurale et fonctionnelle des interfaces entre le champignon et la plante-hôte. Rev. For. Fr., 43- 56. Diallo A. T., Samb P. I., Ducousso M., 1999 . Arbuscular mycorrhizal fungi in the semi-arid areas of Senegal. Eur. J. Soil Biot., 35 (2):65-75.
Dickson S. 2004. The Arum–Paris continuum of mycorrhizal symbioses. New Phytol., 163:187–200.
Dickson S., Kolesik P., 1999.Visualisation of mycorrhizal fungal structures and quantification of their surface area and volume using laser scanning confocal microscopy. Mycorrhiza, 9:205–213. Dickson S., Smith F. A., Smith S. E., 2007. Structural differences in arbuscular mycorrhizal symbioses: more than 100 years after Gallaud, where next?. Mycorrhiza, 17:375–393.
Diop T. A., 2003. In vitro culture of arbuscular mycorrhizal fungi: advances and future prospects. Afr. J.Biotec., 2 (12) : 692-697.
Diop T. A., Plenchette C., Strullu D. G. 1994. Dual axenic culture of sheared-rood inocula of vesicular-arbuscular mycorrhizal fungiassociated with tomato roots. Mycorrhiza, 5:17-22.
Djili K., Daoud Y., Ayache N., 1999. Analyse de la distribution verticale du calcaire dans les sols de l‟Algérie septentrionale.Etude et Gestion des Sols. I.N.A., Département de science des sols El-Harrach, Alger, Algérie, pp. 202-213.
Dodd J. C., 2000. The role of arbuscular mycorrhizal fungi in agro and natural ecosystems. Agricul., 29 (1): 63-70. Dodd J. C., Boddington C. L., Rodriguez A., Gonzalez-Chavez C., Mansur I., 2000. Mycelium of arbuscular mycorrhizal fungi (AMF) from different genera: form, function and detection. Plant Soil, 226: 131-151. Dotzler N., Krings M., Taylor T. N., Agerer R., 2006. Germination shields in Scutellospora (Glomeromycota:Diversisporales, Gigasporaceae) from the 400 million-year-old Rhynie chert. Mycol. Progress., 5:178–184.
Douds D. D., 1997. A procedure for the establishment of Glomus mosseae in dual culture with Ri T-DNA-transformed carrot roots. Mycorrhiza, 7:57–61. Douds D.D., Schenck N.C., 1991. Germination and hyphal growth of VAM fungi during and after storage in soil at five matric potentials. Soil Biol. Biochem., 23:177–183.
Douds D.D., Millner P., 1999. Biodiversity of arbuscular mycorrhizal fungi in agroecosystems. Agric. Ecosyst. Environ., 74:77–93.
Douds D.D., Nagahashi G., Abney G. D., 1996. The differential effects of cell wall associated phenolics,cell walls, and cytosolic phenolics of host and non host roots on the growth of two species of AM fungi. New Phytol., 133:289–29.

142
Dreyer B., Morte A., López J. Á., Honrubia M., 2009. Comparative study of mycorrhizal susceptibility and anatomy of four palm species. Mycorrhiza, pp.1-13.
Ducousso M., Colonna J.P., 1993. Infection endomycorhizienne chez de jeunnes Faidherbia albida : influence sur la croissance et le développement. ICRAF, Niamey, Niger, pp. 151-156.
DDuummaass --Gaudot EE..,, GGoolllloottttee AA..,, CCoorrddiiee rr CC..,, GGiiaanniinnaazzzzii SS..,, GGiiaanniinnaazzzzii--PPee aarrssoonn VV..,, 22000000.. MMoodduullaatt iioonn ooff hhoosstt ddeeffeennssee ssyysstteemmss.. IInn:: AArrbbuussccuullaarr MMyyccoorrrrhhiizzaass :: PPhhyyssiioollooggyy aanndd FFuunnccttiioonn.. EEddss.. KKaappuullnniikk YY..,, DDoouuddss YY.. DD..DD.. JJrr,,,, KKlluuwweerr AAccaaddeemmiicc PPrreessss,, DDoorrddrreecchhtt,, TThhee NNeetthheerrllaannddss:: 117733--220000..
Echairi A., Nouaim R., Chaussod R., 2008. Intérêt de la mycorhization contrôlée pour la production de plants d‟arganier (Argania spinosa) en conditions de pépinière. Article de recherche. Sécheresse, 19 (4) : 277-81.
Egger K. N., Sigler L., 1993. Relatedness of the ericoid endophytes Scytalidium vaccinii and Hymenoscyphus ericae inferred from analysis of ribosomal DNA. Mycologia, 85: 219–230.
Egli S., Brunner I., 2002. Les mycorhizes Une fascinante biocénose en forêt. Not. Prat., 35 : 1-8.
Elias K., Safir G., 1987. Hyphal elongation of Glomus fasciculatus in response to root exudates. Appl. Environ. Microbiol., 53: 1928–1930.
EEllsseenn AA..,, DDeeccllee rrcckk SS.. ,, DDee WWaaee llee DD..,, 22000011.. EEffffeecctt ooff GGlloommuuss iinnttrraarraaddiicceess oonn tthhee rreepprroodduuccttiioonn ooff tthhee bbuurrrroowwiinngg nneemmaattooddee ((RRaaddoopphhoolluuss ssiimmiilleess)) iinn ddiixxeenniicc ccuulltt uurree.. MMyyccoorrrrhhiizzaa ,, 1111:: 4499-- 5511..
EEllsseenn AA..,, DDeeccllee rrcckk SS.. ,, DDee WWaaee llee DD..,, 22000022 ..EEffffeett ddee ttrrooiiss cchhaammppiiggnnoonnss mmyyccoorrhhiizziieennss aarrbbuussccuullaaiirreess ssuurr
ll‟‟ iinnffeeccttiioonn dduu bbaannaanniieerr ppaarr llee nnéémmaattooddee àà ggaallllee ddeess rraacciinneess ((MMeellooiiddooggyynnee sspppp..))..IInnffoommuussaa,, 1111 ((11)) :: 2211--
2233..
EEllsseenn AA..,, BB aaiimmee yy HH..,, SSwwee nnnnee nn RR..,, DDee WWaaee llee DD..,, 22000033.. RReellaattiivvee mmyyccoorrrrhhiizzaa ll ddeeppeennddeennccyy aanndd mmyyccoorrrrhhiizzaa-- nneemmaattooddee iinntteerraaccttiioonn iinn bbaannaannaa ccuulltt iivvaarrss ((MMuussaa sspppp..)) ddiiffffeerriinngg iinn nneemmaattooddee ssuusscceeppttiibbiilliitt yy..
PPllaanntt SSooiill,, 225566:: 330033--331133.. Enkhtuya B., Vosatka M., 22000055.. IInntteerraaccttiioonn bbeettwweeeenn ggrraassss aanndd ttrreeeess mmeeddiiaatteedd bbyy eexxttrraarraaddiiccaall
mmyycceelliiuumm ooff ssyymmbbiioott iicc aarrbbuussccuullaarr mmyyccoorrrrhhiizzaall ffuunnggii.. Symbiosis,, 3388((33)) :: 226611--227766
Fabbri A., Bartolini G., Lambardi M., Kailis S.G., 2004. Olive propagation manual. Landlinks Press, Espagne, 160 p. Fang Y.C., Huang Z., Liu Y.R., 2000. Study on VA mycorrhizal flora of tobacco (Nicotiana tobacum). Acta. Tob. Sinica., 6:26–31 Fantanazzo G., Baldoni L., 1999 – Propositions pour un programme d‟amélioration génétique de l‟olivier, Rév. Olivae, 34: 27 – 38. Ferrol N., Barea J. M., Azcon-Aguilar C., 2002. Mechanisms of nutrient transport across interfaces in arbuscular mycorrhizae. Plant Soil, 244:231-237.
Fester T., Hause B., Schmidt D., Halfmann K., Schmidt J., Wray V., Hause G., Strack D., 2002. Occurrence and localization of apocarotenoids in arbuscular mycorrhizal plant roots. Plant Cell. Physiol., 43:256–265.
Fidelibus M.W., Martin C.A., Wright G.C., Stutz J.C., 2000. Effect of arbuscular mycorrhizal (AM) fungal communities on growth of „Volkamer‟ lemon in continually moist or periodically dry soil. Scienta Hort., 84: 127-140.

143
FFiilliioonn MM ..,, SStt AArrnnaauudd MM..,, FFoorrttiinn JJ.. AA..,, 11999999.. DDiirreecctt iinntteerraaccttiioonn bbeettwweeeenn tthhee aarrbbuussccuullaarr mmyyccoorrrrhhiizzaall
ffuunngguuss GGlloommuuss iinnttrraarraaddiicceess aanndd ddiiffffeerreenntt rrhhiizzoosspphheerree mmiiccrroooorrggaanniissmmss.. NNeeww PPhhyyttooll.. ,, 114411:: 221111-- 222211..
Filion M., St-Arnaud M., Guillon C., Hamel C., Jabaji-Hare S. H., 2001. Suitability of Glomus intraradices in vitro-produced spores and root segment inoculumfor the establishment of a mycorrhizosphere in an experimental microcosms. Can. J. Bot., 79:879–888.
Fisher J. B., Jayachandran K., 2005. Presence of arbuscular mycorrhizal fungi in South Florida native plants. Mycorrhiza, 15:580–588.
Flores S., Saxena D., Stotzky G., 2005. Transgenic Bt plants decompose less in soil than non-Bt plants. Soil Biol. Biochem., 37:1073–1082. Fortas Z., 1990. Etude de trois espèces de terfez : caractères culturaux et cytologie du mycélium isolé et associé à Hélianthémum gutatum thèse de Doctorat d‟état, Université d‟Oran Es-Senia (Algérie) INRA de Clermont- ferrand (france), 166p.
Fortas Z., Chevalier G., 1992a. Caractéristiques de la germination des ascospores de Terfezia arenaria (Moris) Trappe, récolté en Algérie. Cryptogamie, Mycol., 13: 21-29.
Fortas Z., Chevalier G., 1992 b. Effet des conditions de culture sur la mycorhization de l’heliantthemum guttatum par trois espèces de Terfez des genres Terfezia et Tirmania d‟Algérie. Can. J. Bot., 70: 2453-2460.
Fortin J.A., Plenchette C., Piché Y., 2008. Les mycorhizes. La nouvelle révolution verte. MultiMonde Quae. (Eds.), Québec, 131 p.
Fortin J.A., Bécard G., Declerck S., Dalpé Y., St-Arnaud M., Goughlan A.P., Piché Y., 2002. Arbuscular mycorrhiza on root-organ Culture. Can. J. Bot., 80:1-20. Frank A.B., 1885. Ueber die auf Wurzelsymbiose beruhende Ernahrung gewisser Baume durch unterirdische Pilze.Ber Deutch Bot. Gessel., 3: 128-145.
Franke-Snyder M., Douds D.D. Jr., Galvez L., Phillips J.G., Wagoner P., Drinkwater L., Morton J.B., 2001. Diversity of communities of arbuscular mycorrhizal (AM) fungi present in conventional versus low-input agricultural sites in eastern Pennsylvania, USA. Appl. Soil Ecol., 16:35–48. Fries E.M., 1829. Beobachtungen iiber die thamniscophage Mykorrhiza einiger Halophyten. Bot. Not. , 255-264. Friese C. F., Allen M. F., 1991. The spread ofVAmycorrhizal fungal hyphae in the soil: inoculums types and external hyphal architecture. Mycologia, 83:409–418.
Gadkar V., Adholeya A., 2000. Intraradical sporulation of AM Gigaspora margarita in longterm axenic cultivation in Ri T-DNA carrot root. Mycol. Res., 104:716–721.
Gai J. P., Liu R. J., 2003. Effects of soil factors on AMF in the rhizosphere of wild plants. Chin J.Appl. Ecol., 14:18–22.
Gai J. P., Christie P., Feng G., Li X. L., 2006. Twenty years of research on community composition and species distribution of arbuscular mycorrhizal fungi in China: a review Mycorrhiza, 16: 229–239. Gallaud I., 1905. Études sur les mycorrhizes endotrophes. Rev. Génér. Bot., 17:5–48.

144
Gaouar-Benyelles N., 1996. Apport de la biologie des populations de la mouche de l‟olivier Bactrocera (Dacus) oleae Gmel (Ditera : Tephiritdae) à l‟optimisation de son contrôle dans la région de Tlemcen. Thèse de Doctorat. Université de Tlemcen, Algérie, 116 p Garcia-Romera I., Garcia-Garrido J. M., Ocampo J.A., 1991. P ectolytic enzymes in the vescicular arbuscular mycorrhizal fungus Glomus mosseae. F.E.M.S. Microbiol. Lett., 78: 343– 346.
Gaspar L., Pollero R.J., Cabello M.N., 1994. Triacylglycerol consumption during spore germination of vesicular arbuscular mycorrhizal fungi. J. Am. Oil Chem. Soc., 71:449–452.
Gaur A., Adholeya A., 2004. Prospects of arbuscular mycorrhizal fungi in phytoremediation of heavy metal contaminated soils. Curr. Sci., 86 : 528–534.
Gazey C., Abbott L.K., Robson A.D., 1993. VA mycorrhizal spores from three species of Acaulospora – Germination, longevity and hyphal growth. Mycol. Res., 97:785–790.
Gemma J. N., Koske R. E. 1989. Field inoculation of American beachgrass (Ammophila breviligulata) with V-A mycorrhizal fungi. J. Environm. Manag. , 29: 173-182.
Gemma J. N., Koske R. E., Carreiro M. 1989. Seasonal dynamics of selected species of VA mycorrhizal fungi in a sand dune. Mycol. Res., 92 : 317-321.
Genre A., Chabaud M., Timmers T., Bonfante P., Barker D.G., 2005. Arbuscular mycorrhizal fungi elicit a novel intracellular apparatus in Medicago truncatula root epidermal cells before infection. Plant Cell., 17:3489–3499. Gerdemann J.W., Nicolson T.H., 1963. Spores of mycorrhizal endogone extracted from soil by wet seiving and decanting. Trans. Br. Mycol. Soc., 46:235–244.
Gerdemann J. W., Trappe J. M. 1974. The Endogonaceae in the Pacific Northwest. Myc. Memoir., 5: 1-76.
Gianinazzi S., Trouvelot A., Gianinazzi-Pearson V., 1989. Conceptual approaches for the rational use of VA endomycorrhizae in agriculture: possibilities and limitations. Agric. Ecosyst. Env., 29: 153-161.
Gianinazzi-Pearson V., Gianinazzi S., 1983. The physiology of vesicular-arbuscular mycorrhizal roots. Plant Soil, 71: 197-209.
Gianinazzi-Pearson V. Gianinazzi S., Trouvelot A., 1985. Evaluation of the infectiveness of indigenous vesicular arbuscular fungal population in some agricultural soils of Burgundy. Can. J. Bot., 63:1521-1524.
Gianinazzi-Pearson V., Branzanti B., Gianinazzi S., 1989. In vitro enhancement of spore germination and early hyphal growth of a vesicular-arbuscular mycorrhizal fungus by host root exudates and plant flavonoids. Symbiosis, 7:243–255.

145
Giasson P., Karam A., Jaouich A., 2008. Arbuscular mycorrhizae and allevation of soil stresses on plant growth. ZA. . Siddiqui et al. (eds.), Mycorrhizae: Sustainable Agriculture and Forestry: 99–134.
Giovannetti M., 1983. Establishment and growth effects of Glomus mosseae on the legume
Hedysarum coronarium L. growing in poor alkaline soils. Soil Biol. Biochem.,15:385–387.
Giovannetti M., Avio L., Salutini L., 1991. Morphological, cytochemical, and ontogenetic characteristics of a new species of vesicular-arbuscular mycorrhizal fungus. Can. J. Bot., 69:161–167
Giovannetti M., Sbrana C., Logi C., 2000. Microchambers and video-enhanced light microscopy for monitoring cellular events in living hyphae of arbuscular mycorrhizal fungi. Plant Soil, 226:153–159.
Giovannetti M., Avio L., Sbrana C., 2010. Fungal Spore Germination and Pre-symbiotic Mycelial Growth – Physiological and Genetic Aspects. In: Arbuscular Mycorrhizas. eds. Koltai H., Kapulnik Y. Physiol. Funct.: 3 -32 .
Giovannetti M., Avio L., Sbrana C., Citernesi A.S., 1993. Factors affecting appressorium development in the vesicular-arbuscular mycorrhizal fungus Glomus mosseae (Nicol. & Gerd.) Gerd. & Trappe. New Phytol., 123: 115 – 122.
Giri B., Kapoor R., Mukerji K. G., 2005. Effect of the arbuscular mycorrhizae Glomus fasciculatum and G. macrocarpum on the growth and nutrient content of Cassia siamea in a semi-arid Indian wasteland soil. New Forests, 29:63–73
Godfrey R. M., 1957. Studies of British species of Endogone. I. Morphology and taxonomy. Trans. Br. Mycol. Soc. , 40 : 117-135.
GGooss lliigg PP..,, HHooddggee AA..,, GGooooddllaassss GG..,, BBeennddiinngg GG.. DD..,, 22000066.. AArrbbuussccuullaarr mmyyccoorrrrhhiizzaall ffuunnggii aanndd oorrggaanniicc
ffaarrmmiinngg.. AAggrr iiccuulltt.. EEccoossyysstt.. EEnnvv..,,111133:: 1177-- 3355..
GGoossss MM.. GG..,, ddee VVaarree nnnneess AA..,, 22000022.. SSooiill ddiissttuurrbbaannccee rreedduucceess tthhee eeffffiiccaassssiittyy ooppff mmyyccoorrrrhhiizzaa ll
aassssoocciittiioonnssffoorr eeaarrllyy ssooyybbeeaann ggrroowwtthh aanndd NN22 ffiixxaatt iioonn..SSooiill BB iiooll.. BB iioocchh..,, 3344:: 11116677--11117733..
Govindarajulu M., Pfeffer P.E., Jin H., Abubaker J., Douds D. D., Allen J.W. , Bucking H., Lammers P.J., Shachar-Hill Y., 2005. Nitrogen transfer in the arbuscular mycorrhizal symbiosis. Nature, 435: 8419-823.
Green C. D., Stodola A., Auge R.M., 1998. Transpiration of detached leaves from mycorrhizal and nonmycorrhizal cowpea and rose plants given varying abscisic acid, pH, calcium and phosphorus. Mycorrhiza, 8: 93–99.
Green J.J., Baddeley J.A.,Cortina J., Watson C.A., 2005. Root development in the Mediterranean shrub Pistacia lentiscus as affected by nursery treatments.J. Arid Env., 61:1–12.
Green N.E., Graham J.H., Schenck N.C., 1976. The influence of pH on the germination of vesicular arbuscular mycorrhizal spores. Mycologia, 68: 929–934.
Greipsson S., El-mayas H., 2002. Synergistic effect of soil pathogenic fungi and nematodes reducing bioprotectoin of arbuscular mycorrhizal fungi on the grass leymus arenarius. Biocontrol, 47: 715-727. Greipsson S., El-Mayas H., Vestberg M., Walker C. 2002. Arbuscular mycorrhizal fungi in sandy soils in Iceland. Arctic Antarctic Alpine Res., 34 : 419-427.

146
Greny A., 1973. Étude anatomo-morphologique des endomycorrhizes constitue par le Maïs, l‟Avoine, le Blé, l‟Orge et diverses Graminées prairiales et adventices. Mémorie presenté au Conservatoire des Arts et Métiers pour obtenir le Diplome d‟Ingenieur C.N.A.M., Paris. 126p. Guether M., Balestrini R., Hannah M.A., Udvardi M.K., Bonflote P., 2009. Genome-wide reprogramming of regulatory networks, transport ,cell wall and membrane biogenesis during arbuscular mycorrhizal symbiosis in Lotus japonicus. New Phytol., 182: 200-212. Guechi A., Girre L., 2002. Recherche et analyse d‟un effet mutagène des extraits de feuilles
d‟olivier parasitées par le champignon Cycloconium oleaginum Cast. Sciences et Technologie,
Algérie, 18: 96-100.
Guillemin J.P., Gianinazzi S., 1994. Contribution des endomycorhizes a la production de plantes micropropagées d‟ananas (Ananas comosus (L.).Thèse de doctorat, Université de Dijon, France, 200 p.
Guissou T., 2009. Contribution of arbuscular mycorrhizal fungi to growth and nutrient uptake by jujube and tamarind seedlings in a phosphate (P)-deficient soil. Afr. J. Microbiol. Res., 3 (5) : 297-304.
Guissou T., Ba A. M., Plenchette C., Guinko S., Duponnois R., 2001. Effets des mycorhizes à arbuscules sur la tolérance à un stress hydrique de quatre arbres fruitiers : Balanites aegyptiaca (L.) Del., Parkia biglobosa (Jacq.) Benth., Tamarindus indica L. et Zizyphus mauritiana Lam. Note de recherche. Science et changements planétaires / Sécheresse, 12 (2): 121-127.
Gunasekaran P., Sundaresan P., Ubalthoose R. N., Lakshmanan M. , 1987. Effect of pH, temperature and nutrients on the germination of a vesicular arbuscular mycorrhizal fungus, Glomus fasciculatum in vitro. Proc. Indian AS-Plant Sci., 97:231–234. Goltapeh E. M., Danesh Y. R., Prasad R., Varma A., 2008. Mycorrhizal Fungi: What We Know and What Should We Know?. A. Varma (ed.) Mycorrhiza, n°??: 3-27 .
Guttenberger M., 2000. A rapid staning procedure for arbuscules of living arbuscular mycorrhizas using neutral red as acidotropic dye.Plant Soil , 226:211-218.
Hahn A. , Bonfante P., Horn K., Pausch F., Hock B., 1993. Production of monoclonal antibodies against surface antigens of spores from arbuscular mycorrhizal fungi by an improved immunization and screening procedure. Mycorrhiza, 4:69-78.
Hamlat M. , 1995. Influence des phyto hormones sur les embryons, les micro boutures d‟olivier (Olea europea L.). Thèse de magistère, Inst. Biol. Tizi Ouzou, Algérie. 136p.
HHaaoo ZZ..PP..,, CChhrriiss ttiiee PP.. ,, QQiinn LL..,, WWaanngg CC..XX..,, LLii,, XX..LL..,, 22000055.. CCoonnttrrooll ooff FFuussaarr iiuumm ww iilltt ooff ccuuccuummbbeerr sseeeeddlliinnggss bbyy iinnooccuullaatt iioonn wwiitthh aann aarrbbuussccuullaarr mmyyccoorrrrhhiizzaall ffuunngguuss.. JJ.. PPllaanntt NNuuttrr.. ,, 2288:: 11996611––11997744.. Harinikumar K. M., Bagyaraj D., 1989. Effect of cropping sequence, fertilizers and farmyard manure on vesicular-arbuscular mycorrhizal fungi in different crops over three consecutive seasons. Biol. Fertil. Soils, 7:173–175.
Harley JL. 1989. The fourth benefactors‟ lecture the significance of mycorrhiza. Mycological Research 92: 129 – 139
Harrison M. J., 1997. The arbuscuar mycorrhizal symbiosis: an underground association. Trends Plant Sci., 2 : 54-60.

147
Harrison M. J., 1999. Molecular and cellular aspects of the arbuscular mycorrhizal symbiosis. Plant Physiol., 50:361–89.
Hart M. M., Klironomos J.N., 2002. Diversity of arbuscular mycorrhizal fungi and ecosystem functioning In: van der Heijden MGA, Sanders IR (eds) Mycorrhizal ecology, vol 157. New York, pp 225–242.
HHaarrtt MM..MM..,, RRee aaddee rr RR..JJ..,, KKlliirroonnoommooss JJ..NN..,, 22000033.. PPllaanntt ccooeexxiisstteennccee mmeeddiiaatteedd bbyy aarrbbuussccuullaarr mmyyccoorrrrhhiizzaa ll ffuunnggii.. TTrreennddss EEccooll.. EEvvooll.. ,, 1188:: 441188 –– 442233..
HHaassee llwwaannddttee rr KK,, RRee aadd DD.. JJ.. ,, 11998800.. FFuunnggaall aassssoocciiaattiioonn ooff rroooott ooff ddoommiinnaanntt aanndd ssuubb--ddoommiinnaanntt ppllaannttss
iinn hhiigghh AA llppiinneessvveeggeettaattiioonn ssyysstteemmss wwiitthh ssppeecciiaall rreeffeerreennccee ttoo mmyyccoorrrrhhiizzaa.. OOeeccooll.. ,, 4455:: 5577--6622.. Hausse B., Fester T., 2005. Molecular and cell biology of arbuscular, mycorrhizal symbiosis. Planta, 221: 184-196. Hausse B., Mrock C., Isayenkov S., Strack D., 2007. Jasmonates in arbuscular mycorrhizal interactions. Phytochem., 68(1):101-110.
Helgason T., Daniell T.J., Husband R., Fitter A.H., Young J.P.W., 1998. Ploughing up the wood-wide. Nature, 394:431-439.
HHee llggaass oonn TT..,, MMaarryywwee aatthhee rr JJ..WW..,, DDee nniissoonn JJ.. ,, WWiillss oonn PP..,, YYoouunngg JJ..PP..WW..,, FFiittttee rr AA.. HH..,, 22000022 .. SSeelleeccttiivviitt yy aanndd ffuunncctt iioonnaa ll ddiivveerrssiitt yy iinn arbuscular mycorrhizas of occurrence fungi and plant from temperate deciduouswoodland. J. Ecol., 90:371-384.
Henrion B., Chevalier G., Martin F., 1994. Typing truffle species by PCR amplification of ribosomal DNA spacer. Mycol. Res. 98: 37–43.
Hepper C. M., 1984. Regulation of spore germination of the vesicular-arbuscular mycorrhizal fungus Acaulospora laevis by soil pH.. Trans. Br. Mycol. Soc., 83:154–156.
Hepper C.M., Smith G.A., 1976. Observation on the germination of Endogone spores. Trans. Br. Mycol. Soc., 66:189–194.
Hepper C. M., Jakobsen I., 1983. Hyphal growth from spores of the mycorrhizal fungus Glomus caledonius: effect of amino acids. Soil Biol. Biochem., 15:55–58. Herrera-Medina M. J., Tamayo M. I., Vierheilig H., Ocampo J. A., Garcıa-Garrido J. M.. 2008. The Jasmonic Acid Signalling Pathway Restricts the Development of the Arbuscular Mycorrhizal Association in Tomato. J. Plant Growth Regul. 27:221–230.
Herrmann P., 1980. Travaux pratiques. D.A.A. Science du sol- A mangement. D.E.A. Agronomic (option pédologie). Ecole Nationale Supérieur Agronomique. Montpellier. Chaire de géologie. Science du Sol, 63 p. Heywood, V. H., 1978. Flowering Plants of the World. Oxford, London, Melbourne: Oxford University Press, pp.18-25.
Hibbett D. S., Matheny P. B., 2009. The relative ages of ectomycorrhizal mushrooms and their plant hosts estimated using Bayesian relaxed molecular clock analyses. B.M.C. Biol.: 7-13 .
Hibbett D.S., Gilbert L-B, Donoghue M. J., 2000. Evolutionary instability of ectomycorrhizal symbioses in basidiomycetes. Nature, 407:506–508.

148
Hibbett D. S., Binder M., Bischoff J., 2007. A higher-level phylogenetic classification of the Fungi. Mycol. Res., 111:509–547. Hijri M., Sanders I.R., 2005. Low gene copy number shows that arbuscular mycorrhizal fungi inherit genetically different nuclei. Nature, 433: 160-163
Hijri M., Kuhn G., Sanders I.R., 2001. Evidence for the evolution of multiple genomes in arbuscular mycorrhizal fungi. Nature, 414: 745-748
HHiijjrrii M.,, SSyyccoorroovvaa ZZ..,, OOee hhll FF..,, IInnee iicchheenn KK..,, MM aaddee rr PP..,, WWiiee mmkkee nn AA..,, RReeddeecckkee rr DD..,, 22000066.. CCoommmmuunniitt iieess ooff arbuscular mycorrhizal fungi iinn aarraabbllee ssooiillss aarree nnoott nneecceessssaarriillyy llooww iinn ddiivveerrssiitt yy.. MMooll..
EEccooll.. ,, 1155:: 22227777--22228899..
Hoad, T.F., 1991. The concise Oxford Dictionary of English Etymology. The Clarendon Press, pp. 22-26. Ian A. D., Guza R. C., Krazewski S. E., Peter B., 2004. Reich Shared ectomycorrhizal fungi between a herbaceous perennial ( Helianthemum bicknellii ) and oak ( Quercus ) seedlings. New Phytol., 164: 375–382.
Istambouli A., 1976. Etude expérimentale sur la nature des périodes de repos des semences et des bourgeons d‟olivier (Olea europea L). Thèse. Université D‟Aix, Marseille III, France. 200p.
Iwaniuk A., Blaszkowski J. 2004. Arbuscular fungi and mycorrhizae of agricultural soils of the Western Pomerania. Part I. Occurrence of arbuscular fungi and mycorrhizae. Acta Mycol. 39(1), 59-84. Jaizme-Vega M.C., Azcon R., 1991. Effect of vesicular –arbuscular myorrhizal fungi on pineapple (Ananas comosus L. Merr.) in the Canary Island. Fruits, 46: 155-167.
Jaizme-Vega M. C., Azcon R., 1995. Response to some tropical and subtropical cultures to endomycorrhizal fungi. Mycorrhiza, 5: 213-217.
Jaizme-Vega MC., Esquivel Delamo M., Dominguez P. T., Rodriguez Romero A.S., 2002. Effets de la mycorhization sur le développement de deux cultivars de bananiers issus de la micropropagation. Infomusa, 11(1): 25-28.
Jaizme-Vega M. C., Rodriguez-Romeo A.S., Hermoso C.M., Declerck S., 2003. Growth of micrpropagated bananas colonized by root-organ culture produced arbuscular mycorrhizal fungi entrapped in Ca-alginate beads. Plant and Soil, 254:329-335.
Jakobsen I., 1995. Transport of phosphorus and carbon in VA mycorrhizas. In: Mycorrhiza. Eds. Varma A , Hock. B, Germany, pp. 297–324. Jansa J., Mozafar A., Anken T., Ruh R., Sanders I.R., Frossard E., 2002 . Diversity and structure of AMF Communities as affected by tillage in a temperate soil. Mycorrhiza, 12: 225-234.
Jansa J., Mozafar A., Kuhn G., Anken T., Ruh R,. Sanders I. R., Frossard E., 2003. Soil tillage affects the community structure of mycorrhizal fungi in maize roots. Ecol. Appl., 13:1164–1176.
Janse J. M. 1897. Les endophytes radiaux de quelques plantes Javanaises. Ann. Jardin Bot. Buitenzorg, 14 : 53-201. Janos, D. P. 2007. Plant responsiveness to mycorrhizae differs from dependence upon mycorrhizae. Mycorrhiza 17:75–91.

149
Janoušková M., Seddas P., Mrnka L., van Tuinen D., Dvořáčková A., Tollot M., Gianinazzi-Pearson V., Vosátka M., Gollotte A., 2009. Development and activity of Glomus intraradices as affected by co-existence with Glomus claroideum in one root system. Mycorrhiza, 19:393–402. Jasper D.A., Abbot L.K., Robson A.D., 1989. Soil disturbance reduces the infectivity of the external hyphae of vesicular–arbuscular mycorrhizal fungi. New Phytol., 112:101–107
Jeffries P., Boyer L. R., Rice P., Ray J. N., Dodd J. C., 2007. Ultrastructure of spore development in Scutellospora heterogama. Mycorrhiza, 17:395–403.
Johansson M., 2001. Fungal associations of Danish Calluna vulgaris roots with special reference to ericoid mycorrhiza. Plant Soil, 231: 225–232.
Johansson J. F., Paul L. R., Finlay R. D., 2004. Microbial interactions in the mycorrhizosphere and their significance for sustainable agriculture. FEMS. Microbiol. Ecol., 48: 1-13.
Johnson D., Vandenkoornhuyse P.J., Leake J.R., Gilbert L., Booth R.E. Grime J.P., Young J.P.W., Read D.J., 2004. Plant communities affect arbuscular mycorrhizal fungal diversity and community composition in grassland microcosms. New Phytol., 161(2):503-515.
Jones F.R., 1924. A mycorhizal fungus in the root of legumes and some other plants. J. Agric. Res., 29: 459-470. Jose S., Gillespie A. R., Seifert J. R., Mengel D. B., Pope P. E., 2000. Defining competition vectors in a temperate alley cropping system in the Midwestern USA -3. Competition for nitrogenand litter decompositiondynamics. Agroforest. Syst. 48: 61-77. Juge C., Samson J., Bastien C., Vierheilig H., Coughlan A., Piché Y., 2002. Breaking dormancy in spores of the arbuscularmycorrhizal fungus Glomus intraradices: a critical cold-storage period. Mycorrhiza, 12:37–42.
JJuummppoonnee nn AA..,, 22000011.. DDaarrkk sseeppttaattee eennddoopphhyyttee-- AArree tthheeyy mmyyccoorrrrhhiizzaa ll.. MMyyccoorrrrhhiizzaa,, 1111::220077-- 221111
KKaabbiirr ZZ..,, 22000055.. TTiillllaaggee oorr nnoo ttiillllaaggee :: IImmppaacctt oonn mmyyccoorrrrhhiizzaaee.. CCaann.. JJ.. ppllaanntt SSccii.. ,, 8855:: 2233--2299..
Kabir Z., O’Halloran I.P., Hamel C., 1997. Overwinter survival of arbuscular mycorrhizal hyphae is favored by attachment to roots but diminished by disturbance. Mycorrhiza, 7: 197-200.
Kahiluoto H., Ketoja E., Vestberg M., Saarela I., 2001. Promotion of AM utilization through reduced P fertilization Field studies. Plant Soil, 231:65–79.
Kaldorf M., Ludwig-Müller J., 2000. AM fungi might affect the root morphology of maize by increasing indole- 3-butyric acid biosynthesis. Physiol. Plant, 109 (1): 58-67.
Kamienski F., 1881. Die vegetations organe der Monotropa hypopitys L. Bot. Zeitchr., 39: 225-234.
Kanno T., Saito M., Ando Y., Macedo M.C.M., Nakamura T., Miranda C.H.B., 2006. Importance of indigenous arbuscular mycorrhiza for growth and phosphorus uptake in tropical forage grasses growing on an acid, infertile soil from the Brazilian savannas. Tropical Grasslands, 40: 94–101.
Kapoor R., Giri B., Mukerji K.G., 2002. Glomus macrocarpum: a potential bioinoculant to improve essential oil quality and concentration in dill (Anethum graveolens L.) and carum (Trachyspermum ammi (Linn.) Sprague). World J. Microbiol. Biotechnol., 18:459-463.

150
Karandashov V.E., Kuzovkina I.N., George E., Marschner H., 1999. Monoxenic culture of arbuscular mycorrhizal fungi and plant hairy roots. Russian J. Plant Physiol., 46:87–92.
KKaarraass aawwaa TT..,, KKaass aahhaarraa YY..,, TTaakkeebbee MM..,, 22000022.. DDiiffffeerreennccee iinn ggrroowwtthh rreessppoonnssee ooff mmaaiizzee ttoo pprreecceeddiinngg
ccrrooppppiinngg ccaauusseedd bbyy fflluuccttuuaatt iioonniinn tthhee ppooppuullaatt iioonn ooff iinnddiiggeennuuoouuss aarrbbuussccuullaarr mmyyccoorrrrhhiizzaall ffuunnggii.. SSooiill
BBiiooll.. BBiioocchh..,, 3344::885511-- 885577..
Kernaghan G., 2005. Mycorrhizal diversity: cause and effect ?. Pedobiol., 49: 511-520. Khade, S. W., Adholeya, A., 2007. Feasible bioremediation through arbuscular mycorrhizal fungi imparting heavy metal tolerance: A retrospective. Bioremedia. J., 11: 1–33.
Khade S., W., Adholeya A., 2009. Arbuscular Mycorrhizal Association in Plant Growing on Metal- Contaminated Sopils Adjoining Kanpur Tanneries, Uttar Pradesh, India Water Air Soil Pollu., 202: 45-56. Khade S. W., Rodrigues B. F., 2003. Incidence of arbuscular mycorrhizal colonization in tubers of Gloriosa superba L. Mycorrhiza News, 15(3): 14 – 16
Khasa D.P., Hambling B., Kernaghan G., Fung M., Ngimbi E., 2002. Genetic variability in salt tolerance of selected boreal woody seedlings. Forest Ecology and Management 165:257-269.
Kliromonos J.N., McCune J., Hart M., Neville J., 2000. The influence of arbuscular mycorrhizae on the relationship between plant diversity and productivity. Ecol. Letters, 3: 137-141.
KKllooee ppppee rr JJ.. WW..,, RRyyuu CC.. MM..,, ZZhhaanngg SS..,, 22000044.. IInndduucceedd rreessiissttaannccee aanndd pprroommoott iioonn ooff ppllaanntt ggrroowwtthh bbyy
BBaacciilllluuss sssspp.. PPhhyyttooppaatthhooll.. ,, 9944:: 11225599--11226666.. Kormanik P.P., McGraw A.C., 1982. Quantification of vesicular-arbuscular mycorrhizal in plants roots. In: Methods and principles of mycorrhizal research, Schenck N.C. (éd.). Amer. Phytop. Soc. Minnesota, pp. 37-45. Koske R. E., 1981. Gigaspora gigantea: observations on spore germination of a VA-mycorrhizal fungus. Mycologia, 73:288–300. Koske R.E., Testier B., 1983. A convenient permanent slide mounting medium. Mycol. Soc. Am. Newsl., 34: 59-63. Koske R. E, Gemma J. N, Jackson N., 1997. Mycorrhizal fungi associated with three species of turfgrass. Can. J. Bot. , 75: 320-332.
Kosuta, S., Chabaud M., Lougnon G., Gough C., Denarie J., Barker D., Bécard G., 2003. A Diffusible Factor from Arbuscular Mycorrhizal Fungi Induces Symbiosis-Specific MtENOD11 Expression in Roots of Medicago truncatula. Plant Physiol., 131(3): 952-962.
Kubikova E., Moore, J.L., Ownlew B.H., Mullen M.D., Augé R.M., 2001. Mycorrhizal impact on osmotic adjustment in Ocimum basilicum during a lethal drying episode. J Plant Physiol 158: 1227–1230.
Land S., von Alten H., Schönbeck F., 1993. The influence of host plant, nitrogen fertilization and fungicide application on the abundance and seasonable dynamics of vesicular-arbuscular mycorrhizal fungi in arable soils in northern Germany. Mycorrhiza, 2:157–166.
Langeron M., 1952. Précis de mycologie. Masson (Ed.), Paris, pp. 367-397.

151
Le Tacon F., 1985. Les mycorrhizes: une coopération entre plantes et champignons. La Recherche, 166: 624-632.
LLee TTaaccoonn FF.. ,, LLee TTaaccoonn TT..,, MMaauurroonn VV..,, RRoouusssseeaauu YY..,, BB aacckkee rr MM ..,, BBoouucchhaarrdd DD..,, 11999999.. FFeerrttiilliissaattiioonn
rraaiissoonnnnééee eett mmyycciirrhhiizzee.. 44 èèmmee rreennccoonnttrree ddee llaa ffeerrttiilliissaattiioonn rraaiissoonnééee.. BBllooiiss,, nnoovveemmbbrree-- ddéécceemmbbrree 11999999,,
pppp.. 221111--222222..
LLee aakkee JJ..,, JJoohhnnssoonn DD..,, DDoonnnnee llllyy DD..,, MM uucckkllee GG..,, BBooddyy LL..,, RRee aadd DD..,, 22000044.. NNeettwwoorrkkss ooff ppoowweerr aanndd iinnff lluueennccee:: tthhee rroollee ooff mmyyccoorrrrhhiizzaall mmyycceelliiuumm iinn ccoonnttrroolllliinngg ppllaanntt ccoommmmuunniitt iieess aanndd aaggrroossyysstteemm
ffuunnccttiioonniinngg.. CCaann.. JJ.. BBoott.. ,, 8822::11001166-- 11004455..
Leyval, C., Joner E.J., del Val C., Haselwandter K., 2002 . Potential or arbuscular mycorrhizal fungi for bioremediation. In: Mycorrhiza Technology in Agriculture, from Genes to Bioproducts. Eds.Gianinazzi, S, Schuepp, H., Barea, J.M., Haselwandter, K., Birkhauser Verlag, Basel, 175–186. Li L. F., Tao L., Zhiwei Z., 2007. Differences of arbuscular mycorrhizal fungal diversity and community between a cultivated land, an old field, and a never-cultivated field in a hot and arid ecosystem of southwest China. Mycorrhiza,17: 655–665.
Liddell H.G., Scott R., 1991. A Greek-English Lexicon. Clarendon Press, pp. 21-27.
Ligrone R., Carafa A., Lumini E., Bianciotto V., Bonfante P., Duckett J. G., 2007. Glomeromycotean associations in liverworts: a molecular cellular and taxonomic analysis. Am. J. Bot., 94:1756–1777. Linderman, R.G., 1994. Role of VAMfungi in biocontrole. In : Mycorrhizae and plant health. Eds. Pfleger F.L., Linderman R. G. The American phytopathological society, St. Paul, MN , pp. 1-25. Linderman, R.G., 2000. Effects of mycorrhizas on plant tolerance to disease. In: Arbuscular Mycorrhizas: Physiology and Function. Ed. Kapulnik Y. Kluwer Academic Publishers, Dordrecht, the Netherlands, pp. 345-367. Liu W., 2010. Do genetically modified plants impact arbuscular mycorrhizal fungi?. Ecotoxicology, 19:229–238.
Logi C., Sbrana C., Giovannetti M., 1998. Cellular events involved in survival of individual arbuscular mycorrhizal symbionts growing in the absence of the host. Appl. Environ. Microbiol., 64:3473–3479.
Lohman M.L., 1927. Occurrence of mycorrhiza in Iowa forest plants. University of Iowa Studies in Natural History 11:33-58.
Lopez-Raez J.A. , Charnikhova T. , Gomez-Roldan V. , Matusova R. , Kohlen W. , De Vos
R., Verstappen F., Puech-Pages V., Becard G., Mulder P., Bouwmeester H.,
. 2008. Tomato
strigolactones are derived from carotenoids and their biosynthesis is promoted by phosphate
starvation. New Phytol., 178 (4): 863–874.
Louis I., Lim G., 1988. Effect of storage of inoculum on spore germination of a tropical isolate of Glomus clarum. Mycologia, 80:157–161.
Loussert R. Brousse G., 1978. L‟olivier. Techniques agricoles et productions méditerranéennes. (Eds.) Maisonneuve et Larousse, Paris, France, 480 p.

152
Lovelock C.E., Andersen K., Morton J.B., 2003. Influence of host tree species and environmental variables on arbuscular mycorrhizal communities in tropical forests. Oecol., 135: 268 – 279. Lugo M.A., Cabello M.N., 2002. Native arbuscular mycorrhizalfungi (AMF) from mountain grassland (Cόrdoba, Argentinia) I. Seasonal variation of fungal spore density. Mycologia, 94: 579-586.
LLuuggoo MM..AA..,, FFee rrrree rroo MM..,, MMeennooyyoo EE..,, EEssttéévvee zz MM..CC..,, SSiiññee rriizz FF..,, AAnnttoonn AA..,, 22000088.. AArrbbuussccuullaarr mmyyccoorrrrhhiizzaa ll ffuunnggii aanndd rrhhiizzoosspphheerriicc bbaacctteerriiaa ddiivveerrssiitt yy aalloonngg aa aalltt iittuuddiinnaall ggrraaddiieenntt iinn SSoouutthh AAmmeerriiccaann
PPuunnaa ggrraassssllaanndd.. MMiiccrroobbiiaa ll EEccooll.. ,, 5555:: 770055––771133..
Madan R., Pankhirst C. ,Bawke B., Smith S., 2002. Use of fatty acids of for identification of AM fungi and estimation of biomass of AM spores in the soil. Soil Bio. and Biochem., 34:125-128.
Mangan S. A.,Eom A. H., Adler G. H., Yavitt J. B., Herre E. A., 2004. Diversity of Mycorrhizal fungi across a fragmented forest in Panama: insular spore communities differ from mainland communities. Oecol., 141: 687-700.
Martin F., Perotto S., Bonfante P., 2007. Mycorrhizal fungi: a fungal community at the interface between soil and roots. In: Pinton R, Varanini Z, Nannipieri P (eds) The Rhizosphere: biochemistry and organic substances at the soil-plant interface. CRC Press, Boca Raton, pp. 201 – 136.
MMaarruullaannddaa AA..,, AAzzccóónn RR..,, RRuuiizz--LLoozzaannoo JJ..MM..,, 22000033..CCoonnttrriibbuutt iioonn ooff ssiixx aarrbbuussccuullaarr mmyyccoorrrrhhiizzaa ll ffuunnggaall iissoollaatteess ttoo wwaatteerr uuppttaakkee bbyy LLaaccttuuccaa ssaattiivvaa LL.. ppllaannttss uunnddeerr ddrroouugghhtt ssttrreessss.. PPhhyyssiiooll.. PPllaanntt.. 111199,,
552266––553333..
Matheny P.B., Wang Z., Binder M., 2007. Contributions of rpb2 and tef1 to the phylogeny of mushrooms and allies (Basidiomycota, Fungi). Mol. Phylogenet. Evol., 43:430–45.
MMaatthhiimmaarraann NN..,, RRuuhh RR..,, VVuulllliioouudd PP..,, FFrroossss aarrdd EE..,, jjaannss aa JJ.. ,, 22000055.. GGlloommuuss iinnttrraarraaddiicceess ddoommiinnaatteess aarrbbuussccuullaarr mmyyccoorrrrhhiizzaa ll ccoommmmuunniitt iieess iinn aa hheeaavvyy tteexxttuurreedd aaggrriiccuulltt uurraall ssooiill.. MMyyccoorrhhiizzaa ,, 1166:: 6611-- 6666.. Matsuda Y., Noguchi Y., Ito S., 2009. Ectomycorrhizal fungal community of naturally regenerated Pinus thunbergii seedlings in a coastal pine forest. J. For. Res., 14:335–341.
Mazliak P., 1982. Physiologie végétale, Tome II, Croissance et Développement, Ed. Coll. Méthode Herman, Paris, 465 p. McGonigle T., Fitter A.H., 1990. Ecological specificity of vesicular-arbuscular mycorrhizal associations. Mycol. Res., 94:120–122.
Meier R., Charvat I., 1992. Gemination of Glomus mosseae spores: procedure and ultrastructural analysis. Int. J. Plant Sci., 153:541–549.
Mendil M., Sebri A., 2006. L‟olivier en Algérie. ITAF, Alger, Algérie, 99 p. Merryweather J., 2001. Comment: Meet the Glomales- the ecology of mycorrhiza. British Wildlife. 13:2 86-93. Merryweather J.W., Fitter A.H., 1998. The arbuscular mycorrhizal fungi of Hyacinthoides non-scripta-I.Diversity of fungal taxa. New Phytol., 138:117–129.
Meyen J., 1829. Ueber das Herauswachsen parasitischer Gewachse aus den Wurzeln anderer Pflanzen. Flora, 12:49-63.

153
Miller D. D., Domoto P. A., Walker C., 1985. Mycorrhizal fungi at eighteen apple rootstock plantings in the United States. New Phytol., 100: 379-391.
Miller M. H., 2000. Arbuscular mycorrhizae and the phosphorus nutrition of maize: A review of Guelph studies. Can.J. Plant Sc., 80: 47-52. Monreal M., Berch S.M. and Berbee M., 1999. Molecular diversity of ericoid mycorrhizal fungi. Can. J. Bot., 77: 1580–1594.
Moreira M., Nogueira M. A., Tsai S. M.,Gomes-da-Costa S. M., Cardoso E. J. B. N., 2007. Sporulation and diversity of arbuscular mycorrhizal fungi in Brazil Pine in the field and in the greenhouse. Mycorrhiza,17:519–526. Moreira-Souza M., Trufem S.F.B., Gomes-da-Costa S.M., Cardoso E.J.B.N., 2003. Arbuscular mycorrhizal fungi associated with Araucaria angustifolia (Bert.) O. Ktze. Mycorrhiza, 13:211–215 . Morton J.B., Benny G.L., 1990. Revised classification of arbuscular mycorrhizal fungi (Zygonycetes): A New order, Glomales, Two New busorders, Glomineae and Gigasporineae, with an emendation of Glomaceae. Mycotaxon, 37: 471-491.
Morton J. B., Redecker D. 2001. Two families of Glomales, Archaeosporaceae and Paraglomaceae, with two new genera Archaeospora and Paraglomus, based on concordant molecular and morphological characters. Mycologia 93: 181–195.
Mosse B. 1959. The regular germination of resting spores and some observations on the growth requirements of an Endogone sp. causing vesicular-arbuscular mycorrhiza. Trans. Br. Mycol. Soc,. 42: 273-286.
Mosse B. 1970. Honey-coloured, sessile Endogone spores. I. Life history. Arch. Microbiol., 70: 167-175.
Mosse B., 1988. Some studies relating to “independent” growth of vesicular-arbuscular endophytes. Can. J. Bot., 66:2533–2540.
Mosse B., Bowen G. D. 1968. A key to the recognition of some Endogone spore types. Trans. Br. Mycol. Soc. 51, 469-483.
Mugnier J., Mosse B., 1987. Vesicular-arbuscular mycorrhizal infections in transformed rootinducing T-DNA roots grown axenically. Phytopathol., 77:1045–1050.
MMuummmmee yy DD.. LL.. ,, RRiilllliigg MM..CC..,, HHoollbbee nn WW.. EE..,, 22000055.. NNeeiibboorr iinngg ppllaanntt iinnff lluueenncceess oonn aarrbbuussccuullaarr
mmyyccoorrrrhhiizzaa ll ffuunnggaa ll ccoommmmuunniitt iieess ccoommppooss iitt iioonn aass aassssiisssseedd bbyy TT--RRFFLLPP aannaallyysseess.. PPllaanntt SSooiill,, 227711:: 8833-- 9900..

154
MMuunnggaaii NN.. WW..,, MMoottaavvaallllii PP..PP..,, KKrree mmee rr RR..II..,, NNeellssoonn KK.. AA..,, 22000055.. SSppaattiiaall vvaarriiaatt iioonn ooff ssooiill eennzzyymmee aaccttiivviitt iieess aanndd mmiiccrroobbiiaa ll ffuunncctt iioonnaa ll ddiivveerrssiittyy iinn tteemmppeerraattee aalllleeyy ccrrooppppiinngg ssyysstteemmss.. BBooiill..FFeerrtt iilliittyy ooff
SSooiill.. ,, 4422:: 112299-- 113366..
Muthukumar T., Udaiyan K., 2000. Arbuscular mycorrhizas of plants growing in the western Ghats region, Southern India. Mycorrhiza, 9:297–313. Muthukumar T., Prakash S., 2009. Arbuscular mycorrhizal morphology in crops and associated weeds in tropical agro-ecosystems. Mycoscience, 50:233–239 .
Muthukumar T., Sha L., Yang X., Cao M., Tang J., Zheng Z., 2003 . Mycorrhiza of plants in different vegetation types in tropical ecosystems of Xishuangbanna, southwest China. Mycorrhiza 13:289–297. Muthukumar T., Senthilkumar M., Rajangam M., Udaiyan K., 2006 . Arbuscular mycorrhizal morphology and dark septate fungal association in medicinal and aromatic plants of Western Ghats, Southern India. Mycorrhiza, 17:11–24.
Nagahashi G., Douds D.D., 2000. Partial separation of root exudate components and their effects upon the growth of germinated spores of AM fungi. Mycol .Res., 104: 1453–1464.
Nagahashi G., Douds D. D., 2004. Synergism between blue light and root exudate compounds and evidence for a second messenger in the hyphal branching response of Gigaspora gigantean. Mycologia, 96(5) : 948–954. Nagahashi G., Douds D.D., 2005. Environmental Factors That Affect Presymbiotic Hyphal Growth and Branching of Arbuscular Mycorrhizal Fungi. In: In Vitro Culture of Mycorrhizas. eds. Declerck S.,. Strullu D.-G, Fortin A., Soil Biology, 4:95-109. Nagahashi G., Douds D.D. Jr, Buee M., 2000. Light-induced hyphal branching of germinated AM fungal spores. Plant Soil, 219:71–79.
Nasr H., Diem H., 1988. Effer de l‟endomycorhization Vésiculo-arbusculaire sur la croissance et la fixation biologique de l‟azote par Acacia cyanophylia Lind.Institut National de RecherchesForestières.Tunis., pp. 232-242. Norman, J.R., Atkinson, D., Hooker, J.E., 1996. Arbuscular mycorrhizal fungal-induced alteration to root architecture in strawberryand induced resistance to the root pathogen Phytophthora fragariae. Plant Soil, 185: 191–198
Nouaïm R, Chaussod R., 1996. Rôle des mycorhizes dans l‟alimentation hydrique des plantes, notamment des ligneux en zones arides. Options Méditerranéennes, 20 : 9-26.
Novero M., A. Genre , K. Szczyglowski , P. Bonfante 2008. Root Hair Colonization by Mycorrhizal Fungi. Plant Cell. Monographe, 12: 1-24. NuutilaA.M., Vestberg M., KauppinenV., 1995. Infection of hairy roots of strawberry (Fragaria × Ananassa Duch.) with arbuscular mycorrhizal fungus. Plant Cell. Rep., 14:505–509.

155
O’Connor P. J., Smith S. E., Smith F.A., 2001. Arbuscular mycorrhizal association in the southern Simpson Desert. Aust. J. Bot., 49:493–499.
Oláh, B., Brière C., Bécard G., Dénarié D., Gough C., 2005. Nod factors and a diffusible factor from arbuscular mycorrhizal fungi stimulate lateral root formation in Medicago truncatula via the DMI1/DMI2 signalling pathway. Plant J., 44:195–207. Oehl F., Sieverding E., 2004. Pacispora, a new vesicular arbuscular mycorrhizal fungal genus in the Glomeromycetes. J. Appl. Bot. 78, 72-82. Oehl F., de Souza F.A., Sieverding E., 2008. Revision of Scutellospora and description of five new genera and three new families in the arbuscular mycorrhiza-forming Glomeromycetes. Mycotaxon, 106: 311–360.
Oehl F., Sieverding E., Ineichen K., Mäder P., Boller T., Wiemken A., 2003. Impact of land use intensity on the species diversity of arbuscular mycorrhizal fungi in agroecosystems of Central Europe. Appl. Envir. Microbiol., 69:2816–2824.
Oehl F., Sieverding E., Ineichen K., Ris E-A., Boller T., Wiemken A., 2005. Community structure of arbuscular mycorrhizal fungi at different soil depths in extensively and intensively managed agroecosystems. New Phytol., 165:273–283.
Oehl F., Sieverding E., Mäder P., Dubois D., Ineichen K., Boller T., Wiemken A., 2004. Impact of long-term conventional and organic farming on the diversity of arbuscular mycorrhizal fungi. Oecologia, 138:574–583
Oihabi A., 1991. Etude de l'influence des mycorrhizes a vesicules et arbuscules sur le bayoud et la nutrition du palmier dattier. Ph.D. thesis, Université Cadi Ayyad, Marrakech, Morocco, 200 p. Oihabi A., Meddich A., 1996. Effet des mycorhizes à arbuscules (MA) sur la croissance et la composition minérale du trèfle.Cahier Agricultures, 5 : 382-386.
Olivier B., Bertheau Y., Diem H. G., Gianinazzi-pearson V., 1983. Influence de la variété de Vigna unguiculata dans l‟expression de trios association endomycorhiziennes à vésicules et à arbuscules. Can. J. Bot., 61 : 354-358
OOmmrraannii--SSaabbbbaagghhii AA.. OO..,, SShhaahhrriiaarrii MM ..,, AAnnbbaarraann MM.. FF,, MMoohhaammmmaaddii SS.. AA..,, NNaannkkaallii AA..,, MM aarrddii MM..,, GGhhaarree yyaazziiee BB ..,, 22000077.. MMiiccrroossaatteelllliittee mmaarrkkeerrss bbaasseedd aasssseessssmmeenntt ooff ggeenneettiicc ddiivveerrssiittyy iinn IIrraanniiaann oolliivvee
((OOlleeaa eeuurrooppeeaa LL..)) ..CCoolllleeccttiioonnss..SSccii.. HHoorrttiiccuulltt.. ,, pppp..11--99..
Onguene N.A., Tsimi J.P.M., Balla M.J.E., 2002. Statut mycorhizien de l‟okoumé (Aucoumea klaineana P.) en régénération artificielle au sud Cameroun. Tropicultura, 20(3): 104-108.
Ozgenturk N. O., Oruc F., Sezerman U., Kucukural A., Korkut S. V., Toksoz F., Un C., 2010. Generation and Analysis of Expressed Sequence Tags from Olea europea L. Corporation Comparative and Functional Genomics, 9 : 1-9.
Pagano M. C., Bellote A. F., Scotti M. R., 2009. Aboveground nutrient components of Eucalyptus camaldulensis and E. grandis in semiarid Brazil under the nature and the mycorrhizal inoculation conditions. J. Forestry Res., 20(1):15–22.
Palenzuela J., Ferrol N., Boller T., Azcón-Aguilar C., Oehl F.. 2008. Otospora bareai, a new fungal species in the Glomeromycetes from a dolomitic shrub land in Sierra de Baza National Park (Granada, Spain). Mycologia, 100:296-305.

156
Panna D. , Highland K., 2010. Mycorrhizal colonization and distribution of arbuscular mycorrhizal fungi associated with Michelia champaca L. under plantation system in northeast India . J. For. Res., 21(2): 137-142. Pasquini, G., Baldoni L., Ferretti L., Pannelli G., Faggioli F., Barba M., 2002. Evaluation of the strawberry latent ringspot virus (SLRSV) in some olive cultivars. Atti Convegno Internazionale Olivicoltura, Spoleto, Italy, pp. 462-465.
Pawlowska T.E., Douds D.D., Charvart I., 1999. In vitro propagation and life cycle of the arbuscular mycorrhizal fungus Glomus etunicatum. Mycol. Res., 103:1549–1556.
Peretto R., Bettini V., Favaron F., Alghisi P., Bonfante P., 1995. Polygalacturonase activity and location in arbuscular mycorrhizal roots of Allium porrum L. Mycorrhiza, 5: 157-163.
Perotto S., Girlanda M., Martino E., 2002. Ericoid mycorrhizal fungi: some new perspectives on old acquaintances. Plant Soil, 244: 41–53.
Petri L., 1903. Ricerche sul signifacyo morfologica del prosporoidi (sporangioli di Janse) nelle mmicorrize endotrofiche. Nuovo G. Bot.ital., 10 : 541-550.
Peyronel B., 1923. Fructification de l‟endophyte à vésicules des mycorhizes endotrophes. Bull. Soc. Myc. Fr., 39 : 119-126.
Peyronel B., 1924. Specie di « Endogone » productricci di micorrize endotrofiche. Boll. Stn. Patol. Veg. Roma 5, 73-75
Pfeffer W. 1877. Ueber fleischfressende Planzen und über die ern hrung durch Aufnahme organischer Stoffe überhaupt. Landwirtschaftliche Jahrbücher 6: 969–988.
PPffee iiffffee rr CC..MM ..,, BB lloossss HH..EE..,, 11998888.. GGrroowwtthh aanndd nnuuttrriitt iioonn ooff gguuaayyuullee ((PPaarrtthheenniiuumm aarrggeennttaattuumm)) iinn aa ssaalliinnee ssooiill aass iinnff lluueenncceedd bbyy vveessiiccuullaarr--aarrbbuussccuullaarr mmyyccoorrrrhhiizzaa aanndd pphhoosspphhoorr uuss ffeerrttiilliizzaatt iioonn.. NNeeww PPhhyyttooll.. ,,
110088::331155--332211
Phillips J.M., Hayman D.S. ,1970. Improved procedures for clearing roots and staining parasitic and vesicular–arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc., 55:158–16.
Plenchette C., 2005. Mycorhizes et nutrition phosphatée des plantes. Journées Techniques Fruits et Légumes et Viticulture Biologiques, 6-7 décembre, Beaune, pp. 103-109
Plenchette C., Perrin R., 1989. The concept of soil infectivity and a method for its determination as applied to endomycorhizas. Can. J. Bot., 67: 112-115.
Plenchette C., Fortin J.A., Furlan V., 1983. Growth responses of several plant species to mycorrhizae in a soil of moderate P-fertility. Part I. Mycorrhizal dependency under field conditions.Plant Soil, 70:199–209.
PPllee nncchhee ttttee CC..,, DDeeccllee rrcckk SS..,, DDiioopp TT..AA..,, SSttrruulllluu DD..GG..,, 11999966.. IInnffeeccttiivviitt yy ooff mmoonnooaaxxeenniicc ssuubbccuullttuurreess ooff
tthhee aarrbbuussccuullaarr mmyyccoorrhhhhiizzaa ll ffuunngguuss GGlloommuuss vveerrssiiffoorrmmee aassssoocciiaatteedd wwiitthh RRii--TT--DDNNAA--ttrraannssffoorrmmeedd ccaarrrroott
rroooott.. AAppppll.. MMiiccrroobbiiooll.. BB iiootteecchh.. ,, 4466:: 554455--554488
Plenchette C., Bois J. F., Duponnois R., Cadet P., 2000. La mycorhization (Glomus aggregatum) du mil (Pennisetum glaucum). Note technique. Étude et gestion des sols, 7 (4) : 379-384.

157
Plenchette C., Clermont-Dauphin C., Meynard J.M, Fortin J.A., 2005. Managing arbuscular mycorrhizal fungi in cropping systems. Can. J. Plant Sci., 85: 31-40.
Pons F., Gianinazzi-Pearson V., 1984. Effect of phosphorus, potassium, nitrogen and pH on the in vitro behaviour of vesicular-arbuscular endomycorrhizal fungi. Cryptogamie Mycol. 5:87–100.
Pope S., Smith S.E., Christophersen H.M., 2007. Arsenic uptake by Medicago truncatula: P supply and arbuscular mycorrhizal (AM) colonization do not reduce specific uptake from soil. In: Biogeochemistry of Trace Elements: Environmental Protection. Eds. Zhu Y.G., Lepp N., Naidu R . Remediation and Human Health, Germany, pp. 863–864. Porcel R., Ruiz-Lozano J.M., 2004. Arbuscular mycorrhizal influence on leaf water potential, solute accumulation and oxidative stress in soybean plants subjected to drought stress. J.Exp.Bot., 55:1743–1750.
Porcel R, Barea J. M., Ruiz-Lozano J. M., 2003. Antioxidant activities in mycorrhizal soybean plants under drought stress and their possible relationship to the process of nodule senescence. New Phytol., 157: 135–143. Porcel R., Aroca R., Cano C., Bago A., Ruiz-Lozano J. M., 2006. Identification of a Gene from the Arbuscular Mycorrhizal Fungus Glomus intraradices Encoding for a 14-3-3 Protein that Is Up-Regulated by Drought Stress During the AM Symbiosis. Microb. Ecol., 52: 575–582.
Porras Piedra A., Soriano Martín M.L., Fernández Izquierdo G., 2005. Application de mycorhizes à la culture de l‟olivier. Influence sur le développement des jeunes plants de la variété „Cornicabra‟. Olivæ, 104: 46-54 .
Porras Soriano A., Domench Menor B., Castillo Rubio J., Soraino Martin M L, Porras Piedra A., 2002. Influence des mycorhizes vésico-arbusculaires sur la croissance des boutures d‟olivier multipliées sous nébulation. Olivæ, 92: 33-37. Porter W.M., Robson A.D., Abbott L.K. , 1987. Field survey of the distribution of vesicular-arbuscular mycorrhizal fungi in relation to soil pH. J. Appl. Ecol., 24:659–662.
Pozo M. J., Slezack-Deschaumes S., Dumas-Gaudo E., Gianinazzi S., Azcón-Aguilar C., 2002. Plant defense responses induced by arbuscular mycorrhizal fungi. In S. Gianinazzi, H. Schüepp, J. M. Barea and K. Haselwandter (Eds.), Mycorrhiza Technology in Agriculture, from Genes to Bioproducts, pp. 103-111.
Pringle A., Bever J.D., 2002. Divergent phonologies may facilitate the coexistence of arbuscular mycorrhizal fungi in a North Carolina grassland. Am. J. Bot., 89:1439–1446. Püschel D., Rydlová J., Vosátka M., 2007. Mycorrhiza influences plant community structure in succession on spoil banks. Basic Appl. Ecol., 8: 510-520.
PPuutthhuurr JJ ..TT..,, PPrraassss aadd KK.. VV..SS.. KK..,, SShhaarrmmiillaa PP..,, PPaarrddaa SSaarraaddhhii PP..,, 11999988.. VVeessiiccuullaarr aarrbbuussccuullaarr
mmyyccoorrrrhhiizzaa ll ffuunnggii iimmpprroovveess eessttaabblliisshhmmeenntt ooff mmiiccrroopprr ooppaaggaatteedd LLeeuuccaaeennaa lleeuuccoocceepphhaallaa ppllaannttlleettss..
PPllaanntt CCeellll.. TT iissss.. OOrrgg.. CCuulltt.. ,, 5533:: 4411--4477
Radhika K.P., Rodrigues B. F., 2010.Arbuscular mycorrhizal fungal diversity in some commonly occurring medicinal plants of Western Ghats, Goa region. J. For. Res., 21(1): 45–52.
Rai M. K., 2001 Current advances in mycorhization in mycropropagation. In Vitro Cell. Dev. Biol. Plant, 37:158-167.

158
RRaammaammoooorrtthhyy VV..,, VViiss wwaannaatthhaann RR..,, RRaagguucchhee rraannddee rr TT..,,PPrraakkaass aamm VV..,, SSaammiiyyaappppaann RR..,, 22000011.. IInndduucctt iioonn ooff ssyysstteemmiicc rreessiissttaannccee bbyy ppllaanntt ggrroowwtthh pprroommoott iinngg rrhhiizzoobbaacctteerriiaa iinn ccrroopp ppllaanntt aaggaaiinnsstt ppeessttss
aanndd ddiisseeaasseess.. CCrroopp PPrrootteecctt.. ,, 2200:: 11-- 1111 ..
Ravikumar .R., Ananthakrishan S., Appasany T., Ganapathi A., 1997 . Effect of endomycorhizae (VAM) on bamboo seedling growth and biomass productivity . Forest Ecol. Manag., 98:205-208.
RRee aadd DD.. JJ.. ,, HHaassee llwwaannddttee rr KK..,, 11998811.. OObbsseerrvvaattiioonn oorr tthhee mmyyccoorrhhiizzzzaa ll ssttaattuuss ooff ssoommee aallppiinnee ppllaanntt ccoommmmuunniitt iieess.. NNeeww PPhhyyttooll.. ,, 8888::334411--335522..
Read D.J., Kianmehr H., Malibari A., 1977. The biology of mycorrhiza in Helianthemum Mill. New Phytol., 78: 305–312.
Redecker D, Raab P, 2006. Phylogeny of the Glomeromycota (arbuscular mycorrhizal fungi): recent developments and new gene markers. Mycologia, 98: 885-895. Redecker D., Morton J.B., Bruns T.D., 2000. Ancestral lineages of arbuscular mycorrhizal fungi (Glomales). Mol. Phyloge. Evol., 14(2): 276-284
Régis S., 2007. L‟œil de paon : six ans de suivit pour bâtir un modèle. Phytoma : 12-17.
Reinhardt D., 2007. Programming good relations development arbuscular mycorrhizal symbiosis. Current Opinion in Plant Biology, 10: 98-105. Ren X., 2006. Effect of Bt transgenic rice (KMD) on soil bacterial community and rhizosphere AM fungi. Master degree thesis of Zhejiang University. HangZhou, Chine. pp. 322-330.
Requena N., Serrano E., Ocόn A., Breuninger M., 2007. Plant signals and fungal perception during arbuscular mycorrhiza establisnment. Phytochemistry ,68: 33-40.
Rillig M.C., 2004. Arbuscular mycorrhizae and terrestrial ecosystem processes. Ecol. Lett., 7: 740-754.
Rillig M.C., Mummey D.L. 2006. Mycorrhizas and soil structure. New Phytol., 171:41–53.
Rillig M.C., Mummey D.L., Ramsey P.W., Klironomos J.N., Gannon J.E., 2006. Phlogeny of arbuscular mycorhizal fungi predicts community composition of symbiosis associated bacteria. FEMS Microbiol. Ecol., 57(3): 389-395. Rinaldi A. C., Comadini O., Kuyper T. W., 2008. Ectomycorrhizal fungal diversity: separating the wheat from the chaff. Fungal Divers., 33:1–45.
Rhizopoulou S., 2007. Olea europaea L. A Botanical Contribution to Culture. Am.-Eur. J. Agric. & Environ. Sci., 2 (4): 382-387.
Rodríguez-Echeverrìa. S., Freitas H., 2006. Diversity of AMF associated with Ammophila arenaria ssp. arundinacea in Portuguese sand dunes Mycorrhiza, 16:543–552
Roldán- Fajardo B.E., Barea J.M., 1986. Mycorhizal dependency in the olive tree (Olea europaea L.) Physiological and genetical aspects of the mycorrhizae. Proc. of the 1st Europ. Symp. on Mycorrhizae, Dijon, INRA, Paris, France, pp. 323-326.
Rony C., Baalbaki R., Kalaitzis P. , Talhouk S. N., 2009. Molecular characterization of Lebanese olive germplasm. Tree Genetics and Genomes, 5:109–115.

159
Rothwell, F. M., Trappe J. M., 1979. Acaulospora bireticulata sp. nov. Mycotaxon, 8:471-475.
Rugini E., 1984. In vitro propagation of some olive (Olea europea sativa L.) cultivars with different root-ability, and medium development using analytical data from developing shoots and embryos. Sci. Hortic., 24: 123-134.
Rugini E., 2002. Miglioramento genetico ed applicazioni biotecnologiche per la salvaguardia della biodiversità in olivo (Genetic improvement and biotechnology application for safeguard biodiversity in olive. Atti Convegno Internazionale di Olivicoltura , pp. 74-81.
Rugini E., Baldoni L., 2004. Olea europea Olive. In: Biotechnology of Fruit and Nut Crops. Chap. 15 (R.E. Litz ). CABI Publishing, Noworty Way, Wallingford, Oxfordshire OX10 8DE, UK. Pag 404-428.
Rugini E., Pesce P. G., 2006. Genetic improvement of olive. Pomol. Croit., 12: 43- 74. Rugini E. Mencuccini M., Biasi, R., Altamura M. M., 2005. Olive (Olea europea L.) S.M. Jain and P.K. Gupta (eds.), Protocol for Somatic Embryogenesis in Woody Plants, Netherlands, pp. 345–360.
Rugini E., Gutierrez-Pesce P., Spampinato P.L., Ciaramiello A., D’Ambrosio C., 1999. New perspective for biotecnologies in olive breeding: morphogensis, in vitro selection and gene transformation. Acta Horticult., 474: 107-110.
Ruiz Lozano J., 2003. Arbuscular mycorrhizal symbiosis and alleviation of osmotic stress. Nero perspectives for molecular studies. Mycorrhiza 13: 309- 317. Ruíz-Lozano J. M., Roussel H., Gianinazzi S., Gianinazzi-Pearson V., 1999. Defense genes are differentially induced by a mycorrhizal fungus and Rhizobium sp. in wild-type and symbiosis-defective pea genotypes. Mol. Plant-Microb. Interact. 12:976–984.
Ryan M. H., van Herwaarden A. F., Angus J.F., Kirkegaard J.A., 2005. Reduced growth of autumn-sown wheat in a low-P soil is associated with high colonisation by arbuscular mycorrhizal fungi. Plant Soil, 270: 275–286.
Saad D., 2009. Études des endomycorhizes de la variété Sigoise d‟olivier (Olea europea L.) et essai de leur application à des boutures semi-ligneuses multipliées sous nébulisation. Mémoire de Magister en Biotechnologie, Univ. Oran, 124 p.
Saad D., Sidhoum W., Fortas Z., Bellahcene M., 2010. Essais d‟endomycorhization des boutures herbacées de la variété Sigoises d‟olivier d‟Algérie. 1
er congrés international. Symbioses
Mycorhiziennes : Ecosystèmes et Environnement en milieu Méditerranéen (MYCOMED).les 11 -13 octobre 2010.Marrakech, Maroc.pp.191.
SSaahhllii ZZ..,, 22000099.. Produits de terroir et développement local en Algérie Cas des zones rurales de montagnes et de piémonts. Options méditerranéennes, 89:305-338. Sancholle M., Dalpé Y., Grandmougin-Ferjani A., 2001. Lipids of Mycorrhizae. In: Hock (ed) The Mycota. IX. Fungal associations. Springer, Berlin Heidelberg , New York, pp. 63–93. Sanders I.R. 2002. Ecology and evolution of multigenomic arbuscular mycorrhizal fungi.Am. Nat ., 160: 128–141. Sanders I.R., 2004. Intraspecific genetic variation in arbuscular mycorrhizal fungi and its consequences for molecular biology, ecology, and development of inoculum. Can. J. Bot., 82:1057–1062.

160
Sasanelli N., D’Addabbo N., Dell’Orco P., Mencuccini M., 2000. The in vitro use of olive explants in screening trials for resistance to the root-knot nematode, Meloidogyne incognita. Nematropica, 30: 101-106.
Sayhan S., Ozzambak E., 1994. Culture de tissus de deux variétés d‟olivier turques.Olivae, 52 :28-29.
Scatena F.N., Moya S., Estrada C. and Chinea J.D., 1996. The first five years in the reorganization of aboveground biomass and nutrient use following hurricane Hugo in the Bisley Experimental Watersheds, Luquillo Experimental Forest, Puerto Rico. Biotropica, 28: 424–440.
Schellenbaum, L., Berta, G., Ravolanirina, F., Tisserant, B., Gianinazzi, S., Fitter, A.H., 1991. Influence of endomycorrhizal infection on root morphology in a micropropagated woody plant-species (Vitis vinifera L.). Annals of Bot., 68: 135–141 Scheltema M. A., Abbott L. K., Robson A. D., 1987. Seasonal variation in the infectivity of VA mycorrhizal fungi in annual pastures in a Mediterranean environment. Aust. J. Agric. Res. 38:707–715.
Schenck N. C., Smith G. 1982. Additional new and unreported species of mycorrhizal fungi (Endogonaceae) from Florida. Mycologia 74, 77-92.
Schenck, N.C., Pérez, Y., 1990. Manual for the identification of VA mycorrhizal (VAM) fungi. Univ. of Florida Press, Florida, USA, pp. 241. Schenck N.C., Graham S.O., Green N.E., 1975. Temperature and light effect on contamination and spore germination of vesicular-arbuscular mycorrhizal fungi. Mycologia, 67:1189–1192.
Schlicht A., 1889. Beitrag zur Kenntniss der Verbreitung und Bedeutung der Mycorhizen. Landwirtschaftliche Jahrbcher, 18:478–506.
Schüßler A., Wolf E., 2005. Geosiphonpyriformis– a Glomeromycotan Soil Fungus Forming Endosymbiosis with Cyanobacteria. In: In Vitro Culture of Mycorrhizas. eds. Declerck S., Strullu D.-G., Fortin A.. Soil Biology, 4: 271-289.
Schüßler A., Schwarzott D., Walker C., 2001. A new fungal phylum, the Glomeromycota: phylogeny and evolution. Mycol. Res. 105:1413–1421.
Schultz P. A., Bever J. D., Morton J. B. 1999. Acaulospora colossica sp. nov. from an old field in North Carolina and morphological comparisons with similar species, A. laevis and A. koskei. Mycologia, 91: 676-683. Schwarzott D., Walker C., Schüßler A., 2001. Glomus, the largest genus of the arbuscular mycorrhizal fungi (Glomales), is non-monophyletic. Mol. Phylogen. Evol. ,21:190–197.
Sharma S., Parkash V., Aggarwal A., 2008. Glomales I: amonograph of Glomus spp.(Glomaceae) in the sunflower rhizospher of Haryana, India. Helia, Nr. 49: 13-18.
Sharples J. M., Chambers S. M., Meharg A. A., Cairney J. W. G., 2000. Genetic diversity of root-associated fungal endophytes from Calluna vulgaris at contrasting field sites. New Phytol. 148: 153–162.
Sheikh N. A., Sanders F. E. 1988. Effect of temperature on germination mycorrhizal spores and VAM infection in roots. Biologia, 34:337–350.

161
Shi . Z. Y., Chen Y. L., Feng G., Liu R. J., Christie P., Li X. L., 2006. Arbuscular mycorrhizal fungi associated with the Meliaceae on Hainan Island, China. Mycorrhiza, 16: 81–87. Siddiqui Z.A., Akhtar M.S., 2008. Effects of OrganicWastes, Glomus intraradices and Pseudomonas putida on the Growth of Tomato and on the Reproduction of the Root-knot Nematode Meloidogyne Incognita. Phytoparasitica, 36(5):460-471. Siddiqui Z. A., Pichtel J., 2008. Mycorrhizae: an overview. In: Mycorrhizae. Eds. A. Siddiqui et al. Sustainable Agricul. Fores., Berlin, pp. 1–35.
SSiieevvee rrddiinngg EE..,, 11998899.. EEccoollooggyy ooff VVAAMM ffuunnggii iinn ttrrooppiiccaall aaggrrooeeccoossyysstteemmss.. AAggrriicc.. EEccoossyysstt.. EEnnvviirroonn.. ,, 2299::
336699--339900
Sieverding E., 1991. Vesicular-arbuscular mycorrhiza management in tropical agrosystems. GTZ publishers, Germany, 371 p.
Sieverding E., Oehl F. , 2006. Revision of Entrophospora and description of Kuklospora and Intraspora, two new genera in the arbuscular mycorrhizal Glomeromycetes. J. Appl. Bot. Food Qual. 80: 69-81.
Simon L., Levesque R., C., Lalonde M., 1993.Origin and diversification of endomyccorrhizal fungi and coincidence with vascular land plants. Nature, 363: 67-69.
Simoneau P., Louisy-Louis N., Plenchette C., Strullu D. G., 1994. Accumulation of new polypeptides in Ri T-DNA transformed roots of tomato (Lycopersicon esculentum) during the development of vesicular arbuscular mycorrhizae. Appl. Environ. Microbiol., 60:1810–1813.
SSiinngghh,, AA..,, SShhaarrmmaa,, JJ.. ,, RReexxee rr,, KK..HH..,, VVaarrmmaa,, AA..,, 22000000.. PPllaanntt pprroodduucctt iivviitt yy ddeetteerrmmiinnaannttss bbeeyyoonndd
mmiinneerraallss,, wwaatteerr aanndd lliigghhtt.. PPiirriiffoorrmmoossppoorraa iinnddiiccaa :: aa rreevvoolluutt iioonnaarryy ppllaanntt ggrroowwtthh pprroommoott iinngg ffuunngguuss.. CCuurr..
SSccii.. ,, 7799:: 110011--110066
Siqueira J.O., Hubbell D.H., 1984. Effect of nutrition on germination and growth of vesicular arbuscular mycorrhizal (VAM) fungi. In: Proc 6th North American Conf Mycorrhizae (NACOM), Oregon, USA, 368 pp.
Siqueira J.O., Hubbell D.H., Schenck N.C., 1982 . Spore germination and germ tube growth of a vesicular arbuscular mycorrhizal fungus in vitro. Mycologia, 74:952–959.
Siqueira J.O., Colozzi-Filho A.,oliveira E., 1989. Ocurrencia de micorrizas vesiculo arbusculares em agro e ecossistemasnaturaes de estadode minas gerais . Pesqui. Agropecu. Bras., 24 : 11499-1506
Siqueira J.O., Sylvia D.M., Gibson J., Hubbell D.H., 1985. Spores, germination, and germ tubes of vesicular arbuscular mycorrhizal fungi. Can. J. Microbiol. 31:965–971.
Smith F.A., Smith S.E., 1996. Mutualism and parasitism: Diversity in function and structure in the arbuscular (VA) mycorhzal symbiosis. Adv. Bot. Res., 22: 143.

162
Smith F. A., Smith S. E., 1997. Structural diversity in (vesicular)- arbuscular mycorrhizal symbioses. New Phytol. 137:373–388.
Smith S., E., Barker S., J., Zhu Y, G., 2006. Fast moves in arbuscular mycorrhizal symbiotic signaling.Trends Plant Sc., 11(8):369-371.
Smith S. E., Christophersen H. M., Pope S., Smith F. A., 2010. Arsenic uptake and toxicity in plants: integrating mycorrhizal influences. Plant Soil, 327:1–21. Smith S.E., Read D.J., 1997. Mycorrhizal symbiosis. Academic Press. London, pp.301-310. Smith S.E., Read D.J., 2008. Mycorrhizal symbiosis. 3rd edn. Academic Press, San Diego, pp.16-32.
Solaiman M. Z., Abbott L.K., 2004. Functional Diversity of Arbuscular Mycorrhizal Fungi on Root Surfaces Plant Surface. Eds. Varma A., Abbott L., Werner D., Hampp R., Berlin. Plant Surface Microbiology, pp.331-349. Sorensen J.N., Larsen J., Jakobsen I., 2005. Mycorrhiza formation and nutrient concentration in leeks (Allium porrum) in relation to previous crop and cover crop management on high P soils. Plant Soil, 273: 101–114 Sow H.A., Diop T.A., Ndiaye F., Manga A.G.B., Diallo A., 2008. Influence de la mycorhization arbusculaire sur la culture intensive de l‟oignon (Allium cepa L.) au Senegal. Journal des Science, 8(1): 1-6. Spain J. L., 1992. Patency of shields in water mounted spores of four species in Acaulosporaceae (Glomales). Mycotaxon, 28:331–339.
Spain J. L., 2003. Emendation of Archeospora and its type species, Archaeospora trappei. Mycotaxon, 87: 109-112.
Spain J. L., Sieverding E., Oehl F. 2006. Appendicispora: a new genus in the arbuscular mycorrhiza-forming Glomeromycetes, with a discussion of the genus Archaeospora. Mycotaxon, 97: 163-182.
Starr, F., K. Martz, and L.L. Loope. 1999. New plant records from East Maui for 1998. Bishop Mus. Occ. Pap., 59(2):11-15.
Strikic F. , Mavsar D. B., Perica S., Cmelik Z., Satovic Z., Javornik B., 2010.Genetic variation within the olive (Olea europaea L.) cultivar Oblica detected using amplified fragment length polymorphism (AFLP) markers. Afr. J. Biotec., 9(20 ): 2880-2883.
Strullu D.G., Diop T.A., Plenchette C., 1997. Réalisation de collections in vitro de Glomus intraradices (Schenck et Smith) et Glomus versiforme (Karsten et Berch) et proposition d‟un cycle de développement. C.R. Acad. Sci., 320: 41-47.
Strullu D.G., Romand C., 1986. Méthodes d‟obtention d‟endomycorhize à vésicules et arbuscules en cordition axeniques C.R .Acad. Sci. Paris, 303 : 245-250.
Strullu D. G., Romand C., 1987. Culture axénique de vésicules isolées à partir d‟endomycorhizes et ré-association in vitro des racines de tomate. C. R. Acad. Sci., Paris, 305:15–19.
Stukenbrock E. H.. Rosendahl S., 2005. Distribution of dominant arbuscular mycorrhizal fungi among five plant species in undisturbed vegetation of a coastal grassland. Mycorrhiza, 15: 497–503.

163
Stürmer S.L., 1998. Chacarcterization of diversity of fungi forming arbuscular endomycorrhizae in selected plant communities. Thesis in Forestry and Consumer Science. December, Morgantown, West Virginia. USA, 94p.
Stürmer S. L., Morton J. B. 1997. Developmental patterns defining morphological characters in spores of four species in Glomus. Mycologia, 89: 72–81. Stürmer S. L., Morton J. B. 1999. Scutellospora rubra, a new arbuscular mycorrhizal species from Brazil. Mycol. Res. 103 (8) : 949-954..
Stürmer S. L., Siqueira J. O., 2010. Species richness and spore abundance of arbuscular mycorrhizal fungi across distinct land uses in Western Brazilian Amazon. Mycorrhiza, 1-12.
Sward R.J., 1981. The structure of the spores of Gigaspora margarita. III. Germ tube emergence and growth. New Phytol., 88:667–673.
Sylvia D. MM.., Williams S. E., 1992. Vesicular-arbuscular mycorrhizae and environmental stresses. In: Bethlenfalvay GJ, Linderman RG (eds) Mycorrhizae in sustainable agriculture. Agronomy Society of America, Madison, WI, pp: 328-336.
SSyyllvviiaa DD.. MM..,, CChhee llllee mmii DD..OO..,, 22000011.. IInntteerraaccttiioonn aammoonngg rroooott iinnhhaabbiitt iinngg ffuunnggii aanndd tthhee iirr iimmpplliiccaatt iioonn ffoorr
bbiioollooggiiccaall ccoonnttrrooll ooff rroooott ppaatthhooggeennss.. AAddvv.. AAggrroonn.. ,, 7733::11-- 3333..
Tadych M., Blaszkowski J. 2000. Arbuscular mycorrhizal fungi of the Brda river valley in the Tuchola Forests. Acta Mycol. , 35: 3-23.
Tao L., Zhiwei Z., 2005. Arbuscular mycorrhizas in a hot and arid ecosystem in southwest China. Appl. Soil Ecol., 29: 135-141.
Tardieu F., Katerji N., Bethenod O., 1990. Relation entre l‟état hydrique de sol et quelques indicateurs de l‟état hydriques du maïs après floraison. Agronomie, 10: 617-626.
Taxter R., 1922. A revision of Ondogonaceae. Proc. Am. Acad. Arts Sci., 57: 291-351.
Taylor J., Harrier L. A., 2001. A comparison of development and mineral nutrition of micropropagated Fragaria x ananassa cv. Elvira (strawberry) when colonized by nine species of arbuscular mycorrhizal fungi. Appl. Soil Ecol., 18:205-215. Taylor T. N., Remy W., HASS H., Kerp H., 1995. Fossil arbuscular mycorrhizae from the early Devonian. Mycologia, 9: 560-573.
Tchabi A., Coyne D., Hountondji F., Lawouin L., Wiemken A., Oehl F., 2008. Arbuscular mycorrhizal fungal communities in sub-Saharan Savannas of Benin, West Africa, as affected by agricultural land use intensity and ecological zone. Mycorrhiza, 18:181–195.
Tedersoo L., May T. W., Smith,M. E., 2010. Ectomycorrhizal lifestyle in fungi: global diversity, distribution, and evolution of phylogenetic lineages. Mycorrhiza, 20:217–263.
Thomas S.R., Dunstan W.A., Dell B., Trappe J.M., Malajczuk N., 2003. Pisolithus hypogaeus sp nov.: A hypogeous representative of the genus Pisolithus from western Australia. Mycotaxon. 87:405-410.
Timonen S., Smith F. A., Smith S. E. 2001. Microtubules of mycorrhizal fungus Glomus intraradices in symbiosis with tomato roots. Can. J. Bot., 79:307–313.

164
Tiwari P., Adholeya A., 2002. In vitro co-culture of two AMF isolates Gigaspora margarita and Glomus intraradices on Ri T-DNA transformed roots. FEMS Microbiology Letters 206: 39-43.
Tommerup I.C., 1983. Spore dormancy in vesicular-arbuscular mycorrhizal fungi. Trans. Br .Mycol. Soc., 81:37–45.
Tommerup I., Kidby D. K., 1980. Production of aseptic spores of vesicular arbuscular endophytes and their viability after chemical and physical stress. Appl. Environ. Microbiol., 39:1111–1119.
Toth R., Miller R.M., 1984. Dynamics of arbuscule development and degeneration in a Zea mays mycorrhiza. Am. J. Bot.,71: 449–460.
Troech Z.I., Loynachan T.E. 2003. Endomycorrhizal fungal survival in continuous corn, soybean and fallow. Agron. J., 95:224–230.
Trouvelot A., Kouch J., Gianninazzi-pearson V. 1986. Mesure du taux de mycorhization VA d‟un système radiculaire : recherche de méthodes d‟estimation ayant une signification fonctionelle.Les mycorhizes :Physiologie et génétique.1er Séminaire Dijon. Paris , INRA , pp. 217-221.
Tulasne L. R. Tulasne C. 1844.Fungi nonnulli hipogaei, novi v. minus cogniti auct. G. Bot. Ital., 2 : 55-63. Turrini A., Avio L., Bedini S., Giovannetti M., 2008. In situ collection of endangered arbuscular mychorrhizal fungi in a Mediterranean UNESCO Biosphere Reserve. Biodivers. Conserv., 17:643–657.
Turrini A., Sbrana C., Strani P., Pezzarossa B., Risaliti R., Giovannetti M., 2010. Arbuscular mycorrhizal fungi of a Mediterranean island (Pianosa), within a UNESCO Biosphere Reserve. Biol. Fertil. Soils, 46:511–520.
Turrini A., Sbrana C., Pitto L., Ruffini C. M., Giorgetti L., Briganti R., Bracci T., Evangelista M., Nuti M. P., Giovannetti M. 2004. The antifungal Dm-AMP1 protein from Dahlia merckii expressed in Solanum melongena is released in root exudates and differentially affects pathogenic fungi and mycorrhizal symbiosis. New Phytol., 163:393–403.
Uhlmann E., Görke C., Petersen A., Oberwinkler F., 2006. Arbuscular mycorhizae from arid parts of Namibia. J. Arid Envir., 64: 221-237. Ultra V.U., Tanaka S., Sakurai K., 2007. Effects of arbuscular mycorrhiza and phosphorus application on arsenic toxicity in sunflower (Helianthus annuus L.) and on the transformation of arsenic in the rhizosphere. Plant Soil 290:29–24.
Vandenkoornhuyse P., Husband R., Daniell T. J., Watson I. J., Duck J. M., Fitter A. H., Young J. P. W., 2002. Arbuscular mycorrhizal community composition associated with two plant species in a grassland ecosystem. Mol. Ecol., 11: 1555–1564. Van der Heijden M. G. A., Bardgett R. D., Van Straalen N. M., 2008. The unseen majority: soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecol. Lett. 11:296–310.
Van der Heijden M. G. A., Kliromonos J. N., Ursic M., Moutoglis P., Streiwolf-Engel R., Boller T., Wiemken A., Sanders I. R., 1998. Mycorhizal fungal diversity determines plant biodiversity, ecosystem variability and productivity. Nature, 396: 69-72.

165
Van Tuinen D., Jacquot E., Zhao B., Gollotte A., Gianinazzi-Pearson V., 1998. Characterization of root colonization profiles by a microcosm community of arbuscular mycorrhizal fungi using 25S rDNA-targeted nested PCR. Mol. Ecol., 7:879–887.
Vestberg M., Saari K., Kukkonen S., Hurme T., 2005. Mycotrophy of crops in rotation and soil amendment with peat influence the abundance and effectiveness of indigenous arbuscular mycorrhiza l fungi in field soil. Mycorrhiza,15:447–458. Vestberg M., Cassells A., C., 2009. The use of AMF and PGPR inoculants Singly and Combined to promote Microplant Establishment, Growth and Health. A. Varma and A. C. Kharkwal (eds.), Berlin, Chapter 21, Symbiotic Fungi, Soil Biol., 18:337-360.
Vidović B., Petanović R., 2008. A New Eriophyid Mite (Acari: Eriophyidae) and A New Record from Olea europaea L. (Oleaceae) in Montenegro. Ann. Zoolog., 58(4):825-829.
Vierheilig H., 2004. Regulatory mechanism during the plant arbuscular mycoryzal fungus interaction . Can J . Bot., 82: 1166-1176 .
Vierheilig H., Piché Y., 2002. Signaling an arbuscular mycorrhiza : fact and hypothesis .In :Buslig B . manthey J . (eds) .flavonoid in cell function kluwer academic publisher plenum Press New York , pp. 23-39 .
Vierheilig H., Alt-Hug M., Engel-Streitwolf R., Mäder P., Wiemken A., 1998. Studies on the attractional effect of root exudates on hyphal growth of an arbuscular mycorrhizal fungus in a soil compartment-membrane system. Plant Soil, 203: 137–144.
Vilarino A., Sainz M.J. , 1997. Treatment of Glomus mosseae propagules with 50% sucrose increases spore germination and inoculum potential. Soil Biol. Biochem., 29:1571–1573.
Vimard B., St-Arnaud M., Furlan V., Fortin J.A., 1999. Colonization potential of in vitroproduced arbuscular mycorrhizal fungus spores comparedwith root-segment inoculums from open pot culture. Mycorrhiza, 8:335–338.
Vivas A., Barea J.M., Azcon R., 2005. Brevibacillus brevis Isolated from Cadmium- or Zinc-Contaminated Soils Improves in Vitro Spore Germination and Growth of Glomus mosseae under High Cd or Zn Concentrations. Microbiol Ecol., 49 : 416–424. Vivas A., Voros I., Biro B., Barea J.M., Ruiz-Lozano J.M., Azcon R.,2003. Beneficial effects of indigenous Cd-tolerant and Cd-sensitive Glomus mosseae associated with a Cd-adapted strain of Brevibacillus sp. in improving plant tolerance to Cd contamination. Appl. Soil Ecol., 24: 177–786.
Vosatka M., Albrechtova J., 2009. Benefits of Arbuscular Mycorrhizal Fungi to Sustainable Crop Production. Chapter 10. M.S. Khan et al. (eds.), Microbial Strategies for Crop Improvement, 205- 225
Weber R. W.S., 2009. Recent Developments in the Molecular Taxonomy of Fungi Physiology and Genetics, 1st EditionThe Mycota XVT. Anke and D. Weber (Eds.) Berlin, pp.1-15. Walker C., 1983. Taxonomic concepts in the Endogonaceae: Spore wall characteristics in species descriptions. Mycotaxon 18: 443–455

166
Walker C., 1985. Endogone lactiflua forming ectomycorrhizas with Pinus contorta. Trans. Br. Mycol. Soc., 84: 353-355.
Walker C., Sanders F. E. 1986. Taxonomic concepts in the Endogonaceae: III. The separation of Scutellospora gen. nov. from Gigaspora Gerd. & Trappe. Mycotaxon, 27 :169-182.
Walker C., Trappe J. M., 1993. Names and epithets in the Glomales and Endogonales. Mycol. Res. ,97: 339–344. Walker C., Schüßler A., 2004. Nomenclatural clarifications and new taxa in the Glomeromycota. Mycol. Res., 108: 979–982.
Walker C., Vestberg M., Schubler A., 2007a.Nomenclatural classification of in Glomeromycota. Mycol. Res., 111: 253-255.
Walker C., Giovannetti M., Avio L., Citernesi A.S., Nicolson T.H., 1995. A new fungal species forming arbuscular mycorrhizas: Glomus viscosum. Mycol. Res., 99:1500–1506.
Walker C., Vestberg M., Demircik F., Stockinger H., Sauito M., Sawaki H., Nishmura I., Schubler A., 2007b. Molecular phylogeny and new taxa in the Archaeosporales 5Glomeromycota) Ambispor. fennica gen. sp. nov., Ambisporaceae fam. nov., and emendation of Archaeospora and Archaeosporaceae. Mycol. Res. ,11: 137-13. Wang F. Y, Lin X., Yin R., 2005. Heavy metal uptake by arbuscular mycorrhizas of Elsholtzia splendens and the potential for phytoremediation of contaminated soil. Plant Soil, 269: 225–232.
Wang, F.Y., Liu R.J., Lin X.G., Zhou J.M., 2004. Arbuscular mycorrhizal status of wild plants in saline-alkaline soils of the Yellow River Delta. Mycorrhiza, 14: 133-137.
Wang G.M., Stribley D.P., Tinker P.B., Walker C., 1993. Effects of pH on arbuscular mycorrhiza I. Field observations on the long-term liming experiments at Rothamsted and Woburn. New Phytol., 124:465–472.
Wang H., Charles A.S., Cornell J. D., Myrna H.P., 2002 Spatial dependence and the relationship of soil organic carbon and soil moisture in the Luquillo Experimental Forest, Puerto Rico. Landscape Ecology, 17: 671–684. Wang S.G., Lin X.G., Yin R., Hou Y.L., 2003. Effects of Di-n-Butyl Phthalate on Mycorrhizal and Non-Mycorrhizal Cowpea. Plants Biol. Plant., 47( 4): 637-639,
Wang Y. Y., Vestberg M., Walker C., Hurme T., Zhang X., Lindström K., 2008. Diversity and infectivity of arbuscular mycorrhizal fungi in agricultural soils of the Sichuan Province of mainland China. Mycorrhiza, 18:59–68.
Weidmann, S., Sanchez L., Descombin J., Chatagnier O., Gianinazzi S., Gianinazzi-Pearson V.,2004. Fungal Elicitation of signal transduction-related plant genes precedes mycorrhizal establishment and requires the dmi3 gene in Medicago truncatula. Mol. Plant-Microbe Int., 17: 1385-1393.
Whipps J.M., 2004. Prospects and limitations for mycorrhizas in biocontrol of root pathogens. Can.J. Bot., 82: 1198-1227.

167
Wilson J.M., Trinick M.J., 1982. Factors affecting the estimation of numbers of infective propagules of vesicular arbuscular mycorrhizal fungi. Aust. J. Soil Res., 21:73–81.
Wright S.F., Upadhyaya A., 1998. A survey of soils for aggregate stability and glomalin, a glycoprotein produced by hyphae of arbuscular mycorrhizal fungi.Plant Soil,198: 97–107.
Wu T., Hao W., Lin X., Shi Y., 2002. Screening of arbuscular mycorrhizal fungi for the revegetation of eroded red soils in subtropical China. Plant Soil. , 239: 225–235. Wubet T., Kottke I., Teketay D., Oberwinkler F., 2003 . mycorrhizal status of indigenous trees in dry Afromontane forests of Ethiopia. For. Ecol. Manage., 179: 387-399.
Xavier L.J.C., Boyetchko S.M., 2002. Arbuscular mycorrhizal fungi as biostimulants and bioprotectants of crops. Appl. Mycol. Biotechnol. ,2 : 311-340. Xavier, L. J. C., Germida, J. J., 2002. Response of lentil under controlled conditions to coinoculation with arbuscular mycorrhizal fungi and rhizobia varying in efficacy. Soil Biol. Biochem.,34, 181-188. Xia Y. S., Chen B. D., Christie P., 2007. Arsenic uptake by arbuscular mycorrhizal maize (Zea mays L.) grown in an arsenic-contaminated soil with added phosphorus. J. Environ Sci., 19:1245–1251. Yakoub-Bougdal S., Chérifi D., Bonaly J., 2007. Production de vitroplants d‟Olea europea var Chemlal. Cahiers Agricultures, 16(2): 125-127.
Yamato M., Iwasaki M., 2002. Morphological types of arbuscular mycorrhizal fungi in roots of understory plants in Japanese deciduous broadleaved forests. Mycorrhiza, 12:291–296.
Yamato M., 2004. Morphological types of arbuscular mycorrhizal fungi in roots of weeds on vacant land. Mycorrhiza ,14:127–131.
Yamato M., 2005. Morphological types of arbuscular mycorrhizas in pioneer woody plants growing in an oil palm in Sumatra, Indonesia. Mycoscience, 46:66–68.
Yang Y.F., Yuan H.X., Liu Y.L., Xu X.P., Li B.J., 2002. Research on root microorganism community of „„RCH‟‟ transgenic rice. Chin. J. Agric. Econ., 10:29–31. Zhao Z.W., Wang G.H., Yang L. 2003. Biodiversity of arbuscular mycorrhizal fungi in a tropical rainforest of Xishuangbanna, southwest China. Fungal Diversity, 13:233–242
Zhao Z. W., Xia Y.M., Qin X.Z., Li X.W., Cheng L.Z., Sha T., Wang G.H., 2001. Arbuscular
mycorrhizal status of plants and the spore density of arbuscular mycorrhizal fungi in the tropical rain
forest of Xishuangbanna, Southwest China. Mycorrhiza, 11(3): 159–162
Zhang, Y., Guo L.D., 2007. Arbuscular mycorrhizal structure and fungi associated with mosses. Mycorrhiza, 17(4): 319-325.
Zohary D., Spiegel R.P., 1975. Beginnings of fruit growing in the old world. Science, 187: 319-327.

168
Zubek S., Turnau K., Blaszkowski J., 2008. Arbuscular mycorrhiza of endangered plants from the Tatra MTS. Acta Soc. Tis Bot. Polonia, 77 (2): 148-156 .
Zycha H. 1935. Mucorineae. Kryptogamenfl. Mark Brandenburg 6a, Leipzig, 264 p.
Biblio net [1]. http://fr.wikipedia.org/wiki/Sig_(Alg%C3%A9rie) [2]. www.Google earth.fr
[3].http://maps.google.fr//
[4].http://fr.wikipedia.org/wiki/Béchar.
RÉSUMÉ
L‟étude de l‟association mycorhizienne arbusculaire a ét é effectuée sur la variété Sigoise d‟olivier (Olea europea L.) cultivée dans
des vergers d‟olivier de la région de Sig et de Béchar. Plusieurs aspects écologiques et biologiques ont été mis en évidence.
Le sol des vergers de Sig est sablo-limoneux à texture fine, son taux d‟humidité relative est de 18.3 à 21.8%, son pH est alcalin
(8.03 et 8.43) ; il est pauvre en azote (0.09 à 0.14%) , en phosphore (0.008% à 0.02%) et relativement faible en matières organiques (1.69 à
2.00%).
Les examens microscopiques des racines naturelles d‟olivier ont révélé la présence la de mycorhizes arbusculaires de type « Arum »
à des taux très élevés (82% à 100 selon l‟âge des oliviers) ; ces résultats confirment le caractère mycotrophique de cet arbre. L‟estimation
du nombre le plus probable de propagules indique que le sol rhizosphérique des oliveraies est riche en champignons mycorhiz iens
arbusculaires (5.500 et 7000 propagules par Kg de sol). La caractérisation morphologique des spores de CMA isolées du sol rhizosphérique
révèle la présence de plusieurs espèces appartenant à trois genres de Glomeromycota : Glomus , Acaulospora et Scutellospora avec une
prédominance du genre Glomus (Gloméracées).
Deux isolats de CMA (I1 et I2) et deux complexes mycorhiziens (CMS et CMB) indigènes aux sols étudiés ont été inoculés à des
boutures herbacées de la variété Sigoise d‟olivier, en conditions contrôlées et élevés en serre, pendant 3 mois. L‟inoculation des plants
favorise significativement le développement des plants mycorhizés; ces résultats confirment que l‟endomycorhization contrôlée de l‟ol ivier
améliore la croissance des plants d‟où l‟intérêt de son utilisation en pépinière.
Les essais de culture axénique des CMA à partir de mycorhizes ou de spores en association avec des racines de plantules de tomate
in vitro nous a permis de suivre le développement du mycélium extraracinaire et les structures issues de ce dernier (spores, sporocarpes, VLS
et BAS) et d‟étudier les différents stades de développement d‟une spore isolée.
Mots clés : Variété Sigoise d‟olivier, Sig, et Béchar, CMA indigènes, mycorhizes arbusculaire , boutures herbacées d‟olivier ,
endomycorhization contrôlée, culture axénique.

169
ABSTRACT
The study of the arbuscular mycorrhizal association was done on the Sigoise cultivar (Olea europaea L.) of olive tree, originary of
two stations Sig( in two olive plantations and Béchar.. Several ecological and biological traits were evidenced.
Pedological analysis in these two areas of Sig reveal a sandy limon-like soil with a fine granulation, a relative humidity ranging from
18.3 to 21.8%, an alkaline pH (8.03 to 8.43), weak in nitrogen (0.09 to 0.14%) and phosphorus (0.008% to 0.02%) concentrations, and poor
in organic matter (1.69 to 2.00%).
Microscopical observations of olive root fragments revealed that this variety forms “Arum” type of arbuscular mycorrhizae with very
high infection rates (82 to 100% according the tree ages) this result confirms the mycotrophic characteristic of this tree. The estimatio n of the
most probable number indicates that the olive rhizospheric soil is rich with AMF (5500 and 7000 propagules/kg of soil).
Morphological characterization of spores of AMF isolated from olive rhizospher, shows the presence of three genus that belong to
the phylum of Glomeromycota: Glomus sp., Acaulospora sp. and Scutellospora sp. with the dominance of Glomus species.
Two AMF isolates (I1 and I2) and two indigenous mixture of AMF (CMS and CMB) were inoculated to young olive plantlets from
Sigoise cultivar, in control conditions. After three months in green house, the mycorrhizal plantlets have better development than non
mycorrhizal ones (roots and leaves), this result confirms that the mycorrhization improves the growth of plantlets that is interesting for its use
in nurseries.
The axenic culture of AMF using spores or mycorrhized roots in association with tomatoes roots has lets us to follow the
development of extraradical mycelium and some structures formed by it ( spores, sporocarps, VLS and BAS), and in other parts to study
different stages of development of an isolate spore.
Keywords : Olea europea, Sigoise cultivar of olive tree, Sig, Béchar, AMF, “Arum” type, mycorrhization, axenic culture.
europea
183218803843009
0140008002169200
« Arum »82100

170
GlomeromycotaspGlomusAcaulospora spScutellospora sp
55007000
I1, I2,CMS, CMB
Olea europea« Arum »

171
Annexe 1: Composition des colorants et réactifs utilisés.
-FAA : Mélange éthanol-acide acétique-formol (Phillips et Hayman, 1970) :
Formol à 37%
Acide acétique glacial
Alcool éthylique 70°
5mL
5mL
90mL
- Bleu de trypan au lactophénol (Phillips et Hayman ,1970)
Acide lactique
Glycérol
Bleu trypan
Phénol
Eau distillée
100mL
100mL
0,3g
100mL
100mL
- Fuschine acide au lactophénol
Acide lactique
Glycérol
Fuschine acide
Phénol
Eau distillée
100mL
100mL
0,3g
100mL
100mL
-Lactophénol (Langeron, 1952) :
Acide lactique
Glycérol
Phénol pur
Eau distillée
Les liquides doivent être pesés et non mesurés.
1g
2g
1g
1g
-Polyvinyl-lactoglycérol (PVLG) (Locquin et Langeron, 1978)

172
Alcool polyvinylique aq. à 15%
Acide lactique
Glycérol
Alcool polyvinylique aq. à 15%: dissoudre au bain marie à 80°C
56g
22g
22g
-Réactif de Melzer (Locquin et Langeron, 1978)
Hydrate de chlorale 100g
Iodine 1.5g
Potassium iodide 5g
Eau distillée 100 mL

173
Annexe 2 : Carte de couleur INVAM des spores ((BBllaasszzkkoo wwsskk ii,, 22000033 ))..

174
Annexe 3: Mesures des paramètres de croissance des plants l‟olivier.
Plants d’olivier inoculés par I1
Inc H (cm) LR (cm) Nbr F Nbr T Pf (g) Ps (g)
I 1 20 28,5 20 3 4,4 2,13
I 1 15 2 9 2 4,55 2,28
I 1 17 8 9 1 2,7 1,19
I 1 16 15 13 2 4,8 2,6
I 1 15,5 9,5 4 1 4,6 2,79
I 1 11,5 11 13 2 2,14 0,98
I 1 14 10 31 3 7 4,12
I 1 14 7 13 2 3,33 1,73
I 1 14 10,5 19 3 3,24 1,62
I 1 15 2 5 1 4,2 2,11
Plants d’olivier inoculés par I2
Inc2 H (cm) LR (cm) Nbr F Nbr T Pf (g) Ps (g)
Inc2 18 11 42 4 18 11,3
Ins2 19 13,5 14 1 5,08 2,53
Inc2 13 8,5 16 2 3,02 1,51
Inc2 23,5 14 31 2 7,3 3,5
Inc2 19 15,5 19 2 4,8 2,39
Inc2 28,5 11,5 13 1 4,5 1,97
Inc2 14,5 13,5 14 1 6 4,5
Inc2 13 13 6 2 3 1,52
Inc2 16,5 17 15 1 3,8 2,13
Inc2 17 15 15 1 4 2,5
Plants d’olivier inoculés par CMS
Inc H (cm) LR (cm) Nbr F Nbr T Pf (g) Ps (g)
CMS 14 9 11 1 2,78 1,46
CMS 25 17 28 3 7,6 4,2
CMS 13 6,5 10 1 3,4 1,78
CMS 13 6 7 1 7,05 4,66
CMS 11 9 29 3 6 3,065
CMS 16 8 12 2 6,05 3,53
CMS 12 16,5 9 1 3,38 1,87
CMS 14 13 16 2 3,25 1,6
CMS 15 14 19 2 6 4,5
CMS 13 12 17 2 5 3,5

175
Plants d’olivier inoculés par CMB
Inc H (cm) LR (cm) Nbr F Nbr T Pf (g) Ps (g)
CMB 17 22 39 3 10,6 7
CMB 14 6 7 1 4,16 3,12
CMB 20 30 20 2 10 5,93
CMB 11 25 13 2 3 1,73
CMB 18,5 22 17 1 6,7 2,67
CMB 17 20 27 2 6 3,24
CMB 14 12 10 3 2,5 1,55
CMB 20 11 17 2 5 2,67
CMB 21 9 32 4 10,5 6,12
CMB 15 13 16 2 8 4,84
Plants d’olivier non inoculés (témoins)
Tém H (cm) LR (cm) Nbr F Nbr T Pf (g) Ps (g)
Tém 10,5 5 2 1 2,78 1,9
Tém 13 16,5 8 1 2,85 1,51
Tém 11 18,5 8 1 3,3 2,35
Tém 13,5 10 21 1 5,62 3,52
Tém 12 10 15 2 5 3
Tém 11 17 10 1 3,8 1,9
Tém 13 10 8 1 3,6 2
Tém 14 12 10 1 4 2,5
Tém 13,5 11 13 1 3,2 2,3
Tém 12 9 6 1 3 1,5
Tém et T : témoin. ; H auteur de la partie aérienne ; LR : longueur des racines ;
Nbr : nombre de feuilles ; Nbr T : nombre de tiges ; Pf : poids frais ; Ps : poids sec.
Inc : inoculé.

176
Annexe 4 : Résultats des analyses de variance (ANOVA) et du test
de Newman-Keuls
Tableau 1 : Analyse de variance à un facteur de développement (inoculation) pour variable :
Hauteur de la partie aérienne des plants (cm).
Analyse de la Variance (new.sta) :
Effets significatifs marqués à p < 0,05000
Effet
Erreur Err Erreur
SC dl MC SC dl MC F p
Intergroupe 197,141111 4 39,4282222 497,538889 44 11,307702 3,48684659 0,00967388
SC : somme des carrés; MC : moyenne des carrés; p : probabilité.
Tableau 2: Test de Newman-Keuls ; Variable: Hauteur (cm) (new.sta) : Différences
significatives marquées à p < 0,05000.
Effet principal : Inoculation
{1} {2} {3} {4} {5}
I1 {1}
I2 {2} 0,120112
CMS {3} 0,688844 0,087963
CMB {4} 0,303279 0,335202 0,327125
Témoin {5} 0,146224 0,002677 0,137637 0,024669

177
Tableau 3: Analyse de variance à un facteur de développement (inoculation) pour
variable : Longueur de la partie racinaire des plants (cm).
Analyse de la Variance (new.sta) Effets significatifs marqués à p <
0,05000
Effet
Erreur Erreur Erreur
SC dl MC SC dl MC F p
Intergroupe 275,199444 4 55,0398889 1416,88056 44 32,2018308 1,70921614 0,15247693
Tableau 4 : Test de Newman-Keuls ; Variable: Longueur des racines (cm) (new.sta)
Différences significatives marquées à p < 0,05000.
Effet principal : Inoculation
{1} {2} {3} {4} {5}
I1 {1}
I2 {2} 0,657537
CMS {3} 0,766550 0,670088
CMB {4} 0,078261 0,142178 0,101799
Témoin {5} 0,811331 0,593372 0,751487 0,116245
Tableau 5 : Analyse de variance à un facteur de développement (inoculation) pour
variable : Nombre de feuilles des plants .
Analyse de la Variance (new.sta)
Effets significatifs marqués à p < 0,05000
Effet
Erreur Erreur Erreur
SC dl MC SC dl MC F p
Intergroupe 625,82 4 125,164 3208,5 44 72,9204545 1,71644569 0,15079995

178
Tableau 6: Test de Newman-Keuls ; Variable: Nombre de feuilles : Différences
significatives marquées à p < 0,05000
Effet principal : Inoculation
{1} {2} {3} {4} {5}
I1 {1}
I2 {2} 0,406500
CMS {3} 0,564557 0,479966
CMB {4} 0,369308 0,733278 0,546347
Témoin {5} 0,360744 0,134341 0,298695 0,095619
Tableau 7: Analyse de variance à un facteur de développement (inoculation) pour
variable : Nombre de tiges des plants.
Analyse de la Variance
(new.sta) :
Effets significatifs marqués à p
<0 ,05000
Effet
Erreur Err Erreur
SC dl MC SC dl MC F p
Intergroupe 7,46444444 4 1,49288889 27,6555556 44 0,62853535 2,37518682 0,05422268
Tableau 8 : Test Newman-Keuls ; Variable: Nombre de tiges: Différences
significatives marquées à p <0 ,05000
Effet principal : Inoculation
{1} {2} {3} {4} {5}
I1 {1}
I2 {2} 0,675878
CMS {3} 0,575045 0,779010
CMB {4} 0,575045 0,498503 0,501046
Témoin {5} 0,066959 0,097155 0,129665 0,025868

179
Tableau 9: Analyse de variance à un facteur de développement (inoculation) pour
variable : poids frais de la partie aérienne des plants (g).
Analyse de la Variance
(new.sta)
Effets significatifs
marqués à p <0,05000
Effet
Erreur Erreur Erreur
SC dl MC SC dl MC F p
Intergroupe 61,250052 4 12,2500104 311,19702 44 7,07265955 1,7320232 0,14724601
Tableau 10 : Test de Newman-Keuls; Variable: Poids frais (g) (new.sta) :
Différences significatives marquées à p < 0,05000
Effet principal : Inoculation
{1} {2} {3} {4} {5}
I1 {1}
I2 {2} 0,267176
CMS {3} 0,421813 0,449352
CMB {4} 0,148766 0,557593 0,373296
Témoin {5} 0,747921 0,243531 0,498275 0,111396
Tableau 11: Analyse de variance à un facteur de développement (inoculation) pour
variable : Poids sec de la partie aérienne des plants (g).
Analyse de la variance (new.sta) :
Effet significati f marqué
à p< 0.05000
Effet
SC
dl MC Erreur
dl
Erreur
MC
F p
Intergroupe 61,250052 4 5,489402 45 3,018306 1,818703 ,141833

180
Tableau 12 : Test Newman-Keuls; Variable: Poids sec (g) (new.sta)
Différences significatives marquées à p <0,05000
Effet principal : Inoculation
{1} {2} {3} {4} {5}
I1 {1}
I2 {2} 0.398405
CMS {3} 0,513766 0,637708
CMB {4} 0,187888 0,521623 0,506755
Témoin {5} 0,905359 0,318057 0,328032 0,165758

181
Annexe 5 : Composition des milieux de culture utilisés
Milieu de Strullu et Romand modifié (SRM)
Macroéléments(mM)
Mg SO4, 7H2O KNO3
Ca (NO3)2, 4H2O KH2PO4
KCl Na Fe EDTA
3 0.75
1.4 0.03
0.8 0.02
Oligoélément (µM)
MnSO4, 4H2O CuSO4, 5H2O
ZnSO4, 7 H2O H3BO3 (NH4)6Mo7O24
Na2MoO4, 2H2O
11 0.9
1 30
0.0 28
0.09
Vitamines (µM)
Panthoténate de Ca Biotine Acide nicotinique
Pyrodoxine Thiamine
Cyanocobalamine Saccharose (mM) Agar (g)
Eau distillée
pH avant autoclavage
2 4
8.1
4.6
2.6
0.3
29.3 8
1000mL
5.5
Eau gélosée à 1%
Agar
Eau distillée pH avant autoclavage
10
1000mL 6.8
g

182
Milieu de Crush et Hay(1981)
NH4NO3
Ca(NO3)2, 4H2O. KNO3 MgSO4, 7H2O
Na2SO4, 10H2O CuSO4, 5H2O
MnCl2, 4H2O H3BO3 KCl
KH2PO4 K2HPO4, 3H2O
FeSO4,7H2O Na2EDTA Charbon actif
Agar Eau distillée
pH avant autoclavage
0.080
0.159 0.064
0.039
0.086 0.00001
0.0004 0.00085 0.0236
0.0001 0.000045
0.017 0.022
5
15 1000mL
5.6
g
g g g
g g
g g g
g g
g g g
g