Embed Size (px)
Citation preview

1
UNIVERSITE DE LIMOGES
Ecole doctorale « Science-Technologie-Santé » ED 258
Faculté des Sciences et Techniques
UMR CNRS 6101
Laboratoire "Physiologie moléculaire de la
réponse immune et des lymphoproliférations"
TTTHHHEEESSSEEE
N° …………. Pour soutenir le grade de
DDDOOOCCCTTTEEEUUURRR DDDEEE LLL’’’UUUNNNIIIVVVEEERRRSSSIIITTTEEE DDDEEE LLLIIIMMMOOOGGGEEESSS
Discipline : Biologie, Sciences, Santé
Spécialité : Immunologie
Présentée et soutenue publiquement par
Sophie Raynal-Duchez
Le 19 Octobre 2007
Directeurs de Thèse : Pr. Michel Cogné
Pr. Raymond Julien
Jury :
Président : Abderrahman Maftah – Professeur, Limoges (UMR INRA 1061)
Rapporteurs : Claudine Schiff – Directrice de recherche INSERM, Marseille (CIML)
Hervé Watier - Professeur, Tours (IPGA)
Examinateur : Bernardo Reina – Chargé de recherche 1 CNRS, Strasbourg (IGBMC)
Modèles transgéniques pour l’étude de la fonction
des récepteurs des cellules B et de leur glycosylation

2

3
Remerciements
Je tiens tout d’abord à remercier Claudine Schiff, Hervé Watier et Bernardo Reina qui me font
l’honneur de juger mon travail. Je remercie également Abdou Maftah d’avoir accepté de présider
ce jury de thèse et Raymond Julien pour être l’instigateur du projet « cellules B et
glycosylation ».
Je tiens à remercier Michel pour m’avoir accueillie au sein de son laboratoire dès mon stage de
maîtrise et de m’avoir acceptée en thèse, sous sa direction. Merci de m’avoir dirigée, motivée et
stressée (ça, ce n’était pas le plus facile pour moi !).
Je remercie tous mes collègues du 2ème et du 3ème pour tous les bons moments que nous avons
partagés : les soirées de Noël, les repas de midi et apéros divers (de même qu’une fameuse
omelette aux girolles après une soirée éprouvante au cyto !!!)… une sacrée équipe de joyeux
lurons qui savent travailler, s’investir mais également prendre du bon temps quand il faut. Merci
au « labo du fond », à la Denizot’s team, Eric-le roi de la recup’, Anne-Gaëlle, la Laurent’s team,
Vincent, Seb, Cathy, « la caisse », Cendrine, Chahrazed, ZO-la reine des southern, Jean- ses
clusters et son petit monde du 2ème, Bernadette… Sans oublier le personnel du CHU des
laboratoires d’Hématologie et d’Immunologie qui ont également contribué au bon déroulement
de cette thèse.
Un petit mot pour Rada, Cécile et V2 qui m’ont aidée dans les manips surtout à la fin… vous êtes
trop mignonnes !!! mais également pour mes « serial killer »… de souris : Rémi, tu restes une
énigme pour moi et Virgo : « t’es vraiment trop cool ». Je remercie Armelle pour les multiples
immunisations, Nadine pour les jolies chimères et Sylvie, « ma petite souris préférée »,
continuellement souriante, qui s’est toujours occupée avec respect de mes diverses lignées de
souris transgéniques…
Le petit bureau « Tempus »… nous sommes 4 (Claire, Laulau, Krap et moi) et je pense que je ne
retrouverai jamais « meilleur bureau » !! Vous êtes plus dans mon cœur que des collègues ou
copains de cerveau (voir freezbe VWR), ami(e)s ??? Outre les bonnes blagues, les « remontages
de moral » et les grandes conversations-manip, j’ai beaucoup appris auprès de vous. Merci Krap

4
de m’avoir donnée le goût pour la recherche, c’est avec toi que tout a commencé et je t’en suis
reconnaissante (même si tu m’avais dit que j’allais en … baver !!!).
Je n’ai pas oublié mes copines « soirées – Grey’s » (Virginie, Nathalie et Christelle), on a passé
de bons moments ensemble et j’espère en passer plein d’autres… sans oublier les soirées
« Pécharmant » avec Virgo qui seront suivies, j’espère, de superbes soirées « piῆa colada » dans
un paysage de rêve !!!!!
Merci à Chantal, Jean-Luc et Claire, les Pro de la cytométrie de m’avoir donner le goût et les
bases de cette technique, et surtout d’avoir passé pas mal d’heures à trier mes cellules (petit
rappel douloureux pour Chantal !!!!)
Je tiens à remercier toute ma famille et ma belle-famille pour m’avoir soutenue et surtout mes
parents pour leur présence, leur amour et leurs encouragements… sans oublier mes amies :
Hélène, Emilie, Mylène et Vinciane qui sont continuellement à mon écoute, et toujours en attente
de mes appels téléphoniques… je vais me rattraper… et toutes les personnes que j’oublie
certainement de citer, qui me sont chères.
Enfin le meilleur pour la fin, mon Jordi… merci de m’avoir épaulée, soutenue, encouragée,
réconfortée dans cette longue aventure et de t’être même transformé en homme à tout faire
pendant quelques semaines… Tu as su respecter mon travail tout en supportant les contraintes…
je t’aime fort. Je suis fière d’être ta femme et cette thèse, c’est un peu la tienne.

5
Abréviations
-/- : homozygote négatif -/+ : hétérozygote +/+ : homozygote positif Ac : anticorps ADCC : « Antibody Dependent Cell-mediated Cytotoxicity » ADN : acide désoxyribonucléique Ag : antigène AID : cytidine désaminase induite après activation APE1 : endonucléase apyrimidique 1 ARN : acide ribonucléique ARNm : acide ribonucléique messager B : lymphocyte B BCR : récepteur des cellules B BLNK : B-cell linker protein BSAP : B-cell specific activation protein C : région constante CAT : chloramphénicol acétyltransférase CD40-L : ligand de CD40 CDC : « Complement-Dependent Cytotoxicity » CDGs : « Congenital Disorders of Glycosylation » (désordres congénitaux de la glycosylation) CDR : complementary determining region (protéine se liant aux imunoglobulines) CG : centre germinatif CH : région constante de chaîne lourde CL : région constante de chaîne légère CSH : cellule souche hématopoïétique CSPG : Chondroitin Sulfate Proteoglycan CSR : commutation de classe D : segment de diversité D.O : Densité Optique Da : dalton DH : segment de diversité de chaîne lourde DNase I : désoxyribonucléase I dNTPs : désoxyribonucléosides triphosphates DO : absorbance EGF : « epidermal growth factor » Eiκ : séquence stimulatrice intronique des chaînes légères kappa
ELISA : enzyme linked immunosorbent assay (test avec un immunoadsorbant lié à une enzyme) ES : Embryonic stem (cell) Eµ : séquence stimulatrice intronique des chaînes lourdes Fab : domaine de liaison à l’antigène d’une immunoglobuline FACS : Fluorescence Activated Cell Sorter Fc : domaine effecteur d’une immunoglobuline FDC : Follicular dendritic cells (cellules dendritiques folliculaires) FR : framework (région « armature ») Fuc : fucose GAGs : Glycosaminoglycannes Gal : galactose GalNAc : N-acétylgalactosamine GalNac-T : N-actétylgalactosaminyltransférase Gal-T : Galactosyltransférase Glc : glucose GlcA : acide glucuronique GlcNAc : N-acétylglucosamine GlcNAc-T : N-Acétylglucosaminyltranférase GlcT : Glucosyltransférase GPI : Glycosyl Phosphatidyl Inositol HAT: histone acétyltransférase HES : Hairy/Enhancer of Split hs : site hypersensible à la DNase I HSA : heat stable antigene HSPG : Heparan Sulfate Proteoglycan ICN : Intracellular Notch IdoA : acide iduronique IFN : interféron Ig : immunoglobuline IgH : locus de chaîne lourde d’immunoglobuline IgL : locus de chaîne légère d’immunoglobuline IL : interleukine J : segment de jonction Kb : kilobase kDa : kilodalton

6
KO : « knock-out » : délétion de gène par recombinaison homologue LCR : région de contrôle du locus Lfng : Lunatic Fringe LPS : lipopolysaccharide bactérien Man : mannose MAR : région d’ancrage à la matrice MEC : Matrice extracellulaire member Mfng : Manic Fringe Néo
r : gène de résistance à la néomycine NeuAc : N-acetyl-D-neuraminic acid NeuAc : acide N-acétylneuraminique ou acide sialique NHEJ : jonction d’extrémités non homologues NMD : Nonsense-mediated mRNA decay (dégradation des ARNm non-sens) ORF : Open Reading Frame PALS : manchons périartériolaires spléniques Pax : Paired box pb : paire de bases pb, Kpb : paire de bases, Kilo paires de bases PBS : « phosphate buffered saline » PCR : réaction de polymérisation en chaîne Pgk : promoteur de la phosphoglycérate kinase
PLC : phospholipase C PolyA : site de polyadénylation pVH : promoteur des régions variables des chaînes d’immunoglobulines RAG : Recombination activating gene (gène activant la recombinaison) REG : réticulum endoplasmique rugueux RPMI : Rosewell Park Memorial Institut (milieu de culture pour cellules eucaryotes) RSS : séquence signal de recombinaison S : région switch Shh : Sonic Hedgehog SHM : hypermutation somatique SiaT : Sialyltransférase SVF : sérum de veau fœtal TCR : récepteur des cellules T TdT : déoxynucléotidyltransférase TGF: transforming growth factor TGFβ : Transforming Growth Factor β TLDA : TaqMan Low Density micro-Array TNF : tumor necrosis factor UNG : uracil glycosilase V : région variable VH : région variable de chaîne lourde VL : région variable de chaîne légère Wnt : Wingless-type MMTV integration site family Xyl : Xylose

7
Table des figures et des tableaux
Figure 1 : Structure des immunoglobulines. 19
Figure 2 : Juxtaposition des régions hypervariables CDR en « 3D » 20
Figure 3 : Les Récepteurs de la cellule B (BCR) et de la cellule préB (préBCR) sont associés aux
deux molécules de signalisation Igα et Igβ. 21
Figure 4 : Représentation schématique des loci codant les Ig chez la souris. 23
Figure 5 : Les séquences signal de recombinaison (RSS) 25
Figure 6 : Réarrangements des gènes variables du locus IgH. 26
Figure 7 : Résolution des coupures double brin induites par les enzymes RAG. 27
Figure 8 : Les nucléotides N et P. 29
Figure 9 : L’accessibilité du locus des gènes d’Ig lors des réarrangements V(D)J par remodelage
de la chromatine. 30
Figure 10 : L’exclusion allélique. 32
Figure 11 : Structure des loci IgH chez la souris et l’homme. 34
Figure 12 : Les différentes étapes de l’ontogénèse des lymphocytes B conventionnels (B2). 40
Figure 13 : PAX5 et lineage B 42
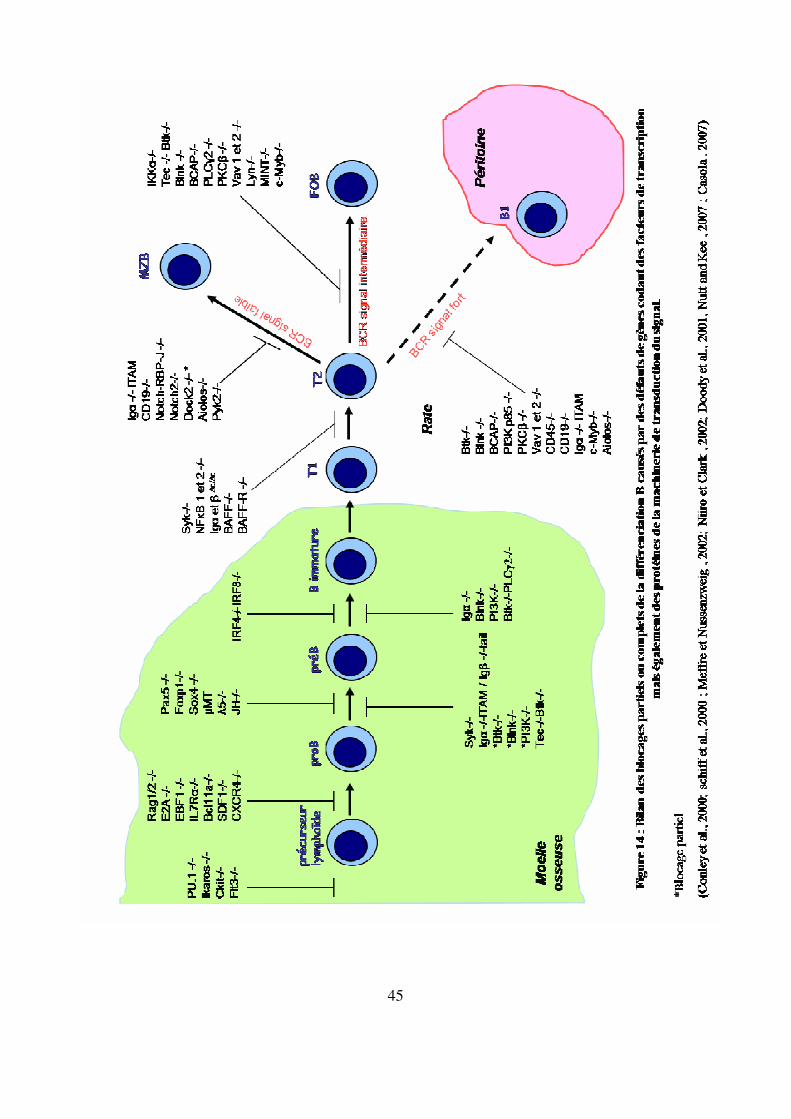
Figure 14 : Bilan des blocages partiels ou complets de la différenciation B causés par des défauts
de gènes codant des facteurs de transcription mais également des protéines de la
machinerie de transduction du signal. 45
Figure 15 : Maturation des B périphériques au niveau des centres germinatifs 47
Figure 16 : BLIMP-1 contrôle la différenciation plasmocytaire 49
Figure 17 : Schéma récapitulatif de la maturation des B périphériques 51
Figure 18 : Exemple de « switch » (commutation de classe) vers IgE. 52
Figure 19 : Structures générées par la transcription lors de la commutation de classe. 56
Figure 20 : Rôle de AID dans la commutation isotypique. 58
Figure 21 : Translocation des BCR dans les rafts après crosslinking avec un antigène. 60
Figure 22 : Signalisation médiée par le BCR. 61
Figure 23 : Représentation schématique de la transduction du signal par le BCR. 62
Figure 24 : Comparaison des régions spécifiques aux mIg 69
Figure 25 : Séquences des domaines cytoplasmiques. 70
Tableau 1 : Les principales glycosyltransférases de vertébrés greffant des sucres
monosaccharidiques sur des substrats mono- ou polysaccharidiques. 79
Figure 26 : Synthèse des N-glycoprotéines 80

8
Figure 27 : Les trois types de N-glycannes présents chez les Eucaryotes. 82
Figure 28 : Représentation schématique des principaux monosaccharides O-liés sur des résidus
Sérine et Thréonine. 83
Figure 29 : Liste non exhaustive des CSPGs et HSPGs 86
Figure 30 : La famille des glycosaminoglycannes 88
Figure 31 : Biosynthèse des HS et des CS. 90
Tableau 2 : Les différents motifs des CS 92
Figure 32 : Structure et fonction d’une IgG. 93
Figure 33 : Voie de signalisation Wnt. 98
Figure 34 : Voie de signalisation Notch. 100
Figure 35 : Les différentes structures de glycannes O-fucosylés observées sur la protéine Notch.
102
Figure 36 : Les HSPGs jouent de nombreux rôles dans la physiologie cellulaire. 105

9
Sommaire
Introduction Générale 11
Rappels Bibliographiques 17
I. Organisation et expression des gènes d’immunoglobuline 21
1.1 Le locus des chaînes lourdes (IgH) 22 1.2 Les loci des chaînes légères (IgL) 22
1.2.1 Le Locus Igκ 22 1.2.2 Le Locus Igλ 23
II. Génération du répertoire des cellules B 24
2.1 Les recombinaisons V(D)J 24 2.2 La régulation de la recombinaison V(D)J 29
2.2.1 Modèle ou théorie de l’accessibilté 29 2.2.2 La théorie de l’exclusion allélique 31 2.2.3. La régulation transcriptionnelle 33
a. Les promoteurs 33 b. Les enhancers introniques (Eμ, DQ52, Eκ) 35 c. Les enhancers 3’ (3’IgH, 3’kappa et 3’ lambda) 36 d. Les éléments atténuateurs « silencers » et les « MAR » 37
2.3 L’épissage des transcrits d’Ig 38
III. L’ontogenèse B 39
3.1 La phase indépendante des antigènes 41 3.2 La phase dépendante des antigènes 44 3.3 Le mécanisme de la commutation de classe 51
a. Les régions S 53 b. La transcription germinale et le rôle des cytokines 54 c. Le rôle de AID dans la commutation de classe 56
IV. L’activation des lymphocytes B où la signalisation via le BCR 59
4.1. La signalisation par le BcR et sa modulation 59 4.2. La signalisation en fonction des différents isotypes du BCR 68
V. « Glycannes et glycoconjugués » 77
5.1. La glycosylation 78 5.1.1. La N-glycosylation des protéines 80 5.1.2. La O-glycosylation des protéines 83 5.1.3. La O-Glycosylation de type Glycosaminoglycanne (GAG) 85
a. Les différentes sous-familles de GAGs 87 b. La biosynthèse des GAGs 89
5.2. Les « sucres » et leur implication dans le système immunitaire 92 5.2.1. La glycosylation des Ig 92

10
5.2.2. Les galectines, les voies Notch et Wnt : importance de la glycosylation. 95 a. Les galectines 95 b. Les WNTs 97 c. Notch 99
5.2.3. Les différentes fonctions associées aux GAGs 102
Résultats - Discussion 107
Première partie : les BCR modifiés 109
Article 1 111 Article 2 113 Demande de dépôt de brevet 115
Deuxième partie : les glycosaminoglycannes 131
Article 3 133
Perspectives 135
Annexes 145
Article 4 147 Article 5 149
Références Bibliographiques 151
Résumé 15180

11
Introduction Générale

12

13
Afin d’assurer la défense de l’organisme, le système immunitaire dispose d’une capacité
d’apprentissage permettant de spécifier la tolérance des antigènes du « soi » (ce qui est propre à
l’organisme) et l’élimination des antigènes du « non-soi » (ce qui est étranger). Cette dichotomie
est nuancée par la nécessité de co-signaux d’activation de l’immunité innée (signaux de
« danger ») pour la mise en place d’une réponse immunitaire efficace. Les bases de la réponse
adaptative reposent ensuite principalement sur les deux lignées lymphoïdes T et B et sur leurs
récepteurs membranaires spécifiques d’antigènes, ainsi que sur la possibilité pour la lignée B de
produire des formes solubles de leurs récepteurs pour l’antigène : les anticorps. Ces anticorps
sont capables de se lier spécifiquement à un antigène dans le but de le neutraliser et de l’éliminer.
Les cellules participant à cette immunité peuvent circuler dans l’ensemble de l’organisme et se
localiser, à des étapes spécifiques de leur maturation, au niveau d’aires spécialisées (moelle
osseuse, organes lymphoïdes secondaires…). Le système immunitaire inclut donc, non seulement
les lymphocytes T ou B mais aussi des cellules accessoires telles que les macrophages et autres
cellules présentatrices d'antigènes, et des cellules épithéliales de soutènement ou des cellules
stromales qui constituent le « microenvironnement » de la maturation lymphocytaire et de la
réponse immune.
La capacité des lymphocytes à répondre de manière spécifique à un grand nombre
d’antigènes étrangers, est le résultat d’un processus de différenciation à l’issue duquel chaque
lymphocyte exprime un récepteur unique à l’antigène.
Une longue série de phénomènes d’activation et de coopération cellulaire est nécessaire
avant d’aboutir à la sécrétion d’un anticorps spécifique par une cellule de la lignée lymphoïde B.
Cette réponse présuppose l’expression à la surface cellulaire d’une forme membranaire de
l’immunoglobuline au sein du récepteur des cellules B pour l’antigène (BCR), dont la spécificité
est définie pour chaque clone B. La population des lymphocytes B est donc formée d’un vaste
répertoire de clones produisant des BCR spécifiques des différents antigènes. Ce processus de
génération de la diversité des immunoglobulines a été proposé par Burnet en 1959 dans sa théorie
de la sélection clonale. Les cellules B subissent au cours de l’ontogenèse une sélection
directement dépendante de la spécificité de leur récepteur pour l’antigène.
Chez l’homme et chez la souris, les cellules B se différencient à partir de cellules souches
hématopoïétiques d’abord dans le foie fœtal puis dans la moelle osseuse par un processus
ordonné qui comprend un nombre limité d’étapes et nécessite une série de réarrangements intra-
géniques permettant l’assemblage de gènes d’immunoglobulines fonctionnels. Ces

14
réarrangements aboutissent à la mise en place d’un récepteur pré-B à la surface des précurseurs B
puis d’un récepteur B à la surface des cellules B immatures puis matures.
Immunocompétent, le lymphocyte B naïf va alors quitter la moelle osseuse via la
circulation sanguine et gagner les organes lymphoïdes secondaires tels la rate et les ganglions
lymphatiques. Si un lymphocyte B interagit avec un antigène dont son BCR est spécifique, la
cellule entreprendra alors une expansion clonale et pourra subir les processus de commutation de
classe et d’hypermutation somatique pour obtenir un récepteur de haute affinité dont l’expression
sera ensuite durablement conservée par des cellules B mémoires, leur conférant une capacité de
réponse rapide et adaptée en cas de nouveau contact avec le même antigène. La stimulation B
pourra également bifurquer vers la différenciation terminale plasmocytaire, soit précocement en
aboutissant à la génération de plasmocytes à courte durée de vie au niveau de foyers extra-
folliculaires (capables de sécréter rapidement de grandes quantités d’Ig de faible affinité), soit
plus tardivement au niveau des centres germinatifs, aboutissant alors à des plasmocytes qui
produisent des anticorps de haute affinité, certains de ces plasmocytes étant en outre à longue
durée de vie et capables de maintenir une production très prolongée d’anticorps spécifiques
même en l’absence de nouveau challenge antigénique.
A chaque grande étape de la maturation B, des mécanismes de sélection ont lieu et
permettent d’une part de ne conserver que les cellules exprimant un BCR fonctionnel et d’autre
part de limiter l’apparition ou l’activation de cellules exprimant un récepteur auto-réactif
(reconnaissant les antigènes du soi). Ces cellules auto-réactives sont en grande majorité éliminées
ou « anergisées » pendant la différenciation B précoce, durant laquelle elles peuvent mourir par
apoptose (délétion clonale), entrer en anergie, ou réinitier le réarrangement des gènes
d’immunoglobulines (« BCR editing »).
Au final, tant au niveau de l’activation B que du contrôle de l’autoréactivité, on conçoit
donc que la sélection d’un répertoire adapté de BCR (ou d’anticorps) via la sélection de clones B,
n’est possible que grâce au caractère clonotypique du BCR, bénéficiant donc du fait que chaque
cellule ne possède qu’un seul type de récepteur à sa surface grâce au phénomène d’exclusion
allélique qui régit l’expression des chaînes lourdes et des chaînes légères d’immunoglobulines.
Au cours de ma thèse, je me suis intéressée à la fonction même du BCR et notamment du
BCR commuté vers IgA et IgE afin de savoir si une chaîne lourde α ou une chaîne lourde ε
étaient capables de jouer un rôle similaire à celui d’une chaîne lourde µ et donc apporter des
éléments de réponse concernant la capacité d’un récepteur B (soit de type IgA soit de type IgE) à

15
conduire les cellules B au long de leur développement jusqu’au stade final plasmocyte-sécréteur
d’anticorps. Pour cela, nous avons réalisé plusieurs modèles de knock-in portant des BCR
modifiés où le gène Cα1 humain dans un cas et Cε humain dans un autre, ont été intégrés par
recombinaison homologue au niveau de la région switch µ afin d’obtenir une lignée de souris
transgéniques n’exprimant que des IgA ou des IgE. Nous avons choisi des gènes constants
humains pour palier au problème de switch résiduel endogène.
Dans une autre perspective d’étude des phénomènes pouvant modifier la réactivité des
récepteurs de surface de la cellule B, nous nous sommes intéressés à la modulation de la
glycosylation au cours de la différenciation B. La glycosylation est un des principaux événements
de maturation post-traductionnelle des protéines et des lipides. Elle consiste en une succession de
réactions enzymatiques aboutissant à l’élaboration de motifs glycanniques greffés sur des
glycoprotéines ou des glycolipides. La diversité des combinaisons glycanniques permet de
multiples interactions. Sachant que chaque modification (le greffage ou le clivage d’un sucre)
dépend de l’activité d’un gène, il nous a semblé intéressant d’étudier les variations de
l’expression de gènes impliqués dans la glycosylation afin d’être capables, par la suite, de
différencier des sous-populations B correspondant à différents états de maturation, d’activation
ou de localisation. Cette étude du glycotranscriptome au cours de la maturation B, nous a permis
de cibler puis d’étudier d’une manière plus approfondie 2 gènes d’intérêt CsGalNacT1 et Extl1
jouant un rôle dans la biosynthèse des glycosaminoglycannes. Nous avons donc réalisé des
lignées de souris transgéniques surexprimant de façon B-spécifique ces gènes afin de comprendre
plus précisément le rôle de ces enzymes in vivo au cours du développement B.

16

17
Rappels Bibliographiques

18
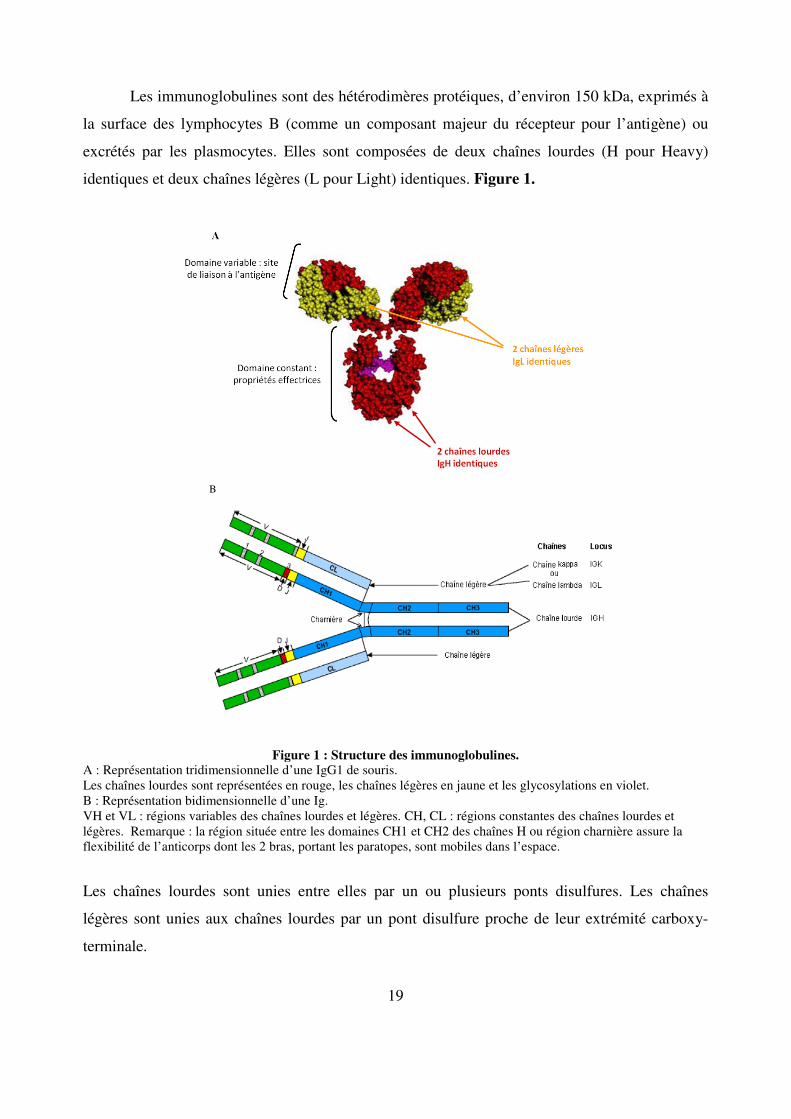
19
Les immunoglobulines sont des hétérodimères protéiques, d’environ 150 kDa, exprimés à
la surface des lymphocytes B (comme un composant majeur du récepteur pour l’antigène) ou
excrétés par les plasmocytes. Elles sont composées de deux chaînes lourdes (H pour Heavy)
identiques et deux chaînes légères (L pour Light) identiques. Figure 1.
Figure 1 : Structure des immunoglobulines.
A : Représentation tridimensionnelle d’une IgG1 de souris. Les chaînes lourdes sont représentées en rouge, les chaînes légères en jaune et les glycosylations en violet. B : Représentation bidimensionnelle d’une Ig. VH et VL : régions variables des chaînes lourdes et légères. CH, CL : régions constantes des chaînes lourdes et légères. Remarque : la région située entre les domaines CH1 et CH2 des chaînes H ou région charnière assure la flexibilité de l’anticorps dont les 2 bras, portant les paratopes, sont mobiles dans l’espace.
Les chaînes lourdes sont unies entre elles par un ou plusieurs ponts disulfures. Les chaînes
légères sont unies aux chaînes lourdes par un pont disulfure proche de leur extrémité carboxy-
terminale.
B

20
Chaque chaîne est composée d'une région constante C et d'une région variable V.
L'association des domaines variables des chaînes lourdes et légères définit le site de fixation à
l'antigène (« niche antigénique ») tandis que les domaines constants des chaînes lourdes confèrent
ses propriétés effectrices à l'immunoglobuline. Chacun des domaines variables VL et VH possède
trois zones hypervariables ou « régions déterminant la complémentarité » (CDR) séparées par des
régions plus conservées (régions « charpente » ou FR pour « framework ») et la juxtaposition de
ces zones CDR forme le paratope (site de liaison). Figure 2.
Figure 2 : Juxtaposition des régions hypervariables CDR en « 3D »
L’homme et la souris possèdent deux types de chaînes légères, appelés κ et λ, qui peuvent
s'associer à chacune des cinq classes de chaînes lourdes (µ, δ, γ, ε et α) définissant respectivement
les IgM, IgD, IgG, IgE et IgA. Les IgG sont subdivisées en 4 sous-classes : IgG1, IgG2a, IgG2b,
IgG3 chez la souris et IgG1, IgG2, IgG3 et IgG4 chez l’homme. Les IgA existent en outre chez
l’homme comme deux sous-classes IgA1 et IgA2.
Il est à noter que les Ig sécrétées n’existent pas uniquement sous des formes
monomériques mais peuvent se présenter sous formes dimériques (IgA) voir multimériques
(pentamériques pour IgM) grâce à l’association avec un petit polypeptide de 16 kDa appelé
chaîne J. Dans le cas des IgA sécrétoires retrouvées dans les compartiments muqueux, les IgA
dimériques s’associeraient secondairement avec un composant sécrétoire (polyIgR) de 70 kDa
synthétisé par les cellules épithéliales des muqueuses.
Le récepteur de la cellule B (BCR) est constitué tout d’abord d’une Ig possédant à
l’extrémité C-terminale des deux chaînes lourdes, une région transmembranaire d’une vingtaine
d’acides aminés hydrophobes permettant l’ancrage des Ig de membrane, suivie d’une courte
région cytoplasmique qui semble jouer un rôle (encore très mal connu) dans l’internalisation des

21
antigènes et dans la transduction du signal. Les capacités les plus évidentes et les mieux
explorées de transduction de signal viennent en fait non pas directement de l’immunoglobuline
membranaire mais de son association avec des modules de transduction du signal Igα/Igβ
contenant des motifs ITAMs (Immunoreceptor Tyrosine Based Activation Motif) dans leur
domaine intracytoplasmique (Hombach et al., 1988; Schamel and Reth, 2000a; Schamel and
Reth, 2000b). Figure 3.
Figure 3 : Les Récepteurs de la cellule B (BCR) et de la cellule préB (préBCR) sont associés aux deux
molécules de signalisation Igα et Igβ. Schéma de Martenson et al., 2007.
I. Organisation et expression des gènes d’immunoglobuline
La formation des chaînes lourdes et des chaînes légères des Ig résulte de l’association de
plusieurs segments de gènes qui sont organisés en loci sur des chromosomes différents.
Les régions variables des chaînes lourdes sont obtenues par l’association, dans un premier
temps, d'un segment de jonction JH avec un segment de diversité DH (induisant la délétion de
l'ADN compris entre ces segments) puis le réarrangement de cette association D-JH avec un
segment variable VH, le tout aboutissant à la formation d'un exon VDJ. Les régions variables des
chaînes légères sont quant à elles, générées seulement par jonction des segments VL et JL. Les
parties comprises entre les différents segments sont délétées lors de ces réarrangements sous
forme d'un ADN circulaire ou épisome. Les segments géniques codant les régions variables des
chaînes lourdes et des chaînes légères sont ensuite associées aux exons codant la région constante
des chaînes correspondantes par épissage, formant ainsi des ARN messagers matures prêts à être
traduits en protéines. De plus, les exons V sont précédés d'une petite séquence de 60 à 90 paires

22
de bases (pb), l’exon signal ou leader (L), codant la majeure partie (N-terminale) du peptide
signal de la protéine (Huber et al., 1993; Weichhold et al., 1993; Zachau, 1993). Le passage de la
forme membranaire à la forme sécrétée des Ig s'effectue par épissage alternatif d'un même
transcrit primaire de chaîne lourde (Early et al., 1980; Maki et al., 1981; Rogers et al., 1981;
Rogers et al., 1980).
1.1 Le locus des chaînes lourdes (IgH)
Le locus des chaînes lourdes est situé sur le chromosome 12 chez la souris et sur le
chromosome 14 chez l’homme. Il comprend chez la souris 97 segments VH (dont environ 55
pseudogènes), regroupés en 15 sous-familles, 12 à 14 segments DH (DQ52 étant le segment DH
localisé le plus en 3’), regroupés en 4 sous-familles, et 4 segments JH suivis de huit gènes codant
les régions constantes des 8 classes et sous-classes d'immunoglobulines, répartis selon l'ordre
suivant: µ, δ, γ3, γ1, γ2a, γ2b, ε, et α (Figure 4). Chez l’homme, le locus des chaînes lourdes
comprend 38 à 46 segments VH regroupés en 7 sous-familles, 23 segments DH, 6 segments JH
suivis de neuf gènes codant les régions constantes des 9 classes et sous-classes
d'immunoglobulines, répartis selon l'ordre suivant: µ, δ, γ3, γ1, α1, γ2, γ4, ε, et α2.
Chaque gène constant est composé de multiples exons codant les domaines structuraux
propres à chaque chaîne lourde ainsi que les régions charnières pour certaines immunoglobulines.
Enfin, des exons codent les régions intra-cytoplasmiques et transmembranaires pour chaque
isotype.
1.2 Les loci des chaînes légères (IgL)
1.2.1 Le Locus Igκ
Les gènes des chaînes légères sont situés sur le chromosome 6 chez la souris et sur le
chromosome 2 chez l’homme. Le locus Ig κ murin, en configuration germinale, comporte 94 à 96
segments Vκ (dont environ 70 pseudogènes) et 5 segments Jκ (dont seulement 4 fonctionnels)
tandis que le locus Ig κ humain comporte quant à lui, 31 à 35 segments Vκ fonctionnels répartis
en 5 sous-familles ainsi que 5 segments Jκ.
Les segments Vκ et Jκ codent la partie variable de la chaîne légère. Un seul segment Cκ
code la partie constante que ce soit chez l’homme ou la souris.

23
1.2.2 Le Locus Igλ
Les gènes de chaînes légères sont situés sur le chromosome 16 chez la souris et sur le
chromosome 22 chez l’homme. Ils sont répartis en 4 familles comprenant chacune une paire de
segments Jλ et Cλ (Jλ1 à Jλ4 et Cλ1 à Cλ4). Le locus de la souris ne comporte que trois segments
Vλ : le segment Vλ1 s’associe préférentiellement à Jλ1 et Jλ3 tandis que les segments Vλ2 et Vλ3
s’associent préférentiellement à Jλ2 (Jλ4 et Cλ4 étant défectifs) (Figure 4). Le répertoire Vλ très
restreint de la souris explique probablement en partie le faible pourcentage de lymphocytes B
exprimant une chaîne légère λ (environ 5% de cellules λ+ par rapport à 95% de cellules κ+ soit
un rapport de 1/20). Ce rapport est largement supérieur chez l'homme (1/3) qui possède presque
autant de gènes codant les régions variables λ que de gènes codant les régions variables κ
(Lefranc et al., 1999).
Figure 4 : Représentation schématique des loci codant les Ig chez la souris. Les segments variables V sont représentés en bleu. Les segments de diversité D sont en violet. Les segments de jonction J en vert et les gènes constants C en beige.

24
II. Génération du répertoire des cellules B
2.1 Les recombinaisons V(D)J
Les régions variables des immunoglobulines sont codées par l’association d’un segment
V, d’un segment D, spécifique des chaînes lourdes, et d’un segment J, par un mécanisme appelé
« recombinaison V(D)J ». Le grand nombre de segments V, D et J disponibles, les multiples
combinaisons entre ces éléments (diversité combinatoire) ainsi que l’imprécision de leurs
jonctions (diversité jonctionnelle) contribuent considérablement à la diversité des
immunoglobulines.
La recombinaison V(D)J est ciblée vers de courtes séquences spécifiques RSS
(Recombination Signal Sequence) non codantes et relativement conservées, qui bordent tous les
segments V, D et J. Les séquences RSS sont coupées de façon précise et forment des bouts francs
alors que l’autre moitié du site de coupure située du côté des segments codants, doit passer par
une structure intermédiaire en épingle à cheveux qui devra être ouverte, générant une extrémité
simple brin traitée secondairement. Des modifications aléatoires de ces extrémités simple brin,
avant leur jonction, vont générer des imprécisions jonctionnelles majeures (mutations, délétions
ou insertions nucléotidiques). Les mécanismes de réarrangement demeurent relativement
identiques pour les gènes variables des chaînes lourdes et légères (ou des gènes du TCR) (Cobb
et al., 2006; Tonegawa, 1983; Yancopoulos and Alt, 1986).
Chaque RSS est constituée d’un motif consensuel très conservé de sept nucléotides
(heptamère 5’-CACAGTG) et d’un autre de neuf nucléotides (nonamère 5’-ACAAAAACC). Ces
deux motifs sont séparés par une séquence peu conservée de 12 ou 23 nucléotides (Tonegawa,
1983) nommée « espaceur ». Selon la longueur de ce motif, deux classes de RSS sont définies :
les 12-RSS et les 23-RSS. Ces séquences sont présentes en 3’ des gènes V, en 5’ des gènes J et
flanquent les gènes D. La recombinaison ne peut s’effectuer qu’entre RSS possédant un
séparateur de taille différente (règle 12/23) permettant d’éviter des réarrangements non désirés.
Ainsi, chaque segment D est flanqué du même type de RSS en 5’ et en 3’, ne l’autorisant à
réarranger qu’avec les RSS compatibles d’un V en 5’ et d’un J en 3’ et évitant la formation
d’exons VH-JH ou VH-D-D-JH (Meek et al., 1989). De même, tous les segments V d’un même
locus étant flanqués de la même RSS, il ne peut survenir de réarrangement V-V. Figure 5. Dans
des locus comme ceux du TCR, où l’arrangement des RSS ne suffirait théoriquement pas à
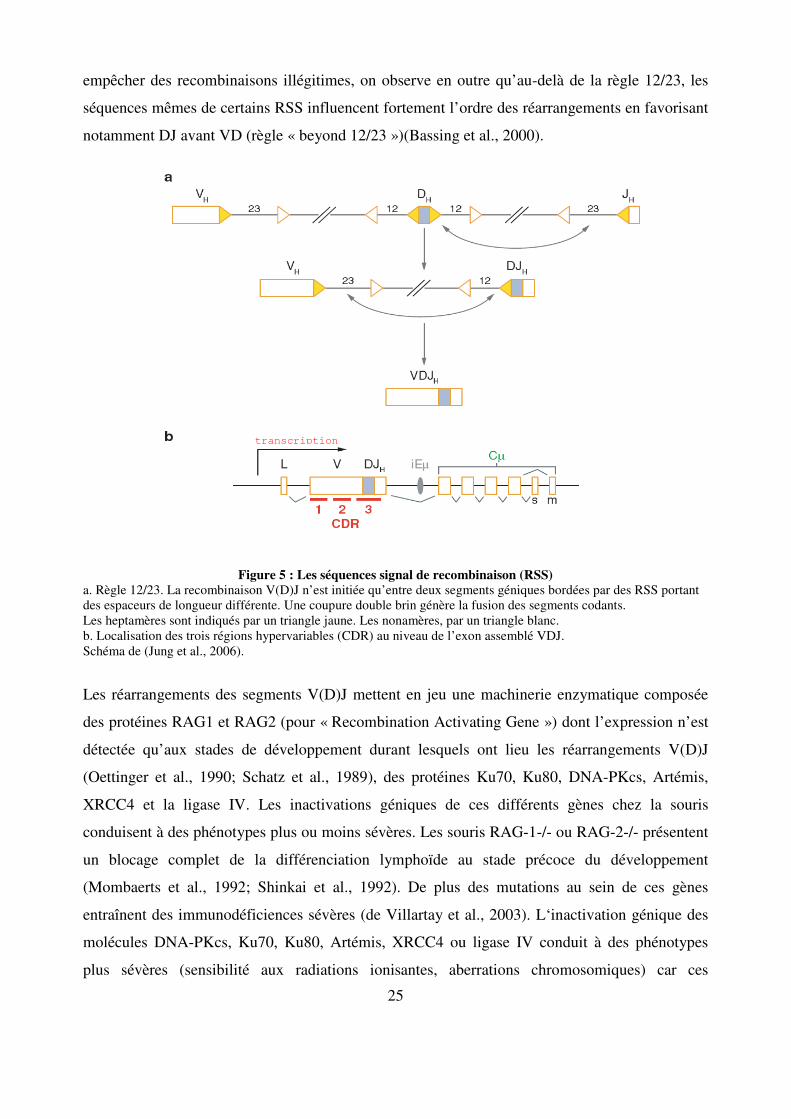
25
empêcher des recombinaisons illégitimes, on observe en outre qu’au-delà de la règle 12/23, les
séquences mêmes de certains RSS influencent fortement l’ordre des réarrangements en favorisant
notamment DJ avant VD (règle « beyond 12/23 »)(Bassing et al., 2000).
Figure 5 : Les séquences signal de recombinaison (RSS) a. Règle 12/23. La recombinaison V(D)J n’est initiée qu’entre deux segments géniques bordées par des RSS portant des espaceurs de longueur différente. Une coupure double brin génère la fusion des segments codants. Les heptamères sont indiqués par un triangle jaune. Les nonamères, par un triangle blanc. b. Localisation des trois régions hypervariables (CDR) au niveau de l’exon assemblé VDJ. Schéma de (Jung et al., 2006).
Les réarrangements des segments V(D)J mettent en jeu une machinerie enzymatique composée
des protéines RAG1 et RAG2 (pour « Recombination Activating Gene ») dont l’expression n’est
détectée qu’aux stades de développement durant lesquels ont lieu les réarrangements V(D)J
(Oettinger et al., 1990; Schatz et al., 1989), des protéines Ku70, Ku80, DNA-PKcs, Artémis,
XRCC4 et la ligase IV. Les inactivations géniques de ces différents gènes chez la souris
conduisent à des phénotypes plus ou moins sévères. Les souris RAG-1-/- ou RAG-2-/- présentent
un blocage complet de la différenciation lymphoïde au stade précoce du développement
(Mombaerts et al., 1992; Shinkai et al., 1992). De plus des mutations au sein de ces gènes
entraînent des immunodéficiences sévères (de Villartay et al., 2003). L‘inactivation génique des
molécules DNA-PKcs, Ku70, Ku80, Artémis, XRCC4 ou ligase IV conduit à des phénotypes
plus sévères (sensibilité aux radiations ionisantes, aberrations chromosomiques) car ces
transcription

26
molécules sont impliquées de façon ubiquitaire dans les mécanismes de réparation de l’ADN
(Rooney et al., 2004).
L’expression des gènes RAG-1 et RAG-2 lors du développement lymphocytaire B est un
processus strictement contrôlé de façon spatio-temporelle: une première vague d’expression
survient au stade pro-B permettant les réarrangements au niveau du locus IgH, puis au stade pré-
B pour les réarrangements des gènes de chaînes légères, sans oublier une intervention dans
l’édition du BCR des cellules immatures. Figure 6.
Figure 6 : Réarrangements des gènes variables du locus IgH. Les exons codant sont représentés par des rectangles gris, les introns par des traits fins, les signaux de recombinaisons par des triangles, les promoteurs par des flèches et les activateurs par des cercles. D’après (Dudley et al., 2005).
Pendant les premières étapes de la réaction de recombinaison, le complexe RAG-1/ RAG-
2 s’associe à une séquence RSS (12 ou 23) et induit, en présence de cations divalents métalliques,
une coupure endonucléolytique précisément à l’extrémité de l’heptamère. Grâce à une courbure
de l’ADN, la deuxième séquence RSS (23 ou 12) est rapprochée de la première séquence RSS et
forme ainsi le complexe synaptique. C’est au niveau de ce complexe que la coupure double brin
ARNm µ épissé
Réarrangement D vers J
Réarrangement V vers DJ
Expression
ADN cellule proB
ADN cellule préB
transcription
Eµ
VH1 VH2 Vn JH DH Cµ
Sµ23 23 23 23 2312 12 12 12 2323 23
ARNm µ épissé
Réarrangement D vers J
Réarrangement V vers DJ
Expression
ADN cellule proB
ADN cellule préB
transcription
Eµ
VH1 VH2 Vn JH DH Cµ
Sµ23 23 23 23 2312 12 12 12 2323 23
Recombinaison-règle 12/23
Recombinaison-règle 12/23
Epissage
Alternatif
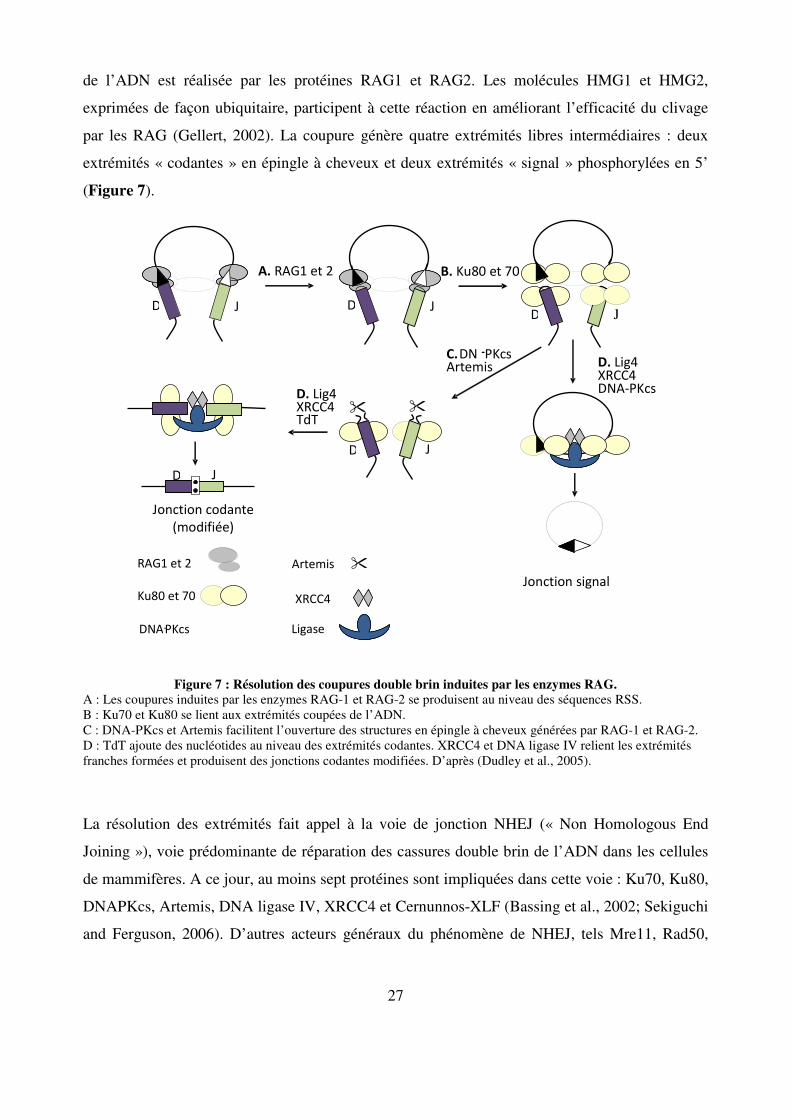
27
de l’ADN est réalisée par les protéines RAG1 et RAG2. Les molécules HMG1 et HMG2,
exprimées de façon ubiquitaire, participent à cette réaction en améliorant l’efficacité du clivage
par les RAG (Gellert, 2002). La coupure génère quatre extrémités libres intermédiaires : deux
extrémités « codantes » en épingle à cheveux et deux extrémités « signal » phosphorylées en 5’
(Figure 7).
Figure 7 : Résolution des coupures double brin induites par les enzymes RAG. A : Les coupures induites par les enzymes RAG-1 et RAG-2 se produisent au niveau des séquences RSS. B : Ku70 et Ku80 se lient aux extrémités coupées de l’ADN. C : DNA-PKcs et Artemis facilitent l’ouverture des structures en épingle à cheveux générées par RAG-1 et RAG-2. D : TdT ajoute des nucléotides au niveau des extrémités codantes. XRCC4 et DNA ligase IV relient les extrémités franches formées et produisent des jonctions codantes modifiées. D’après (Dudley et al., 2005).
La résolution des extrémités fait appel à la voie de jonction NHEJ (« Non Homologous End
Joining »), voie prédominante de réparation des cassures double brin de l’ADN dans les cellules
de mammifères. A ce jour, au moins sept protéines sont impliquées dans cette voie : Ku70, Ku80,
DNAPKcs, Artemis, DNA ligase IV, XRCC4 et Cernunnos-XLF (Bassing et al., 2002; Sekiguchi
and Ferguson, 2006). D’autres acteurs généraux du phénomène de NHEJ, tels Mre11, Rad50,
D J
A. RAG1 et 2
D J
B. Ku80 et 70
D J
D. Lig4 XRCC4 DNA-PKcs
D J
� � D. Lig4 XRCC4 TdT
C. DN
- PKcs
Artemis
D J
�
RAG1 et 2 Ku80 et 70 DNA -PKcs
Artemis XRCC4
Ligase
Jonction codante
(modifiée)
Jonction signal
(précise)

28
γH2AX ou Nbs1, ont également une implication dans le processus particulier de NHEJ que
représente la recombinaison VDJ (Chen et al., 2000; Clatworthy et al., 2005).
Les deux extrémités signal sont liguées par la DNA ligase IV tandis que les extrémités codantes
sont d’abord reconnues par Ku70 qui interagit avec Ku80. L’hétérodimérisation des protéines
Ku70 et Ku 80 produit une structure en anneau dont la partie centrale forme un canal qui encercle
les extrémités d’ADN libre (Walker et al., 2001). La protéine DNA-PKcs représente la sous-unité
catalytique qui constitue, avec les protéines Ku70 et Ku80, les composants de la protéine kinase
dépendante de l’ADN, la DNA-PK. La DNA-PKcs va recruter et former un complexe avec
Artemis facilitant l’ouverture des structures en épingle à cheveux (Ma et al., 2002). La protéine
XRCC4 s’associe avec la DNA ligase IV, complexe qui sera recruté aux sites de cassures par des
interactions avec la DNA-PK (Drouet et al., 2005). Cernunnos, protéine récemment identifiée, va
interagir avec ce complexe et relier les extrémités codantes (Ahnesorg et al., 2006; Buck et al.,
2006). Figure 7.
La jonction codante est imprécise et peut comporter de courtes délétions, additions de
duplications palindromiques (appelées « P ») ou de courtes insertions GC-riches (appelées « N »)
ajoutées par la terminal désoxynucléotidyltransférase ( TdT ) avant la ligation des extrémités. La
TdT insère des nucléotides de manière aléatoire et ne nécessite pas de matrice (Alt and
Baltimore, 1982). Cette cascade de réactions aboutit à la fusion des segments recombinants avec
une importante variabilité jonctionnelle (et donc des boucles CDR3) augmentant ainsi la diversité
du répertoire (Schlissel, 2003; Sekiguchi and Ferguson, 2006). Figure 8.
La recombinaison V(D)J permet donc in fine de générer un vaste répertoire d’Ig à partir
d’un nombre restreint de gènes. En effet, grâce à l’utilisation des différents gènes, des coupures
de l’ADN quelquefois imprécises, ainsi que la « P et la N diversité », il est possible de générer
théoriquement jusqu’à 109 Ig différentes (Tonegawa, 1983). Cependant, le tribut à payer pour
cette variabilité particulièrement importante, est la répercussion aléatoire sur le cadre de lecture
des protéines, avec seulement une jonction codante sur trois en phase correcte de lecture et
pouvant coder une protéine fonctionnelle.

29
Figure 8 : Les nucléotides N et P.
Ils sont présents en nombre aléatoire et contribuent à la diversité jonctionnelle. Ils peuvent conduire à des réarrangements non-productifs (incapacité à produire une chaîne d’Ig fonctionnelle).
2.2 La régulation de la recombinaison V(D)J
L’existence de séquences de recombinaison (RSS) où d’enzymes (RAG) spécifiques aux
lignées lymphoïdes B et T n’explique pas à elle seule le haut niveau de régulation des
réarrangements V(D)J.
2.2.1 Modèle ou théorie de l’accessibilté
Il a été établi que le ciblage spécifique de la recombinase RAG vers les séquences RSS
s’exerçait grâce à des modifications de l’accessibilité des différents loci à la recombinaison des
différents segments géniques (Yancopoulos and Alt, 1985). Les mécanismes physiques et
biochimiques de cette accessibilité mettent en jeu plusieurs phénomènes entre lesquels tous les
liens n’ont pas encore été clairement établis. On parle ainsi de remodelage chromatinien par
hyper-acétylation et méthylation des histones, de repositionnement et ouverture des nucléosomes
(Roth and Roth, 2000). Figure 9.

30
Figure 9 : L’accessibilité du locus des gènes d’Ig lors des réarrangements V(D)J par remodelage de la
chromatine. HAT : histones acétyltransférases. Modifié à partir de (Roth and Roth, 2000).
La structure de la chromatine n’est pas figée. Elle est soumise à un remodelage qui permet
le passage entre formes condensée et décondensée. Le degré de condensation est faible dans
l’euchromatine définie comme ouverte et accessible aux protéines agissant directement sur
l’ADN : polymérase, enzymes diverses et facteurs de transcription, et est élevé dans
l’hétérochromatine définie comme fermée et inaccessible (Felsenfeld, 1992; Kadonaga, 1998). Il
a été démontré que la méthylation des histones réprimait les réarrangements des gènes d’Ig :
l'analyse du profil de méthylation des gènes des chaînes lourdes et légères montre que les loci
sont méthylés dans les tissus non lymphoïdes et que l'initiation de la recombinaison V(D)J au
cours du développement des précurseurs lymphocytaires est associée à une déméthylation des
CChhrroommaattiinnee
ccoonnddeennssééee
HATs
DDéémméétthhyyllaattiioonn
ddee ll’’AADDNN
AAccééttyyllaattiioonn
ddeess hhiissttoonneess
RRééaarrrraannggeemmeenntt
ddeess
nnuuccllééoossoommeess
HATs
CChhrroommaattiinnee aacccceessssiibbllee

31
séquences Ig (Goodhardt et al., 1993; Mather and Perry, 1983; Mostoslavsky et al., 1998; Storb
and Arp, 1983).
Remodelage chromatinien et accessibilité transcriptionnelle sont liés : il a longtemps été
admis que la présence de transcrits germinaux reflétait l’ouverture des loci des Ig et donc leur
accessibilité aux enzymes de la recombinaison (Sleckman et al., 1996). Enfin des modifications
d’interactions avec la matrice nucléaire mais aussi le regroupement des loci accessibles dans des
compartiments nucléaires (foci) spécialisés contribuent à l’accessibilité des loci et donc aux
réarrangements V(D)J (Sen, 2005).
2.2.2 La théorie de l’exclusion allélique
Afin que chaque lymphocyte B généré exprime un récepteur unique pour l’antigène, il est
nécessaire de contrôler/surveiller la recombinaison V(D)J au sein du locus. Ce mécanisme,
appelé « exclusion allélique » apparaît comme essentiel dans la génération de cellules B
fonctionnelles. Il implique que pour chaque composant des chaînes d’Ig (HC et LC) ne soit
exprimé qu’un seul allèle. Trois modèles ont été proposés afin d’expliquer ce phénomène :
- le modèle de « toxicité » postulant que la présence de chaînes lourdes d’Ig différentes dans une
même cellule induirait un signal d’apoptose – modèle infirmé par les travaux de Sonoda et al.
(Sonoda et al., 1997).
- le modèle « stochastique » postulant que la probabilité de deux réarrangements productifs est
très faible (Coleclough et al., 1981).
- le modèle « régulé » qui avance que lorsqu’un réarrangement est productif, permettant la
transcription d’un ARNm en phase, non aberrant, codant une chaîne lourde µ (ou plus tard une
chaîne légère κ ou λ), va induire un rétrocontrôle négatif bloquant les réarrangements sur les
autres allèles. Figure 10. Ce modèle s’expliquerait par une perte de l’accessibilité du deuxième
allèle (mécanismes épigénétiques cités ci-dessus) (Mostoslavsky et al., 2001).

32
Figure 10 : L’exclusion allélique. L’inhibition par rétroaction (feedback inhibition) : l’expression d’une chaîne µ fonctionnelle (ou L) va inhiber les réarrangements du second allèle. Schéma de (Jung et al., 2006).
Ce modèle est fortement remis en cause au regard du processus d’édition du BCR où une cellule
B immature mais exprimant un BCR autoréactif (chaîne lourde et chaîne légère déjà réarrangées
mais induisant un signal fort), va continuer d’exprimer les RAG, donc réaliser des
réarrangements secondaires des gènes des chaînes légères afin de remplacer la chaîne
autoréactive (Gay et al., 1993; Tiegs et al., 1993). Ce mécanisme d’induction de la tolérance
montre qu’aux loci des chaînes légères (particulièrement le locus κ), la théorie de l’exclusion
allélique est sérieusement mise à mal de même que la théorie de la sélection clonale de Burnet,
selon laquelle chaque cellule B présenterait un récepteur défini dont l’autoréactivité serait
sanctionnée par une délétion clonale. En fait, il semble aujourd’hui que les cellules B subissent
un apprentissage du soi au cours duquel elles vont pouvoir modifier la spécificité de leur
récepteur. Le processus d’édition et/ou révision du BCR peut être vu comme un moyen important
de limiter les pertes occasionnées par cette autoréactivité en donnant des « secondes chances » à
la cellule avant d’induire sa délétion.
En conclusion, l’exclusion allélique est un processus complexe mettant en jeu de
nombreuses régulations qui vont aboutir à la monospécificité des cellules B impliquées dans la
réponse immune. Ce phénomène est très efficace puisque malgré les nombreux réarrangements
successifs que peuvent subir les gènes d’immunoglobulines lors du développement des cellules
Réarrangement
sur les 2 allèles
Configuration germinale

33
B, le nombre de cellules exprimant plusieurs BCR distincts en périphérie est extrêmement faible.
Nos travaux suggèrent en fait qu’une part importante de ce phénomène repose sur l’avantage
sélectif que représente pour un clone B l’expression d’une seule chaîne et donc d’un BCR
clonotypique, sans que pour autant il n’existe d’interdiction mécanique absolue à une inclusion
allélique (Sirac et al., 2006).
2.2.3. La régulation transcriptionnelle
La régulation de la transcription des gènes d’Ig est similaire à celle d’autres gènes, et fait
intervenir principalement des séquences nucléotidiques cis-régulatrices et des protéines se fixant
à ces séquences (éléments trans régulateurs). Les interactions multiples entre ces différents
éléments sont complexes et permettent un contrôle rigoureux de l’expression et de la spécificité
cellulaire B des gènes d’Ig. Plusieurs types d’éléments cis-régulateurs ont été identifiés : les
promoteurs (responsables de la transcription appropriée et efficace de ces gènes), les activateurs
transcriptionnels (« enhancers »), les atténuateurs (« silencers ») et les régions d’ancrage à la
matrice (« MAR : Matrix Association Region ») (Ernst and Smale, 1995) Figure 11. Toutes ces
séquences régulatrices comportent des sites de fixation pour les facteurs de transcription et
agissent de façon concertée. La majorité de ces facteurs de transcription ne sont pas spécifiques
de la lignée lymphoïde B mais l’action synergique de plusieurs d’entre eux peut, elle, s’avérer B-
spécifique.
a. Les promoteurs
Les promoteurs des gènes de chaînes lourdes et légères d’Ig se situent en amont de chaque
région V et assurent un niveau basal de transcription. Leur activité est comprise dans une région
d’environ 250 pb en amont du site d’initiation de la transcription. L'ensemble des promoteurs des
gènes d'Ig possèdent une séquence octamérique très conservée en amont de la TATA box :
ATGCAAAT dans les promoteurs des gènes de chaînes lourdes (pVH), ou la séquence inversée
ATTTGCAT dans les promoteurs de chaînes légères (Falkner and Zachau, 1984; Parslow et al.,
1984).
Ces séquences sont entre autres reconnues par les facteurs Oct-1, exprimé de façon
ubiquitaire, et Oct-2, spécifique à la lignée B. La forme en « hélice-boucle-hélice » du domaine
de liaison à l'ADN permet une forte affinité pour l'ADN. Ces deux protéines activent

34
efficacement la transcription à partir d’un promoteur d’immunoglobuline mais uniquement dans
la lignée B (LeBowitz et al., 1988; Pfisterer et al., 1994; Pierani et al., 1990).
Un co-activateur appelé OCA-B interagit physiquement avec les facteurs Oct-1 et Oct-2. Il
facilite l’activation transcriptionnelle d’un promoteur d’Ig par Oct-1 ou Oct-2 (Luo et al., 2006;
Pierani et al., 1990). Cependant, des souris déficientes en Oct-2 et OCA-B ont un développement
B précoce normal (Schubart et al., 2001).
En plus de l’octamère, chez la souris, les promoteurs pVH contiennent une séquence
heptamérique, nécessaire pour assurer en association avec l’octamère, une activation spécifique et
optimale du promoteur (Eaton and Calame, 1987). L’octamère et l’heptamère lient in vitro les
mêmes facteurs de transcription Oct et semblent avoir une interaction coopérative (Kemler et al.,
1989; Poellinger et al., 1989). D’autres éléments ont été localisés en amont de l’heptamère : une
région riche en pyrimidines (Eaton and Calame, 1987) de fonction inconnue, un motif reconnu
par le facteur de transcription Ig/EBP-1 et un motif µE3. Chez l’homme, l’heptamère n’est
présent que dans le promoteur des régions VH appartenant à la famille VH1(Kemler et al., 1989).
Figure 11 : Structure des loci IgH chez la souris et l’homme. Les gènes constants sont représentés par des rectangles gris, les activateurs par des ellipses rouges, les régions répétées en tandem (TR) et répétées inversées (IR) sont positionnées en 3’ des gènes α. A : Locus murin. Les ellipses jaunes représentent les régions S et les rectangles verts, les régions d’ancrage à la matrice (MAR). Les promoteurs germinaux sont signalés par des flèches coudées. B : Locus humain. Les régions en 3’ des gènes α1 et α2 contiennent des activateurs et des régions riches en G+C (ellipses bleues).
Cγ3 Cγ1 Cα1 Cγ4

35
Des promoteurs ont également été décrits en amont de chaque gène des régions constantes
des chaînes lourdes d'Ig, à l'exception de Cδ. Ce sont les sites d'initiation de la transcription
germinale. Les activateurs situés en amont des promoteurs germinaux jouent également un rôle
important dans la commutation isotypique.
b. Les enhancers introniques (Eµ, DQ52, Eκ)
L’activateur intronique Eµ fut le premier découvert au sein du locus IgH. Il est localisé
dans l’intron entre le dernier segment JH et le gène Cµ (Banerji et al., 1983). Il est entouré par
deux régions d’attachement à la matrice (MAR : « Matrix Associated Region »). Cet élément
semble actif tout au long du développement B. Sa localisation en amont de Sµ lui permet d’être
préservé de tous les évènements de recombinaisons survenant dans le locus. Eµ régule
positivement les réarrangements V(D)J et la transcription initiée au niveau de multiples
promoteurs du locus IgH (Chen et al., 1993; Engler et al., 1991; Serwe and Sablitzky, 1993). On
retrouve au sein de cette séquence, une séquence octamérique identique à celles des promoteurs
VH ainsi qu’un noyau dur « core Eµ », responsable de l’essentiel de l’activité de l’enhancer,
contenant les sites µE5, µE2, µA, µE3 et µB. Eµ fixe de nombreux facteurs parmi lesquels Oct-1
et Oct-2 (Ernst and Smale, 1995) mais aussi les protéines activatrices de la famille « hélice-
boucle-hélice » dont les produits du gène E2A (Murre et al., 1989). Ces derniers sont essentiels
dans la lymphopoïèse B, les animaux E2A -/- présentant un blocage précoce de l’ontogenèse B
(avant même les premiers réarrangements D-JH) (Bain et al., 1994). On peut remarquer que des
sites consensuels pour la fixation des produits du gène E2A sont présents sur de nombreux
éléments cis-régulateurs du locus IgH (Eµ, DQ52 et la LCR en 3’) de même que dans d’autres
loci B-spécifiques.
DQ52 présente plusieurs caractéristiques qui font de lui un segment de gène particulier. Il
est préférentiellement utilisé lors des premiers réarrangements DJ au cours de l'ontogénie (Bangs
et al., 1991; Tsukada et al., 1990) et est le seul segment D à être transcrit bien avant la survenue
des premiers réarrangements V(D)J (Alessandrini and Desiderio, 1991).
Il a été longtemps considéré que les transcrits germinaux (µ0) étaient des indicateurs les plus
précoces de l'engagement vers la lignée B et que leur transcription pouvait être un préalable
nécessaire à l'accessibilité de la région D-J à la recombinase V(D)J (Thompson et al., 1995)
L’activateur situé en 5’ de DQ52 constitue un promoteur-activateur synergique de Eµ (Kottmann
et al., 1994) impliqué dans l’activation des réarrangements précoces aux stades initiaux de

36
l’ontogenèse B. Sa délétion, chez la souris, n’inhibe pas le réarrangement DJH et ne bloque pas la
transcription µ0. En revanche, une diminution des réarrangements impliquant JH3 et JH4 est
observée (Nitschke et al., 2001). La délétion conjointe de cet élément avec Eµ a révélé que DQ52
n’était pas requis pour le réarrangement DJH et pour la transcription µ0 (Afshar et al., 2006).
Ceci suggère l’existence d’autres éléments de contrôle dans le locus IgH qui régulent le
réarrangement DJH et la transcription.
Des régions stimulatrices introniques ou « amont » ont également été caractérisées au
niveau du locus κ comme par exemple : les éléments en 5’ des Jκ et l’enhancer intronique κ
(EIκ).
c. Les enhancers 3’ (3’IgH, 3’kappa et 3’ lambda)
Différents groupes ont identifié, chez la souris, quatre activateurs transcriptionnels en 3’
du locus IgH (Figure 11), nommés hs1,2 (Lieberson et al., 1991), hs3a (Matthias and Baltimore,
1993), hs3b (Giannini et al., 1993) et hs4 (Michaelson et al., 1995) – « hs » pour site
hypersensible à la DNase I. Cette région régulatrice présente une structure palindromique
contenant de longues séquences répétées inversées qui flanquent hs1,2, centre de cet immense
palindrome. Les activateurs hs3a et hs3b présentent 97% d’homologies entre eux mais sont en
orientation opposée dans le locus (Chauveau et al., 1998). La région 3’ du locus IgH humain est
assez similaire dans sa structure, sauf en ce qui concerne hs3b qui n’a pas d’homologue chez
l’homme. De plus, il existe, suite à une duplication ancestrale du locus IgH, deux gènes α (α1 et
α2) et donc deux régions cis-activatrices en 3’.
L’activité de la région 3’ du locus IgH a surtout été appréciée par des expériences de
transfections dans des lignées cellulaires établies correspondant à des stades B définis, et plus
récemment par des modèles transgéniques. Ainsi, il a été montré que hs1,2 était actif aux stades B
tardifs (Lieberson et al., 1991). Les éléments hs3a et hs3b sont décrits comme des activateurs
plus faibles en expression transitoire ; leur activité n’est détectée qu’aux stades B mature et
plasmocyte (Madisen and Groudine, 1994; Matthias and Baltimore, 1993) Hs4 est également
considéré comme assez faible mais son activité se retrouve tout au long de l’ontogénie B, depuis
le stade pro-B jusqu’au stade plasmocyte (Madisen and Groudine, 1994; Michaelson et al., 1995).
Considérés séparément, ces quatre activateurs sont faibles. Cependant, leur combinaison conduit
à une forte activité de transcription. Des études de transfections transitoires combinant Eµ et les
différents activateurs 3’ murins ont montré des effets synergiques dépendant du stade de

37
maturation : aux stades précoces, Eµ et hs4 semblent prépondérants ; hs3a, hs1,2 et hs3b sont
actifs aux stades tardifs, mais leurs effets continuent d’être accentués par Eµ et hs4 jusqu’au stade
plasmocytaire, au cours duquel l’efficacité des séquences régulatrices semble être optimale
(Chauveau et al., 1998). Par ailleurs, il a été suggéré que la combinaison de ces quatre éléments
leur conférait les propriétés d’une LCR (Locus Control Region) (Chauveau et al., 1999; Madisen
and Groudine, 1994). Une LCR est définie comme une séquence d’ADN capable, dans un
système transgénique ou de transfection stable, de conférer au gène associé une expression tissu-
spécifique avec un taux transcriptionnel élevé dépendant du nombre de copies et indépendant du
site d’intégration (Ernst and Smale, 1995). Le rôle in vivo de la LCR en 3’ du locus IgH n’est pas
encore totalement élucidé. En effet, la redondance des séquences activatrices complique
considérablement l’évaluation de leur implication individuelle dans les processus touchant le
locus IgH (réarrangements, transcription, etc...). Ainsi, le remplacement, par recombinaison
homologue, de hs1,2 par le gène de résistance à la néomycine induit une forte diminution de la
commutation de classe (Cogne et al., 1994; Manis et al., 1998). L’inactivation individuelle de
hs3a et hs1,2 par le système Cre-loxP n’a pas révélé de phénotype remarquable (Manis et al.,
1998). Ce n’est que récemment que l’action redondante de ces différents activateurs à été mise en
évidence dans une expérience de délétion où deux de ces éléments étaient éliminés du locus. Ces
travaux ont clairement démontré le rôle essentiel des deux éléments les plus distaux dans la
transcription germinale des gènes constant de chaînes lourdes et dans la commutation isotypique
(Pinaud et al., 2001).
En 3’ du locus κ existe en outre un activateur « 3’κ », qui fonctionne en synergie avec
l’activateur intronique afin de permettre l’expression et les réarrangements normaux au niveau du
locus Igκ (Inlay et al., 2002). Plus loin encore en 3’, un enhancer « downstream » (« Ed ») a été
récemment identifié chez la souris et chez l’homme, avec une activité spécifique du stade
plasmocytaire (Liu et al., 2002). Il existe également en 3’ du locus λ, un enhancer intronique
« 3’λ » qui va permettre l’expression et les réarrangements au niveau du locus lambda de même
que l’hypermutation somatique (Popov et al., 1999).
d. Les éléments atténuateurs « silencers » et les « MAR »
Ils ont notamment été caractérisés dans les régions intervenantes entre segments V et J, ainsi que
dans les régions d’ancrage à la matrice nucléaire (régions MAR). Comme les promoteurs et les
enhancers, ils possèdent des sites de liaison à des facteurs de régulation trans (Liu et al., 2002).

38
En ce qui concerne les MAR, elles semblent avoir une certaine dualité et leur fonction précise à
proximité des enhancers introniques reste controversée : il semblerait qu’elles soient inhibitrices
dans les cellules non B et participent à la spécificité cellulaire B (Kadesch et al., 1986; Pierce et
al., 1991) ; en contrepartie dans la lignée B, elles modifieraient la structure de la chromatine pour
amplifier la propagation d’un état d’accessibilité du locus au voisinage de l’activateur (Forrester
et al., 1994). Probablement du fait de redondances biologiques, la délétion ciblée des MARs
flanquant l’enhancer Eµ n’a révélé aucun phénotype (Sakai et al., 1999).
2.3 L’épissage des transcrits d’Ig
Les gènes de chaînes lourdes d’Ig incluent des exons susceptibles de subir un épissage alternatif
et codant soit des régions d’ancrage membranaire soit des peptides spécifiques des chaînes
sécrétées. La régulation de cet épissage alternatif des ARN pré-messagers est donc capitale pour
la maturation des lymphocytes B. Elle intervient d’une part lors de la différenciation des
lymphocytes B immatures (exprimant une IgM membranaire sans IgD associée) en cellules B
matures co-exprimant alors IgM et IgD membranaires. Cette régulation est aussi impliquée dans
la maturation des cellules en plasmocytes excréteurs.
Dans les cellules B immatures, il semble que le gène δ soit méthylé et que la transcription initiée
à partir du promoteur VH ne s’étende pas au-delà du gène µ, donnant lieu à un transcrit mature
qui est épissé de façon à ne coder que la forme membranaire de la chaine µ (Maki et al., 1981).
La situation devient plus complexe dans les cellules B matures, où un seul transcrit primaire
inclut l’ensemble des séquences µs, µm, δm et δs (m pour membranaire et s pour sécrété) (Wall
and Kuehl, 1983). Les mécanismes exacts de la régulation post-transcriptionnelle des ARN pré-
messagers µ/δ ainsi que des formes membranaires et sécrétées des Ig restent encore mal connus.
L’épissage alternatif des formes membranaires semble être orienté par le clivage ou le non-
clivage préalable des ARNm au niveau du site de polyadénylation « de type sécrété », mais
semble aussi contrôlé par une structure spécifique des introns, capables de promouvoir l’une ou
l’autre forme d’épissage indépendamment et avant même la polyadénylation (Bruce et al., 2003;
Danner and Leder, 1985; Peterson and Perry, 1986). Pour la régulation µ/δ, la terminaison de
transcription qui survient en 3’ du site de polyadénylation de Cµ pourrait jouer un rôle, mais il a
été montré que sa délétion, tout en favorisant la production de transcrit primaires longs allant

39
jusqu’à Cδ, ne supprime pas pour autant la production exclusive de transcrits épissés µ plutôt que
δ dans les cellules immatures (Yuan et al., 1996).
Par ailleurs, la stabilité des ARNm augmente lors de la différenciation terminale des cellules B, et
la synthèse importante de ribosomes au sein du plasmocyte ralentit probablement la dégradation
enzymatique des ARNm des Ig, s’accompagnant d’une augmentation de la demi-vie des ARNm
qui codent les formes sécrétées des chaînes lourdes (Jack and Wabl, 1988; Mason et al., 1988).
Enfin, il a été montré que l’équipement global en facteurs régulateurs d’épissage variait entre le
stade lymphocytaire et le stade plasmocytaire, avec un effet s’exerçant même sur des transcrits
primaires non-Ig (Bruce et al., 2003).
III. L’ontogenèse B
Les gènes d’immunoglobulines sont spécifiquement exprimés dans la lignée
lymphocytaire B. Spontanément non fonctionnels dans leur disposition germinale, ils devront
d’abord subir au cours du développement B, une série de réarrangements somatiques ordonnés.
Codées par ces réarrangements, des chaînes d’Ig membranaires fonctionnelles pourront s’associer
de manière non covalente à deux molécules transmembranaires : Igα (CD79a ou mb-1) et Igβ
(CD79b ou B29), et former successivement les récepteurs pré-BCR puis BCR, clés de la
maturation, de l’activation et de la survie de la lignée B. On peut séparer l’ontogenèse des
lymphocytes B en deux phases principales :
Une phase de différentiation et de maturation indépendante de l’antigène. Elle se déroule
principalement dans la moelle osseuse et aboutit à la formation de lymphocytes B immatures
exprimant une Ig de surface qui vont pouvoir alors répondre aux antigènes.
Une phase d’activation et de différentiation finale dépendante des antigènes du soi d’abord puis
du non-soi en « périphérie » au niveau des organes lymphoïdes secondaires (rate, ganglions
lymphatiques et formations lymphoïdes associées aux muqueuses MALT pour Mucosae
Associated Lymphoid Tissue). Elle aboutit à la formation de plasmocytes et de cellules B
mémoires spécifiques d’un antigène étranger.
Les différentes étapes de ce processus de différenciation vont amener les cellules à exprimer où à
ne plus exprimer un ensemble de récepteurs (marqueurs de surface) et à répondre à divers
médiateurs directement impliqués dans le processus ordonné aboutissant à l’expression à la
surface et/ou la sécrétion d’une immunoglobuline spécifique d’un antigène (Figure 12).
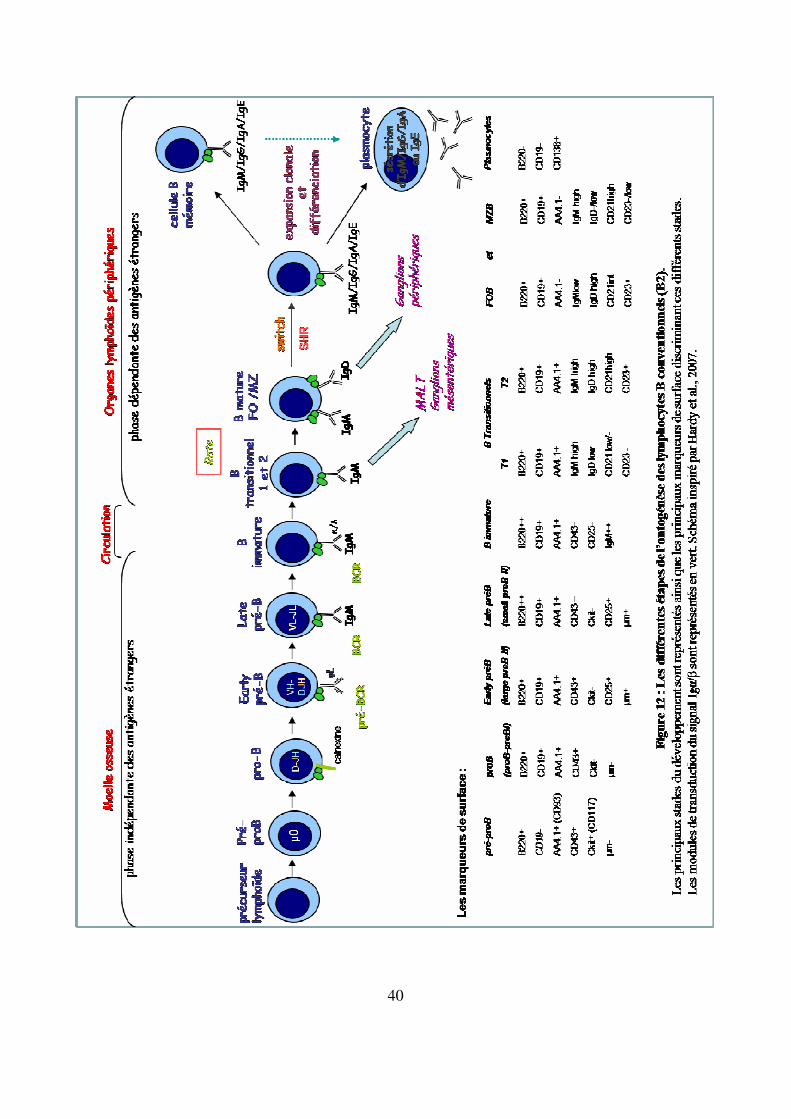
40
Figure 12 : Les différentes étapes de l’ontogénèse des lymphocytes B conventionnels (B2).

41
3.1 La phase indépendante des antigènes
La différenciation B débute dès le stade embryonnaire et se poursuit tout au long de la vie.
Les mammifères ne possèdent pas d'organe spécialisé dans la lymphopoïèse B et celle-ci se
déroule au sein des tissus hématopoïétiques. Les cellules B se développent ainsi à partir des
cellules souches lymphoïdes dans le tissu hématopoïétique du foie fœtal à partir de 8 à 9
semaines de gestation chez l’homme et vers le 14ème jour chez la souris. Le foie fœtal est ensuite
relayé par la moelle osseuse, qui conserve cette fonction chez l'adulte.
La phase indépendante des antigènes se déroule dans la moelle osseuse et correspond à la
différenciation de cellules précurseurs hématopoïétiques en cellules B matures
immunocompétentes. Cette étape implique les réarrangements des gènes d’immunoglobulines,
respectant le plus souvent un ordre précis (le modèle " ordonné " décrit par Alt et Baltimore (Alt
et al., 1984) : d’abord dans le locus IgH puis dans le locus Igκ ou Igλ. Ces réarrangements
séquentiels se produisent en l’absence d’antigène exogène et caractérisent les divers stades de la
maturation.
Le stade pré-proB est caractérisé par la forte expression du transcrit germinal µ0 (ou JH1)
(Li et al., 1996). Ces cellules n’ont pas encore engagé de réarrangements et expriment faiblement
les enzymes RAG. Elles présentent à leur surface le marqueur de la lignée B, B220 (Hardy et al.,
1991). Différents facteurs de transcription sont précocement impliqués dans la différenciation des
progéniteurs hématopoïétiques en progéniteurs communs lymphoïdes (CLP) puis en cellules B
(Fuxa and Skok, 2007; Nutt and Kee, 2007).
C’est au stade proB que débutent les réarrangements des gènes d’Ig par le rapprochement
d'une région DH vers une région JH sur les deux allèles (les enzymes RAG et la TdT sont
fortement exprimés). Vient ensuite se recombiner sur cet assemblage DJ un gène VH afin de
former l'exon codant la région variable de la chaîne lourde. Pax-5, l’interleukine 7 (IL-7) et E2A
jouent un rôle dans le maintien du lignage B. Le facteur de transcription Pax5 (BSAP) activerait
les gènes spécifiques du lignée B et inhiberait simultanément les gènes spécifiques des autres
populations médullaires (Cobaleda et al., 2007). Figure 13. Durant la lymphopoïèse B précoce, la
recombinaison V(D)J est régulée de différentes façons dont le repositionnement du locus IgH en
périphérie du noyau (accessibilité du locus). Pax5 jouerait un rôle dans la contraction du locus et
permettrait la recombinaison des VH les plus distaux sur le DJH déjà assemblé (Fuxa et al.,
2004). Il aurait également un rôle transactivateur dans la recombinaison VH-DJH en se liant au
niveau des régions codantes des VH (mais seulement sur certaines familles de VH, ce qui

42
pourrait influencer la diversification du répertoire) et en interagissant physiquement avec le
complexe RAG1/RAG2 pour faciliter la recombinaison V(D)J (Zhang et al., 2006). Si ces
réarrangements permettent l'expression d'un ARN messager fonctionnel (sans codons stop, et
avec une phase de lecture correcte), la chaîne lourde µ est alors synthétisée et exportée à la
membrane en association avec d’une part la " pseudo-chaîne légère " (composée de deux
protéines invariantes λ5 et VpreB chez la souris ou SLC pour Surrogate Light Chain chez
l’homme) et d’autre part, les molécules de transduction du signal intracellulaire Ig-α et Ig-β,
formant ainsi le pré-BCR (stade pre-B).
Figure 13 : PAX5 et lineage B L’engagement vers la lignée B est encore réversible jusqu’au stade proB. La cellule est capable de se différencier, in vitro, en présence de cytokines choisies, en différents types cellulaires et in vivo après transplantation dans des souris receveuses. La délétion de PAX5 induit une rétrodifférenciation des B en précurseurs lymphoïdes. Schéma de (Cobaleda et al., 2007).
Contrairement au stade pro-B où les réarrangements D-J ont lieu sur les deux
chromosomes, les réarrangements V-DJ ne se produisent de façon fonctionnelle que sur un seul
allèle («exclusion allélique »). Le rôle de ce pré-BCR est indispensable au bon déroulement du
développement des cellules B et a, en particulier, été souvent considéré comme essentiel dans
l'induction des réarrangements des gènes des chaînes légères. Il est en effet très habituel que les
gènes des chaînes légères soient réarrangés après ceux des chaînes lourdes et de façon
séquentielle, gènes κ d'abord puis gènes λ, lorsque les réarrangements κ n'ont pas permis
d'aboutir à un exon VJ fonctionnel (Brauninger et al., 2001; Hieter et al., 1981; Korsmeyer et al.,
1982). Cependant, de nombreuses exceptions sont venues en partie infirmer cette théorie du
modèle ordonné, qui ne semble en fait correspondre qu’à une préférence stochastique. En
particulier, il a été démontré que certains réarrangements κ pouvaient avoir lieu avant la
formation d'une chaîne lourde fonctionnelle (Chen et al., 1993; Ehlich et al., 1993;

43
Novobrantseva et al., 1999) et que des réarrangements des gènes λ pouvaient avoir lieu en même
temps, ou même avant les gènes κ. (Berg et al., 1990; Hauke et al., 1988; Liu et al., 1997;
Oberdoerffer et al., 2003; Pauza et al., 1993). Le pré-BCR fournit surtout un signal prolifératif
qui va permettre la multiplication des cellules (« small » pré-B) avant qu’elles n’engagent les
réarrangements au niveau des loci des chaînes légères (réinduction des enzymes de
recombinaison RAG1 et RAG2 mais cette fois sans expression de la TdT), poursuivant ainsi la
différenciation B (Melchers et al., 1999). La question des ligands du pré-BCR est aussi un point-
clé. Même si certains travaux montrent qu’un certain degré de différenciation B est possible en
l’absence d’association chaîne lourde - λ5/VpréB ou en substituant aux pseudo-chaînes légères
des chaînes légères normales (by-passant ainsi le stade pré-B), cette différentiation B perd en
efficacité de par la diminution de la prolifération du stade pré-BI (Corcos et al., 1995; Corcos et
al., 1991; Ohnishi and Melchers, 2003; Shaffer and Schlissel, 1997). La différenciation précoce
se fait au contraire de façon optimale lorsque l’expression d’un pré-BCR normal (incluant
« l’extra-boucle de la molécule λ5), permet aux cellules pré-B d’interagir physiquement avec les
niches des cellules stromales et d’y recevoir un signal prolifératif. Les ligands du pré-BCR
exprimés à la surface des cellules stromales sont notamment des intégrines, dont la liaison au pré-
BCR se fait grâce à une molécule de couplage soluble, la galectine-1 (Gauthier et al., 2002; Rossi
et al., 2006). Un rôle de ligand stromal non confirmé à ce jour a aussi été évoqué pour les
héparansulfate protéoglycannes (HSPGs) chez la souris (Bradl et al., 2003). Dans tous les cas,
l'expression des gènes recombinés des chaînes lourdes et légères aboutit in fine à l'expression à la
membrane de la cellule B d'une IgM composée de la chaîne lourde µ et d'une chaîne légère κ ou λ
(stade B immature). Toute cellule incapable d’assembler son pré-BCR ou d’associer et de
transporter à la membrane une chaîne lourde µ avec une chaîne légère sera éliminée, et la
sélection des lymphocytes B implique un taux très élevé d’apoptose tout au long de leur
développement. A cette sélection positive, s’ajoute un processus de sélection négative selon
lequel les cellules immatures possédant des Ig membranaires spécifiques pour les antigènes du
soi (autoréactifs – induisant un signal fort) seront également éliminées (de 40 à 60% des cellules)
(Melchers et al., 1993). Des réarrangements secondaires produisant de nouvelles chaînes légères
pourront aussi au stade B immature sauver un clone B en le dotant d’un BCR qui ne soit plus
autoréactif (Hertz and Nemazee, 1997; Melamed and Nemazee, 1997; Pelanda et al., 1997; Tiegs
et al., 1993). Une cellule immature conservant son autoréactivité malgré l’editing peut encore
entrer en anergie (diminution de la prolifération, désensibilisation et sous-expression du BCR
(Goodnow et al., 1988; Pike et al., 1982; Vilen et al., 1997) ou être éliminée par apoptose

44
(Hartley et al., 1993; Norvell et al., 1995). Il faut noter que si ce système de prévention de
l’autoréactivité vaut pour le compartiment principal des cellules B périphériques (dites B2), un
autre compartiment plus minoritaire dit B1 ou CD5+ (se localisant préférentiellement dans le
péritoine et dans la zone marginale de la rate), est au contraire sélectionné positivement lors de sa
maturation via l’expression de BCRs auto-réactifs (Cariappa and Pillai, 2002).
Les cellules qui survivent quittent la moelle osseuse vers les organes lymphoïdes périphériques
où elles pourront subir les dernières étapes de maturation.
Par recombinaison homologue (délétion partielle ou complète de certains gènes), on a pu mettre
en évidence un certain nombre de blocages à différentes étapes de la maturation B qui sont
récapitulés dans la figure 14.
3.2 La phase dépendante des antigènes
Les cellules B immatures, qui ont quitté la moelle osseuse, passent par un stade
intermédiaire, le stade B transitionnel durant lequel vont avoir lieu des modifications de la
réponse induite par le BCR. Ce stade constitue la dernière étape avant la rencontre avec des
antigènes exogènes. Les cellules B transitionnelles font donc le lien entre les cellules B
immatures de la moelle osseuse et les cellules B matures périphériques. Elles représentent une
étape importante de la maturation B puisque c’est à ce stade que la cellule va perdre sa sensibilité
à la sélection négative par le BCR, gagner les follicules de la rate et devenir sensible aux signaux
induits pas les lymphocytes T (Chung et al., 2003). Loder et al., en 1999, ont été les premiers à
montrer que le compartiment B immature de la rate pouvait se subdiviser en deux populations
bien distinctes : les B transitionnelles de type 1 (T1) et les B transitionnels de type 2 (T2)
exprimant toutes les deux fortement HSA (Loder et al., 1999). Le phénotype des cellules T1 est
très proche de celui des cellules immatures de la moelle avec pas ou peu d’IgD membranaire, une
forte expression d’IgM membranaire et une absence d’expression des récepteurs CD21, CD23 et
de la molécule anti-apoptotique Bcl2. La population des cellules T2 est quant à elle un
intermédiaire phénotypique entre les cellules T1 et les cellules matures folliculaires. Elles
expriment IgD ainsi que CD21 et CD23 (marqueurs utilisés comme témoin d’activation des
cellules B) (Rolink et al., 2004).
45
Figure 14 : Bilan des blocages partiels ou complets de la différenciation B causés par des défauts de gènes
codant des facteurs de transcription mais également des protéines de la machinerie de transduction du signal.

46
Au sein de la lignée B une marque distinctive de l’ensemble des cellules transitionnelles,
T1 comme T2, est l’expression de CD93 (une lectine transmembranaire qui lie le facteur C1q du
complément), couramment reconnue par l’anticorps monoclonal AA4.1. Allman et al., en 2001
ont subdivisé la population T2 en deux selon l’expression de l’IgM membranaire (les T2 sont
IgMhigh alors que les T3 sont IgMint)(Allman et al., 2001). Cette nouvelle population T3,
caractérisée tout d’abord comme transitionnelle, pourrait en fait correspondre à une population
anergique indépendante (compartiment An1) qui serait maintenue dans son état anergique par la
présence d’autoantigènes dans la rate (Merrell et al., 2006).
Les cellules T1 circulent dans le sang et se situent dans la partie externe des manchons
lymphoïdes péri-artériolaires (PALS) de la rate tandis que les T2 sont uniquement retrouvées
dans la rate aussi bien dans les PALS que dans les follicules où ont lieu la présentation des
antigènes exogènes et l’activation T-dépendante des cellules B matures. A la différence des T1,
les cellules T2 sont capables de proliférer suite à la stimulation par le BCR (Petro et al., 2002; Su
and Rawlings, 2002) et d’intégrer les signaux de costimulation par les lymphocytes T (Chung et
al., 2002). La population T1 est la cible de la sélection négative en périphérie : seulement un petit
pourcentage de cellules T1 se différenciera en cellules T2 (délétion clonale et anergie). BAFF (B
cell activating factor) jouerait un rôle crucial dans l’homéostasie des B périphériques. Les B
transitionnels qui reçoivent un signal de BAFF via BAFF-R, sont sauvées de la mort cellulaire et
peuvent se différencier pour devenir des B matures (Sasaki et al., 2004; Schiemann et al., 2001).
La différenciation des cellules transitionnelles semble également être dépendantes des signaux
délivrés par le BCR. L’étude de nombreux modèles de souris transgéniques où des mutations
affectent la signalisation par le BCR a permis de montrer qu’une altération du signal empêche
soit le passage du stade T1 au stade T2 soit le passage du stade T2 au stade B mature (Figure
14). Ces cellules semblent en outre pouvoir s’orienter vers plusieurs « destins » (cellules B1, B2
folliculaires ou cellules de la zone marginale). Casola et al. en 2004 ont développé la théorie de
« la force du signal » (signal strentgh) en utilisant des souris dépourvues de BCR et qui
expriment à la place la protéine de latence du virus d’Epstein Barr LMP2A. Il apparaît que des
signaux faibles, intermédiaires et forts favorisent la formation de cellules B de la zone marginale,
de cellules B folliculaires et de cellules B1 respectivement.
Les cellules T2 vont finir d’émigrer dans les zones centrales des follicules où vont leur
être présentés des antigènes exogènes dans un micro-environnement constitué notamment par des
lymphocytes T helpers (TH) et des cellules présentatrices d’antigènes, les FDC ( pour « Follicular
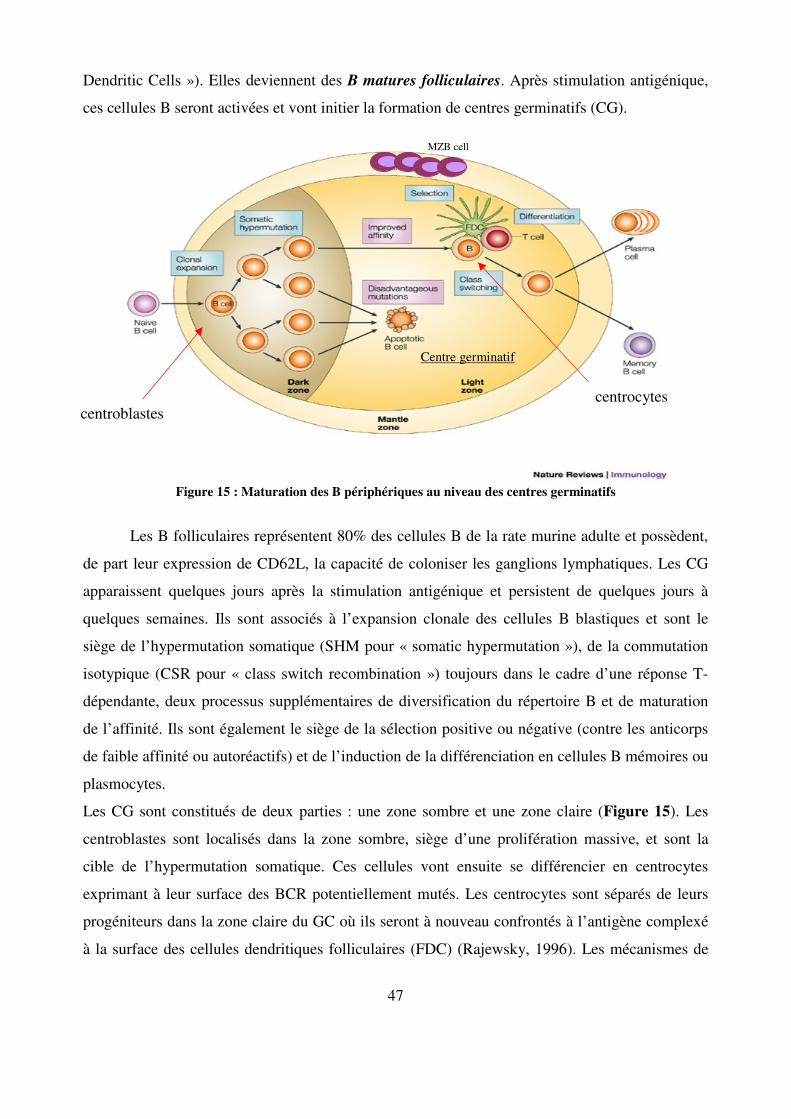
47
Dendritic Cells »). Elles deviennent des B matures folliculaires. Après stimulation antigénique,
ces cellules B seront activées et vont initier la formation de centres germinatifs (CG).
Figure 15 : Maturation des B périphériques au niveau des centres germinatifs
Les B folliculaires représentent 80% des cellules B de la rate murine adulte et possèdent,
de part leur expression de CD62L, la capacité de coloniser les ganglions lymphatiques. Les CG
apparaissent quelques jours après la stimulation antigénique et persistent de quelques jours à
quelques semaines. Ils sont associés à l’expansion clonale des cellules B blastiques et sont le
siège de l’hypermutation somatique (SHM pour « somatic hypermutation »), de la commutation
isotypique (CSR pour « class switch recombination ») toujours dans le cadre d’une réponse T-
dépendante, deux processus supplémentaires de diversification du répertoire B et de maturation
de l’affinité. Ils sont également le siège de la sélection positive ou négative (contre les anticorps
de faible affinité ou autoréactifs) et de l’induction de la différenciation en cellules B mémoires ou
plasmocytes.
Les CG sont constitués de deux parties : une zone sombre et une zone claire (Figure 15). Les
centroblastes sont localisés dans la zone sombre, siège d’une prolifération massive, et sont la
cible de l’hypermutation somatique. Ces cellules vont ensuite se différencier en centrocytes
exprimant à leur surface des BCR potentiellement mutés. Les centrocytes sont séparés de leurs
progéniteurs dans la zone claire du GC où ils seront à nouveau confrontés à l’antigène complexé
à la surface des cellules dendritiques folliculaires (FDC) (Rajewsky, 1996). Les mécanismes de
Centre germinatif
centroblastes
MZB cell
centrocytes

48
sélection des cellules exprimant des anticorps de haute affinité sont particulièrement stringents et
font intervenir des interactions spécifiques avec des cellules T (coopération B/T). De plus ces
cellules rentrent en compétition pour une interaction la plus affine possible avec les FDC (Berek
et al., 1991; Jacob et al., 1991; Kuppers et al., 1993).
Les cellules B folliculaires vont accumuler des mutations somatiques au niveau des séquences de
leur domaine variable modifiant ainsi leur affinité pour l’antigène (processus d’« hypermutation
somatique »). Ces cellules expriment faiblement la protéine anti-apoptotique bcl-2 et fortement le
récepteur Fas (CD95) (Yoshino et al., 1994) et semblent donc destinées à mourir sauf si des
signaux de survie leur sont adressés par les lymphocytes TH (H pour Helpers) reconnaissant le
même antigène (Liu and Arpin, 1997). Ainsi, si les mutations accumulées améliorent l’affinité
pour l’antigène, les cellules seront sauvées et pourront proliférer avant d’entrer dans les
compartiments B mémoires ou plasmocytes (Liu et al., 1991). Au contraire, si les mutations ne
changent pas, voire diminuent l’affinité, ou si les interactions avec les lymphocytes TH
n’ont pas
lieu (cas d’anticorps autoréactifs), ces cellules sont alors éliminées par apoptose (MacLennan,
1994).
Les cellules B mémoires, exprimant fortement le gène bcl-2, constituent un groupe
minoritaire de cellules à longue durée de vie, capable de persister à l’état quiescent sans proliférer
(de plusieurs mois à plusieurs dizaines d’années chez l’homme). Elles n’expriment en général
plus d’IgD, ont commutées et peuvent avoir des localisations préférentielles (telles que les
muqueuses pour les cellules ayant « switché » vers IgA) (Liu et al., 1995). Elles ont la faculté de
répondre très rapidement à des pathogènes externes. En effet, elles peuvent présenter rapidement
et efficacement l’antigène aux lymphocytes T lors d’une réponse secondaire et se différencier en
plasmocytes. Cette différenciation peut se faire en dehors des centres germinatifs (Liu et al.,
1991) bien que les cellules B mémoires puissent aussi recommencer de nouveaux cycles de
sélection et d’hypermutation dans les centres germinatifs, permettant ainsi d’optimiser la
mémoire immunitaire de l’individu (Berek and Milstein, 1988). Par ailleurs, il a été montré que
des cellules B mémoires pouvaient se développer en l’absence de CG, notamment celles portant
des IgM et des IgG1 dépourvues de mutations somatiques (Toyama et al., 2002).
Les plasmocytes, exprimant le récepteur de surface CD138 (syndecan-1), sont les cellules
effectrices de la réponse immunitaire humorale. Ce sont de vraies usines de production et de
sécrétion d’anticorps à destination de l’ensemble de l’organisme. La durée de vie de ces cellules

49
sécrétrices est controversée. Deux sortes de plasmocytes peuvent être générés : à durée de vie
longue ou à durée de vie courte en fonction de l’antigène et des signaux reçus suite à la
stimulation antigénique. Certains pourraient également naître de la transformation rapide d’une
cellule B mémoire suite à une stimulation antigénique (Ochsenbein et al., 2000). La mise en place
du programme de différenciation plasmocytaire est dépendante de deux facteurs de transcription :
Blimp-1 et Xbp-1 (Calame et al., 2003; Lin et al., 2003). La délétion conditionnelle du gène
Prdm1 (codant Blimp-1) dans les cellules B provoque un blocage sévère de la différenciation
plasmocytaire qui se traduit par un défaut de sécrétion d’Ig et par l’absence de plasmocytes de vie
longue et courte (Shapiro-Shelef et al., 2003). Le mode d’action de Blimp-1 consiste à réprimer
plusieurs autres facteurs de transcription indispensables à la voie B tels que Pax5 et Bcl6 (Shaffer
et al., 2002) Figure 16. Le facteur de transcription IRF4 est également requis pour la génération
de plasmocytes. Son KO conditionnel dans les B du CG montre un défaut en plasmocytes post-
CG et montre également que les B mémoires sont incapables de se différencier en plasmocytes.
De plus, les B des souris IRF4-/- perdent l’expression d’AID et ne peuvent plus « switcher »
(Klein et al., 2006). Benson et al. en 2007, ont montré qu’une expression forte d’ IRF4 dans les
cellules B des CG orienterait ces cellules vers une différenciation en plasmocytes à longue durée
de vie et non pas en B mémoires (Benson et al., 2007).
Figure 16 : BLIMP-1 contrôle la différenciation plasmocytaire Schéma de Kallies and Nutt, 2007. Blimp-1 exprimé dès le stade pré-plasmablast va réprimer les facteurs de transcription du lignage B, Bcl6 et pax5 et permettre l’expression de Xbp-1 et IRF-4.

50
D’autres cellules B périphériques interviennent dans les réponses immunes T-indépendantes.
Leurs origines restent encore controversées mais leur action est essentielle puisque ce sont ces
cellules qui vont constituer la première ligne de défense contre les micro-organismes (Hardy,
2006) : ce sont les cellules B de la zone marginale folliculaire (MZ) de la rate et les cellules B1
présentes notamment dans la cavité péritonéale. Ces cellules B périphériques présentent
l’originalité d’être positivement sélectionnées par des antigènes du soi, indiquant que des
différences dans le niveau de signalisation du BCR en réponse à ces antigènes peuvent
déterminer la localisation des cellules B et leur rôle en périphérie (Cariappa and Pillai, 2002).
Elles sont à l’origine des autoanticorps « naturels » polyréactifs de faible affinité dont les
fonctions sont multiples : élimination des débris cellulaires, transports de cytokines ou encore
formation des complexes antigènes/anticorps présentés aux cellules B folliculaires par les FDC
dans les centres germinatifs (Coutinho et al., 1995; Stall et al., 1996).
Les cellules B MZ expriment fortement IgM et CD21, faiblement IgD et CD23 et ne
montrent chez la souris aucun signe de maturation de l’affinité, ce qui les différencie des autres
populations de cellules B présentes dans la rate (transitionnelles, matures folliculaires et
mémoires) (Dammers et al., 2000). Elles ne semblent pas nécessiter la présence de lymphocytes
T pour être sélectionnées et activées puisqu’elles sont présentes chez les souris nudes ou
thymectomisées (Kumararatne and MacLennan, 1982). Elles sont aussi caractérisées par un état
d’activation élevé leur permettant de répondre rapidement à un antigène T-indépendant par
différenciation en cellules sécrétrices d’anticorps (Oliver et al., 1997; Oliver et al., 1999). La voie
Notch et notamment Notch2 jouerait un rôle important dans la formation de cette population
(Hozumi et al., 2004; Kuroda et al., 2003; Saito et al., 2003; Tanigaki et al., 2002). Il est
important de signaler que la physiologie des cellules B MZ semble radicalement différente chez
l’homme puisque cette fois les cellules B IgM+ IgD+ y portent majoritairement le marqueur B
mémoire CD27 et expriment un BCR muté. Une partie des cellules B mémoires (CD27+) du sang
périphérique humain semble d’ailleurs être un équivalent circulant de ces B MZ. A la différence
des cellules B folliculaires, les B MZ humains mutent leur BCR de façon indépendante de
l’interaction CD40-CD40L et ont un rôle dans la réponse aux antigènes polysaccharidiques
(capsules bactériennes notamment) (Weller et al., 2005).
Les cellules B1 sont, elles, situées en dehors de tout organe lymphoïde dans les cavités
péritonéales. Leur origine est controversée : il semblerait qu’elles dérivent de précurseurs issus
du foie foetal mais de récentes études tendent à montrer qu’elles pourraient dériver aussi des
précurseurs médullaires ou des cellules B périphériques « classiques » (B2) chez l’adulte (Arnold

51
et al., 2000; Chumley et al., 2000; Wortis and Berland, 2001). Elles expriment, pour la plupart, le
marqueur de surface CD5 qui les différencient des autres populations de cellules B.
Figure 17 : Schéma récapitulatif de la maturation des B périphériques Ce modèle proposé par Casola et al., en 2007, montre 2 voies de maturation partant de deux progéniteurs différents (foie fœtal et moelle osseuse) et aboutissant à la génération de cellules B1 dans un cas et de cellules B2/conventionnelles dans un autre (Casola, 2007).
3.3 Le mécanisme de la commutation de classe
La spécificité antigénique des Ig est déterminée par les régions variables des chaînes lourdes et
des chaînes légères des Ig. Les fonctions effectrices, en revanche, dépendent des régions
constantes des chaînes lourdes et varient selon les isotypes. Les IgM sont majoritairement
produites au cours de la réponse primaire, alors que les IgG, IgA ou IgE sont majoritairement
produites au cours d'une réponse secondaire ou tertiaire. Au cours du changement d'isotype ou
commutation de classe, un même gène réarrangé VDJ (éventuellement remanié par
l'hypermutation somatique) va se rapprocher d’un nouveau segment constant de classe différente
par un processus de recombinaison somatique. Les cellules B matures peuvent alors exprimer et
sécréter des IgG, IgA ou IgE spécifiques de l’antigène. Ce processus se révèle donc primordial
pour la diversité fonctionnelle des réponses anticorps.

52
Figure 18 : Exemple de « switch » (commutation de classe) vers IgE. Les exons codants sont représentés par des rectangles beiges, les introns par des traits fins, les régions S par des ellipses et les activateurs par des cercles.
L’IgG est l’isotype prédominant dans le sang et la lymphe. Les différentes sous-classes d’IgG ont
des fonctions effectrices différentes dues à leur capacité variable à lier les différents récepteurs Fc
(FcγRI, FcγRII, FcγRIII and FcγRIV), à induire l’ADCC (Antibody-Dependant cell-mediated
Cytotoxicity) et/ou à activer le complément (Nimmerjahn and Ravetch, 2005). Les IgG3 jouent
un rôle important dans la réponse antibactérienne et sont très efficaces dans la phagocytose. Les
IgG2a sont prédominantes dans les réponses antivirales. Dans les réponses T-dépendantes, les
IgG1 représentent l’isotype dominant dans la lutte contre les infections virales et parasitaires. La
voie du complément n’est pas activée mais les IgG1 stimulent la phagocytose efficacement.
Les IgA sont prédominantes dans les sécrétions (respiratoires, digestives ou génitales) et sont
fortement résistantes à la protéolyse enzymatique. Les IgA mucosales sécrétées dans la lumière
intestinale protègeraient l’organisme contre les virus et les bactéries à tropisme muqueux.
Les IgE sont impliquées dans la défense parasitaire, mais peuvent également médier les réactions
d’hypersensibilité. Les mastocytes (tissus) et basophiles (circulation) expriment des récepteurs à
haute affinité pour IgE (FcεRI) qui peuvent lier les IgE monomériques en absence d’Ag. Les Ag
spécifiques induisent l’agrégation des complexes IgE-FcεR entraînant la sécrétion d’histamine
impliquée dans les réactions d’hypersensibilité. Il existe en revanche une voie d'hypersensibilité
indépendante d'IgE. Chez des souris délétées pour Cε, on note la persistance d'une réaction
anaphylactique en l’absence d'IgE (Oettgen et al., 1994).

53
La commutation isotypique, permettant à une cellule d'exprimer une chaîne lourde
différente d'IgM est un mécanisme complexe et hautement régulé. Elle cible des points chauds de
recombinaisons appelés région « switch » ou S, situées en amont de tous les domaines constants à
l'exception de Cδ. Plusieurs éléments cis-régulateurs du locus IgH sont également impliqués dans
le contrôle ce processus. Alors que les éléments situés les plus en 5’et notamment l’activateur
transcriptionnel intronique Eµ, semblent n’avoir que peu de rôle ou un rôle redondant (Perlot et
al., 2005), les promoteurs germinaux situés en amont des régions switch jouent un rôle majeur et
les activateurs transcriptionnels qui les contrôlent semblent pour l’essentiel regroupés en aval du
locus IgH, où ils constituent la « région régulatrice 3'IgH » (éléments pour la plupart détaillés
dans le chapitre 2).
a. Les régions S
Les régions S ou régions « switch » sont des sites de recombinaison composés de motifs répétés,
riches en G, situés dans l’intron qui précède chacun des gènes constants, à l’exception de Cδ
(Figure 18). Elles sont composées de répétitions de courtes séquences en tandem (les pentamères
GAGCT, GGGGT ou GGGCT) en aval d’un court exon I précédé de son promoteur. Les
répétitions courtes peuvent elles-même s’organiser en motifs se répétant sous la forme d’unités
plus longues, comme par exemple dans les régions Sγ caractérisées par des motifs répéts de 49
pb. La longueur des régions S varie de 1 kb (Sε) à 10 kb (Sγ1) (Dunnick et al., 1993).
La recombinaison a lieu de façon imprécise entre différents sites localisés dans ou au voisinage
des régions S. Les étapes de reconnaissance et de clivage des régions S demeurent mal comprises.
Des cassures à la fois double et simple brin pourraient être impliquées dans l’initiation de la
commutation de classe (Chen et al., 2001b), et de fréquentes mutations, stigmates de
l’intervention d’AID, sont retrouvées au voisinage des points de cassures (Kenter, 2003; Manis et
al., 2002a; Manis et al., 2002b). Les cassures d’ADN situées dans les régions S sont suivies d’une
étape de réparation et de ligation impliquant le mécanisme NHEJ et les processus de réparation
ubiquitaires, tels que les mécanismes de réparation des « mistmatchs ». L’ADN intermédiaire,
sous forme circulaire non réplicative ne subsiste que transitoirement sous forme d’épisome et se
perd au cours de la division celulaire, tandis que la réparation chromosomique place la région
constante cible à proximité de la région VDJ.
La délétion quasi complète de Sµ aboutit à un blocage sévère mais incomplet de la commutation
de classe vers tous les domaines constants en aval (Khamlichi et al., 2004; Luby et al., 2001). Par
contre, la délétion de Sγ1 aboutit à un blocage quasi-complet de la commutation vers IgG1 mais

54
pas vers les gènes constants en aval (Shinkura et al., 2003). L’inversion de la région Sγ1, diminue
son efficacité en tant que cible du switch, de mêm que toute amputation de sa longueur et il est
donc clair que ces longues séquences répétées contribuent par leur orientation, leur longueur et
leur caractère répétitif à cibler les recombinaisons, sans qu’elles soient pour autant indispensables
à ces recombinaisons (Zarrin et al., 2005). Le caractère GC-riche des régions switch de
smammifères ne semble pas fondamental pour la mécanique du switch et on a pu leur substituer
avec efficacité une région Sµ AT-riche provenant de Xenopus laevis (Zarrin et al., 2004) au sein
de laquelle le processus de recombinaison s’est révélé cibler des motifs AGCT.
b. La transcription germinale et le rôle des cytokines
Avant la commutation, les cellules B activées expriment un ou plusieurs ARN appelés
transcrits germinaux codés par l’un ou l’autre gène constant. Le caractère obligatoire de cette
transcription préalable au switch a permis de considérer très tôt qu’une grande partie de la
régulation du switch portant sur l’accessibilité des gènes cibles, pouvait être ramenée pour
l’essentiel à une régulation de la transcription germinale (Lutzker et al., 1988). Ce phénomène de
transcription germinale se déroule aussi bien sur l’allèle exprimé que sur l’allèle exclu et il est
fréquent que les deux allèles recombinent vers le même gène constant (Delpy et al., 2003). Tous
les transcrits germinaux ont une structure analogue présentant un site d’initiation de la
transcription localisé en 5’ de chaque région S, appelé promoteur I. Après initiation, la
transcription se poursuit à travers un exon I non traductible puis à travers la région S et les
différents exons constants CH jusqu’aux sites de polyadénylation (Dudley et al., 2005).
L’épissage se fait normalement entre l’exon I et le site accepteur de CH1. La présence de
nombreux codons stop rend ces transcrits stériles (Chaudhuri and Alt, 2004; Goodman et al.,
1993).
Différents modèles d’études ont mis en évidence le contrôle de la production des
différents isotypes par les cytokines et différents mitogènes (Stavnezer, 2000). Pour les
principaux, l’addition d’interleukine 4 (IL-4) à des cellules activées par le LPS induit la
commutation de classe vers IgG1 et IgE tandis que l’addition d’interféron-γ (IFN-γ) induit la
commutation vers IgG2a. Le « transforming growth factor » (TGF-β) induit quant à lui la
commutation de classe vers IgA et IgG2b. En plus des cytokines, les signaux issus des contacts
entre cellules T et B sont impliqués dans la commutation de classe. L’élément le plus important
dans ces contacts est CD40-L exprimé sur les cellules T activées. Le signal médié via CD40
induit la commutation de classe vers la majorité des isotypes.

55
L’action des cytokines semble s’exercer essentiellement au niveau des promoteurs
germinaux. Des sites de fixation de certains facteurs de transcription dans les promoteurs
germinaux ont été identifiés et agissent en réponse à l’action des cytokines. Les cytokines
induiraient l’expression ou l’activité de facteurs transcriptionnels souvent multiples et capables
de former des complexes activateurs en se liant à leurs promoteurs germinaux cibles (Delphin
and Stavnezer, 1995; Warren and Berton, 1995). Par exemple, la stimulation in vitro des cellules
B par le l’IL-4 en présence de LPS induit la transcription germinale de γ1 et ε par le recrutement
de Stat6, NF-κB, PU.1, BSAP(Pax5), C/EBP et AP-1 (Geha et al., 2003).
Le fait que tous les transcrits germinaux aient la même structure suggère qu’ils exercent,
par eux-mêmes, une fonction importante. Des mutations ciblées qui abolissent ou altèrent la
structure de ces transcrits indiquent que non seulement leur existence mais encore leur épissage
correct sont nécessaires au mécanisme de commutation isotypique. La transcription en elle-même
n’est pas suffisante pour obtenir la commutation de classe, puisque le remplacement de l’exon Iε
par un promoteur fort mais non inductible par l’IL4 (Eµ associé au promoteur pVH) conduit à
une transcription germinale forte, mais diminue cependant la commutation de classe (Bottaro et
al., 1994). La corrélation entre la synthèse des transcrits germinaux et la commutation isotypique
a conduit à proposer un modèle basé sur l’accessibilité de la chromatine. Grâce à leur remodelage
chromatinien et à la transcription germinale qui lui est associée, les régions S deviendraient
accessibles à des facteurs agissant en trans (Stavnezer-Nordgren and Sirlin, 1986). Au regard de
cette hypothèse, il a été montré que les gènes constants, en cours de commutation, étaient
hypométhylés, hyperacétylés et présentaient des sites d’hypersensibilité à la DNase I dans le
promoteur germinal (Berton and Vitetta, 1990).Cependant, l’hyperacétylation des histones des
régions S et des promoteurs germinaux ne suffit pas à induire la commutation isotypique sans
transcription germinale (Nambu et al., 2003).
La transcription germinale pourrait également aboutir à la formation de structures
particulières, au niveau de l’ADN, qui seraient la cible de AID pour initier le clivage de l’ADN et
déclencher la commutation de classe (Chaudhuri and Alt, 2004). Quatre modèles de structures ont
été proposés : les quartets G (Dempsey et al., 1999), les stem-loops (Tashiro et al., 2001), la
boucle R (Yu et al., 2003) et la bulle stabilisée par RPA (protéine se liant à l’ADN simple brin et
impliquée dans la réplication et la réparation) (Chaudhuri et al., 2004). Figure 19. De ces quatre
structures, seul le modèle de la boucle R simple brin a été démontré in vivo. Ces boucles
contiennent un hybride stable ARN-ADN sur le brin riche en G et une région simple brin au
niveau de la séquence riche en C.

56
Figure 19 : Structures générées par la transcription lors de la commutation de classe. Les transcrits des régions S peuvent s’associer de façon stable au brin d’ADN matrice pour former des hybrides ARN-ADN, dans lesquels le brin non matrice peut, en théorie, adopter différentes structures (quartets G, stems loops ou bulles) ou peut rester simple brin (R loop). RPA : protéine de réplication A. Schéma de (Chaudhuri and Alt, 2004).
c. Le rôle de AID dans la commutation de classe
AID (« Activation-induced cytidine deaminase ») a été découverte par criblage soustractif
de banques d’ADNc, dans une lignée B (CH12-F3) pouvant commuter in vitro. Cette protéine de
24kDa est exclusivement exprimée dans les centres germinatifs in vivo, ou après stimulation de
splénocytes in vitro (Muramatsu et al., 1999). AID est absolument indispensable pour la SHM et
la CSR. En cas de déficit en AID, le développement B est normal (expression d’IgM et d’IgD)
mais on peut voir un blocage complet de la commutation de classe (souris déficientes pour tous
les autres isotypes malgré induction de la transcription germinale) et de l’hypermutation
somatique (Muramatsu et al., 2000; Revy et al., 2000).
L’analyse de la séquence codante d’AID a révélé une forte homologie avec APOBEC-1.
APOBEC-1 est une sous-unité catalytique d’un complexe multiprotéique d’édition de l’ARN ;
elle agit sur l’ARNm de l’apolipoprotéine Apob. AID a ainsi été considérée dans un premier
temps comme une nouvelle protéine d’édition de l’ARN agissant sur un ou plusieurs ARNm
codant des endonucléases impliquées dans l’introduction de cassures de l’ADN au cours de

57
l’hypermutation (Muramatsu et al., 1999). Ce modèle est encore aujourd’hui soutenu par l’équipe
d’Honjo. Cependant, un autre courant et des données solides suggèrent plutôt qu’AID agit
directement sur l’ADN (Bransteitter et al., 2003; Chaudhuri et al., 2003; Dickerson et al., 2003;
Petersen-Mahrt et al., 2002; Pham et al., 2003).
AID est essentiellement localisée dans le cytoplasme des cellules B, probablement pour
limiter son effet mutagène. Après activation des cellules B, elle est transloquée dans le noyau
grâce à un signal de localisation nucléaire présent dans se partie N-terminale puis est exportée du
noyau grâce à une séquence-signal d’export localisée dans sa partie C-terminale (Ito et al., 2004;
McBride et al., 2004). Le mécanisme qui régule le ratio nucléaire/cytoplasmique est inconnu
mais certaines données suggèrent une corrélation avec la phosphorylation de AID par la protéine
kinase A (PKA) dépendante de l’AMPc (Basu et al., 2005). L’analyse mutationnelle de AID a
révélé que la protéine avait une structure bipartite au regard de son activité mutagénique. Les
mutations affectant sa partie N-terminale bloquent la SHM mais pas la CSR alors que les
mutations affectant sa partie C-terminale bloquent la CSR mais pas la SHM suggérant fortement
l’existence de cofacteurs distincts pour la SHM et la CSR, mais leur identité reste pour l’instant
inconnue (Shinkura et al., 2004).
AID semble intervenir dans l’initiation de la commutation de classe en générant une
déamination des cytosines (dC) en uraciles (dU) au niveau des régions S riches en motifs AGCT
considérés comme des « hots spots » de déamination, comparables à ceux de l’hypermutation
somatique (Chaudhuri and Alt, 2004; Zarrin et al., 2004). La génération d’ADN simple brin au
cours de la transcription germinale des régions S semble être impliquée dans le ciblage de
l’action d’ AID vers les régions riches en GC (Figure 20). A la suite de la déamination des
cytosines par AID, l’action de l’uracile glycosilase (UNG) (et peut-être du complexe Msh2-
Msh6) est d’éliminer la mutation entraînant ainsi la formation d’un site abasique (gap) :
processus pré-requis dans le modèle de réparation des « mismatch » par la voie de réparation
BER (« Base Excision Repair »). Il est à noter que les souris déficientes en UNG montrent un
blocage sévère dans la commutation de classe (Rada et al., 2002; Schrader et al., 2005). APE1,
endonucléase apurinic/apyrimidique ayant un rôle majeur dans cette voie, génère une coupure au
niveau de ce site. Cependant, des données récentes impliquent plutôt le complexe MRN
(Mre11/Rad50/Nbs1) dans ce clivage (Larson et al., 2005). Les coupures sont ensuite réassociées
par un processus nécessitant l’intervention de nombreux facteurs protéiques tels que H2AX
(histone 2A family member X), 53BP1 (p53 binding protein), mutL homologue 1 (MLH1),
l’ataxia telangiesctasia mutated (ATM) et DNA-PKcs (Petersen et al., 2001; Reina-San-Martin et
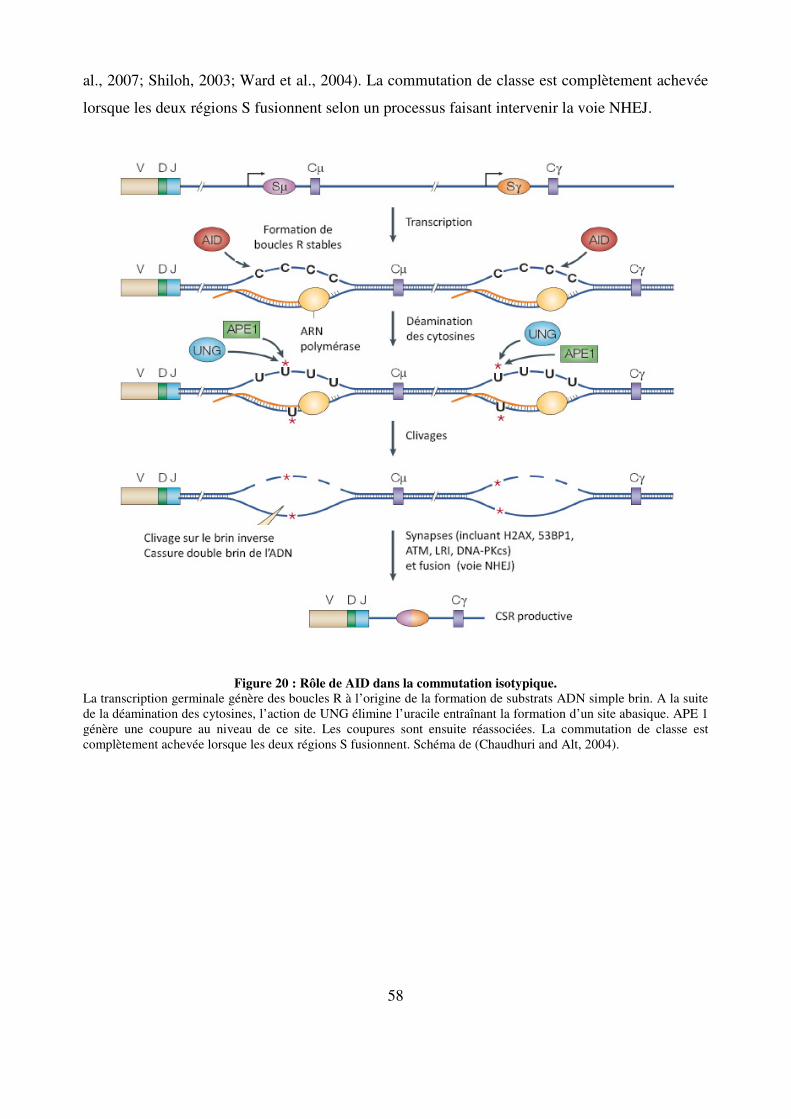
58
al., 2007; Shiloh, 2003; Ward et al., 2004). La commutation de classe est complètement achevée
lorsque les deux régions S fusionnent selon un processus faisant intervenir la voie NHEJ.
Figure 20 : Rôle de AID dans la commutation isotypique. La transcription germinale génère des boucles R à l’origine de la formation de substrats ADN simple brin. A la suite de la déamination des cytosines, l’action de UNG élimine l’uracile entraînant la formation d’un site abasique. APE 1 génère une coupure au niveau de ce site. Les coupures sont ensuite réassociées. La commutation de classe est complètement achevée lorsque les deux régions S fusionnent. Schéma de (Chaudhuri and Alt, 2004).

59
IV. L’activation des lymphocytes B où la signalisation via le BCR
4.1. La signalisation par le BcR et sa modulation
La liaison de l'antigène au BCR (B cell antigen Receptor) permet l'activation de la
cascade des signaux de transduction, l'internalisation et l’apprêtement ("processing") des
complexes BCR/antigènes. Le complexe BCR, identifié pour la première fois en 1970 (Raff et
al., 1970), est un complexe multimérique. Il comprend une immunoglobuline membranaire (mIg),
structure de reconnaissance de l'antigène, un hétérodimère associé de manière stable, non
covalente, Igα (CD79A) et Igβ (CD79B), élément de transduction du signal.
Igα et Igβ sont des glycoprotéines de 22kDa et de 26kDa codées respectivement par le
gène mb1 (Hombach et al., 1988; Sakaguchi et al., 1988) et par le gène B29 (Hombach et al.,
1990; Parkhouse, 1990), appartenant à la superfamille des immunoglobulines et composées
chacune d’un domaine extracellulaire (114 et 132 acides aminés), d’un domaine
transmembranaire (22 acides aminés) et d’un domaine cytoplasmique (61 et 48 acides aminés)
(Wang and Clark, 2003). Le domaine extracellulaire de Igα comporte deux cystéines et un
tryptophane qui sont retrouvés dans les protéines de la super-famille des Ig. Ces deux cystéines
semblent permettre la formation d'un pont di-sulfure intra-chaîne et une troisième cystéine serait
probablement engagée dans la liaison di-sulfure interchaîne entre Igα et Igβ. La protéine Igβ est
quant-à elle, dotée de 5 cystéines dont 3 seraient engagées dans des liaisons analogues à Igα, et
les deux dernières pourraient former un autre pont di-sulfure intra-chaîne (Williams and Barclay,
1988). Tout comme les Ig de surface, les séquences murines et humaines de ces deux protéines
révèlent une très grande conservation dans les domaines transmembranaires (100% pour Igα et
85% pour Igβ) et intra-cytoplasmiques (87% pour Igα et 90% pour Igβ) mais faible dans la partie
extracellulaire (56% d'identité pour Igα et 59% pour Igβ). Le pourcentage élevé d'identité de
séquence des domaines transmembranaires suggère un rôle fonctionnel très important, lié à
l'interaction avec les mIg et/ou avec d'autres protéines membranaires.
Cet hétérodimère est indispensable à la transduction du signal grâce à l'existence, au
niveau de leurs queues cytoplasmiques, de motifs ITAM (Immunoreceptor Tyrosine-based
Activation Motif) qui servent de substrat pour des protéines tyrosine kinases (PTKs). Décrit pour
la première fois en 1989 (Reth, 1989), le cœur de ce motif très conservé,
(D/E(X)7D/EXXYXXI/L(X)7YXXI/L), comprend deux tyrosines séparées par onze résidus et
suivies chacune par une leucine(L) ou isoleucine (I) en position +3.
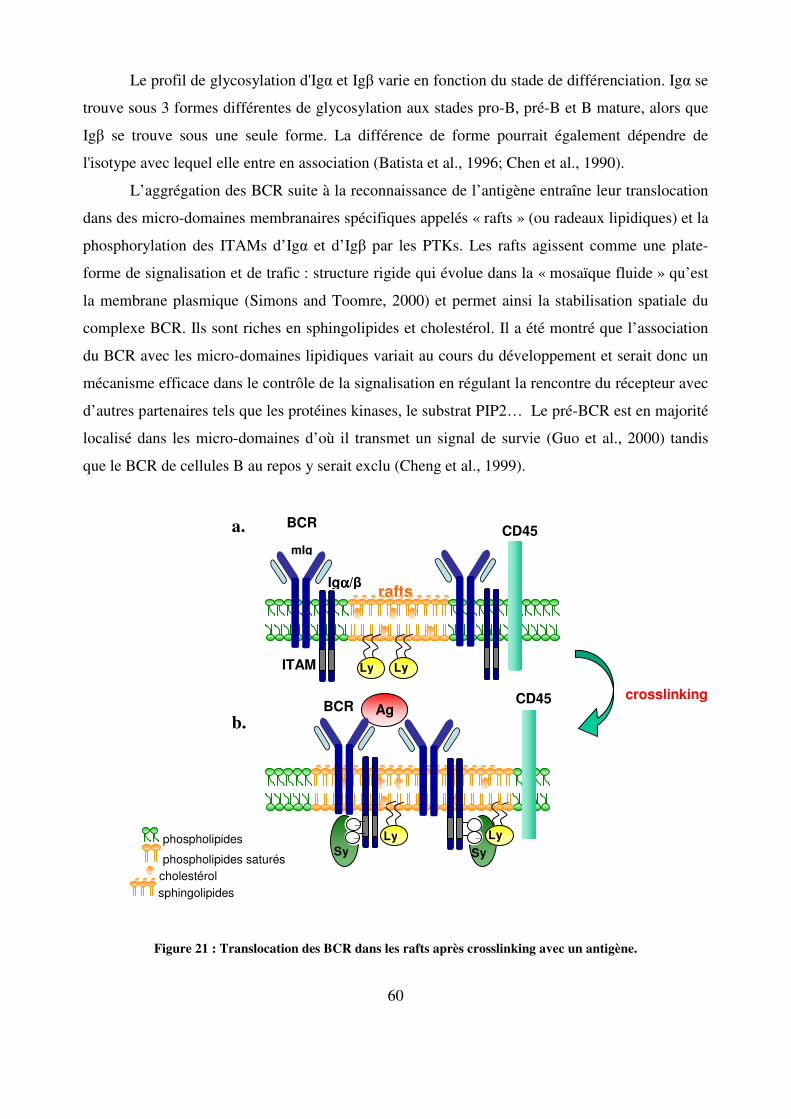
60
Le profil de glycosylation d'Igα et Igβ varie en fonction du stade de différenciation. Igα se
trouve sous 3 formes différentes de glycosylation aux stades pro-B, pré-B et B mature, alors que
Igβ se trouve sous une seule forme. La différence de forme pourrait également dépendre de
l'isotype avec lequel elle entre en association (Batista et al., 1996; Chen et al., 1990).
L’aggrégation des BCR suite à la reconnaissance de l’antigène entraîne leur translocation
dans des micro-domaines membranaires spécifiques appelés « rafts » (ou radeaux lipidiques) et la
phosphorylation des ITAMs d’Igα et d’Igβ par les PTKs. Les rafts agissent comme une plate-
forme de signalisation et de trafic : structure rigide qui évolue dans la « mosaïque fluide » qu’est
la membrane plasmique (Simons and Toomre, 2000) et permet ainsi la stabilisation spatiale du
complexe BCR. Ils sont riches en sphingolipides et cholestérol. Il a été montré que l’association
du BCR avec les micro-domaines lipidiques variait au cours du développement et serait donc un
mécanisme efficace dans le contrôle de la signalisation en régulant la rencontre du récepteur avec
d’autres partenaires tels que les protéines kinases, le substrat PIP2… Le pré-BCR est en majorité
localisé dans les micro-domaines d’où il transmet un signal de survie (Guo et al., 2000) tandis
que le BCR de cellules B au repos y serait exclu (Cheng et al., 1999).
Figure 21 : Translocation des BCR dans les rafts après crosslinking avec un antigène.
cholestérol
sphingolipides
phospholipides
phospholipides saturés
Ag crosslinking
BCR
Igαααα/β rafts
CD45
Lyn
Lyn
Lyn
Lyn
ITAM
Syk
mIg
Syk
PP
PP
a.
b. BCR
CD45
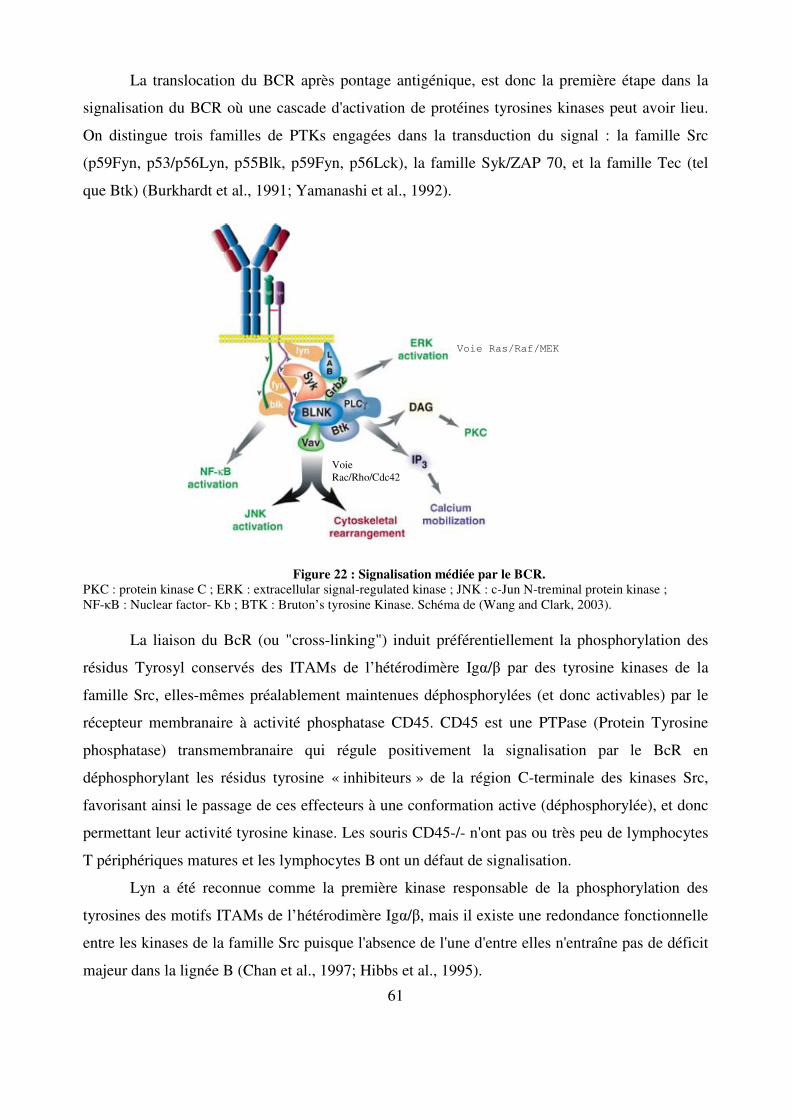
61
La translocation du BCR après pontage antigénique, est donc la première étape dans la
signalisation du BCR où une cascade d'activation de protéines tyrosines kinases peut avoir lieu.
On distingue trois familles de PTKs engagées dans la transduction du signal : la famille Src
(p59Fyn, p53/p56Lyn, p55Blk, p59Fyn, p56Lck), la famille Syk/ZAP 70, et la famille Tec (tel
que Btk) (Burkhardt et al., 1991; Yamanashi et al., 1992).
Figure 22 : Signalisation médiée par le BCR. PKC : protein kinase C ; ERK : extracellular signal-regulated kinase ; JNK : c-Jun N-treminal protein kinase ; NF-κB : Nuclear factor- Κb ; BTK : Bruton’s tyrosine Kinase. Schéma de (Wang and Clark, 2003).
La liaison du BcR (ou "cross-linking") induit préférentiellement la phosphorylation des
résidus Tyrosyl conservés des ITAMs de l’hétérodimère Igα/β par des tyrosine kinases de la
famille Src, elles-mêmes préalablement maintenues déphosphorylées (et donc activables) par le
récepteur membranaire à activité phosphatase CD45. CD45 est une PTPase (Protein Tyrosine
phosphatase) transmembranaire qui régule positivement la signalisation par le BcR en
déphosphorylant les résidus tyrosine « inhibiteurs » de la région C-terminale des kinases Src,
favorisant ainsi le passage de ces effecteurs à une conformation active (déphosphorylée), et donc
permettant leur activité tyrosine kinase. Les souris CD45-/- n'ont pas ou très peu de lymphocytes
T périphériques matures et les lymphocytes B ont un défaut de signalisation.
Lyn a été reconnue comme la première kinase responsable de la phosphorylation des
tyrosines des motifs ITAMs de l’hétérodimère Igα/β, mais il existe une redondance fonctionnelle
entre les kinases de la famille Src puisque l'absence de l'une d'entre elles n'entraîne pas de déficit
majeur dans la lignée B (Chan et al., 1997; Hibbs et al., 1995).
Voie Ras/Raf/MEK
Voie Rac/Rho/Cdc42
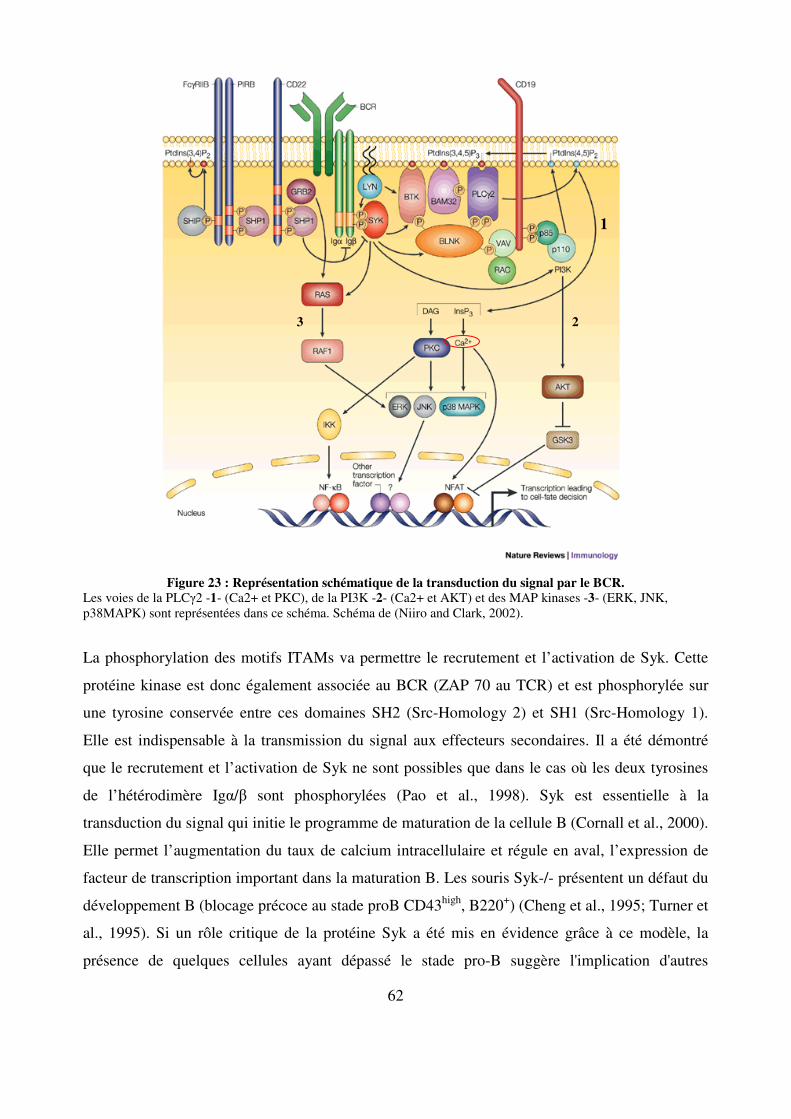
62
Figure 23 : Représentation schématique de la transduction du signal par le BCR.
Les voies de la PLCγ2 -1- (Ca2+ et PKC), de la PI3K -2- (Ca2+ et AKT) et des MAP kinases -3- (ERK, JNK, p38MAPK) sont représentées dans ce schéma. Schéma de (Niiro and Clark, 2002).
La phosphorylation des motifs ITAMs va permettre le recrutement et l’activation de Syk. Cette
protéine kinase est donc également associée au BCR (ZAP 70 au TCR) et est phosphorylée sur
une tyrosine conservée entre ces domaines SH2 (Src-Homology 2) et SH1 (Src-Homology 1).
Elle est indispensable à la transmission du signal aux effecteurs secondaires. Il a été démontré
que le recrutement et l’activation de Syk ne sont possibles que dans le cas où les deux tyrosines
de l’hétérodimère Igα/β sont phosphorylées (Pao et al., 1998). Syk est essentielle à la
transduction du signal qui initie le programme de maturation de la cellule B (Cornall et al., 2000).
Elle permet l’augmentation du taux de calcium intracellulaire et régule en aval, l’expression de
facteur de transcription important dans la maturation B. Les souris Syk-/- présentent un défaut du
développement B (blocage précoce au stade proB CD43high, B220+) (Cheng et al., 1995; Turner et
al., 1995). Si un rôle critique de la protéine Syk a été mis en évidence grâce à ce modèle, la
présence de quelques cellules ayant dépassé le stade pro-B suggère l'implication d'autres
1
2 3

63
protéines. La protéine ZAP-70 était décrite comme étant exclusivement exprimée dans les
cellules T et dans les cellules NK (Chan et al., 1992), mais son implication dans la transduction
du signal des cellules B a récemment été démontrée (Schweighoffer et al., 2003). En effet, les
cellules déficientes à la fois pour Syk et ZAP-70 présentent un blocage complet au stade Pré-B.
ZAP-70 semble donc être exprimée pendant le développement B et interviendrait dans la
transduction du signal.
Syk participe à la voie de la PLC-γ2. Elle va former un complexe avec la PLC-γ2
(phospholipase C gamma2), isoforme prédominante dans les lymphocytes B, par l’association de
la tyrosine phosphorylée de Syk au domaine SH2 de la PLC-γ2. Syk phosphoryle la Tyrosine 783
de la PLC-γ2 ce qui va aboutir, au final, à l'augmentation de la concentration du Ca2+
cytosolique. La PLC-γ2 est recrutée à la membrane cellulaire par l’intéraction de son domaine PH
(phospholipid-binding pleckstrin-homology domain) et réalise l'hydrolyse du PIP2 (Phosphatidyl
Inositol 4-5 biphosphate) en IP3 (inosito 1-4-5 triphosphate) et DAG (diacylglycérol). L'IP3
permet la libération dans le cytosol du Ca2+ stocké dans les vésicules intra-cytoplasmiques.
L’augmentation du niveau de la concentration intracellulaire du calcium est nécessaire à
l’activation de facteurs de transcription comme NF-κB (Nuclear Factor-κB) et NFAT (Nuclear
Factor of activated T-cells). Le DAG active quant à lui, la protéine kinase C (PKC) qui peut ainsi
être transloquée dans le cytoplasme où elle pourra phosphoryler les résidus sérine/thréonine
d'autres protéines. Les phosphoprotéines devront agir ensuite comme des facteurs de transcription
de différents gènes, tels que les oncogènes c-fos et c-myc. Les PKC semblent également
participer à l'activation de la voie Ras par le BCR, que l'on décrira plus loin, puisque un
inhibiteur des PKC bloque la voie de l'activation des MAP (Microtubule Associated Protein)
kinases au cours de l'activation des cellules immatures (Gold et al., 1992). Les GTPases de la
famille Vav jouent également un rôle crucial dans la signalisation via le BCR, notamment dans
l’activation de rac1, la réorganisation du cytosquelette après l’engagement du BCR et la
mobilisation du Ca2+ (Tedford et al., 2001). Néanmoins, l'activation de PLC-γ2 et le flux de
Ca2+ intracellulaire sont insuffisants pour déclencher la prolifération des lymphocytes B. L'influx
de Ca2+ extracellulaire apparaît indispensable pour entraîner la prolifération et la synthèse de
cytokines induites par le BcR. C'est sous l'action de la tyrosine kinase de la famille Tec, Btk pour
Bruton agammaglobulinemia Tyrosine Kinase (Hashimoto et al., 1999) (Itk et Rlk dans les
lymphocytes T), relayée par la molécule adaptatrice Blnk/SLP-65 (pour B-cell linker protein) que
se fera cet influx de Ca2+ extracellulaire. Des mutations de Btk sont la cause de déficits

64
immunitaires : XLA (X-linked agammaglobulinemia ou Bruton agammaglobulinemia) chez
l'homme et XID (X-linked immunodeficiency) chez la souris. Il a été également montré que Btk
se liait directement à Igα au niveau de sa queue cytoplasmique et jouerait certainement un rôle
dans l’initiation de l’activation du signal (Kabak et al., 2002).
La deuxième voie de signalisation est la voie de la PI3-Kinase. On distingue 3 groupes
dans cette famille : PI3KI, PI3KII, et PI3KIII. Le mode d'action des acteurs de cette voie reste
controversé, mais son importance a été validée par l'utilisation d'inhibiteurs de PI3K (Beckwith et
al., 1996). La PI3K est impliquée dans de nombreux processus biologiques cellulaires comme la
différenciation, la survie, la migration ou le métabolisme. Dans le système immunitaire, des
dérèglements de la signalisation de la PI3K entraînent des immunodéficiences ou leucémies et
des maladies auto-immunes dans le cas d’une hyperactivation de la PI3K. Elle régule le
développement et la différenciation des précurseurs de la moelle osseuse ainsi que l’activation et
la prolifération des cellules B matures. Les souris déficientes pour la sous-unité régulatrice p85α
présentent des défauts profonds dans le fonctionnement des cellules B : prolifération diminuée et
survie altérée avec un blocage partiel au stade pro-préB accompagné d’un nombre de B matures
diminué en périphérie (Fruman et al., 1999; Suzuki et al., 2003).
Le co-récepteur CD19 semble jouer un rôle important dans cette voie. En effet, au cours
de l'engagement du BCR, il peut y avoir co-ligation avec plusieurs autres récepteurs, tel que
CD19, entraînant la phosphorylation de leurs parties cytoplasmiques. CD19 pourrait alors
recruter la PI3K. Cette association se fait grâce au domaine SH2 de la sous-unité p85, ce qui
entraîne la phosphorylation de tyrosines impliquées dans l'activation de cette protéine (Fearon
and Carter, 1995). Cette protéine voit également son activité augmenter par la liaison de sa sous-
unité avec le domaine SH3 des PTK, Fyn et Lyn (Yamanashi et al., 1991). Il y aurait donc
formation d'un complexe multiprotéique après l'engagement du BCR, dans lequel PI3K serait liée
simultanément avec CD19 et Fyn ou Lyn (DeFranco, 1997). Activée, la PI3K va phosphoryler en
position 3’ les phosphatidylinositol 4-5 biphosphate (PIP2) pour générer les phosphatidylinositol
3-4-5 triphosphate (PIP3). La production de PIP3 est un élément central dans la signalisation du
BCR car beaucoup d’effecteurs impliqués dans la cascade, contiennent des domaines PH pouvant
se lier aux PIP3. Dans la signalisation du BCR, une des cibles de la PI3K activée est la
sérine/thréonine kinase Akt, également appelée protéine kinase B (PKB) (Franke et al., 1997;
Klippel et al., 1997). Une fois recrutée et activée, Akt est transloquée dans le noyau. Elle favorise
la survie cellulaire en régulant par phosphorylation des facteurs de transcription de « survie »

65
comme NF-κB (Krappmann et al., 2001) et E2-F mais peut aussi transmettre un signal anti-
apoptotique en phosphorylant Bad, protéine de la famille Bcl-2 (del Peso et al., 1997) ou en
inhibant GSK-3 (glycogen synthase kinase-3). L’activation rapide et transitoire de NF-κB en
réponse à tout type de stimuli, implique généralement la phosphorylation de IκB par le complexe
kinase IKK, qui permet la libération de NF-κB et sa translocation dans le noyau afin d’initier les
transcriptions.
Une autre voie initiée après l'engagement du BCR est la voie des MAP-kinases qui
regroupe trois membres : ERK (Extracellulaire signal-regulated kinase), JNK/SAPK (c-Jun NH2-
terminal kinase) et p38 MAPK (Johnson and Lapadat, 2002). Après leur activation en cascade,
ces kinases vont jouer leur rôle dans la signalisation en phosphorylant différents groupes de
facteurs de transcription : Elk-1 et c-Myc (pour ERK), c-jun et ATF-2 pour activating
transcription factor 2 (pour JNK) et ATK-2 et MAX (pour p28 MAPK) qui pourront se fixer sur
les régions promotrices des gènes impliqués dans l'activation B.
Des enzymes ayant une activité GTPasique comme Ras et Rac-1 vont intervenir en amont de
cette voie de signalisation. Par exemple, Ras contrôle la cascade Raf-1/MEK/ERK. L’activation
de cette voie est essentielle au développement des cellules B matures (Iritani et al., 1997). La
phosphorylation des ITAMs va permettre le recrutement indirect de la molécule adaptatrice Shc
qui va former un complexe d’activation de Ras avec les molécules Grb2 et SOS : passage de la
forme Ras-GDP à la forme Ras-GTP induisant ainsi la voie classique des MAP kinases.
La signalisation du BCR est donc une voie régulée par :
- la concentration de l’antigène et la force de liaison entre l’antigène et le récepteur (Casola et
al., 2004; Pillai et al., 2004).
- l’auto-régulation du BCR : la phosphorylation de l’hétérodimère Igα/β peut dicter la qualité
et la quantité du signal (Choquet et al., 1994; Luisiri et al., 1996).
- la présence de co-récepteurs membranaires.
On peut détailler une partie des corécepteurs membranaires du BCR jouant un rôle dans la
régulation de la signalisation tout en sachant que cette régulation est sans doute encore plus vaste
et plus complexe…
CD19, co-récepteur, associé au BCR, favorise son entrée dans les rafts. Il contribue
positivement à la signalisation induite par Igα/Igβ en favorisant le recrutement de Lyn, Fyn, PI3K
(Buhl and Cambier, 1999; Cherukuri et al., 2001) mais également de Vav. CD19 forme un

66
complexe avec les récepteurs CD81 et CD21, le récepteur du C3d (Otero et al., 2001). Ce
complexe collabore avec le BCR pour permettre aux lymphocytes B de répondre à de faibles
concentrations d'antigènes. CD45 n’est jamais recruté dans les rafts après l’engagement du BCR
et son absence permet une signalisation du BCR via une phosphorylation de Lyn plus importante
(Cheng et al., 1999; Katagiri et al., 1999).
CD22 est une glycoprotéine transmembranaire de type I spécifique des lymphocytes B qui
lie des résidus acides sialiques liés en α2,6. Elle appartient à la famille des Siglecs (Sialic acid-
binding Ig-like lectins). Cette molécule est exprimée très tôt au cours du développement B
(transition pro-B/pré-B) et son taux d’expression à la surface des cellules B augmente durant la
maturation, atteignant un maximum au stade B mature (Moyron-Quiroz et al., 2002) et
disparaissant au stade plasmocyte. Ce récepteur est aussi retrouvé à la surface de lymphocytes B
isolés des différents compartiments lymphoïdes. Il est associé au BCR et est connu pour réguler
plutôt négativement la transduction du signal (Nitschke et al., 1997), constituant une boucle de
rétro-contrôle. Il a été démontré que les 3 motifs ITIMs (Immunoreceptor Tyrosine-based
Inhibitory Motifs) de son domaine cytoplasmique sont phosphorylés par Lyn au niveau de leur
tyrosine juste après l’engagement du BCR. Ceci permettrait l’accrochage de plusieurs protéines
(contenant des domaines SH2) telles que les tyrosines phosphatases SHP-1 et SHP-2 qui auraient
alors une activité inhibitrice sur le BCR (Nitschke, 2005). Les souris déficientes en CD22
répondent fortement à une stimulation du BCR par une augmentation du taux de Ca2+ et par une
forte prolifération cellulaire (O'Keefe et al., 1996) : « hyper-réponse » des cellules B. De plus,
cette réponse exagérée est associée à une production importante d'auto-anticorps (O'Keefe et al.,
1999). Les substrats de SHP-1 regroupent entre autre, Syk, Blnk, CD22 lui-même (Mizuno et al.,
2000). La déphosphorylation de Blnk par SHP-1 empêche le recrutement de la PLC-γ2 et la
déphosphorylation de Syk et des PTKs de la famille Src diminue leur activité et empêche la
phosphorylation des motifs ITAMs du BCR. Donc en conclusion, l’activation de Lyn va jouer
deux rôles : activateur, en initiant la signalisation du BCR via la phosphorylation des ITAMs de
l’hétérodimère Igα/β et inhibiteur par phosphorylation des motifs ITIMs de CD22 aboutissant au
recrutement de SHP-1. Le phénotype « hyper-immun » des souris CD22-/- est proche de celui
observé chez les souris déficientes pour la phosphatase SHP-1 de même que l’étude de plusieurs
lignées B déficientes en CD22 conforte la thèse du rôle inhibiteur de CD22. Dans ces lignées, une
réponse exagérée est également observée à la suite d'une stimulation antigénique avec un flux
calcique augmenté de manière significative (Nadler et al., 1997). Razi et Varki (1998) ont

67
démontré que CD22 était continuellement « masquée » par des ligands en cis présents à la surface
des lymphocytes B et qu’il pouvait être « démasqué » suite à une stimulation avec un anticorps
anti-IgM/CD40 (Razi and Varki, 1998). Les interactions avec les ligands en cis peuvent dominer
d’autres interactions avec des ligands en trans ce qui modulerait l’activité de CD22. La structure
sialoside terminale reconnue par CD22 est synthétisée in vivo par l’enzyme ST6GalI
(sialyltransférase). Les souris déficientes en ST6GalI suggèrent un rôle complexe ou double de
CD22 puisqu’elles ont une immunodépression B (après immunisation avec des antigènes T-
dépendants ou T-indépendants), présentent de faibles taux sériques d’IgM, et une prolifération B
diminuée (Collins et al., 2002), alors que le double KO CD22/ST6gal1 a montré une restauration
de la réponse B (flux calcique et prolifération après pontage du BCR) (Collins et al., 2006b).
CD22 est localisé préférentiellement au niveau des domaines riches en clathrine et est exclu des
rafts après activation et translocation du BCR mais, se rapprocherait du BCR (pour jouer son rôle
de régulateur négatif) lors de la fusion rafts avec les domaines riches en clathrine pour
l’internalisation (Collins et al., 2006a). Il a été montré que dans les souris ST6Gal1-/-, CD22 était
associé avec des IgM de surface dans les B au repos. On peut donc suggérer que la suppression
du signal B dans les souris ST6Gal1-/- est médiée par une colocalisation ou un rapprochement de
CD22 avec le BCR. Le domaine lectine de CD22 et sa liaison à des ligands glycosylés ne
semblent donc pas à ce jour être clairement associés à sa fonction inhibitrice de l’activation (Poe
et al., 2004).
Le récepteur de faible affinité pour les fragments Fc des IgG, appelé FcγRIIB ou CD32
peut également exercer un effet négatif sur la signalisation du BCR et inhibe la prolifération. De
façon similaire à CD22, le signal inhibiteur est assuré par Lyn qui va également phosphoryler les
motifs ITIMs de FcγRIIB favorisant le recrutement des tyrosines phosphatases SHP-1 et SHP-2.
Ces dernières agissent directement en hydrolysant les PIP3 en PIP2 ou indirectement en
déphosphorylant CD19 bloquant le recrutement et l’activation de la PI3K (Fong et al., 2000;
Hippen et al., 1997).
Le récepteur membranaire CD40, quant à lui, appartient à la famille des récepteurs de
TNF. Il induit un signal mitogénique par interaction avec son ligand CD40L (CD154) exprimé
par les lymphocytes T activés.
Dernièrement, un nouveau récepteur membranaire inhibiteur spécifique des cellules B1 a
été caractérisé (Hoffmann et al., 2007). Siglec-G (orthologue de la siglec-10 humaine) est un
membre de la famille des siglec tout comme CD22 mais plutot relié à la sous-famille de CD33

68
(siglec3). Siglec-G ne serait exprimé que dans les cellules B avec une plus forte expression dans
les cellules B1. Les souris Siglec-G-/- présentent une expansion du compartiment B1 dans la
cavité péritonéale, une plus forte mobilisation du calcium intracellulaire après pontage avec un
anti-IgM dans les cellules B1 et un fort taux d’IgM sérique.
CD72 est également un co-récepteur membranaire inhibiteur du BCR, du fait de la
présence d’un motif ITIM dans son domaine cytoplasmique. En réponse à une stimulation
antigénique, CD72 régulerait négativement toute la voie de transduction du signal en aval du
BCR incluant NF-AT, NF-κB, ERK, JNK, MAPK mais aussi AKT ce qui aboutirait à un arrêt du
cycle cellulaire et à la mort des cellules primaires B matures (Li et al., 2006).
Les mécanismes de transduction du signal semblent être partagés par les récepteurs pré-B et B
compte tenu des différents blocages démontrés dans les souris déficientes en Syk, Btk, Blnk,
PI3K, Lyn, CD19…. récapitulés dans la Figure 14 du chapitre II.
4.2. La signalisation en fonction des différents isotypes du BCR
Quelqu’en soit l’isotype, la présence d’un BCR à la membrane du lymphocyte B est
indispensable à sa survie (Lam et al., 1997). La signalisation via le BCR ne se limite cependant
pas à la survie mais contrôle au contraire de multiples choix (notamment entre apoptose, anergie,
prolifération et différenciation) tout au long de l’ontogénie B. En contraste avec cette complexité
et cette variété des choix cellulaires, la description d’une structure univoque du BCR, associant
toujours une molécule d’immunoglobuline membranaire à un hétérodimère Igα/Igβ, et
l’identification des motifs de signalisation ITAM portés par les régions intracellulaires de ces
deux peptides (Venkitaraman, Reth, etc…) pourraient donner à penser que toutes les
immunoglobulines membranaires transduisent le même type de signal « limité » à une
signalisation α/β lors de leur pontage par un ligand. Une comparaison des différentes classes de
chaînes lourdes entre elles et au cours de l’évolution suggère cependant que la structure des trois
domaines (extra-cellulaire / transmembranaire et intra-cellulaire) de l’immunoglobuline elle-
même pourrait moduler cette signalisation.
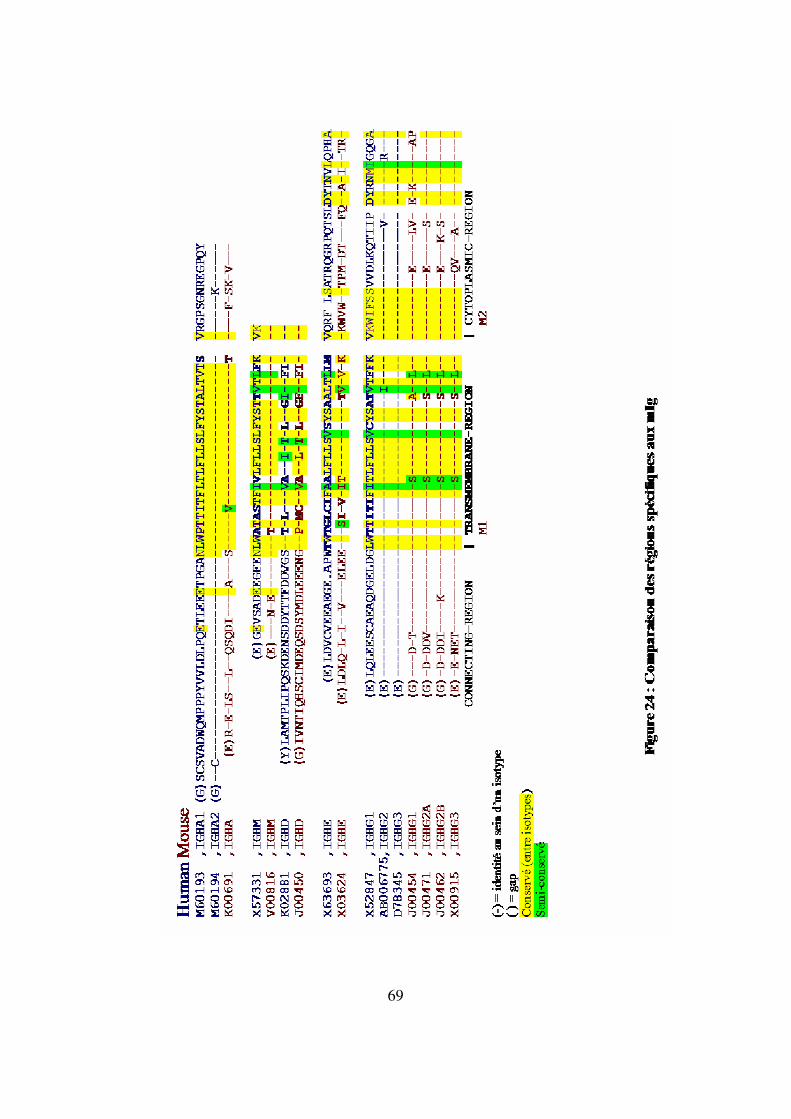
69
Figure 24 : Comparaison des régions spécifiques aux mIg

70
La partie transmembranaire des immunoglobulines membranaires est très conservée au cours de
l'évolution, ce qui témoigne de son importance fonctionnelle (Bensmana and Lefranc, 1990;
Rogers and Wall, 1984). Elle est quasiment identique entre la souris, le lapin, et l'homme (Figure
24). Cette conservation se retrouve également entre les différents isotypes (Rogers and Wall,
1984; Yamawaki-Kataoka et al., 1982) et il semble donc vraisemblable que ce domaine
transmembranaire ait une fonction unique d’une espèce à l’autre et d’un isotype à l’autre. Sur les
25 acides aminés composant le domaine transmembranaire, 10 sont des résidus Sérine/Thréonine
ou Tyrosine. Ces résidus hydrophiles sont impliqués dans l'interaction avec les domaines
transmembranaires d'autres molécules, notamment l'hétérodimère Igα/Igβ.
À l’inverse, une importante variabilité inter-isotypes, suggestive de variations
fonctionnelles, apparaît au niveau des domaines intra-cytoplasmiques. Leur taille varie en
fonction des isotypes : IgM et IgD comptent seulement trois résidus dans cette partie, IgA 14,
IgG et IgE en comptent 28. La taille des isotypes commutés, nettement plus importante, peut
donc suggérer que cette queue intra-cellulaire serait d’une façon ou d’une autre gênante pendant
le développement précoce de la cellule B et serait donc absente des deux isotypes qui s’expriment
à ces stades précoces. De plus, on note la présence d’une tyrosine conservée dans un motif
commun à γ et ε.
Figure 25 : Séquences des domaines cytoplasmiques.
IgM ---VTLF KVK
IgD ---VTFI KVK
IgG1 ---LTVT KVKWIFSSVVELKQTLVPEYKNMIGQAP
IgG2a ---LTVT KVKWIFSSVVELKQKISPDYRNMIGQGA
IgG2b ---VTLF KVKWIFSSVVELKQKISPDYRNMIGQGA
IgG3 ---VTLF KVKWIFSSVVQVKQTAIPDYRNMIGQGA
IgA ---LTVT TVRGPFGSKEVPQY
IgE ---LTVT KVKWVLSTPMQDTPQTFQDYANILQTRA
domaines transmembranaires domaines cytoplasmiques
IgM ---VTLF KVK
IgD ---VTFI KVK
IgG1 ---LTVT KVKWIFSSVVELKQTLVPEYKNMIGQAP
IgG2a ---LTVT KVKWIFSSVVELKQKISPDYRNMIGQGA
IgG2b ---VTLF KVKWIFSSVVELKQKISPDYRNMIGQGA
IgG3 ---VTLF KVKWIFSSVVQVKQTAIPDYRNMIGQGA
IgA ---LTVT TVRGPFGSKEVPQY
IgE ---LTVT KVKWVLSTPMQDTPQTFQDYANILQTRA
domaines transmembranaires domaines cytoplasmiques

71
Au cours du développement B, le premier isotype à être exprimé est l'IgM. Le domaine
cytoplasmique de cet isotype, ainsi que celui d'IgD est court et n'est composé que de 3 aa. IgM et
IgD sont donc entièrement dépendantes de l'hétérodimère Igα/Igβ pour assurer les fonctions de
transduction du signal. L’intérêt pour des cellules matures au repos (resting B cells) d’exprimer à
la fois deux isotypes différents de même spécificité antigénique, IgM et IgD, n’est pas encore
bien compris. Comme tous les isotypes, le cross-linking d’IgM et d’IgD entraîne l'activation des
PTK et la phosphorylation de leurs substrats. Mais on sait que l'intensité du signal d’activation est
dépendante de l'isotype. La réponse médiée via IgD est plus intense et plus durable que celle
médiée via IgM et cette différence serait due à son extrémité C-proximale ainsi qu’à un profil de
phosphorylation différent (Kim and Reth, 1995). La similitude de leurs domaines cytoplasmiques
leur permet de se remplacer l'un l'autre. Des souris déficientes pour IgM (délétion de Cµ et de
l’intron µ-δ) présentent un développement B quasi-normal avec expression membranaire et
sécrétée d’IgD. Les différents compartiments B de la moelle et des organes lymphoïdes
secondaires (compartiments B conventionnels - B2) sont normaux. On note une légère différence
dans le péritoine où le nombre de cellules B1 (B220+ CD5+) est diminué dans les souris IgM-/-
par rapport aux souris WT. Après stimulation antigénique (T-dépendante ou T-indépendante), il y
a formation de centres germinatifs, commutation de classe et réponse spécifique à l’antigène. La
seule anomalie rapportée est un retard des réponses antigène-spécifiques. IgD est donc capable de
se substituer en grande partie à IgM (Lutz et al., 1998). C’est en fait l’expression conjointe d’IgM
et d’IgD qui semble présenter un certain degré de redondance non indispensable aux réponses
immunes mais capable de les optimiser. En effet et réciproquement, les souris déficientes pour
IgD (Roes and Rajewsky, 1993), ont un nombre normal de cellules B (conventionnels B2 ou les
B1), une expression membranaire des deux recepteurs CD23 et CD22 (marqueurs d’activation) et
une réponse quasiment aussi efficace que chez les souris sauvages aux antigènes T-dépendants ou
T-indépendants. Par contre, les IgD-/- ont un retard dans la maturation d’affinité après
stimulation avec un Ag T-dépendant. IgD semblerait être important pour accélérer, optimiser le
recrutement de cellules B hautement spécifiques dans les étapes précoces d’une réponse primaire
T-dépendante. En conclusion, l’expression précoce de µ ou de δ est suffisante (mais non
optimale) pour induire le développement B. La formation du pré-BCR semble possible que ce
soit avec la chaîne µ ou avec la chaîne δ. Ceci est certainement du à la présence du court domaine
cytoplasmique identique que ce soit pour IgM ou IgD. IgM comme IgD peut médier l'activation
(Sieckmann et al., 1982), et les deux récepteurs semblent transduire le signal par le même
mécanisme (Cambier et al., 1987).

72
L’absence conjointe de µ et δ aboutit à un phénotype bien plus délétère pour la lignée B.
Elle est observée chez les souris µMT où un codon stop (suivi d’une cassette NéoR) a été inséré
dans le premier exon de membrane µM1 de Cµ (Kitamura et al., 1991). Ces souris présentent
lorsqu’elles sont sur fond génétique mixte C57Bl6/129 un blocage complet au stade préB.
Aucune expression membranaire d’IgM et d’IgD n’est détectée. Il n’y a pas de B matures, de
plasmocytes ni d’Ig dans le sérum. L’étude des réarrangements des chaînes lourdes a montré que
de nombreuses cellules possédaient des réarrangements VDJ productifs sur les deux allèles et que
les cellules B sécrétrices d’anticorps issues d’animaux hétérozygotes exprimaient deux chaînes
lourdes (Kitamura and Rajewsky, 1992). Une étude plus approfondie de ces animaux a en fait
permis de montrer qu’au bout de plusieurs mois, ils finissaient par peupler leur tractus digestifs
de cellules commutées vers la synthèse d’IgA, qui avaient pu quitter la moelle osseuse sous
forme de progéniteurs B et bénéficier ensuite d’un milieu cytokinique propice à la commutation
de classe IgA pour récupérer de cette façon l’expression d’un BCR fonctionnel (Macpherson et
al., 2001). Au-delà, Hasan et al en 2002 puis Orinska et al, en 2002, ont montré, chez les souris
µMT sur fond génétique BALB/c, la présence d’une petite population B à IgG, IgA ou IgE de
membrane, capable de reconnaître spécifiquement des antigènes et de se différentier en
plasmocytes sécréteurs d’anticorps (Hasan et al., 2002; Orinska et al., 2002). Les auteurs ont émis
l’hypothèse qu’une petite partie des précurseurs µ-/- s’échappait de la moelle (en échappant à la
sélection « positive » pour l’expression d’un BCR fonctionnel) et pouvait switcher en périphérie
vers les différents isotypes (avec le risque que ce chemin alternatif de différenciation ne laisse
émerger des clones auto-réactifs potentiellement responsables de maladies auto-immunes…).
Comme nous l’avons vu dans le chapitre 3, lors d'une réponse secondaire, la commutation
de classe permet l'expression d'un nouvel isotype chez la souris : IgG3, IgG1, IgG2b, IgG2a, IgE
ou IgA. La taille des domaines cytoplasmiques de ces isotypes est plus grande que celle d’IgM et
d’IgD et comporte notamment une tyrosine conservée. Sans que son rôle dans un BCR complet
assemblé ne soit clarifié, un motif commun à toutes les IgG et aux IgE a pu être défini autour de
cette tyrosine (motif Tyr-X-X-Met/Ile) comme jouant un rôle dans l’internalisation du BCR IgG
en l’absence d’association à Igα/Igβ (Knight et al., 1997; Weiser et al., 1997). Le rôle de ces
domaines cytoplasmiques et leur degré de dépendance vis-à-vis de l'hétérodimère Igα/Igβ sont
mal connus mais on sait que tous les isotypes sont normalement exprimés à la surface de la
cellule en association avec l'hétérodimère Igα/Igβ, même si une stabilité du récepteur

73
membranaire s’avère possible expérimentalement pour les IgG en absence de l’hétérodimère
(Abney et al., 1978; Knight et al., 1997; Venkitaraman et al., 1991; Weiser et al., 1994).
Au cours de ce travail de thèse, nous avons voulu savoir si un BCR commuté à IgA où à
IgE pouvait permettre un développement B normal chez la souris, ainsi qu’une réponse
spécifique à l’antigène. Avant d’aborder la partie plus expérimentale de mon travail, je vais vous
détailler dans ce chapitre, les différents modèles de souris transgéniques (KO, KI et transgenèse)
déjà établis dans le but de susbstituer à l’expression d’un BCR normal IgM/IgD celle d’un BCR
muté, commuté ou pas.
Roth et al., en 1993, ont montré qu’une chaîne lourde transgénique γ2b bloquait
partiellement le réarrangement et l’expression de la chaîne µ endogène mais ne pouvait ensuite se
substituer à cette chaîne µ. Les souris porteuses du transgène γ2b présentent donc un déficit en
cellules B. Les jeunes souris n’ont pratiquement pas de cellules B. A 4 semaines, les souris ont
environ 10% de B dans la rate et n’arrivent à accumuler un nombre normal de cellules B qu’à
partir de 16 semaines. Toutes les cellules B qui se développent dans ces conditions expriment la
chaine µ endogène (comme c’est le cas dans les souris transgéniques human γ1 qui n’ont pas de
cellules B exprimant uniquement le transgène γ1 (Yamamura et al., 1986)). γ2b ne peut donc pas
promouvoir la maturation B et ceci ne serait pas dû à un effet toxique du transgène puisqu'un
croisement avec des souris transgéniques µ restaurerait la maturation B et un croisement avec les
souris µ-membrane KO, ne restaurerait pas le compartiment B. L'expression d'IgG2b est
retrouvée dès les stades précoces du développement et induirait un fort « feed-back » négatif sur
l'expression des gènes des chaînes lourdes endogènes (Denis et al., 1990; Roth et al., 1993; Shen
et al., 1999). Une fois ce stade dépassé, le rôle anti-apoptotique d'IgM ainsi que le signal de
différenciation cellulaire ne pourraient donc être assurés par cette chaîne γ2b transgénique. Ces
différents résultats ont été montrés dans plusieurs lignées transgéniques fondatrices différentes,
porteuses du même transgène. Une seule lignée transgénique (« C line »), exprimant le même
transgène γ2b, permettrait un développement normal après croisement avec les souris µ-
membrane KO). Le site d'intégration et/ou le niveau d’expression du transgène semblent donc
moduler ce phénotype. On ne peut pas exclure non plus dans ces expériences de transgenèse, un
rôle de la région variable, la restriction du répertoire à un VHDJH particulier pouvant peut-être
avoir des effets imprévisibles sur la maturation B. Mais si la maturation B dans ces souris (« C
line ») ne nécessite pas l'expression d'IgM, elle semble dépendante de l'expression de la chaîne λ5
en association avec un facteur inconnu (Kurtz et al., 1997; Roth et al., 1995).

74
Certains auteurs (Achatz et al., 1997; Kaisho et al., 1997; Martin and Goodnow, 2002) ont
suggéré l'implication de certains domaines cytoplasmiques d'Ig commutées dans la mémoire
immunologique. Achatz et al. en 1997 ont établi deux lignées de souris différentes portant des
mutations au niveau des domaines constant ε. La lignée ∆M1M2 est délétée des domaines
transmembranaires et cytoplasmiques d’IgE alors que la lignée KVK∆tail est quant à elle délétée
du domaine cytoplasmique ε et a, à la place, le domaine cytoplasmique d’IgM/d’IgD « KVK ».
Dans les deux cas la réponse spécifique après immunisation est fortement compromise. En effet,
la sécrétion d'IgE spécifique est négligeable dans les souris sans domaine
transmembranaire/cytoplasmique ε. Lorsque seul le domaine cytoplasmique est absent, la
production d'IgE sérique est diminuée de moitié. On observe une nette diminution de la réponse
secondaire à l’Ag chez ces souris. Cette baisse de production d'IgE sérique est le résultat d'une
baisse de l'activité biologique associée à l'absence du domaine transmembranaire/cytoplasmique
d'IgE et non à un défaut de la commutation isotypique vers IgE.
Une autre étude (Kaisho et al., 1997) s'est intéressée au domaine cytoplasmique d'IgG1. A
partir de lignées de souris où le gène γ1 a été amputé des exons codant pour le domaine d’ancrage
membranaire de l’IgG1, elle montre la dépendance de l'expression membranaire de l'Ig pour
obtenir une réponse primaire et secondaire IgG1. Ces souris sont naturellement incapables de
produire des cellules B mémoires IgG1+. Comme pour les IgE, il est donc établi que le passage
par un stade d’expression membranaire de l’IgG1 est indispensable pour la production efficace de
plasmocytes sécréteurs de cette IgG1, en particulier en réponse secondaire.
Puisque, grâce à la commutation de classe, les cellules B vont exprimées un BCR
commuté à Ig contenant une queue intra-cytoplasmique que les cellules naïves ne possèdent pas
(IgM), la question qui reste en suspens est celle d’un rôle régulateur spécifique, modifiant le
« destin » des cellules commutées, pour cette région additionnelle du BCR commuté.
Cette question a d’abord été abordée par l’étude de trois lignées de souris transgéniques
exprimant des BCR spécifiques pour un même antigène (HEL : Hen-Egg Lysozyne), et mettant
en avant l'implication du domaine cytoplasmique d'IgG1 dans le développement B et dans la
mémoire immunologique. Sur ces 3 lignées exprimant une même région variable, une exprime le
transgène IgM, une autre le transgène IgG1, et une troisième un récepteur chimérique dont la
partie extracellulaire est celle de l'IgM et les parties transmembranaires et cytoplasmiques, celles
d'IgG1. Il est à noter qu’une chaîne légère anti-HEL kappa a également été co-transfectée. En
croisant ces différentes lignées transgéniques (comportant de faibles nombres de copies du
transgène) avec des souris RAG2-/-, comme ça a déjà été réalisé pour de simples transgènes

75
IgM/IgD (Brink et al., 1992; Spanopoulou et al., 1994), dans les 3 cas, il y a restauration du
développement B et maturation B complète (population mature CD21+ en périphérie). Dans ce
cas, IgG1 pourrait donc se substituer à IgM non seulement pour médier l’exclusion allélique (pas
ou peu de cellules co-exprimant IgM et IgG1) mais aussi pour assurer les signaux de maturation
des cellules B (Pogue and Goodnow, 2000). De plus, il apparaît que, dans ce cas, ce transgène
IgG1 anti-HEL via son domaine cytoplasmique, permettrait d'augmenter significativement
l’expansion clonale, la production d'Ac ainsi que le nombre de plasmocytes au cours d'une
réponse T-dépendante. Le domaine cytoplasmique d'IgG1 interviendrait certainement dans la
survie des cellules au cours des divisions cellulaires successives. Les auteurs suggèrent que pour
obtenir une réponse secondaire efficace, la présence de quelques cellules spécifiques de
l’antigène et exprimant de l’IgG membranaire suffit (Martin and Goodnow, 2002). Après
initiation et transduction du signal, l'activation doit cesser. Le signal médié par IgM est régulé
négativement, en partie par le co-récepteur inhibiteur CD22 comme nous l’avons vu dans le sous-
chapitre ci-dessus. La présence d’un BCR à IgG n’induirait pas la phosphorylation des motifs
ITIMs de CD22, ne permettrait donc pas le recrutement et l'activation de la phosphatase SHP-1 et
donc l'inactivation de protéines de signalisation. La queue cytoplasmique d’IgG serait donc
responsable à la fois de l'inhibition du signal médié via CD22 et de l'augmentation de la
transduction du signal observée dans les cellules IgG+. Ce qui a amené à proposer le modèle
selon lequel IgG serait indépendante du rétro-contrôle inhibiteur du signal par la phosphorylation
de CD22 afin de rendre l’IgG membranaire plus sensible à la stimulation antigénique, et obtenir
une réponse secondaire plus forte que celle obtenue avec IgM (Wakabayashi et al., 2002). Cette
hypothèse très séduisante a conduit à proposer l’existence de 2 catégories de BCR : un BCR
« affecteur » IgM exprimé précocément et très efficace pour la mise en place du répertoire, la
sélection négative et la maturation précoce de la lignée B, et un BCR « effecteur » (notamment
IgG) hyper-excitable et capable de contribuer à la forte réactivité du système immunitaire et à la
génération rapide de cellules sécrétrices d’anticorps de haute affinité lors des réponses
secondaires à l’antigène (Manser, 2002).
Sato et al., (2007) ont également montré, en établissant différents transfectants exprimant
chacun un BCR commuté (IgM, IgG, IgA et IgE) que, comme IgG, après pontage du BCR-IgE, il
y avait augmentation du taux de calcium intracellulaire associée à une plus forte phosphorylation
de ERK, évènements qui ne sont pas retrouvés après pontage d’un BCR à IgA ou IgM. Ils ont
également montré que CD22 jouerait son rôle de co-récepteur membranaire inhibiteur du signal
de prolifération, d’une manière isotype –dépendant. Il régulerait le signal médié par un BCR IgA

76
alors qu’il n’aurait pas d’effet inhibiteur sur un BCR IgE, du à une mauvaise association de
CD22 avec la queue cytoplasmique de l’IgE, générant ainsi une augmentation de signal. Par
contre, ils montrent que CD72 a un même effet inhibiteur que ce soit un BCR IgM, IgG, IgE ou
IgA. Ils ont par la suite montré que le motif conservé Tyr-X-X-(Met/Ile) commun à toutes les IgG
et aux IgE, n’intervenait pas dans cette augmentation du signal.
Si ces hypothèses sont séduisantes pour rendre compte de la physiologie des réponses humorales,
elles ont cependant, récemment, été remises en cause par Horikawa et al. et Waisman et al., en
avril 2007. L’augmentation du taux de calcium intracellulaire et de la réponse antigénique dans
les cellules B à BCR IgG ne serait pas due à une diminution de la phosphorylation de CD22 ou à
une diminution de sa fonction inhibitrice. Horikawa et al., ont montré que le signal médié par la
queue cytoplasmique de l’IgG n’induisait pas l’expression de certains gènes retrouvés beaucoup
plus exprimés dans des B à IgM de membrane (ces gènes seraient plutôt inducteurs d’une
différenciation MZ) et ils montrent que c’est cette absence ou ce faible signal qui induirait une
réponse B mémoire augmentée et orienterait les cellules vers la différenciation plasmocytaire
(théorie du « less is more ») (Horikawa et al., 2007). Waisman et al., quand à eux, ont démontré
en réalisant une lignée de souris γ1 KI, qu’un BCR IgG pouvait se substituer à un BCR IgM et
permettre un développement B normal. Ils montrent quand même, un léger blocage au stade pro-
préB mais n’identifient ni hyper-différenciation plasmocytaire (non explorable dans leur modèle),
ni indépendance par rapport à CD22 (Waisman et al., 2007).

77
V. « Glycannes et glycoconjugués »
Le dogme central de la biologie moderne a quelque temps reposé sur le concept d’un flux
d’informations biologiques se déplaçant de l’ADN vers l’ARN puis vers la protéine. Il est
cependant maintenant clair que la compréhension des mécanismes de biogenèse d’un organisme
ne repose pas uniquement sur la colinéarité de ces 3 molécules, mais aussi notamment sur 2
autres types de structures majeures que sont les lipides et les sucres, en particulier dans le cadre
des modifications post-traductionnelles des protéines. Polyosides et lipides servent de réserves
énergétiques, de signaux transducteurs, ou encore d’éléments structuraux. Toutes les cellules
vivantes portent sur leurs lipides ou leurs protéines un réseau complexe et dense
d’oligosaccharides (ou glycannes) formant ainsi des glycoprotéines et des glycolipides (dont nous
ne ferons pas état dans ce manuscrit). Localisés pour la plupart à la surface cellulaire, les
glycannes jouent un rôle fondamental dans les interactions “cellule-cellule” et “cellule-matrice
extracellulaire”, cruciales pour le développement d’organismes multicellulaires complexes. Ils se
trouvent également dans une position privilégiée pour l’établissement d’interactions avec
différents pathogènes. Ainsi, au cours des dernières décennies, les chercheurs ont par exemple
mis en évidence, leur importance dans la détermination des groupes sanguins (Kabat, 1956), leurs
rôles dans les interactions cellulaires (Roseman, 1970), leurs implications dans la fécondation
(Jego et al., 1980) et la différenciation erythrocytaire (Liu et al., 1981)...
La grande complexité des structures glycanniques, ajoutée au fait que leur séquence n’est pas
déterminable simplement en décodant une matrice d’acide nucléique, a considérablement freiné
les recherches, et ce n’est que depuis ces vingt dernières années que de nouvelles technologies,
destinées à étudier de façon approfondie ces molécules, ont permis l’essor d’une nouvelle
discipline connue sous le nom de « glycobiologie ». La glycobiologie allie les fondements de la
biochimie et de la biologie moléculaire des glucides. Son but est donc de décoder le sens de ce
troisième alphabet des sucres, qui s'ajoute à celui des protéines et des acides nucléiques. Ainsi,
bien que l’activité biologique des glycoprotéines soit généralement portée par la partie protéique,
les glycannes détiennent des informations supplémentaires permettant notamment de moduler la
fonction biologique des glycoprotéines, déterminer leur durée de vie, et signaler leur adressage
aux différents compartiments cellulaires.

78
5.1. La glycosylation
La glycosylation, généralement définie comme le principal événement de maturation post-
traductionnelle des protéines et des lipides, consiste en une succession de réactions enzymatiques
se déroulant, chez les eucaryotes, principalement dans le réticulum endoplasmique et dans
l’appareil de Golgi. Les glycoprotéines sont constituées d’une chaîne peptidique sur laquelle sont
greffés des sucres. Il est à noter que plusieurs résidus d’une même chaîne polypeptidique peuvent
être substitués par des sucres. Les enzymes impliquées, les glycosyltransférases, font preuve, à
l’image de leurs nombreux substrats, d’une très grande diversité fonctionnelle. Leur action
principale consiste à greffer des mono- ou des polysaccharides sur différents substrats
(monosaccharides, polysaccharides, lipides, ou peptides), en des sites bien définis. Les substrats
donneurs sont utilisés sous une forme énergétique (ou activée) de nucléotide-sucre (GDP-fucose,
UDP-galactose…), et plus rarement sous une forme lipide-sucre (dolichol-P-Glc) (Kleene and
Berger, 1993). Les glycosyltransférases eucaryotes sont en très grande majorité des
glycoprotéines membranaires de type 2, associées aux compartiments cellulaires
exocytoplasmiques, en particulier le réticulum endoplasmique et l’appareil de Golgi (Paulson and
Colley, 1989). Après un court domaine cytoplasmique N-terminal (≈10 acides aminés) et une
portion transmembranaire unique (≈ 15 acides aminés), se déploie la portion luminale de la
protéine formée d’un domaine intermédiaire flexible de longueur très variable (70 à 320 acides
aminés), assurant la liaison avec un domaine catalytique globulaire dont la taille varie de 200 à
250 résidus. Les glycosyltransférases sont nommées selon le sucre qu’elles transfèrent, par
exemple « mannosyltransférase » s’il s’agit du mannose, ou « fucosyltransférase », s’il s’agit du
fucose, mais aussi et surtout en fonction de la liaison formée (α ou β) entre le sucre transféré et le
substrat accepteur, ce dernier restant, dans la majorité des cas de nature mono- ou
polysaccharidique (Tableau 1). Dans les cellules animales, de nombreux travaux soulignent
l’importance de la régulation spatio-temporelle de l’expression de ces enzymes dans la définition
des épitopes glycanniques. Grâce au séquençage des génomes, nous savons aujourd'hui
qu'environ 1% des gènes d'un organisme leur est dédié (Lowe and Marth, 2003).

79
Tableau 1 : Les principales glycosyltransférases de vertébrés greffant des sucres monosaccharidiques sur des substrats mono- ou polysaccharidiques.
A la série d’enzymes « classiques » s’ajoutent des enzymes plus singulières, telle que
l’oligosaccharidyltransférase, qui utilise comme substrat donneur un oligosaccharide lié à une
molécule de dolichol-phosphate, ultérieurement transféré sur une protéine au cours des premières
étapes de la biosynthèse des N-glycannes, ou encore telles que les mannosyltransférases et
glucosyltransférases, utilisant du mannose ou du glucose liés également au dolichol-phophate
(Kornfeld and Kornfeld, 1985). Il existe aussi des glycosyltransférases d’origine procaryote qui
transfèrent des sucres liés à l’undecaprényl-phosphate (Shaper and Shaper, 1992). Enfin,
certaines glycosyltransférases sont en mesure de lier des monosaccharides à des résidus Thr ou
Ser, la protéine servant alors directement de substrat accepteur. C’est le cas pour certaines N-
acétylgalactosaminyltransférases, qui initient la synthèse des O-glycannes, ainsi que pour des
enzymes plus récemment caractérisées, que sont les O-fucosyltransférases ou les O-N-
acétylglucosaminyltransférases (Van den Steen et al., 1998).
Pour les glycoprotéines, deux grandes familles de glycannes ont donc été définies selon la
nature de la liaison covalente établie entre le glycanne et la partie peptidique : les N- et les O-
glycannes. Les protéines N-glycosylées portent un glycanne sur l’atome d’azote de la chaîne
latérale d’une asparagine comprise dans un motif consensus de type N-X-S/T (X pouvant être
n’importe quel résidu autre que la proline). Les protéines O-glycosylées portent un
monosaccharide ou un glycanne fixé par l’intermédiaire de l’atome d’oxygène de la chaîne
latérale d’une sérine ou d’une thréonine. Ces deux types de glycosylation peuvent se retrouver
simultanément sur une même protéine (comme par exemple, les récepteurs de la famille Notch).
α1,2 ; α1,3 ; α1,4 ; α1,6 ; β1,4
GDP-ManMannoseMannosyltransférase
α1,3 ; β1,4 UDP-GalNacN-acétylgalactosamineN-acétylgalactosaminyltransférase
β1,2 ; β1,4 ; β1,6 UDP-GlcNacN-acétylglucosamineN-acétylglucosaminyltransférase
β1,3 ; β1,4 ; α1,3UDP-GalGalactoseGalactosyltransférase
α1,2 ; α1,3 ; α1,4 ; α1,6GDP-FucFucoseFucosyltransférase
α2,3 ; α2,6 ; α2,8CMP-NeuAcAcide N-acétylneuraminiqueSialyltransférase
Liaisons formées entre sucre donneur et sucre accepteur
Substrat donneur
Sucre transféréGroupe d'enzymes

80
5.1.1. La N-glycosylation des protéines
Les N-glycoprotéines représentent la classe la plus abondante de glycoprotéines (Vijay, 1998). La
N-glycosylation des protéines chez les Eucaryotes est un processus métabolique hautement
conservé. En effet, on retrouve des N-glycannes aussi bien chez les organismes unicellulaires
(Levures, Trypanosomes, etc.) que chez les organismes pluricellulaires (Plantes, Insectes,
Mammifères, etc.) (Spiro, 2002). Pour tous les Eucaryotes, cette modification post-
traductionnelle est indispensable au bon fonctionnement des cellules (Kukuruzinska and Lennon-
Hopkins, 1999). Ainsi, un défaut dans la machinerie de la N-glycosylation peut conduire, chez
l’Homme, à un syndrome de type CDG (Congenital Disorder of Glycosylation) (Freeze and Aebi,
2005; Lowe and Marth, 2003).
Figure 26 : Synthèse des N-glycoprotéines
La synthèse des N-glycoprotéines (Figure 26) est un processus très complexe qui se
déroule dans trois compartiments cellulaires : le cytoplasme, le réticulum endoplasmique rugueux
(RER) et le Golgi. Le nombre et la taille des branchements varient en fonction de la protéine, du
tissu et de l’espèce considérée (Kornfeld and Kornfeld, 1985).
La translocation co-traductionnelle des protéines dans la lumière du RER constitue la première
étape du transport intracellulaire des protéines N- glycosylées (Roth, 2002). A leur arrivée dans le
réticulum, les protéines subissent de nombreuses modifications, dont la glycosylation. Une fois
leur maturation achevée, au sein de l’appareil de Golgi, les glycoprotéines non résidentes de ce
compartiment sont triées puis prises en charge par des vésicules golgiennes afin d’assurer leur
transport vers leurs zones d’activité (Roth, 2002).
Membrane du RE
Dolichol-phosphate N-acetylglucosamine Mannose Glucose Fucose Galactose Acide sialique Chaîne polypeptidique Protéine
Membrane de l’appareil de Golgi

81
L’édification d’une structure polymannosylée, préalable à la N-glycosylation des
protéines, débute sur la face cytosolique du RER par l’addition séquentielle de deux résidus N-
acétylglucosamine et de cinq résidus mannose sur le dolichol-phosphate (1), un lipide
membranaire. Par un mécanisme de flip-flop (Vishwakarma and Menon, 2005), ce complexe est
déplacé vers la face luminale de la membrane du RER (2), où quatre mannoses et trois autres
glucoses sont ajoutés (3). L’oligosaccharide de 14 résidus (Glc3Man9GlcNAc2), structure
consensuelle à tous les N-glycannes, est ensuite transféré « en bloc » du dolichol-phosphate vers
un résidu asparagine d’une chaîne polypeptidique naissante (4) via une liaison β-N-glycosidique,
si et seulement si ce dernier est inclus dans la séquence consensuelle Asn-X-Ser/Thr (X désignant
tout acide aminé excepté la proline) (Pan and Elbein, 1990). Cette réaction est catalysée par un
complexe enzymatique membranaire appelé oligosaccharyltransférase (OST) (Silberstein and
Gilmore, 1996). Il a été estimé que 90% de ces asparagines consensuelles sont glycosylées
(Gavel and von Heijne, 1990). Alors que la protéine acquiert sa structure native, trois glucoses et
un mannose sont éliminés par différentes enzymes de déglycosylation (5, 6, 7). La glycoprotéine,
est ensuite adressée, via des vésicules de transport, vers le Golgi (8) afin d’y poursuivre sa
maturation (dans le sens cis-trans). Après quelques démannosylations et transfert de trois résidus
N-acétylglucosamine (9, 10, 11, 12), la synthèse du glycanne est achevée par divers ajouts de
molécules de fucose à sa base, de galactose et d’acides sialiques à sa périphérie (13, 14).
Au cours de leur maturation golgienne, les N-glycannes évoluent structuralement en 3
groupes (Figure 27) (Geisow, 1991). Les N-glycannes riches en mannoses ou
oligomannosidiques, qui pourront éventuellement se diversifier dans les différents compartiments
golgiens et acquérir une structure hybride, car conservant des branches mannosidiques et
présentant de nouvelles substitutions par des GlcNAc. Enfin, le dernier niveau de diversification
aboutit à des glycannes complexes, ne possédant plus aucune branche mannosidique. Quelle que
soit leur structure, ces glycannes possèdent un noyau pentasaccharidique commun GlcNAc2Man3.
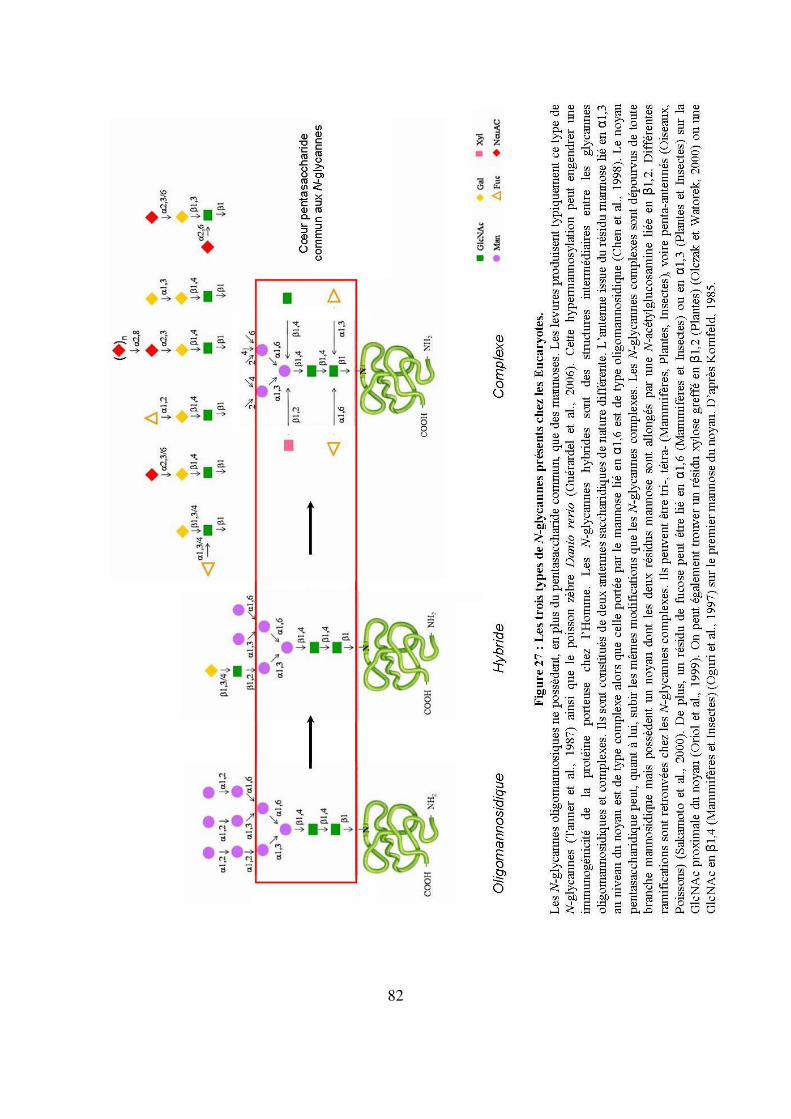
82
Figure 27 : Les trois types de N-glycannes présents chez les Eucaryotes.

83
5.1.2. La O-glycosylation des protéines
Contrairement à la N-glycosylation qui débute par l’assemblage d’un oligosaccharide dans
le RER qui sera ensuite transféré en bloc sur la protéine naissante, la O-glycosylation se fait par
additions successives de monosaccharides sur un acide aminé hydroxylé d’une protéine
(Brockhausen et al., 1995). Elle est initiée dans la lumière du RER et se poursuit dans les
compartiments de l’appareil de Golgi. Le lieu de synthèse des O-glycannes reste à ce jour très
controversé, se situant dans une zone allant du RE au Golgi proximal, en passant par le
compartiment intermédiaire de ces 2 organites, appelé ERGIC, « ER-Golgi compartment » (Roth
et al., 1994). Dans une grande majorité des cas, l’acide aminé impliqué est un résidu sérine ou
thréonine, mais on rencontre également l’hydroxyproline et l’hydroxylysine (dans le cas du
collagène) (Yamauchi et al., 1982) ou la tyrosine (dans le cas de la glycogénine) (Kao et al.,
1999). Les principaux monosaccharides constituant les O-glycannes sont la N-
acétylgalactosamine, la N-acétylglucosamine, le galactose, le glucose, le xylose, le mannose, le
fucose et l’acide sialique. Alors que pour les N-glycoprotéines, la liaison entre glycanne et
squelette peptidique est unique (liaison d’une asparagine à un GlcNAc), il existe différents types
de liaisons pour les O-glycoprotéines en fonction de la nature du glycanne et de l’acide aminé
engagé (Figure 28).
Figure 28 : Représentation schématique des principaux monosaccharides O-liés sur des résidus Sérine et
Thréonine.
Les signaux qui déterminent quel acide aminé sera O-glycosylé ne sont pas encore bien connus.
Le site de O-glycosylation ne fait pas forcément intervenir un acide aminé compris dans une
séquence peptidique consensus, sauf dans les cas de la O-glucosylation et de la O-fucosylation.
Dans le domaine de la glycobiologie, la O-glycosylation est un thème de recherche relativement
récent (fin des années 70) par rapport à l’intérêt porté à la N-glycosylation (début des années 60),
les données disponibles sur les O-glycannes sont donc beaucoup moins conséquentes. Les
structures O-glycanniques sont extrêmement variées et présentent de nombreuses différences,

84
aussi bien au niveau de leurs synthèses qu’au niveau de leurs rôles biologiques. On peut les
classer en différentes « famille » :
• La O-glycosylation de type mucine est la plus répandue. Elle est initiée par l’ajout d’un
GalNac (par des ppGaNtases) sur les résidus Ser/Thr de protéines transmembranaires ou
sécrétées. Le résidu GalNAc peut rester sous la forme de monosaccharide ou être allongé
par d’autres enzymes (ajout de GlcNAc et/ou de Gal, de Fuc et de NeuAc). Un glycanne
de type mucine peut contenir jusqu’à 15 sucres. Les mucines sont des macromolécules
recouvrant les cellules en contact avec le milieu extérieur et protégeant les épithéliums
contre toutes sortes d’agressions d’origine endogène ou exogène (sucs digestifs, micro-
organismes, polluants, toxines…). Elles représentent une grande famille de protéines
fortement glycosylées. L'"enrobage sucré" dense des mucines les rend plus résistantes à la
protéolyse assurant ainsi la protection de nos épitheliums digestifs ou respiratoires. Les
O-glycannes de type mucine se retrouvent majoritairement sur des protéines de
l’épithélium, de l’endothélium vasculaire et à la surface des globules rouges (tel que
CD43 (leukosialin), CD45RA ; CD34, glyCAM-1, MadCAM-1 qui sont des ligands de la L-
selectine exprimée sur les leucocytes…). Ces stuctures sucrées définissent différents types
d’antigène : elles constituent les déterminants des groupes sanguins (A, B, O) et
tissulaires (Lewis). Le dissaccharide Galβ1,3GalNAc appelée antigène T et le simple
résidu GalNAc, appelé antigène Tn sont présents à la surface de nombreuses cellules (la
« Peanut Agglutinin » pour PNA, serait connue pour se lier à l’antigène T). Cette 0-
glycosylation permet également la synthèse des ligands des galectines.
• la O-mannosylation, qu’on retrouve sur des glycoprotéines du cerveau, du système
nerveux et des muscles squelettiques. La plupart des structures 0-mannosylées comportent
le « core glycanne » Galβ1,4GlcNAcβ1,2Man1-O-S/T (Haltiwanger and Lowe, 2004)
• la O-N-acétylglucosaminylation, qui est catalyséee par l’enzyme OGT (O-GlcNAc
Transférase). Cette modification, par un seul sucre, est fréquemment retrouvée sur des
protéines telles que l’ARN polymérase II, les protéines des pores nucléaires, les facteurs
de transcription (SP1- specificity protein-1 et SRF –Serum response Factor), le récepteur
aux oestrogènes, les protéines de proto-oncogènes (c-myc, c-fos, c-jun…), les protéines
« suppresseurs de tumeur » (p53), et les protéines du cytosquelette (Tau, Vinculine,
Ankyrine, Clathrine…). Une délétion du gène OGT provoque l’apoptose des cellules ES
murines (Shafi et al., 2000).

85
• la O-glucosylation qui a lieu sur les domaines EGF (Epidermal growth factor) et EGF-like
que comportent de nombreuses protéines de surface (Harris and Spellman, 1993).
• la O-fucosylation affecte également les domaines EGF mais aussi les domaines TSR
(Thrombospondin type I repeat) et dépend respectivement de deux enzymes. Celle
responsable de la O-fucosylation des EGF est Pofut1 (Protein Ofucosyltransferase 1)
(Wang et al., 1996a) alors que celle greffant le fucose sur les TSR est Pofut2 (Protein O-
fucosyltransferase 2) (Luo et al., 2006). Notch et ses ligands seraient régulés par o-
fucosylation.
• et enfin la O-Glycosylation de type Glycosaminoglycanne (GAG) que je développerai
plus particulièrement dans ce chapitre.
5.1.3. La O-Glycosylation de type Glycosaminoglycanne (GAG)
Au cours de ma thèse, je me suis particulièrement intéressée aux GAGs, auxquels je vais donc
dédier un paragraphe indépendant afin d’être le plus exhaustif possible.
Les GAGs sont de longues chaînes polysaccharidiques non ramifiées d’unités
disaccharidiques répétées, généralement un hexosamine (glucosamine GlcN ou N-
acétylgalactosamine GalNAc) suivi d’un acide hexuronique (acide D-glucuronique GlcA ou
acide L-iduronique IdoA) d’où la dénomination actuelle : glyco pour l’acide hexuronique, amino
pour l’hexosamine, et glycanne pour signifier qu’il s’agit de chaînes glucidiques en général
greffées sur une protéine. Les GAGs sont généralement sulfatés de façon variable. Ces longues
chaînes polysaccharidiques sont liées par liaison covalente à une protéine centrale (ou « core
protein ») par l’intermédiaire d’un résidu sérine, formant ainsi les protéoglycannes, qui
constituent une famille hétérogène de macromolécules (Figure 29). Il est difficile d’établir une
classification des protéoglycannes. On distingue toutefois selon leur localisation cellulaire des
protéoglycannes intracellulaires, membranaires et extracellulaires.
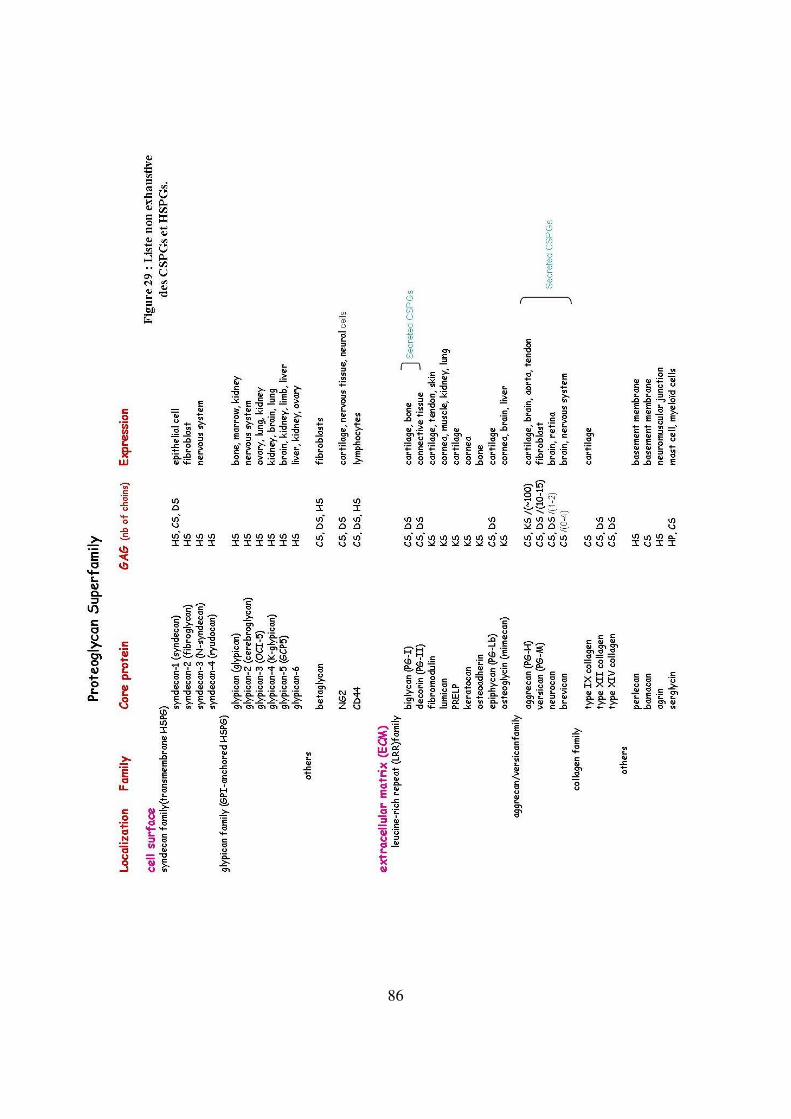
86
Figure 29 : Liste non exhaustive des CSPGs et HSPGs

87
Les protéoglycannes peuvent varier considérablement par leur composition protéique, leur
taille moléculaire ainsi que le nombre et le type de chaînes GAGs par molécule. On ne connaît
pas encore le mécanisme qui dicte à la cellule la glycosylation d’une protéine donnée par une
chaîne de GAGs. Ceci est d’autant plus vrai que certaines protéines existent temporairement, soit
sous forme de protéoglycannes, soit sous forme de protéines sans chaînes GAGs, et sont appelées
ainsi des « protéoglycannes à temps partiel ». Il semble que toutes les cellules diploïdes
possèdent un système de synthèse de GAGs, mais on ignore encore si la synthèse de composants
polysaccharidiques constitue pour la cellule une condition nécessaire à sa survie bien que chez le
nématode, Caernorhabditis elegans, il ait été récemment démontré que les CSPGs (pour
Chondroitin Sulfate Proteoglycans) étaient nécessaires à la survie et à la division cellulaire
(Hwang et al., 2003; Mizuguchi et al., 2003).
Bien qu’il existe toujours un motif disaccharidique de base, la longueur et la composition
des chaînes de GAGs sont extrêmement variables, tout comme l’arrangement spatial des
groupements latéraux hydroxyle, sulfate et carboxyle, le long des chaînes. Les chaînes de GAGs
étant parfois fortement sulfatées, les protéoglycannes ont donc généralement une assez forte
charge anionique. Leur polarité, trait physicochimique le plus marquant, les fait implicitement
participer à certaines fonctions biologiques comme l’hydratation des tissus, la fixation des cations
ou le rôle de barrière à filtration ionique. Les chaînes polysacharidiques ont tendance à adopter
des conformations très étirées, repliées au hasard qui occupent un volume considérable par
rapport à leur masse.
a. Les différentes sous-familles de GAGs
La nature des sucres formant le motif disaccharidique tout comme la nature et le degré de
sulfatation permettent de classer les différents GAGs en sous familles (Perrimon and Bernfield,
2001). Figure 30 :
- les galactosaminoglycannes comprennent les Chondroïtines Sulfate (CS) et les
Dermatans Sulfates (DS), caractérisés par une Galactosamine N-Acétylée (GalNAc) attachée à
GlcA (pour CS) ou à GlcA/IdoA (pour DS).
o Le DS est en fait un isomère du CS où le résidu D-glucuronate est remplacé par
un épimère, le L-iduronate. La quantité d’acide iduronique dans la chaîne de DS peut varier de 0
à 90%. De plus, l’épimérisation se fait en « bloc », ce qui donne des régions riches en L-iduronate
et d’autres riches en D-glucuronate dans la même chaîne de DS. Le DS est associé
principalement à la peau et aux tissus non cartillagineux.

88
o Les chondrocytes sont les cellules spécialisées dans la synthèse des CS. Les
groupements sulfates se trouvent en général sur les carbones C-4 et C-6 de la galactosamine et
sur les carbones C-2 des iduronates, pour les DS. Les C4S et C6S sont liés aux protéoglycannes
du cartilage, du matériel squelettique, de la peau et de la cornée. Ces GAGs assurent au cartilage
une fonction protectrice. Alors que le C4S est un composant important pour les processus de
calcification, le C6S serait responsable de l’intégrité des surfaces articulaires.
Figure 30 : La famille des glycosaminoglycannes
- les glucosaminoglycannes, qui sont eux-même divisés en trois catégories :
o L’acide hyaluronique ou hyaluronane (HA) est le seul GAG non fixé à une
protéine porteuse et non sulfaté (Iozzo, 1998). Il possède une structure homogène d’unités
répétées GlcA-GlcNAc et est beaucoup plus long que les autres GAGs. L’HA est un composant
très représenté dans l’organisme (tissu conjonctif, peau, cartilage, fluide synovial). Du fait de son
haut degré d’hydratation, l’HA est probablement responsable de la grande teneur en eau de
certains tissus. Ce polysaccharide gigantesque (masse moléculaire moyenne voisine de 1000
kDa…) possède une viscosité intrinsèque très élevée, assurant l’hydratation et l’assemblage des
différents éléments du tissu conjonctif par formation de complexes supramoléculaires et donnant
au liquide synovial sa viscosité.
o Le Keratane Sulfate (KS) n’est pas considéré comme un GAG à part entière car
il est dépourvu d’acides hexuroniques mais étant combiné à une protéine porteuse de la même
manière que les proteoglycannes, il est communément classé parmi les GAGs. Il est constitué
d’une alternance de galactose et de N-acétylglucosamine. Les groupes sulfates se situent
généralement en C-6 de la glucosamine et occasionnellement en C-6 du galactose. Le KS est
représenté dans le cartilage, le matériel squelettique et la cornée.
Glycosaminoglycannes
Galactosaminoglycannes Glucosaminoglycannes
CS DS
HS/HP
KS
HA
Glycosaminoglycannes
Galactosaminoglycannes Glucosaminoglycannes
CS DS
HS/HP
KS
HA

89
o L’Héparane Sulfate (HS) et l’Héparine (HP, notamment produite dans les
granulations des mastocytes et basophiles). Ils font partie d’une même sous-famille. Ce sont les
formes les plus complexes et les plus sulfatées de la famille des GAGs dans le cas de l’héparine.
La distribution des groupements sulfates le long des chaînes osidiques va déterminer si le produit
final est classé parmi les héparines (très fortement sulfatées – 80% des glucosamines) ou les HS
(moins sulfatées- 40 à 50% des glucosamines). Ils ont été énormément étudiés à cause de leur
implication dans de multiples fonctions biologiques proteoglycans (Gallagher et al., 1986;
Salmivirta et al., 1996), les HS étant retrouvés dans le milieu extracellulaire de nombreux tissus,
et de l’application thérapeutique majeure de l’HP comme anticoagulant (Bourin and Lindahl,
1993).
b. La biosynthèse des GAGs
La biosynthèse des GAGs (Lindahl et al., 1998; Perrimon and Bernfield, 2000) est un
processus multi-étapes qui a lieu dans l’appareil de Golgi. La première étape implique
l’attachement d’un tétrasaccharidique unique, commun à tous les GAGs, au niveau du
groupement hydroxyle d’une sérine d’une « core protein » (Figure 31). Le tétrasaccharide
toujours rencontré est GlcAβ(1,3)-Galβ(1,3)-Galβ(1,4)-Xylβ1-0-Ser. Ce processus est catalysé
par quatre enzymes qui ajoutent les monosaccharides de manière séquentielle à l’extrémité non
réductrice de la chaîne en croissance.
Il y a ensuite, élongation de la chaîne par ajout d’unités saccharidiques puis modification
de la chaîne polysaccharidique par N-déacétylation, N et O-sulfatations, épimérisation… faisant
intervenir un large répertoire d’enzymes spécifiques : N-desacétylase/N-sulfotransférase (NDST),
C5-épimérase, 6-O-sulfotransferase, 3-O-sulfotransferase…. Les enzymes ne modifient pas
toutes les unités saccharidiques de la chaîne ce qui crée un degré d’hétérogénéité structurale. Les
GAGs sont modifiés variablement en fonction des conditions cellulaires (Sugahara and Kitagawa,
2002). Le degré de sulfatation semble lui être déterminé par le type de cellules et leur état
physiologique (Tumova et al., 2000). La variabilité de ces modifications le long du polymère crée
la diversité structurale et fonctionnelle des chaînes de GAGs (Zimmermann and David, 1999). Il
est à noter que dans le cas des HS, la réaction d’initiation, catalysée par la xylosyltranférase, a
lieu au niveau d’une région d’ancrage spécifique définie par deux ou trois séquences consécutives
de résidus « Serine-Glycine » (Esko and Zhang, 1996) encadrées par des résidus hydrophobes et
acides. Le nombre de sites d’attachement de chaînes varie en fonction de la nature de la « core
protein », mais typiquement, il y en a entre deux et quatre pour HS et beaucoup plus pour HP.
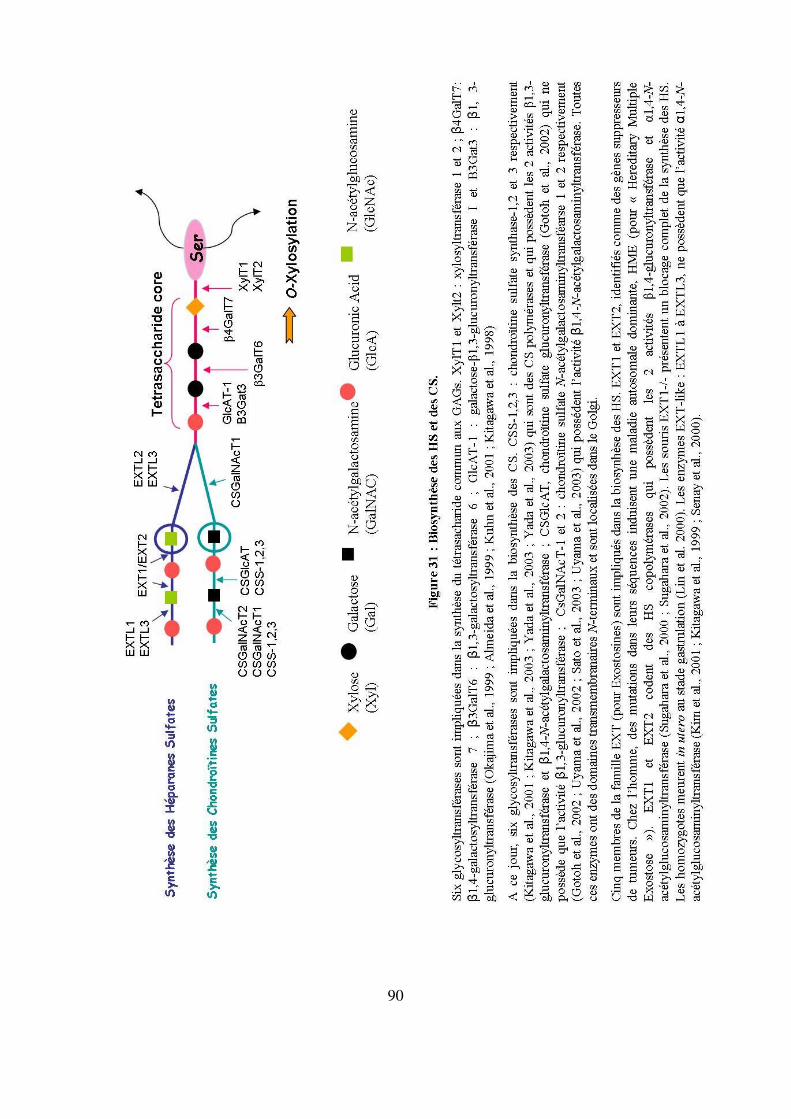
90
Figure 31 : Biosynthèse des HS et des CS.

91
C’est le premier hexosamine greffé sur l’extrémité non-réductrice du tétrasaccharide
d’attachement (dans ce cas, l’acide glucuronique) qui va déterminer la nature de la chaîne de
GAG. Si c’est un N-acétylglucosamine (GlcNAc), il y aura alors biosynthèse de
glucosaminoglycanne (HS ou HP) et si c’est un N-acétylgalactosamine (GalNAc), il y aura alors
biosynthèse de galactosaminoglycanne (CS ou DS). Il a été suggéré que les motifs de la séquence
peptidique proche du site d’attachement du tétrasaccharide jouaient un rôle de signal pour
l’addition de l’un ou l’autre des monosaccharides (Zhang et al., 1995). De plus la modification du
tétrasaccharide commun aux GAG par phosphorylation et sulfatation pourrait avoir un rôle
important dans l’assemblage des ces chaînes. Effectivement, la phosphorylation du résidu xylose
est retrouvée pour les chaînes HS et CS alors que la sulfatation du deuxième galactose n’est
visible que pour les chaînes CS. Cette sulfatation semble donc dicter à la cellule la synthèse de
chaînes CS spécifiquement.
Après le premier saccharide attaché, un transfert alternatif de GlcA et de GlcNAc (pour
HS) ou GalNAc (pour CS), à partir de leurs nucléotides UDP-sucres correspondants, forme le
reste de la chaîne GAG. Approximativement, 20 à 300 monosaccharides sont additionnés au
polysaccharide linéaire avant la fin de synthèse. Cette hétérogénéité de longueur crée un premier
niveau de complexité. Une fois la chaîne formée, les disaccharides subissent une série de
modifications. On estime qu’environ 10% des disaccharides constituants les chaînes HS subissent
des modifications contre 80% pour les chaînes héparines (Salmivirta et al., 1996).
Dans le cas des glucosaminoglycannes, la modification du polymère est initiée par une N-
désacétylation/N-sulfatation des GlcNAc par une enzyme la N-désacétylase/N-sulfotransférase.
Les étapes suivantes ont lieu de façon séquentielle sur les résidus adjacents des N-
sulfoglucosamines (GlcNS) qui sont reconnus par la C5- épimérase qui va alors catalyser la
transformation de certains acides D-glucuroniques (GlcA) en acides L-iduroniques (IduA)
(Salmivirta et al., 1996). Ensuite a lieu une O-sulfatation des acides iduroniques en position C-2
par une 2-O-sulfotranférase. Cette sulfatation est suivie par l’action d’une glucosamine-6-O-
sulfotransférase, qui va alors transfèrer un groupe O-sulfate sur la position C-6 de GlcNac ou du
GlcNS. Des sulfatations peuvent également avoir lieu en position C3 de certains GlcNS6S
(Tumova et al., 2000).
Dans le cas des galactosaminoglycannes, des groupements ester-sulfates peuvent être
greffés sur les galactosamines. La place de ces groupements détermine la nature des
chondroïtines sulfates. Si ce dernier est fixé à l’alcool porté par le C-4 du GalNAc, il s’agit de

92
chondroïtine-4-sulfate (C4S) ; s’il est fixé sur le C-6, il s’agit de chondroïtine-6-sulfate (C6S)
(Tableau 2).
CSA = [-4)GlcA(β1-3)GalNAc4S(β1-]n
CSB = [-4)IdoA(β1-3)GalNAc4S(β1-]n (ou DS)
CSC = [-4)GlcA(β1-3)GalNAc6S(β1-]n
CSD = [-4)GlcA2S(β1-3)GalNAc6S(β1-]n
CSE = [-4)GlcA(β1-3)GalNAc4S,6S(β1-]n
Tableau 2 : Les différents motifs des CS
La fixation des sulfates n’est pas d’une régularité absolue, certains résidus des chaînes HS et CS
peuvent donc en être dépourvus. On a donc une alternance de domaines hautement sulfatés de
séquences variables (appelés domaines S ou NS) avec des domaines peu ou pas sulfatés de
séquences d’unités saccharidiques de base (appelés domaines NA). Toutes ces réactions de
sulfatation font intervenir des sulfotransférases qui utilisent le 3’-phosphoadénosine-5’-
phosphosulfate (PAPS) comme donneur de groupements sulfates. D’une manière générale, les
chaînes CS sont plus sulfatées donc plus chargées négativement que les chaînes HS. De plus, les
chaînes HS (co-polymères) possèdent des résidus IduA. Elles présentent donc, contrairement aux
chaînes CS (polymères simples), une très grande flexibilité conformationnelle qui leur permet de
s’auto-associer et d’interagir avec de multiples molécules extracellulaires et cellulaires. La
structure dans l’espace des CS est celle d’une simple hélice, les groupements sulfates chargés
étant dirigés vers l’extérieur de l’hélice. Il est également à noter qu’il existe des enzymes de
dégradation comme les héparitinases et les chondroïtinases qui éliminent certains groupements
sulfates et contribuent de ce fait à la formation de domaines plus ou moins sulfatés adoptant ainsi
des structures 3D différentes.
5.2. Les « sucres » et leur implication dans le système immunitaire
5.2.1. La glycosylation des Ig
Toutes les IgG possèdent un site de N-glycosylation situé sur l’asparagine 297 du
domaine CH2 de la chaîne lourde d’Ig et 20% d’entre elles possèdent un site potentiel de N-
glycosylation sur la région variable. Nous nous contenterons ici d’évoquer la glycosylation de la
CS4
CS6
CS4/6

93
région constante. L’analyse des différentes structures oligosaccharidiques rencontrées sur l’IgG
humaine (Jefferis, 2005; Raju et al., 2001) nous révèle la présence d’une structure bi-antennée de
type complexe GlcNAc2Man3GlcNAc2 présentant un haut degré d’hétérogénéité quant à la
présence de certains sucres terminaux comme le galactose, fucose, l’acide sialique et un résidu
GlcNAc intercalaire (Figure 32). Ces différentes glycoformes rencontrées constituent une
microhétérogénéité tout en sachant que la présence de ce N-glycanne est déterminante, et capable
de fortement moduler l’activité de l’anticorps notamment au niveau de fonctions effectrices
comme l’activation du complément et l’ADCC.
Figure 32 : Structure et fonction d’une IgG. Schéma de Carter et al., 2006.
L’absence anormale de structure N-glycosylée au niveau de la région Fc des Ig est ainsi capable
d’altérer sa conformation et de perturber les liaisons Fc / facteur C1q du complément ou Fc /
récepteurs FcγR (FcγRI ou CD64, FcγRIIa ou CD32a ou FcγRIIIa ou CD16a) exprimés à la
surface des cellules cytotoxiques communément appelées « tueuses » (NK, monocytes,
macrophages).

94
L’ADCC est une réaction de cytotoxicité dans laquelle des cellules cytotoxiques porteuses de
récepteurs FcγR reconnaissent les cellules cibles recouvertes d’un anticorps spécifique. La lyse
est induite seulement si l’oligosaccharide greffé sur l’asparagine 297 de l’anticorps humain est
correctement glycosylé. De la même manière, il a été montré que la présence de sucres sur
l’Asparagine 297 était indispensable à la fixation au complément, le complément étant constitué
d’un groupe de protéines sériques (C1q, C1r et C1s) impliqué dans l’inflammation, l’activation
de cellules phagocytaires et la lyse cellulaire (on parle également d’activité CDC pour
Complement-Dependent Cytotoxicity).
Un grand nombre d’études ont montré qu’un anticorps non glycosylé aurait une affinité de liaison
aux récepteurs FcγR très fortement diminuée (Lund et al., 1995; Mimura et al., 2001; Radaev and
Sun, 2001; Watt et al., 2003). Par contre, l’absence de fucose à la base du N-glycanne du Fc des
IgG (ARNi ou Fut8-/-) améliore nettement l’affinité pour le FcγRIIIa et augmente fortement
l’ADCC (sans modifier l’activation du complément) (Niwa et al., 2005; Shields et al., 2002;
Shinkawa et al., 2003; Yamane-Ohnuki et al., 2004). L’addition d’un résidu N-acétylglucosamine
(GlcNAc) intercalaire, catalysée par la Gnt-III, qui réduirait le niveau de fucosylation,
augmenterait également la fixation au récepteur FcγRIIIa et donc la lyse par ADCC (Umana et
al., 1999). Ces observations prennent de l’importance à la lumière de travaux montrant qu’un
variant du récepteur FcγRIIIa, FcγRIIIa-158V qui lie fortement l’IgG1, est associé à une
meilleure réponse à l’anticorps thérapeutique Rituximab (IgG1 chimérique anti-CD20 utilisée
dans le traitement des lymphomes B) comparé à des patients qui ont l’allotype FcγRIIIa-158F
ayant une faible affinité de liaison pour l’IgG1 (Cartron et al., 2002). D’autres modifications de la
structure du pentasaccharide greffé sur l’Asn297 peuvent avoir des effets sur la lyse. Par exemple,
une diminution de la galactosylation entraînerait une diminution de la CDC (Boyd et al., 1995;
Hodoniczky et al., 2005). Par contre, un fort degré de sialylation diminuerait l’ADCC du fait
d’une faible affinité de liaison pour le récepteur FcγRIIIa sur les cellules NK (Scallon et al.,
2007). En parallèle, il a également été démontré que la présence d’acide sialique sur ce N-
glycanne du Fc des IgG donnait des propriétés anti-inflammatoires à l’Ig, dépendantes de
l’expression accrue de FcγRIIB à la surface des cellules effectrices, et qu’après un challenge
antigénique, les IgG spécifiques de l’antigène pouvaient passer d’un état sialylé à un état
hyposialylé afin de générer une réponse adaptative pro-inflammatoire (Kaneko et al., 2006).
L’ingénierie des anticorps monoclonaux, désormais utilisés en clinique humaine, peut ainsi
permettre le développement d’une nouvelle génération d’anticorps, dont les propriétés
fonctionnelles ont été optimisées, permettant des avancées significatives dans le traitement de

95
pathologies pour lesquelles l’arsenal thérapeutique est limité. L’ingénierie du profil de
glycosylation de la région Fc des IgG1 humaines (l’isotype utilisé dans la très grande majorité
des anticorps monoclonaux commercialisés) est donc une stratégie importante, utilisée pour
optimiser les fonctions effectrices de ces anticorps.
5.2.2. Les galectines, les voies Notch et Wnt : importance de la
glycosylation.
a. Les galectines
Les galectines appartiennent à une famille de lectines très conservée à travers les espèces, qui
comprend 15 membres chez les mammifères. Ce sont des lectines de type S (thiol-dépendantes)
capables de lier les galactoses ou les N-acétylgalactosamines de nombreuses protéines N ou O-
glycosylées intra- ou extra-cellulaires. Ces lectines, molécules solubles, ne requièrent pas la
présence de calcium pour leur activité de liaison. Les différentes galectines ont un profil
d’expression spécifique, hautement régulé (les galectines 1, 3, 7 et 9 sont connues comme
s’exprimant dans les cellules de l’immunité). Ces protéines peuvent être exprimées dans le noyau
et dans le cytoplasme. Certaines sont sécrétées et interagissent avec de nombreuses molécules
glycosylées (appelées « contre-récepteurs ») exprimées à la surface des cellules (Hughes, 1999).
Les galectines peuvent interagir avec des intégrines, des molecules de la matrice extra cellulaire
(telles que la laminine et la fibronectine), le ganglioside GM1 et avec des molécules exprimées
par les lymphocytes comme CD3, CD7, CD43 et CD45… (Fukumori et al., 2003; Gu et al., 1994;
Hadari et al., 2000; Moiseeva et al., 2003; Pace et al., 1999; Sturm et al., 2004). La famille des
galectines peut se diviser en 3 sous-groupes selon le nombre de domaines de reconnaissance aux
hydrates de carbone qu’elles possèdent. Ces CRD pour Carbohydrate Recognition Domain, sont
longs d’environ 130 acides aminés et sont donc responsables de la liaison aux résidus β-
galactosides. Les galectines prototypes (galectines 1, 2, 5, 7, 10, 11, 13, 14 et 15) contiennent un
CRD mais existent soit sous forme monomérique, soit sous forme homodimérique. Les galectines
chimériques ne sont représentées que par un seul membre, la galectine 3 qui est composée de
courtes répétitions proline-glycine d’environ 120 acides aminés, fusionnéees au CRD. Les
galectines tandem (galectines 4, 6, 8, 9 et 12) contiennent 2 CRD connectés par l’intermédiaire
d’un « linker » d’environ 70 acides aminés.

96
Des rôles variés ont été démontrés pour les galectines dans l’induction ou la protection vis-à-vis
de l’apoptose : la galectine 3 serait anti-apoptotique alors que les galectines 7 et 12 induiraient
l’apoptose lorsqu’elles sont surexprimées (Hoyer et al., 2004; Yang and Liu, 2003). Les
galectines interviendraient dans des domaines aussi variés que la signalisation cellulaire et la
stabilisation des microdomaines lipidiques, l’adhésion cellulaire, la régulation de la prolifération,
la différenciation cellulaire, la migration et la transformation cellulaire (Honjo et al., 2001;
Lagana et al., 2006; Liu and Rabinovich, 2005). L’expression des galectines est un facteur
pronostic clé dans de nombreux cancers (estomac, foie, colon, reins, sein, prostate, ovaires,
thyroïde et système nerveux central).
La fonction des galectines est dépendante des contre-récepteurs sur lesquels elles peuvent se
fixer. Un certain nombre de glycosyl-transférases ont déjà été identifiées pour leurs capacités à
synthétiser les galactosides reconnus par les galectines. Ainsi dans les lymphocytes T,
l’expression de la core 2 beta-1, 6-N-acetylglucosaminyltransferase (core2GnT) est à l’origine
d’une structure O-liée qui peut-être allongée et présenter de multiples résidus lactosaminiques
permettant la liaison de la galectine-1, qui joue alors un rôle pro-apoptotique pour ces cellules
(Galvan et al., 2000) A l’inverse, les galactosides peuvent être spécifiquement masqués par
l’addition d’acides sialiques. Des sialyltransférases telles que la ST6GalI peuvent ainsi empêcher
la liaison de la galectine-1 à des glycoprotéines des cellules T immatures telles que CD45. Cette
sialylation semble protéger les cellules T les plus immatures du thymus contre une apoptose et
une inhibition de l’activité phosphatase de CD45 induites par la galectine-1 (Amano et al., 2003).
Il a récemment été démontré que les galectines 1 et 3 induisaient la mort des cellules T en se liant
à différents récepteurs membranaires comme CD45 et CD7, respectivement (Stillman et al.,
2006). Mgat5 (beta1,6 N-acetylglucosaminyltransferase V), enzyme de branchement qui catalyse
le transfert d’un GlcNAc en β(1,6) sur le Man lié en α(1,6) du pentasaccharide cœur commun à
tous les N-glycannes, a également été démontrée comme capable d’augmenter le seuil
d’activation du TCR des cellules T matures via l’augmentation d’expression des galactosides liés
par les galectines (Demetriou et al., 2001), de telle sorte que le déficit en Mgat5 s’accompagne de
réponses auto-immunes exacerbées.
Les galectines reconnaissent, via leur CRD, une famille de molécules d’adhésion cellulaire très
étudiée : les intégrines. Il a récemment été démontré que la galectine-1, sécrétée par les cellules
stromales, intéragissait avec des contre-récepteurs glycosylés exprimés à la fois à la surface des
lymphocytes préB et à la surface des cellules stromales de la moelle osseuse, formant ainsi une
synapse entre les préB et les cellules stromales. Cette liaison se ferait par l’intermédiaire des

97
intégrines VLA-4, VLA-5 et α4β7, la synapse permettant la re-localisation des pré-BCR et des
intégrines et induisant ainsi une transduction du signal (Gauthier et al., 2002; Rossi et al., 2006).
b. Les WNTs
Les Wnts constituent une famille de molécules de signalisation impliquées dans le contrôle du
développement. Ce sont des glycoprotéines de poids moléculaires compris entre 39 et 45kDa,
riches en cystéine. Il existe 19 membres de la famille Wnt chez l’homme et seulement 18 chez la
souris (Miller, 2002). Les Wnts stimulent différentes activités cellulaires : prolifération, migration
et polarité cellulaire. Beaucoup de Wnts sont essentiels pendant l’embryogenèse, mais sont aussi
actifs dans la régénération des tissus adultes tels que la peau, les follicules pileux et l’os (Alonso
and Fuchs, 2003; Staal and Clevers, 2003). Chez les vertébrés, les Wnts activent trois voies
différentes, la voie Wnt/ß-caténine, la voie Wnt/Ca2+ et le mécanisme du contrôle de la polarité
cellulaire (Huelsken and Birchmeier, 2001). La voie Wnt/ß-caténine, connue sous le nom de
« voie canonique », intervient dans le contrôle de la prolifération et de la survie cellulaire. La ß-
caténine est une molécule multifonctionnelle agissant en particulier au cours de la signalisation
par Wnt et de l'adhésion cellule-cellule (Hecht and Kemler, 2000; Obara and Lesot, 2004; Willert
and Nusse, 1998). Phosphorylée par la GSK-3β (glycogène synthase kinase), elle-même associée
à la protéine APC (Adenomatous Polyposis Coli), la βcaténine est rapidement dégradée dans les
protéasomes après ubiquitinylation. En présence d’un signal Wnt, la phosphorylation est inhibée,
ce qui permet à la ß-caténine de s’accumuler dans le noyau, de s’associer au complexe de
transcription LEF/TCF et d’activer la transcription de gènes cibles (Figure 33). Lors de
processus tumoraux, la βcaténine va induire la transcription des gènes c-myc, c-jun, Fra et
cycline-D1 (Karim et al., 2004). La voie Wnt/Ca2+ augmente le niveau du calcium intracellulaire
par la voie d’activation de la phospholipase C, aboutissant à l’activation de la PKC et de la
calcineurin. Il y aura alors accumulation de facteurs de transcription tels NF-AT dans le noyau, et
donc transcription de gènes cibles (Wang and Malbon, 2003). La voie Wnt/polarité cellulaire
(« PCP pathway » pour planar cell polarity) est activée par la liaison de Wnt à des petites
GTPases (RhoA, Rac) permettant l’activation de JNK… ayant des effets sur la polarité cellulaire
et sur l’organisation du cytosquelette (Habas et al., 2003).

98
Figure 33 : Voie de signalisation Wnt. Wnt se lie au récepteur Frizzled via un domaine riche en cystéine, CRD, faisant également intervenir les corécepteurs LRP5 et LRP6 (b). Schéma de (Staal and Clevers, 2003).
Wnt1, premier membre de la famille, a été décrit comme un proto-oncogène induisant le
développement de tumeurs mammaires quand on le surexprimait chez la souris (Nusse and
Varmus, 1982). De même, Wnt5a, tout d’abord décrit comme induisant la transformation
cellulaire via son potentiel oncogénique, serait surexprimé dans de nombreuses tumeurs et
faciliterait l’invasion des métastases. A l’opposé, il régulerait négativement la prolifération B et
supprimerait les tumeurs hématopoïétiques via la voie non-canonique Ca2+ (Liang et al., 2003).
Il est également connu que Wnt1 interagirait avec des chaînes HS à la surface cellulaire mais leur
rôle n’est pas encore bien établi (Reichsman et al., 1996). Il a par ailleurs été démontré, chez la
drosophile (D.melanogaster), que les glypicans (HSPGs) liés à la membrane cellulaire par un
ancrage glycosylphosphatidylinositol - ancre GPI) jouaient un rôle dans la distribution tissulaire
de Wingless et Hedgehog au cours du développement (Hacker et al., 2006). Le rôle de la voie
Wnt n’est pas encore bien étudié dans la lignée lymphoïde. On sait que l’absence de TCF-1
aboutit à une détérioration précoce du compartiment T (atrophie thymique avec déficit en

99
thymocytes double négatifs CD4- CD8-) sans répercussion sur la fonction immune ni sur le
nombre de cellules T en périphérie (Schilham et al., 1998). Un blocage complet de la maturation
T est obtenu dans les double KO TCF/LEF (Galceran et al., 1999).
Les WNTs se lient à des récepteurs membranaires au nombre de 10 chez l’homme et 9 chez la
souris, connues sous le nom de Frizzled (Fzds) (Wang et al., 1996b). Peu de données existent sur
la spécificité ou l’affinité WNT/ligand sur leur récepteur/frizzled, mais il doit exister des
redondances puisqu’il y a deux fois plus de membres Wnts que de membres Fzds.
De nombreuses souris KO pour les récepteurs Frizzled (3, 4, 5, 6, 9) ont été réalisées mais les
conséquences de ces déficiences sévères (qui touchent principalement le système nerveux central)
n’ont pas été regardées dans le système hématopoïétique et encore moins dans le développement
B. Ranheim et al., en 2005, ont cherché vainement à reproduire le syndrome humain de Williams-
Beuren (délétion de 1,4 Mb dans la région chromosomique 7 q11.23) chez la souris en réalisant le
KO du gène Frizzled 9. Par contre, ils ont obtenu une lignée de souris présentant un phénotype
sévère touchant le système lymphoïde. Les souris Fzd9-/- présentent une splénomégalie, une
atrophie thymique et une lympho-adénopathie, selon leur âge, avec accumulation de plasmocytes
dans les ganglions. Elles ont un développement B immature altéré (diminution du nombre de pré-
B et de B immatures mais pas d’accumulation pro-B). Par contre, elles présentent un nombre de
B normal dans la rate et les ganglions. Ces données suggèrent donc un rôle pour la signalisation
wnt/fzd dans le développement lymphoïde.
c. Notch
La voie de signalisation Notch intervient dans de nombreux processus développementaux comme
la neurogénèse, la gliogénèse, la myogénèse mais également dans le développement des
lymphocytes (Tanigaki and Honjo, 2007). Chez les mammifères, la famille des récepteurs
transmembranaires Notch est hautement conservée et se compose de 4 membres : Notch 1, 2, 3 et
4. Ils peuvent interagir avec au moins 5 ligands (Serrate/Jagged 1 et 2 ; Delta-like 1, 3 et 4).
La liaison ligand-récepteur, via des motifs glycanniques, induit une série de clivages
protéolytiques au niveau du domaine transmembranaire de Notch aboutissant à la libération de
son domaine intracellulaire (ICD), qui transloqué dans le noyau, ira s’associer avec RBP-Jκ
(aussi appelé CSL/ Rbpsuh). Figure 34. Il y aura alors recrutement des coactivateurs PCAF et
GCN5 (ce seraient des histones acétyltransférases) et initiation de la transcription de gènes cibles
comme Hes1, Hes5 et NF-κB (Artavanis-Tsakonas et al., 1999; Kurooka and Honjo, 2000;
Mumm and Kopan, 2000). MINT, régulateur négatif de Notch, va rentrer en compétition avec
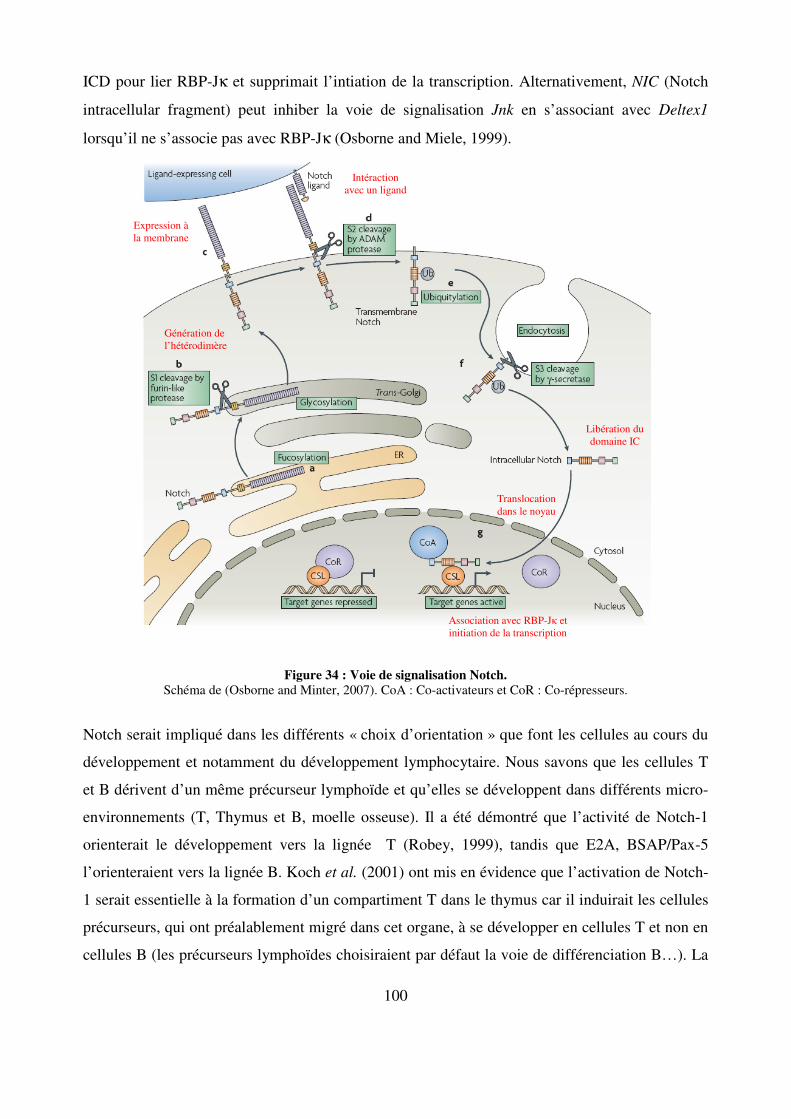
100
ICD pour lier RBP-Jκ et supprimait l’intiation de la transcription. Alternativement, NIC (Notch
intracellular fragment) peut inhiber la voie de signalisation Jnk en s’associant avec Deltex1
lorsqu’il ne s’associe pas avec RBP-Jκ (Osborne and Miele, 1999).
Figure 34 : Voie de signalisation Notch. Schéma de (Osborne and Minter, 2007). CoA : Co-activateurs et CoR : Co-répresseurs.
Notch serait impliqué dans les différents « choix d’orientation » que font les cellules au cours du
développement et notamment du développement lymphocytaire. Nous savons que les cellules T
et B dérivent d’un même précurseur lymphoïde et qu’elles se développent dans différents micro-
environnements (T, Thymus et B, moelle osseuse). Il a été démontré que l’activité de Notch-1
orienterait le développement vers la lignée T (Robey, 1999), tandis que E2A, BSAP/Pax-5
l’orienteraient vers la lignée B. Koch et al. (2001) ont mis en évidence que l’activation de Notch-
1 serait essentielle à la formation d’un compartiment T dans le thymus car il induirait les cellules
précurseurs, qui ont préalablement migré dans cet organe, à se développer en cellules T et non en
cellules B (les précurseurs lymphoïdes choisiraient par défaut la voie de différenciation B…). La
Génération de l’hétérodimère
Expression à la membrane
Intéraction avec un ligand
Libération du domaine IC
Translocation dans le noyau
Association avec RBP-Jκ et initiation de la transcription

101
délétion conditionnelle des gènes Notch1 et RBP-Jκ dans la moelle osseuse de souris adultes,
aboutit à un défaut de développement T et induit la différenciation B (Han et al., 2002; Radtke et
al., 1999; Wilson et al., 2001). De plus, une expression ectopique de Lunatic Fringe (Lfng) dans
le thymus régulerait négativement l’activité de Notch-1, ce qui induirait les précurseurs
lymphoïdes à choisir la voie de maturation B. Cette enzyme golgienne catalyse la liaison d’un
GlcNAc en β(1,3) sur un fucose O-lié. Sa seule activité connue à ce jour s’exerce vis-à-vis des
domaines EGF-like O-fucosylés du récepteur Notch (Moloney et al., 2000) et de ses ligands,
Delta et Jagged (Panin et al., 1997). Il est important d’ajouter que Fringe (et ses homologues,
manic et radical) a un effet variable sur Notch, dépendant des ligands de Notch. Fringe activerait
Notch par l’intermédiaire de Delta et inhiberait Notch par l’intermédiaire de Serrate/Jagged
(Panin et al., 1997). Donc, le fait que Lfng régule négativement Notch-1 dans les précurseurs
lymphoïdes reflète la probable implication de Serrate/Jagged dans la décision T ou B. Des études
structurales ont montré que les di- (GlcNAcβ1,3Fucα1-O-Ser), tri- (Galβ1,4GlcNAcβ1,3Fucα1-
O-Ser) ou tétra-saccharides (NeuAcα2,3Galβ1,4GlcNAcβ1,3 Fucα1-O-Ser) construits sur le
fucose O-lié (Figure 35) au niveau d’une sérine d’un domaine EGF-like moduleraient l’activité
de Notch (Chen et al., 2001a). L’enzyme qui réalise la liaison du fucose est la O-
fucosyltransférase I, est une enzyme golgienne codée par le gène POFUT1 (Wang et al., 2001).
Une souris KO pour POFUT1 meurt à l’état embryonnaire suite à de nombreux
dysfonctionnements dans divers processus développementaux. Le greffage de résidus O-fucoses
sur les domaines EGF de Notch et de ses ligands est donc un évènement incontournable dans le
déclenchement des voies de signalisation, puisque c’est leur allongement (ou non) par les
enzymes de la famille Fringe qui va moduler leurs interactions. Parmi les 6 enzymes de la même
famille (β4GalT-I à VI), seule β4GalT-I (Zhou et al., 1998), une β-1,4-galactosyltransférase, a la
capacité d’allonger le disaccharide pour former le motif Galβ1,4GlcNAcβ1,3Fucα1-O-Ser qui
semble être indispensable à l’activation de la voie de signalisation de Notch (Chen et al., 2001a).
Une souris mutante pour la β4GalT-I, montre des altérations de la somitogénèse (Chen et al.,
2006) bien que cette lignée soit viable. Le phénotype discret de l’adulte mutant est sans doute dû
à une activité redondante des β4GalTs. Pour compléter le motif, une β-galactoside-α-2,3-
sialyltransférase, grefferait un acide sialique en α(2,3) sur le Gal d’un motif Galβ1,3GalNAc
(Harduin-Lepers et al., 2001).

102
Figure 35 : Les différentes structures de glycannes O-fucosylés observées sur la protéine Notch. La maturation du O-fucose est sous le contrôle de 2 glycosyltransférases, Fringe et β4GalT1. Le 4ème sucre greffé est un acide sialique en position α(2,3). D’après Moloney et al., 2000.
L’inactivation de RBP-Jκ dans le lignage B aboutit à une défaut en cellules B MZ avec une
augmentation du nombre de B folliculaires, indiquant que la voie Notch peut intervenir
également dans le « choix d’orientation » MZB versus FOB à partir d’un même précurseur, ici :
la cellule B transitionnelle 2. La délétion conditionnelle de Notch2 dans les cellules
hématopoïétiques cause également la perte du compartiment B MZ (Saito et al., 2003). De plus
l’inactivation du ligand delta-like 1, dans les cellules hématopoïétiques, aboutit également à une
perte du compartiment B, ce qui n’est pas le cas lors de son inactivation B-spécifique. Notch2
exprimé sur les cellules B pourrait interagir avec Delta-like 1 qui serait exprimé sur les cellules
dendritiques afin d’induire la différenciation des B de la zone marginale (Hozumi et al., 2004;
Kuroda et al., 2003). Thomas et al., en 2007, ont montré que la co-stimulation de B folliculaires
avec le ligand de Notch, delta-like 1, stimulait l’activation et la prolifération B médiées par le
BCR et CD40 et augmentait le switch vers IgG1 et la production d’IgG1. La voie Notch serait
donc très importante dans l’apparition et la maturation de la lignée B.
5.2.3. Les différentes fonctions associées aux GAGs
Les protéoglycannes sont des constituants majeurs de la membrane plasmique, du
glycocalyx et de la matrice extracellulaire (MEC) (Vidal y Plana and Karzel, 1980). Ils peuvent
interagir avec un grand nombre de molécules/macromolécules dans différents tissus et peuvent
donc être associés à différentes fonctions, différents processus (Bandtlow and Zimmermann,
2000; Coombe and Kett, 2005).
Fucα1-O-Ser
GlcNAcβ1,3-Fucα1-O-Ser
Galβ1,4-GlcNAcβ1,3-Fucα1-O-Ser
NeuAcα2,3-Galβ1,4-GlcNAcβ1,3-Fucα1-O-Ser
B4galactosyltransférase-1 (β4GalT I)
Β3-N-acétylglucosaminyltransférase (Fringe)
β-galactoside α-2,3-sialyltransférase (ST3Gal II)
Fucα1-O-Ser
GlcNAcβ1,3-Fucα1-O-Ser
Galβ1,4-GlcNAcβ1,3-Fucα1-O-Ser
NeuAcα2,3-Galβ1,4-GlcNAcβ1,3-Fucα1-O-Ser
Fucα1-O-Ser
GlcNAcβ1,3-Fucα1-O-Ser
Galβ1,4-GlcNAcβ1,3-Fucα1-O-Ser
NeuAcα2,3-Galβ1,4-GlcNAcβ1,3-Fucα1-O-Ser
B4galactosyltransférase-1 (β4GalT I)
Β3-N-acétylglucosaminyltransférase (Fringe)
β-galactoside α-2,3-sialyltransférase (ST3Gal II)

103
Dans la MEC, les protéoglycannes forment un gel hydraté dont la taille des pores et la densité des
charges varient selon la composition moléculaire des GAGs. Ils jouent ainsi un rôle dans la
viscosité et la résistance aux compressions des tissus (Schonherr et al., 1995). L’existence d’un
tel gel contrôle la vitesse de diffusion ainsi que la disponibilité des molécules hydrosolubles
(Kolset and Gallagher, 1990). Ils facilitent la migration des cellules au cours de la morphogenèse
(Lin and Perrimon, 2002; Sanes, 2003). Lors des processus tumoraux, ils sont impliqués dans la
modulation des interactions cellulaires (Ma and Geng, 2000; Wegrowski and Maquart, 2004),
dans la formation de plaques amyloïdes comme dans la maladie d'Alzheimer (Gupta-Bansal et
al., 1995; van Horssen et al., 2003). Ils jouent également un rôle dans d’autres maladies sévères
comme les arthropathies (Schwartz and Domowicz, 2002).
Les protéoglycannes de la MEC agiraient sur l’adhésion, la migration et la prolifération cellulaire
et joueraient aussi un rôle important dans le système immunitaire en retenant du fait de leur
charge et en augmentant donc la concentration locale au voisinage de la membrane plasmique de
nombreux facteurs de croissance comme les fibroblast growth factors (FGFs, impliqués dans de
nombreux processus physiologiques incluant la prolifération cellulaire, la différenciation, la
morphogenèse et l’angiogenèse) (Nishimura et al., 2000; Ornitz et al., 1992), ou encore les HGFs
(Hepatocyte Growth Factor), les TGFs (Transforming Growth Factor), les membres de la famille
TNFs (Tumor necrosis Factor) notammant APRIL et GM-CSF en initiant des cascades de
signalisation intracellulaire (Haltiwanger and Lowe, 2004; Ingold et al., 2005). Par exemple, les
FGFs en s’associant fortement avec les HS des protéoglycannes de la membrane cellulaire,
induiraient la dimérisation de leurs récepteurs FGFR initiant ainsi un signal de transduction
(Guerrini et al., 2002; Thompson et al., 1994). Très récemment, Sakurai et al. (2007), ont
démontré que TACI, membre de la famille des récepteurs TNF régulerait la production d’IgA
dans les cellules B par APRIL via son association avec des HSPGs. Les protéoglycannes présents
à la surface cellulaire et dans la MEC interagiraient également avec des interleukines (IL3, IL-6,
IL8, IL7, IFNγ…) et des chimiokines (CXCL4 ou facteur plaquettaire 4, SDF-1 pour Stromal
cell-Derived Factor, RANTES…) souvent produites au niveau des sites inflammatoires (Ashikari
et al., 1995; Proudfoot et al., 2001; Spillmann et al., 1998; Stringer and Gallagher, 1997; Tanaka
et al., 1998). Ils retiendraient ces molécules à la surface cellulaire augmentant de ce fait leur
concentration effective (gradient) au voisinage des récepteurs (Figure 36). Les CSPGs sont
également connus pour jouer des rôles importants dans l’adhésion cellulaire et la migration
cellulaire mais surtout au niveau des cellules neuronales et/ou au niveau du cartilage.

104
Enfin, leurs multiples fonctions peuvent être exploitées à des fins thérapeutiques. Par exemple,
l'héparine est depuis longtemps utilisée pour son activité anticoagulant (Petitou et al., 1988) grâce
à son interaction avec l’antithrombine III stabilisant la liaison avec la thrombine, les
chondroïtines sulfates sont, quant à elles, testées pour tenter de ralentir voire stopper le
développement de l'arthrose (Leeb et al., 1996).
Il a été démontré que le profil de sulfatation des unités disaccharidiques constituant les chaînes
GAGs et surtout CS influençait les propriétés de liaison des GAGs et donc la fonction des
CSPGs. Les monocytes/macrophages sécrètent spécifiquement des CSPGs et 80 à 90% de ces
CSPGs seraient des C4S (CSA ou CSB) (Laskin et al., 1991; Uhlin-Hansen et al., 1993).
Rachmilewitz et Tykocinski, en 1998 ont regardé les effets des C4S sur des PBMC (Human
Peripheral Blood Mononuclear Cell) et ont montré que CSA activerait les monocytes qui alors,
sécrèteraient de l’IL-1β (cytokine pro-inflammatoire) quand à CSB, il activerait les cellules B
(pas les T) par le biais de CD44 (prolifération qui peut-être inhibée en présence de TGFβ et
d’anti-CD44). Il a en outre été démontré que HA induit la production d’IL-1β (de même que
TNFα et IGF-1) par les macrophages et la différenciation/prolifération des cellules B dépendant
de son interaction avec CD44 (Noble et al., 1993; Rafi et al., 1997). De plus, Aoyama et al. 2005,
ont montré que CSB induit la prolifération des B murins dépendants de la voie de la PKC et
PI3K, induisant la phosphorylation de Akt mais pas de ERK. La serglycine est un CSPG (C4S),
sécrétée par les cellules hématopoïétiques. On en retrouve dans les granules de sécrétions des
mastocytes, des basophiles et des cellules NK (Natural Killer). Elle serait également un ligand de
CD44 et contribuerait à activer les T cytotoxiques (Toyama-Sorimachi et al., 1995). Elle serait
également constitutivement sécrétée dans des cellules plasmatiques myélomateuses (Theocharis
et al., 2006). Akiyama et al, en 2004, ont montré que la présence de CS dans les cultures
cellulaires en présence d’ovalbumine, induisait la sécrétion de cytokines de type TH1 (IFNγ, IL2
et IL12) et supprimait ou diminuait fortement la sécrétion de cytokines de type TH2 (IL5 et
d’IL10) par des splénocytes préalablement sensibilisés à l’ovalbumine. Cette activité semble être
renforcée lorsqu’ils utilisent des dissaccharides CSA (sulfate en position 4 et pas d’IdoA) et CSE
comparés aux autres séquences. Le degrè de sulfatation ainsi que la position des sulfates semblent
donc être importants dans l’activité des CS. De plus, l’association de CSA avec la L-selectin
exprimée à la surface des lymphocytes T jouerait également un rôle important. Il a également été
montré que le Versican (CSPG contenant des chaînes CS et DS) interagirait avec la L-, la P-
selectin (acteurs moléculaires de l’adhésion cellulaire qui, en reconnaissant leurs différents
ligands, vont autoriser un contact transitoire des leucocytes avec la couche monocellulaire

105
endothéliale, de façon à permettre leur attachement, leur déplacement, puis en dernier lieu leur
migration trans-endothéliale) (McEver et al., 1995), mais également avec certaines chemokines
comme SLC, IP-10 et SDF-1β grâce à des motifs disaccharidiques hautement sulfatés (CSE). Par
contre l’interaction Versican-CD44 requiert des motifs disaccharidiques moins voir pas du tout
sulfatés comme peut l’être, HA (Kawashima et al., 2002). Vander Voort et al. (2000) ont
démontré, chez l’homme, que les lymphocytes B d’amygdales fraîchement isolés, exprimaient
peu de HSPGs et qu’après activation du BCR et/ou du CD40, il y avait une forte augmentation de
l’expression des HSPGs membranaires comme par exemple, CD44 qui via ses chaînes HS, allait
interagir avec HGF et induire la transduction d’un signal (phosphorylation de Met, Akt/PKB).
Les HSPGs peuvent donc être considérés comme des co-récepteurs fonctionnels intervenant dans
la régulation B spécifique de l’antigène. Il a également été démontré que différents mitogènes
induisaient la synthèse de GAGs (CS et HS) que ce soit dans le milieu de culture ou à la surface
cellulaire dans 3 lignées humaines Jurkat (lignée T), Daudi (lignée B) et THP-1 (Makatsori et al.,
2001).
Figure 36 : Les HSPGs jouent de nombreux rôles dans la physiologie cellulaire.
Schéma de Bishop et al. Nature 2007.

106

107
Résultats - Discussion

108

109
Première partie : les BCR modifiés

110

111
Article 1
“Premature expression of α Ig heavy chains delays B cell differentiation and
promotes plasma cell maturation but not mucosal homing”
Sophie Raynal-Duchez, Rada Amin, Nadine Cogné, Christophe Sirac, Laurent Delpy, Micael
Bardel, Blaise Corthesy and Michel Cogné.
(Manuscrit soumis à Journal of Experimental Medecine)
Cet article est consacré à l’étude par un modèle de knock-in chez la souris, de la fonction
d’un BCR commuté à IgA afin de savoir si une chaîne α peut se substituer et promouvoir un
développement B ainsi qu’une réponse B-spécifique à l’antigène comme le fait normalement une
chaîne lourde µ. On sait en effet, qu’une chaîne lourde δ peut fonctionner de façon quasi-
identique à µ (Lutz et al., 1998) alors qu’une chaîne γ se comporte au contraire, de façon
spécifique, moins efficace que µ pour la différenciation B précoce mais hyperstimulatrice dans
les cellules B matures en cas de pontage du BCR IgG même si les modalités moléculaires
précises de cette hyper-réactivité restent l’objet de controverses (Horikawa et al., 2007; Pogue
and Goodnow, 2000; Waisman et al., 2007; Wakabayashi et al., 2002). Alors que de nombreuses
études ont été consacrées au BCR IgG, très peu ont exploré la fonction du BCR IgA (Leduc and
Cogne, 1996; Sato et al., 2007) et nous avons donc décidé d’aborder cette question dans un
modèle de knock-in où le gène Cα1 humain a été intégré par recombinaison homologue au niveau
de la région switch µ afin d’obtenir une lignée de souris transgéniques n’exprimant que des IgA.
Nous avons choisi le gène constant humain pour être en mesure de distinguer l’expression de
notre transgène de celle du gène Cα endogène, en particulier chez les hétérozygotes, mais aussi
chez les homozygotes puisque nous savions d’expérience que la délétion de Sµ ne bloque que
partiellement le switch sur le locus muté (Khamlichi et al., 2004).
Cette étude nous permet de dire que la chaîne lourde α est capable de jouer un rôle
similaire à celui d’une chaîne lourde µ pendant les étapes précoces de la différenciation, à savoir
son expression à la membrane et son association avec les modules de signalisation Igα/Igβ afin
de permettre un développement B. Mais on note certaines différences qui permettent de dire que,
comme pour la chaîne γ, le pré-BCR α est moins efficace qu’un pré-BCR µ car on observe un
blocage partiel à la transition pro/préB ainsi qu’une augmentation de l’éditing du BCR

112
manifestée par un ratio κ/λ plus élevé. Malgré ce blocage partiel, une population B à IgA de
membrane est retrouvée dans tous les compartiments B périphériques. Ces B peuvent être activés
après rencontre avec un antigène et être à l’origine d’une réponse spécifique. On remarque
également la présence d’anticorps murins d’isotypes différents dans le sérum de ces animaux,
indiquant que les cellules à IgA de membrane forcée peuvent subir la commutation de classe en
l’absence de Sµ, comme des B à IgM dans le cas de la délétion isolée de Sµ.
Le trait phénotypique le plus marqué reste l’hyperprolifération plasmocytaire. Ces B à IgA de
membrane seraient beaucoup plus activables à l’état basal et ont une grande facilité à se
transformer en plasmocytes après stimulation avec un mitogène d’où l’abondance de plasmocytes
observés dans les différents compartiments lymphoïdes périphériques. Ce trait constitue un point
commun avec le BCR IgG lui aussi hyperactivable (même si l’hyperdifférenciation plasmocytaire
dans tous les modèles d’expression forcée d’un BCR IgG est apparue comme un trait inconstant).
L’IgA rejoindrait donc selon notre étude l’IgG parmi les BCR surtout effecteurs (c'est-à-dire très
efficaces dans les cellules matures stimulées), tandis que la chaîne µ comme la chaîne δ semblent
les plus efficaces composants des pre-BCR et BCR affecteurs qui sous-tendent la maturation B
précoce et permettent à chaque clone B de valider son paratope et de rejoindre, en fonction du
niveau de stimulation que lui confère ce paratope, l’un ou l’autre des compartiments B.
Un dernier point important de notre équipe concerne l’absence d’effet positif de l’expression
d’une IgA membranaire sur le homing muqueux des lymphocytes B alors que les tissus
lymphoïdes associés aux muqueuses sont pourtant le lieu préférentiel de résidence des cellules B
normales à IgA, validant donc l’hypothèse que le homing muqueux des cellules B normales est
initialement réalisé par des cellules à IgM membranaires, qui commutent ensuite sur place vers la
classe IgA.
Contribution personnelle à ce travail :
Ma part de cette étude, en collaboration avec Rada Amin, a consisté en un phénotypage très
détaillé (cytométrie de flux, immunisations, stimulations, tris cellulaires et préparation d’ARN,
d’ADNc, PCR quantitative…) d’animaux préalablement établis au laboratoire mais caractérisés
de façon très préliminaire.

113
Article 2
“B cells carying an IgE BCR are intrisically short-lived”
Sophie Raynal-Duchez, Hezhong Liu, Nadine Cogné, Claire Carrion, Yves Denizot, Jean Sainte-
Laudy and Michel Cogné.
(Manuscrit en préparation)
Cet article préliminaire et encore « en travaux », est consacré à l’étude, par des modèles de
transgenèse et de transfection, de la fonction des IgE de forme membranaire. Il rapporte deux
conclusions principales :
- la chaîne lourde ε des IgE n’est pas capable de jouer un rôle similaire à celui d’une chaîne
µ pendant les étapes précoces de la différenciation, malgré ses homologies de structure
avec les chaînes µ (en particulier pour ce qui concerne l’ancrage membranaire et
l’association avec les modules de signalisation Igα/Igβ du BCR)
- lors de son expression dans une lignée B de phénotype B mature, cette chaîne lourde ε
assemblée au sein d’un BCR, est à l’origine d’un signal constitutif pro-apoptotique.
Ces observations suggèrent fortement que les propriétés mêmes de l’IgE membranaire sont à
l’origine de la quasi-absence des cellules à BCR IgE in vivo chez l’individu sain. Cette absence
est d’autant plus surprenante que in vitro et sur des cultures à court terme, des taux élevés de
cellules à IgE membranaire (voisin des taux de cellules à IgG1 membranaire) peuvent être induits
en présence d’activateurs B polyclonaux et d’IL4.
L’idée que l’expression d’un BCR IgE ne soit possible que de façon transitoire et puisse aboutir à
une différenciation plasmocytaire mais non à une différenciation B mémoire cadre donc bien
avec les observations faites chez l’homme comme chez la souris, et place l’IgE membranaire
dans le rôle d’une molécule auto-régulatrice de la réponse IgE.
Un certain nombre d’expériences restent en cours pour compléter cet article en préparation,
avec en particulier :
- la détermination de la voie d’apoptose en cause dans les transfectants (en particulier en
vérifiant sa sensibilité aux inhibiteurs des caspases)

114
- la décontamination de la lignée εKI par transfert d’embryons afin de pouvoir élever des
animaux homozygotes de façon stable (alors qu’ils meurent actuellement très vite du fait
de déficit immunitaire)
- le croisement des animaux dans un fond p53-/-, pour tester la possibilité de lever ainsi le
blocage de différenciation précoce
- l’établissement de lignées d’animaux transgéniques porteuses de « la construction ε
réarrangée » déjà utilisée pour transfecter la lignée A20.
Le résultat attendu de ces deux expériences serait l’obtention de deux types de lignées : celles
ayant réussi à bloquer l’expression du transgène et possédant donc un système immunitaire
normal, et celles exprimant fortement le transgène IgE dont nous prédirions qu’il devrait alors
induire une importante attrition de l’ensemble des compartiments B.
Contribution personnelle à ce travail :
Ma part de cette étude a été le suivi des animaux εKI depuis le stade F1 et la caractérisation
complète du phénotype, ainsi que la réalisation des constructions utilisées dans la lignée A20,
l’obtention et l’étude des transfectants.

115
Demande de dépôt de brevet
Etablissement et caractérisation d’une lignée de souris transgéniques homozygotes pour la mutation gammaprim
(introduction du gène de chaîne lourde d’immunoglobuline γ1 humaine au sein du locus
IgH murin) Inventeurs : Michel Cogné, Sophie Raynal-Duchez, Eric Pinaud, Nadine Cogné.
Une partie de mon travail de recherche a été la mise au point d’une lignée de souris portant un
BCR modifié codant une chaîne lourde γ1 humaine (criblage des clones ES, suivi des animaux
chimériques puis F1, caractérisation phénotypique complète). Un gène Cγ1 humain a été intégré par
recombinaison homologue au niveau de la région switch µ du locus IgH endogène. L’obtention de
cette lignée présente un intérêt biotechnologique en permettant de produire des IgG1 humanisées
monoclonales chez la souris, la souris étant un modèle idéal pour la production ultérieure
d’hybridomes (Linenberger et al., 2002). L’avantage de ces souris « modifiées » réside dans la
conservation de leur répertoire B, et de leur capacité à générer des réponses spécifiques de forte
affinité vis-à-vis d’antigènes choisis. Les souris « IgG1 » permettront de produire des anticorps
monoclonaux humanisés de classe IgG1 dont les applications seront multiples : par exemple dans le
domaine des bioréactifs (réactifs de diagnostic) et surtout dans des applications thérapeutiques et
pharmacologiques (synthèse d’anticorps thérapeutiques destinés à la prévention et au traitement de
maladies infectieuses, de cancers). Les anticorps humanisés présentant une très bonne tolérance chez
l’homme constituent à l’heure actuelle une voie thérapeutique en plein essor.
Même s’il existe d’autres voies pour produire des IgG1 humaines actuellement (humanisation
in vitro de monoclonaux murins, utilisation de la « xenomouse »), l’un des intérêts majeurs de notre
modèle réside dans la possibilité d’obtenir d’emblée, dès le stade hybridome, des anticorps dont la
chaîne lourde est une chaîne γ1 humaine. Ils permettent donc, devant un pool d’anticorps, de choisir
d’emblée celui qui est le plus efficace dans les tests (alors que lors d’une humanisation in vitro,
prévoir si un anticorps donné restera actif ou sera le plus actif d’un groupe, relève de la divination… )
B Cell Design, start-up en cours de création, rattachée au laboratoire, valorise déjà la lignée
de souris productrices d’IgA monoclonales humanisées (Cogné M. et al., 2003) et se propose de
valoriser cette lignée de souris transgéniques IgG1.

116
Ma contribution personnelle à ce travail :
Ma contribution à ce travail a été la transfection de la construction γ1 KI dans les cellules ES, le
criblage des clones positifs et le phénotypage complet des animaux.
L’aspect biotechnologique mis de côté, il est intéressant de regarder le phénotype de ces souris
sous l’aspect immunologique fondamental. Bien que la question de la capacité d’un BCR IgG à
se substituer à un BCR IgM lors du développement B ait déjà été posée par plusieurs équipes
décrivant des souris transgéniques ou knock-in… elle reste en suspens… Il y a quelques années,
les travaux de Tsubata et Goodnow avaient démontré une fonction hyper-activatrice du BCR IgG,
avec hyperplasmocytose accompagnée d’un fort taux d’IgG sérique, suggérant la notion d’un
BCR IgG « effecteur ». Il avait alors été démontré que ce BCR IgG transgénique n’était pas
sensible à la régulation négative de CD22 via la phosphatase SHP-1… mais ces données viennent
d’être contestées par Waisman et al., début 2007, n’identifiant ni hyper-différenciation
plasmocytaire, ni indépendance par rapport à CD22 dans des cellules B « knock-in pour l’IgG1
murine » qui cette fois sont, en plus, capables de construire un vrai répertoire B diversifié car les
autres modèles étaient des modèles transgéniques utilisant un VDJ déjà réarrangé. Il faut
cependant noter que le modèle de Waisman est biaisé car le gène γ1 utilisé est délété de son site
de polyadénylation « de type sécrété… »… on peut donc imaginer que cette délétion est assez
indigeste pour des cellules voulant se différencier en plasmocytes !!!. Les travaux d’Horikawa
(2007) vont cependant dans le même sens que ceux de Waisman alors que la mutation qu’ils
étudient, ne s’accompagne pas d’une délétion du site polyA de type sécrété. Dans leur modèle, il
s’agit d’un simple remplacement de la partie trans-membranaire et intracellulaire de l’IgM par
celle d’une IgG1 dans un modèle de transgène anti-lysozyme. On peut cependant noter que ce
modèle présente un autre biais : celui de présumer que la spécificité de signal d’une IgG par
rapport à une IgM se résume au rôle de sa queue intra-cellulaire (dans notre modèle, nous avons
l’avantage de travailler avec une chaîne γ1 complète !). Même s’il n’est pas inducteur de
plasmocytose et s’il est soumis à l’inhibition par CD22, le BCR IgG1 pour Waisman et Horikawa
apparaît plus activateur qu’une IgM (en matière de flux calciques) et pouvant lui être substitué
précocément pendant la différenciation des progéniteurs B dans la moelle osseuse mais avec un
ralentissement de la transition proB vers pré-B.
Le modèle KI IgG1 humaine que nous avons décrit dans cette thèse, avec une application
potentiellement plutôt biotechnologique, est encore partiellement étudié et nécessite des
explorations poussées pour qu’on puisse l’opposer aux précédents et comprendre de quels

117
mécanismes relèvent les différences observées. Quoi qu’il en soit, ce modèle manifeste
clairement une hyper-différenciation plasmocytaire attestée par un ratio d’expression Blimp-1 (ou
Xbp-1) / CD79a d’environ 30 dans la rate des animaux homozygotes pour la mutation, et de 75
dans les ganglions drainants et pouvant atteindre des records de 130 après immunisation des
souris. Par ailleurs, un blocage sévère de la maturation B à la transition pro-B/pre-B est observé
mais malgré l’absence d’expression d’IgM/d’IgD, un compartiment B périphérique reste capable
de se différencier chez ces animaux et représente environ 6% des splenocytes totaux. Ces B
peuvent donc être activés après rencontre avec un antigène et être à l’origine d’une réponse
spécifique. Comme pour les souris αKI, on remarque la présence d’anticorps murins endogènes
d’isotypes variés dans le sérum, indiquant que les B IgG1 peuvent subir la commutation de classe
en l’absence de Sµ.
Les différences notables entre les deux lignées γ1KI (Waisman et la nôtre) restent 1) à
notre désavantage, l’utilisation dans notre cas d’une région constante humaine qui peut donc
compliquer la réflexion même si on sait que les régions transmembranaires et cytoplasmiques
IgG murines et humaines sont fortement homologues… (Figure n°24, chapitre IV) et 2) à notre
avantage, l’utilisation d’un gène complet capable de participer à une différenciation
plasmocytaire. En réponse aux doutes que peut susciter le point 1), il est en tout cas frappant de
retrouver dans notre modèle l’hyper-différenciation plasmocytaire d’abord décrite avec des
transgéniques γ1 murine.
Il est à noter que les double-hétérozygotes α/γ, issues de croisement entre souris γ1KI et
αKI, après immunisation antigénique présentent une réponse spécifique IgG1 et IgA à des taux
élevés. La réponse IgA est identique à celle obtenue dans des souris homozygotes αKI mais on
obtient après immunisation une meilleure réponse IgG1 spécifique des souris α/γ par rapport aux
souris homozygotes γ1KI.
De plus amples manipulations doivent donc être réalisées pour comprendre les différences
de phénotypes dans les différents modèles de transgenèse et de knock-in IgG1 mais également
comprendre pourquoi les doubles hétérozygotes α/γ répondent mieux à une stimulation
antigénique que les simples homozygotes γ1… ( la capacité de ces cellules, non seulement à se
différencier, mais aussi à présenter l’antigène aux cellules T et à bénéficier alors de leur aide est
sans doute à cet égard un point qui méritera une attention particulière).

118

119
Etablissement et caractérisation d’une lignée de souris transgéniques homozygotes pour la mutation gammaprim
(introduction du gène de chaîne lourde d’immunoglobuline γ1 humaine au sein du locus
IgH murin)
A. METHODOLOGIE POUR L’OBTENTION DES ANIMAUX MUTANTS ET LEUR
CARACTERISATION
1) Construction du vecteur de ciblage de la recombinaison homologue
Les constructions plasmidiques ont été réalisées à partir du plasmide bluescript SK (pSK)
(Stratagene, LaJolla, CA, USA) et de la souche bactérienne E. coli TG1 (Stratagene, LaJolla,
CA, USA), en utilisant les protocoles classiques de préparation, de clonage et d’analyse de
l’ADN tels que ceux décrits dans Current Protocols in Molecular Biology (Frederick M.
AUSUBEL, 2000, Wiley and son Inc, Library of Congress, USA).
Le vecteur de recombinaison homologue ou vecteur de ciblage dérivé de pSK, dénommé p-
gamma1KI, comprend : i) un fragment de 6860 pb du gène d’immunoglobuline gamma 1 humain
incluant plusieurs exons codant pour les domaines constants CH1, hinge, CH2 et CH3 et les
exons dits « de membrane » (m1 et m2) de la chaîne lourde d’immunoglobuline γ1 ii) une
cassette néo (fragment de 1,6 kb), iii) des séquences flanquantes homologues à une portion du
locus murin des chaînes lourdes d’immunoglobulines, avec en amont un fragment d’environ 5 kb
correspondant à la région JH-Eµ (fragment DQ 52/JH) en aval d’un autre fragment d’environ 5
kb correspondant au gène Cµ (fragment Cµ).
De manière plus précise, les différents fragments ont été insérés dans le plasmide
bluescript SK, selon les étapes suivantes :
- Dans une première étape, le fragment Cµ a été cloné au site Xho1 de pSK pour donner le
plasmide pA.
- Dans une seconde étape, le fragment DQ 52/JH a été cloné en 5’ du fragment Cµ dans le
plasmide B, aux sites EcoRV- Cla I, pour donner le plasmide pB.
- Dans une troisième étape, une cassette néo a été insérée au site Sal1 entre DQ52/JH et Cµ, pour
donner le plasmide pC.
- Enfin, dans une dernière étape, le fragment Cγ1 de 6860 pb flanqué d’adaptateurs Cla1 et
contenant le gène gamma 1 humain a été inséré entre le fragment JH et la cassette néo au site Cla
I du plasmide pC pour donner le vecteur de ciblage dénommé p-gamma1KI.
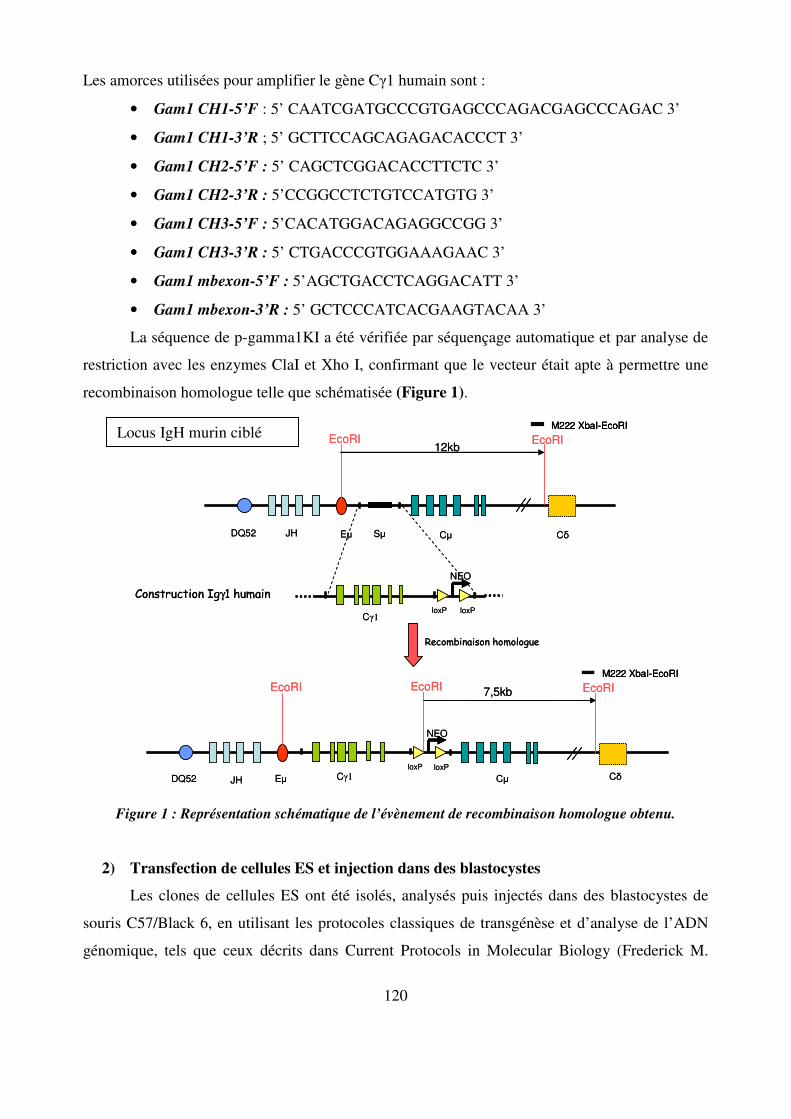
120
Les amorces utilisées pour amplifier le gène Cγ1 humain sont :
• Gam1 CH1-5’F : 5’ CAATCGATGCCCGTGAGCCCAGACGAGCCCAGAC 3’
• Gam1 CH1-3’R ; 5’ GCTTCCAGCAGAGACACCCT 3’
• Gam1 CH2-5’F : 5’ CAGCTCGGACACCTTCTC 3’
• Gam1 CH2-3’R : 5’CCGGCCTCTGTCCATGTG 3’
• Gam1 CH3-5’F : 5’CACATGGACAGAGGCCGG 3’
• Gam1 CH3-3’R : 5’ CTGACCCGTGGAAAGAAC 3’
• Gam1 mbexon-5’F : 5’AGCTGACCTCAGGACATT 3’
• Gam1 mbexon-3’R : 5’ GCTCCCATCACGAAGTACAA 3’
La séquence de p-gamma1KI a été vérifiée par séquençage automatique et par analyse de
restriction avec les enzymes ClaI et Xho I, confirmant que le vecteur était apte à permettre une
recombinaison homologue telle que schématisée (Figure 1).
Figure 1 : Représentation schématique de l’évènement de recombinaison homologue obtenu.
2) Transfection de cellules ES et injection dans des blastocystes
Les clones de cellules ES ont été isolés, analysés puis injectés dans des blastocystes de
souris C57/Black 6, en utilisant les protocoles classiques de transgénèse et d’analyse de l’ADN
génomique, tels que ceux décrits dans Current Protocols in Molecular Biology (Frederick M.
Targeted mouse IgH locus
Recombinaison homologue
JH Eµ CµDQ52
NEO
loxP loxP
M222 XbaI-EcoRI
EcoRI7,5kbEcoRI EcoRI
Cδ
Construction Igγ1 humain
JH Eµ Sµ CµDQ52
loxP loxP
M222 XbaI-EcoRI
EcoRI EcoRI
Cδ
12kb
Cγ1
Cγ1
NEO
Targeted mouse IgH locus
Recombinaison homologue
JH Eµ CµDQ52
NEO
loxP loxP
M222 XbaI-EcoRIM222 XbaI-EcoRI
EcoRI7,5kbEcoRI EcoRI
Cδ
Construction Igγ1 humain
JH Eµ Sµ CµDQ52
loxP loxP
M222 XbaI-EcoRIM222 XbaI-EcoRI
EcoRI EcoRI
Cδ
12kb
Cγ1
Cγ1
NEO
Locus IgH murin ciblé

121
AUSUBEL, 2000, Wiley and son Inc, Library of Congress, USA). De manière plus précise, des
cellules ES ont été transfectées par électroporation de l’ADN de p-gamma1KI linéarisé au site
Not I. Les clones sélectionnés en présence de généticine ont été prélevés et leur ADN digéré par
EcoRI puis analysé par Southern Blot à l’aide d’une sonde radioactive s’hybridant en dehors du
site de recombinaison homologue, en 5’ du gène constant IgH Delta (δ) et de son site EcoR I.
Figure 2 : A. linéarisation du vecteur avant transfection (ND, non digéré ; NotI, linéarisé par l’enzyme
NotI); B. Southern blot d’un clone mutant (1) et de ses sous-clones (3 et 4) par comparaison à un ADN
non muté (2)
La présence d’un allèle recombinant est visualisée par un fragment d’environ 7,5 kb
(représentant le fragment µ murin et la cassette néo) alors que l’allèle sauvage correspond à un
fragment de 12 kb. Dans ces conditions, sur 192 clones analysés, 1 s’est révélé positif (n°153).
La vérification du caryotype n’a montré aucune anomalie chromosomique (aneuploïdie).
Ce clone a été injecté dans des blastocystes de souris C57/Black 6 en utilisant les
protocoles classiques de transgénèse tels que ceux décrits dans Transgenic Mouse: Methods and
Protocols (Methods in Molecular Biology (Clifton, N.J.), V. 209.). Un ♂ chimère, présentant un
degré de chimérisme élevé, a été obtenu et mis en accouplemement avec des ♀ C57/Black 6 afin
d’obtenir des animaux hétérozygotes qui ont été analysés par PCR et ELISA. Une lignée de
souris homozygotes pour le locus IgH recombiné, dénommée ci-après lignée gamma 1 knock-in
ou γ1KI, a ensuite été obtenue par croisement des animaux hétérozygotes.
153Sous-clones
12 kb
7,5 kb
153Sous-clones
12 kb
7,5 kb
λ Hind III N.D. Not Iλ Hind III N.D. Not I
1 2 3 4

122
3) Détection du locus IgH recombiné portant le gène Cγ1 humain (allèle gamma 1
knock-in ou γ1KI) et du locus IgH sauvage (allèle µµµµ sauvage) par PCR
L’ADN génomique d’un échantillon de queue d’animaux homozygotes obtenus comme
précisé ci-dessus, a été analysé par PCR à l’aide des deux couples d’amorces suivants :
- couple spécifique du locus IgH murin non muté (allèle µ sauvage) :
- amorce UpstreamSpe I Smu : 5’ GAG TAC CGT TGT CTG GGT CAC 3’
- amorce SacI-3’Imu : 5’ GAG CTC TAT GAT TAT TGG TTA AC 3’
La réaction d’amplification a été réalisée avec une température d’hybridation de 61°C. Cette PCR
amplifie en 30 cycles un fragment de 91 paires de bases-encadrant le site Spe I-spécifique du
locus IgH murin non muté.
- couple spécifique du locus IgH recombiné portant le gène Cγ1 humain (allèle
gamma1 knock-in ou γ1KI ) :
- amorce NeoI : 5’ GCA TGA TCT GGA CGA AGA GCA T 3’
- amorce Neo2 : 5’ TCC CCT CAG AAG AAC TCG TCA A 3’
La réaction d’amplification a été réalisée avec une température d’hybridation de 55°C. Cette PCR
amplifie en 30 cycles un fragment de 120 paires de bases spécifique du locus IgH recombiné
portant le gène Cγ1 humain.
Une lignée de souris homozygotes pour la mutation γ1KI, dénommée ci-après lignée gamma
1 knock-in ou gammaprim a été établie ; les animaux de cette lignée sont systématiquement et
simultanément négatifs en PCR avec les amorces spécifiques de l’allèle µ sauvage et positifs en
PCR avec les amorce spécifiques de l’allèle gamma 1 knock-in.
Figure 3: Southern Blot sur ADN
génomique de queue de souris
WT (piste A)
hétérozygote γ1-KI (piste B)
homozygote γ1-KI (piste C)
homozygote γ1-KI floxé (piste D)

123
4) Dosage du taux des IgG1 sériques totales en néphélométrie et en ELISA
Les IgG1 humaines sériques ont été dosées par néphélométrie sur un automate BNII de la
société Behring en utilisant le kit Behring IgG1 selon les recommandations du fournisseur.
Ces résultats ont été confirmés en ELISA suivant la technique suivante :
- coating de plaques 96 puits (Maxisorb, NUNC) par incubation une nuit à 4° avec un
anticorps non marqué anti-IgG humaine (monoclonal anti-human IgG, Sigma) dilué au
1/1000ème en tampon Carbonate 0,1 M pH 8,3 (100 microlitres/puits)
- 3 lavages en PBS Tween 0,1%
- saturation des plaques en milieu complet {PBS, 3% BSA}(100 microlitres/puits)
- 3 lavages en PBS Tween 0,1%
- distribution des sérums à tester dilués au 1/100ème et au 1/1000 ème en tampon {PBS, 1%
BSA}(100 microlitres/puits)
- incubation 3 heures à 37°C
- 3 lavages en PBS Tween 0,1%
- distribution d’un antisérum anti-IgG humaines marqué à la phosphatase alcaline
(monoclonal anti-human IgG-AP, SIGMA) dilué au 1/1000ème (100 microlitres/puits)
- 3 lavages en PBS Tween 0,1%
- réaction enzymatique de révélation par addition de PNP en tampon TRIS 0,2M (Sigma)
- blocage de la réaction par addition de soude 3N (30 microlitres/puits)
- lecture en spectrométrie à une longueur d’onde d’absortion de 405 nm.
5) Recherche de l’expression d’un récepteur membranaire de la classe des IgG
humaines à la surface des lymphocytes périphériques des animaux mutants
Préparation des cellules lymphoïdes : La rate, organe lymphoïde secondaire, a été choisie pour
cet expérimentation. Cet organe a été prélevé chez différents animaux mutants homozygotes γ1KI
puis dilacéré en tampon VERSENE (Invitrogen), et filtré sur tamis (40 microns) afin d’obtenir
une suspension de cellules individuelles débarrassée des agégats cellulaires. Les cellules
spléniques ont alors été centrifugées et soumises à une étape supplémentaire de choc osmotique
afin d’obtenir la lyse des globules rouges par reprise du culot cellulaire dans 1 ml d’eau distillée.
Les cellules ont ensuite été immédiatement reprises en milieu complet (RPMI + 10% sérum de
veau fœtal), numérées et conservées sur glace.

124
Marquage à l’aide d’anticorps fluorescents :
- 105 cellules ont été incubées pendant 30 minutes à 4° avec une dilution au 1/100ème d’un
anticorps anti-IgM de souris marqué à l’isothiocyanante de fluorescéïne (Southern
Biotechnologies) et un anticorps anti-IgG humain couplé Alexa 633.
- Les cellules ont ensuite été lavées dans 5ml de PBS.
- Le surnageant a été décanté et les cellules ont été reprises dans 100 microlitres de PBS,
0,5% BSA, 0,1 mM EDTA
Analyse en cytofluorimétrie : sur un cytomètre de flux COULTER XL.
6) Immunisation des animaux
Les animaux ont été immunisés une première fois par injection intra-péritonéale de 20
microgrammes d’antigène P24 (protéine de la pepside VIH-1) diluée dans 200 microlitres de
sérum physiologique et mise en émulsion avec 200 microlitres d’adjuvant complet de Freund
(SIGMA)
Toutes les 2 semaines, les animaux ont subi un rappel vaccinal par injection intra-péritonéale de
20 microgrammes d’antigène P24 diluée dans 200 microlitres de sérum physiologique et mise en
émulsion avec 200 microlitres d’adjuvant incomplet de Freund (SIGMA).
7) Dosage des anticorps spécifiques de l’antigène P24
La présence d’anticorps spécifiques de l’antigène P24 a été recherchée par ELISA 2 semaines,
puis 4 semaines après la deuxième injection de l’antigène suivant la technique suivante :
- coating de plaques 96 puits (Maxisorb, NUNC) par incubation une nuit à 4° avec de
l’antigène P24 à la concentration de 1 microgrammes/ml en tampon Carbonate 0,1 M pH
8,3 (100 microlitres/puits)
- 3 lavages en PBS Tween 0,1%
- saturation des plaques en milieu complet {PBS, 3% BSA}(100 microlitres/puits)
- 3 lavages en PBS Tween 0,1%
- distribution des sérums à tester dilés au 1/100ème et au 1/200 ème en tampon {PBS, 1%
BSA}(100 microlitres/puits)
- incubation 3 heures à 37°C
- 3 lavages en PBS Tween 0,1%

125
- distribution d’un antisérum anti-IgG humaines marqué à la phosphatase alcaline
(monoclonal anti-human IgG-AP, SIGMA) dilué au 1/1000ème (100 microlitres/puits)
- 3 lavages en PBS Tween 0,1%
- réaction enzymatique de révélation par addition de PNP en tampon TRIS 0,2M (Sigma)
- blocage de la réaction par addition de soude 3N (30 microlitres/puits)
- lecture en spectrométrie à une longueur d’onde d’absortion de 405 nm.
- Le taux des anticorps IgG anti-P24 a été exprimé en unités arbitraires établies pour la
moyenne des sérums dilués au 1/100ème et 1/200ème en fonction du rapport Densité optique
sérum testé / Densité optique du sérum témoin.

126
B. PHENOTYPE DES ANIMAUX MUTANTS
1. Mise au point de tests PCR spécifiques de l’allèle muté et de l’allèle non muté
Une lignée homozygote pour la mutation γ1-KI a d’abord été établie. Les animaux de cette lignée
sont systématiquement et simultanément négatifs pour les test « PCR allèle µ sauvage » et
positifs pour le test « PCR γ1-KI».
2. Dosage du taux des IgM murines et des IgG « humaines » sériques totales en
néphélométrie et en ELISA
a. Par néphélométrie, les dosages d’IgG1 humaines sériques ont donné des résultats
totalement corrélés avec ceux du génotypage réalisé par les tests PCR :
- les animaux « contrôles « non mutants » ont un taux nul d’immunoglobulines humaines
de classe IgG1
- les animaux homozygotes γ1-KI ont toujours un taux significatif d’IgG1 humaines, ce
taux variant entre 0,5 et 1 g/l dans le sérum. Par contre, les IgM murines sont
indétectables dans le sérum de ces animaux.
b. En ELISA, des résultats très tranchés permettent également de distinguer hétérozygotes
et homozygotes en ELISA avec des variations très franches : valeurs situées à 0,8 g/l d’IgG1
humaines pour les sérums d’homozygotes dilués au 1/500ème ; valeurs nettement plus faibles
(0,03) pour les sérums d’hétérozygotes même à la dilution la plus faible (au 1/100ème). A l’inverse
lorsque les IgM murines sont dosées en ELISA, on observe un taux normal d’IgM murines (de
l’ordre de 1 g/l) chez les souris contrôles « non mutantes » de même que chez les animaux
hétérozygotes pour la mutation gammaprim. Par contre, le taux des IgM murines est nul chez les
animaux homozygotes gammaprim.
3. Recherche de l’expression d’un récepteur membranaire de la classe des IgG
humaines à la surface des lymphocytes B périphériques des animaux mutants
Les animaux homozygotes porteurs de la mutation γ1-KI ont été phénotypés grâce à la cytométrie
en flux, avec des résultats en complet accord avec ceux donnés par le dosage des
immunoglobulines sériques. Chez ces animaux, on ne détecte aucune expression d’IgM murines.
Pourtant en l’absence d’expression d’IgM, un compartiment de cellules B périphériques CD19+
est capable de se différencier et représente 6 % des lymphocytes spléniques. Ces cellules B
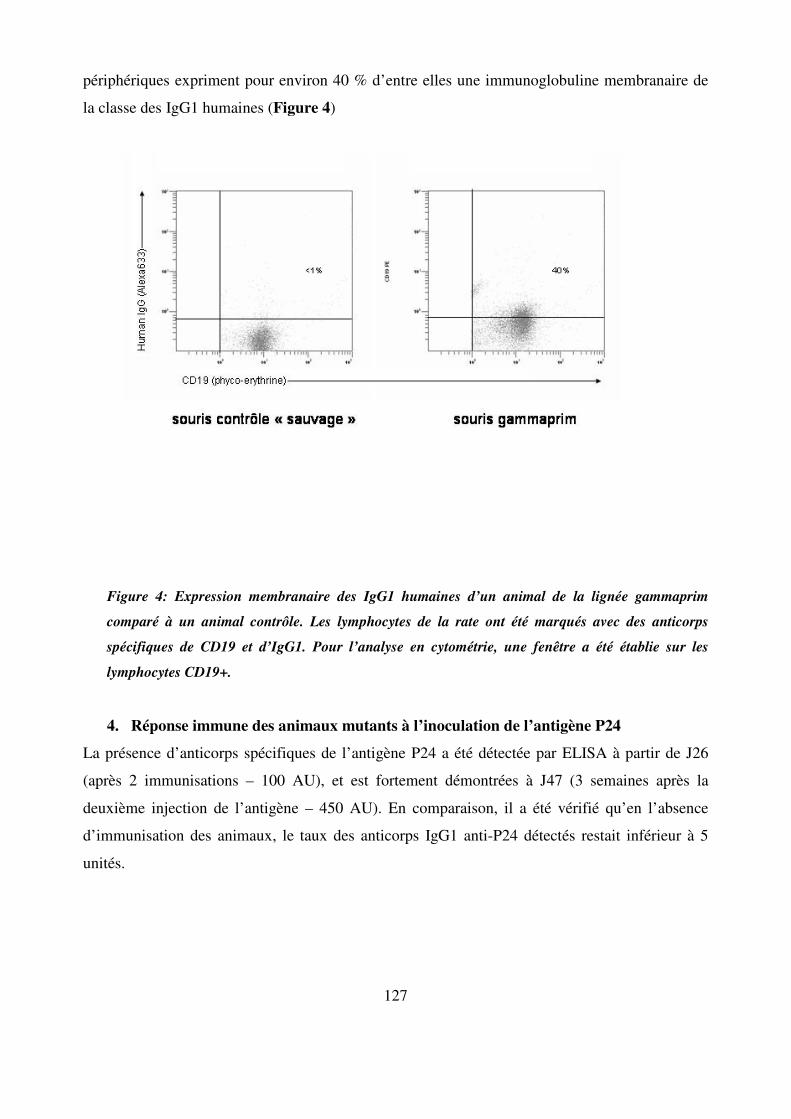
127
périphériques expriment pour environ 40 % d’entre elles une immunoglobuline membranaire de
la classe des IgG1 humaines (Figure 4)
Figure 4: Expression membranaire des IgG1 humaines d’un animal de la lignée gammaprim
comparé à un animal contrôle. Les lymphocytes de la rate ont été marqués avec des anticorps
spécifiques de CD19 et d’IgG1. Pour l’analyse en cytométrie, une fenêtre a été établie sur les
lymphocytes CD19+.
4. Réponse immune des animaux mutants à l’inoculation de l’antigène P24
La présence d’anticorps spécifiques de l’antigène P24 a été détectée par ELISA à partir de J26
(après 2 immunisations – 100 AU), et est fortement démontrées à J47 (3 semaines après la
deuxième injection de l’antigène – 450 AU). En comparaison, il a été vérifié qu’en l’absence
d’immunisation des animaux, le taux des anticorps IgG1 anti-P24 détectés restait inférieur à 5
unités.
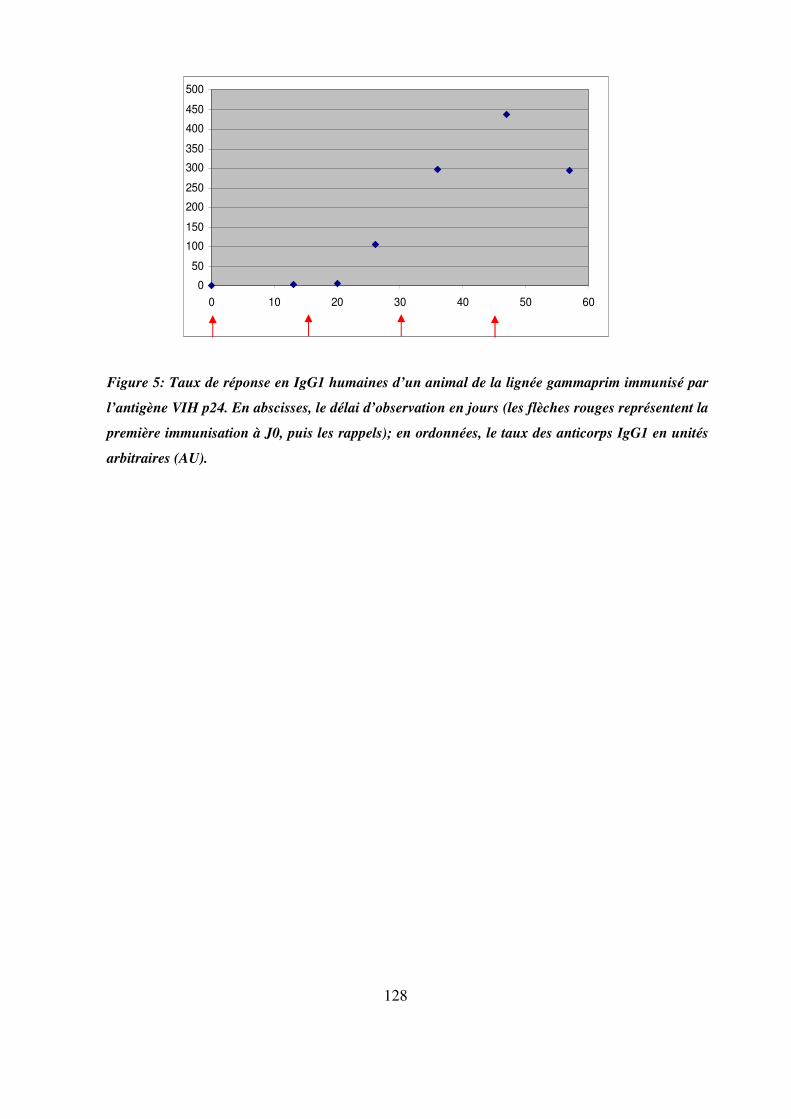
128
Figure 5: Taux de réponse en IgG1 humaines d’un animal de la lignée gammaprim immunisé par
l’antigène VIH p24. En abscisses, le délai d’observation en jours (les flèches rouges représentent la
première immunisation à J0, puis les rappels); en ordonnées, le taux des anticorps IgG1 en unités
arbitraires (AU).
0
50
100
150
200
250
300
350
400
450
500
0 10 20 30 40 50 60

129
C. CONCLUSION
Ce travail a reposé sur la réalisation et la caractérisation d’une lignée d’animaux chez lesquels la
mutation γ1-KI a été introduite dans le locus IgH murin avec l’espoir que ces animaux
produiraient des immunoglobulines sériques incluant une chaîne lourde humaine gamma1.
L’obtention de mutants homozygotes pour l’insertion γ1 dans le locus IgH permet donc d’obtenir
des souris produisant des lymphocytes B qui n’expriment que la chaîne γ1 humaine à l’exclusion
des chaînes lourdes murines.
Cette lignée « gammaprim », a été caractérisée comme capable de produire des IgG1 humanisées
au niveau de la chaîne lourde et comme capable de produire des anticorps spécifiques après
immunisation. Nous proposons donc de l’utiliser pour la production d’anticorps monoclonaux
humanisés de classe IgG1 qui pourront trouver des applications spécifiques soit en temps que
réactifs, soit en temps que moyen d’immunothérapie (anti-infectieuse ou anti-tumorale), soit
enfin en temps que système de criblage rapide d’anticorps monoclonaux ayant les propriétés
effectrices d’une IgG1 humaines dans des tests d’activité in vitro ou in vivo.

130

131
Deuxième partie : les glycosaminoglycannes

132

133
Article 3
“Modulation of glycotranscriptome and of glycosaminoglycan biogenesis genes
along B cell lineage differentiation”
Sophie Raynal-Duchez, Virginie Pascal, Nadine Cogné, Chantal Jayat-Vignoles, Virginie
Lespinet, Raymond Julien and Michel Cogné.
(Manuscrit en préparation)
Dans une autre perspective d’étude des phénomènes pouvant modifier la réactivité des récepteurs
de surface de la cellule B, nous nous sommes intéressés à la modulation de la glycosylation au
cours de la différenciation B par l’étude des variations de l’expression de gènes impliqués dans la
glycosylation, afin d’être capables par la suite, de différencier des sous-populations B
correspondant à différents états de maturation, d’activation ou de localisation.
Cette étude du glycotranscriptome, nous a notamment permis de cibler puis d’étudier d’une
manière plus approfondie deux gènes d’intérêt CsGalNacT1 et Extl1 jouant tous deux un rôle
dans la biosynthèse des glycosaminoglycannes. Ces deux gènes sont exprimés de façon plus ou
moins souvent inverse dans la maturation B. CsGalNacT1 étant absent aux stades B au repos
mais exprimé aux stades immatures et B activés alors qu’Extl1 n’est quand à lui pas exprimé aux
stades B immatures, alors qu’il est exprimé au stade mature au repos avec une expression qui
diminue aux stades B activés.
Dans le but de discerner si cette régulation d’expression présente un rôle fonctionnel ou pas, nous
avons choisi de la compromettre et de guetter ce que seraient, s’il y en a, les effets de cette
dérégulation. Alors que peu de données sont disponibles quant au rôle de ces enzymes in vivo,
nous avons donc réalisé des lignées de souris transgéniques surexprimant de façon B-spécifique
ces 2 gènes. Lors de l’amplification des ADNc respectifs de CsGalNAcT1 et Extl1 dans le but de
construire les vecteurs de transgenèse, nous avons amplifié une forme courte du transcrit
CsGalNAcT1 issue d’un épissage alternatif que nous appelons CsGalNAcT1∆. Ce transcrit,
encore inconnu, apparaît spécifique à la lignée lymphoïde (il est seulement détecté dans la moëlle
osseuse, la rate, les ganglions et le thymus) et code une protéine tronquée ayant perdu son site
catalytique et pouvant donc jouer le rôle de dominant négatif.
Ces différentes souris transgéniques CsGalNAcT1FL, Extl1 et CsGalNAcT1∆ ont récemment été
réalisées. Deux lignées de souris transgéniques issues de deux clones ES différents pour Extl1 et

134
CsGalNAcT1FL ont été obtenues alors qu’on ne compte, à ce jour, qu’une seule lignée pour
CsGalNAcT1∆. L’analyse très récente de ces animaux (CsGalNAcT1FL et Extl1) sur une seule
lignée montre certaines différences mais ces différentes pistes restent préliminaires et ne
permettent encore pas de conclure quant à l’importance de ce phénotype, sa reproducibilité, et
donc de déterminer les rôles de ces enzymes dans la lignée B.
Cet article, « en gros travaux » mérite beaucoup plus de manipulations à la paillasse afin de
mettre en évidence les rôles précis et incontestables de ces enzymes in vivo au cours de la
maturation B et donc comprendre de façon plus large, le rôle des GAGs le long de la
différenciation B, du stade B immature au stade très mature, plasmocyte-sécréteur d’anticorps.
Contribution personnelle au travail :
Ma contribution à ce travail a été :
1) de réaliser les purifications de cellules, les préparations d’ARN et de cDNA, de créer deux
« micro-fluidic cards » d’intérêt en choissant des gènes impliqués dans la glycosylation, mais
également l’analyse, le traitement et l’interprétation des données de PCR quantitative
2) d’effectuer les clonages et les constructions B-spécifiques permettant la surexpression des
gènes Extl1 et CsGalNAcT1 (FL et ∆)
3) de transfecter ces constructions dans les cellules ES et de cribler les clones positifs
4) de phénotyper les animaux.

135
Perspectives

136

137
L’état des réflexions sur les isotypes du BCR et leur fonction :
Quelqu’en soit l’isotype, la présence d’un BCR à la membrane du lymphocyte B est
indispensable à sa survie (Lam et al., 1997). La signalisation via le BCR ne se limite cependant
pas à la survie mais contrôle au contraire de multiples choix, notamment entre apoptose, anergie,
prolifération et différenciation tout au long de l’ontogénie B.
La description d’une structure univoque du BCR, associant toujours une molécule d’Ig
membranaire à l’hétérodimère Igα/Igβ, complexe de transduction du signal portant des motifs
ITAM au niveau des deux régions cytoplasmiques, pourrait donner à penser que toutes les Ig
membranaires transduisent le même type de signal « limité » à une signalisation α/β lors de leur
pontage par un ligand. Une comparaison des différentes classes de chaînes lourdes entre elles et
au cours de l’évolution, suggère cependant que la structure des trois domaines (extra-cellulaire /
transmembranaire et intra-cellulaire) de l’Ig elle-même, pourrait moduler cette signalisation. La
question du rôle spécifique des isotypes commutés du BCR reste imparfaitement résolue. C’est
une question importante car l’idée d’un rôle spécialement « affecteur » de l’IgM dans la mise en
place et la sélection initiale du répertoire (où à la fois des signaux positifs et négatifs sont
nécessaires) cadre bien avec la physiologie des cellules immatures, alors que l’idée d’un caractère
plus spécialement « effecteur » des BCR commutés apparaît comme une explication spécialement
séduisante de la dualité entre des réponses humorales primaires, basales et des réponses
secondaires, explosives aboutissant à la génération rapide de cellules sécrétrices d’anticorps de
haute affinité (Manser, 2002). De nombreuses questions méritent cependant clairement d’être
explorées plus avant.
La question du BCR « IgE » est à part, du fait de l’implication des IgE dans une
pathologie très fréquente et parfois gravissime, l’anaphylaxie. Capables de susciter de fortes
réactions immunitaires alors même qu’elles peuvent n’être présentes qu’à des concentrations
inférieures au ng/ml, les IgE spécifiques se doivent d’être contrôlées d’une façon particulièrement
stringente et aucune explication claire ne justifie à ce jour que des cellules facilement
différentiables et identifiables in vitro en présence d’IL4 (Coffman et al., 1993), ne soient par
contraste virtuellement indétectables in vivo. La nature et la localisation des cellules-mémoire à
IgE restent ainsi obscures, alors même que leur existence est indirectement attestée par la
permanence des IgE spécifiques d’allergènes dans le sérum des patients parfois même plusieurs
dizaines années après un contact sensibilisant.

138
En ce qui concerne le BCR IgE, un nombre relativement limité d’études a été publié jusqu’ici,
avec une cohérence imparfaite. Une première étude de transfectants associant la région constante
Cε à un domaine variable défini (sans tester un répertoire) a montré une fonction de l’IgE
similaire mais moins forte que celle de l’IgM dans la lignée immature WEHI-231 : en
l’occurrence une fonction pro-apoptotique en cas de pontage du BCR comme on peut l’attendre
dans une lignée pré-B ! (Batista et al., 1996). Dans une autre étude, le BCR IgE s’est révélé au
contraire (comme l’IgG) insensible à l’inhibition par CD22 et pourrait donc être ainsi considéré
de façon simpliste, comme « hyperactivateur » (Sato et al., 2007). Enfin, des constructions basées
sur des ADNc ε transfectés dans la lignée A20 en association à un seul type de domaine variable
(donc induisant un biais V potentiel) ont suggéré que la forme majeure de l’IgE membranaire
humaine (à espaceur extracellulaire long) était modérement apoptotique en cas de pontage du
BCR, alors que l’isoforme membranaire mineure (à espaceur extracellulaire court) l’était plus
nettement (Poggianella et al,. 2006). Nos résultats avec un fragment génomique entier libre de
s’épisser naturellement vont au-delà puisqu’ils révèlent un signal tonique pro-apoptotique en
l’absence même de pontage du BCR.
Même si de plus amples démonstrations sont nécessaires, l’idée qui ressort de nos travaux, dans
le contexte de la littérature précédente, est que l’IgE membranaire est plutôt inductrice de
différenciation plasmocytaire que de différenciation B mémoire, ce qui cadre bien avec le fait
qu’on ne retrouve les cellules productrices d’IgE qu’en dehors des centres germinatifs
(notamment chez des souris hyper-IgE), là où se localisent normalement les plasmocytes
récemment différenciés (Erazo et al., 2007). Nous pensons donc que l’expression d’un BCR IgE
est une étape transitoire nécessaire à la différenciation des plasmocytes à IgE (qui pourront
constituer la mémoire des réponses anti-infectieuses IgE comme la mémoire de l’anaphylaxie),
mais qu’il s’agit d’une étape forcément brève, l’IgE membranaire condamnant les cellules qui
l’expriment à un suicide rapide. Nos résultats seraient alors en accord avec ceux d’Eraso qui
identifient dans le compartiment mémoire, non pas des B à IgE membranaires, mais plutôt des B
à IgM ou IgG1 membranaire comme capables de participer à des réponses secondaires (sans
préjuger de l’existence de plasmocytes à longue durée de vie). Clairement, beaucoup est encore à
attendre de l’exploitation d’autres modèles transgéniques dans une situation où les cellules
primaires à IgE sont sans doute trop peu abondantes pour se prêter à une analyse in vivo chez
l’homme ou la souris sauvage.
Bien que largement plus explorée par de nombreux articles décrivant des souris transgéniques ou
knock-in, la question de fonction spécifique du BCR IgG n’est pas clarifiée. Les travaux des

139
équipes de Tsubata et Goodnow avaient, il y a quelques années, fait accepter l’idée d’une
fonction hyper-activatrice du BCR IgG, clairement hyper-stimulatrice de la différenciation
plasmocytaire et hors du contrôle de CD22 /SHP-1… (mais là encore, dans des modèles biaisés
par l’utilisation d’un V unique et non pas d’un répertoire diversifié). Ce modèle séduisant vient
de s’écrouler au début de l’année 2007 avec les travaux de Waisman n’identifiant ni hyper-
différenciation plasmocytaire, ni indépendance par rapport à CD22 dans des cellules B « knock-
in pour l’IgG1 murine » et cette fois capables de construire un vrai répertoire B. Le rôle de l’IgG1
dans ces travaux apparaît en fait comme quasi-identique à celui d’une IgM et pouvant lui être
substitué précocément pendant la différenciation des progéniteurs B dans la moelle osseuse. Le
modèle knock-in IgG1 humaine que nous avons décrit dans cette thèse avec une application
potentiellement plutôt biotechnologique est encore compris de façon trop préliminaire pour qu’on
puisse l’opposer aux précédents, mais il manifeste clairement une hyper-différenciation
plasmocytaire attestée par la surexpression de Blimp1 et Xbp1 et un blocage précoce de
maturation à la transition pro-B/pre-B.
Les points qui compliquent la réflexion peuvent être l’utilisation d’une région constante humaine
dans notre cas (même si elle est hautement similaire à son homologue murine dans la région
transmembranaire et intracellulaire)… on peut ainsi s’interroger quant à la bonne association de
la chaîne lourde humaine avec les chaînes légères ou pseudo-légères murines et quant au risque
d’une conformation anormale du récepteur en résultant et perturbant la signalisation en aval ?? la
question du ligand ou des ligands du préBCR reste également posée : une mauvaise association
avec la pseudo-chaîne légère pourrait empêcher l’association avec un ligand (galectine, HSPG…)
tandis que la présence prématurée d’un isotype commuté possédant un long domaine
cytoplasmique pourrait moduler les interactions avec des partenaires intra-cellulaires à préciser.
Par delà d’hypothétiques barrières d’espèces, il est cependant frappant de retrouver dans notre
modèle l’hyperdifférenciation plasmocytaire d’abord décrite avec des transgènes γ1 d’origine
murine. La diversité des phénotypes des multiples lignées transgéniques γ publiées laisse
finalement perplexe puisque certaines lignées réalisent l’exclusion allélique, d’autres pas… leur
niveau d’expression variable comme leur lien avec une seule région V peuvent être à l’origine de
biais incontournables. Enfin, il ne faut sans doute pas sous-estimer le rôle que peuvent jouer les
domaines extracellulaires CH1 à CH3/CH4 du BCR dans sa fonction en tant que récepteur de
membrane. Les travaux qui ont précédemment cherché à apprécier les différences de signalisation
intracellulaire d’IgM et d’IgG simplement en greffant à une IgM la région intracellulaire d’une
IgG ne reflètent sans doute pas la fonction d’un vrai BCR IgG complet (la différence observée à

140
cet égard, entre les formes des mIgE à espaceur extracellulaire court ou long sont
particulièrement illustratives de ce niveau supplémentaire de complexité du récepteur). De plus, il
a été démontré que les simples KO λ5, VpréB1, le double KO VpréB1/VpréB2 et le triple KO λ5/
VpréB1/VpréB2 montraient un blocage au stade préBI-préBII donc les domaines extracellulaires
jouent bien un rôle important dans la signalisation et donc l’induction de la maturation B
(Martensson et al., 1999; Mundt et al., 2001; Shimizu et al., 2002).
Enfin, la question du BCR IgA constitue un terrain encore plus vierge que celui de l’IgE. Notre
laboratoire avait montré dans le passé que les IgA membranaires humaines et en particulier
l’IgA2, s’associaient moins bien que l’IgM au co-récepteur CD19 dans des expériences de co-
modulation après pontage. Nos travaux sur les souris knock-in IgA vont dans le même sens et ne
révèlent pas de phosphorylation de CD19. Si notre modèle IgA peut souffrir de la même critique
que le modèle IgG parce qu’il est hétérologue, les éléments de réponse à cette objection se
trouvent là-aussi dans la très forte identité de séquences entre l’homme et la souris en ce qui
concerne les régions d’ancrage membranaire et intra-cellulaire de l’IgA. Ce modèle manifeste
ainsi comme notre modèle IgG1, une tendance à l’hyperdifférenciation plasmocytaire et un
blocage de maturation précoce à la transition proB/préB. Ces résultats cadrent assez bien avec ce
que l’on sait de la physiologie normale du système immunitaire humain (et murin) ou les cellules
à mIgM dominent très nettement les cellules à mIgA (que ce soit dans le sang périphérique, la
moelle, mais aussi dans les plaques de Peyer ou les amygdales), mais où pourtant, le taux de
synthèse quotidien de l’IgA soluble est très supérieur à celui de l’IgM soluble (proche pour l’IgA
voire supérieur à celui de l’IgG, tout en aboutissant à des concentrations plasmatiques inférieures
du fait de l’exportation et de la déperdition permanente au niveau des muqueuses et de la demie
vie 5 à 6 fois plus courte dans le milieu interne). Même si ces données sont très difficiles à
quantifier in vivo, il n’est cependant pas choquant pour un histologiste ou un pathologiste de
considérer que les cellules primaires à IgA observées in vivo chez le sujet sain sont plus souvent
des plasmocytes (en particulier muqueux) et que les cellules à IgM sont plus souvent des
lymphocytes à Ig de membrane. Même si les expériences basées sur des transfectants (Sato et al.,
2007) n’ont pas montré pour l’IgA l’indépendance par rapport à l’inhibition que peut médier
CD22 (comme cela a été clamé puis contesté pour mIgG), nous pensons donc que le BCR IgA est
constitutivement hyperactivateur (cf l’activation constitutive de phospho-ERK dans notre
modèle) et promoteur de différenciation plasmocytaire (d’où la facilité de réponses IgA T-
dépendantes comme T-indépendantes).

141
L’ouverture d’un nouveau champ d’étude quant à l’importance des glycosaminoglycannes
dans la physiologie de la lignée B :
Toutes les cellules vivantes portent sur leurs lipides ou leurs protéines un réseau complexe et
dense de glycannes formant ainsi des glycoprotéines ou des lipides. Localisés pour la plupart à la
surface cellulaire, les glycannes jouent un rôle fondamental dans les interactions “cellule-cellule”
et “cellule-matrice extracellulaire”, cruciales pour le développement d’organismes
multicellulaires complexes. Ils se trouvent également dans une position privilégiée pour
l’établissement d’interactions avec différents pathogènes. Ainsi, au cours des dernières décennies,
les chercheurs ont mis en évidence leur importance dans la détermination des groupes sanguins
(Kabat, 1956), leurs rôles dans les interactions cellulaires (Roseman, 1970), leurs implications
dans la fécondation (Jego et al., 1980), dans la différenciation erythrocytaire (Liu et al., 1981),
mais également dans certaines maladies où un défaut dans la machinerie de la N-glycosylation
peut conduire, chez l’Homme, à un syndrome de type CDG (Congenital Disorder of
Glycosylation) (Freeze, 2006)…
Bien que la glycosylation soit un vaste domaine, peu de choses sont encore connues dans
la maturation B. Nous savons par exemple, que les B activés des centres germinatifs deviennent
PNA high, que certaines molécules B-spécifiques, comme par exemple les lectines de la famille
des Siglecs, CD22 et récemment Siglec-G sont connues pour moduler le signal du BCR après
activation, que certains glycosaminoglycannes greffés sur des core-protéiques (syndécan comme
CD138 exprimé à la surface des plasmocytes) ou ancrés à la membrane (glypican) interviennent
dans l’adhésion cellulaire, la migration, que d’autres sont connus pour intervenir dans la
signalisation en jouant un rôle dans l’oligomérisation des récepteurs de surface (TNF R)… Il est
également connu que les différents isotypes d’Ig sont glycosylés (O et/ou N-glycosylation)
permettant ainsi le repliement et l’assemblage correct des Ig, améliorant leur transport intra-
cellulaire et les protègeant de la dégradation intracellulaire (pour revue : Wall et Kuehl, 1983).
Par des expériences d’inhibition de la glycosylation, il a été montré que la glycosylation n’était
pas nécessaire pour l’expression des Ig à la membrane des cellules B mais qu’elle était
obligatoire pour la sécrétion par les plasmocytes de certains isotypes. De plus, il est maintenant
clairement établi que les sucres présents dans la région Fc des Ig participent directement à
certaines fonctions effectrices des anticorps comme la fixation du complément, l’ADCC ou les
interactions avec les récepteurs Fc. Des anomalies de glycosylation des Ig sont également
associées à certaines pathologies telles que la polyarthrite rhumatoïde ou certaines néphropathies

142
(néphropathies à IgA, amylose AL, syndrome de Randall) (Allen et al., 1999; Preud'homme et al.,
1994; Stevens, 2000).
Dans une perspective d’étude des phénomènes pouvant modifier la réactivité des
récepteurs de surface de la cellule B, nous nous sommes intéressés à la modulation de la
glycosylation au cours de la différenciation B par l’étude des variations de l’expression de
certains gènes ciblés, impliqués dans la glycosylation, afin d’être capables par la suite, de
différencier des sous-populations B correspondant à différents états de maturation, d’activation
ou de localisation. Cette étude du glycotranscriptome au cours de la maturation B, nous a
notamment permis de cibler puis d’étudier d’une manière plus approfondie deux gènes d’intérêt
CsGalNacT1 et Extl1 jouant tous deux un rôle dans la biosynthèse des glycosaminoglycannes et
étant exprimés différemment au cours de l’ontogénie B. Dans le but de discerner si cette
régulation d’expression présente un rôle fonctionnel ou pas, nous avons choisi de la
compromettre et de regarder les effets éventuels de cette dérégulation en réalisant des lignées de
souris transgéniques qui surexpriment de façon B-spécifique ces 2 gènes. Ces résultats encore
très préliminaires nous montrent certaines pistes d’intérêts mais demandent de plus amples
manipulations. Nous nous attendons à ce que la surexpression des enzymes intervenant
directement dans la biosynthèse des GAGs (que ce soit CsGalNAct1 et Extl1) au cours de la
maturation B induise une plus forte expression de GAGs à la surface des cellules. L’identification
des glycoprotéines porteuses de ces GAGs et de leurs fonctions serait alors l’objectif majeur
suivant. Au moins en théorie, un accroissement d’expression des GAGs à la surface de nos
cellules pourrait avoir des conséquences sur la signalisation/l’oligomérisation de certains
récepteurs induisant une signalisation trop forte ou au contraire masquant des sites d’interactions
avec différents ligands. Les GAGs étant connus pour être des éponges à facteurs de croissance et
chimiokines, le fait de moduler leur présence pourrait avoir un impact sur la concentration de ces
différents facteurs au voisinage des cellules et donc avoir un impact sur la signalisation, les
interactions cellulaires… que les GAGs soient membranaires ou sécrétés. Il est déjà connu que
les GAGs joueraient un rôle dans la différenciation B précoce. Des HSPGs sécrétés par les
cellules stromales ont été mis en évidence en tant que potentiels ligands du pré-BCR chez la
souris (Bradl et al., 2003) mais une seule étude a cependant été réalisée et reste sans suite à ce
jour.
La caractérisation des patrons d’expression du « glycotranscriptome B » correspondant
aux différents états d’activation, de localisation ou de maturation B a non seulement un intérêt
fondamental, mais aussi « valorisable ». Ainsi, dans le cadre d’une collaboration avec le groupe

143
« Lymphomagenèse » dirigé par le professeur J. Feuillard au sein de notre UMR CNRS, nous
essayons de déterminer selon les patrons d’expression de différentes sous-populations B humaines,
quels sont les équivalents normaux de différents types de lymphomes B (folliculaires, LLC,
plasmocytomes) dans la perspective de mettre au point des outils diagnostiques, pronostiques et de
suivi thérapeutique. Pour l’instant une sélection de 27 gènes dont les produits auraient un rôle dans
l’établissement du microenvironnement cellulaire ont été identifiés et permettent de séparer ces trois
types de proliférations (extrèmement différentes et déjà très étudiées mais intéressantes ici pour
valider le pouvoir discriminant du glycotranscriptome). Ces différents patrons d’expression B
normaux peuvent également servir de comparaison à d’autres pathologies de l’immunité B comme la
maladie de Berger qui est une néphropathie à dépôt d’IgA (où les IgA seraient volontiers
hyposialylées et hypogalactosylées).

144

145
Annexes

146

147
Article 4
“RNA surveillance down-regulates expression of nonfunctional κ alleles and
detects premature termination within the last κ exon”
Laurent Delpy, Christophe Sirac, Emmanuelle magnoux, Sophie Raynal-Duchez and Michel
Cogné.
www.pnas.org/cgi/doi/10.1073/pnas.0305586101
(Manuscrit accepté le 22 Mars 2004 à Proceedings of the National Academy of Sciences, USA)
La recombinaison V(D)J peut induire l’addition ou la délétion de nucléotides de façon aléatoire.
Dans certains cas, ces réarrangements génèrent un décalage du cadre de lecture pouvant induire la
présence de codons stop prématurés (PTC pour Premature Termination Codon) et par
conséquence, la possibilité de produire une chaîne d’immunoglobuline tronquée. Bien qu’une
forte proportion de lymphocytes matures périphériques possède un allèle non fonctionnel aux loci
IgH et/ou IgL, l’expression de chaînes tronquées n’est pas observée dans les cellules B normales.
Laurent Delpy et Christophe Sirac ont voulu montré que les transcrits de chaînes légères κ
possédant des décalages du cadre de lecture provoqués par les recombinaisons V-J (et induisant
ainsi des mutations non-sens) étaient sujets à un processus de surveillance des ARN pouvant
engendrer leur élimination ou l’inhibition de leur épissage et ainsi, permettre le maintien de
l’exclusion allélique des chaînes légères dans des cellules possédant deux allèles réarrangés dont
l’un « hors-phase ».
Contribution personnelle à ce travail :
Ce travail, initié lors de mon stage de maîtrise et achevé au début de ma première année de thèse
a consisté en l’amplification par PCR des jonctions Vκ-Jκ, leur clonage dans des vecteurs PCRII-
TOPO puis le séquençage et l’analyse de ces différents produits de PCR.

148

149
Article 5
“Light chain inclusion permits terminal B cell differentiation and does not
necessarily result in autoreactivity”
Christophe Sirac, Claire Carrion*, Sophie Raynal-Duchez*, Isabelle Comte and Michel Cogné.
(* contribution égale)
www.pnas.org/cgi/doi/10.1073/pnas.0509121103
(Manuscrit accepté le 23 Mars 2006 à Proceedings of the National Academy of Sciences, USA)
Lors du développement des cellules B, l’assemblage du BCR se fait normalement selon un
processus ordonné de réarrangement (détaillé dans le chapitre II) aboutissant à ce que chaque
cellule B ne possède qu’un seul type de récepteur à sa surface grâce au phénomène d’exclusion
allélique, essentiel dans la génération du répertoire B. Cependant, plusieurs travaux ont démontré
l’existence d’un faible taux de cellules B périphériques exprimant deux chaînes lourdes distinctes
(Barreto et Cumano, 2000) ou, plus fréquemment, deux chaînes légères, une κ et une λ (Giachino
et coll., 1995). Dans ce travail, nous avons étudié les effets de l’insertion génique d’un gène VκJκ
humain réarrangé en lieu et place des domaines Jκ du locus endogène de la souris. Cette étude a
démontré l’apparition de cellules « inclues » dans les animaux hétérozygotes et homozygotes
pour le transgène, c’est-à-dire exprimant une chaîne légère λ endogène et la chaîne chimérique κ.
Ces cellules participent normalement aux réponses immunitaires et peuvent se différencier en
cellules B mémoires commutées ou en plasmocytes sécréteurs d’anticorps hybrides κ et λ non-
autoréactifs. Cela indique que l’exclusion allélique des chaînes légères n’est pas un pré-requis au
développement de cellules B normales et que la sélection négative des BCR auto-réactifs peut
s’effectuer même sous des conditions d’inclusion.
Contribution personnelle à ce travail :
J’ai essentiellement participé à cette étude lors des révisions demandées pour l’acceptation de ce
papier alors que Christophe avait quitté le laboratoire pour son stage post-doctoral. Mon travail a
en particulier consisté en la mise au point de RT-PCR Quantitative–sybrGreen sur des
échantillons d’ARN de cellules B matures que j’ai préalablement triées afin de quantifier
l’expression des transcrits κ et µ dans les souris hétérozygotes et homozygotes pour la mutation.

150

151
Références Bibliographiques

152

153
Abney, E. R., Cooper, M. D., Kearney, J. F., Lawton, A. R. and Parkhouse, R. M. (1978). Sequential expression of immunoglobulin on developing mouse B lymphocytes: a systematic survey that suggests a model for the generation of immunoglobulin isotype diversity. J
Immunol 120, 2041-9. Achatz, G., Nitschke, L. and Lamers, M. C. (1997). Effect of transmembrane and cytoplasmic domains of IgE on the IgE response. Science 276, 409-11. Afshar, R., Pierce, S., Bolland, D. J., Corcoran, A. and Oltz, E. M. (2006). Regulation of IgH gene assembly: role of the intronic enhancer and 5'DQ52 region in targeting DHJH recombination. J Immunol 176, 2439-47. Ahnesorg, P., Smith, P. and Jackson, S. P. (2006). XLF interacts with the XRCC4-DNA ligase IV complex to promote DNA nonhomologous end-joining. Cell 124, 301-13. Alessandrini, A. and Desiderio, S. V. (1991). Coordination of immunoglobulin DJH transcription and D-to-JH rearrangement by promoter-enhancer approximation. Mol Cell Biol 11, 2096-107. Allen, A. C., Bailey, E. M., Barratt, J., Buck, K. S. and Feehally, J. (1999). Analysis of IgA1 O-glycans in IgA nephropathy by fluorophore-assisted carbohydrate electrophoresis. J
Am Soc Nephrol 10, 1763-71. Allman, D., Lindsley, R. C., DeMuth, W., Rudd, K., Shinton, S. A. and Hardy, R. R. (2001). Resolution of three nonproliferative immature splenic B cell subsets reveals multiple selection points during peripheral B cell maturation. J Immunol 167, 6834-40. Alonso, L. and Fuchs, E. (2003). Stem cells in the skin: waste not, Wnt not. Genes Dev 17, 1189-200. Alt, F. W. and Baltimore, D. (1982). Joining of immunoglobulin heavy chain gene segments: implications from a chromosome with evidence of three D-JH fusions. Proc Natl Acad
Sci U S A 79, 4118-22. Alt, F. W., Yancopoulos, G. D., Blackwell, T. K., Wood, C., Thomas, E., Boss, M., Coffman, R., Rosenberg, N., Tonegawa, S. and Baltimore, D. (1984). Ordered rearrangement of immunoglobulin heavy chain variable region segments. Embo J 3, 1209-19. Amano, M., Galvan, M., He, J. and Baum, L. G. (2003). The ST6Gal I sialyltransferase selectively modifies N-glycans on CD45 to negatively regulate galectin-1-induced CD45 clustering, phosphatase modulation, and T cell death. J Biol Chem 278, 7469-75. Arnold, L. W., McCray, S. K., Tatu, C. and Clarke, S. H. (2000). Identification of a precursor to phosphatidyl choline-specific B-1 cells suggesting that B-1 cells differentiate from splenic conventional B cells in vivo: cyclosporin A blocks differentiation to B-1. J Immunol 164, 2924-30. Artavanis-Tsakonas, S., Rand, M. D. and Lake, R. J. (1999). Notch signaling: cell fate control and signal integration in development. Science 284, 770-6. Ashikari, S., Habuchi, H. and Kimata, K. (1995). Characterization of heparan sulfate oligosaccharides that bind to hepatocyte growth factor. J Biol Chem 270, 29586-93. Bain, G., Maandag, E. C., Izon, D. J., Amsen, D., Kruisbeek, A. M., Weintraub, B. C., Krop, I., Schlissel, M. S., Feeney, A. J., van Roon, M. et al. (1994). E2A proteins are required for proper B cell development and initiation of immunoglobulin gene rearrangements. Cell 79, 885-92. Bandtlow, C. E. and Zimmermann, D. R. (2000). Proteoglycans in the developing brain: new conceptual insights for old proteins. Physiol Rev 80, 1267-90. Banerji, J., Olson, L. and Schaffner, W. (1983). A lymphocyte-specific cellular enhancer is located downstream of the joining region in immunoglobulin heavy chain genes. Cell 33, 729-40.

154
Bangs, L. A., Sanz, I. E. and Teale, J. M. (1991). Comparison of D, JH, and junctional diversity in the fetal, adult, and aged B cell repertoires. J Immunol 146, 1996-2004. Bassing, C. H., Alt, F. W., Hughes, M. M., D'Auteuil, M., Wehrly, T. D., Woodman, B. B., Gartner, F., White, J. M., Davidson, L. and Sleckman, B. P. (2000). Recombination signal sequences restrict chromosomal V(D)J recombination beyond the 12/23 rule. Nature 405, 583-6. Bassing, C. H., Swat, W. and Alt, F. W. (2002). The mechanism and regulation of chromosomal V(D)J recombination. Cell 109 Suppl, S45-55. Basu, U., Chaudhuri, J., Alpert, C., Dutt, S., Ranganath, S., Li, G., Schrum, J. P., Manis, J. P. and Alt, F. W. (2005). The AID antibody diversification enzyme is regulated by protein kinase A phosphorylation. Nature 438, 508-11. Batista, F. D., Anand, S., Presani, G., Efremov, D. G. and Burrone, O. R. (1996). The two membrane isoforms of human IgE assemble into functionally distinct B cell antigen receptors. J Exp Med 184, 2197-205. Beckwith, M., Fenton, R. G., Katona, I. M. and Longo, D. L. (1996). Phosphatidylinositol-3-kinase activity is required for the anti-ig-mediated growth inhibition of a human B-lymphoma cell line. Blood 87, 202-10. Bensmana, M. and Lefranc, M. P. (1990). Gene segments encoding membrane domains of the human immunoglobulin gamma 3 and alpha chains. Immunogenetics 32, 321-30. Benson, M. J., Erickson, L. D., Gleeson, M. W. and Noelle, R. J. (2007). Affinity of antigen encounter and other early B-cell signals determine B-cell fate. Curr Opin Immunol 19, 275-80. Berek, C., Berger, A. and Apel, M. (1991). Maturation of the immune response in germinal centers. Cell 67, 1121-9. Berek, C. and Milstein, C. (1988). The dynamic nature of the antibody repertoire. Immunol Rev 105, 5-26. Berg, J., McDowell, M., Jack, H. M. and Wabl, M. (1990). Immunoglobulin lambda gene rearrangement can precede kappa gene rearrangement. Dev Immunol 1, 53-7. Berton, M. T. and Vitetta, E. S. (1990). Interleukin 4 induces changes in the chromatin structure of the gamma 1 switch region in resting B cells before switch recombination. J Exp Med 172, 375-8. Bottaro, A., Lansford, R., Xu, L., Zhang, J., Rothman, P. and Alt, F. W. (1994). S region transcription per se promotes basal IgE class switch recombination but additional factors regulate the efficiency of the process. Embo J 13, 665-74. Bourin, M. C. and Lindahl, U. (1993). Glycosaminoglycans and the regulation of blood coagulation. Biochem J 289 (Pt 2), 313-30. Boyd, P. N., Lines, A. C. and Patel, A. K. (1995). The effect of the removal of sialic acid, galactose and total carbohydrate on the functional activity of Campath-1H. Mol Immunol 32, 1311-8. Bradl, H., Wittmann, J., Milius, D., Vettermann, C. and Jack, H. M. (2003). Interaction of murine precursor B cell receptor with stroma cells is controlled by the unique tail of lambda 5 and stroma cell-associated heparan sulfate. J Immunol 171, 2338-48. Bransteitter, R., Pham, P., Scharff, M. D. and Goodman, M. F. (2003). Activation-induced cytidine deaminase deaminates deoxycytidine on single-stranded DNA but requires the action of RNase. Proc Natl Acad Sci U S A 100, 4102-7. Brauninger, A., Goossens, T., Rajewsky, K. and Kuppers, R. (2001). Regulation of immunoglobulin light chain gene rearrangements during early B cell development in the human. Eur J Immunol 31, 3631-7.

155
Brink, R., Goodnow, C. C., Crosbie, J., Adams, E., Eris, J., Mason, D. Y., Hartley, S. B. and Basten, A. (1992). Immunoglobulin M and D antigen receptors are both capable of mediating B lymphocyte activation, deletion, or anergy after interaction with specific antigen. J
Exp Med 176, 991-1005. Brockhausen, I., Reck, F., Kuhns, W., Khan, S., Matta, K. L., Meinjohanns, E., Paulsen, H., Shah, R. N., Baker, M. A. and Schachter, H. (1995). Substrate specificity and inhibition of UDP-GlcNAc:GlcNAc beta 1-2Man alpha 1-6R beta 1,6-N-acetylglucosaminyltransferase V using synthetic substrate analogues. Glycoconj J 12, 371-9. Bruce, S. R., Dingle, R. W. and Peterson, M. L. (2003). B-cell and plasma-cell splicing differences: a potential role in regulated immunoglobulin RNA processing. Rna 9, 1264-73. Buck, D., Malivert, L., de Chasseval, R., Barraud, A., Fondaneche, M. C., Sanal, O., Plebani, A., Stephan, J. L., Hufnagel, M., le Deist, F. et al. (2006). Cernunnos, a novel nonhomologous end-joining factor, is mutated in human immunodeficiency with microcephaly. Cell 124, 287-99. Buhl, A. M. and Cambier, J. C. (1999). Phosphorylation of CD19 Y484 and Y515, and linked activation of phosphatidylinositol 3-kinase, are required for B cell antigen receptor-mediated activation of Bruton's tyrosine kinase. J Immunol 162, 4438-46. Burkhardt, A. L., Brunswick, M., Bolen, J. B. and Mond, J. J. (1991). Anti-immunoglobulin stimulation of B lymphocytes activates src-related protein-tyrosine kinases. Proc Natl Acad Sci U S A 88, 7410-4. Calame, K. L., Lin, K. I. and Tunyaplin, C. (2003). Regulatory mechanisms that determine the development and function of plasma cells. Annu Rev Immunol 21, 205-30. Cambier, J. C., Newell, M. K., Justement, L. B., McGuire, J. C., Leach, K. L. and Chen, Z. Z. (1987). Ia binding ligands and cAMP stimulate nuclear translocation of PKC in B lymphocytes. Nature 327, 629-32. Cariappa, A. and Pillai, S. (2002). Antigen-dependent B-cell development. Curr Opin
Immunol 14, 241-9. Carter, P. J. (2006). Potent antibody therapeutics by design. Nat Rev Immunol 6, 343-57. Cartron, G., Dacheux, L., Salles, G., Solal-Celigny, P., Bardos, P., Colombat, P. and Watier, H. (2002). Therapeutic activity of humanized anti-CD20 monoclonal antibody and polymorphism in IgG Fc receptor FcgammaRIIIa gene. Blood 99, 754-8. Casola, S. (2007). Control of peripheral B-cell development. Curr Opin Immunol 19, 143-9. Casola, S., Otipoby, K. L., Alimzhanov, M., Humme, S., Uyttersprot, N., Kutok, J. L., Carroll, M. C. and Rajewsky, K. (2004). B cell receptor signal strength determines B cell fate. Nat Immunol 5, 317-27. Chan, A. C., Iwashima, M., Turck, C. W. and Weiss, A. (1992). ZAP-70: a 70 kd protein-tyrosine kinase that associates with the TCR zeta chain. Cell 71, 649-62. Chan, V. W., Meng, F., Soriano, P., DeFranco, A. L. and Lowell, C. A. (1997). Characterization of the B lymphocyte populations in Lyn-deficient mice and the role of Lyn in signal initiation and down-regulation. Immunity 7, 69-81. Chaudhuri, J. and Alt, F. W. (2004). Class-switch recombination: interplay of transcription, DNA deamination and DNA repair. Nat Rev Immunol 4, 541-52. Chaudhuri, J., Khuong, C. and Alt, F. W. (2004). Replication protein A interacts with AID to promote deamination of somatic hypermutation targets. Nature 430, 992-8. Chaudhuri, J., Tian, M., Khuong, C., Chua, K., Pinaud, E. and Alt, F. W. (2003). Transcription-targeted DNA deamination by the AID antibody diversification enzyme. Nature 422, 726-30.

156
Chauveau, C., Decourt, C. and Cogne, M. (1998). Insertion of the IgH locus 3' regulatory palindrome in expression vectors warrants sure and efficient expression in stable B cell transfectants. Gene 222, 279-85. Chauveau, C., Jansson, E. A., Muller, S., Cogne, M. and Pettersson, S. (1999). Cutting edge: Ig heavy chain 3' HS1-4 directs correct spatial position-independent expression of a linked transgene to B lineage cells. J Immunol 163, 4637-41. Chen, H. T., Bhandoola, A., Difilippantonio, M. J., Zhu, J., Brown, M. J., Tai, X., Rogakou, E. P., Brotz, T. M., Bonner, W. M., Ried, T. et al. (2000). Response to RAG-mediated VDJ cleavage by NBS1 and gamma-H2AX. Science 290, 1962-5. Chen, J., Lu, L., Shi, S. and Stanley, P. (2006). Expression of Notch signaling pathway genes in mouse embryos lacking beta4galactosyltransferase-1. Gene Expr Patterns 6, 376-82. Chen, J., Moloney, D. J. and Stanley, P. (2001a). Fringe modulation of Jagged1-induced Notch signaling requires the action of beta 4galactosyltransferase-1. Proc Natl Acad Sci
U S A 98, 13716-21. Chen, J., Trounstine, M., Kurahara, C., Young, F., Kuo, C. C., Xu, Y., Loring, J. F., Alt, F. W. and Huszar, D. (1993). B cell development in mice that lack one or both immunoglobulin kappa light chain genes. Embo J 12, 821-30. Chen, J. Z., Stall, A. M., Herzenberg, L. A. and Herzenberg, L. A. (1990). Differences in glycoprotein complexes associated with IgM and IgD on normal murine B cells potentially enable transduction of different signals. Embo J 9, 2117-24. Chen, X., Kinoshita, K. and Honjo, T. (2001b). Variable deletion and duplication at recombination junction ends: implication for staggered double-strand cleavage in class-switch recombination. Proc Natl Acad Sci U S A 98, 13860-5. Cheng, A. M., Rowley, B., Pao, W., Hayday, A., Bolen, J. B. and Pawson, T. (1995). Syk tyrosine kinase required for mouse viability and B-cell development. Nature 378, 303-6. Cheng, P. C., Dykstra, M. L., Mitchell, R. N. and Pierce, S. K. (1999). A role for lipid rafts in B cell antigen receptor signaling and antigen targeting. J Exp Med 190, 1549-60. Cherukuri, A., Cheng, P. C., Sohn, H. W. and Pierce, S. K. (2001). The CD19/CD21 complex functions to prolong B cell antigen receptor signaling from lipid rafts. Immunity 14, 169-79. Choquet, D., Ku, G., Cassard, S., Malissen, B., Korn, H., Fridman, W. H. and Bonnerot, C. (1994). Different patterns of calcium signaling triggered through two components of the B lymphocyte antigen receptor. J Biol Chem 269, 6491-7. Chumley, M. J., Dal Porto, J. M., Kawaguchi, S., Cambier, J. C., Nemazee, D. and Hardy, R. R. (2000). A VH11V kappa 9 B cell antigen receptor drives generation of CD5+ B cells both in vivo and in vitro. J Immunol 164, 4586-93. Chung, J. B., Sater, R. A., Fields, M. L., Erikson, J. and Monroe, J. G. (2002). CD23 defines two distinct subsets of immature B cells which differ in their responses to T cell help signals. Int Immunol 14, 157-66. Chung, J. B., Silverman, M. and Monroe, J. G. (2003). Transitional B cells: step by step towards immune competence. Trends Immunol 24, 343-9. Clatworthy, A. E., Valencia-Burton, M. A., Haber, J. E. and Oettinger, M. A. (2005). The MRE11-RAD50-XRS2 complex, in addition to other non-homologous end-joining factors, is required for V(D)J joining in yeast. J Biol Chem 280, 20247-52. Cobaleda, C., Schebesta, A., Delogu, A. and Busslinger, M. (2007). Pax5: the guardian of B cell identity and function. Nat Immunol 8, 463-70. Cobb, R. M., Oestreich, K. J., Osipovich, O. A. and Oltz, E. M. (2006). Accessibility control of V(D)J recombination. Adv Immunol 91, 45-109.

157
Cogne, M., Lansford, R., Bottaro, A., Zhang, J., Gorman, J., Young, F., Cheng, H. L. and Alt, F. W. (1994). A class switch control region at the 3' end of the immunoglobulin heavy chain locus. Cell 77, 737-47. Coleclough, C., Perry, R. P., Karjalainen, K. and Weigert, M. (1981). Aberrant rearrangements contribute significantly to the allelic exclusion of immunoglobulin gene expression. Nature 290, 372-8. Collins, B. E., Blixt, O., Bovin, N. V., Danzer, C. P., Chui, D., Marth, J. D., Nitschke, L. and Paulson, J. C. (2002). Constitutively unmasked CD22 on B cells of ST6Gal I knockout mice: novel sialoside probe for murine CD22. Glycobiology 12, 563-71. Collins, B. E., Blixt, O., Han, S., Duong, B., Li, H., Nathan, J. K., Bovin, N. and Paulson, J. C. (2006a). High-affinity ligand probes of CD22 overcome the threshold set by cis ligands to allow for binding, endocytosis, and killing of B cells. J Immunol 177, 2994-3003. Collins, B. E., Smith, B. A., Bengtson, P. and Paulson, J. C. (2006b). Ablation of CD22 in ligand-deficient mice restores B cell receptor signaling. Nat Immunol 7, 199-206. Coombe, D. R. and Kett, W. C. (2005). Heparan sulfate-protein interactions: therapeutic potential through structure-function insights. Cell Mol Life Sci 62, 410-24. Corcos, D., Dunda, O., Butor, C., Cesbron, J. Y., Lores, P., Bucchini, D. and Jami, J. (1995). Pre-B-cell development in the absence of lambda 5 in transgenic mice expressing a heavy-chain disease protein. Curr Biol 5, 1140-8. Corcos, D., Iglesias, A., Dunda, O., Bucchini, D. and Jami, J. (1991). Allelic exclusion in transgenic mice expressing a heavy chain disease-like human mu protein. Eur J Immunol 21, 2711-6. Cornall, R. J., Cheng, A. M., Pawson, T. and Goodnow, C. C. (2000). Role of Syk in B-cell development and antigen-receptor signaling. Proc Natl Acad Sci U S A 97, 1713-8. Coutinho, A., Kazatchkine, M. D. and Avrameas, S. (1995). Natural autoantibodies. Curr Opin Immunol 7, 812-8. Dammers, P. M., Visser, A., Popa, E. R., Nieuwenhuis, P. and Kroese, F. G. (2000). Most marginal zone B cells in rat express germline encoded Ig VH genes and are ligand selected. J Immunol 165, 6156-69. Danner, D. and Leder, P. (1985). Role of an RNA cleavage/poly(A) addition site in the production of membrane-bound and secreted IgM mRNA. Proc Natl Acad Sci U S A 82, 8658-62. de Villartay, J. P., Poinsignon, C., de Chasseval, R., Buck, D., Le Guyader, G. and Villey, I. (2003). Human and animal models of V(D)J recombination deficiency. Curr Opin
Immunol 15, 592-8. DeFranco, A. L. (1997). The complexity of signaling pathways activated by the BCR. Curr Opin Immunol 9, 296-308. del Peso, L., Gonzalez-Garcia, M., Page, C., Herrera, R. and Nunez, G. (1997). Interleukin-3-induced phosphorylation of BAD through the protein kinase Akt. Science 278, 687-9. Delphin, S. and Stavnezer, J. (1995). Regulation of antibody class switching to IgE: characterization of an IL-4-responsive region in the immunoglobulin heavy-chain germline epsilon promoter. Ann N Y Acad Sci 764, 123-35. Delpy, L., Le Bert, M., Cogne, M. and Khamlichi, A. A. (2003). Germ-line transcription occurs on both the functional and the non-functional alleles of immunoglobulin constant heavy chain genes. Eur J Immunol 33, 2108-13. Demetriou, M., Granovsky, M., Quaggin, S. and Dennis, J. W. (2001). Negative regulation of T-cell activation and autoimmunity by Mgat5 N-glycosylation. Nature 409, 733-9.

158
Dempsey, L. A., Sun, H., Hanakahi, L. A. and Maizels, N. (1999). G4 DNA binding by LR1 and its subunits, nucleolin and hnRNP D, A role for G-G pairing in immunoglobulin switch recombination. J Biol Chem 274, 1066-71. Denis, K. A., Provost, S., Witte, O. N., Brinster, R. L. and Storb, U. (1990). Delay of early B-lymphocyte development by gamma 2b immunoglobulin transgene: effect on differentiation-specific molecules. Dev Immunol 1, 105-12. Dickerson, S. K., Market, E., Besmer, E. and Papavasiliou, F. N. (2003). AID mediates hypermutation by deaminating single stranded DNA. J Exp Med 197, 1291-6. Drouet, J., Delteil, C., Lefrancois, J., Concannon, P., Salles, B. and Calsou, P. (2005). DNA-dependent protein kinase and XRCC4-DNA ligase IV mobilization in the cell in response to DNA double strand breaks. J Biol Chem 280, 7060-9. Dudley, D. D., Chaudhuri, J., Bassing, C. H. and Alt, F. W. (2005). Mechanism and control of V(D)J recombination versus class switch recombination: similarities and differences. Adv Immunol 86, 43-112. Dunnick, W., Hertz, G. Z., Scappino, L. and Gritzmacher, C. (1993). DNA sequences at immunoglobulin switch region recombination sites. Nucleic Acids Res 21, 365-72. Early, P., Rogers, J., Davis, M., Calame, K., Bond, M., Wall, R. and Hood, L. (1980). Two mRNAs can be produced from a single immunoglobulin mu gene by alternative RNA processing pathways. Cell 20, 313-9. Eaton, S. and Calame, K. (1987). Multiple DNA sequence elements are necessary for the function of an immunoglobulin heavy chain promoter. Proc Natl Acad Sci U S A 84, 7634-8. Ehlich, A., Schaal, S., Gu, H., Kitamura, D., Muller, W. and Rajewsky, K. (1993). Immunoglobulin heavy and light chain genes rearrange independently at early stages of B cell development. Cell 72, 695-704.
Engel, H., Rolink, A. and Weiss, S. (1999). B cells are programmed to activate kappa and lambda for rearrangement at consecutive developmental stages. Eur J Immunol, 29, 2167-2176. Engler, P., Roth, P., Kim, J. Y. and Storb, U. (1991). Factors affecting the rearrangement efficiency of an Ig test gene. J Immunol 146, 2826-35. Erazo, A., Kutchukhidze, N., Leung, M., Christ, A. P., Urban, J. F., Jr., Curotto de Lafaille, M. A. and Lafaille, J. J. (2007). Unique maturation program of the IgE response in vivo. Immunity 26, 191-203. Ernst, P. and Smale, S. T. (1995). Combinatorial regulation of transcription. I: General aspects of transcriptional control. Immunity 2, 311-9. Esko, J. D. and Zhang, L. (1996). Influence of core protein sequence on glycosaminoglycan assembly. Curr Opin Struct Biol 6, 663-70. Falkner, F. G. and Zachau, H. G. (1984). Correct transcription of an immunoglobulin kappa gene requires an upstream fragment containing conserved sequence elements. Nature 310, 71-4. Fearon, D. T. and Carter, R. H. (1995). The CD19/CR2/TAPA-1 complex of B lymphocytes: linking natural to acquired immunity. Annu Rev Immunol 13, 127-49. Felsenfeld, G. (1992). Chromatin as an essential part of the transcriptional mechanism. Nature 355, 219-24. Fong, D. C., Brauweiler, A., Minskoff, S. A., Bruhns, P., Tamir, I., Mellman, I., Daeron, M. and Cambier, J. C. (2000). Mutational analysis reveals multiple distinct sites within Fc gamma receptor IIB that function in inhibitory signaling. J Immunol 165, 4453-62. Forrester, W. C., van Genderen, C., Jenuwein, T. and Grosschedl, R. (1994). Dependence of enhancer-mediated transcription of the immunoglobulin mu gene on nuclear matrix attachment regions. Science 265, 1221-5.

159
Franke, T. F., Kaplan, D. R., Cantley, L. C. and Toker, A. (1997). Direct regulation of the Akt proto-oncogene product by phosphatidylinositol-3,4-bisphosphate. Science 275, 665-8. Freeze, H. H. (2006). Genetic defects in the human glycome. Nat Rev Genet 7, 537-51. Freeze, H. H. and Aebi, M. (2005). Altered glycan structures: the molecular basis of congenital disorders of glycosylation. Curr Opin Struct Biol 15, 490-8. Fruman, D. A., Snapper, S. B., Yballe, C. M., Alt, F. W. and Cantley, L. C. (1999). Phosphoinositide 3-kinase knockout mice: role of p85alpha in B cell development and proliferation. Biochem Soc Trans 27, 624-9. Fukumori, T., Takenaka, Y., Yoshii, T., Kim, H. R., Hogan, V., Inohara, H., Kagawa, S. and Raz, A. (2003). CD29 and CD7 mediate galectin-3-induced type II T-cell apoptosis. Cancer Res 63, 8302-11. Fuxa, M., Skok, J., Souabni, A., Salvagiotto, G., Roldan, E. and Busslinger, M. (2004). Pax5 induces V-to-DJ rearrangements and locus contraction of the immunoglobulin heavy-chain gene. Genes Dev 18, 411-22. Fuxa, M. and Skok, J. A. (2007). Transcriptional regulation in early B cell development. Curr Opin Immunol 19, 129-36. Galceran, J., Farinas, I., Depew, M. J., Clevers, H. and Grosschedl, R. (1999). Wnt3a-/--like phenotype and limb deficiency in Lef1(-/-)Tcf1(-/-) mice. Genes Dev 13, 709-17. Gallagher, J. T., Lyon, M. and Steward, W. P. (1986). Structure and function of heparan sulphate proteoglycans. Biochem J 236, 313-25. Galvan, M., Tsuboi, S., Fukuda, M. and Baum, L. G. (2000). Expression of a specific glycosyltransferase enzyme regulates T cell death mediated by galectin-1. J Biol Chem 275, 16730-7. Gauthier, L., Rossi, B., Roux, F., Termine, E. and Schiff, C. (2002). Galectin-1 is a stromal cell ligand of the pre-B cell receptor (BCR) implicated in synapse formation between pre-B and stromal cells and in pre-BCR triggering. Proc Natl Acad Sci U S A 99, 13014-9. Gavel, Y. and von Heijne, G. (1990). Sequence differences between glycosylated and non-glycosylated Asn-X-Thr/Ser acceptor sites: implications for protein engineering. Protein Eng 3, 433-42. Gay, D., Saunders, T., Camper, S. and Weigert, M. (1993). Receptor editing: an approach by autoreactive B cells to escape tolerance. J Exp Med 177, 999-1008. Geha, R. S., Jabara, H. H. and Brodeur, S. R. (2003). The regulation of immunoglobulin E class-switch recombination. Nat Rev Immunol 3, 721-32. Geisow, M. J. (1991). The coming of age of glycobiology. Trends Biotechnol 9, 221-5. Gellert, M. (2002). V(D)J recombination: RAG proteins, repair factors, and regulation. Annu Rev Biochem 71, 101-32. Giannini, S. L., Singh, M., Calvo, C. F., Ding, G. and Birshtein, B. K. (1993). DNA regions flanking the mouse Ig 3' alpha enhancer are differentially methylated and DNAase I hypersensitive during B cell differentiation. J Immunol 150, 1772-80. Gold, M. R., Sanghera, J. S., Stewart, J. and Pelech, S. L. (1992). Selective activation of p42 mitogen-activated protein (MAP) kinase in murine B lymphoma cell lines by membrane immunoglobulin cross-linking. Evidence for protein kinase C-independent and -dependent mechanisms of activation. Biochem J 287 (Pt 1), 269-76. Goodhardt, M., Cavelier, P., Doyen, N., Kallenbach, S., Babinet, C. and Rougeon, F. (1993). Methylation status of immunoglobulin kappa gene segments correlates with their recombination potential. Eur J Immunol 23, 1789-95. Goodman, D. J., Gaff, C. and Gerondakis, S. (1993). The IL-4 induced increase in the frequency of resting murine splenic B cells expressing germline Ig heavy chain gamma 1 transcripts correlates with subsequent switching to IgG1. Int Immunol 5, 199-208.

160
Goodnow, C. C., Crosbie, J., Adelstein, S., Lavoie, T. B., Smith-Gill, S. J., Brink, R. A., Pritchard-Briscoe, H., Wotherspoon, J. S., Loblay, R. H., Raphael, K. et al. (1988). Altered immunoglobulin expression and functional silencing of self-reactive B lymphocytes in transgenic mice. Nature 334, 676-82. Gu, M., Wang, W., Song, W. K., Cooper, D. N. and Kaufman, S. J. (1994). Selective modulation of the interaction of alpha 7 beta 1 integrin with fibronectin and laminin by L-14 lectin during skeletal muscle differentiation. J Cell Sci 107 (Pt 1), 175-81. Guerrini, M., Agulles, T., Bisio, A., Hricovini, M., Lay, L., Naggi, A., Poletti, L., Sturiale, L., Torri, G. and Casu, B. (2002). Minimal heparin/heparan sulfate sequences for binding to fibroblast growth factor-1. Biochem Biophys Res Commun 292, 222-30. Guo, B., Kato, R. M., Garcia-Lloret, M., Wahl, M. I. and Rawlings, D. J. (2000). Engagement of the human pre-B cell receptor generates a lipid raft-dependent calcium signaling complex. Immunity 13, 243-53. Gupta-Bansal, R., Frederickson, R. C. and Brunden, K. R. (1995). Proteoglycan-mediated inhibition of A beta proteolysis. A potential cause of senile plaque accumulation. J Biol
Chem 270, 18666-71. Habas, R., Dawid, I. B. and He, X. (2003). Coactivation of Rac and Rho by Wnt/Frizzled signaling is required for vertebrate gastrulation. Genes Dev 17, 295-309. Hacker, H., Redecke, V., Blagoev, B., Kratchmarova, I., Hsu, L. C., Wang, G. G., Kamps, M. P., Raz, E., Wagner, H., Hacker, G. et al. (2006). Specificity in Toll-like receptor signalling through distinct effector functions of TRAF3 and TRAF6. Nature 439, 204-7. Hadari, Y. R., Arbel-Goren, R., Levy, Y., Amsterdam, A., Alon, R., Zakut, R. and Zick, Y. (2000). Galectin-8 binding to integrins inhibits cell adhesion and induces apoptosis. J
Cell Sci 113 (Pt 13), 2385-97. Haltiwanger, R. S. and Lowe, J. B. (2004). Role of glycosylation in development. Annu
Rev Biochem 73, 491-537. Han, H., Tanigaki, K., Yamamoto, N., Kuroda, K., Yoshimoto, M., Nakahata, T., Ikuta, K. and Honjo, T. (2002). Inducible gene knockout of transcription factor recombination signal binding protein-J reveals its essential role in T versus B lineage decision. Int Immunol 14, 637-45. Harduin-Lepers, A., Vallejo-Ruiz, V., Krzewinski-Recchi, M. A., Samyn-Petit, B., Julien, S. and Delannoy, P. (2001). The human sialyltransferase family. Biochimie 83, 727-37. Hardy, R. R. (2006). B-1 B cell development. J Immunol 177, 2749-54. Hardy, R. R., Carmack, C. E., Shinton, S. A., Kemp, J. D. and Hayakawa, K. (1991). Resolution and characterization of pro-B and pre-pro-B cell stages in normal mouse bone marrow. J Exp Med 173, 1213-25. Harris, R. J. and Spellman, M. W. (1993). O-linked fucose and other post-translational modifications unique to EGF modules. Glycobiology 3, 219-24. Hartley, S. B., Cooke, M. P., Fulcher, D. A., Harris, A. W., Cory, S., Basten, A. and Goodnow, C. C. (1993). Elimination of self-reactive B lymphocytes proceeds in two stages: arrested development and cell death. Cell 72, 325-35. Hasan, M., Polic, B., Bralic, M., Jonjic, S. and Rajewsky, K. (2002). Incomplete block of B cell development and immunoglobulin production in mice carrying the muMT mutation on the BALB/c background. Eur J Immunol 32, 3463-71. Hashimoto, S., Iwamatsu, A., Ishiai, M., Okawa, K., Yamadori, T., Matsushita, M., Baba, Y., Kishimoto, T., Kurosaki, T. and Tsukada, S. (1999). Identification of the SH2 domain binding protein of Bruton's tyrosine kinase as BLNK--functional significance of Btk-SH2 domain in B-cell antigen receptor-coupled calcium signaling. Blood 94, 2357-64.

161
Hauke, G., Epplen, J. T., Chluba, J., Hollmann, A. and Peter, H. H. (1988). IgD/lambda plasmocytoma with immunoglobulin kappa light-chain genes in the germ-line configuration. J Clin Immunol 8, 407-13. Hecht, A. and Kemler, R. (2000). Curbing the nuclear activities of beta-catenin. Control over Wnt target gene expression. EMBO Rep 1, 24-8. Hertz, M. and Nemazee, D. (1997). BCR ligation induces receptor editing in IgM+IgD- bone marrow B cells in vitro. Immunity 6, 429-36. Hibbs, M. L., Tarlinton, D. M., Armes, J., Grail, D., Hodgson, G., Maglitto, R., Stacker, S. A. and Dunn, A. R. (1995). Multiple defects in the immune system of Lyn-deficient mice, culminating in autoimmune disease. Cell 83, 301-11. Hieter, P. A., Korsmeyer, S. J., Waldmann, T. A. and Leder, P. (1981). Human immunoglobulin kappa light-chain genes are deleted or rearranged in lambda-producing B cells. Nature 290, 368-72. Hippen, K. L., Buhl, A. M., D'Ambrosio, D., Nakamura, K., Persin, C. and Cambier, J. C. (1997). Fc gammaRIIB1 inhibition of BCR-mediated phosphoinositide hydrolysis and Ca2+ mobilization is integrated by CD19 dephosphorylation. Immunity 7, 49-58. Hodoniczky, J., Zheng, Y. Z. and James, D. C. (2005). Control of recombinant monoclonal antibody effector functions by Fc N-glycan remodeling in vitro. Biotechnol Prog 21, 1644-52. Hoffmann, A., Kerr, S., Jellusova, J., Zhang, J., Weisel, F., Wellmann, U., Winkler, T. H., Kneitz, B., Crocker, P. R. and Nitschke, L. (2007). Siglec-G is a B1 cell-inhibitory receptor that controls expansion and calcium signaling of the B1 cell population. Nat Immunol 8, 695-704. Hombach, J., Leclercq, L., Radbruch, A., Rajewsky, K. and Reth, M. (1988). A novel 34-kd protein co-isolated with the IgM molecule in surface IgM-expressing cells. Embo J 7, 3451-6. Hombach, J., Lottspeich, F. and Reth, M. (1990). Identification of the genes encoding the IgM-alpha and Ig-beta components of the IgM antigen receptor complex by amino-terminal sequencing. Eur J Immunol 20, 2795-9. Honjo, Y., Nangia-Makker, P., Inohara, H. and Raz, A. (2001). Down-regulation of galectin-3 suppresses tumorigenicity of human breast carcinoma cells. Clin Cancer Res 7, 661-8. Horikawa, K., Martin, S. W., Pogue, S. L., Silver, K., Peng, K., Takatsu, K. and Goodnow, C. C. (2007). Enhancement and suppression of signaling by the conserved tail of IgG memory-type B cell antigen receptors. J Exp Med 204, 759-69. Hoyer, K. K., Pang, M., Gui, D., Shintaku, I. P., Kuwabara, I., Liu, F. T., Said, J. W., Baum, L. G. and Teitell, M. A. (2004). An anti-apoptotic role for galectin-3 in diffuse large B-cell lymphomas. Am J Pathol 164, 893-902. Hozumi, K., Negishi, N., Suzuki, D., Abe, N., Sotomaru, Y., Tamaoki, N., Mailhos, C., Ish-Horowicz, D., Habu, S. and Owen, M. J. (2004). Delta-like 1 is necessary for the generation of marginal zone B cells but not T cells in vivo. Nat Immunol 5, 638-44. Huber, C., Huber, E., Lautner-Rieske, A., Schable, K. F. and Zachau, H. G. (1993). The human immunoglobulin kappa locus. Characterization of the partially duplicated L regions. Eur J Immunol 23, 2860-7. Huelsken, J. and Birchmeier, W. (2001). New aspects of Wnt signaling pathways in higher vertebrates. Curr Opin Genet Dev 11, 547-53. Hughes, R. C. (1999). Secretion of the galectin family of mammalian carbohydrate-binding proteins. Biochim Biophys Acta 1473, 172-85.

162
Hwang, H. Y., Olson, S. K., Esko, J. D. and Horvitz, H. R. (2003). Caenorhabditis elegans early embryogenesis and vulval morphogenesis require chondroitin biosynthesis. Nature 423, 439-43. Ingold, K., Zumsteg, A., Tardivel, A., Huard, B., Steiner, Q. G., Cachero, T. G., Qiang, F., Gorelik, L., Kalled, S. L., Acha-Orbea, H. et al. (2005). Identification of proteoglycans as the APRIL-specific binding partners. J Exp Med 201, 1375-83. Inlay, M., Alt, F. W., Baltimore, D. and Xu, Y. (2002). Essential roles of the kappa light chain intronic enhancer and 3' enhancer in kappa rearrangement and demethylation. Nat Immunol 3, 463-8. Iozzo, R. V. (1998). Matrix proteoglycans: from molecular design to cellular function. Annu Rev Biochem 67, 609-52. Iritani, B. M., Forbush, K. A., Farrar, M. A. and Perlmutter, R. M. (1997). Control of B cell development by Ras-mediated activation of Raf. Embo J 16, 7019-31. Ito, S., Nagaoka, H., Shinkura, R., Begum, N., Muramatsu, M., Nakata, M. and Honjo, T. (2004). Activation-induced cytidine deaminase shuttles between nucleus and cytoplasm like apolipoprotein B mRNA editing catalytic polypeptide 1. Proc Natl Acad Sci U S A 101, 1975-80. Jack, H. M. and Wabl, M. (1988). Immunoglobulin mRNA stability varies during B lymphocyte differentiation. Embo J 7, 1041-6. Jacob, J., Kassir, R. and Kelsoe, G. (1991). In situ studies of the primary immune response to (4-hydroxy-3-nitrophenyl)acetyl. I. The architecture and dynamics of responding cell populations. J Exp Med 173, 1165-75. Jefferis, R. (2005). Glycosylation of recombinant antibody therapeutics. Biotechnol Prog 21, 11-6. Jego, P., Joly, J. and Boisseau, C. (1980). [Amphibian jelly envelopes (proteins secreted by the oviduct) and their role in fertilization]. Reprod Nutr Dev 20, 557-67. Johnson, G. L. and Lapadat, R. (2002). Mitogen-activated protein kinase pathways mediated by ERK, JNK, and p38 protein kinases. Science 298, 1911-2. Jung, D., Giallourakis, C., Mostoslavsky, R. and Alt, F. W. (2006). Mechanism and control of V(D)J recombination at the immunoglobulin heavy chain locus. Annu Rev Immunol 24, 541-70. Kabak, S., Skaggs, B. J., Gold, M. R., Affolter, M., West, K. L., Foster, M. S., Siemasko, K., Chan, A. C., Aebersold, R. and Clark, M. R. (2002). The direct recruitment of BLNK to immunoglobulin alpha couples the B-cell antigen receptor to distal signaling pathways. Mol Cell Biol 22, 2524-35. Kabat, E. A. (1956). Heterogeneity in extent of the combining regions of human antidextran. J Immunol 77, 377-85. Kadesch, T., Zervos, P. and Ruezinsky, D. (1986). Functional analysis of the murine IgH enhancer: evidence for negative control of cell-type specificity. Nucleic Acids Res 14, 8209-21. Kadonaga, J. T. (1998). Eukaryotic transcription: an interlaced network of transcription factors and chromatin-modifying machines. Cell 92, 307-13. Kaisho, T., Schwenk, F. and Rajewsky, K. (1997). The roles of gamma 1 heavy chain membrane expression and cytoplasmic tail in IgG1 responses. Science 276, 412-5. Kallies, A. and Nutt, S. L. (2007). Terminal differentiation of lymphocytes depends on Blimp-1. Curr Opin Immunol 19, 156-62. Kaneko, Y., Nimmerjahn, F. and Ravetch, J. V. (2006). Anti-inflammatory activity of immunoglobulin G resulting from Fc sialylation. Science 313, 670-3.

163
Kao, Y. H., Lee, G. F., Wang, Y., Starovasnik, M. A., Kelley, R. F., Spellman, M. W. and Lerner, L. (1999). The effect of O-fucosylation on the first EGF-like domain from human blood coagulation factor VII. Biochemistry 38, 7097-110. Karim, R., Tse, G., Putti, T., Scolyer, R. and Lee, S. (2004). The significance of the Wnt pathway in the pathology of human cancers. Pathology 36, 120-8. Katagiri, T., Ogimoto, M., Hasegawa, K., Arimura, Y., Mitomo, K., Okada, M., Clark, M. R., Mizuno, K. and Yakura, H. (1999). CD45 negatively regulates lyn activity by dephosphorylating both positive and negative regulatory tyrosine residues in immature B cells. J
Immunol 163, 1321-6. Kawashima, H., Atarashi, K., Hirose, M., Hirose, J., Yamada, S., Sugahara, K. and Miyasaka, M. (2002). Oversulfated chondroitin/dermatan sulfates containing GlcAbeta1/IdoAalpha1-3GalNAc(4,6-O-disulfate) interact with L- and P-selectin and chemokines. J Biol Chem 277, 12921-30. Kemler, I., Schreiber, E., Muller, M. M., Matthias, P. and Schaffner, W. (1989). Octamer transcription factors bind to two different sequence motifs of the immunoglobulin heavy chain promoter. Embo J 8, 2001-8. Kenter, A. L. (2003). Class-switch recombination: after the dawn of AID. Curr Opin
Immunol 15, 190-8. Khamlichi, A. A., Glaudet, F., Oruc, Z., Denis, V., Le Bert, M. and Cogne, M. (2004). Immunoglobulin class-switch recombination in mice devoid of any S mu tandem repeat. Blood 103, 3828-36. Kim, K. M. and Reth, M. (1995). The B cell antigen receptor of class IgD induces a stronger and more prolonged protein tyrosine phosphorylation than that of class IgM. J Exp Med 181, 1005-14. Kitamura, D. and Rajewsky, K. (1992). Targeted disruption of mu chain membrane exon causes loss of heavy-chain allelic exclusion. Nature 356, 154-6. Kitamura, D., Roes, J., Kuhn, R. and Rajewsky, K. (1991). A B cell-deficient mouse by targeted disruption of the membrane exon of the immunoglobulin mu chain gene. Nature 350, 423-6. Kleene, R. and Berger, E. G. (1993). The molecular and cell biology of glycosyltransferases. Biochim Biophys Acta 1154, 283-325. Klein, U., Casola, S., Cattoretti, G., Shen, Q., Lia, M., Mo, T., Ludwig, T., Rajewsky, K. and Dalla-Favera, R. (2006). Transcription factor IRF4 controls plasma cell differentiation and class-switch recombination. Nat Immunol 7, 773-82. Klippel, A., Kavanaugh, W. M., Pot, D. and Williams, L. T. (1997). A specific product of phosphatidylinositol 3-kinase directly activates the protein kinase Akt through its pleckstrin homology domain. Mol Cell Biol 17, 338-44. Knight, A. M., Lucocq, J. M., Prescott, A. R., Ponnambalam, S. and Watts, C. (1997). Antigen endocytosis and presentation mediated by human membrane IgG1 in the absence of the Ig(alpha)/Ig(beta) dimer. Embo J 16, 3842-50. Kolset, S. O. and Gallagher, J. T. (1990). Proteoglycans in haemopoietic cells. Biochim
Biophys Acta 1032, 191-211. Kornfeld, R. and Kornfeld, S. (1985). Assembly of asparagine-linked oligosaccharides. Annu Rev Biochem 54, 631-64. Korsmeyer, S. J., Hieter, P. A., Sharrow, S. O., Goldman, C. K., Leder, P. and Waldmann, T. A. (1982). Normal human B cells display ordered light chain gene rearrangements and deletions. J Exp Med 156, 975-85.

164
Kottmann, A. H., Zevnik, B., Welte, M., Nielsen, P. J. and Kohler, G. (1994). A second promoter and enhancer element within the immunoglobulin heavy chain locus. Eur J
Immunol 24, 817-21. Krappmann, D., Patke, A., Heissmeyer, V. and Scheidereit, C. (2001). B-cell receptor- and phorbol ester-induced NF-kappaB and c-Jun N-terminal kinase activation in B cells requires novel protein kinase C's. Mol Cell Biol 21, 6640-50. Kukuruzinska, M. A. and Lennon-Hopkins, K. (1999). ALG gene expression and cell cycle progression. Biochim Biophys Acta 1426, 359-72. Kumararatne, D. S. and MacLennan, I. C. (1982). The origin of marginal-zone cells. Adv Exp Med Biol 149, 83-90. Kuppers, R., Zhao, M., Hansmann, M. L. and Rajewsky, K. (1993). Tracing B cell development in human germinal centres by molecular analysis of single cells picked from histological sections. Embo J 12, 4955-67. Kuroda, K., Han, H., Tani, S., Tanigaki, K., Tun, T., Furukawa, T., Taniguchi, Y., Kurooka, H., Hamada, Y., Toyokuni, S. et al. (2003). Regulation of marginal zone B cell development by MINT, a suppressor of Notch/RBP-J signaling pathway. Immunity 18, 301-12. Kurooka, H. and Honjo, T. (2000). Functional interaction between the mouse notch1 intracellular region and histone acetyltransferases PCAF and GCN5. J Biol Chem 275, 17211-20. Kurtz, B. S., Witte, P. L. and Storb, U. (1997). Gamma 2b provides only some of the signals normally given via mu in B cell development. Int Immunol 9, 415-26. Lagana, A., Goetz, J. G., Cheung, P., Raz, A., Dennis, J. W. and Nabi, I. R. (2006). Galectin binding to Mgat5-modified N-glycans regulates fibronectin matrix remodeling in tumor cells. Mol Cell Biol 26, 3181-93. Larson, E. D., Cummings, W. J., Bednarski, D. W. and Maizels, N. (2005). MRE11/RAD50 cleaves DNA in the AID/UNG-dependent pathway of immunoglobulin gene diversification. Mol Cell 20, 367-75. Laskin, J. D., Dokidis, A., Sirak, A. A. and Laskin, D. L. (1991). Distinct patterns of sulfated proteoglycan biosynthesis in human monocytes, granulocytes and myeloid leukemic cells. Leuk Res 15, 515-23. LeBowitz, J. H., Kobayashi, T., Staudt, L., Baltimore, D. and Sharp, P. A. (1988). Octamer-binding proteins from B or HeLa cells stimulate transcription of the immunoglobulin heavy-chain promoter in vitro. Genes Dev 2, 1227-37. Leduc, I. and Cogne, M. (1996). Regulatory elements of the mb-1 gene encoding the Ig-alpha component of the human B-cell antigen receptor. Mol Immunol 33, 1277-86. Leeb, B. F., Petera, P. and Neumann, K. (1996). [Results of a multicenter study of chondroitin sulfate (Condrosulf) use in arthroses of the finger, knee and hip joints]. Wien Med
Wochenschr 146, 609-14. Lefranc, M. P., Pallares, N. and Frippiat, J. P. (1999). Allelic polymorphisms and RFLP in the human immunoglobulin lambda light chain locus. Hum Genet 104, 361-9. Li, D. H., Tung, J. W., Tarner, I. H., Snow, A. L., Yukinari, T., Ngernmaneepothong, R., Martinez, O. M. and Parnes, J. R. (2006). CD72 down-modulates BCR-induced signal transduction and diminishes survival in primary mature B lymphocytes. J Immunol 176, 5321-8. Li, Y. S., Wasserman, R., Hayakawa, K. and Hardy, R. R. (1996). Identification of the earliest B lineage stage in mouse bone marrow. Immunity 5, 527-35. Liang, H., Chen, Q., Coles, A. H., Anderson, S. J., Pihan, G., Bradley, A., Gerstein, R., Jurecic, R. and Jones, S. N. (2003). Wnt5a inhibits B cell proliferation and functions as a tumor suppressor in hematopoietic tissue. Cancer Cell 4, 349-60.

165
Lieberson, R., Giannini, S. L., Birshtein, B. K. and Eckhardt, L. A. (1991). An enhancer at the 3' end of the mouse immunoglobulin heavy chain locus. Nucleic Acids Res 19, 933-7. Lin, K. I., Tunyaplin, C. and Calame, K. (2003). Transcriptional regulatory cascades controlling plasma cell differentiation. Immunol Rev 194, 19-28. Lindahl, U., Kusche-Gullberg, M. and Kjellen, L. (1998). Regulated diversity of heparan sulfate. J Biol Chem 273, 24979-82. Linenberger, M. L., Maloney, D. G. and Bernstein, I. D. (2002). Antibody-directed therapies for hematological malignancies. Trends Mol Med 8, 69-76. Liu, D., Jenab, J. and Rosenberg, N. (1997). Kappa and lambda rearrangement occur simultaneously in transformed pre-B cells. J Immunol 159, 6061-9. Liu, F. T. and Rabinovich, G. A. (2005). Galectins as modulators of tumour progression. Nat Rev Cancer 5, 29-41. Liu, P. I., Poon, K. C. and Liu, S. S. (1981). In vitro effect of glucan on mouse hemopoietic committed progenitor cells. Life Sci 29, 1027-32. Liu, Y. J. and Arpin, C. (1997). Germinal center development. Immunol Rev 156, 111-26. Liu, Y. J., Barthelemy, C., de Bouteiller, O., Arpin, C., Durand, I. and Banchereau, J. (1995). Memory B cells from human tonsils colonize mucosal epithelium and directly present antigen to T cells by rapid up-regulation of B7-1 and B7-2. Immunity 2, 239-48. Liu, Y. J., Zhang, J., Lane, P. J., Chan, E. Y. and MacLennan, I. C. (1991). Sites of specific B cell activation in primary and secondary responses to T cell-dependent and T cell-independent antigens. Eur J Immunol 21, 2951-62. Liu, Z. M., George-Raizen, J. B., Li, S., Meyers, K. C., Chang, M. Y. and Garrard, W. T. (2002). Chromatin structural analyses of the mouse Igkappa gene locus reveal new hypersensitive sites specifying a transcriptional silencer and enhancer. J Biol Chem 277, 32640-9. Loder, F., Mutschler, B., Ray, R. J., Paige, C. J., Sideras, P., Torres, R., Lamers, M. C. and Carsetti, R. (1999). B cell development in the spleen takes place in discrete steps and is determined by the quality of B cell receptor-derived signals. J Exp Med 190, 75-89. Lowe, J. B. and Marth, J. D. (2003). A genetic approach to Mammalian glycan function. Annu Rev Biochem 72, 643-91. Luby, T. M., Schrader, C. E., Stavnezer, J. and Selsing, E. (2001). The mu switch region tandem repeats are important, but not required, for antibody class switch recombination. J
Exp Med 193, 159-68. Luisiri, P., Lee, Y. J., Eisfelder, B. J. and Clark, M. R. (1996). Cooperativity and segregation of function within the Ig-alpha/beta heterodimer of the B cell antigen receptor complex. J Biol Chem 271, 5158-63. Lund, J., Takahashi, N., Pound, J. D., Goodall, M., Nakagawa, H. and Jefferis, R. (1995). Oligosaccharide-protein interactions in IgG can modulate recognition by Fc gamma receptors. Faseb J 9, 115-9. Luo, Y., Koles, K., Vorndam, W., Haltiwanger, R. S. and Panin, V. M. (2006). Protein O-fucosyltransferase 2 adds O-fucose to thrombospondin type 1 repeats. J Biol Chem 281, 9393-9. Lutz, C., Ledermann, B., Kosco-Vilbois, M. H., Ochsenbein, A. F., Zinkernagel, R. M., Kohler, G. and Brombacher, F. (1998). IgD can largely substitute for loss of IgM function in B cells. Nature 393, 797-801. Lutzker, S., Rothman, P., Pollock, R., Coffman, R. and Alt, F. W. (1988). Mitogen- and IL-4-regulated expression of germ-line Ig gamma 2b transcripts: evidence for directed heavy chain class switching. Cell 53, 177-84.

166
Ma, Y., Pannicke, U., Schwarz, K. and Lieber, M. R. (2002). Hairpin opening and overhang processing by an Artemis/DNA-dependent protein kinase complex in nonhomologous end joining and V(D)J recombination. Cell 108, 781-94. Ma, Y. Q. and Geng, J. G. (2000). Heparan sulfate-like proteoglycans mediate adhesion of human malignant melanoma A375 cells to P-selectin under flow. J Immunol 165, 558-65. MacLennan, I. C. (1994). Germinal centers. Annu Rev Immunol 12, 117-39. Macpherson, A. J., Lamarre, A., McCoy, K., Harriman, G. R., Odermatt, B., Dougan, G., Hengartner, H. and Zinkernagel, R. M. (2001). IgA production without mu or delta chain expression in developing B cells. Nat Immunol 2, 625-31. Madisen, L. and Groudine, M. (1994). Identification of a locus control region in the immunoglobulin heavy-chain locus that deregulates c-myc expression in plasmacytoma and Burkitt's lymphoma cells. Genes Dev 8, 2212-26. Makatsori, E., Karamanos, N. K., Papadogiannakis, N., Hjerpe, A., Anastassiou, E. D. and Tsegenidis, T. (2001). Synthesis and distribution of glycosaminoglycans in human leukemic B- and T-cells and monocytes studied using specific enzymic treatments and high-performance liquid chromatography. Biomed Chromatogr 15, 413-7. Maki, R., Roeder, W., Traunecker, A., Sidman, C., Wabl, M., Raschke, W. and Tonegawa, S. (1981). The role of DNA rearrangement and alternative RNA processing in the expression of immunoglobulin delta genes. Cell 24, 353-65. Manis, J. P., Dudley, D., Kaylor, L. and Alt, F. W. (2002a). IgH class switch recombination to IgG1 in DNA-PKcs-deficient B cells. Immunity 16, 607-17. Manis, J. P., Tian, M. and Alt, F. W. (2002b). Mechanism and control of class-switch recombination. Trends Immunol 23, 31-9. Manis, J. P., van der Stoep, N., Tian, M., Ferrini, R., Davidson, L., Bottaro, A. and Alt, F. W. (1998). Class switching in B cells lacking 3' immunoglobulin heavy chain enhancers. J Exp Med 188, 1421-31. Manser, T. (2002). Effector BCRs: inside information on IgG. Nat Immunol 3, 114-6. Martensson, A., Argon, Y., Melchers, F., Dul, J. L. and Martensson, I. L. (1999). Partial block in B lymphocyte development at the transition into the pre-B cell receptor stage in Vpre-B1-deficient mice. Int Immunol 11, 453-60. Martensson, I. L., Keenan, R. A. and Licence, S. (2007). The pre-B-cell receptor. Curr
Opin Immunol 19, 137-42. Martin, S. W. and Goodnow, C. C. (2002). Burst-enhancing role of the IgG membrane tail as a molecular determinant of memory. Nat Immunol 3, 182-8. Mason, J. O., Williams, G. T. and Neuberger, M. S. (1988). The half-life of immunoglobulin mRNA increases during B-cell differentiation: a possible role for targeting to membrane-bound polysomes. Genes Dev 2, 1003-11. Mather, E. L. and Perry, R. P. (1983). Methylation status and DNase I sensitivity of immunoglobulin genes: changes associated with rearrangement. Proc Natl Acad Sci U S A 80, 4689-93. Matthias, P. and Baltimore, D. (1993). The immunoglobulin heavy chain locus contains another B-cell-specific 3' enhancer close to the alpha constant region. Mol Cell Biol 13, 1547-53. McBride, K. M., Barreto, V., Ramiro, A. R., Stavropoulos, P. and Nussenzweig, M. C. (2004). Somatic hypermutation is limited by CRM1-dependent nuclear export of activation-induced deaminase. J Exp Med 199, 1235-44. McEver, R. P., Moore, K. L. and Cummings, R. D. (1995). Leukocyte trafficking mediated by selectin-carbohydrate interactions. J Biol Chem 270, 11025-8.

167
Meek, K. D., Hasemann, C. A. and Capra, J. D. (1989). Novel rearrangements at the immunoglobulin D locus. Inversions and fusions add to IgH somatic diversity. J Exp Med 170, 39-57. Melamed, D. and Nemazee, D. (1997). Self-antigen does not accelerate immature B cell apoptosis, but stimulates receptor editing as a consequence of developmental arrest. Proc Natl
Acad Sci U S A 94, 9267-72. Melchers, F., Karasuyama, H., Haasner, D., Bauer, S., Kudo, A., Sakaguchi, N., Jameson, B. and Rolink, A. (1993). The surrogate light chain in B-cell development. Immunol
Today 14, 60-8. Melchers, F., ten Boekel, E., Yamagami, T., Andersson, J. and Rolink, A. (1999). The roles of preB and B cell receptors in the stepwise allelic exclusion of mouse IgH and L chain gene loci. Semin Immunol 11, 307-17. Merrell, K. T., Benschop, R. J., Gauld, S. B., Aviszus, K., Decote-Ricardo, D., Wysocki, L. J. and Cambier, J. C. (2006). Identification of anergic B cells within a wild-type repertoire. Immunity 25, 953-62. Michaelson, J. S., Giannini, S. L. and Birshtein, B. K. (1995). Identification of 3' alpha-hs4, a novel Ig heavy chain enhancer element regulated at multiple stages of B cell differentiation. Nucleic Acids Res 23, 975-81. Miller, J. R. (2002). The Wnts. Genome Biol 3, REVIEWS3001. Mimura, Y., Sondermann, P., Ghirlando, R., Lund, J., Young, S. P., Goodall, M. and Jefferis, R. (2001). Role of oligosaccharide residues of IgG1-Fc in Fc gamma RIIb binding. J
Biol Chem 276, 45539-47. Mizuguchi, S., Uyama, T., Kitagawa, H., Nomura, K. H., Dejima, K., Gengyo-Ando, K., Mitani, S., Sugahara, K. and Nomura, K. (2003). Chondroitin proteoglycans are involved in cell division of Caenorhabditis elegans. Nature 423, 443-8. Moiseeva, E. P., Williams, B., Goodall, A. H. and Samani, N. J. (2003). Galectin-1 interacts with beta-1 subunit of integrin. Biochem Biophys Res Commun 310, 1010-6. Mombaerts, P., Iacomini, J., Johnson, R. S., Herrup, K., Tonegawa, S. and Papaioannou, V. E. (1992). RAG-1-deficient mice have no mature B and T lymphocytes. Cell 68, 869-77. Mostoslavsky, R., Singh, N., Kirillov, A., Pelanda, R., Cedar, H., Chess, A. and Bergman, Y. (1998). Kappa chain monoallelic demethylation and the establishment of allelic exclusion. Genes Dev 12, 1801-11. Mostoslavsky, R., Singh, N., Tenzen, T., Goldmit, M., Gabay, C., Elizur, S., Qi, P., Reubinoff, B. E., Chess, A., Cedar, H. et al. (2001). Asynchronous replication and allelic exclusion in the immune system. Nature 414, 221-5. Moyron-Quiroz, J. E., Partida-Sanchez, S., Donis-Hernandez, R., Sandoval-Montes, C. and Santos-Argumedo, L. (2002). Expression and function of CD22, a B-cell restricted molecule. Scand J Immunol 55, 343-51. Mumm, J. S. and Kopan, R. (2000). Notch signaling: from the outside in. Dev Biol 228, 151-65. Mundt, C., Licence, S., Shimizu, T., Melchers, F. and Martensson, I. L. (2001). Loss of precursor B cell expansion but not allelic exclusion in VpreB1/VpreB2 double-deficient mice. J Exp Med 193, 435-45. Muramatsu, M., Kinoshita, K., Fagarasan, S., Yamada, S., Shinkai, Y. and Honjo, T. (2000). Class switch recombination and hypermutation require activation-induced cytidine deaminase (AID), a potential RNA editing enzyme. Cell 102, 553-63. Muramatsu, M., Sankaranand, V. S., Anant, S., Sugai, M., Kinoshita, K., Davidson, N. O. and Honjo, T. (1999). Specific expression of activation-induced cytidine deaminase

168
(AID), a novel member of the RNA-editing deaminase family in germinal center B cells. J Biol
Chem 274, 18470-6. Murre, C., McCaw, P. S., Vaessin, H., Caudy, M., Jan, L. Y., Jan, Y. N., Cabrera, C. V., Buskin, J. N., Hauschka, S. D., Lassar, A. B. et al. (1989). Interactions between heterologous helix-loop-helix proteins generate complexes that bind specifically to a common DNA sequence. Cell 58, 537-44. Nadler, M. J., McLean, P. A., Neel, B. G. and Wortis, H. H. (1997). B cell antigen receptor-evoked calcium influx is enhanced in CD22-deficient B cell lines. J Immunol 159, 4233-43. Nambu, Y., Sugai, M., Gonda, H., Lee, C. G., Katakai, T., Agata, Y., Yokota, Y. and Shimizu, A. (2003). Transcription-coupled events associating with immunoglobulin switch region chromatin. Science 302, 2137-40. Niiro, H. and Clark, E. A. (2002). Regulation of B-cell fate by antigen-receptor signals. Nat Rev Immunol 2, 945-56. Nimmerjahn, F. and Ravetch, J. V. (2005). Divergent immunoglobulin g subclass activity through selective Fc receptor binding. Science 310, 1510-2. Nishimura, T., Nakatake, Y., Konishi, M. and Itoh, N. (2000). Identification of a novel FGF, FGF-21, preferentially expressed in the liver. Biochim Biophys Acta 1492, 203-6. Nitschke, L. (2005). The role of CD22 and other inhibitory co-receptors in B-cell activation. Curr Opin Immunol 17, 290-7. Nitschke, L., Carsetti, R., Ocker, B., Kohler, G. and Lamers, M. C. (1997). CD22 is a negative regulator of B-cell receptor signalling. Curr Biol 7, 133-43. Nitschke, L., Kestler, J., Tallone, T., Pelkonen, S. and Pelkonen, J. (2001). Deletion of the DQ52 element within the Ig heavy chain locus leads to a selective reduction in VDJ recombination and altered D gene usage. J Immunol 166, 2540-52. Niwa, R., Natsume, A., Uehara, A., Wakitani, M., Iida, S., Uchida, K., Satoh, M. and Shitara, K. (2005). IgG subclass-independent improvement of antibody-dependent cellular cytotoxicity by fucose removal from Asn297-linked oligosaccharides. J Immunol Methods 306, 151-60. Noble, P. W., Lake, F. R., Henson, P. M. and Riches, D. W. (1993). Hyaluronate activation of CD44 induces insulin-like growth factor-1 expression by a tumor necrosis factor-alpha-dependent mechanism in murine macrophages. J Clin Invest 91, 2368-77. Norvell, A., Mandik, L. and Monroe, J. G. (1995). Engagement of the antigen-receptor on immature murine B lymphocytes results in death by apoptosis. J Immunol 154, 4404-13. Novobrantseva, T. I., Martin, V. M., Pelanda, R., Muller, W., Rajewsky, K. and Ehlich, A. (1999). Rearrangement and expression of immunoglobulin light chain genes can precede heavy chain expression during normal B cell development in mice. J Exp Med 189, 75-88. Nusse, R. and Varmus, H. E. (1982). Many tumors induced by the mouse mammary tumor virus contain a provirus integrated in the same region of the host genome. Cell 31, 99-109. Nutt, S. L. and Kee, B. L. (2007). The transcriptional regulation of B cell lineage commitment. Immunity 26, 715-25. O'Keefe, T. L., Williams, G. T., Batista, F. D. and Neuberger, M. S. (1999). Deficiency in CD22, a B cell-specific inhibitory receptor, is sufficient to predispose to development of high affinity autoantibodies. J Exp Med 189, 1307-13. O'Keefe, T. L., Williams, G. T., Davies, S. L. and Neuberger, M. S. (1996). Hyperresponsive B cells in CD22-deficient mice. Science 274, 798-801. Obara, N. and Lesot, H. (2004). Subcellular localization of beta-catenin and cadherin expression in the cap-stage enamel organ of the mouse molar. Histochem Cell Biol 121, 351-8.

169
Oberdoerffer, P., Novobrantseva, T. I. and Rajewsky, K. (2003). Expression of a targeted lambda 1 light chain gene is developmentally regulated and independent of Ig kappa rearrangements. J Exp Med 197, 1165-72. Ochsenbein, A. F., Pinschewer, D. D., Sierro, S., Horvath, E., Hengartner, H. and Zinkernagel, R. M. (2000). Protective long-term antibody memory by antigen-driven and T help-dependent differentiation of long-lived memory B cells to short-lived plasma cells independent of secondary lymphoid organs. Proc Natl Acad Sci U S A 97, 13263-8. Oettgen, H. C., Martin, T. R., Wynshaw-Boris, A., Deng, C., Drazen, J. M. and Leder, P. (1994). Active anaphylaxis in IgE-deficient mice. Nature 370, 367-70. Oettinger, M. A., Schatz, D. G., Gorka, C. and Baltimore, D. (1990). RAG-1 and RAG-2, adjacent genes that synergistically activate V(D)J recombination. Science 248, 1517-23. Ohnishi, K. and Melchers, F. (2003). The nonimmunoglobulin portion of lambda5 mediates cell-autonomous pre-B cell receptor signaling. Nat Immunol 4, 849-56. Oliver, A. M., Martin, F., Gartland, G. L., Carter, R. H. and Kearney, J. F. (1997). Marginal zone B cells exhibit unique activation, proliferative and immunoglobulin secretory responses. Eur J Immunol 27, 2366-74. Oliver, A. M., Martin, F. and Kearney, J. F. (1999). IgMhighCD21high lymphocytes enriched in the splenic marginal zone generate effector cells more rapidly than the bulk of follicular B cells. J Immunol 162, 7198-207. Orinska, Z., Osiak, A., Lohler, J., Bulanova, E., Budagian, V., Horak, I. and Bulfone-Paus, S. (2002). Novel B cell population producing functional IgG in the absence of membrane IgM expression. Eur J Immunol 32, 3472-80. Ornitz, D. M., Yayon, A., Flanagan, J. G., Svahn, C. M., Levi, E. and Leder, P. (1992). Heparin is required for cell-free binding of basic fibroblast growth factor to a soluble receptor and for mitogenesis in whole cells. Mol Cell Biol 12, 240-7. Osborne, B. and Miele, L. (1999). Notch and the immune system. Immunity 11, 653-63. Osborne, B. A. and Minter, L. M. (2007). Notch signalling during peripheral T-cell activation and differentiation. Nat Rev Immunol 7, 64-75. Otero, D. C., Omori, S. A. and Rickert, R. C. (2001). Cd19-dependent activation of Akt kinase in B-lymphocytes. J Biol Chem 276, 1474-8. Pace, K. E., Lee, C., Stewart, P. L. and Baum, L. G. (1999). Restricted receptor segregation into membrane microdomains occurs on human T cells during apoptosis induced by galectin-1. J Immunol 163, 3801-11. Pan, Y. T. and Elbein, A. D. (1990). Control of N-linked oligosaccharide synthesis: cellular levels of dolichyl phosphate are not the only regulatory factor. Biochemistry 29, 8077-84. Panin, V. M., Papayannopoulos, V., Wilson, R. and Irvine, K. D. (1997). Fringe modulates Notch-ligand interactions. Nature 387, 908-12. Pao, L. I., Famiglietti, S. J. and Cambier, J. C. (1998). Asymmetrical phosphorylation and function of immunoreceptor tyrosine-based activation motif tyrosines in B cell antigen receptor signal transduction. J Immunol 160, 3305-14. Parkhouse, R. M. (1990). Three B-cell surface molecules associating with membrane immunoglobulin. Immunology 69, 298-302. Parslow, T. G., Blair, D. L., Murphy, W. J. and Granner, D. K. (1984). Structure of the 5' ends of immunoglobulin genes: a novel conserved sequence. Proc Natl Acad Sci U S A 81, 2650-4. Paulson, J. C. and Colley, K. J. (1989). Glycosyltransferases. Structure, localization, and control of cell type-specific glycosylation. J Biol Chem 264, 17615-8. Pauza, M. E., Rehmann, J. A. and LeBien, T. W. (1993). Unusual patterns of immunoglobulin gene rearrangement and expression during human B cell ontogeny: human B

170
cells can simultaneously express cell surface kappa and lambda light chains. J Exp Med 178, 139-49. Pelanda, R., Schwers, S., Sonoda, E., Torres, R. M., Nemazee, D. and Rajewsky, K. (1997). Receptor editing in a transgenic mouse model: site, efficiency, and role in B cell tolerance and antibody diversification. Immunity 7, 765-75. Perlot, T., Alt, F. W., Bassing, C. H., Suh, H. and Pinaud, E. (2005). Elucidation of IgH intronic enhancer functions via germ-line deletion. Proc Natl Acad Sci U S A 102, 14362-7. Perrimon, N. and Bernfield, M. (2000). Specificities of heparan sulphate proteoglycans in developmental processes. Nature 404, 725-8. Perrimon, N. and Bernfield, M. (2001). Cellular functions of proteoglycans--an overview. Semin Cell Dev Biol 12, 65-7. Petersen-Mahrt, S. K., Harris, R. S. and Neuberger, M. S. (2002). AID mutates E. coli suggesting a DNA deamination mechanism for antibody diversification. Nature 418, 99-103. Petersen, S., Casellas, R., Reina-San-Martin, B., Chen, H. T., Difilippantonio, M. J., Wilson, P. C., Hanitsch, L., Celeste, A., Muramatsu, M., Pilch, D. R. et al. (2001). AID is required to initiate Nbs1/gamma-H2AX focus formation and mutations at sites of class switching. Nature 414, 660-5. Peterson, M. L. and Perry, R. P. (1986). Regulated production of mu m and mu s mRNA requires linkage of the poly(A) addition sites and is dependent on the length of the mu s-mu m intron. Proc Natl Acad Sci U S A 83, 8883-7. Petitou, M., Lormeau, J. C. and Choay, J. (1988). Interaction of heparin and antithrombin III. The role of O-sulfate groups. Eur J Biochem 176, 637-40. Petro, J. B., Gerstein, R. M., Lowe, J., Carter, R. S., Shinners, N. and Khan, W. N. (2002). Transitional type 1 and 2 B lymphocyte subsets are differentially responsive to antigen receptor signaling. J Biol Chem 277, 48009-19. Pfisterer, P., Annweiler, A., Ullmer, C., Corcoran, L. M. and Wirth, T. (1994). Differential transactivation potential of Oct1 and Oct2 is determined by additional B cell-specific activities. Embo J 13, 1654-63. Pham, P., Bransteitter, R., Petruska, J. and Goodman, M. F. (2003). Processive AID-catalysed cytosine deamination on single-stranded DNA simulates somatic hypermutation. Nature 424, 103-7. Pierani, A., Heguy, A., Fujii, H. and Roeder, R. G. (1990). Activation of octamer-containing promoters by either octamer-binding transcription factor 1 (OTF-1) or OTF-2 and requirement of an additional B-cell-specific component for optimal transcription of immunoglobulin promoters. Mol Cell Biol 10, 6204-15. Pierce, J. W., Gifford, A. M. and Baltimore, D. (1991). Silencing of the expression of the immunoglobulin kappa gene in non-B cells. Mol Cell Biol 11, 1431-7. Pike, B. L., Boyd, A. W. and Nossal, G. J. (1982). Clonal anergy: the universally anergic B lymphocyte. Proc Natl Acad Sci U S A 79, 2013-7. Pillai, S., Cariappa, A. and Moran, S. T. (2004). Positive selection and lineage commitment during peripheral B-lymphocyte development. Immunol Rev 197, 206-18. Pinaud, E., Khamlichi, A. A., Le Morvan, C., Drouet, M., Nalesso, V., Le Bert, M. and Cogne, M. (2001). Localization of the 3' IgH locus elements that effect long-distance regulation of class switch recombination. Immunity 15, 187-99. Poe, J. C., Fujimoto, Y., Hasegawa, M., Haas, K. M., Miller, A. S., Sanford, I. G., Bock, C. B., Fujimoto, M. and Tedder, T. F. (2004). CD22 regulates B lymphocyte function in vivo through both ligand-dependent and ligand-independent mechanisms. Nat Immunol 5, 1078-87.

171
Poellinger, L., Yoza, B. K. and Roeder, R. G. (1989). Functional cooperativity between protein molecules bound at two distinct sequence elements of the immunoglobulin heavy-chain promoter. Nature 337, 573-6. Pogue, S. L. and Goodnow, C. C. (2000). Gene dose-dependent maturation and receptor editing of B cells expressing immunoglobulin (Ig)G1 or IgM/IgG1 tail antigen receptors. J Exp
Med 191, 1031-44. Popov, A. V., Zou, X., Xian, J., Nicholson, I. C. and Bruggemann, M. (1999). A human immunoglobulin lambda locus is similarly well expressed in mice and humans. J Exp Med 189, 1611-20. Preud'homme, J. L., Aucouturier, P., Touchard, G., Striker, L., Khamlichi, A. A., Rocca, A., Denoroy, L. and Cogne, M. (1994). Monoclonal immunoglobulin deposition disease (Randall type). Relationship with structural abnormalities of immunoglobulin chains. Kidney Int 46, 965-72. Proudfoot, A. E., Fritchley, S., Borlat, F., Shaw, J. P., Vilbois, F., Zwahlen, C., Trkola, A., Marchant, D., Clapham, P. R. and Wells, T. N. (2001). The BBXB motif of RANTES is the principal site for heparin binding and controls receptor selectivity. J Biol Chem 276, 10620-6. Rada, C., Williams, G. T., Nilsen, H., Barnes, D. E., Lindahl, T. and Neuberger, M. S. (2002). Immunoglobulin isotype switching is inhibited and somatic hypermutation perturbed in UNG-deficient mice. Curr Biol 12, 1748-55. Radaev, S. and Sun, P. D. (2001). Recognition of IgG by Fcgamma receptor. The role of Fc glycosylation and the binding of peptide inhibitors. J Biol Chem 276, 16478-83. Radtke, F., Wilson, A., Stark, G., Bauer, M., van Meerwijk, J., MacDonald, H. R. and Aguet, M. (1999). Deficient T cell fate specification in mice with an induced inactivation of Notch1. Immunity 10, 547-58. Raff, M. C., Sternberg, M. and Taylor, R. B. (1970). Immunoglobulin determinants on the surface of mouse lymphoid cells. Nature 225, 553-4. Rafi, A., Nagarkatti, M. and Nagarkatti, P. S. (1997). Hyaluronate-CD44 interactions can induce murine B-cell activation. Blood 89, 2901-8. Rajewsky, K. (1996). Clonal selection and learning in the antibody system. Nature 381, 751-8. Raju, T. S., Briggs, J. B., Chamow, S. M., Winkler, M. E. and Jones, A. J. (2001). Glycoengineering of therapeutic glycoproteins: in vitro galactosylation and sialylation of glycoproteins with terminal N-acetylglucosamine and galactose residues. Biochemistry 40, 8868-76. Razi, N. and Varki, A. (1998). Masking and unmasking of the sialic acid-binding lectin activity of CD22 (Siglec-2) on B lymphocytes. Proc Natl Acad Sci U S A 95, 7469-74. Reichsman, F., Smith, L. and Cumberledge, S. (1996). Glycosaminoglycans can modulate extracellular localization of the wingless protein and promote signal transduction. J
Cell Biol 135, 819-27. Reina-San-Martin, B., Chen, J., Nussenzweig, A. and Nussenzweig, M. C. (2007). Enhanced intra-switch region recombination during immunoglobulin class switch recombination in 53BP1-/- B cells. Eur J Immunol 37, 235-9. Reth, M. (1989). Antigen receptor tail clue. Nature 338, 383-4. Revy, P., Muto, T., Levy, Y., Geissmann, F., Plebani, A., Sanal, O., Catalan, N., Forveille, M., Dufourcq-Labelouse, R., Gennery, A. et al. (2000). Activation-induced cytidine deaminase (AID) deficiency causes the autosomal recessive form of the Hyper-IgM syndrome (HIGM2). Cell 102, 565-75. Robey, E. (1999). Regulation of T cell fate by Notch. Annu Rev Immunol 17, 283-95.

172
Roes, J. and Rajewsky, K. (1993). Immunoglobulin D (IgD)-deficient mice reveal an auxiliary receptor function for IgD in antigen-mediated recruitment of B cells. J Exp Med 177, 45-55. Rogers, J., Choi, E., Souza, L., Carter, C., Word, C., Kuehl, M., Eisenberg, D. and Wall, R. (1981). Gene segments encoding transmembrane carboxyl termini of immunoglobulin gamma chains. Cell 26, 19-27. Rogers, J., Early, P., Carter, C., Calame, K., Bond, M., Hood, L. and Wall, R. (1980). Two mRNAs with different 3' ends encode membrane-bound and secreted forms of immunoglobulin mu chain. Cell 20, 303-12. Rogers, J. and Wall, R. (1984). Immunoglobulin RNA rearrangements in B lymphocyte differentiation. Adv Immunol 35, 39-59. Rolink, A. G., Andersson, J. and Melchers, F. (2004). Molecular mechanisms guiding late stages of B-cell development. Immunol Rev 197, 41-50. Rooney, S., Sekiguchi, J., Whitlow, S., Eckersdorff, M., Manis, J. P., Lee, C., Ferguson, D. O. and Alt, F. W. (2004). Artemis and p53 cooperate to suppress oncogenic N-myc amplification in progenitor B cells. Proc Natl Acad Sci U S A 101, 2410-5. Roseman, S. (1970). The synthesis of complex carbohydrates by multiglycosyltransferase systems and their potential function in intercellular adhesion. Chem Phys Lipids 5, 270-97. Rossi, B., Espeli, M., Schiff, C. and Gauthier, L. (2006). Clustering of pre-B cell integrins induces galectin-1-dependent pre-B cell receptor relocalization and activation. J
Immunol 177, 796-803. Roth, D. B. and Roth, S. Y. (2000). Unequal access: regulating V(D)J recombination through chromatin remodeling. Cell 103, 699-702. Roth, J. (2002). Protein N-glycosylation along the secretory pathway: relationship to organelle topography and function, protein quality control, and cell interactions. Chem Rev 102, 285-303. Roth, J., Wang, Y., Eckhardt, A. E. and Hill, R. L. (1994). Subcellular localization of the UDP-N-acetyl-D-galactosamine: polypeptide N-acetylgalactosaminyltransferase-mediated O-glycosylation reaction in the submaxillary gland. Proc Natl Acad Sci U S A 91, 8935-9. Roth, P. E., Doglio, L., Manz, J. T., Kim, J. Y., Lo, D. and Storb, U. (1993). Immunoglobulin gamma 2b transgenes inhibit heavy chain gene rearrangement, but cannot promote B cell development. J Exp Med 178, 2007-21. Roth, P. E., Kurtz, B., Lo, D. and Storb, U. (1995). lambda 5, but not mu, is required for B cell maturation in a unique gamma 2b transgenic mouse line. J Exp Med 181, 1059-70. Saito, T., Chiba, S., Ichikawa, M., Kunisato, A., Asai, T., Shimizu, K., Yamaguchi, T., Yamamoto, G., Seo, S., Kumano, K. et al. (2003). Notch2 is preferentially expressed in mature B cells and indispensable for marginal zone B lineage development. Immunity 18, 675-85. Sakaguchi, N., Kashiwamura, S., Kimoto, M., Thalmann, P. and Melchers, F. (1988). B lymphocyte lineage-restricted expression of mb-1, a gene with CD3-like structural properties. Embo J 7, 3457-64. Sakai, E., Bottaro, A. and Alt, F. W. (1999). The Ig heavy chain intronic enhancer core region is necessary and sufficient to promote efficient class switch recombination. Int Immunol 11, 1709-13. Salmivirta, M., Lidholt, K. and Lindahl, U. (1996). Heparan sulfate: a piece of information. Faseb J 10, 1270-9. Sasaki, Y., Casola, S., Kutok, J. L., Rajewsky, K. and Schmidt-Supprian, M. (2004). TNF family member B cell-activating factor (BAFF) receptor-dependent and -independent roles for BAFF in B cell physiology. J Immunol 173, 2245-52.

173
Sato, M., Adachi, T. and Tsubata, T. (2007). Augmentation of signaling through BCR containing IgE but not that containing IgA due to lack of CD22-mediated signal regulation. J
Immunol 178, 2901-7. Scallon, B. J., Tam, S. H., McCarthy, S. G., Cai, A. N. and Raju, T. S. (2007). Higher levels of sialylated Fc glycans in immunoglobulin G molecules can adversely impact functionality. Mol Immunol 44, 1524-34. Schamel, W. W. and Reth, M. (2000a). Monomeric and oligomeric complexes of the B cell antigen receptor. Immunity 13, 5-14. Schamel, W. W. and Reth, M. (2000b). Stability of the B cell antigen receptor complex. Mol Immunol 37, 253-9. Schatz, D. G., Oettinger, M. A. and Baltimore, D. (1989). The V(D)J recombination activating gene, RAG-1. Cell 59, 1035-48. Schiemann, B., Gommerman, J. L., Vora, K., Cachero, T. G., Shulga-Morskaya, S., Dobles, M., Frew, E. and Scott, M. L. (2001). An essential role for BAFF in the normal development of B cells through a BCMA-independent pathway. Science 293, 2111-4. Schilham, M. W., Wilson, A., Moerer, P., Benaissa-Trouw, B. J., Cumano, A. and Clevers, H. C. (1998). Critical involvement of Tcf-1 in expansion of thymocytes. J Immunol 161, 3984-91. Schlissel, M. S. (2003). Regulating antigen-receptor gene assembly. Nat Rev Immunol 3, 890-9. Schonherr, E., Witsch-Prehm, P., Harrach, B., Robenek, H., Rauterberg, J. and Kresse, H. (1995). Interaction of biglycan with type I collagen. J Biol Chem 270, 2776-83. Schrader, C. E., Linehan, E. K., Mochegova, S. N., Woodland, R. T. and Stavnezer, J. (2005). Inducible DNA breaks in Ig S regions are dependent on AID and UNG. J Exp Med 202, 561-8. Schubart, K., Massa, S., Schubart, D., Corcoran, L. M., Rolink, A. G. and Matthias, P. (2001). B cell development and immunoglobulin gene transcription in the absence of Oct-2 and OBF-1. Nat Immunol 2, 69-74. Schwartz, N. B. and Domowicz, M. (2002). Chondrodysplasias due to proteoglycan defects. Glycobiology 12, 57R-68R. Schweighoffer, E., Vanes, L., Mathiot, A., Nakamura, T. and Tybulewicz, V. L. (2003). Unexpected requirement for ZAP-70 in pre-B cell development and allelic exclusion. Immunity 18, 523-33. Sekiguchi, J. M. and Ferguson, D. O. (2006). DNA double-strand break repair: a relentless hunt uncovers new prey. Cell 124, 260-2. Sen, R. (2005). A move to exclude. Nat Immunol 6, 128-30. Serwe, M. and Sablitzky, F. (1993). V(D)J recombination in B cells is impaired but not blocked by targeted deletion of the immunoglobulin heavy chain intron enhancer. Embo J 12, 2321-7. Shaffer, A. L., Lin, K. I., Kuo, T. C., Yu, X., Hurt, E. M., Rosenwald, A., Giltnane, J. M., Yang, L., Zhao, H., Calame, K. et al. (2002). Blimp-1 orchestrates plasma cell differentiation by extinguishing the mature B cell gene expression program. Immunity 17, 51-62. Shaffer, A. L. and Schlissel, M. S. (1997). A truncated heavy chain protein relieves the requirement for surrogate light chains in early B cell development. J Immunol 159, 1265-75. Shafi, R., Iyer, S. P., Ellies, L. G., O'Donnell, N., Marek, K. W., Chui, D., Hart, G. W. and Marth, J. D. (2000). The O-GlcNAc transferase gene resides on the X chromosome and is essential for embryonic stem cell viability and mouse ontogeny. Proc Natl Acad Sci U S A 97, 5735-9.

174
Shaper, J. H. and Shaper, N. L. (1992). Enzymes associated with glycosylation. Current
Opinion Structural Biology 2, 701-709. Shapiro-Shelef, M., Lin, K. I., McHeyzer-Williams, L. J., Liao, J., McHeyzer-Williams, M. G. and Calame, K. (2003). Blimp-1 is required for the formation of immunoglobulin secreting plasma cells and pre-plasma memory B cells. Immunity 19, 607-20. Shen, X., Bozek, G., Pinkert, C. A. and Storb, U. (1999). The C(H)1 and transmembrane domains of mu in the context of a gamma2b transgene do not suffice to promote B cell maturation. Int Immunol 11, 1663-71. Shields, R. L., Lai, J., Keck, R., O'Connell, L. Y., Hong, K., Meng, Y. G., Weikert, S. H. and Presta, L. G. (2002). Lack of fucose on human IgG1 N-linked oligosaccharide improves binding to human Fcgamma RIII and antibody-dependent cellular toxicity. J Biol Chem 277, 26733-40. Shiloh, Y. (2003). ATM and related protein kinases: safeguarding genome integrity. Nat
Rev Cancer 3, 155-68. Shimizu, T., Mundt, C., Licence, S., Melchers, F. and Martensson, I. L. (2002). VpreB1/VpreB2/lambda 5 triple-deficient mice show impaired B cell development but functional allelic exclusion of the IgH locus. J Immunol 168, 6286-93. Shinkai, Y., Rathbun, G., Lam, K. P., Oltz, E. M., Stewart, V., Mendelsohn, M., Charron, J., Datta, M., Young, F., Stall, A. M. et al. (1992). RAG-2-deficient mice lack mature lymphocytes owing to inability to initiate V(D)J rearrangement. Cell 68, 855-67. Shinkawa, T., Nakamura, K., Yamane, N., Shoji-Hosaka, E., Kanda, Y., Sakurada, M., Uchida, K., Anazawa, H., Satoh, M., Yamasaki, M. et al. (2003). The absence of fucose but not the presence of galactose or bisecting N-acetylglucosamine of human IgG1 complex-type oligosaccharides shows the critical role of enhancing antibody-dependent cellular cytotoxicity. J
Biol Chem 278, 3466-73. Shinkura, R., Ito, S., Begum, N. A., Nagaoka, H., Muramatsu, M., Kinoshita, K., Sakakibara, Y., Hijikata, H. and Honjo, T. (2004). Separate domains of AID are required for somatic hypermutation and class-switch recombination. Nat Immunol 5, 707-12. Shinkura, R., Tian, M., Smith, M., Chua, K., Fujiwara, Y. and Alt, F. W. (2003). The influence of transcriptional orientation on endogenous switch region function. Nat Immunol 4, 435-41. Sieckmann, D. G., Finkelman, F. D. and Scher, I. (1982). IgD as a receptor in signaling the proliferation of mouse B-lymphocytes. Ann N Y Acad Sci 399, 277-89. Silberstein, S. and Gilmore, R. (1996). Biochemistry, molecular biology, and genetics of the oligosaccharyltransferase. Faseb J 10, 849-58. Simons, K. and Toomre, D. (2000). Lipid rafts and signal transduction. Nat Rev Mol Cell
Biol 1, 31-9. Sirac, C., Carrion, C., Duchez, S., Comte, I. and Cogne, M. (2006). Light chain inclusion permits terminal B cell differentiation and does not necessarily result in autoreactivity. Proc Natl Acad Sci U S A 103, 7747-52. Sleckman, B. P., Gorman, J. R. and Alt, F. W. (1996). Accessibility control of antigen-receptor variable-region gene assembly: role of cis-acting elements. Annu Rev Immunol 14, 459-81. Sonoda, E., Pewzner-Jung, Y., Schwers, S., Taki, S., Jung, S., Eilat, D. and Rajewsky, K. (1997). B cell development under the condition of allelic inclusion. Immunity 6, 225-33. Spanopoulou, E., Roman, C. A., Corcoran, L. M., Schlissel, M. S., Silver, D. P., Nemazee, D., Nussenzweig, M. C., Shinton, S. A., Hardy, R. R. and Baltimore, D. (1994).

175
Functional immunoglobulin transgenes guide ordered B-cell differentiation in Rag-1-deficient mice. Genes Dev 8, 1030-42. Spillmann, D., Witt, D. and Lindahl, U. (1998). Defining the interleukin-8-binding domain of heparan sulfate. J Biol Chem 273, 15487-93. Spiro, R. G. (2002). Protein glycosylation: nature, distribution, enzymatic formation, and disease implications of glycopeptide bonds. Glycobiology 12, 43R-56R. Staal, F. J. and Clevers, H. C. (2003). Wnt signaling in the thymus. Curr Opin Immunol 15, 204-8. Stall, A. M., Wells, S. M. and Lam, K. P. (1996). B-1 cells: unique origins and functions. Semin Immunol 8, 45-59. Stavnezer-Nordgren, J. and Sirlin, S. (1986). Specificity of immunoglobulin heavy chain switch correlates with activity of germline heavy chain genes prior to switching. Embo J 5, 95-102. Stavnezer, J. (2000). Immunology. A touch of antibody class. Science 288, 984-5. Stevens, F. J. (2000). Four structural risk factors identify most fibril-forming kappa light chains. Amyloid 7, 200-11. Stillman, B. N., Hsu, D. K., Pang, M., Brewer, C. F., Johnson, P., Liu, F. T. and Baum, L. G. (2006). Galectin-3 and galectin-1 bind distinct cell surface glycoprotein receptors to induce T cell death. J Immunol 176, 778-89. Storb, U. and Arp, B. (1983). Methylation patterns of immunoglobulin genes in lymphoid cells: correlation of expression and differentiation with undermethylation. Proc Natl
Acad Sci U S A 80, 6642-6. Stringer, S. E. and Gallagher, J. T. (1997). Specific binding of the chemokine platelet factor 4 to heparan sulfate. J Biol Chem 272, 20508-14. Sturm, A., Lensch, M., Andre, S., Kaltner, H., Wiedenmann, B., Rosewicz, S., Dignass, A. U. and Gabius, H. J. (2004). Human galectin-2: novel inducer of T cell apoptosis with distinct profile of caspase activation. J Immunol 173, 3825-37. Su, T. T. and Rawlings, D. J. (2002). Transitional B lymphocyte subsets operate as distinct checkpoints in murine splenic B cell development. J Immunol 168, 2101-10. Sugahara, K. and Kitagawa, H. (2002). Heparin and heparan sulfate biosynthesis. IUBMB Life 54, 163-75. Suzuki, H., Matsuda, S., Terauchi, Y., Fujiwara, M., Ohteki, T., Asano, T., Behrens, T. W., Kouro, T., Takatsu, K., Kadowaki, T. et al. (2003). PI3K and Btk differentially regulate B cell antigen receptor-mediated signal transduction. Nat Immunol 4, 280-6. Tanaka, Y., Fujii, K., Hubscher, S., Aso, M., Takazawa, A., Saito, K., Ota, T. and Eto, S. (1998). Heparan sulfate proteoglycan on endothelium efficiently induces integrin-mediated T cell adhesion by immobilizing chemokines in patients with rheumatoid synovitis. Arthritis Rheum 41, 1365-77. Tanigaki, K., Han, H., Yamamoto, N., Tashiro, K., Ikegawa, M., Kuroda, K., Suzuki, A., Nakano, T. and Honjo, T. (2002). Notch-RBP-J signaling is involved in cell fate determination of marginal zone B cells. Nat Immunol 3, 443-50. Tanigaki, K. and Honjo, T. (2007). Regulation of lymphocyte development by Notch signaling. Nat Immunol 8, 451-6. Tashiro, J., Kinoshita, K. and Honjo, T. (2001). Palindromic but not G-rich sequences are targets of class switch recombination. Int Immunol 13, 495-505. Tedford, K., Nitschke, L., Girkontaite, I., Charlesworth, A., Chan, G., Sakk, V., Barbacid, M. and Fischer, K. D. (2001). Compensation between Vav-1 and Vav-2 in B cell development and antigen receptor signaling. Nat Immunol 2, 548-55.

176
Theocharis, A. D., Seidel, C., Borset, M., Dobra, K., Baykov, V., Labropoulou, V., Kanakis, I., Dalas, E., Karamanos, N. K., Sundan, A. et al. (2006). Serglycin constitutively secreted by myeloma plasma cells is a potent inhibitor of bone mineralization in vitro. J Biol
Chem 281, 35116-28. Thompson, A., Timmers, E., Schuurman, R. K. and Hendriks, R. W. (1995). Immunoglobulin heavy chain germ-line JH-C mu transcription in human precursor B lymphocytes initiates in a unique region upstream of DQ52. Eur J Immunol 25, 257-61. Thompson, L. D., Pantoliano, M. W. and Springer, B. A. (1994). Energetic characterization of the basic fibroblast growth factor-heparin interaction: identification of the heparin binding domain. Biochemistry 33, 3831-40. Tiegs, S. L., Russell, D. M. and Nemazee, D. (1993). Receptor editing in self-reactive bone marrow B cells. J Exp Med 177, 1009-20. Tonegawa, S. (1983). Somatic generation of antibody diversity. Nature 302, 575-81. Toyama-Sorimachi, N., Sorimachi, H., Tobita, Y., Kitamura, F., Yagita, H., Suzuki, K. and Miyasaka, M. (1995). A novel ligand for CD44 is serglycin, a hematopoietic cell lineage-specific proteoglycan. Possible involvement in lymphoid cell adherence and activation. J
Biol Chem 270, 7437-44. Toyama, H., Okada, S., Hatano, M., Takahashi, Y., Takeda, N., Ichii, H., Takemori, T., Kuroda, Y. and Tokuhisa, T. (2002). Memory B cells without somatic hypermutation are generated from Bcl6-deficient B cells. Immunity 17, 329-39. Tsukada, S., Sugiyama, H., Oka, Y. and Kishimoto, S. (1990). Estimation of D segment usage in initial D to JH joinings in a murine immature B cell line. Preferential usage of DFL16.1, the most 5' D segment and DQ52, the most JH-proximal D segment. J Immunol 144, 4053-9. Tumova, S., Woods, A. and Couchman, J. R. (2000). Heparan sulfate chains from glypican and syndecans bind the Hep II domain of fibronectin similarly despite minor structural differences. J Biol Chem 275, 9410-7. Turner, M., Mee, P. J., Costello, P. S., Williams, O., Price, A. A., Duddy, L. P., Furlong, M. T., Geahlen, R. L. and Tybulewicz, V. L. (1995). Perinatal lethality and blocked B-cell development in mice lacking the tyrosine kinase Syk. Nature 378, 298-302. Uhlin-Hansen, L., Wik, T., Kjellen, L., Berg, E., Forsdahl, F. and Kolset, S. O. (1993). Proteoglycan metabolism in normal and inflammatory human macrophages. Blood 82, 2880-9. Umana, P., Jean-Mairet, J., Moudry, R., Amstutz, H. and Bailey, J. E. (1999). Engineered glycoforms of an antineuroblastoma IgG1 with optimized antibody-dependent cellular cytotoxic activity. Nat Biotechnol 17, 176-80. Van den Steen, P., Rudd, P. M., Dwek, R. A., Van Damme, J. and Opdenakker, G. (1998). Cytokine and protease glycosylation as a regulatory mechanism in inflammation and autoimmunity. Adv Exp Med Biol 435, 133-43. van Horssen, J., Wesseling, P., van den Heuvel, L. P., de Waal, R. M. and Verbeek, M. M. (2003). Heparan sulphate proteoglycans in Alzheimer's disease and amyloid-related disorders. Lancet Neurol 2, 482-92. Venkitaraman, A. R., Williams, G. T., Dariavach, P. and Neuberger, M. S. (1991). The B-cell antigen receptor of the five immunoglobulin classes. Nature 352, 777-81. Vidal y Plana, R. R. and Karzel, K. (1980). [Glucosamine: its value for the metabolism of articular cartilage. 1. Biochemistry of proteoglycans, studies on in-vitro cultures of embryonal mouse fibroblasts and bone germs]. Fortschr Med 98, 557-62. Vijay, I. K. (1998). Developmental and hormonal regulation of protein N-glycosylation in the mammary gland. J Mammary Gland Biol Neoplasia 3, 325-36.

177
Vilen, B. J., Famiglietti, S. J., Carbone, A. M., Kay, B. K. and Cambier, J. C. (1997). B cell antigen receptor desensitization: disruption of receptor coupling to tyrosine kinase activation. J Immunol 159, 231-43. Vishwakarma, R. A. and Menon, A. K. (2005). Flip-flop of glycosylphosphatidylinositols (GPI's) across the ER. Chem Commun (Camb), 453-5. Waisman, A., Kraus, M., Seagal, J., Ghosh, S., Melamed, D., Song, J., Sasaki, Y., Classen, S., Lutz, C., Brombacher, F. et al. (2007). IgG1 B cell receptor signaling is inhibited by CD22 and promotes the development of B cells whose survival is less dependent on Ig alpha/beta. J Exp Med 204, 747-58. Wakabayashi, C., Adachi, T., Wienands, J. and Tsubata, T. (2002). A distinct signaling pathway used by the IgG-containing B cell antigen receptor. Science 298, 2392-5. Walker, J. R., Corpina, R. A. and Goldberg, J. (2001). Structure of the Ku heterodimer bound to DNA and its implications for double-strand break repair. Nature 412, 607-14. Wall, R. and Kuehl, M. (1983). Biosynthesis and regulation of immunoglobulins. Annu
Rev Immunol 1, 393-422. Wang, H. Y. and Malbon, C. C. (2003). Wnt signaling, Ca2+, and cyclic GMP: visualizing Frizzled functions. Science 300, 1529-30. Wang, L. D. and Clark, M. R. (2003). B-cell antigen-receptor signalling in lymphocyte development. Immunology 110, 411-20. Wang, Y., Lee, G. F., Kelley, R. F. and Spellman, M. W. (1996a). Identification of a GDP-L-fucose:polypeptide fucosyltransferase and enzymatic addition of O-linked fucose to EGF domains. Glycobiology 6, 837-42. Wang, Y., Macke, J. P., Abella, B. S., Andreasson, K., Worley, P., Gilbert, D. J., Copeland, N. G., Jenkins, N. A. and Nathans, J. (1996b). A large family of putative transmembrane receptors homologous to the product of the Drosophila tissue polarity gene frizzled. J Biol Chem 271, 4468-76. Wang, Y., Shao, L., Shi, S., Harris, R. J., Spellman, M. W., Stanley, P. and Haltiwanger, R. S. (2001). Modification of epidermal growth factor-like repeats with O-fucose. Molecular cloning and expression of a novel GDP-fucose protein O-fucosyltransferase. J Biol
Chem 276, 40338-45. Ward, I. M., Reina-San-Martin, B., Olaru, A., Minn, K., Tamada, K., Lau, J. S., Cascalho, M., Chen, L., Nussenzweig, A., Livak, F. et al. (2004). 53BP1 is required for class switch recombination. J Cell Biol 165, 459-64. Warren, W. D. and Berton, M. T. (1995). Induction of germ-line gamma 1 and epsilon Ig gene expression in murine B cells. IL-4 and the CD40 ligand-CD40 interaction provide distinct but synergistic signals. J Immunol 155, 5637-46. Watt, G. M., Lund, J., Levens, M., Kolli, V. S., Jefferis, R. and Boons, G. J. (2003). Site-specific glycosylation of an aglycosylated human IgG1-Fc antibody protein generates neoglycoproteins with enhanced function. Chem Biol 10, 807-14. Wegrowski, Y. and Maquart, F. X. (2004). Involvement of stromal proteoglycans in tumour progression. Crit Rev Oncol Hematol 49, 259-68. Weichhold, G. M., Ohnheiser, R. and Zachau, H. G. (1993). The human immunoglobulin kappa locus consists of two copies that are organized in opposite polarity. Genomics 16, 503-11. Weiser, P., Muller, R., Braun, U. and Reth, M. (1997). Endosomal targeting by the cytoplasmic tail of membrane immunoglobulin. Science 276, 407-9. Weiser, P., Riesterer, C. and Reth, M. (1994). The internalization of the IgG2a antigen receptor does not require the association with Ig-alpha and Ig-beta but the activation of protein tyrosine kinases does. Eur J Immunol 24, 665-71.

178
Weller, S., Reynaud, C. A. and Weill, J. C. (2005). Splenic marginal zone B cells in humans: where do they mutate their Ig receptor? Eur J Immunol 35, 2789-92. Willert, K. and Nusse, R. (1998). Beta-catenin: a key mediator of Wnt signaling. Curr
Opin Genet Dev 8, 95-102. Williams, A. F. and Barclay, A. N. (1988). The immunoglobulin superfamily--domains for cell surface recognition. Annu Rev Immunol 6, 381-405. Wilson, A., MacDonald, H. R. and Radtke, F. (2001). Notch 1-deficient common lymphoid precursors adopt a B cell fate in the thymus. J Exp Med 194, 1003-12. Wortis, H. H. and Berland, R. (2001). Cutting edge commentary: origins of B-1 cells. J
Immunol 166, 2163-6. Yamamura, K., Kudo, A., Ebihara, T., Kamino, K., Araki, K., Kumahara, Y. and Watanabe, T. (1986). Cell-type-specific and regulated expression of a human gamma 1 heavy-chain immunoglobulin gene in transgenic mice. Proc Natl Acad Sci U S A 83, 2152-6. Yamanashi, Y., Fukui, Y., Wongsasant, B., Kinoshita, Y., Ichimori, Y., Toyoshima, K. and Yamamoto, T. (1992). Activation of Src-like protein-tyrosine kinase Lyn and its association with phosphatidylinositol 3-kinase upon B-cell antigen receptor-mediated signaling. Proc Natl Acad Sci U S A 89, 1118-22. Yamanashi, Y., Kakiuchi, T., Mizuguchi, J., Yamamoto, T. and Toyoshima, K. (1991). Association of B cell antigen receptor with protein tyrosine kinase Lyn. Science 251, 192-4. Yamane-Ohnuki, N., Kinoshita, S., Inoue-Urakubo, M., Kusunoki, M., Iida, S., Nakano, R., Wakitani, M., Niwa, R., Sakurada, M., Uchida, K. et al. (2004). Establishment of FUT8 knockout Chinese hamster ovary cells: an ideal host cell line for producing completely defucosylated antibodies with enhanced antibody-dependent cellular cytotoxicity. Biotechnol
Bioeng 87, 614-22. Yamauchi, M., Noyes, C., Kuboki, Y. and Mechanic, G. L. (1982). Collagen structural microheterogeneity and a possible role for glycosylated hydroxylysine in type I collagen. Proc
Natl Acad Sci U S A 79, 7684-8. Yancopoulos, G. D. and Alt, F. W. (1985). Developmentally controlled and tissue-specific expression of unrearranged VH gene segments. Cell 40, 271-81. Yancopoulos, G. D. and Alt, F. W. (1986). Regulation of the assembly and expression of variable-region genes. Annu Rev Immunol 4, 339-68. Yang, R. Y. and Liu, F. T. (2003). Galectins in cell growth and apoptosis. Cell Mol Life
Sci 60, 267-76. Yoshino, T., Kondo, E., Cao, L., Takahashi, K., Hayashi, K., Nomura, S. and Akagi, T. (1994). Inverse expression of bcl-2 protein and Fas antigen in lymphoblasts in peripheral lymph nodes and activated peripheral blood T and B lymphocytes. Blood 83, 1856-61. Yu, K., Chedin, F., Hsieh, C. L., Wilson, T. E. and Lieber, M. R. (2003). R-loops at immunoglobulin class switch regions in the chromosomes of stimulated B cells. Nat Immunol 4, 442-51. Yuan, D., Witte, P. L., Tan, J., Hawley, J. and Dang, T. (1996). Regulation of IgM and IgD heavy chain gene expression: effect of abrogation of intergenic transcriptional termination. J
Immunol 157, 2073-81. Zachau, H. G. (1993). The immunoglobulin kappa locus-or-what has been learned from looking closely at one-tenth of a percent of the human genome. Gene 135, 167-73. Zarrin, A. A., Alt, F. W., Chaudhuri, J., Stokes, N., Kaushal, D., Du Pasquier, L. and Tian, M. (2004). An evolutionarily conserved target motif for immunoglobulin class-switch recombination. Nat Immunol 5, 1275-81.

179
Zarrin, A. A., Tian, M., Wang, J., Borjeson, T. and Alt, F. W. (2005). Influence of switch region length on immunoglobulin class switch recombination. Proc Natl Acad Sci U S A 102, 2466-70. Zhang, L., David, G. and Esko, J. D. (1995). Repetitive Ser-Gly sequences enhance heparan sulfate assembly in proteoglycans. J Biol Chem 270, 27127-35. Zhang, Z., Espinoza, C. R., Yu, Z., Stephan, R., He, T., Williams, G. S., Burrows, P. D., Hagman, J., Feeney, A. J. and Cooper, M. D. (2006). Transcription factor Pax5 (BSAP) transactivates the RAG-mediated V(H)-to-DJ(H) rearrangement of immunoglobulin genes. Nat
Immunol 7, 616-24. Zhou, D., Chen, C., Jiang, S., Shen, Z., Chi, Z. and Gu, J. (1998). Expression of beta1,4-galactosyltransferase in the development of mouse brain. Biochim Biophys Acta 1425, 204-8. Zimmermann, P. and David, G. (1999). The syndecans, tuners of transmembrane signaling. Faseb J 13 Suppl, S91-S100.

180
RESUME
La capacité des lymphocytes à répondre de manière spécifique à un grand nombre
d’antigènes étrangers, est le résultat d’un processus de différenciation à l’issue duquel chaque
lymphocyte exprime un récepteur unique à l’antigène. Une longue série de processus d’activation
et de coopération cellulaire faisant intervenir de nombreuses protéines glycosylées ou pas, est
nécessaire avant d’aboutir à la sécrétion d’un anticorps spécifique par une cellule de la lignée
lymphoïde B. Cette réponse présuppose l’expression à la surface cellulaire d’une forme
membranaire de l’immunoglobuline au sein du récepteur des cellules B pour l’antigène (BCR),
dont la spécificité est définie pour chaque clone B.
Au cours de ma thèse, je me suis intéressée à la fonction même du BCR et notamment du
BCR commuté vers IgA et IgE afin de savoir si une chaîne lourde α ou ε était capable de jouer un
rôle similaire à celui d’une chaîne lourde µ et de promouvoir au sein du récepteur B (soit de
classe IgA soit de classe IgE) le développement des cellules jusqu’au stade final du plasmocyte
sécréteur d’anticorps. Pour cela, nous avons réalisé plusieurs modèles de knock-in portant des
BCR modifiés où le gène Cα1 humain dans un cas et Cε humain dans un autre, ont été intégrés
par recombinaison homologue au niveau de la région switch µ afin d’obtenir une lignée de souris
transgéniques n’exprimant que des IgA ou des IgE.
Dans une autre perspective d’étude des phénomènes pouvant modifier la réactivité des
récepteurs de surface de la cellule B, nous nous sommes intéressés à la modulation au cours de la
différenciation B des phénomènes de glycosylation, ceux-ci constituant les éléments majeurs de
maturation post-traductionnelle des protéines et des lipides. Après une première étude
glycotranscriptomique, mettant en valeur l’expression de deux gènes distincts tous deux
impliqués dans la biosynthèse des glycosaminoglycannes, CsGalNacT1 et Extl1, nous avons
voulu étudier d’une manière plus approfondie le rôle de ces enzymes in vivo au cours du
développement B en réalisant des lignées de souris transgéniques surexprimant de façon B-
spécifique ces deux gènes d’intérêt.





























