-
This article appeared in a journal published by Elsevier. The
attachedcopy is furnished to the author for internal non-commercial
researchand education use, including for instruction at the authors
institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling
orlicensing copies, or posting to personal, institutional or third
party
websites are prohibited.
In most cases authors are permitted to post their version of
thearticle (e.g. in Word or Tex form) to their personal website
orinstitutional repository. Authors requiring further
informationregarding Elseviers archiving and manuscript policies
are
encouraged to visit:
http://www.elsevier.com/authorsrights
-
Author's personal copy
Research paper
Systematics of a palaeoecologically signicant boreal Mesozoic
fossil wood genus,Xenoxylon Gothan
Marc Philippe a,, Frdric Thvenard a, Natalya Nosova b, Kyungsik
Kim c, Serge Naugolnykh d
a UMR 5276 of the CNRS and Universit Lyon 1, 7 rue Dubois,
F69622 Villeurbanne, Franceb Komarov Botanical Institute of the
Russian Academy of Sciences, ul. Prof. Popova, 2, St. Petersburg,
197376, Russiac Division of Biological Sciences, Chonbuk National
University, Jeonju, 561-756, South Koread Geological Institute of
Russian Academy of Sciences, 119017, Moscow, Pyzhevsky per. 7,
Russia
a b s t r a c ta r t i c l e i n f o
Article history:Received 19 October 2012Received in revised form
23 January 2013Accepted 29 January 2013Available online 20 February
2013
Keywords:fossil
woodXenoxylonLaurasiaMesozoicpalaeobiogeography
The genus Xenoxylon was established for gymnosperm-like fossil
wood with xenoxylean radial pitting andlarge oopores in its
cross-elds. It is restricted to the Mesozoic and to the Northern
Hemisphere, where itis commonly found. As an indicator of cool-wet
boreal climates, as suggested by its distribution, Xenoxylonis of
special interest to palaeoclimatology. Its systematics, however, is
poorly known with several speciesneeding re-evaluation. Xenoxylon
nomenclature is also still unresolved, with various synonyms used
for thesame taxa even in recent literature. Having reviewed several
types and a large number of specimens weperformed a taxonomical and
nomenclatural reappraisal of all species known to date. The results
of thisreview are presented here, together with a taxonomical key.
For most taxa stratigraphical and geographicaldistributions have
also been re-evaluated.
2013 Elsevier B.V. All rights reserved.
1. Introduction
When, in 1905, Gothan coined the name Xenoxylon for a fossil
woodfrom Svalbard which puzzled him, he probably did not imagine
that thisname had a prolic future. At this time Xenoxylon was known
only bysome poorly dated specimens from the Arctic. At present this
genus isknown to occur from the late Triassic through to the latest
Cretaceous,throughout the Northern Hemisphere (Laurasia), with the
noticeableexception of most of North America (Philippe et al.,
2009). No less than31 Xenoxylon species have been proposed (Table
1), and the genus is ofpalaeobiogeographical and palaeoecological
interest (Philippe andThvenard, 1996; Ding et al., 2000; Philippe
et al., 2009). Indeed its dis-tribution is restricted to Laurasia,
mostly at high palaeolatitudes, and ex-panded southwards during
cool intervals only, suggesting thatXenoxylonfavoured cooler/wetter
climates. Reliable indicators of terrestrial cli-mates are greatly
required for the understanding of Mesozoic climaticevolution.
Mineralogical, sedimentological and geochemical evidenceconrm that
Xenoxylon was restricted to cool-wet temperate
climates(Tchoumatchenco et al., 2008; Amiot et al., 2011; Selmeier
and Grosser,2011).
The taxonomy, systematics and nomenclature of the genus
are,however, largely unsorted. From a taxonomic view point, some of
theanatomical features of the genus that were originally considered
to be
of little signicance have been re-evaluated (Philippe et al.,
2009). Asfor the systematics, unexpected synonymies were recently
discovered(Philippe and Hayes, 2010), whereas some names published
by Chinesecolleagues have been overlooked. Few types have been
reviewed. Finallythe nomenclature of Xenoxylon needs reappraisal as
a putative earliersynonym (Sciadopityoxylon Schmalhausen; Philippe
and Bamford,2008) and several species names are still controversial
(e.g. Xenoxylonbarberi Seward).
Herewe present a reviewof the genus, with full systematic
treatment,taxonomical key and distribution data.
2. Material and methods
The material for Xenoxylonwas studied at several institutional
repos-itories listed with their respective abbreviations below and
between pa-rentheses the number of Xenoxylon samples studied
there:
BIN Komarov Botanical Institute of the Russian Academy
ofSciences (RAS), St. Petersburg, Russia (30)
BMNH Natural History Museum, London, UK (17)BUPM Berkeley
University Palaeontological Museum, USA (1)CNU Chonbuk National
University, South Korea (70)CU Chuo University, Tokyo, Japan (2)HMB
Humboldt Museum, Berlin, Germany (9)LPUL Laboratoire de
Palobotanique de l'Universit de Lyon,
France (30)MNHN Musum national d'Histoire naturelle, Paris,
France (2)NHMD Natural History Museum of Denmark (38)
Review of Palaeobotany and Palynology 193 (2013) 128140
Corresponding author.E-mail addresses: [email protected]
(M. Philippe), [email protected]
(F. Thvenard), [email protected] (N. Nosova),
[email protected] (K. Kim),[email protected] (S.
Naugolnykh).
0034-6667/$ see front matter 2013 Elsevier B.V. All rights
reserved.http://dx.doi.org/10.1016/j.revpalbo.2013.01.013
Contents lists available at SciVerse ScienceDirect
Review of Palaeobotany and Palynology
j ourna l homepage: www.e lsev ie r .com/ locate / revpa lbo
-
Author's personal copy
SMNH Swedish Museum of Natural History, Stockholm, Sweden
(29)TUMS Tohoku University Museum in Sendai, Japan (32)UTM
University of Tokyo Museum (15).
Other abbreviations used:
ISBT Institut fr spezielle Botanik der Universitt
Tbingen,Germany
NIGPAS Nanjing Institute of Geology and Palaeontology,
AcademiaSinica, China
SIGMR Shenyang Institute of Geology andMineral Resources,
Liaoning,China.
More than 285 samples of Xenoxylon were studied
microscopically,either with scanning electronic microscopy, or
light microscopy, usingthin-sections or Collodion casts (Philippe
et al., 2006). For all the taxamentioned here protologues were
accessed. An extensive bibliographi-cal surveywas performed,which
to the best of our knowledge covers allpublished data for
Xenoxylon.
3. Results
3.1. Circumscription of Xenoxylon
Since the seminal work by Gothan (1905) the circumscription
ofXenoxylon has been little questioned. This genus is based on
theoccurrence of at least partially xenoxylean radial pitting in
thetracheids (strongly attened contiguous areolated pits;
Mller-Stoll,1951; Fig. 1) and a large fenestriform oopore in the
early wood cross-elds (characteristically two when a ray cell
terminal wall divides thecross-eld; Fig. 2). Krusel (1949) included
in Xenoxylon, woods witha mixed type of radial pitting and with 24
rounded oopores percross-eld. As clearly demonstrated by
Vogellehner (1967, 1968),such woods belong to distinct genera such
as Protocircoporoxylon
Vogellehner or Protopolyporoxylon Vogellehner. The term
xenoxyleanapplies to a type of tracheid radial pittingwhere pits
are contiguous andappear much attened. In order to quantify this
attening from slideS1765 at the SMNH (holotype for Xenoxylon
latiporosum; Philippe andCantrill, 2007) we measured the
longitudinal diameter/radial diameterratio for 34 radial pits. The
maximumwas 0.69 and the minimum 0.45,whilst the mean-value was 0.57
and the standard-deviation 0.05. Thisratio, also known as the
height/width ratio, is rarely lower than 0.6in the wood of most
extant species with araucarian radial pitting(sensu IAWA, 2004).
The 0.6 value is thus a limit below which a radialpit can be
characterised as xenoxylean (see illustration in Figs. 1 and
3).
Table 1The thirty-one species names proposed within the genus
Xenoxylon; those of which type was studied are underlined.
Species name Authority Reference Notes
arcticum Shilkina Shilkina and Yatsenko-Khmelevsky (1980) Nomen
nudumbarberi Seward (as Cupressinoxylon) Krusel (1949) p.p. a
synonym of X. huttonianumcanoasense Rau (as Cedroxylon) Krusel
(1949) Not a Xenoxylonconchylianum Fliche Fliche (1910) Type
lostellipticum Mller-Stoll Vogellehner (1968) A synonym of X.
huttonianumfuxinense Ding Ding et al. (2000)hopeiense Chang Chang
(1929)huolinhense Ding Ding et al. (2000)huttonianum Witham (as
Peuce) Philippe and Hayes (2010)jakutiense Shilkina Shilkina (1986)
A nomen nudum in Shilkina & Yatsenko-Khmelevsky, 1980japonicum
Vogellehner (not validly published in 1968) Suzuki and Terada
(1992) A synonym of X. meisterijurassicum Eckhold (as
Protopodocarpoxylon) Krusel (1949) Not a Xenoxylon; topotypes
seen[kurumaense] Tsunada Unpublished thesis (1980) Not a validly
published namelatiporosum Cramer Gothan (1905)[liaoningense] Duan
& Wang Duan et al. (1995) Not a validly published namemeisteri
Palibin & Jarmolenko Palibin and Jarmolenko (1932)moorei
Tidwell Tidwell et al. (1998) Not a Xenoxylonmorrisonii Medlyn
& Tidwell Medlyn and Tidwell (1975) Not a Xenoxylonnariwaense
Yamazaki, Tsunada & Koike Yamazaki et al. (1980) Type
lostparvipunctatum Vogellehner Vogellehner (1965)peideensis Zheng
& Zhang Zheng and Zhang (1982) As peidensephyllocladoides
Gothan Gothan (1906) Type lost, topotypes seenpseudoellipticum
Yamazaki & Tsunada Yamazaki and Tsunada (1981) Type
lostsaadawii Youssef Youssef (2002) Not a Xenoxylonshimakurai
Yamazaki & Tsunada Yamazaki and Tsunada in Yamazaki et al.
(1984) = X. meisteri; type lostsuljuctense Shilkina &
Khudayberdyev Shilkina and Khudaiberdyev (1971) Type losttomiense
Larichev Larishchev (1948) Probably not a Xenoxylon; type
losttsuruokai Nishida, Nishida, & Suzuki Nishida et al. (1993)
Not a Xenoxylonwattarianum Nishida & Nishida Nishida and
Nishida (1986)yatsenkoi Yunusov Yunusov in Gomolitzky et al. (1981)
Not a Xenoxylonyixianense Zhang & Shang Zhang and Shang
(1996)
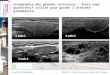
Fig. 1. Xenoxylon latiporosum (Cramer) Gothan; sample 1991/38 at
the HMB, Jurassic,Poland; typical xenoxylean radial pitting on
tracheids; note that on the narrow radialface of a tracheid, pits
are locally round and distant (a very rare feature for thisspecies
and the latiporosum group).
129M. Philippe et al. / Review of Palaeobotany and Palynology
193 (2013) 128140
-
Author's personal copy
3.2. Validity of previously published material
Among the 31 binomials proposed to date within Xenoxylon,
someare based on material which we consider dubious, are misplaced,
orare nomenclaturally invalid. Xenoxylon arcticum, described in
1980by Shilkina and Yatsenko-Khmelevsky, is considered to be a
nomennudum. Nothing in this publication identies a particular
specimenin Shilkina's collection, housed at the BIN
(St.-Petersburg), and wedid notnd any record of that namewhilst
checking Shilkina's collection.Xenoxylon (?) tomiense Larishchev
has not been considered as it wasassigned with doubt to the genus
Xenoxylon (Larishchev, 1948, 1957;Mller-Stoll and Schultze-Motel,
1988). As they have several pits pereld,Xenoxylon canoasense
(Rau)Krusel,Xenoxylon jurassicum (Eckhold)Krusel, Xenoxylon moorei
Tidwell, Xenoxylon morrisonense Medlyn etTidwell and Xenoxylon
yatsenkoi Yunusov in Gomolitzky et al. do not be-long to Xenoxylon
(Philippe and Thvenard, 1996 and references therein).Moreover
Xenoxylon saadawii Youssef from the late Jurassicearly Creta-ceous
of Gebel Kamil, Egypt, does not have xenoxylean radial
pitting(Youssef, 2002), and its cross-elds are typical for
Metapodocarpoxylon
libanoticum. Yamazaki and Tsunada (in Yamazaki et al., 1984)
describedXenoxylon shimakurai, a taxon in which the diagnosis
includes pith fea-tures. If only the secondary xylem is considered,
then X. shimakurai is an-atomically similar to Xenoxylon meisteri
because of its radial pitting anddense tangential pitting.
Xenoxylon yatsenkoi Yunusov in Gomolitzkyet al. (1981) has pointed
oopores in the early-wood cross-elds (pl. 30g. 4 of the protologue)
and although areolated pits in its tracheids areat-elliptic they
are not xenoxylean (Gomolitzky et al., 1981, pl. 28g. 4, pl. 30 g.
2). With kind permission of Prof. Harufumi Nishida thetype specimen
of Xenoxylon tsuruokai Nishida, Nishida et al. (1993) wasreviewed
(although published as tsuruokae, this must be spelled astsuruokai
as this is dedicated to Mr. Shigeru Tsuruoka); we observedonly
araucarian radial pitting, and although cross-elds sometimes
dis-play a large pale window (presumably poorly preserved cell
wall) wellpreserved cross-elds with only one small bordered
oculipore were alsoobserved in the early wood. This wood also has
much axial parenchyma,whereas such parenchyma is generally
exceptional in Xenoxylon.
3.3. Sub-generic morphogroups
Two informal groups are usually recognised within Xenoxylon:
thelatiporosum-group and the phyllocladoides-group. These groupscan
be separated on the type of pitting on the radial walls of
earlywood tracheids (Figs. 3 and 4; Table 2). In the
latiporosum-groupthe radial pitting is strictly xenoxylean, with
only few exceptions. Inthe phyllocladoides-group pits are locally
xenoxylean, but otherwisespaced and rounded. Recent discoveries in
north-eastern China ledDing et al. (2000) to the proposal of a
third group of species, theliaoningense-group. Since [Xenoxylon
liaoningense] is not a validlypublished name and as this informal
X. liaoningense group hasbeen mentioned only by Ding et al., we
prefer to use here the nameof meisteri-group, which reects the name
of the rst describedand validly published species of this group.
These three informalgroups have no legitimate nomenclatural
status.
Radial pitting in the meisteri-group usually appears
latiporosum-like at rst glance, but close observation reveals
biseriate pitting and/orpit clusters. In addition to these
characteristic pit arrangements sometracheid portions appear
unpitted (Fig. 5). Pit clusters and unpittedtracheid portions (but
not biseriate pits) are also observed in thelatiporosum-group, even
in the holotype, but are always extremelyrare and local. The
meisteri-group is also characterised by frequent
Fig. 2. Xenoxylon phyllocladoides Gothan, sample 117 in Gothan,
slide S00192-02 at theSMNH; cross-elds at the late wood/early wood
transition; note that in the late woodpits are bordered, and a eld
with two pits.
Fig. 3. Interpretative diagram of typical tracheid radial
pitting of the latiporosumgroup.
Fig. 4. Interpretative diagram of typical tracheid radial
pitting of the phyllocladoidesgroup.
130 M. Philippe et al. / Review of Palaeobotany and Palynology
193 (2013) 128140
-
Author's personal copy
abundant tangential pitting. Although in the latiporosum-group
tan-gential pits occasionally occur, these are always uniseriate,
round anddistant. In the meisteri-group the tangential pitting,
when present, iscommonly bi-tri-seriate with crowded alternate
pits. For woods of themeisteri-group, however, even within a single
specimen both radialand tangential pittings are usually
variable.
Within each of the three morphogroups described in Table
2,species are distinguished mainly on the basis of features
usuallyregarded as of little signicance ray height, pit size,
occurrence ofaxial parenchyma or of tangential pitting. Diameter of
tracheid radialpits was once considered to be a feature with
systematic value(Vogellehner, 1965), but it is now clear that it
varies considerablywith both the tracheid width and the nature of
fossilisation. The pitdiameter/tracheid width ratio might be a more
reliable feature, butagain it is variable, even within a single
specimen. Here we adopt aconservative approach andmerge taxa only
if they are found to be an-atomically completely similar. Several
morphospecies differ only inone character state. Although it would
have been more rigorous to beable to distinguish species on two or
three specic character states, ifsuch policy is applied to the set
of the estimated 350 Xenoxylon speci-mens we considered (rsthand
experience plus literature), only threespecies would be recognised.
The data set yet available (350 specimensfor the whole Northern
Hemisphere and a ca. 155 My time interval) istoo limited to
convincingly support such merging.
Finally, several Asian specimens related toXenoxylondisplay
abundantsepta-like structures. They have been variously interpreted
as attenedtylose-like structures, septae, cracks or fungal hyphae
(Boureau, 1950;Arnold, 1953; Watari, 1960). Although thin (ca. 1 or
2 m in thickness),they obstruct the whole tracheid, as
evidencedwhen fossilisation slightlyturned them (Fig. 6). As yet we
have been unable to interpret them andfeel that they do not carry
any taxonomical interest.
3.4. A taxonomical key for Xenoxylon species
Ding Qiu-Hong (in Ding et al., 2000) published a key to
Xenoxylon inChinese that included information from some previously
publishedkeys (e.g. Krusel, 1949; Nishida and Nishida, 1986). Ding
et al., howev-er, did not have the opportunity to study the type of
material outside ofChina, and relied for most taxa on original
publication (protologue)only.
Table 2Three groups recognised within Xenoxylon Gothan; these
groups have no legitimate nomenclatural status.
Tracheid pitting Latiporosum group Meisteri group
Phyllocladoides group
Normal radial tracheid pitting Strictly xenoxylean, in
longuninterrupted uniseriate rows
Generally xenoxylean, uniseriate rowslocally interrupted,
clusters
Locally xenoxylean, otherwise mixed type,mostly uniseriate
Biseriate radial pitting Absent In some cases, alternate or
opposite, spaced orcontiguous
Tangential Rare, uniseriate, spaced Occasional to common, up to
triseriate, alternate,generally contiguous
Rare, uniseriate, spaced
Fig. 5. Interpretative diagram of typical tracheid radial
pitting of the meisteri group.
Fig. 6. Xenoxylon meisteri Palibin et Jarmolenko, material
studied by Suzuki and Terada(1992), at the TUMS; A sample 53907,
septae, hyphae and/or thylosis?; B sample53903, tilted septa
(arrow) showing its bidimensional nature.
131M. Philippe et al. / Review of Palaeobotany and Palynology
193 (2013) 128140
-
Author's personal copy
Here we provide a translation of Ding Qiu-Hong's key withseveral
updates and modications, since some species were notincluded by
Ding et al. (2000). We also took into account otherprevious
reappraisals (Vogellehner, 1965; Serra, 1966; Shilkina
andKhudaiberdyev, 1971). Although we have not been able to
studyrecently described Chinese species, we reviewed most of the
availableXenoxylon type material (Table 1).
Features described in the revised key presented below should
onlybe used for non-traumatic areas where tracheids are opposite
betweentwo adjacent rows, because when tracheids alternate they
developnarrow radial faces which can bear atypical pitting (Fig.
1).
1 Early wood bordered pits on radial tracheid walls: small,
typically1016 mhigh1221 mwide; pits: uniseriate in long
uninterruptedrows, not much attened; biseriate radial pits:
alternate andcrowded; tangential tracheid pitting: completely
lacking Xenoxylonparvipunctatum Vogellehner
1 Early wood bordered pits on radial tracheid walls: large,
typically1538 m high2040 mwide 2, 2, and 2
2 Early wood bordered pits on radial tracheid walls:
alwaysuniseriate, contiguous and compressed (xenoxylean radial
pitting),very rarely spaced (only in the narrowest tracheids);
tangential tracheidwalls: rarely pitted, when present: uniseriate,
round and distant Xenoxylon latiporosum group (Fig. 3) 3 and 3
3 Early wood bordered pits: generally covering only half of
tracheidradial wall width Xenoxylon peideensis Zheng &
Zhang
3 Early wood bordered pits: generally covering the whole widthof
tracheid radial walls 4 and 4
4 Some rays with traumatic resin canals/pockets
Xenoxylonyixianense Zhang & Shang
4 All rays uniseriate 5, 5, and 5 (similar species)5 Rays (1) 26
(10) cells high Xenoxylon watarianum Nishida &
Nishida5 Rays mostly 510 cells high, at most 16 cells
Xenoxylon
conchylianum Fliche5 Rays mostly 1020 cells high, rarely up to
25 cells Xenoxylon
latiporosum (Cramer) Gothan
2 Early wood bordered pits on radial tracheid walls: contiguous
orspaced, round or elliptic, only locally xenoxylean; tracheid
tangentialwalls: rarely pitted, when present these tangential pits
are uniseriateand spaced Xenoxylon phyllocladoides group (Fig. 4) 6
and 6
6 Rims of Sanio: present 7 and 77 Axial parenchyma: present;
radial tracheid pitting: uni-biseriate
Xenoxylon hopeiense Chang7 Axial parenchyma: absent; radial
tracheid pitting: always
uniseriate Xenoxylon jakutiense Shilkina6 Rims of Sanio: absent
8 and 88 Early wood cross-eld pits usually 12 (3) times as wide
than
high Xenoxylon phyllocladoides Gothan8 At least some early wood
cross-eld pits: 45 times as wide as
high Xenoxylon huttonianum (Witham) Philippe & Hayes
2 Early wood bordered pits on radial tracheid walls: clearly
inter-mediate; xenoxylean type frequent, intermingled with clusters
of 39radial pits (those at tips being smaller andmore rounded)
and/or signif-icant proportion of bi-triseriate pit rows and/or
area of unpitted tra-cheid wall; tangential tracheid wall: often
pitted, usually with someuni-triseriate contiguous alternate pits
Xenoxylon meisteri group(Fig. 5) 9 and 9
9 Bordered pits on radial tracheid walls: always uniseriate
(exceptfor traumatic areas) 10 and 10
10 Short clusters of 35 pits frequent Xenoxylon huolinhenseDing
in Ding, Zheng & Zhang
10 Clusters: usually much longer; radial pitting:
appearinglatiporosum-like at rst glance 11 and 11
11" Rays: (1) 38 (18) cells high Xenoxylon meisteri Palibin
&Jarmolenko
11 Rays: up to 4050 cells high Xenoxylon japonicumVogellehner ex
Suzuki & Terada
9 Bordered pits on radial tracheid walls of untraumatised
wood:sometimes biseriate or even triseriate 12, 12, and 12
12 Uniseriate radial pit areolae: mostly round or elliptical,
andspaced; biseriate pits: mostly opposite 13 and 13
13 Rays: (5) 3040 (50) cells high [Xenoxylon liaoningense]Duan
& Wang
13 Rays: (1) 26 (12) cells high Xenoxylon nariwaenseYamazaki,
Tsunada & Koike
12 Uniseriate radial pit areolae: generally compressed;
biseriatepits: generally alternate Xenoxylon fuxinense Ding in
Ding, Zheng& Zhang
12 Uniseriate radial pit areolae: very compressed; bi-triseriate
pits:always alternate Xenoxylon suljuctense Shilkina &
Chudajberdiev
4. Systematic classication
The systematic classication presented here relies as much as
pos-sible on the reappraisal of types and published specimens. In
the fewcases for which it has not been possible to study the
material we havehad to rely on the protologue. Names between square
brackets arenot validly published.
For distribution data we used only well documented data
(eitherfrom published or unpublished sources), which was not
entirely sat-isfactory since many publications do not provide
information on all ofthe points mentioned in the key.
For each entry at specic level we use the following format:
Species name
Basionym and full bibliographical reference to the
protologueSynonymyNote(s)Type: repository; reappraised or not;
later typication(s) if anyDistribution: 1) source for the type;
2)rst appearance datum(FAD) lastappearance datum (LAD); and 3)
palaeogeographical range
4.1. Generic level
FOSSIL-GYMNOSPERMAEPINOPHYTA?Morphogenus Xenoxylon Gothan, Abh.
Knigl. Preuss. Geol.Landesanst. ser. 2. 44: 38. 1905.Type:
Xenoxylon latiporosum (Cramer) Gothan, Abh. Knigl. Preuss.Geol.
Landesanst. ser. 2, 44: 108. 1905. (Pinites latiporosus Cramer)
note that the combination is made by Gothan only in the Register(a
plant name index) of his thesis (1905).Diagnosis: Tracheidoxyls
without axial resin channels; axialparenchyma: occasional and then
sparse; radial tracheid pitting: at leastlocally xenoxylean, i.e.
with contiguous pits much wider than high;ray-cell walls: thin,
smooth, unpitted except for cross-elds where thereis one (rarely
two) window-like unbordered oopores that occupy thewhole eld in the
early-wood, sometimes more restricted and narrowlybordered in both
the late-wood and the reaction wood (slightly modi-ed from
Vogellehner, 1968).Synonymy: The oldest description of a fossil
wood with a Xenoxylonanatomy is that of Peuce huttoniana in 1833 by
Witham of Lartington(Philippe and Hayes, 2010). The name Peuce
Lindley et Hutton is apoorly dened generic name (unused for more
than a century) with di-vergent Mesozoic and Palaeozoic syntypes,
neotypied by Palaeozoicmaterial (Andrews, 1955), which should not
be used for Mesozoic
132 M. Philippe et al. / Review of Palaeobotany and Palynology
193 (2013) 128140
-
Author's personal copy
tracheidoxyls (Philippe and Bamford, 2008). The wood P.
huttonianawas subsequently assigned to genera Pinites Lindley et
Hutton,Elatoxylon Hartig and Cedroxylon Kraus in Schimper, but none
of thesenames should be used for fossil wood as their diagnoses,
their circum-scriptions, and their syntypes (several are polytypic)
are unclear(Bamford and Philippe, 2001; Philippe and Bamford, 2008;
Philippeand Hayes, 2010). Corresponding nomenclatural and
taxonomicalsynonymies are too uncertain to question the largely
automaticlectotypication of these generic names by Andrews (1955),
despitethe fact that they potentially have priority over
Xenoxylon.
In 1877 Schmalhausen briey described a wood collected by Gbelin
the Jurassic of the Mangyshlak Peninsula (Caspian Sea,
westernKazakhstan) as Sciadopityoxylon vetusta. Given the date,
this genericname is validly published (ICBN, art. 38.1; McNeill et
al., 2012) andshould be considered for synonymy. Schmalhausen's
description isstrongly reminiscent of Xenoxylon. In 1879
Schmalhausen gave amore complete description of the features of
extant Sciadopityswood (and hence those features he considered to
be diagnostic forhis Sciadopityoxylon): The Sciadopitys-wood is
very similar to that ofcommon pine, but it does not have the resin
canals of the latter, and inthe upper and lower rows of ray-cells
the irregular tooth-like thickeningsof pinewood are lacking. By
Sciadopitys the tracheids have pits on theirradial walls, which are
uniseriate, rarely biseriate in very lax woods,sometimes so densely
distributed that they are contiguous, sometimesmore or less
irregularly distant from one another. The ray-cells haveon their
lateral side, like in pinewood, large pores which are
oval-transversal in spring wood, almost oblique and slit-like in
the autumnwood, and single on the width of the tracheid. In
tangential view therays are always uniseriate, (Schmalhausen, 1879:
40; our transla-tion). A Jurassic wood tting this description is
likely to belong toXenoxylon. Moreover this genus was later
documented in the Jurassicof Mangyshlak (Philippe and Thvenard,
1996), and Schmalhausen(1879) reported other samples with similar
anatomy from the Juras-sic of Murayevnya (Ryazan oblast', Russia).
Unfortunately, Gbel'swood specimen has not been illustrated and is
apparently lost. At theBIN the only specimenwe located
fromMangyshlak, a typicalXenoxylonlatiporosum, was collected in
1956 by R.I. Vyalova. It is therefore impos-sible, at present, to
consider priority of Sciadopityoxylon over Xenoxylon.A neotype
should, however, be designated for Sciadopityoxylon.
The recently described Protosciadopityoxylon Zhang, Zheng et
Ding(Zhang et al., 1999; Zheng et al., 2008) is similar
toXenoxylon. It displaysxenoxylean radial pitting and fenestriform
oopores, but the latter arenarrowly bordered or even sub-taxodioid
(sic). Several authors haveassigned samples to Xenoxylon that
sometimes exhibit bordered ooporesin the cross-elds, for example
Gothan and his material from Knig KarlLand (Fig. 7), and Dorofeyuk
and Filin (1969) and their Siberianmaterial.In
Protosciadopityoxylon even early-wood oopores in
non-compressionwood are bordered. This justies the separation of
Protosciadopityoxylonfrom Xenoxylon (Zhang Wu & Zheng Shaolin,
in litt., 2010).
Range: Xenoxylon has been recorded from the Carnian
(LateTriassic; Fliche, 1910; Vogellehner, 1965; Yamazaki and
Tsunada,1981, 1982) to the Maastrichtian (Late Cretaceous; Spicer
andParrish, 1990; Terada et al., 2011). Palaeobiogeographically it
hasbeen found through all Eurasia (including southern plates such
asGeorgia, Northern Iran, South China and Indosinia, but has not
yetbeen recorded from Sibumasu, India, Arabian Peninsula),
Greenland,Alaska and Canada (Alberta).
4.2. Species
Xenoxylon conchylianum FlicheXenoxylon conchylianum Fliche,
Flore fossile du Trias en Lorraine etFranche-Comt: 234, pl. 23 gs.
45. 1910.
Note: In his protologue Fliche emphasises the presence of
radiallyelongated cross-elds and low rays. Age is also used by
Fliche as an
argument to distinguish this species, since Xenoxylon had then
onlybeen recorded from the Jurassic. The original illustration is
crude andof little diagnostic value, the original description is
limited, and type re-vision has long been recommended (Krusel,
1919). Vogellehner statedthat this species is in nahe Verwandschaft
(closely related) to hisXenoxylon parvipunctatum (Vogellehner,
1965: 46), but although bothtaxa have some rounded pits
intermingled among usual xenoxyleanpits, in Xenoxylon conchylianum
all pits are said to be contiguous.Type:We searched for the
specimen described by Fliche inNancy (whereFliche spent most of his
professional life, and where part of hispalaeobotanical collection
is now housed) and in other FrenchMuseumswhere the Fliche's fossil
wood collection could have been housed. Thissearch was
unsuccessful. To the best of our knowledge this collectionhas never
been reviewed. The protologue is so restricted in termsof
char-acteristic features that it is not possible to design a
neotype.
Distribution: 1) type Carnian (not Muschelkalk) of Vaubexy,
Vosges,France; 2) FAD/LAD Schilfsandstein, Early Carnian, Lorraine,
France;this taxon was also reported from the Middle Jurassic of
Hebei,China (Wang, 1995) but with a poor illustration; in his
unpublishedthesis, Khudaiberdyev mentioned (Table 1, p. 19) the
occurrenceof this taxon in the Jurassic of Middle Asia but later
omitted thisdata (Shilkina and Khudaiberdyev, 1971; Khudaiberdyev,
1993);unpublished material from Baikal area at the BIN is labelled
Xenoxyloncf. conchylianum, but the preservation is poor; 3)
geographical rangeWestern Europe.
Xenoxylon fuxinense Ding in Ding, Zheng et Zhang
Xenoxylon fuxinense Ding in Ding, Zheng et Zhang,
ActaPalaeontologica sinica, 39: 245, pl. 1 gs. 16. 2000.
1933 Xenoxylon latiporosum (Cramer) Gothan in Gothan et Sze1980
[Xenoxylon kurumaense] Tsunada, unpublished MSc Thesis,Waseda
University1981 Xenoxylon cf. suljuctense Shilkina et Khudaiberdyev
in Yamazakiet Tsunada
Note: This species appears similar to Xenoxylon suljuctense,
differingonly in details of the tracheid pitting pattern
(occurrence of some
Fig. 7. Xenoxylon phyllocladoides Gothan, slide S1827/03 at the
SMNH; material seen byGothan, with locally bordered oopores
(arrow).
133M. Philippe et al. / Review of Palaeobotany and Palynology
193 (2013) 128140
-
Author's personal copy
opposite to sub-opposite pit pairs); sample S4846 seen at the
SMNH,from Fengtien, Hsi-Hsien and determined by Gothan and Sze
(1933)as Xenoxylon latiporosum, has inated tips bearing both
alternate andopposite biseriate pits in untraumatised wood.
Type: Not seen by us but studied by Ding et al. (2000), sample
Fx-3,stored at the SIGMR.
Distribution: 1) type Haizhou Coal Mine, Fuxin, Liaoning, Fuxin
Fm.,AptianAlbian, Early Cretaceous; 2) FAD Liassic, Negoya Fm.,
KurumuaGroup, Toyama Prefecture, Japan, LAD-type; and 3)
geographical rangeJapan and Liaoning, north-eastern China.
Xenoxylon hopeiense Chang
Xenoxylon hopeiense Chang, Bull. Geol. Soc. China, 7: 250, pl. 1
gs.14, text gs. 17. 1929.
1936 Phyllocladoxylon heizoense Shimakura1967 Xenoxylon barberi
(Seward) Krusel in Shilkina (p.p., sample674 at the BIN)2011
Dacrydioxylon sp. in Selmeier et Grosser
Note 1: According to Chang (1929) similar to
Xenoxylonphyllocladoides, but featuring Rims of Sanio; assigned
toX. phyllocladoidesby Gothan and Sze (1933); good illustration
provided by Ding et al.(2000, pl. 4 gs. 15); X. phyllocladoides
topotypes from Poland neverhave Rims of Sanio and thus it is worth
separating this species.
Note 2: The material described as Xenoxylon phyllocladoides
byJiang et al. (2008) from the Lanqi Formation, Liaoning, might
belongto Xenoxylon hopeiense as it displays Rims of Sanio.
Note 3: The wood from North Korea (Botandai of Heizyo)described
as Phyllocladoxylon heizoense Shimakura (Shimakura, 1936)is also
similar to Xenoxylon hopeiense (syntypes seen at the TUMS). Itis
illustrated in Kim et al. (2005, pl. 19).
Note 4: The wood described as Dacrydioxylon sp. by Selmeier
andGrosser (2011) belongs to Xenoxylon hopeiense; it has axial
parenchy-ma and Rims of Sanio (Selmeier and Grosser, 2011, pl. 6 g.
2).Type: Not seen; Professor C.Y. Chang (Beijing University)
described thetype specimen, recorded as block n 188 from the
Geological Survey;this large block, over 3 tons in weight, had been
on exhibition (Hsiao,1935) but was since lost, probably during the
Sino-Japanese war;recent researches in this town failed to locate
this type specimen(Jiang Zikun, in litt.). Among the syntypes of
Phyllocladoxylonheizoense Shimakura, sample 6874 at the TUMS is
designated hereas the lectotype for that name, and as the neotype
of Xenoxylonhopeiense Chang.
Distribution: 1) type Late Jurassic, Hopei, North-Eastern China
(Chang,1929); 2) FAD Beipiao Fm., Early Jurassic, Liaoning (Wang et
al.,2006), LAD Aptian, Shahai Fm., Yixian, Western Liaoning
(Zhangand Shang, 1996), and Lipovtsy Fm., Southern Primorye,
Russian FarEast (Afonin, 2008); and 3) geographical range
North-eastern China(Beijing, Heilongjiang, Liaoning, Xinjiang),
Eastern Siberia and Arctic(Alexandra I., Franz-Josef Land, Canadian
Arctic Archipelago).
Xenoxylon huolinhense Ding in Ding, Zheng et Zhang
Xenoxylon huolinhense Ding in Ding, Zheng et Zhang,
ActaPalaeontologica sinica, 39: 246, pl. 2 gs. 16. 2000.
Note 1: From the original illustration (pl. 2 g. 4) it seems
that theclustering of radial pits is an artefact due to wavy wood
grain (an un-dulating tracheid wall renders some pits out of
focus); the smallersize and different shapes of the pits at the
tips of the pit row showsit is no artefact; it is observed that pit
clusters usually occur on the
widest tracheid portions, and that pit rows are often
interruptedwhere tracheids are narrow.
Note 2: This wood is similar to Xenoxylon nariwaense, except
forthe occurrence in the latter of some opposite biseriate pit
pairs,which might be a local feature only.
Type: Not seen by us but studied by Ding et al. (2000), sample
H14,stored at the SIGMR.
Distribution: 1) type Huolinhe Coal Mine, Inner Mongolia
AutonomousRegion, Early Cretaceous, Huolinhe Fm.; 2) FAD/LAD this
taxon isknown only from its type specimen; and 3) geographical
range InnerMongolia Autonomous region, China.
Xenoxylon huttonianum (Witham) Philippe et Hayes
Peuce huttonianum Witham of Lartington, Internal Structure of
FossilVegetables, 70, pl. 14 g. 9, pl. 15 gs. 4 et 5 (huttoniana et
hutonii).1833.
1833 Peuce huttoniana (p. 70)"huttonii" (p. 63) Witham1848
Elatoxylon huttoniana Hartig1849 Peuce huttonii Brongniart1849
Pinites huttonianus Gppert in Bronn1870 Cedroxylon huttonianum
Kraus in Schimper1904 Cupressinoxylon barberi Seward1949 Xenoxylon
barberi (Seward) Krusel1968 Xenoxylon ellipticum Schultze-Motel ex
Vogellehner2010 Xenoxylon huttonianum (Witham) Philippe et
Hayes
Note: Wang's datum for Xenoxylon ellipticum from theHongweikeng
Formation, Carnian, Northern Guangdong (Wang,1991) exhibits some
biseriate radial pitting and therefore belongsto the
meisteri-group.
Type: Seen; slide 51498 at the MNH London, Nicol's collection,
chosen asthe neotype for Peuce huttonianum (Philippe and Hayes,
2010); in Berlin,at the HMB, a fossil wood labelled Araucarioxylon
wrttembergicus be-longs to Xenoxylon huttonianum, but this is
probably not the type ofPinites wuerttembergicus Goeppert
(indicated as kept in Mnchen bySchuster, 1909).
Distribution: 1) type Whitby, Yorkshire, UK; 2) FAD
EarlyPliensbachian (Davoei Zone) of Lincolnshire, UK (unpublished
data,sample V25987 at BMNH), LAD Humphresianum zone,
Bajocian,Osnabruck, Germany (unpublished data, leg. F. Wittler);
and 3) geo-graphical range endemic in Western Europe.
Xenoxylon jakutiense Shilkina
Xenoxylon jakutiense Shilkina in Takthajan, A.L. (Ed.),
Problemypaleobotaniki, 154, pl. 4 gs. 14.1986.
Note: This name was quoted as jakutense in Shilkina
andYatsenko-Khmelevsky (1980: 115) but was only validly pub-lished
in 1986; Ding et al. (2000) assigned this wood to
theirliaoningense-group (here meisteri-group) on the basis of
someround tracheid radial pits appearing somewhat spaced out along
thetracheid; however, this arrangement is the result of crassulae,
andXenoxylon jakutiense is therefore more similar to Xenoxylon
hopeiense.Its radial pitting is strictly uniseriate, but Shilkina's
protologue (1986)emphasises that the specimen tracheids are
narrow.
Type: Seen, Coll. 691, spec. 1/335 in wood slides collection of
BIN,three slides.
134 M. Philippe et al. / Review of Palaeobotany and Palynology
193 (2013) 128140
-
Author's personal copy
Distribution: 1) type AptianAlbian of Olenek River Basin,
Siberia;2) FAD/LAD other materials have not yet been reported to
ourknowledge; and 3) geographical range Eastern Siberia.
Xenoxylon japonicum Vogellehner ex Suzuki et Terada
[Xenoxylon japonicum] Vogellehner, Palaeontographica, Abteilung
B,124: 145. 1968.Xenoxylon japonicum Vogellehner ex Suzuki et
Terada, J. Phytogeogra-phy Taxonomy 40: 96. 1992.
1934 Xenoxylon latiporosum (Cramer) Gothan in Shimakura,
deSuzuki et Terada, 1992: 961936 Xenoxylon latiporosum (Cramer)
Gothan in Shimakura pro parte
Note 1: Distinguished from Xenoxylon latiporosum by Vogellehner
onthe basis of frequent tangential pitting and high rays (in this
order);Vogellehner did not typify his new taxon (his name is thus
not validlypublished according to ICBN, art. 37.1), but referred to
all the specimensdescribed collectively by Shimakura (1936) as X.
latiporosum. Thesesyntypes were reviewed by Suzuki and Terada
(1992: 96) who selecteda lectotype (sample n 6870 in Shimakura,
1936) on the basis of preserva-tion and of ray height. This
lectotype does not display tangential pittingand exhibits the
typicalmeisteri-type of radial pitting. Suzuki and Terada(1992)
noted that some of the Shimakura's samples have much lowerrays. For
one of them e.g., n 57601 from Northeast China, housed at theTUMS,
we observed: ray height 211 cells, mean 4.22, standard
deviation1.9; 27 counted. Using the above key n 57601 keys out here
as Xenoxylonmeisteri, Xenoxylon japonicum and X. meisteri often
co-occur in the samelocalities, and thus might originate from the
same plant species.
Note 2: In his protologue Vogellehner (1968) also questioned
theattribution of some samples to Xenoxylon japonicum; the material
de-scribed in 1950 by Boureau (probably now in Paris, not yet
located)from the Liassic? of Vietnam similarly has high rays (up to
36 cells)and some round pits, Boureau's material is probably
referable toX. japonicum along with the late Early Cretaceous
(AptianAlbian?)specimen from Alaska described by Arnold (1953)(a
review of theslides should be undertaken).
Type: Lectotype seen, slide no. 6870 deposited at the TUMS.
Distribution: 1) type Late Triassic, the quarry of
Botandai,Pyongyang, North Korea; 2) FAD/LAD to our knowledge
othermaterialshave not yet been reported; and 3) geographical range
Eastern Siberia.
Xenoxylon latiporosum (Cramer) Gothan
Xenoxylon latiporosum (Cramer) Gothan in Abhandl. Preuss.
Geol.Landesanst. 44: 108. 1905.Pinites latiporosus Cramer in O. von
Heer (Ed.), Flora Fossilis Arctica,176, pl. 40 gs. 18. 868.
1868 Pinites pauciporosus Cramer in O. von Heer, p. 1761870
Araucarioxylon latiporosum (Cramer) Kraus in Schimper, p. 3841887
Araucarioxylon koreanum Felix with doubt, vide Philippe, 19951911
Araucarites latiporosus (Cramer) Tuzson p. 31
Type: Seen, sample S1764 and the three associated slides
S1765,S1766, and S1767 at the SMNH are the holotypes (Philippe and
Cantrill,2007).
Distribution: 1) Holotype Brentskardhaugen Bed,
ToarcianBajocian(Reolid et al., 2010), Green Harbour, Spitzberg; 2)
FAD UpperKokuibelsu Subsuite, Norian, Central Pamir, Tajikistan
(Khudaiberdyev,1993), but there are several other data only dated
as Late Triassic,
LAD Yuliangzi Fm., EarlyMid Maastrichtian, Heilongjiang (Sun et
al.,2007); and 3) geographical range probably covering the
northernmostboreal hemisphere, in Asia, Europe and northernNorth
America (south toAlberta), but several datamust be reappraised. In
particular a reviewof allthe Japanese material found them to t
within Xenoxylon meisteri.
[Xenoxylon liaoningense Duan et Wang in Li et Cui]
[Xenoxylon liaoningense Duan etWang in Duan, Cui etWang], Proc.
Inter.Symp. Tree anatomy & Wood Formation, Tianjin, China (Wu
Shuminged.), International Academic Publishers, Beijing, p. 168,
gs. 79 p. 169(unnumbered plate), gs. 14 p. 170 (unnumbered plate).
1995.[Xenoxylon liaoningense Duan et Wang in Li et Cui], Atlas of
fossil plantanatomy in China, Science Press, Beijing, p. 114115,
plates only, notext. 1995.
Note 1: This name is not validly published as no type is
selectedwhereas two specimens are mentioned; as there is no
indication ofa repository; and as the different diagnoses in this
paper are not clear-ly related to taxa (ICBN, arts. 32.1 d, 37.1,
37.7; McNeill et al., 2012).This name was published a second time
during the same year (in Liand Cui, 1995), but with no protologue.
It was further described andillustrated (copy of original
illustration) in Zheng et al. (2008) butagain without diagnosis and
type designation. As there are some dis-crepancies in the
protologue and as the type details are lacking it wasjudged
importunate to validate the name here.
Note 2: Anatomically similar to Xenoxylon nariwaense, which
haspriority and differs only by ray height; but since we have not
yetseen any of the types and as ages are different we are reluctant
to es-tablish this synonymy.
Type: Not seen, stored in Beijing, at Institute of Botany,
Chinese Acad-emy of Sciences (Zhang Wu & Zheng Shaolin pers.
com.).
Distribution: 1) type Shahai Fm., Aptian, Yixian, Western
Liaoning;2) FAD/LAD to our knowledge no othermaterials have been
reported;and 3) geographical range North-eastern China.
Xenoxylon meisteri Palibin et Jarmolenko
Xenoxylon meisteri Palibin et Jarmolenko, Trudy VGRO, 51:
177,text-g. 1 (Meisterii). 1932.
1936 Xenoxylon latiporosum (Cramer) Gothan in Shimakura, pro
parte1981 Xenoxylon pseudoellipticum Yamazaki et Tsunada
Note 1: The holotype for Xenoxylon pseudoellipticum,
supposedlykept at Waseda University, Japan, is in fact lost (Profs.
Mitsuo Suzuki& Harufumi Nishida pers. com.); it is, however,
well illustrated anddescribed in its protologue; Yamazaki &
Tsunada assigned it to thephyllocladoides-group but early wood
radial pits are neither roundnor spaced.
Note 2: The diagnosis of Xenoxylon shimakurai includes
pithfeatures (Yamazaki et al., 1984); as long as only secondary
xylemis considered, it ts within Xenoxylon meisteri;
unfortunatelyX. shimakurai type is lost (Profs. Mitsuo Suzuki &
Harufumi Nishidapers. com.).
Note 3: The taxonomic signicance of tangential radial pitting
withinthe genus Xenoxylon was questioned (Shilkina and
Khudaiberdyev,1971: 131); within material from the same provenance
heavily tangen-tially pitted wood and tangentially unpitted wood
generally co-occur(Shimakura, 1936; our observations). Dense
tangential pitting inXenoxylon latiporosum-like wood is, however,
encountered only in woodfrom the Cretaceous of Asia (including the
Arctic) making justiable thedistinction of a morphospecies. The
correct name for this taxon isXenoxylon meisteri, a name minimally
illustrated in its protologue butthe holotype of which is available
at the BIN. The original diagnosis for
135M. Philippe et al. / Review of Palaeobotany and Palynology
193 (2013) 128140
-
Author's personal copy
X. meisteri reads: Wood rings distinct, about 5 mmwide, late
wood thinwith thick-walled cells, early wood cells (up to 5 times)
larger,subparenchymatous; resin canals lacking; medullary ray
uniseriate,composed of 311 superposed cells, with a single (very
rarely two)large simple lateral pore; prosenchymatous cells
thin-walled, atgrowth-ring limit somewhat thickened, divided by
sub-tyloses (sic),pits only radial, uniseriate or very rarely
sub-biseriate, elliptical-compressed in dense rows, 1518 m high and
3033 m wide (ourtranslation, prosenchymatous cellsmeans here
tracheids).
Note 4: Specimens showing the characteristics of
Xenoxylonmeisteri have usually been assigned to Xenoxylon
latiporosum; a com-plete review is necessary, and all literature
data for X. latiporosummust be reconsidered, especially those from
Asia; most, if not all,specimens from the Tetori series (Cretaceous
of Japan) belong toX. meisteri, as is, for example, the wood
illustrated by Kim et al.(2005), pl. 6.
Note 5: In traumatic wood biseriate opposite pairs and axial
paren-chyma can be observed (sample 44490 in Suzuki and Terada,
1992,seen at the TUMS).
Type: Seen, sample 507/N224 at the BIN, three slides; other
materialslabelled as Xenoxylon meisteri at the BIN include no.
508/no. 224 (5slides) and wood no. 6 (3 slides).
Distribution: 1) type Vitichsk Nagorie, Vitim Plateau, east of
BaikalLake, Eastern Siberia, Cretaceous (Palibin and Jarmolenko,
1932);2) FAD Beipiao Fm., Early Jurassic of Liaoning (Ding et al.,
2000)and Kuruma Group (cf. Nagoya Fm.), Toyama Prefecture,
Japan(Yamazaki et al., 1984), older material might belong here
(Xenoxyloncf. japonicum, Late Triassic, southwestern Japan,
Yamazaki et al.,1980), LAD Late Cretaceous of Dauria, Transbaikalia
(Dorofeyukand Filin, 1969); and 3) geographical range Japan,
north-easternChina, Eastern Siberia, but several Asian specimens
assigned toXenoxylon latiporosum probably belong here.
Xenoxylon nariwaense Yamazaki, Tsunada et Koike
Xenoxylon nariwaense Yamazaki, Tsunada et Koike, Memoirs of
theSchool for Science and Engineering of Waseda University, 44: 93,
pl.1 g. 1, pl. 2 gs. 12, pl. 3 gs. 16, text-g. 2. 1980.
Note1: Xylologically similar to [Xenoxylon liaoningenseDuan
etWang],which is probably an earlier taxonomical synonym;
unfortunately theconfused and poorly illustrated protologue for [X.
liaoningense] makes itdifcult to judge whether it is a synonym and
material reappraisal isnecessary.
Note 2: Shimakura and Fujiyama (1962) studied Xenoxylon woodfrom
the same provenance as the type of Xenoxylon nariwaense;they
assigned it to Xenoxylon latiporosum.Type: Not seen, no. 79040303-a
(holotype) and 79040303-b(paratype), at Waseda University, Japan;
both lost according to Prof.Mitsuo Suzuki (pers.
com.).Distribution: 1) types Hinabata Fm, Nariwa Group, Norian,
LateTriassic, Hinabata, Okayama Prefecture, Japan (Yamazaki et
al.,1980); 2) FAD/LAD other materials have not yet been
reported;and 3) geographical range Japan.
Xenoxylon parvipunctatum Vogellehner
Xenoxylon parvipunctatum Vogellehner, Erlanger
geologischenAbhandlungen 59: 43, pl. 9 g. 81, pl. 10 gs. 8691, pl.
11 g. 92. 1965.
Note: Where a cross-eld is crossed by a ray parenchyma
trans-verse wall in some cases it has two oopores (Vogellehner, in
a letterquoted by Selmeier, 1968); in establishing this taxon,
Vogellehnerpointed out mostly the small size of radial pits;
however, it is also
worth noting that radial pits are not particularly attened,
sometimesbiseriate alternate, sometimes spaced (protologue, pl. 10
g. 88); theradial pitting conforms to that of the meisteri-group,
whereas thetangential pitting is absent.Type: Not seen, ISBT,
Mgdefrau collection, SMF 31, SMS 31ac, SMSV31ac.Distribution: 1)
type Schilfsandstein, Early Carnian, Simmerhausen,southern
Thuringia, Germany (Vogellehner, 1965); 2) FAD/LAD other materials
have not yet been reported to our knowledge; and 3)geographical
range Western Europe.
Xenoxylon peideensis Zheng et Zhang
Xenoxylon peideensis Zheng et Zhang, Bulletin Shenyang Institute
ofGeology and Mineral resources 5: 332, pl. 31 gs. 112.
1982(peidense).
Note 1: This species was described in Chinese (Zheng and
Zhang,1982) without clear indication of a type specimen and no
diagnosisyet at this time, this was in accordance with the rules of
the ICBN(art. 32.1); subsequently, it was mentioned by He Dechang
(1995)as X. peideense but this is probably more a cacography than
anorthographic variant as the correct form is X. peideensis (ICBN,
rec.60D1); authors put emphasis on a mesh-like structure which
mightbe due to alteration.
Note 2: Radial pit rows covering only half of the width of the
tracheidwall are often observed inwoods of both the
latiporosum-group and themeisteri-group. By Xenoxylon peideensis,
however, such half-covering issupposed to be the dominant
gure.Type: Not seen by us but studied by Ding et al. (2000), HP39
(Zheng andZhang, 1982: 349), stored at the SIGMR, Shenyang, China
(ZhangWu &Zheng Shaolin, in litt.).Distribution: 1) typeMishan,
Eastern Heilongjiang, Dongshengcun Fm,Middle Jurassic; 2) FAD
Beipiao Fm., Early Jurassic, Kazuo, Liaoning(Ding et al., 2000),
LAD Huolinhe Fm, Late Jurassic/Early Cretaceous,Huolinhe mine,
Jarud, Inner Mongolia (He Dechang, 1995); and 3) geo-graphical
range from our database Xenoxylon peideensis is endemicin
North-eastern China (Liaoning, Heilongjiang).
Xenoxylon phyllocladoides Gothan
Xenoxylon phyllocladoides Gothan, Verh. Russ.-Kais. Mineral.
Gesell.44: 454, g. 4. 1906.
1882 Araucarioxylon latiporosum (Cramer) Kraus, in Conwentz1940
Mesembrioxylon sp. in Sitholey1955 Mesembrioxylon sp. in Jacob et
ShuklaXenoxylon barberi auctorem plurime, sed vide Xenoxylon
huttonianumibidem1988 Xenoxylon jurassicum Gothan in Matyjasik et
Gierlinski sednon Kraeusel
Note 1: For a long while this taxon was referred to as
Xenoxylonbarberi (Seward) Krusel, a name based on Cupressinoxylon
barberiSeward, the syntypes of which are all from the Toarcian of
Whitby(Yorkshire, UK), and are stored at the BMNH (as 51495, 51496
and51498); these syntypes were assigned to Xenoxylon
huttonianum(Philippe and Hayes, 2010), and thus the name X. barberi
is not rele-vant to Xenoxylon phyllocladoides synonymy.
Note 2: Matyjasik and Gierliski (1988) use the name
XenoxylonjurassicumGothan, but Gothan never erected such name;
unfortunate-ly the only published Xenoxylon jurassicum is that of
Krusel (1949)based on Protopodocarpoxylon jurassicum Eckhold,
itself based onPodocarpoxylon sp. Gothan, which was used for a wood
from theXenoxylon phyllocladoides type locality; investigations by
us of thistype locality yielded numerous topotypes of
Protopodocarpoxylonjurassicum, which are clearly not Xenoxyla
(Philippe et al., 2006).
136 M. Philippe et al. / Review of Palaeobotany and Palynology
193 (2013) 128140
-
Author's personal copy
Type: Not seen, said to be deposited in Warsaw, at Polish
GeologicalInstitute; this and other polish collections with
palaeontologicalsections were contacted thanks to the help of
Michal Zaton andLeszek Marynowski (Sosnowiec University), but
without success.We have, however, examined topotypes (Philippe et
al., 2006).
Among them we propose sample MP1571 at the LPUL as the
neotypeand illustrate it here (Fig. 8) since Gothan's protologue is
minimallyillustrated.Description of the neotype: tracheidoxyl; no
axial parenchyma or resincanals; growth-rings marked, early- to
late-wood transition abrupt,late-wood limited to 25 layers of
cells; transverse section of tracheidssquare to hexagonal; rays
average 7 cells high (mean 7.6, standard devia-tion 5.3, n=38);
tangential pitting limited to rare round isolated andsmall
areolated pits; ray cell walls thin and smooth, pitted only
incross-elds; transverse ray cell walls perpendicular, ray cells 24
tra-cheids wide; cross-elds with a single unbordered oopore
occupyingthe whole eld, with width three times that of the height
in the widesttracheids, square to rectangular (width 0.5 times that
of the height) inthe late-wood; two oopores occur in about 2% of
the cross-elds, eitherin high marginal cells or in widest
cross-elds when they are divided bythewall of a transverse ray
cell; when tracheids alternate from one radialrow to the other, the
oopore sometimes does not occupy the whole eld;
Fig. 8. Xenoxylon phyllocladoides Gothan neotype (sample MP1571
at the LPUL, Bathonian, Anna brick pit, Gnaszyn, south-central
Poland) in radial section; A three alternatingtracheids on the
left, radial pitting variable; B wide early wood tracheids with
biseriate (on the right) and uniseriate radial pitting; C
cross-elds (on tracheid casts) with largefenestriform oopores,
variable in size and usually larger in marginal cells; D cross-elds
(on ray cell casts), oopores not always occupying the whole eld; E
late woodcross-elds, relatively round, slightly bordered (a local
gure); F showing the rare condition of two-pits per cross-elds in a
ray marginal cell.
Table 3Radial pitting exhibited by the neotype of Xenoxylon
phyllocladoides Gothan. Wood isconsidered normal when the tracheids
of two adjacent radial rows are opposite, and al-ternate when
otherwise. Pitting in the tips of the tracheid is also
differentiated. Per-centages are based on counts of 120 pits.
Radial pitting Normal wood Alternate wood Tracheid tips
Uniseriate distant 78 87 0Uniseriate contiguous round 14 13
8Uniseriate xenoxylean 3 0 1Biseriate opposite 4 0 86Biseriate
alternate 1 0 5
137M. Philippe et al. / Review of Palaeobotany and Palynology
193 (2013) 128140
-
Author's personal copy
ray marginal cells usually not differentiated, locally up to
twice as high asinner ray cells; radial pitting variable, from
uniseriate spaced to biseriatealternate (Table 3); tracheid tips
obtuse to shortly acuminate, denselypitted.Distribution: 1)
typeMiddle Jurassic (Bajocian or Bathonian) of Poland(Gothan,
1906); 2)FAD Upper Kokuibelsu Subsuite, Norian, CentralPamir,
Tajikistan (Khudaiberdyev, 1993)/LAD Kedrova Fm., Albian,Talovka
River Basin, northwestern Kamchatka (Afonin, 2010); and
3)geographical range probably covering the whole of Europe and
Asiaat mid to high palaeolatitudes, but material reappraisal is
needed.
Xenoxylon suljuctense Shilkina et Chudajberdiev
Xenoxylon suljuctense Shilkina et Chudajberdiev,
PalaeontologiaUzbekistana, 2: 132, pl. 80 gs. 14. 1971.
Note: Original diagnosis was written in Russian in 1971, and
thusts ICBN requirements for validity (art. 32.1).Original
diagnosis (our translation): Annual rings are visible. Radial
pitsuniseriate or biseriate. Uniseriate pits large, connected,
strongly at-tened in vertical direction, ovoid in their shape, with
elongated innerapertures. Biseriate pits alternate, covering almost
all the length of thetracheid wall, closely connected, of irregular
pentagonal shape. Innerpit aperture round. Rays are low, from 1 to
10 rows of cells high, com-monly 56 cells. Cross eld pits are
ovoid, elongated in horizontal direc-tion, one pit per cross eld.
Tangential walls of tracheids bear numeroussmall round uniseriate
pits, sometimes biseriate pits with oblique inneraperture. Wood
parenchyma absent.Type: Not seen, indicated as sample 1, stored at
the Botanical Insti-tute of Uzbekistan, Tashkent.Distribution: 1)
type Middle Jurassic of Sulyukta, southernFerghana, Kyrgyzstan; 2)
FAD Late Triassic, Heizyo Middle School,Pyongyang, North-Korea
(sample 62067 at the TUMS, unpublished)/LAD type; and 3)
geographical range Central Asia and Korea.
Xenoxylon wattarianum Nishida et Nishida
Xenoxylon wattarianum Nishida et Nishida, 1986, gs. 56.
1986.
Note: There are some discrepancies in the protologue; the
descrip-tion states that rays are always uniseriate, whilst a
partly biseriate rayis gured (Nishida and Nishida, 1986, g. 5B); on
the same tangentialview a cluster of 12much compressed large pits
is gured, completelydifferent from the other tangential pits
(probably resulting from therotation of some radial pits); dense
tangential pitting was not observedwhilst reviewing the holotype;
this wood is similar to Xenoxylonlatiporosum, but differs in ray
height (Nishida and Nishida, 1986: 200).Type: Seen, sample 11109 at
Chuo University, Tokyo, Japan.Distribution: 1) type
Turonian/Santonian, Nayba River, SouthernSakhaline; 2) FAD/LAD this
taxon is represented only by the typespecimen; and 3) geographical
range Eastern Siberia.
Xenoxylon yixianense Zhang et Shang
Xenoxylon yixianense Zhang et Shang, The Palaeobotanist 45: 46,
pl.12. 1996.
Note: This wood is unique among the Xenoxyla as it features
radialresin canals.
Type: Not seen but studied by Ding et al. (2000); SZ001
deposited atFuxin Mining College, Fuxin, Liaoning, China.
Distribution:1) type Baitazigou, Yixian, Liaoning, Shahai Fm,
EarlyCretaceous (Zhang and Shang, 1996); 2) FAD/LAD this taxonis
represented only by the type specimen; and 3) geographicalrange
north-eastern China.
5. Discussion of Xenoxylon systematic position
On the basis of xylotomy it is impossible to assign Xenoxylon to
aprecise group within the Lignophyta. Indeed, as reected by
Gothan'schoice of the root Xeno (foreign or alien in Greek), this
wood hasno equivalent among modern plants. It has never been found
in con-nection with leafy or reproductive structures.
Xenoxylonwasrst thought to be thewoodof amodern conifer or
per-hapsPodozamitesBraun (Nathorst, 1897), and subsequently
hypothesisedto be thewood of Baiera Braun (Arnold, 1953) or
Sciadopitys (Jarmolenko,1933; Bailey, 1953) or of some extinct
Podocarpaceae (Shilkina andKhudaiberdyev, 1971). In 1957 Larishchev
noticed that Xenoxylon fre-quently co-occurred with
Sciadopitys-like foliages in Siberia. LaterBose and Manum (1990)
demonstrated that these foliages werenot Sciadopitys-related but
members of an extinct conifer family,the Miroviaceae (an extinct
family including the genera TritaeniaMaegdefrau et Rudolf, Mirovia
Reymanwna and SciadopityoidesSveshnikova). Organic geochemistry
ruled out the Pinaceae fromXenoxylon relationships, and evidenced
similarities with extantCupressaceae s.l. and Podocarpaceae
(Marynowski et al., 2008), butbona de Miroviaceae wood has not yet
been studied. Interestinglycross-elds similar to these of Xenoxylon
can be observed in Miroviaeximia (Gordenko) N. Nosova (Natalia
Gordenko, pers. com.) whichmay provide further light on Xenoxylon
relationships.
The stratigraphic range of Miroviaceae also ts well with that
ofXenoxylon, i.e. CarnianMaastrichtian. Interestingly Miroviaceae
occurin the Early Jurassic of Mangyshlak (Nosova and Kiritchkova,
2008)from where Sciadopityoxylon vetustawas described, in the Mid
Jurassicof Polandwhere Xenoxylon also occur (Philippe et al., 2006;
Nosova andWciso-Luraniec, 2007) and in several other localities
where Xenoxylonis documented (such as the Lipovtsy Fm. of Southern
Primorye, RussianFar-East; Afonin, 2008).
More recently Xenoxylon was assigned to the
Protopinaceae(Youssef, 2002). However, this morphotaxon has no
phylogeneticallegitimacy, as it comprises a group of woods with
different afnities.
6. Conclusion
This review based on consulting little known Russian,
Japaneseand Chinese literature as well as fossil material provides
greater clar-ity to Xenoxylon systematics. However as we note above
there is still alot of work that needs to be done: most specimens
need to bereappraised. The stratigraphical and geographical
distributions givenhere for each species is only tentative. It is
likely that hidden in an as-sumed geographically widespread genus
with apparently little anatomi-cal differentiation lies an
interesting biodiversity. In particular Xenoxylonhuttonianum and
Xenoxylonmeisteri are here, for the rst time, evidencedas taxa of
palaeobiogeographical signicance.
Acknowledgements
Prof. Harufumi Nishida is most gratefully acknowledged
forallowing revision of his material and Prof. Mitsuo Suzuki for
lab facil-ities and permission to access Shimakura's material as
well to his ownmaterial. We thank Michal Zaton for the help in
trying to locate theXenoxylon phyllocladoides type in Poland and in
collecting topotypes.Eric Buffetaut, Leszek Marynowski, Platon
Tchoumatchenco, JohannSchnyder, Michal Zaton, Frank Wittler, Zhang
Wu and Zheng Shao-linprovidedmaterial. Jiang Hong-En kindly helped
with Chinese literatureand translated Ding et al.'s key (2000),
adapted here. Jiang Zikun kindlyprovided information on some
Chinese types. David Cantrill and SteveMcloughlin also helped with
accessing and studying the collections inStockholm. Paul Kenrick's
editing is also acknowledged, as well as thatof two anonymous
reviewers.
138 M. Philippe et al. / Review of Palaeobotany and Palynology
193 (2013) 128140
-
Author's personal copy
Appendix A. Supplementary data
Supplementary data to this article can be found online at
http://dx.doi.org/10.1016/j.revpalbo.2013.01.013.
References
Afonin, M., 2008. Pervyie nakhodki iskopayemykh drevesin
Xenoxylon latiporosum(Cramer) Gothan i X. hopeiense Chang na
Dal'nem Vostoke Rossii. Vestnik DVORAN 4, 126132 (in Russian).
Afonin, M., 2010. Xenoxylon fossil woods from the Lower
Cretaceous deposits ofRussian Far-East. Poster at the Sixth
European Palaeobotanical and PalynologicalCongress, Budapest, 610
July 2010.
Amiot, R., Wang, X., Zhou, Z., Wang, X., Buffetaut, E., Lcuyer,
C., Ding, Z., Fluteau, F.,Hibino, T., Kusuhashi, N., Mo, J.,
Suteethorn, V., Wang, Y., Xu, X., Zhang, F., 2011.Oxygen isotopes
of East Asian dinosaurs reveal exceptionally cold Early
Cretaceousclimates. PNAS 108, 51795183.
Andrews, H.N., 1955. Index of generic names of fossil plants,
18201950. U.S. Geologi-cal Survey Bulletin 1013.
Arnold, C.A., 1953. Silicied plant remains from the Mesozoic and
Tertiary of WesternNorth America II. Some fossil woods from Alaska.
Papers of the Michigan Academyof Sciences, Arts and Letters 38,
820.
Bailey, I.W., 1953. Evolution of the tracheary tissue of land
plants. American Journal ofBotany 40, 48.
Bamford, M., Philippe, M., 2001. Gondwanan JurassicEarly
Cretaceous homoxylouswoods: a nomenclatural revision of the genera
with taxonomical notes. Reviewof Palaeobotany and Palynology 113,
287297.
Bose, M.N., Manum, S.B., 1990. Mesozoic conifer leaves with
Sciadopitys-like stomataldistribution. A re-evaluation based on
fossils from Spitsbergen, Greenland andBafn Island. Norsk
Polarinstitut Skrifter 192, 181.
Boureau, E., 1950. Contribution l'tude paloxylologique de
l'Indochine. I: prsencedu Xenoxylon latiporosum (Cramer) Gothan
dans le Lias du Centre-Annam. Bulletindu Service gologique
d'Indochine 29, 116.
Brongniart, A., 1849. Tableau des genres de vgtaux fossiles.
Dictionnaire universeld'Histoire naturelle, Paris, Martinet.
Chang, C.Y., 1929. A new Xenoxylon from North China. Bulletin of
the Geological Societyof China 8, 243251.
Conwentz, H., 1882. Fossile Hlzer aus der Sammlung der
Kniglischen geologischenLandesanstalt zu Berlin. Jahresbericht der
kniglische preuische geologischeLandanstalt und Bergakademie 2,
144171.
Cramer, C., 1868. Fossile Hlzer der arctischen Zone. In: von
Heer, O. (Ed.), Flora fossilisarctica - Die fossile Flora der
Polarlnder. Zrich, pp. 167180.
Dechang, He, 1995. The Coal Forming Plants of Late Mesozoic in
Dan Hinggan Mountains.China Coal industry Publishing House, Beijing
(in Chinese with English abstract).
Ding, Qiu-Hong, Zheng, Shaolin, Zhang, Wu, 2000. Mesozoic fossil
woods of genusXenoxylon from Northeast China and its palaeoecology.
Acta Palaeontologica Sinica39, 237249.
Dorofeyuk, N.I., Filin, V.R., 1969. Xenoxylon latiporosum
(Cramer) Gothan izverkhnemelovykh otlozheniy Zabaikalya. Byulleteny
Moskovskogo obschestvaispytateley prirody - Otdeleniye biologii 74,
7287 (in Russian).
Felix, J., 1887. Untersuchungen ber fossile Hlzer, drittes Stck.
Zeitschrift derDeutschen geologischen Gesellschaft 39, 517528.
Fliche, P., 1910. Flore Fossile du Trias en Lorraine et
Franche-Comt. Berger-Levrault,Paris.
Gomolitzky, N.P., Khudaiberdyev, R.Kh., Yunusov, U.K., 1981.
Materialy k yurskoy oreAngrena. Paleobotanika Uzbekistana 3, 370
(in Russian).
Gppert, H.R., 1849. Vegetabilia. In: Bronn, H.G., Gppert, H.R.,
von Meyer, H. (Eds.),Index Palaeontologicus: Handbuch einer
Geschichte der Natur, 3, pp. 372.
Gothan, W., 1905. Zur Anatomie lebender und fossiler
Gymnospermen-Hlzer.Abhandlungen preuische geologische Landesanstalt
44, 1108.
Gothan, W., 1906. Fossile Hlzer aus dem Bathonian von
Russisch-Polen. VerhandlungenRussische-Kaiserlische Mineralogische
Gesellschaft 44, 435458.
Gothan, W., Sze, H.C., 1933. ber fossile Hlzer aus China.
Memoirs of the National Re-search Institute of Geology 13,
87104.
Hartig, T., 1848. Beitrge zur Geschichte der Panzen und zur
Kenntnis dernorddeutschen Braunkohlen-Flora. Botanische Zeitung 6,
122128 (137141,166172 and 185190).
Hsiao, C.C., 1935. A chemical analysis of the fossil wood
Xenoxylon. Bulletin of the Geo-logical Society of China 14,
7376.
IAWA, 2004. IAWA list of microscopic features for softwood
identication. IAWA Jour-nal 25, 170.
Jacob, K., Shukla, B.N., 1955. Jurassic plants from the Saighan
series of northern Afghanistanand their paleoclimatic and
paleogeographic signicance. Memoirs of the GeologicalSurvey of
India 33, 164.
Jarmolenko, A.V., 1933. The experimental application of stem
secondary wood anatomyto investigation of conifer phylogeny. Soviet
Botany 6, 4663.
Jiang, Hong-En, Ferguson, D.K., Li, Cheng-Sen, Cheng, Ye-Ming,
2008. Fossil coniferouswood from the Middle Jurassic of Liaoning
Province, China. Review ofPalaeobotany and Palynology 150,
3747.
Khudaiberdyev, R., 1993. Fossil woods of the genus Xenoxylon and
their developmentin the Mesozoic of Middle Asia. In: Lucas, S.G.,
Morales, M. (Eds.), The Non-marine Triassic: New-Mexico Museum of
Natural History & Science Bulletin, 3,pp. 22251.
Kim, Kyungsik, Suzuki, M., Oh, Changhwan, 2005. Re-examination
of Prof. Shimakura'sconiferous fossil wood microscope-slides
deposited in Tohoku University Museum.Bulletin of the Tohoku
University Museum 4, 1772.
Kraus, G., 1870. Bois fossiles de conifres. In: Schimper, W.P.
(Ed.), Trait dePalontologie vgtale, tome 2. Baillre, Strasbourg,
pp. 363385.
Krusel, R., 1919. Die fossilen Koniferenhlzer (unter Auschlu von
AraucarioxylonKraus). I: Versuch einer monographischen Darstellung.
Palaeontographica 62,185284.
Krusel, R., 1949. Die fossilen Koniferen-Hlzer (Unter Ausschlu
von AraucarioxylonKraus). II: Kritische Untersuchungen zur
Diagnostik lebender und fossilerKoniferen-Hlzer. Palaeontographica
Abteilung B 89, 83203.
Larishchev, A.A., 1948. Pervaya nakhodka gagata v Kuzbasse.
Uchenyie zapiskiTomskogo universiteta 8, 7387 (in Russian).
Larishchev, A.A., 1957. O nekotorykh iskopaemykh ostatkakh
drevesiny iz ZapadnoySibiri. Doklady VII nauchnoy conferensii,
Tomsk 4, 1315 (in Russian).
Li, Cheng-Sen, Cui, Jin-Zhong (Eds.), 1995. Atlas of Fossil
Plant Anatomy in China. Sci-ence Press, Beijing, China.
Marynowski, L., Philippe, M., Zaton, M., Hautevelle, Y., 2008.
Systematic relationships ofMesozoic wood genus Xenoxylon:
integrative biomolecular and palaeobotanicalapproach. Neues
Jahrbuch fr Geologie und Palontologie Abhandlungen 247,177189.
Matyjasik, A., Gierliski, G., 1988. Drewno jurajskie Xenoxylon
jurassicum z kry ladowcowejw ukowie. Przeglad Geologiczny 38,
106108.
Mcneill, J., Barrie, F.R., Buck, W.R., Demoulin, V., Greuter,
W., Hawksworth, D.L.,Herendeen, P.S., Knapp, S., Marhold, K.,
Prado, J., Prud'homme Van Reine, W.F.,Smith, G.F., Wiersema, J.H.
(Eds.), 2012. International Code of Nomenclature forAlgae, Fungi,
and Plants Melbourne Code Adopted by the Eighteenth
InternationalBotanical Congress Melbourne, Australia, July 2011.
Regnum Vegetabile. A.R.G.Gantner Verlag KG, Rugell, p. 154.
Medlyn, D.A., Tidwell, W.D., 1975. Conifer wood from the Upper
Jurassic of Utah. IXenoxylon morrisonense nov. spec. American
Journal of Botany 62, 203208.
Mller-Stoll, W.R., 1951. Mikroskopie des zersetzen und
fossilierten Holzes. In: Freund,H. (Ed.), Handbuch der Mikroskopie
in der Technik, 5, pp. 727816.
Mller-Stoll, W.R., Schultze-Motel, J., 1988. Gymnospermen Hlzer
des deutschen Jura. I:Xenoxylon und Dadoxylon. Zeitschrift der
deutschen geologischen Gesellschaft 139,6381.
Nathorst, A.G., 1897. Zur mesozoischen Flora Spitzbergens.
Konigliche SvenskaVetenskapsakademiens Handligar 30, 177.
Nishida, M., Nishida, H., 1986. Structure and afnities of the
petried plants from theCretaceous of Northern Japan and Saghalien,
III: petried plants from the UpperCretaceous of Saghalien.
Botanical Magazine 99, 191204.
Nishida, M., Nishida, H., Suzuki, Y., 1993. On some petried
plants from the Cretaceousof Choshi, Chiba prefecture VIII. Journal
of Japanese Botany 68, 289299.
Nosova, N.V., Kiritchkova, A.I., 2008. First records of the
genus Mirovia Reymanwna(Miroviaceae, Coniferales) from the Lower
Jurassic of Western Kazakhstan(Mangyshlak). Paleontological Journal
42, 13831392.
Nosova, N.V., Wciso-Luraniec, E., 2007. A reinterpretation of
Mirovia Reymanwna(Coniferales) based on the reconsideration of the
type species Mirovia szaferiReymanwna from the Polish Jurassic.
Acta Palaeobotanica 47, 359371.
Palibin, I.V., Jarmolenko, A.V., 1932. Novyi vid iskopayemoy
drevesiny s Vitimskogoploskogorya. Izvestiya Vsesoyuznogo
geologo-razvedyvatel'nogo obyedineniya(Trudy VGRO) 51, 177179 (in
Russian).
Philippe, M., 1995. Bois fossiles du Jurassique de Franche-Comt
(nord-est de laFrance): systmatique et biogographie.
Palaeontographica Abteilung B 236,45103.
Philippe, M., Bamford, M., 2008. A key to morphogenera used for
Mesozoic conifer-likewoods. Review of Palaeobotany and Palynology
148, 184207.
Philippe, M., Cantrill, D., 2007. Nomenclatural types and
taxonomy of Gothan's Arcticfossil conifer wood. Taxon 56,
551566.
Philippe, M., Hayes, P., 2010. Reappraisal of two of Witham's
species of fossil wood withtaxonomical and nomenclatural notes on
Planoxylon Stopes, Protocedroxylon Gothanand Xenoxylon Gothan.
Review of Palaeobotany and Palynology 162, 5462.
Philippe, M., Thvenard, F., 1996. Repartition and palaeoecology
of the Mesozoic woodgenus Xenoxylon: palaeoclimatological
implications for the Jurassic of WesternEurope. Review of
Palaeobotany and Palynology 91, 353370.
Philippe, M., Barbacka, M., Gradinaru, E., Iamandei, E.,
Iamandei, S., Kzmr, M., Popa,M., Szakmny, G., Tchoumatchenco, P.,
Zato, M., 2006. Fossil wood and Mid-Eastern Europe terrestrial
palaeobiogeography during the JurassicEarly Cretaceousinterval.
Review of Palaeobotany and Palynology 142, 1532.
Philippe, M., Jiang, H.-E., Kim, K.-S., Oh, C., Gromyko, D.,
Harland, M., Paik, I.-S.,Thvenard, F., 2009. Structure and
diversity of the Mesozoic wood genus Xenoxylonin Far-East Asia:
implications for terrestrial palaeoclimates. Lethaia 42,
393406.
Reolid, M., Philippe, M., Nagy, J., Abad, I., 2010. Exceptional
preservation of phosphaticwood remains of the Brentskardhaugen Bed
(middle Jurassic) from Svalbard(Boreal Realm). Facies 56,
549566.
Schmalhausen, J., 1877. ber Sciadopityoxylon. Arbeiten der
Petersburg Gesellschaft frNaturforschung 8, 114115.
Schmalhausen, J., 1879. Beitrge zur Juraora Russlands. Mmoires
de l'Acadmieimpriale de Saint-Petersbourg 27, 3941.
Schuster, J., 1909. Palobotanische Notizen aus Bayern. I. ber
das Keuper- undLiasholz. Bayerische Botanischen Gesellschaft 22,
120.
Selmeier, A., 1968. Xenoxylon cf. jurassicum (Eckhold)Krusel aus
dem Schilfsandsteinvon Rckensdorf bei Ansbach (Mittel Franken).
Neues Jahrbuch fr Geologie undPalontologie (Abhandlungen) 131,
243251.
Selmeier, A., Grosser, D., 2011. Lower Cretaceous conifer drift
wood from SverdrupBasin, Canadian Arctic Archipelago. Zitteliana
51, 1935.
139M. Philippe et al. / Review of Palaeobotany and Palynology
193 (2013) 128140
-
Author's personal copy
Serra, C., 1966. Etude anatomique et palogographique de quelques
espceshomoxyles du Sud-Vietnam et du Cambodge. Archives Gologiques
du Vietnam8, 59131.
Seward, A.C., 1904. Catalogue of the Mesozoic plants in the
department of geology,British Museum (Natural History). The
Jurassic Flora. II.- Liassic and Oolitic Florasof England
(Excluding the Inferior Oolite Plants of the Yorkshire
Coast).BritishMuseum, London.
Shilkina, I.A., 1967. Iskopayemyie drevesiny Zemli
Frantsa-Iosifa. Paleobotanika 6,2950 (in Russian).
Shilkina, I.A., 1986. Iskopayemyie drevesiny severnoy Yevrazii.
In: Takthajan, A.L. (Ed.),Problemy paleobotaniki. Nauka, Leningrad,
pp. 150155 (in Russian).
Shilkina, I.A., Khudaiberdyev, R.Kh., 1971. Novyie nakhodki i
obzor rodovProtocedroxylon i Xenoxylon. Paleobotanika Uzbekistana
2, 117142 (in Russian).
Shilkina, I.A., Yatsenko-Khmelevsky, A.A., 1980. Nekotoryie
aspekty evolyutsii drevesinykhvoynykh v mezozoye. In: Zhilin, S.G.
(Ed.), Sistematika i evolyutsiya vysshihrasteniy. Nauka, Leningrad,
pp. 110116 (in Russian).
Shimakura, M., 1936. Studies on fossil woods from Japan and
adjacent lands. I: someJurassic woods from Japan and Mandchoukuo.
Science Reports of the Tohoku Im-perial University, series 2, 18,
pp. 267298.
Shimakura, M., Fujiyama, I., 1962. On the Xenoxylon latiporosum
in the Triassic of NariwaGroup. Journal of the Geological Society
of Japan 68, 418419 (in Japanese).
Duan, Shu-Ying, Cui, Jingzhong, Wang, Wangn, Xiong, Bingkun,
Wang, Yongqiang, 1995.Fossil woods from the Early Cretaceous of
Western Liaoning, China. In: Wu, S.M.(Ed.), Wood anatomy Research:
Proceedings of the International Symposium onTree Anatomy and wood
formation, Tianjin, P.R. China, pp. 166171 (1995).
Sitholey, R.V., 1940. Jurassic plants from AfghanTurkistan.
Memoirs of the GeologicalSurvey of India Palaeontologia Indica 4,
157.
Spicer, R.A., Parrish, J.T., 1990. Latest Cretaceous woods of
the central North Slope,Alaska. Palaeontology 33, 225242.
Sun, Ge, Akhmetiev, M., Golovneva, L., Quan, Cheng, Kodrul,
T.M., Nishida, H., Sun, Yue-Wu, Sun, Chun-Lin, Johnson, K.,
Dilcher, D., 2007. Late Cretaceous plants fromJiayin along
Heilongjiang river, northeast China. Courier Forschung
InstitutSenckenberg 258, 7583.
Suzuki, M., Terada, K., 1992. Xenoxylon fossil woods from the
Lower Cretaceous Akaiwasubgroup of Shiramine, Central Japan.
Journal of Phytogeography and Taxonomy40, 9197.
Tchoumatchenco, P., Philippe, M., Yaneva, M., 2008. Did a
glaciation occur during theToarcian (Early Jurassic) in the East
Stara Planina Mts. (East Bulgaria)? Proceedingsof the Bulgarian
Academy of Sciences 61, 13011308.
Terada, K., Nishida, H., Sun, G., 2011. Fossil woods from the
Upper Cretaceous to Paleo-cene of Heilongjang (Amur) River area of
China and Russia. Global Geology 14,192208.
Tidwell, W.D., Britt, B.B., Ash, S.R., 1998. Preliminary oral
analysis of the Mygatt-Moore Quarry in the Jurassic Morrison
Formation, west-central Colorado. ModernGeology 22, 341378.
Tsunada, K., 1980. Studies on the classication of fossil woods.
Master Thesis 1980MS,School of Science and Engineering, Waseda
University, Tokyo, Japan (in Japanese).
Tuzson, J., 1911. Monographie der fossilen Panzenreste der
Balatonseegegend.Resultate der Wissenschaftlichen Erforschung des
Balatonsees, Band 1, Teil 1.Ungarischen Geographischen
Gesellschaft, Wien, pp. 164.
Vogellehner, D., 1965. Untersuchungen zur Anatomie und
Systematik der verkieseltenHlzer aus dem frnkischen und
sdthringischen Keuper. Erlanger GeologischenAbhandlungen 59,
176.
Vogellehner, D., 1967. Zur Anatomie und Phylogenie mesozoischer
Gymnospermenhlzer,Beitrag 7: Prodromus zu einer Monographie der
Protopinaceae. II: Die protopinoidenHlzer der Trias.
Palaeontographica Abteilung B 121, 3051.
Vogellehner, D., 1968. Zur Anatomie und Phylogenie mesozoischer
Gymnospermenhlzer,Beitrag 7: Prodromus zu einer Monographie der
Protopinaceae. II. Die ProtopinoidenHlzer des Jura.
Palaeontographica Abteilung B 124, 125162.
Wang, Shi-Jun, 1991. The occurrence of Xenoxylon ellipticum in
the late Triassic fromNorth Guangdong, China. Acta Botanica Sinica
33, 810812 (in Chinese withEnglish abstract).
Wang, Xin, 1995. Xenoxylon conchylianum Fliche. In: Li, C.S.,
Cui, J.Z. (Eds.), Atlas ofFossil Plant Anatomy in China. Science
Press, Beijing, pp. 105111.
Wang, Yong-Dong, Saiki, K., Zhang, Wu., Zheng, Shaolin, 2006.
Biodiversity andpalaeoclimate of the Middle Jurassic oras from the
Tiaojishan Formation in west-ern Liaoning, China. Progress in
Natural Sciences 16, 222230.
Watari, S., 1960. On some structure and afnity of Xenoxylon
latiporosum. Journal of thefaculty of science, University of Tokyo,
Botany 7, 511521.
Witham of Lartington, T.M., 1833. The Internal Structure of
Fossil Vegetables Foundin the Carboniferous and Oolitic Deposits of
Great Britain. In: A., Black, C. (Eds.),(Edinburgh, UK).
Yamazaki, S., Tsunada, K., 1981. Fossil coniferous woods
belonging to ProtocedroxylonGothan and Xenoxylon Gothan, obtained
from the Upper Triassic Mine Group,Southwest Japan. Bulletin of the
Science and Engineering Research Laboratory ofWaseda University 97,
118.
Yamazaki, S., Tsunada, K., 1982. Paleobotanical study on
fusinites occurring in theLower Jurassic Kuruma Group, Southwest
Japan. Memoirs of the School for Scienceand Engineering of Waseda
University 46, 73123.
Yamazaki, S., Tsunada, K., Koike, N., 1980. Some fossil woods
from the Upper TriassicNariwa Group, south-west Japan. Memoirs of
the School for Science and Engineer-ing of Waseda University 44,
91131.
Yamazaki, S., Tsunada, K., Hagiwara, I., 1984. Comparison
between the Liassic andNeocomian species of Xenoxylon Gothan
occurring in the Hifa Terrane, CentralJapan. Memoirs of the School
of Science and Engineering of Waseda University48, 93115.
Youssef, S.G.M., 2002. Xenoxylon wood from late JurassicEarly
Cretaceous of GebalKmil, Egypt. IAWA Journal 23, 6976.
Zhang, Wu., Shang, Ping, 1996. Xenoxylon yixianense sp. nov.
from Lower Cretaceous ofYixian, Western Liaoning, China. The
Palaeobotanist 45, 389392.
Zhang, Wu, Zheng, Shaolin, Ding, Qiu-Hong, 1999. A new genus
(Protosciadopityoxylongen. nov.) of Early Cretaceous fossil wood
from Liaoning, China. Acta BotanicaSinica 41, 13121316.
Zheng, Shaolin, Zhang, Wu, 1982. Fossil plants from Longzhaogou
and Jixi Groups inEastern Heilongjiang Province. Bulletin of the
Shenyang Institute of Geology andMineral Resources 5, 278349.
Zheng, Shaolin, Li, Yong, Zhang, Wu., Li, Nan, Wang, Yong-Dong,
Yang, Xiao-Ju, Yi,Tie-Mei, Yang, Jia-Ju, Fu, Xiao-Ping, 2008.
Fossil Wood of China. China forestryPublishing House, Beijing,
China.
140 M. Philippe et al. / Review of Palaeobotany and Palynology
193 (2013) 128140









![Pr Philippe Mairiaux Santé au travail et Education pour la santé … · 2013-10-25 · (« lumbago » versus « hernie ou protrusion discale ») [Abenhaimet al Spine1995; 20:791-5]](https://img.pdfslide.fr/doc/110x75/5f6f2f9038548559a315c9ac/pr-philippe-mairiaux-sant-au-travail-et-education-pour-la-sant-2013-10-25.jpg)