Embed Size (px)
Citation preview

LES BRÈVES
PAGE 28
LA RÉPONSE IMMUNITAIRE INNÉE CHEZ DROSOPHILA MELANOGASTER
Par Indoumady Baskara M1 Génétique GCDE, Paris 11
L’émergence de nouvelles maladies infectieuses et la récente résurgence d’anciens pathogènes humains ont souligné l’importance d’une compréhension accrue des mécanismes d’interaction entre les bactéries et leurs hôtes. En effet, certains micro-organismes sont pathogènes et peuvent induire des dysfonctionnements importants chez l’hôte infecté allant parfois jusqu’à provoquer la mort (Beutler, 2004). Les étapes précoces de l’interaction entre l’hôte et le pathogène sont déterminantes pour la suite du processus infectieux. L’immunité adaptative n’est fonctionnelle que quelques jours, voire quelques semaines après l’infection. Entre temps, l’infection a été combattue par l’immunité innée qui, bien que moins spécifique, est néanmoins très efficace (Lemaitre, 1999). Confrontés aux mêmes types de micro-organismes (bactéries, virus, champignons, levures…), les insectes ont développé, au cours de l’évolution, des systèmes de défense performants et conservés. De nombreux outils généti-ques et un système immunitaire simple fondé uniquement sur la réponse innée font de la drosophile un modèle idéal pour disséquer les étapes de l’infection in vivo (Lemaitre, 1999 ; Tzou et al., 2002). Chez la drosophile, l’infection induit l’expression de peptides anti-microbiens (AMPs) présentant de fortes activités antimicrobiennes, permettant de protéger l’organisme des pathogènes. Les gènes codant ces peptides sont régulés par deux voies de signalisation, les voies Toll et imd. Chacune active des facteurs de transcription différents appartenant à la famille Rel/NF-κB. Chez les mammifères, ces facteurs jouent un rôle important dans la régulation de la réponse immunitaire innée et de l’inflammation (Beutler, 2004). La conservation de mécanismes de régulation chez des organismes aussi
distants a souligné l’existence de mécanismes anti-infectieux ancestraux communs à tout le règne animal. Nos connaissances sur ces voies et sur les mécanismes effecteurs de la réponse immunitaire innée sont encore incomplètes. Afin d’approfondir notre connaissance de ce système, des études de génomique ont été réalisées (microarrays, séquençage des génomes de l’hôte et des pathogènes) : elles permettent maintenant de passer à l’ère d’une analyse fonctionnelle post génomique (crible par interférence ARN inductible in vivo). Elle nous permettra d’identifier de nouveaux gènes clés de ce système immunitaire, et de commencer à intégrer les mécanismes moléculaires de l’interaction entre l’hôte et le pathogène. Les mécanismes de défense antimicrobienne de la drosophile Avant l’initiation d’une réponse immunitaire innée, les microorganismes doivent contourner un certain nombre d’éléments de défense (Cf. figure 1). Tout d’abord, ils doivent franchir des barrières naturelles que constituent la cuticule externe, la matrice péri-trophique de l’intestin ou la membrane chitineuse de la trachée (Beutler, 2004 ; Lemaitre, 1999, Tzou et al., 2002). Ensuite ils se heurtent à une barrière chimique. En effet, en ce qui concerne les muqueuses, l’organisme crée des conditions non propices au dévelop-pement du pathogène comme un pH bas dans l’intestin ou encore des enzymes digestifs comme les lysozymes… Lorsqu’un micro-organisme con-tourne ces barrières, tout un ensemble de mécanismes de défense est stimulé. En effet, les insectes, bien que dépourvus de l’arsenal du système adaptatif, présentent une grande résistance aux microorganismes. Ils disposent de mécanismes de défense
proches de ceux mis en jeu dans la réponse innée des mammifères, qui sont fondés sur des réactions cellulaires et humorales (Beutler, 2004 ; Tzou et al., 2002). La réponse cellulaire fait intervenir des cellules équivalentes à celles de la lignée myéloïde des mammifères : les hémocytes (Cf. figure 1). De ces dernières dérivent des cellules spécialisées dans la phago-cytose (plasmatocytes), l’encapsulation de corps étrangers (lamellocytes) et dans la mélanisation (cellules à cristaux) (Tzou et al., 2002). Chez les mammifères, la réponse humorale complète la réponse des cellules du système immunitaire et constitue la meilleure arme contre la propagation des pathogènes. Elle consiste en la synthèse de peptides cationiques qui sont produits essentiellement par le corps gras, l’analogue fonctionnel du foie des mammifères. Ces peptides sont ensuite excrétés dans l’hémolymphe (le sang de la drosophile) : c’est la réponse systémique. Il existe une vingtaine de ces peptides antimicrobiens (AMPs) inductibles qui sont regroupés en sept classes. La diptéricine, la défensine, la drosocine, les cécropines (trois isoformes) ainsi que les attacines ont une action uniquement antibactérienne, la drosomycine a un spectre d’action antifongique, alors que la metchni-kowine a une action mixte (Lemaitre et Hoffmann, 2007). Selon la voie d’entrée du pathogène, les premiers tissus exposés seront les épithélia de l’intestin, des trachées ou de l’épiderme. Ces tissus développent également une réponse immunitaire fondée sur la production locale d’AMPs et de réactifs oxygénés (Cf. figure 1). Cet ensemble de mécanismes constitue la réponse locale. Par exemple, lorsque le pathogène atteint l’intestin, des espèces oxygénées réactives (ROS) sont synthétisées par l’enzyme dual Oxydase

LES BRÈVES
PAGE 31
(duOX) pour l’éliminer (Tzou et al., 2002 ; Bangham et al., 2006). Les gènes codant les AMPs sont peu transcrits normalement, mais sont fortement induits trois à six heures après une infection microbienne ou fongique ou encore une blessure septique. Deux voies de transduction sont mises en évidence comme contrôlant cette transcription, les voies Toll et imd (Hultmark, 2003 ; Lemaitre, 2004). Ces dernières sont activées par la reconnaissance de motifs conservés présents chez les bactéries ou champignons, les PAMPs (Pathogen Associated Molecular Patterns), par des récepteurs spécialisés, les PRRs (Pattern Recognition Receptors). La cascade Toll présente de fortes similarités avec la voie TLR/NFκB des mammifères (Beutler, 2004 ; Lemaitre, 1999). Cette voie est activée essentiellement par les bactéries à Gram positif et les champignons (Cf. figure
2A). La reconnaissance des bactéries à Gram positif se fait par détection de fragments du peptidoglycane de type lysine, caractéristique des bactéries à Gram positif, par les récepteurs PGRP-SA (Peptidoglycan Recognition protein SA) et GNBP1 (Gram Negative Binding Protein 1) qui activent une cascade de clivages et d’activation de protéases à sérine aboutissant à l’activation de spätzle. Cette cytokine, présente dans l’hémolymphe, est le ligand du récepteur Toll et active notamment l’expression de l’AMP drosomycine (Lemaitre, 2004 ; Ferrandon et al., 2004). La voie imd présente des similarités avec la partie intracellulaire de la voie de signalisation du TNF-R (Tumor Necrosis Factor Receptor). Cette voie joue un rôle prédominant dans les infections par des bactéries à Gram négatif (Cf. figure 2A). Le récepteur de la voie imd est une protéine
transmembranaire avec un domaine PGRP (PeptidoGlycane Recognition Protein) : PGRP-LC, qui est capable de détecter les fragments de peptido-glycane de type DAP (Diamino Pimelic peptidoglycane), spécifiquement retrou-vés dans l’enveloppe des bactéries à Gram négatif. Cette cascade conduit par exemple à l’induction de l’expression de la diptéricine (Lemaitre et al., 1997). D’autres PGRP (PGRP-LB, PGRP-SC) ont une activité catalytique qui clive le peptidoglycane, générant des fragments non immuno-réactifs. Ils permettent de définir un seuil d’activation du système immunitaire et régulent négativement ce système pour limiter la durée de la réponse (Cf. figure 2A). Ce système de régulation permet ainsi d’empêcher une activation des défenses contre des bactéries commensales, mais également permet d’éviter d’éventuels dommages aux tissus de l’hôte consécutifs à une
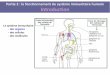
Figure 1 : la réponse immunitaire chez la drosophile : effecteurs et régulation Avant l’initiation d’une réponse immunitaire, l’agent infectieux contourne des barrières physiques (cuticule, mem-brane chitineuse, trachées…) et des barrières chimiques (pH bas, lysozymes...). Puis il initie une réponse immuni-taire comprenant une réponse humorale et cellulaire. La réponse cellulaire fait intervenir les hémocytes (plasmato-cytes, lamellocytes et cellules à cristaux) qui permettent la phagocytose, l’encapsulation et la mélanisation du pa-thogène. La réponse humorale consiste essentiellement en la synthèse de peptides antimicrobiens par le corps gras (réponse systémique) et par les muqueuses (réponse locale).
PAMPs
+Réponse immunitaire
humorale cellulaire
plasmatocytes
lamellocytes
Cellules à cristaux
Régulation
par
Toll
IMD
Barrières physiques
Bactérie gram + ou – et champignons
Phagocytose
Encapsulation
mélanisation
Barrières chimiques
Synthèse AMPs
Autres effecteurs ?
Réponse systémique
CORPS GRAS
Réponse locale
INTESTIN
TRACHEES
pH, lysozymes, ROS…
Cuticule externe

LES BRÈVES
PAGE 31
activation trop importante ou trop longue du système immunitaire (Lemaitre et Hoffmann, 2007). Étude post-génomique de la réponse immunitaire
La majorité des expériences d'infection chez la drosophile ont été réalisées soit par injection des microorganismes directement dans l'hémocœle, soit à l’aide d’une aiguille ou d’un injecteur (Cf. figure 2B). Cette méthode a été utilisée avec succès pour disséquer les mécanismes régulant la réponse immunitaire systémique ou la pathogénie bactérienne (Tzou et al., 2002). Malheureusement, ce type d'infection ne permet pas d'étudier les premières étapes d'une infection naturelle, notamment la colonisation et la persistance de la bactérie dans le tractus digestif. Pour aller au-delà de ces
limites, une méthode d'infection orale a été mise au point afin d'étudier la physiologie des interactions entre l’hôte et les pathogènes (Cf. figure 2B) (Tzou et al., 2002 ; Lemaitre et Hoffmann, 2007). Ainsi on a pu isoler la souche Erwinia carotovora spp. carotovora 15 (Ecc15), un phytopathogène qui utilise la drosophile comme vecteur (Basset et al., 2000).
Récemment, il a été également isolé une bactérie, Pseudomonas entomophila (Pe), qui est capable d'infecter et de tuer les drosophiles. Il s’agit d’une nouvelle espèce de bactérie entomopathogène qui possède un large spectre d’hôte (Liehl et al., 2006). La réponse immunitaire de la drosophile à l’infection naturelle par ces pathogènes est complexe et met en jeu de nombreux acteurs et effecteurs dont certains encore inconnus, sans doute différents de ceux impliqués dans la
réponse systémique. La plupart de ces gènes participent à la réponse de l’hôte, modulent la persistance des bactéries dans l’intestin et représentent donc des cibles d’études prometteuses pour la compréhension du processus infectieux (Tzou et al., 2002 ; Lemaitre et Hoffmann, 2007). Une stratégie pour identifier et caractériser ces gènes clés est l’approche par génétique inverse consistant à inhiber in vivo l’expression d’un gène et à analyser le phénotype associé (Bangham et al., 2006). Nous avons donc débuté au laboratoire un crible pan-génomique destiné à éteindre in vivo, un par un, l’expression de chaque gène du génome de la drosophile par la technique d’interférence ARN inductible in vivo. Plusieurs groupes ont montré que la présence d’ARN double brin (ARNdb) induit la dégradation du transcrit du gène endogène. Ainsi, l’injection d’un
Blessure septique Infection orale
ETAT D’ACTIVATIONDU SYSTÈME IMMUNITAIRE
SUSCEPTIBILITEAU PATHOGENE
Bactéries Gram -
DAPPGRP- LB
PGRP-LC
relish
κB
Diptericin
κBκB
Voie Imd Voie Toll
Bactéries Gram + ou Champignons
Spaetzle
dorsal dif
Dorsal
κB κB κB
Drosomycin
Toll pathwayImd pathway
PGRP-SA
GNBP1
GNBP3
Serine proteases
Figure 2 : régulation de la synthèse des peptides antimicrobiens et étude de la réponse innée de la drosophile A/ Les bactéries à Gram+ et les champignons activent la voie Toll alors que la voie imd est déclenchée par les bac-téries à Gram-. La reconnaissance des bactéries Gram+ par PGRP-SA et GNBP1 et des champignons par GNBP3 entraîne une cascade protéolytique aboutissant au clivage de la cytokine spätzle. Cette dernière est le ligand du récepteur Toll, qui entraîne la translocation nucléaire des facteurs de transcription dorsale et dif conduisant ainsi à l’expression de la drosomycine. La reconnaissance des bactéries à Gram- par PGRP-LC conduit à l’activation de la voie de signalisation Imd, aboutissant à la translocation nucléaire de relish et à l’expression de la diptericin. B/ Afin d’étudier la réponse immunitaire chez la drosophile, différentes lignées sont infectées par blessure septiqueou par infection orale. La susceptibilité de ces lignées aux infections ainsi que l’état d’activation du système immuni-taire sont étudiés.

LES BRÈVES
PAGE 31
fragment d’ARNdb dans l’œuf de drosophile peut mimer un phénotype « perte de fonction du gène endogène » au cours du développement. Pour étudier la fonction d’un gène à un stade plus tardif (larves, adultes), il suffit d’induire l’expression d’ARNdb dans des drosophiles génétiquement modifiées. Notre équipe a donc établi une collaboration avec le laboratoire du Dr Ryu Ueda du National Institute of Genetic, Mishima, Japon. Il s’est lancé dans la production de lignées UAS-IR à
grande échelle, capables de cibler plus de 9000 gènes du génome de la drosophile. Cette collaboration permet pour la première fois d’envisager une étude systématique pan-génomique des gènes impliqués dans le système immunitaire de la drosophile. Les progrès réalisés en génomique et dans le domaine de l’immunité innée permettent de bien appréhender aujourd’hui les mécanismes mis en place par l’hôte dans la reconnaissance et l’élimination d’agents infectieux dans un environnement non septique. Un des
défis majeurs aujourd’hui est d’identifier, au sein des muqueuses, les mécanismes permettant l’élimination spécifique d’un pathogène parmi la flore microbienne. Ainsi on pourra mieux comprendre l’articulation entre réponse locale et systémique ainsi que la physiologie du processus infectieux. Cette approche post-génomique et l’utilisation de pathogènes naturels comme Ecc15 et Pe qui évoluent avec l’insecte rendent possibles de tels projets. ¦
Références : • Beutler B. Innate immunity : an overview. Molecular Immunology 2004 ; 40 : 845-59. • Lemaitre B. La drosophile : un modèle pour l’étude de la réponse immunitaire innée. Médecine Sciences 1999 ; 15 : 15-22. • Tzou P., De Gregorio E., Lemaitre B. How Drosophila combats microbial infection : a model to study innate immunity and
host-pathogen interactions. Curr Opin Microbiol 2002 ; 5 : 102-10. • Bangham J., Jiggins F., Lemaitre B. Insect Immunity : The Post-Genomic Era. UKPMC Funders Group 2006 ; 25(1) : 1-5. • Lemaitre B., Hoffmann J. The Host Defense of Drosophila melanogaster. Annu Rev Immunol 2007 ; 25 : 697-743. • Hultmark D. Drosophila immunity: paths and patterns. Curr Opin Immunol 2003 ; 15 : 12-9. • Lemaitre B. The road to Toll. Nat Rev Immunol 2004 ; 4 : 521-7. • Ferrandon D., Imler J.-L., Hoffmann J.A. Sensing infection in Drosophila: Toll and beyond. Semin Immunol 2004 ; 16 :
43-53. • Lemaitre B., Reichhart J.-M., Hoffmann J. A. Drosophila host defense : Differential induction of antimicrobial peptide
genes after infection by various classes of microorganisms. Proc. Natl. Acad. Sci. USA, 1997 ; 94 : 14614-9. • Basset A., Khush R. S., Braun A., Gardan L., Boccard F., Hoffmann J. A., Lemaitre B. The phytopathogenic bacteria Er-
winia carotovora infects Drosophila and activates an immune response. Proc. Natl. Acad. Sci. USA., 2000 ; 97 (7) : 3376-81.
• Liehl P., Blight M., Vodovar N., Lemaitre B. Prevalence of local immune response against oral infection in a Drosophila / Pseudomonas infection model. PloS Pathogens 2006 ; 2.