Embed Size (px)
Citation preview

Herrn Prof. Dr. Vald. Henriquesan seinem 70. Geburtstag gewidmet.
Reserves azotees musculaires 1
par
Andree Roche.
(Travail du Laboratoire de Chimie Biologique de la Faculte de Medecine et dePharmacie de Marseille, France.)
I. Introduction.
La possibilite de I'existence de reserves azotees musculaires a donnelieu, depuis longtemps deja, a des discussions passionnees. Pour Folinet Denis (1), ces reserves ne seraient pas de nature proteique, maisformees par des acides amines retenus dans Ie muscle par simple adsorption. Abderhalden suppose qu'il existe dans ce tissu une proteinede reserve, distincte de celIe du protoplasme. Terroine, au contraire,admet que l'azote mobilise par I'animal en cas de besoin, est d'origineprotoplasmique. Aucune de ces hypotheses n'est cependant satisfaisante,chacune d'elle s'oppose en effet a certains faits sur lesquels il n'est paspossible d'insister ici. Aussi, afln de concilier toutes Ies donnees connues,Mac Collum (2) a-t-il suppose que, lorsque I'azote indispensable aufonctionnement de l'animal ne lui est plus fourni par sa ration, il seIe procure grace a 131 dislocation partielle de ses proteines museulairesavec mise en liberte de polypeptides simples ou d'acides amines facilementutilisables par I'organisme. Cahn (3) s'est efforce de mettre en evidenceun tel mecanisme au cours du jeune ou de l'atrophie musculaire, maisses resultata ne permettent pas, de son avis meme, de eonsiderer l'hypothese de Mac Collum comme demontree, C'est une telle confirmationexperimentale que je me suis efforcee de realiser, Elle a ete exposee endetails ailleurs (4), aussi me bornerai-je ici, a n'en rapporter que les resul-·tats essentiels indispensables a l'etude du mecanisme par lequel il estpossible de I'expliquer.
1 Der Redaktion am 6. Januar 1934 zugegangen,

76 A.~DREE ROCHE:
II. Recherches personelles,Ces recherches sont de deux ordres, elles consistent:
. A. En une etude des pertes azotees subies par un animal soumisit 1'inanition proteique jusqu'a ce que mort s'ensuive.
B. En une analyse des constituants ssotes du muscle en fin d'experience, dans le but d'etablir la repercussion sur la constitution chimiquede ce tissu, des pertes azotees subies par 1'animal.
A. Pertes azotees a l'inanition totale et a l'inanition proteique.
L'inanition proteique au cours de laquelle l'animal a tous ses besoinscouverts exeepte son besoin en azote, doit permettre la survie de l'animaltant que 1'azote indispensable it son entretien peut lui etre fourni par sespropres tissus. Cette experimentation doit faire subir it l'animalles pertesazotees maxima compatibles avec la vie. Elle realise done la meilleurecondition pour l'etude de la repercussion de ces pertes sur la constitutionchimique du muscle.
Les experiences ont porte sur 40 rats, dont 21 soumis it l'inanitiontotale, et 19 soumis it 1'inanition proteique." Les animaux sont placesdans des cages it metabolisme, 1'azote urinaire est determine tous les deux[ours, l'azote fecal tous les quatre ou cinq [ours. Voici tres brievementresume 1'ensemble des resultats (tableau 1) dont Ie detail est publie dansun travail d'ensemble (4):
Tribleau 1.
Survie Perte Perte azoteePerte en tissu prot.3 100depoids
en mg d'Nlen % d'N(jours) % . total-
Perte de poids X
Inanition totale(21 animaux) 2 a. 8 30 197 a1115 15 50
Inanition pro-teique(19 animaux) 288,72 48 4458,3770 23 8, 55 508, 150
1 Lea animaux au [enne total rC90ivent de l'eau ad libitum; ceux soumisa l'inanition proteiqne, un regime constitue de 10. f&9Qn suivante: amidon, 70;beurre,30; melange salin d'Osbome, 5; sciure, 5; vitamines. Une telIe rationcontient environ O·6 mg. d'azote par gramme.
II La teneur en azote des a.ni:ma.ux au debut de I'experience est calouleea l'aide de valeurs moyennes determinees sur 50 rats environ.
3 Le tissu proMique qui correspondrait a. l'azote est calcule en multipliantIs perte d'azote par 6·25 (tdssu sec), Ies proteines musoulaires ayant une teneuren azote de 16%, puis par 5 (tissu frais), Ie muscle contenant en eHet environ80% d'eau.

RESERVES AZOTEES MUSCULAIRES. 77
Des rats soumis it l'inanition proteique jusqu'a ce que mort s'ensuive,peuvent done perdre de 25 it 55% de leur azote total. En plus desreserves hepatiques (Berg, Noel, Luck, Gautier), insuffisantes pourexpliquer de telles pertes, il doit exister dans l'organisme, une autresource d'azote. Or, l'examen des animaux apres leur mort, et l'aspecthistologique de leur muscle, dont les fibres ont subi une diminution devolume considerable, montrent que ce tissu a contribue pour une partimportante ala couverture du besoin azote de l'animal. Ainsi se trouvepose anouveau le problems de l'existence de reserves azotees au niveaudu muscle.
Si l'on exprime 180 perte en azote sous forme de tissu proteique frais,celle-ci representerait toujours environ 50% de la perte de poids, chezle rat inanitie; 'chez le rat mort d'inanition proteique, elle depasseraitparfois 100% de eelle-ci. Ce resultat, paradoxal en apparence, montreque ce mode de ealeul n'est pas justifie. Le depart global d'une proteinetissulaire n'est done pas le seul mecanisme par lequel l'organisme seprocure l'azote qui lui est neeessaire, et il est permis de penser qu'il y aen outre, depart fragmentaire de 180 molecule. Les polypeptides simplesou les acides amines ainsi liberes, d'origine proteique, ne sont pas euxmemes colloidaux et, leur mobilisation n'altere pas Ioreement le taux eneau de gonflement du tissu. Le coefficient 5 applique au cours du calcuIindique plus bas, ne repond done pas a180 realite. Ainsi l'inanition proteique poursuivie jusqu'a 180 mort, realise une condition experimentaleau cours de 180 quelle l'animal subit une fonte musculaire analogue engrandeur aeelle, bien etudiee par Miescher et par Greene, du Baumanau eours du frai. Les animaux, ainsi prepares, consituent un materiel dechoix pour l'etude du problema qui nous preoeeupe,
Les experiences que nous venons de deerire brievement apportenten outre une serie de faits favorables a l'hypothese proposee parMac Collum.
B. Etude comparee des constitaants azotes du muscle et de sea proteineschez Ie rat normal ou mort d'inanition totale ou proteique.
On trouvera ailleurs (4) une analyse detaillee des principaux con- .stituants cellulaires du muscle chez le rat normal ou mort d'inanitiontotale ou proteique. Je me bornerai ici areproduire les valeurs moyennesdu taux de l'azote extractif (hydrosoluble et soluble dans 180 benzine),et des differents constituants des preteines museulaires, donnees indispensables pour 180 discussion qui va suivre.
Techniques. - Les techniques utilisees sont deerites dans Ie travailcite plus haut (4), je ne puis, faute de place, les exposer ici. L'azote

78 ANDREE ROCHE:
extractif a ete prepare selon Cahn, et tous les dosages relatifs aux proteines musculaires ont ete effeetues sur la poudre musculaire degraisseesans cendres et sans extractif (technique de Janney modifies par Cahn).Une telle preparation contient pratiquement a I'etat pur les proteinesmusculaires et les noyaux cellulaires, ces derniers representant au plus2% du poids total.
Resultats experimentaux.
Les valeurs moyennes obtenues pour les differentes donnees etudieessont reunies dans les tableaux II, III et IV.
a) Azote extractif du muscle.L'azote extractif du muscle comprend deux fractions, l'une soluble
dans la benzine, l'autre soluble dans l'eau. La premiere representspratiquement l'azote des lipoides du tissu, elle n'est retenue que pourpermettre Ie calcul de l'azote extractif total.
L'azote extractif hydrosoluble, au eontraire, doit etre examine iciattentivement. Cette fraction est constituee principalement par de l'uree,de l'ammoniaque, de la creatine, de la creatinine, des nucleotides et desacides amlnes ou des combinaisons de ceux-ci, parmi lesquelles la carnosineet le glutathion occupent une place importante. La fonte musculairesubie au cours de l'inanition proteique montre immediatement que lesacides amines de cette fraction ne constituent pas la seule reserve azoteeau niveau du muscle (Folin et Denis). Par ailleurs, Van Slyke etMeyer (5), Wishart (6), Mitchell, Nevens et Kendall (7), ontmontre que des regimes riches ou pauvres en proteines ne faisaient pasvarier son taux dans les differents tissus. G. D. Cathcart (8), a precisel'existence d'un rapport entre l'azote amide, l'azote amine et l'ammoniaquede l'extractif musculaire. Cette constance remarquable a fait supposeraVan Slyke et Meyer que l'azote extractif du muscle joue non aproprement parler le role d'une reserve, mais devrait ere considere comme letemoin d'un equilibre entre la destruction des proteines tissulaires et leuredification aux depens des acides amines exogenes.
L'etude des variations de cette donnee au cours de l'inanition proteique, doit permettre de lui assigner sa veritable signification. Lesresultats de ce dosage sont consignee dans Ie tableau II.
Si les acides amines de l'extractif representaient aproprement parlerune reserve, lis devraient disparaitre rapidement; or l'examen du tableauIImontre que l'azote extractif du muscle, non seulement ne diminue pas,mais augmente de 21% en moyenne, lors de la mort par inanition proteique, et de 39% lors de la mort par inanition totale.
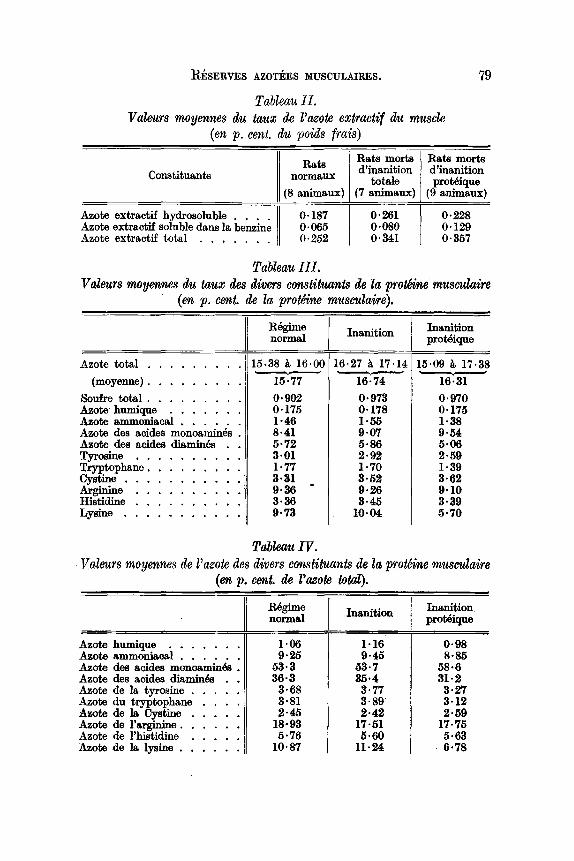
RESERVES AZOTEES MUSCULAIRES.
Tableau 11.Valeurs moyennes du tau» de l'azote extractif du muscle
(en p. cent. du poids (rais)
79
ConstituantsRats
normaux
(8 animaux)
Rats mortsd'inanition
totale(7 animaux)
Rats mortsd'inanitionproMique
(9 animaux)
Azote extractif hydrosolnble . . . '11 0·187Azote extractif soluble dans 180 benzine 0·065Azote extractif total . . . . . .. o· 252
0·2610·0800·341
0·2280·1290·357
Tableau 111.Valeurs moyennes du tau» des divers constituants de la proteine musculaire
(en p. cent. de la proteine musculaire).
Azote total .
(moyenne) .
Soufre total .Azote· humique .Azote ammoniacal • . . . . .Azote des aoides monoamines .Azote des acides diaminasTyrosine ..Tryptophane.Cystine.ArginineHistidineLysine .
Regimenormal
15·38 a 16·00--------15·770·9020·1751·468·415·723·011·773·319·363·369·73
Inanition
16·27 a 17·14----16·740·9730'1781'559·075'862·921·703·529·263·45
10·04
Inanitionproteique
15·09 a 17·38-.--16·310·9700·1751·389·545·062·591·393·629·103·395·70
Tableau IV.Valeurs moyennes de l'azote des divers constituants de la proteine musculaire
(en p. cent. de l'azote total).
Regime Inanition Inanitionnormal proMique
Azote humique . 1·06 1·16 0·98Azote ammoniacal . 9·25 9·45 8·85Azote des acides monoamines • 53·3 53·7 58·6Azote des aoides diaminas 36·3 35·4 31·2Azote de ls tyrosine . 3·68 3·77 3·27Azote du tryptophane 3·81 3·89 3·12Azote de la Cystine 2·45 2'42 2·59Azote de l'arginine. 18·93 17.51 17·75Azote de l'histidine 5·76 5·60 5·63Azote de la lysine . 10·87 11·24 6·78

80 ANDREE ROCHE:
Par ailleurs les acides amines de l'extractif sont plus facilementaccessibles pour l'organisme que ceux des proteines musculaires, dontl'utilisation n'est possible qu'apres une proteolyse prealable, aussi semmesnous conduits a admettre que l'azote extractif hydrosoluble du musclejoue le role d'un intermediaire (Shermann[9], Van Slyke etMeyer[5])dans un equllibre dont nous discuterons plus bas l'existence et lesmodalites.
b) Composition de la proteine musculaire.Je rapporterai ici les valeurs moyennes obtenues pour l'azote total,
l'azote humique, l'azote ammoniacal, l'azote des acides monoamlnes etdes acides diamines, le soufre, la cystine, la tyrosine, Ie tryptophane,l'arginine, l'histidine et la lysine. Les resultats sont rassembles dansles tableaux III et IV, 0\1 ils se trouvent exprimes respectivement enp. cent. des proteines musculaires totales et en p. cent. de l'azote totalde eelles-ei,
Nous allons examiner rapidement l'ensemble des resultats obtenus.
Azote total des proteines musculaires. La teneur en azote total de lapoudre musculaire est en moyenne de 15·77% chez les rats normaux etde 16'74% chez les animaux morts d'inanition totale. Les ecarts autourde la moyenne ne depassent pas 2% dans les deux cas. Chez les animauxmorts d'inanition proteique, la teneur en azote de la poudre est en moyennede 16'31%, mais ici au contraite les ecarts individuels sont considerablea(de 15·09 a17·38%).
Azote des acides monoamines et des acides diamines. Le taux desacides monoaminea augmente Iegerement a l'inanition totale, plus fortement .au cours de l'inanition proteique. En meme temps, on voit Ietaux des acides diamines s'abaisser au [eune total, et surtout au jeftneazote.
Acides amines etudies individuellement. L'inanition totale n'alterepratiquement pas la composition en acides amtnes des proteines museulaires, L'inanition proteique au contraire determine des modificationsprofondes de celle-ci, caracterisees comme il suit: Tandis que certainsacides amines tels que l'arginine, l'histidine,et la cystine, eonservent untaux normal ou legerement augmente, d'autres tels que la tyrosine, .:Ie tryptophane, et surtout la lysine, diminuent notablement.
Ces differentes variations s'accordent pour etayer l'hypothese d'unedislocation partielle du protoplasme, En effet, Ie depart global d'unemolecule proteique, tel que l'ont suppose Terroine, Brenckmann etFeuerbach (10), sans etre exclu a priori, ne peut pas etre Ie seul mecanisme par lequel l'organisme se procure l'azote indispensable a sonentretien, Un tel mode de couverture determinerait en effet une di-

RESERVES AZOTEES'MUSCULAIRES. 81
minution du taux des proteines dans .le muscle, mais non une variationde leur composition.
Les modifications de composition des proteines musculaires sontassez sytematiques pour qu'au premier abord on puisse penser au departd'une proteine de reserve, mais les variations non systematiques du tauxde l'azote total, indiquent au contraireque l'azote soustrait au muscleau cours de ces experiences ne peut provenit que de la dislocation partielleet progressive d'une proteine, et de la mise en Iiberte de polypeptidessimples ou d'acides amines plus ou moins riches en azote.
Rien ne permet de penser que la proteine ainsi fragmentee est uneproteins de reserve. L'examen histologique n'a jamais revele l'existencedans le muscle d'enclaves proteiques eomparablea a celles trouvees parBerg et par Noel dans Ie cas dufoie. Aussi y a-t-il tout lieu de croireque la liberation de polypeptides se fait aux depens du protoplasme, etl'hypothese de Mac Collum se trouve ainsi experimentalement confirmee,
III. Discussion des resultats et conclusions.
A. Reserves azotees. - f:quilibre azote~
Un animal peut done supporter un jeftne azote pendantuntem.pstres long. Une fois ses reserves hepatiques epuisees, il couvre son besoinen azote par dislocation partielle du protoplasme musculaire, et mise enliberte de polypeptides simples ou d'aeides amines facilement utilisablespar l'organisme. Ce mode de eouverture supprime tout phenomenadiscontinu tel que celui realise par la liberll.tion brusque d'une moleculetotale, il explique la constance de l'azote extractif, et permet de concevoirce mecanisme de la maniere suivante: l'azote extractif, intermediaireentre l'edification ou la destruction des tissus et l'azote exogene, estengage dans un double equllibre.
1. Acides amines de l'extractif -~ Acides amines des proteines tissulaires,
2. Acides amines des proteines tissulaires -~ Aeides amines de l'extractif.
Chez l'animal en croissance, ou lorsqu'on passe d'un regime pauvrea un regime riche en azote, l'equilibre 1. predomine, Inversemerit,Ie passage d'un regime riche en azote aun regime pauvre ou au [efmedeplaee l' equilibre en sens inverse. Enfin chez un animal adulte normalen equilibre azote, les deux reactions sont equivalentes. '
Ainsi s'explique tres simplement la loi de Bischoff 'et Voit stirI'equilibre azote, et le fait que la vitesse avec laquelle un 'animal passe
Skandinav. Archiv. LXIX. 6

82 ANDREE ROCHE:
d'un equilibre a un autre depend du regime auquel il a ete anterieurementsoumis.
Est-on en droit de parler de «reserves» azotees musculaires? Non,si l'on donne a ce terme la signification qui lui revient lorsqu'il estapplique a des enclaves telles que le glyeogene hepatique, chezles animauxou l'amidon chez les vegetaux, - Oui, si l'on entend par la que l'azoteainsi libere sert a l'entretien de l'animal et peut-etre pris aces tissus sansmettre, au debut tout au moins, sa vie en danger. Cette discussion perdrad'ailleurs, nous Ie verrons, toute signification lorsque nous aurons etabliIe mecanisme par lequel se fait cette dislocation.
B. Besom mdifferencie et besom differencie d'azote.Les acides amines etudies individuellement au cours de ces ex
periences ont, soit garde un taux constant (arginine histidine, cystine),soit subi nne diminution systematique (tryptophane, tyrosine, lysine),aussi les variations anarchiques du taux de l'azote total doivent-ellesetre imputees a des variations non systematiques des acides aminea queje n'ai pas etudie en detail, c'est a dire des acides monoamines a chainedroite. nest permis de penser que ceux-ei jouent dans la molecule unrole identique (plastique) et sont tous plus ou moins interchengeables,expliquant ainsi l'existence d'un besoin «non differencie» d'azote.
Par ailleurs les acides smines dont le taux diminue regulierementsont justement tous trois indispensables soit a la croissance, soit al'entretien, ils participent au besoin en azote «differencie II de l'individu.
c.Dislocation pariieUe du protoplasme et plasticiie des protemes muscuIaires.La dislocation partielle du protoplasme mise en evidence par ces
experiences suppose nne grande plasticite de la molecule proteique. Onpeut se demander queUes sont les limites de cette plasticite et [usqu'aquel point la composition en acides amines d'une proteins donnee estsusceptible de varier.
Chez le rat normal, la composition des proteines musculaires estsensiblement fixe. Normalenient tout au moins leur edification subit undeterminisme certain. Il parait peu probable, par ailleurs, que la constance du taux de certains acides amines tels que l'arginine oul'histidine,soit le fait d'une simple coincidence. Ne doit-on pas faire intervenir icil'hypothese formulae il y a longtemps deja par Kossel, de l'existeneedans la molecule proteique d'un noyau, veritable groupement prosthetiquede la molecule, constitue principalement par des bases hexoniques (arginine), conditionnant sa grandeur et lui conferant des proprietes physicoehimiqueadetermineea?

RESER':'ES AZOTEES MUSCULAlRES. 83
Enfin, la forte diminution du taux de la lysine, acide amine indispensable it la croissance, nous permet de penser que cette substanceoccupe elle aussi une place particuliere dans la molecule proteique,
Les variations anarchiques du taux des differents acides monoaminesa chaine droite, signalees plus haut, permettent de penser que ces acidespeuvent parfois se substituer les uns aux autres.
D. Plasticite des proteines en general et structure de leur molecule.
Une dislocation partielle in vivo, des proteines tissulaires, telle quecelIe mise en evidence au cours de ces experiences, n'est pas un fait isole,Un calcul simple permet de montrer qu'un phenomene analogue se produitchez Ie Saumon au cours du frai.! Miescher a en effet montre (11) que .l'arginine neeessaire a la formation du testicule de l'animal represente35% de l'arginine musculaire totale. Or, en fin du frai, l'animal quipesait au debut de I'experience 9 kg. environ, n'a perdu que 600 gr. depoids et cela presque uniquement aux depens de son muscle. Une tellefonte musuclaire n'aurait libere que 5 gr. 4 d'arginine, alors que le tissutesticulaire neoforme en contient 23 gr. D doit done se faire chez Iesaumon une dislocation selective de la proteine protoplasmique, seulecapable d'expliquer cette mobilisation massive d'arginine. En accordavec cette maniere de voir il y a lieu de noter que le taux de l'azote dumuscle tombe de 18·5 a 13·5%; (l'arginine contient 32% d'azote).
Les proteines seriqnes et meme l'hemoglobine, presentent uneplasticite analogue, ainsi qu'il ressort des travaux reeents de Fischeret Blankenstein, Lustig, Schenck et Kunstmann (12) pour lespremieres, de Schenck, de Roche et Dubouloz (13) pour la seconde.D est possible que, comme l'avait prevu Kossel, cette notion importantedoive etre generalisee et que toute proteins, tant animale que vegetale,puisse presenter, d'un sujet aun autre, une composition en acides aminesdifferente, tout en conservant des reactions chimiques et des proprietesphysieo-chimiques identiques. Seules des difficultes d'ordre techniqueont empeche jusqu'ici d'etablir ce fait avec certitude.
Les donnees que nous venous d'exposer permettent de remettre endiscussion la conception classique de la molecule. proteique, assignant acette derniere une composition et un poids invariables. Les proteines,substances colloidales a poids moleeulaire eleva, doivent a ce caractereIa plupart de leurs proprietes physleo-ehimlques, C'est lui qui conditionne
1 On sait que lorsque le Saumon remonte Ie Rhin, il reate de 5 it 15 moisdans l'eau douce sans s'alimenter. Pendant cette periods, les testicules de l'animalse developpent aux depens de ses proMinesmusculaires.
6*

84 ANDREE ROCHE:
leur structure. Les travaux reeents, plus specialement ceux de Staudinger (14), de Meyer et Mark (15) de Bergmann (16) pourles hydratesde carbone, ceux de Svedberg (17) de Sorensen (18) pour les proteines,ont permis de montrer que les grosses molecules quelles qu'elles soientpossedent une structure qui leur est propre. L'etude des roentgenogrammes a revele par exemple, que la molecule de cellulose se composaitde 1500 it 2000 molecules de glucose, ces molecules sont groupees enpremier lieu en «chaines de valences primaires» (Meyer et Mark), itraison de 30 it 50molecules de glucose par chaine. La longueur de celle-cifait naitre des forces coordinatives ou forces de van der Waals quirelient entre elles les differentes chaines. Ce mode de liaison differeessentiellement de celui realisepar les covalences primaires par son caractere de re:versibt'lite.
Les proteines se distinguent des autres micelles par la multiplicitedes eltlments simples(acides amines) qui les constituent. Leurs « chainesde valences primaires » ou «constituants independants . (Sorensen)principalement, si nonuniquement, denature polypeptidique, sont Iormeespar un grand nombre d'acides amines differents (27 actuellement isoles).En raisonde cette tres grande eomplexite, la roentgenographie n'a donne[usqu'ici, exeepte pour la fibroine de la soie, qu'une image confuse, aussine posssde-t-on aucun indice ni sur le nombre de ces chaines, ni sur leurcomposition. En particulier, il n'est pas possible de savoir si dans unememe molecule ces chaines sont toutes identiques, ce qui impliqueraitpour chacune d'elles, 1'obligation d'etre tres longues et des posseder tousles aeides amines representes dans la proteine etudiee. Si au contraireunemolecule proteique contient un grand nombre de chaines differentesehaeune d'ellespourra etre soit de constitution complexe, soit destructurerelativement simple, comme c'est le cas pour la partie eristalline de lafibroine de la soie.
.Le faitqu'une meme proteine peut avoir une composition en aeidesaminesdifferente d'un animal it un autre, constitue un argument puissanten faveur de la seconde de ces possibilites. Dans le premier cas, en effet,des variations du taux de certains acides amines impliquent I'ouverturede valences primaires, veritable hydrolyse, difficile a imaginer sans unedislocation totale de la molecule. Dans le second cas, au contraire, il y asimple deplaeement d'un equilibre reversible, liberant des « chaines devalences primaires», La dislocation partielle des proteines du muscle,obervee au coursde I'inanitionproteique, se ferait alors au fur et amesureque les acides amines de 1'extractif sont utilises, suivant la reaction dontil a ete question plus haut:
acides aminesdes proteines du muscle -~ 'acidesaminesde l'extractif.

RESERVES AZOTEES MUSCULAlRES. 85
R~sum6.
II a ete possible d'apporter quelquesprecisions:1. Sur la grandeur de la perte asotee maxima que peut supporter
un animal. .2. Sur la repercussion de, cette perte sur IIi. composition chimique
de muscle.3. Ces donnees ont permis de montrer que, au oours de I'inanition
proteique, Ie muscle contribue pour une part importante ala couverturedu besoin azote de l'animal, et cela grAce a une dislocationpartielle etprogressive du protoplasme. L'hypothese physiologique de Mac Collum,reeoit ici une confirmation experimentale basee sur des faits d'ordrechimique.
4. Les experiences presentees dans ce memoirs montrent que pourune espece animale donnee, la meme proteine tissulaire peut presenterdans. diverses conditions experimentales une composition differente enacides amines, D'autres auteurs ont observe Ie meme fait pour les proteines seriques ou la globine de l'hemoglobine, Sa generalite impliqueun certain nombre de consequences quant a la structure de la moleculeproteique. ElIes sont discutees en detail. '
Bibliographie.
1. Folin, O. et Denis, W., Protein metabolism from the standpoint ofblood and tissue analysis. J. of biol. Ohem. 1912. Vot XI. p. 87~95.
2. Mao Collum, E. V. et Steenbook,H., Wi8c. Arg. Exp. Sta. Yea,Bull. 1912. No.2!. p. 69; oite d'a.pres Mitohell, H. H. et Hamilton, T. S.,The biochemistry of the amino acids. Ed. The Ohemical Oatalog 00. New York.1929. Vol, I. p.619. . .
3. Cahn, Th., Recherches de Physiologie generale sur les variationsdes equilibres deeconstltuants oellulaires aueours du developpement embryonnaireet du jefine, Constitution et evolution de la molecule des protides. TAble deSeiencee, Paris. 1928. Gaston Doin, ed. p. 190 et, Degenerescenoe musculaire.Assimilation fonotionnelle et action nerveuse. Contribution a I'etude des equilibresoellulaires. Thble de Mblecine. Les Editions universitaires de Strasbourg. 1926.p.I45.
4. Roche, A., Inanition proteique, reserves azotees et constitution desproteines musoulaires, ThbJedeMblecine. G. Patissier, Ed., Marseille. 1933. p.229.
5. Van Slyke, D. D. et Meyer, G. M., The fate of protein digestionproducts in the body. V. The effects of feeding and fasting on the amino-acidcontent of the tissues. J. of biol, Okem. 1914. Vol. XVI. p.231-233.
6. Wishart, M. B., Animal calorimetry, IX. The Influence of meatdigestion on the amino-acid content of blood and muscle. Ibidem. 1915. Vol. XX.p. 535--537.

86 ANDREE ROCHE: RESERVES AZOTEES MUSCULAIRES.
7. Mitchell, H. H., N evens, W. B. et Kendall, F. E., The relationbetween the endogenous catabolism and the non protein constituents of thetissues. Ibidem. 1922. Vol. LII. p.417-437.
8. Cathcart, G. D., Nitrogen distribution in the tissues and some ofthe factors which influence it. Biochem. Joum, 1916. Vol. X. p. 197-244.
9. Shermann, H. C., Protein requirement of maintenance in man andthe nutritive efficiency of bread protein. J. of biol, Ohem, 1920. Vol. XLI.p.97-109.
10. Terroine, E. F., Brenckmann, E. et Feuerbach, A., La com·position des organismes et les problemes generaux de 180 nutrition. Arch. Intern.Physiol. 1930. T. XX. p. 446-486.
11. Miescher, F., Statistische und biologische Beitrll.ge zur Kenntnisvom Leben des Rheinlachses im StiBwasser. Die histochemischen uoo physio.logischen Arbeiten von F. Miescher. 1897. Leipzig. 116.
12. Fischer, A. und Blankenstein, A., Untersuchungen tiber die ehemisohe Konstitution der Serum. und OrganeiweiBkorper III und IV. Bioehem,Zeitschr. 1930. Bd. CCXXVIII. p. 437-442 und 1931. Bd. CCXXXI. p. 404bis 411. - Lustig, B., Zur Kenntnis der Unter£raktionen der Globuline undAlbumine im Serum. EbeOOa. 1930. Bd, CCXXV. S. 4-6 und 1931. Bd,CCXXXVIII. S. 307..,-320. - Lustig, B. und Haas, P., Eberula. 1931. Bd.CCXXXI. S. 472-492.
13. Schenck, E. G., Untersuchungen tiber das Globin bei Tieren, gesunden .und kranken Menschen. Ein Beitrag zur Kenntnis der dynamischen Konstitutiondes Hamoglobins. Arch. f. expo Patkol. U. Pharmalcol. 1930. Bd. CL. S.160-172.- Roche, J. et Dubouloz, P., Sur 180 specificiM des globines dans les hemoglobines. O. R. Soo. Biol. 1933. T. CXIII. p.317-319.
14. Staudinger, H., d'epres Meyer, K. et Mark, H., Voir (15) page 55et suivantes.
15. Meyer, K. und Mark, H., Der Aufbau der hoohpolymeren organisc/l,enNaturstoffe. Akademische Verlagsgesellschaft m. b. H. Leipzig. 1930. Bd, I.S.264.
16. Bergmann,M., Allgemeine Strukturchemie der komplexen KohlenJ1ydrate und der Proteine, Ber, der dtsch. Ohem. Ge«. 1926. Bd, LIX. S. 2973pis 2981.. 17. Svedberg, Th., The molecular weight of the proteins in the nativestate. Nature. 1931. Vol. CXXVIlI.· p. 999-1000.
18. Sorensen, S. P. L., The constitution of soluble proteins. (Reversiblydissociable component systems.) C. R. Lab. Carlsberg. 1930. Vol. XVIII. No.5.p.1~124.





























