Embed Size (px)
Citation preview

SARDELLA ANNE
RÔLE DES LYSOPHOSPHOLIPIDES DANS LA PHYSIOPATHOLOGIE DE LA POLYARTHRITE
RHUMATOÏDE
Mémoire présenté à la Faculté des études supérieures de l’Université Laval
dans le cadre du programme de maîtrise en immunologie-microbiologie pour l’obtention du grade de maître ès sciences (M.sc.)
FACULTÉ DE MÉDECINE UNIVERSITÉ LAVAL
QUÉBEC
2010 © Anne Sardella, 2010

ii
Résumé
La polyarthrite rhumatoïde (PR) est une maladie auto-immune touchant environ 1% de la
population Canadienne. Les nombreux traitements disponibles sur le marché présentent
encore des inconvénients majeurs: résistances aux médicaments sur le long terme, et
efficacité partielle ou nulle des traitements. C’est pourquoi il est constamment nécessaire de
développer de nouvelles cibles thérapeutiques. À ce propos, l’acide lysophosphatidique
(LPA) et la sphingosine-1-phosphate (S1P) semblent jouer un rôle dans l’inflammation
chronique impliquée dans l’arthrite. Dans le liquide synovial des patients atteints de la PR,
on retrouve non seulement du LPA, mais aussi certains produits de son catabolisme et
l’enzyme qui le produit. De plus, le TNF-α associé à l’inflammation est capable d’amplifier
les effets du LPA et du S1P. Dans notre étude, on essaie de comprendre les mécanismes
utilisés par ces deux lysophospholipides qui pourraient exacerber l’inflammation chronique
associée à la PR. Les enzymes qui produisent ces lysophospholipides, ou leurs récepteurs,
pourraient être des cibles potentielles dans les futurs traitements de la PR.

iii
Abstract
Rheumatoid arthritis (RA) is an autoimmune disease affecting approximately 1% of
Canadians. The many treatments available still have major disadvantages: drug resistance
in the long term, and partial or no effect of the drug. Therefore it is necessary to constantly
develop new therapeutic targets. In recent years, lysophosphatidic acid (LPA) and
sphingosine-1-phosphate (S1P) have appeared to play a role in the chronic inflammation
involved in arthritis. In synovial fluid of patients with RA, we find not only LPA but also
some products of its catabolism and the enzyme that produces it. Moreover, TNF-α, present
in inflammation, is capable of amplifying the effects of LPA and S1P. In our study, we try
to understand the mechanisms used by these two lysophospholipids which may exacerbate
the chronic inflammation associated with RA. The enzymes that produce these
lysophospholipids or their receptors could be potential targets for future therapies against
RA.

iv
Avant-Propos
Je tiens à remercier toutes les personnes qui m’ont aidé, tant au niveau personnel, que
professionnel, à affronter les difficultés rencontrées lors de ces deux années de Maîtrise.
Merci à Arpita, Fethia, Michel, Élise, Elisandra, Cristina, Nanni, Hugo, Jean-Michel,
Stéphanie, Louis, Sophie, Oana, Emmanuelle, Marie-Thérèse, Joana, Lamia, Isabelle,
Patricia, Aliona, pour votre présence, votre bonne humeur, et votre soutien tout au long de
ma Maîtrise. Merci aussi pour tous les échanges scientifiques et personnels que vous
m’avez offert.
Merci à Delphine, Amine, François, Lynn, et Danielle pour leur soutien psychologique,
indispensable lorsqu’on arrive dans le milieu de la recherche et que l’on a tendance à se
décourager facilement! Merci aussi pour votre aide continue, votre attention, et votre
amitié.
Je remercie aussi Gervais Morissette, sa femme Jacline, et leurs enfants Nicolas et Nadia,
qui m’ont accueilli dès mon arrivée à Québec, et qui m’ont soutenu jusqu’à maintenant. Ils
m’ont fait découvrir les plaisirs de Québec et du Québec, et la joie d’y vivre et de s’y sentir
chez soi.
Merci à mon amoureux, pour son amour, sa patience, et son soutien.
Merci à ma famille, à mes parents qui ont toujours eu confiance en moi et qui m’ont
soutenu malgré leurs problèmes de santé et leurs propres soucis. Merci d’avoir cru en moi,
et de m’avoir donné les moyens et la force d’arriver jusqu’ici.
Je souhaiterai aussi remercier le Dr Philippe Tessier pour son assistance technique sur les
poches d’air et pour nos discussions intéressantes au dernier congrès du CAN.
Merci au Dr Patrice Poubelle pour ses connaissances et surtout pour ses cours passionnants
qui m’ont marqué par leur qualité! (Même si l’on avait trop de travail!)

v
Je remercie Maria Fernandes pour son encadrement, sa rigueur, son aide, son soutien, son
attention régulière à mon égard, et pour le temps qu’elle m’a consacré tout au long de ma
Maîtrise.
Merci à Sylvain Bourgoin, pour son encadrement, son aide précieuse et ses connaissances,
sa patience, sa générosité et sa gentillesse. Merci de m’avoir permis de travailler avec vous,
avec plaisir, sur ces deux projets passionnants. Merci d’avoir été à l’écoute tout au long de
ma Maîtrise et d’avoir été d’une rapidité et d’une attention exceptionnelle à chaque étape de
ma rédaction. Merci d’avoir fait de ma Maîtrise une grande satisfaction!
Un merci spécial à Chenqi Zhao pour toutes ces connaissances que tu m’as transmises et
aidé à acquérir, et pour tout ton enseignement. Merci de m’avoir aidé à améliorer mon
anglais et de m’avoir appris beaucoup, tant au niveau scientifique que personnel. Merci
pour tes conseils, pour les « lift » à 6h du matin, le chandail bleu que je garde
précieusement, tes autres cadeaux, les « tea break » qui me manquent déjà, et surtout merci
d’avoir été à la fois une « boss/co-équipière » exceptionnelle, une amie, et une mère dans
certains moments.
Merci à Louise pour toute son aide et le temps qu’elle a pris pour répondre à mes questions
tout au long de ma maîtrise. J’ai maintenant compris pourquoi elle m’a été présentée
comme la gardienne du « bureau des miracles ». Merci Louise !
Merci à Sonia pour son assistance technique, sa rigueur et son professionnalisme. Merci
d’avoir pris grand soin de nos souris, sans qui nos expériences n’auraient pas été possibles!
Je remercie les organismes boursiers FRAMR (Fonds de recherche sur l’arthrite et les
maladies rhumatismales) ainsi que le CAN (Canadian Arthritis Network) et la Société de
l’arthrite pour leur soutien financier indispensable. Merci particulièrement au CAN pour
m’avoir permis de rencontrer des patients atteints de PR: des rencontres absolument
exceptionnelles, enrichissantes et inoubliables! Merci aussi à Joanne Wright, pour avoir
organisé ces merveilleuses rencontres ainsi que de nombreux évènements instructifs.

vi
Contribution aux articles
Chapitre 2 :
Le prétraitement au TNF-α augmente le recrutement de leucocytes induit par le LPA in vivo, via les récepteurs LPA1 et LPA3: implication des récepteurs CXCR2 des chimiokines, «TNF-α priming enhances in vivo LPA-mediated recruitment of leukocytes through LPA1 and LPA3: implication of CXCR2 ligand chemokines » Chenqi Zhao, Anne Sardella, Jerold Chun, Patrice E. Poubelle, Maria J. Fernandes, and Sylvain G. Bourgoin. Cet article sera soumis très prochainement pour publication dans Journal of Immunology. Chenqi Zhao et moi-même avons travaillé sur ce projet à parts égales, sous la supervision des Docteurs Maria Fernandes et Sylvain Bourgoin. Jerold Chun nous a fourni les souris LPA3 +/- qui nous ont permis de démarrer la colonie LPA3 K.O. utiles pour nos expériences. J’ai pris soin de suivre cette colonie tout au long de ma Maîtrise et d’effectuer les génotypages et le suivi quotidien de cette colonie, c’est-à-dire gérer les accouplements, contrôler les naissances et l’âge des souris pour produire un minimum de souris lors de chaque planification d’expérience, et cependant pouvoir effectuer nos expériences. Chapitre 3 :
La production d’IL-8 induite par le stress hypoxique est dépendante des récepteurs au S1P dans les synoviocytes de type « fibroblast-like » provenant de patients atteints de polyarthrite rhumatoïde, « Hypoxic-like stress-mediated IL-8 production is S1P receptor independent in rheumatoid fibroblast-like synoviocytes » Chenqi Zhao, Anne Sardella, John A. Di Battista, Patrice E. Poubelle, Maria J. Fernandes and Sylvain G. Bourgoin. Cet article sera certainement complété avant d’être soumis. Ma contribution à cet article est principalement reliée aux dernières expériences effectuées concernant l’expression de la sphingosine kinase 1 sur 4 donneurs atteints de polyarthrite rhumatoïde et 4 donneurs sains (3 expériences par donneur). De plus, en début de Maîtrise, j’ai effectué quelques tests ELISA sur certains donneurs, pour mesurer la quantité d’IL-8 produite suite à une stimulation au stress hypoxique induit par le CoCl2. Chenqi Zhao a réalisé toutes les autres expériences in vitro.

vii
À mes parents
La science, comme l'amour, est aveugle. Voilà pourquoi elle se plaît à procéder par
tâtonnements. [Jean O'Neil]

viii
Table des matières
Résumé................................................................................................................................... ii Abstract................................................................................................................................. iii Avant-Propos .........................................................................................................................iv Contribution aux articles........................................................................................................vi Table des matières .............................................................................................................. viii Liste des tableaux....................................................................................................................x Liste des figures .....................................................................................................................xi Liste des abréviations utilisées ............................................................................................ xii Chapitre 1 : Introduction.........................................................................................................1 1.1. La polyarthrite rhumatoïde (PR).................................................................................1
1.1.1. Généralités ..........................................................................................................1 1.1.1.1. Mécanismes et symptômes .............................................................................1 1.1.1.2. Causes probables.............................................................................................4
1.1.2. Cellules et molécules contribuant à l’inflammation des patients atteints de PR 5 1.1.2.1. Les synoviocytes.............................................................................................5 1.1.2.2. Les leucocytes.................................................................................................6 1.1.2.3. Les cytokines et chimiokines........................................................................11
1.1.3. Traitements .......................................................................................................14 1.2. Les lysophospholipides.............................................................................................18
1.2.1. L’acide lysophosphatidique ..............................................................................20 1.2.1.1. Historique......................................................................................................20 1.2.1.2. Métabolisme du LPA....................................................................................21 1.2.1.3. Les récepteurs au LPA..................................................................................22 1.2.1.4. LPA et liquides biologiques..........................................................................25
1.2.2. Le sphingosine-1-phosphate .............................................................................25 1.2.2.1. Historique......................................................................................................25 1.2.2.2. Métabolisme du S1P .....................................................................................26 1.2.2.3. Les récepteurs au S1P ...................................................................................27
1.3. Objectifs de l’étude...................................................................................................30 Chapitre 2 : TNF-α priming enhances in vivo LPA-mediated recruitment of leukocytes through LPA and LPA : implication of CXCR2 ligand chemokines .................................32 1 3
2.1. Résumé en français ...............................................................................................33 2.2. Abstract.................................................................................................................35 2.3. Introduction...........................................................................................................36 2.4. Materials and Methods..........................................................................................39 2.5. Results...................................................................................................................43 2.6. Discussion.............................................................................................................49 2.7. Figure ....................................................................................................................55 2.8. Acknowledgments ................................................................................................69 2.9. Footnotes...............................................................................................................70
Chapitre 3 : Hypoxic-like stress-mediated IL-8 production is S1P receptor independent in rheumatoid fibroblast-like synoviocytes...............................................................................72
3.1. Résumé en français ...............................................................................................73 3.2. Abstract.................................................................................................................74

ix
3.3. Introduction...........................................................................................................75 3.4. Materials and methods ..........................................................................................77 3.5. Results...................................................................................................................80 3.6. Discussion.............................................................................................................83 3.7. Funding .................................................................................................................85 3.8. Acknowledgements...............................................................................................86 3.9. Disclosure statement .............................................................................................87 3.10. Figures ..................................................................................................................88
Chapitre 4 : Interprétation des résultats ................................................................................92 4.1. Discussion.............................................................................................................92 4.2. Perspectives ..........................................................................................................99 4.3. Conclusions.........................................................................................................100
Bibliographie ......................................................................................................................101 Annexes ..............................................................................................................................116
Annexe 1. Répartition des gènes HLA sur le chromosome 6 humain............................116 Annexe 2. Expression des HLA à la surface de la membrane cellulaire ........................117 Annexe 3. Hématopoïèse ................................................................................................118 Annexe 4. Récepteurs aux chimiokines chez les leucocytes humains et leurs ligands spécifiques. .....................................................................................................................119 Annexe 5. Schéma expérimental du modèle de la poche d’air. ......................................121

x
Liste des tableaux
Tableau 1. Agents peptidiques sécrétés par le neutrophile et leurs effets. Tableau 2. Nomenclature des récepteurs aux lysophospholipides LPA et S1P.

xi
Liste des figures
Figure 1. Schéma d’une articulation normale et d’une articulation endommagée par la PR. Figure 2. Photographie de mains déformées par la PR. Figure 3. Radiographie comparative d’une main normale et d’une main d’un patient atteint de PR. Figure 4. Différenciation des précurseurs en cellules immunitaires matures, grâce aux facteurs de croissance. Figure 5. Migration transendothéliale des neutrophiles. Figure 6. Résumé des traitements existant dans la PR et leur(s) cible(s) thérapeutique(s). Figure 7. Schéma structural des lysophospholipides bioactifs LPA et S1P. Figure 8. Principales voies de production du LPA. Figure 9. Signalisation du LPA via ses récepteurs LPA1, LPA2 et LPA3, couplés aux protéines G. Figure 10. Production et dégradation du S1P. Figure 11. Signalisation du S1P via ses récepteurs S1P1, S1P2 et S1P3, couplés aux protéines G. Figure 12. Effect of LPA on leukocyte recruitment into the air pouch. Figure 13. Involvement of LPA receptor(s) in LPA-induced leukocyte recruitment into the air pouch. Figure 14. Effect of TNF-α pre-treatment on LPA- and OMPT-induced leukocyte recruitment into the air pouch. Figure 15. Effect of CXCR2 antagonist on LPA-mediated leukocyte recruitment. Figure 16. Effect of LPA and TNF-α pretreatment on the secretion of cytokines/chemokines in air pouches. Figure 17. Effect of LPA on KC secretion in the air pouch. Figure 18. Effect of LPA and OMPT on the secretion of cytokines/chemokines in the air pouch. Figure 19. CoCl2 and S1P, either alone or in combination, stimulates IL-8 secretion in normal and RA FLS. Figure 20. Differential involvement of S1P receptors in CoCl2-mediated IL-8 secretion by normal and RA FLS. Figure 21. CoCl2 stimulates Sphingosine kinase 1 mRNA expression in normal but not in RA FLS.

xii
Liste des abréviations utilisées
Sigle Signification en anglais Équivalent en français AC Adenylyl Cyclase Adénylyl Cyclase ATX Autotaxin Autotaxine COX Cyclo-oxygenase Cyclo-oxygénase Edg Endothelial differentiation gene « Endothelial differentiation gene » FLS Fibroblast-like synoviocytes Synoviocytes de type B G-CSF Granulocyte Colony-Stimulating Factor Facteur de croissance granulocytaire GM-CSF Granulocyte Macrophage Colony-
Stimulating Factor Facteur de croissance des granulocytes et des macrophages
GPR G protein-coupled receptor Récepteurs couplés aux protéines G HIF Hypoxia-inducible factor Facteur d’induction de l’hypoxie IFN Interferon Interféron IL Interleukin Interleukine IL1-ra IL1-receptor antagonist Antagoniste des récepteurs à l’IL-1 IP-10 Interferon-gamma inducible protein-10
(10kDa) Protéine inductible par l’interféron gamma
KC Keratinocyte-derived chemokines Chimiokines dérivées des kératinocytes LPA Lysophosphatidic acid Acide lysophosphatidique LPP Lipid Phosphate Phosphohydrolases « Lipid Phosphate Phosphohydrolases » LysoPLD Lysophospholipase D Lysophospholipase D MAPK Mitogen-Activated Protein Kinase Kinases activées par les agents mitogènes MCP-1 Monocyte-chemoattractant protein-1 Protéine chimioattractant les monocytes M-CSF Macrophage Colony Stimulating Factor Facteur de croissance des macrophages MIP Macrophage-inflammatory protein « Macrophage-inflammatory protein » MMP Matrix-metalloproteinases Métalloprotéinases matricielles NK Natural killer « Natural killer » PGE Prostaglandin Prostaglandine E PAP Phosphatidic Acid Posphatases Acide phosphatidique phosphatases PI3K Phosphoinositide 3-kinase Phosphoinositide 3-kinase PKB/Akt Protein kinase B/Akt Protéine kinase B/Akt PLA Phospholipase A Phospholipase A PLC Phospholipase C Phospholipase C RA/PR Rheumatoid arthritis Polyarthrite rhumatoïde S1P Sphingosin-1-phosphate Sphingosine-1-phosphate SCF Stem cell factor Facteur des cellules souches SphK Sphingosin kinase Sphingosine kinase TGF Transforming growth factor Facteur de croissance transformant TNF-α Tumor necrosis factor α Facteur de nécrose tumorale

1
Chapitre 1 : Introduction
1.1. La polyarthrite rhumatoïde (PR) La polyarthrite rhumatoïde (PR) est une maladie auto-immune chronique, à tropisme
synovial, touchant près de 1% de la population mondiale. Il s’agit d’une maladie
hétérogène très invalidante, entrainant parfois une destruction totale des articulations, et par
voie de conséquence, des pertes de fonction des membres. Il n’existe aujourd’hui aucun
traitement curatif pour guérir cette maladie. Les causes de cette maladie douloureuse et
destructrice ne sont que partiellement connues. C’est pourquoi, malgré les nombreux
médicaments contribuant à diminuer les douleurs liées à la maladie, il est difficile de
proposer un traitement adéquat et d’obtenir une rémission complète. Dans le but d’en
apprendre d’avantage sur cette mystérieuse maladie et de proposer éventuellement l’espoir
d’un futur traitement plus efficace, nous résumerons dans un premier temps les
connaissances acquises jusqu’à maintenant à ce sujet. Ensuite, nous développerons sur les
deux principaux lysophospholipides LPA et S1P et leurs implications potentielles dans la
polyarthrite rhumatoïde, puisque ces deux molécules semblent avoir un rôle important dans
cette maladie. Enfin, nous exposerons les objectifs et les résultats de nos études portant
respectivement sur le LPA et sur le S1P.
1.1.1. Généralités
1.1.1.1. Mécanismes et symptômes
Un élément déclencheur inconnu, qui pourrait potentiellement être une bactérie selon
certaines hypothèses [1], va stimuler l’activation des lymphocytes T [2], ce qui déclenchera
par conséquent une réaction inflammatoire composée de nombreuses cellules (incluant les
leucocytes et les synoviocytes) et de nombreuses molécules majoritairement pro-
inflammatoires (dont les cytokines). Les cellules inflammatoires, une fois activées,
sécrètent donc des enzymes protéolytiques qui détruisent la matrice cartilagineuse, et
stimulent la prolifération des synoviocytes qui envahissent le cartilage (figure 1). Si
l’équilibre entre les cytokines pro-inflammatoires et anti-inflammatoires n’est pas rétablie,
l’inflammation persiste et devient chronique. C’est ce qui caractérise la PR. La réaction
inflammatoire sera maintenue par l’activation continue des cellules [3, 4] et conduira à

2
l’érosion progressive du cartilage et des os [5]. Plus l’inflammation sera agressive et
chronique, plus les symptômes deviendront visibles physiquement (figures 2 et 3). Le
système immunitaire se met ainsi à agresser notre organisme, qu’il est censé défendre.
Cette maladie s’attaque particulièrement aux articulations. Toutes les articulations peuvent
être touchées et, la plupart du temps, ce phénomène apparaît de manière symétrique [6].
Pour la majorité des malades, la PR affecte d'abord les mains, les poignets et les
articulations des pieds. Avec le temps, beaucoup d’autres articulations peuvent être
touchées.
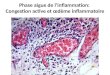
A.
B.
Figure 1. Schéma d’une articulation normale et d’une articulation endommagée par la PR. La membrane synoviale est une paroi très fine, et le cartilage ainsi que les os sont intacts chez un patient normal (A). Chez un patient atteint de PR (B), on observe une hyperplasie de la membrane synoviale (« pannus rhumatoïde ») due à la prolifération des synoviocytes et à l’infiltration de cellules immunitaires (adapté de « Mayo Foundation for Medical Education and Research »).

3
Figure 2. Photographie de mains déformées par la PR. Cette photographie montre des mains déformées par la PR, à un stade avancé de la maladie. En effet, l’inflammation chronique des articulations entraîne l’érosion progressive du cartilage, puis des os. L’inflammation non contrôlée par un traitement adéquat affecte gravement la condition de vie en rendant les articulations moins fonctionnelles, voire non fonctionnelles (www.arthritis.about.com).
Figure 3. Radiographie comparative d’une main normale et d’une main d’un patient atteint de PR. La radiographie de la main du patient non atteint de PR révèle des articulations saines (A) tandis que la radiographie de la main d’un patient atteint de PR (B) révèle une érosion osseuse due à l’inflammation chronique au niveau des articulations (www.health.com).

4
1.1.1.2. Causes probables
Les causes précises déclenchant cette maladie restent inconnues. Cependant, il a été
démontré que certains facteurs génétiques tels que les gènes du système HLA (développé
ci-dessous) [7, 8], environnementaux (ex: infections bactériennes) [9, 10], ou encore
hormonaux [11, 12] pouvaient participer au déclenchement de cette maladie. En effet, la PR
se manifeste de manière plus violente et plus fréquente chez la femme. Ceci serait expliqué
en partie par les effets de plusieurs hormones, principalement les œstrogènes [11, 13].
Différents gènes peuvent intervenir dans la susceptibilité à la PR: gènes codant pour
les chaînes α ou β du récepteur des lymphocytes T [14], gènes des immunoglobulines [15],
gènes du système de l’apoptose (système Fas-ligand) [16], séquences régulatrices du TNF-
α [17] et surtout, les gènes du système HLA (Human Leukocyte Antigen) de classe II.
L’annexe 1 montre la répartition des gènes HLA sur le chromosome 6 humain et l’annexe
2 montre l’expression des HLA, à la surface de la membrane cellulaire. HLA-DR, HLA-DP
ou HLA-DQ de l’humain correspondent au système HLA de classe II. Les prédispositions
génétiques des patients à développer une PR semblent être multiples. Les HLA de classe II
auraient un rôle important à jouer dans le développement de cette pathologie. Le génotype
HLA-DRB1 pourrait en effet influencer le risque et la sévérité de la PR [18]. Le locus HLA
contribue pour environ 30% au risque familial global [19], ce qui nous indique que les
allèles de susceptibilité HLA-DRB1 ne semblent pas être indispensables au développement
de la maladie, mais occupent tout de même une place importante. De plus on retrouve
d’autres locus impliqués dans cette maladie. Cia5 par exemple, est un locus qui régule la
sévérité de la PR induite au collagène ou au pristane [20]. Cia5d, qui constitue une portion
du locus Cia5, va contribuer à réguler l’invasion des FLS [21, 22]. Ces différents locus
impliqués dans la PR confirment donc qu’il existe des prédispositions génétiques liées à la
maladie et montrent le caractère multigénique de cette pathologie.

5
1.1.2. Cellules et molécules contribuant à l’inflammation des patients atteints de PR La cause exacte du déclenchement de l’auto-immunité reste donc encore inconnue à ce
jour. Cependant, de nombreuses cellules participent à l’inflammation chronique associée à
l’arthrite. Dans cette partie, nous présenterons les principales cellules mise en jeu dans la
PR, en s’attardant plus particulièrement sur les synoviocytes et les neutrophiles, deux types
cellulaires importants dans nos travaux de recherche.
1.1.2.1. Les synoviocytes
Les synoviocytes sont les cellules qui tapissent la membrane synoviale. Il en existe deux
types distincts : les synoviocytes de type A ou « macrophage-like synoviocytes » et les
synoviocytes de type B ou « fibroblast-like synoviocytes » (FLS). Ces deux types
cellulaires ont des caractéristiques différentes au niveau morphologique, histochimique et
immunohistologique [23]. Les cellules de type A ont les caractéristiques morphologiques et
phénotypiques d'un macrophage, elles sont donc capables de phagocytose. Les cellules du
type B sont des cellules sécrétrices de nombreuses molécules (incluant des composantes du
liquide synovial) et elles ne sont pas phagocytaires. Seuls les synoviocytes du type B ont la
capacité de croître in vitro. Après trois passages, les cultures cellulaires de synoviocytes
sont donc presque entièrement du type B [24]. De plus, les FLS ont un potentiel prolifératif
bien plus important que les synoviocytes de type A [25].
Les synoviocytes sont connus pour être les cellules les plus actives dans la pathologie de la
PR. La prolifération des FLS dans l’articulation est une des caractéristiques qui définit cette
maladie. Les synoviocytes de type B ont été suggérés comme étant une cible unique pour le
traitement de l’arthrite, à cause de leur rôle central dans la maladie [26, 27]. En effet, ils
sont capables d’interagir avec la majorité des cellules qui contribuent à la destruction du
cartilage et à l’inflammation tels que les lymphocytes B [28] ; les lymphocytes T [29, 30],
mais aussi les monocytes/macrophages [31] et les neutrophiles [32, 33]. Les FLS
arthritiques possèdent des capacités uniques à favoriser la migration des lymphocytes T
CD4+ et CD8+ dans le tissu synovial [34]. Ils interagissent également avec les
neutrophiles, de manière directe et indirecte, pour entrainer la prolifération des cellules
endothéliales vasculaires qui, elles-mêmes, contribuent à la néo-angiogenèse, c’est-à-dire à

6
la formation de nouveaux vaisseaux, produisant ainsi des voies de migration cellulaires
supplémentaires pour la dispersion et la dissémination des cellules immunitaires au niveau
du site inflammatoire [35]. De plus, nous avons démontré que les synoviocytes de type B
étaient capables, in vitro, de produire de l’interleukine-8 (IL-8) qui est un fort chimio-
attractant pour les neutrophiles [36]. Ils sécrètent également de nombreuses autres
cytokines telles que le « monocyte-chemoattractant protein-1 » (MCP-1) [37],
l’interleukine-6 (IL-6) [38] et l’IL-8 qui vont amplifier les phénomènes inflammatoires et la
destruction du cartilage. Filer et al. soulignent que les fibroblastes jouent un rôle important
dans la régulation de la survie des différentes sous-populations de leucocytes dans les tissus
[33]. Hashida et al. suggèrent également que les cellules synoviales sont stimulées
indirectement par les neutrophiles, via la production de collagénase [39]. De cette manière,
les synoviocytes contribuent activement à l’inflammation chronique et à la dégradation du
cartilage chez les patients atteints de PR.
1.1.2.2. Les leucocytes
Les leucocytes sont les globules blancs du sang qui aident aux défenses immunitaires. Ils
sont formés et différenciés dans la moelle osseuse (dans le thymus pour les lymphocytes T),
au même titre que les plaquettes et les hématies lors de l’hématopoïèse (Annexe 3). La
formation des lymphocytes est appelée lymphopoïèse. Toutes les cellules sanguines sont
donc formées à partir d’une seule et même cellule souche, qui va ensuite se diviser et subir
des modifications qui vont entraîner la variété des types cellulaires qu’on peut retrouver
dans le sang. D’une manière générale, les leucocytes sont impliqués dans tous les
phénomènes inflammatoires. L’inflammation chronique, en particulier, est caractérisée par
l’accumulation massive de leucocytes infiltrés dans les tissus [40]. Chez une personne
normale, la majorité des leucocytes circulants (minimum 70%) est composée de
polynucléaires neutrophiles qui sont des granulocytes. Le reste est constitué de monocytes
(différenciés en macrophages dans les tissus), de lymphocytes, et quelques autres
granulocytes (basophiles, éosinophiles) en faible quantité la plupart du temps.
Les polynucléaires neutrophiles sont une sous-population de leucocytes sanguins constitués
puis différenciés dans la moelle osseuse grâce à des facteurs de différenciation qui leur sont
spécifiques (figure 4).

7
SANG
MOELLE OSSEUSE
Lignée lymphoïde
Lignée myéloïde
SANG
Figure 4. Différenciation des précurseurs en cellules immunitaires matures, grâce aux facteurs de croissance. GM-CSF, Granulocyte Macrophage Colony-Stimulating Factor; G-CSF, Granulocyte colony-stimulating factor; IL, Interleukin; M-CSF, Macrophage Colony Stimulating Factor; SCF, Stem cell factor (Adapté de Dr. Janet Fitzakerley, 2009).
Les neutrophiles en circulation meurent en un à deux jours en l’absence d’inflammation. En
revanche, lors d’une inflammation, leur durée de vie est prolongée par des cytokines ou
facteurs de croissance tel que le TNF-α [41] qui inhibent leur apoptose dans le but de leur
laisser un peu plus de temps pour se débarrasser des éléments étrangers à éliminer.
Cependant, il arrive que leur durée de vie soit anormalement trop prolongée, et qu’il y ait

8
par conséquent, une augmentation des dommages causés sur les tissus: l’inflammation
chronique est alors enclenchée. Ces cellules, très importantes dans les défenses
immunitaires sont les premières à migrer au lieu de l’inflammation [42, 43]. En effet, des
cytokines chimioattractantes produites au niveau du site inflammatoire guident les
neutrophiles du sang vers les tissus endommagés : c’est la migration transendothéliale des
neutrophiles (figure 5). Une fois rendu au site inflammatoire, le neutrophile pourra
orchestrer la suite de la réponse immunitaire, influencer les cellules à proximité de
l’inflammation et amener d'autres leucocytes à migrer au site grâce à la production d’une
panoplie de médiateurs peptidiques et lipidiques (tableau 1). En plus de leur rôle central
dans le recrutement d’autres cellules, ces granulocytes ont la capacité de phagocyter les
bactéries dans le but de les détruire, et contribuent ainsi, aussi bien directement
qu’indirectement, aux défenses de l’organisme.

9
1. 2. 3.
Figure 5. Migration transendothéliale des neutrophiles. 1. Rolling. Dans un premier temps, le neutrophile circulant et l’endothélium vont interagir via des molécules d’adhésion appelées sélectines, qui sont les premières étapes de l’adhésion du neutrophile aux cellules endothéliales et qui ne constituent pas une adhésion ferme, mais seulement une adhésion labile dans le but de retenir le neutrophile contre l’endothélium. 2. Polarisation - adhérence ferme. Dans un second temps, une deuxième catégorie de molécules d’adhésion vont intervenir pour renforcer l’accrochage du neutrophile aux cellules endothéliales : il s’agit des intégrines. Le cytosquelette d’actine est localement réorganisé, ce qui entraîne la polarisation morphologique du neutrophile le long de l’endothélium. 3. Transmigration. Le neutrophile va ensuite se faufiler à travers la couche de cellules endothéliales : c’est la migration transendothéliale ou la diapédèse. Ce phénomène de transmigration est favorisé par les facteurs chimiotactiques, par la vasodilatation et par le ralentissement du flux sanguin au voisinage des foyers inflammatoires. L'afflux de neutrophiles y est important et rapide [44].

10
Tableau 1. Agents peptidiques sécrétés par le neutrophile et leurs effets. Résumé des différentes cytokines et chimiokines produites par le neutrophile lors d’une réponse inflammatoire et descriptions de leurs effets sur les cellules immunitaires. IL, interleukine; IP-10, IFN-γ-inductible protein of 10 kDa; MIP, macrophage-inflammatory protein; NK, natural killer; TNF, tumor necrosis factor ; IL-1Ra, IL-1 receptor antagonist ; TGF, transforming growth factor ; GM-CSF, granulocyte macrophage colony-stimulating factor ; IFN, interféron. [45]. Dans la physiopathologie de la PR, les neutrophiles sont les cellules qui contribuent à la
chronicité de l’inflammation. En effet, chez les patients atteints de cette maladie, les
neutrophiles représentent 90% des cellules trouvées dans le liquide synovial. L’importance
et le rôle prépondérant des neutrophiles dans la PR a été reconnu depuis de nombreuses
années [46]. En effet, les neutrophiles ont été plusieurs fois impliqués dans la dégradation
du cartilage dans la PR [47, 48]. D’ailleurs, on les retrouve en grande quantité au niveau du
pannus rhumatoïde [49]. Ils contribueraient aussi au réseau de cytokines inflammatoires
associées à la PR en exprimant notamment l’interleukine-1 (IL-1) qui a un rôle majeur dans
cette maladie [50]. De plus, il a été démontré que la survie des neutrophiles était
significativement prolongée en présence de liquide inflammatoire synovial de patients
atteints de PR [51], ce qui suggère que les neutrophiles participent à l’inflammation

11
chronique de la maladie. Le neutrophile rhumatoïde est également capable d’augmenter la
destruction osseuse articulaire en activant directement les ostéoclastes [52]. Le neutrophile
est donc une cellule centrale dans la pathologie de la PR, car il contribue non seulement à
l’inflammation chronique, mais aussi à la dégradation du cartilage, qui sont deux éléments
caractéristiques de la maladie.
D’autres cellules ont un rôle important à jouer dans le développement de la PR. Le
lymphocyte B par exemple, est responsable de la production du facteur rhumatoïde (FR) et
d’un anticorps anti-peptide citrique citrulliné ou anti-CCP, qui constituent des marqueurs
déterminants pour le diagnostic de la maladie [53]. Le lymphocyte T interagit également
avec les cellules dendritiques pour contribuer à l’inflammation en régulant le recrutement
des cellules immunitaires via certaines chimiokines [54]. On doit aussi considérer le rôle
non négligeable des ostéoblastes et des ostéoclastes qui régulent le remodelage osseux et
qui sont responsables de l’érosion osseuse observée dans cette maladie articulaire [55].
Malgré l’importance de ces autres types cellulaires, nous ne développerons pas d’avantage
leurs rôles dans la PR. En effet, nous avons choisi pour notre première étude in vivo de
nous focaliser sur les leucocytes et plus particulièrement les neutrophiles, qui constituent,
rappelons-le, 90% des cellules contenues dans le liquide synovial de patients atteints de PR.
En ce qui concerne notre seconde étude in vitro, nous travaillerons sur des synoviocytes
provenant soit de patients non malades, soit de patients atteints de PR. La justification
détaillée de l’étude sera développée plus loin.
On vient donc de passer en revue une grande partie des cellules impliquées dans la
physiopathologie de la PR. Mais comment la plupart de ces cellules parviennent-elles au
lieu de l’inflammation et comment cette inflammation va-t-elle être maintenue ?
1.1.2.3. Les cytokines et chimiokines
Les cellules communiquent entre elles grâce à de nombreuses molécules dont les plus
connues sont les hormones, ou encore les neurotransmetteurs. Les cytokines font partie de
ces molécules indispensables à la communication cellulaire. Il s’agit de protéines solubles
de 8 à 50 kDa synthétisées par les cellules et agissant sur d'autres cellules pour en réguler
l'activité et la fonction. Leur action, par l’intermédiaire de récepteurs spécifiques situés sur

12
les cellules, peut être paracrine (sur les cellules environnantes), endocrine (sur les cellules
éloignées) ou autocrine (sur la cellule productrice ou une cellule environnante de même
nature). Toutes les cellules nucléées sont capables de produire des cytokines mais certaines
d’entre elles sont majoritairement sécrétées par les cellules immunitaires pour réguler
l’inflammation. Une réaction immunitaire normale est régulée par les cytokines pro- et anti-
inflammatoires libérées par les cellules. La perte d’équilibre entre les cytokines pro- et anti-
inflammatoires est associée à la physiopathologie de la PR. L’annexe 4 regroupe toutes les
chimiokines et leurs récepteurs chez le leucocyte humain.
Toutes ces cytokines sécrétées par différentes cellules vont donc participer activement à
l’inflammation. Parmi ces cytokines/chimiokines, l’IL-8 est importante car il s’agit d’un
fort chimio-attractant pour les neutrophiles qui contribuent à l’inflammation chronique
associée à la PR. Le rôle de l’IL-8 sur l’activation des neutrophiles a en effet été démontré
dans la PR spécifiquement [56]. L’équivalent de cette chimiokine chez la souris est KC ou
MIP-2, deux chimiokines capables de se fixer sur les récepteurs de l’IL-8 : CXCR2 [43,
57]. Cette notion est importante pour notre projet in vivo sur les souris.
Le TNF-α, quant à lui, est une molécule « clé » dans l’inflammation, et par conséquent dans
la PR. En effet, le TNF-α est une cytokine importante impliquée dans de nombreux cas
pathologiques tels que les cancers, l’auto-immunité, et l’inflammation chronique. On
observe que cette cytokine a de nombreux effets sur une multitude de cellules impliquées
dans la physiopathologie de la PR. Par exemple, celle-ci est capable de stimuler la
prolifération des FLS [58, 59]. Sur le même type cellulaire, nos travaux ont démontré que
le TNF-α augmentait considérablement la production d’IL-8 induite par le LPA [60].
D’ailleurs, le TNF-α a des effets anti-apoptotiques sur ces mêmes cellules [61], et sur les
neutrophiles [51], ce qui signifie qu’il va avoir un rôle de maintien de l’inflammation. En
augmentant l’adhésion des neutrophiles au cartilage, et en favorisant ainsi sa dégradation, le
TNF-α contribue à l’aggravation de la PR [62]. De plus, une équipe de recherche a
démontré que le TNF-α induisait un signal chez le neutrophile qui pourrait promouvoir leur
rétention au site inflammatoire, et ainsi, prolonger l’inflammation pour provoquer une
inflammation chronique [63]. Le TNF-α, en agissant sur le neutrophile, contribue

13
également à la prolifération des lymphocytes B responsables de l’auto-immunité dans la PR
[64].
Parmi ses nombreuses propriétés, le TNF-α a également la capacité d’aider les cellules à
détruire le cartilage. En effet, après une stimulation au TNF-α, les chondrocytes et les
fibroblastes synoviaux libèrent des métalloprotéinases matricielles (MMPs) [65], en
particulier la stromélysine et les collagénases chez des FLS provenant du pannus
rhumatoïde [66]. Ces enzymes dégradent la matrice du tissu conjonctif et sont connues pour
être des médiateurs importants de la destruction des articulations dans la pathologie de la
PR.
Le TNF-α a aussi le pouvoir d’induire la production de cytokine en agissant sur les cellules.
Par exemple, il est capable d’induire la production d’IL-8 et d’IL-6 chez des FLS humains
[67, 68], deux cytokines ayant un rôle important dans la pathologie de la PR. Il induit aussi
la production d’autres cytokines ou facteurs de croissance impliqués dans la pathologie de
la PR, tels que l’IL-1 [69] et le GM-CSF [70].
Non seulement le TNF-α est capable de stimuler l’expression des cytokines dans les
cellules, mais il est aussi connu pour avoir de nombreux effets synergiques avec des
molécules bioactives telles que l’IL-17 [71], l’IL-1 [72], ou encore le S1P [60], et le LPA
[60]. En synergisant avec les molécules présentes dans le milieu inflammatoire de la PR, le
TNF-α aura donc un rôle central et primordial qui lui permettra d’augmenter les effets de
ces molécules.
Récemment, le TNF-α a été proposé comme cible thérapeutique dans le traitement et la
prévention des cancers [73] et depuis quelques années, plusieurs inhibiteurs du TNF-α sont
utilisés comme traitements efficaces de la PR [74]. Cette découverte a d’ailleurs été
révolutionnaire pour les patients résistants aux traitements classiques de la polyarthrite
rhumatoïde, tels que le méthotrexate ou les DMARDs (Disease-modifying antirheumatic
drugs).

14
1.1.3. Traitements Les médicaments existants à ce jour pour les patients atteints de PR ne visent pas tous les
mêmes objectifs. On pourrait les classer deux catégories :
- les traitements symptomatiques ou de 1ère ligne tels que les antalgiques, les
corticostéroïdes à faible dose, ou les anti-inflammatoires non stéroïdiens ou AINSs, qui
réduisent la douleur et la raideur aux articulations. Ils ne freinent pas l’évolution de la
maladie et ne préviennent pas la survenue d’éventuelles déformations articulaires. Ils sont
surtout utilisés au début de la maladie, puis par intermittence.
- les traitements de fonds ou de 2ème ligne (=agents de rémission), appelés aussi
DMARDs, tels que le méthotrexate (le plus couramment utilisé en première intention),
l'hydroxychloroquine ou la sulfasalazine qui visent à prévenir ou retarder les dommages
articulaires et à freiner l’évolution de la maladie.
La figure 6 est un résumé des traitements les plus utilisés dans la PR, qui montre l’endroit
précis où ils agissent. Leur efficacité est en général inconstante et temporaire (phénomène
d’échappement), obligeant à changer de molécule. Ils doivent donc être constamment
adaptés à l'activité inflammatoire, à la réponse du patient, et à la sévérité de la maladie.

15
Figure 6. Résumé des traitements existant dans la PR et leur(s) cible(s) thérapeutique(s) [75].

16
Depuis quelques années, une nouvelle catégorie de médicaments de deuxième ligne appelés
« biologiques » est apparue sur le marché. Contrairement aux autres médicaments
antirhumatismaux, qui combattent l’action du système immunitaire de façon non
spécifique, ces nouvelles thérapies sont conçues pour cibler plus précisément les substances
qu’on croit directement responsables de l’inflammation et de la destruction articulaire. Les
premiers représentants de cette classe de médicaments sont les agents anti-TNF-α. Les
inhibiteurs du TNF se lient au TNF-α, l’empêchant ainsi de se fixer sur ses récepteurs
spécifiques et d’agir sur les cellules. Ceci interfère donc avec la réaction inflammatoire en
diminuant l’inflammation subséquente aux effets du TNF-α. En novembre 1998,
ENBREL® (etanercept) devient le premier inhibiteur du TNF approuvé par la FDA (Food
and drug administration). Une année plus tard, le REMICADE® (infliximab) arrive sur le
marché, et en 2002, HUMIRA® (adalimumab) est commercialisé.
De nombreux patients ont parlé de « traitement miracle » à la suite de l’utilisation de cette
drogue. Ces nouveaux médicaments sont vraiment apparus comme une avancée
révolutionnaire dans le traitement de la PR. Et pourtant, les anti-TNF-α ne sont pas
efficaces sur 100% des patients. En effet, environ 1 patient sur 3 va répondre à ce
traitement, tandis que les autres ne répondront pas, ou partiellement. De plus, plusieurs
études démontrent qu’il y aurait un risque de développement de cancers et une
augmentation du nombre d’infections chez les patients traités aux anti-TNF-α [76, 77].
C’est pourquoi, dans un premier temps, on a cherché à trouver des traitements
complémentaires aux anti-TNF-α. Des chercheurs ont par exemple suggérés de combiner
certains « anciens traitements », comme le méthotrexate, avec les anti-TNF-α pour
améliorer la réponse des patients [78]. Mais les inconvénients que présentent les anti-TNF-
α chez les 2/3 des patients atteints de PR nous ont finalement poussé à proposer de
nouvelles cibles thérapeutiques pour traiter la PR.
Malgré tous ces traitements, dont l’efficacité dépend énormément de la réponse des
patients, et malgré aussi les nouvelles avancées thérapeutiques des 10 dernières années, les
chercheurs continuent d’utiliser toutes leurs connaissances pour proposer un choix encore
plus vaste de traitements de l’arthrite rhumatoïde et éventuellement trouver un moyen de
supprimer définitivement l’apparition de la maladie. Certaines recherches ont récemment

17
démontré qu’un traitement précoce au cours des premiers mois de la maladie augmentait les
chances de rémission prolongée [79, 80].
Dans l’optique de pouvoir nous aussi proposer de nouveaux traitements, alternatifs ou
complémentaires aux traitements déjà existants, nous avons choisi d’étudier les
lysophospholipides, qui semblent être, de plus en plus au cours des années, une cible
potentielle pour le traitement de la PR.

18
1.2. Les lysophospholipides Des phospholipides circulants ne portant qu’une seule chaîne d’acide gras, appelés
lysophospholipides, et connus initialement comme constituants de la membrane plasmique,
sont aujourd’hui considérés comme des médiateurs de la communication intercellulaire.
Leurs principaux représentants sont l’acide lysophosphatidique (LPA) et la sphingosine-1-
phosphate (S1P), deux molécules biologiquement assimilables aux facteurs de croissance et
cytokines [81].
Malgré des voies de biosynthèses différentes, le LPA et le S1P sont très proches sur le plan
structural (figure 7). C’est pourquoi on observe souvent dans la littérature que ces derniers
sont capables d’induire de nombreux effets similaires sur les cellules. Ils sont tous deux
contenus dans la membrane plasmique des cellules et sont libérés par les plaquettes
activées, qui constituent une importante source de ces lysophospholipides [82]. De plus,
leurs rôles de lipides « bioactifs » ont récemment émergé dans la littérature. En effet, on a
reconnu qu’ils étaient impliqués dans l’angiogénèse [83, 84], l’apoptose [85] et le
développement des tumeurs [86]. D’ailleurs le LPA et le S1P sont la cible de nombreux
futurs traitements contre la progression des tumeurs [87].
Figure 7. Schéma structural des lysophospholipides bioactifs LPA et S1P (Dr Jean-Marie BOTTO).

19
Une famille de récepteurs couplés aux protéines G a été identifiée comme récepteurs
spécifiques pour ces deux lysophospholipides. Initialement classés en récepteurs Edg
(Endothelial differenciation gene), ces récepteurs aux lysophospholipides ont récemment
été subdivisés en récepteurs spécifiques au LPA et au S1P, les deux principaux
phospholipides concernés (tableau 2) [88].
Tableau 2
Récepteurs aux lysophospholipides
Ligand IUPHAR
Nomenclature Nom Edg Autres appellations
LPA LPA1 Edg-2 IpA1, vzg-1
LPA LPA2 Edg-4 IpA2
LPA LPA3 Edg-7 IpA3
LPA LPA4 non-Edg GPR23, P2Y9
LPA LPA5 non-Edg GPR92,GPR93
LPA LPA6 non-Edg P2Y5
S1P S1P1 Edg-1 IpB1
S1P S1P2 Edg-5 IpB2, AGR16, H218
S1P S1P3 Edg-3 IpB3
S1P S1P4 Edg-6 IpB4
S1P S1P5 Edg-8 IpB5, nrg-1
Tableau 2. Nomenclature des récepteurs aux lysophospholipides LPA et S1P. IUPHAR: international union of pharmacology ; LPA : acide lysophosphatidique ; S1P : sphingosine-1-phosphate ; vzg : ventricular zone gene ; Edg : endothelial differentiation gene ; ngr : nerve growth factor regulated G protein-coupled receptor [88]. GPR : G protein-coupled receptor [89]. La principale caractéristique de la PR est d’être une maladie inflammatoire chronique, et
qu’il s’agisse de l’angiogenèse, de la prolifération cellulaire, ou de l’apoptose, tous ces
phénomènes également déclenchés par les lysophospholipides sont clairement reliés à
l’inflammation. C’est pourquoi ces deux lysophospholipides sont au centre de nos
recherches et que nous développerons, dans les prochains paragraphes, leurs
caractéristiques spécifiques.

20
1.2.1. L’acide lysophosphatidique
1.2.1.1. Historique
L’acide lysophosphatidique (LPA) est un phospholipide initialement connu pour être un
intermédiaire métabolique de la synthèse des glycérolipides (ex : triglycérides) et des
phospholipides membranaires mais qui apparait actuellement dans la littérature comme un
puissant médiateur biologique : on parle de LPA « bioactif ». Sa formule chimique la plus
couramment retrouvée dans les liquides biologiques et la plus utilisée dans les études
expérimentales est représentée sur la figure 7 vue précédemment.
Les premières propriétés « bioactives » du LPA ont été reconnues dans les années 80. Le
LPA était capable de stimuler le muscle lisse [90], de provoquer l’agrégation des plaquettes
sanguines [91], et de stimuler la prolifération des fibroblastes [92]. Le LPA a donc très vite
été considéré comme une molécule ayant des propriétés semblables aux facteurs de
croissances et aux hormones [93], ce qui s’est par la suite confirmé de manière
exponentielle jusqu’à maintenant. En effet, le LPA active non seulement la prolifération des
cellules, mais aussi leur migration [94], leur survie et l’apoptose [95].

21
1.2.1.2. Métabolisme du LPA
Le LPA peut être produit par plusieurs enzymes dont les principales sont les
phospholipases A (PLA1 et sPLA2) et l’autotaxine/lysophospholipase D (ATX/lysoPLD).
Pour produire du LPA, les phospholipases A vont cliver une des deux chaînes d’acide gras
de l’acide phosphatidique (figure 8). L’autotaxine en revanche, agit sur la
lysophosphatidylcholine (LPC) en libérant la choline, produisant ainsi le LPA (figure 8).
Bien que l’autotaxine puisse cliver tous les lysophospholipides, la LPC est le phospholipide
le plus abondant retrouvé dans les liquides biologiques incluant le plasma et le liquide
synovial [96].
Figure 8. Principales voies de production du LPA [97].
Parallèlement à sa synthèse, le LPA subit des processus de dégradation. La très grande
majorité de la dégradation du LPA extracellulaire est effectuée par des « Lipid Phosphate
Phosphohydrolases (LPP) » présentes sur la face externe de la membrane plasmique [98].
Plus connues sous le nom de « Phosphatidic Acid Phosphatases (PAP) », on en dénombre 3
différentes : PP2A, PP2B ou PP2C (LPP1, LPP2 et LPP3 respectivement) [99]. Les
phosphatases sont importantes dans la dégradation du LPA exogène.

22
1.2.1.3. Les récepteurs au LPA
Le LPA agit sur les cellules via ses récepteurs spécifiques couplés aux protéines G [100].
On en compte 6 actuellement classés en deux catégories. La famille des récepteurs Edg
(Endothelial differenciation gene) comprends les trois principaux récepteurs aux LPA :
LPA1, LPA2 et LPA3 anciennement appelés Edg-2, Edg-4 et Edg-7 respectivement (cf
tableau 2). Trois autres types de récepteurs dits « non-Edg » ont été identifiés
successivement peu après [89] : LPA4, LPA5, et plus récemment LPA6 [101]. Ces deux
sous-catégories de récepteurs au LPA sont connues pour avoir des structures et des
fonctions très différentes. De plus, il a été démontré que les récepteurs Edg et non-Edg
étaient phylogénétiquement éloignés les un des autres [89]. Malgré les similitudes entre les
récepteurs LPA1, LPA2 et LPA3, chacun d’entre eux se distribue différemment dans les
tissus [102, 103], ce qui indiquerait des fonctions spécifiques pour chaque récepteur.
Le récepteur LPA1 a été premièrement identifié sous le nom de vzg-1 (ventricular zone
gene) par le groupe de Jerold Chun à cause de son expression dans la zone ventriculaire du
cortex cérébral embryonnaire de la souris [104]. À la même période, on découvrait
l’existence du récepteur Edg-2 présent sur des cellules embryonnaires humaines de rein qui
répondaient aux stimuli du LPA [105]. Les récepteurs vzg-1 de la souris et Edg-2 chez
l’humain partagent 96% d’homologie, ce qui a permis de conclure qu’il s’agissait du même
récepteur dans deux espèces différentes [102]. Le récepteur LPA1/Edg-2 est localisé
principalement au niveau du système nerveux [106, 107], du foie [108] et des reins [109].
On a retrouvé l’expression des récepteurs LPA1 sur des cellules cancéreuses de la prostate
[110]. Il est exprimé à la surface de cellules microgliales de rongeurs [111]. Ce récepteur a
donc un rôle spécifique au niveau du système nerveux. Le LPA, en se fixant sur le
récepteur LPA1, est capable de provoquer la libération de facteurs pro-inflammatoires [112]
et de stimuler la motilité [113] et la survie cellulaire [114]. Les souris déficientes en LPA1
montrent des défauts semblables à ceux trouvés dans les maladies psychiatriques [115]. La
découverte de ce premier récepteur spécifique au LPA a ouvert les portes pour
l’identification des récepteurs LPA2/Edg-4 [102] et LPA3/Edg-7 [116] qui ont été identifiés
quelques années plus tard.

23
Le récepteur LPA2, découvert peu après LPA1, a donc été identifié et cloné [102]. Le
troisième récepteur au LPA, le récepteur LPA3, fut identifié puis cloné un an plus tard
[103]. La répartition tissulaire des récepteurs LPA2 et LPA3 reste très peu claire et très mal
définie dans la littérature. Récemment, l’expression tissulaire des récepteurs LPA2 et LPA3
a été effectuée chez le cochon, où l’on trouve l’expression des récepteurs LPA2 au niveau
des intestins et des ovaires principalement mais notamment dans tous les autres tissus
exceptés les muscles et la rate. Contrairement au récepteur LPA2, le récepteur LPA3 a une
expression maximale au niveau de la rate, des poumons, et du réseau lymphatique, mais est
absent du cerveau (contrairement à LPA1 et LPA2), de l’estomac, des reins et du gros
intestin. Toujours chez le cochon, le récepteur LPA3, tout comme LPA2 est absent des
muscles, et présent dans les ovaires, le thymus et les organes génitaux [117, 118]. Chez la
souris, le LPA2 semble être exprimé dans la région du cerveau, tandis que le récepteur
LPA3 se trouverait plutôt aux niveaux des fosses nasales et du cœur [107]. Il existe des
souris déficientes en LPA3 qui montrent un problème de reproduction à long terme [117],
ce qui suggère la présence de récepteur LPA3 au niveau de leur appareil génital.
Notre équipe a détecté les récepteurs LPA1, LPA2 et LPA3, chez des FLS humains
provenant de patients atteints de PR[60]. Ceci fait un lien supplémentaire entre le LPA et la
PR. Nous avons aussi démontré que l’acide lysophosphatidique était capable de stimuler la
motilité des synoviocytes arthritiques, via les récepteurs LPA1 et/ou LPA3 [60].
Les effets biologiques du LPA s’observent suite à la liaison du LPA sur ses récepteurs
spécifiques, eux-mêmes couplés aux protéines G hétérotrimériques G12/13, Gi/o et Gq. Ces
protéines G activent à leur tour une vaste gamme de voies intracellulaires en aval
(représentées sur la figure 9). Les récepteurs LPA1 et LPA2 induisent une signalisation via
les trois protéines G : G12/13, Gi/o et Gq, tandis que LPA3 se couple à Gi/o et Gq mais pas à
G12/13. Par exemple, la liaison LPA/LPA récepteur, couplée à la protéine G12/13 va induire
l’activation des petites GTPases, qui sont des enzymes régulatrices du réarrangement du
cytosquelette. De la même manière, Gi/o contrôle l’inhibition de l’adénylyl cyclase, la
stimulation de la cascade des Ras-MAPK (mitogen-activated protein kinase), et l’activation
des PI3K (phosphoinositide 3-kinase) qui activent de manière subséquente Rac et
PKB/Akt. Lorsque Gq est activée via le signal LPA/LPA récepteur, la phospholipase C est

24
activée et conduit à la mobilisation du calcium (C.Zhao, revue acceptée à Investigational
Drugs) (résumé dans la figure 9).
Figure 9. Signalisation du LPA via ses récepteurs LPA1, LPA2 et LPA3, couplés aux protéines G. PKB/Akt, Protein kinase B/Akt; PI3K, phosphoinositide 3-kinase; MAPK, mitogen-activated protein kinase ; AC, Adenylyl cyclase ; PLC, Phospholipase C (la figure résume les informations tirées de C.Zhao, revue acceptée à Investigational Drugs).
Ca2+
ACRho Ras/MAPK PI3K
Rac + PKB/Akt
PLC
LPA LPA1/2 3
G GG12/13 q i/o

25
1.2.1.4. LPA et liquides biologiques
Le LPA est libéré majoritairement des plaquettes activées pour se retrouver sous forme
active dans le sang où il est lié à l’albumine [119]. Sa concentration dans le sang est faible
(en moyenne 0.1 µM chez la femme et 0.7µM chez l’homme) [120] mais peut être
augmentée en cas d’activation plaquettaire. Le LPA a aussi été détecté dans la salive [121].
Dans les cas pathologiques, on peut retrouver le LPA dans divers liquides biologiques. Par
exemple, le LPA a été retrouvé dans les liquides d’ascites des patientes atteintes de cancer
des ovaires [122]. De plus, notre groupe a détecté la présence d’autotaxine dans le liquide
synovial de patients atteints de PR [60]. Plus récemment, le LPA a lui aussi été détecté dans
le liquide synovial inflammatoire [123], ce qui suggère une contribution possible du LPA
dans la PR.
1.2.2. Le sphingosine-1-phosphate
1.2.2.1. Historique
Les effets biologiques de la sphingosine-1-phosphate (S1P), contrairement au LPA, ont été
reconnus un peu plus tard dans la littérature, au début des années 90, où l’on discutait de
ses capacités à induire la prolifération des fibroblastes et où l’on proposait déjà son rôle de
second messager [124]. Ce rôle de médiateur ainsi que ses effets prolifératifs ont d’ailleurs
été confirmés quelques années plus tard, sur des cellules cancéreuses humaines [125]. Le
S1P, tout comme le LPA, a beaucoup été étudié dans le contexte des tumeurs. Néanmoins,
le S1P semble de plus en plus être impliqué fortement dans la PR et contribuer au
phénomène inflammatoire. En effet, le S1P augmente la prolifération des synoviocytes
arthritiques et augmente significativement l’expression de COX-2 et PGE2, qui sont des
molécules cibles dans le traitement de la PR [126]. De plus, d’après nos précédentes
recherches, le S1P a été identifié comme un médiateur potentiel dans la migration
cellulaire, la survie des synoviocytes, et la production de cytokines. Nous avons montré que
le S1P provoquait la migration cellulaire et la sécrétion d’IL-6 et d’IL-8 par les FLS
arthritiques [60].

26
1.2.2.2. Métabolisme du S1P
Malgré les ressemblances dans les effets provoqués par le LPA et le S1P sur les cellules, on
observe des différences entre ces deux phospholipides, notamment leurs modes de
production qui leur sont spécifiques. La sphingosine peut être phosphorylée par la
sphingosine kinase pour former la sphingosine-1-phosphate [127]. Le S1P est dégradé
principalement, et de manière réversible, par les S1P-phosphatases [128, 129]. La S1P
lyase quant à elle, modifie irréversiblement le S1P (figure 10) [130].
Figure 10. Production et dégradation du S1P [131].
Deux isoformes de la sphingosine kinase ont été caractérisées : la sphingosine kinase 1
(SphK1) [132] et la sphingosine kinase 2 (SphK2) [133], deux enzymes présentes dans le
cytosol et le réticulum endoplasmique.
Le S1P a été détecté dans le liquide synovial de patients arthritiques, ce qui suggère une
activité sphingosine kinase bien présente [134]. D’ailleurs la voie de signalisation de la
sphingosine kinase 1 est activée dans la PR [135]. De plus, en inhibant la sphingosine
kinase 1, on diminue considérablement la production de TNF-α, d’IL-1β et d’IL-6, qui sont
des cytokines pro-inflammatoires prépondérantes dans la physiopathologie de la PR car
elles contribuent à l'érosion du cartilage et des os en facilitant la production de
métalloprotéinases matricielles (MMP) et en dérégulant la fonction des
chondrocytes/ostéoclastes [134, 136]. Qu’il s’agisse de l’une ou l’autre des isoenzymes,
chacune a un lien avec les effets reliés au S1P, et surtout, avec la PR. Néanmoins, la SphK1

27
semble avoir le mauvais rôle car elle provoquerait des effets plutôt pro-inflammatoires,
promouvant ainsi la PR. En effet, tandis que la SphK1 stimule la croissance et la survie des
cellules, la SphK2 permet l’apoptose et bloque la prolifération cellulaire. En d’autres
termes, les deux iso-enzymes ont des effets apparemment opposés [137]. En empêchant la
sphingosine kinase 1 d’agir par exemple, on arrive à diminuer considérablement la synthèse
des médiateurs pro-inflammatoires provoquée par l’interaction Monocytes-Lymphocytes T
arthritiques [134], alors que la sphingosine kinase 2 exprimée chez les fibroblastes
synoviaux de patients atteints de PR, contribue à l’apoptose, et par conséquent,
n’amplifierait pas l’inflammation [138, 139].
1.2.2.3. Les récepteurs au S1P
On dénombre aujourd’hui 5 récepteurs du S1P couplés aux protéines G (GPCRs) : S1P1 à
S1P5. Parmi eux, S1P1, S1P2 et S1P3 sont largement exprimés dans de nombreux tissus,
tandis que l’expression de S1P4 est restreinte aux tissus lymphoïdes et hématopoïétiques et
S1P5 au système nerveux central [140]. Que ce soit pour stimuler la migration et l’invasion
cellulaire ou bien pour inhiber l’apoptose, le S1P va interagir avec ses récepteurs pour
amplifier l’inflammation. Hormis certains récepteurs qui sont associés à des effets
communs, chaque récepteur a un rôle spécifique dans l’inflammation.
S1P1/Edg1 couplé à la protéine Gi, a été le premier récepteur spécifique au S1P caractérisé
par le groupe de Takuwa en 1998 [141]. Ce récepteur a été détecté, au niveau ARNm et
protéique, chez des synoviocytes humains provenant de patients atteints de PR [126]. Les
cellules endothéliales des vaisseaux expriment S1P1 (et S1P3) [142]. Récemment, le S1P1 a
été impliqué dans l’angiogenèse, et plus particulièrement dans le processus de migration et
d’invasion. Par ailleurs, la régulation de l’interaction S1P/S1P1 a plusieurs fois été suggérée
comme une cible thérapeutique de la PR [126, 143]. Le récepteur S1P1 semble donc, à
chaque fois, avoir un lien avec la pathologie de la PR et d’une manière plus générale avec
l’inflammation.
Le récepteur S1P2/Edg5 quant à lui, semble posséder des propriétés opposées au S1P1. En
effet, ce récepteur régule négativement la migration cellulaire en réponse à des agents
chimiotactiques [144] et entraine la diminution des effets prolifératifs et de la motilité des

28
fibroblastes embryonnaires de souris à la suite d’une stimulation au S1P [145]. De plus,
S1P2 exerce un effet inhibiteur sur l’angiogenèse in vivo [146].
Le récepteur S1P3/Edg3 est souvent étudié avec le S1P1 à cause de leurs effets similaires.
Par exemple, ces deux récepteurs augmentent la prolifération et la migration des cellules
musculaires lisses des vaisseaux, et de l’endothélium, jouant ainsi un rôle prépondérant
dans l’angiogenèse pathologique. De plus, S1P3 protège les cellules épithéliales de
l’apoptose en utilisant les mêmes voies de signalisation que le S1P1 [147], en d’autres
termes, le S1P agit sur ces deux récepteurs pour favoriser la survie cellulaire. S1P3 agit
donc la plupart du temps de la même manière que S1P1, dans le but de contribuer à
l’inflammation, contrairement à S1P2 qui a des effets inverses.
La répartition tissulaire des récepteurs S1P4 et S1P5 est beaucoup plus restreinte que les
trois autres types de récepteurs au S1P. En effet le S1P4/Edg-6 est retrouvé dans les tissus
lymphoïdes et hématopoïétiques ainsi que dans les poumons. Le mécanisme d’action de
S1P4 est très peu connu, sans doute à cause des difficultés d’expression de ce récepteur à la
surface des cellules [148]. Le récepteur S1P5/Edg-8 est retrouvé dans les tissus
périphériques, la rate et les leucocytes sanguins [149], mais principalement au niveau du
système nerveux central [150] où il semble réguler la migration et la survie des
oligodendrocytes [151, 152]. L’expression de S1P5 au niveau ARNm a été détectée chez
des cellules ostéoblastiques : il s’agit du seul lien éventuel connu à ce jour entre ce
récepteur et la PR.
Pour exercer son action, le S1P, comme le LPA, va se lier à ses récepteurs spécifiques, eux-
mêmes pouvant être couplés à différentes classes de protéines G : G12/13, Gi/o ou Gq
principalement. Par exemple, le récepteur S1P1 va être préférentiellement couplé à la
protéine Gi/o via laquelle est activée la voie des MAPK entrainant l’angiogénèse, mais aussi
la tyrosine kinase (TK) conduisant au réarrangement du cytosquelette mis en jeu lors de la
migration cellulaire ainsi que Akt qui conduit à la chimiotaxie des cellules. Les récepteurs
S1P2 et S1P3 quant à eux sont couplés préférentiellement avec les protéines G12/13 et Gq
qui activent respectivement Rho, impliqué dans la migration cellulaire, et la phospholipase
C qui active de manière subséquente la mobilisation du calcium (C. Zhao, revue acceptée à
Investigational Drugs) (résumé dans la figure 11).

29
Figure 11. Signalisation du S1P via ses récepteurs S1P1, S1P2 et S1P3, couplés aux protéines G. TK, Tyrosine kinase; MAPK, mitogen-activated protein kinase ; PLC, Phospholipase C (la figure résume les informations tirées de C.Zhao, revue acceptée à Investigational Drugs).
Ca2+
Rho MAPK Akt PLC
S1P1
G12/13 Gi/o
S1P2S1P3
Gq
TK

30
1.3. Objectifs de l’étude Comme nous venons de le démontrer, le LPA et le S1P ont de nombreux liens potentiels
avec la pathologie de la PR et ont majoritairement des effets pro-inflammatoires, ce qui
favoriserait le développement et l’agressivité de la maladie. Chacune de ces deux molécules
sera donc étudiée séparément, afin de comprendre leurs particularités et leurs effets
spécifiques sur les cellules, dans des contextes bien définis. Le LPA, qui constitue une
grande partie de mon projet, sera traité en premier.
En ce qui concerne le LPA, notre équipe de recherche a publié en 2008 un article
démontrant la présence d’autotaxine, l’enzyme qui produit le LPA, dans le liquide synovial
de patients atteints de PR [60], ce qui suggérait fortement la présence de LPA dans le
synovium. Ceci a très vite été confirmé par un autre groupe, quelques mois plus tard, qui a
trouvé du LPA directement dans la cavité synoviale des patients malades [123]. De plus,
notre équipe a détecté les récepteurs LPA1, LPA2 et LPA3, chez des FLS humains
provenant de patients atteints de PR [60]. Il est donc raisonnable de penser que le LPA
pourrait contribuer à l’inflammation associée à la PR. En plus de sa présence dans de
nombreux sites inflammatoires comme le synovium et au niveau des tumeurs, le LPA est
considéré comme un véritable facteur de croissance, impliqué dans l’angiogenèse et l’auto-
immunité [153]. En effet, le LPA agit comme un chimio-attractant pour les neutrophiles, in
vitro [154]. De plus, nos résultats précédents démontrent que les FLS de patients atteints de
PR expriment les récepteurs LPA1-3 et que le LPA est capable d’induire la production d’IL-
8 en agissant sur ces FLS [60]. L’IL-8 étant un fort chimio-attractant pour les neutrophiles,
on a émis l’hypothèse que le LPA était capable de recruter les neutrophiles de manière
indirecte en passant par les synoviocytes résidents ou bien de manière directe, en agissant
sur les neutrophiles. Pour étudier les propriétés chimio-attractante du LPA dans un contexte
proche de la PR, nous avons choisi de travailler avec le modèle de la poche d’air chez la
souris, qui constitue d’une part, un très bon outil in vivo pour étudier les propriétés chimio-
attractante d’une molécule, et d’autre part, qui est un modèle proche du synovium [155,
156], nous permettant ainsi d’observer facilement le mécanisme du LPA dans un contexte
proche de la PR. Un schéma explicatif du modèle utilisé se trouve annexe 5.

31
Le S1P, qui est lui aussi impliqué dans la physiopathologie de la PR, a été parallèlement
étudié. Nous avons démontré que les synoviocytes exprimaient 3 des 5 récepteurs au S1P :
S1P1-3 et que le S1P induisait non seulement la migration des FLS, mais aussi la sécrétion
de cytokines inflammatoires telles que l’IL-8 et l’IL-6 [36]. Dans le synovium arthritique,
les FLS subissent d’importants changements qui les transforment en puissants effecteurs de
la réaction inflammatoire [157]. Ils participent aussi au processus inflammatoire en
sécrétant de l’IL-8 [157]. Comme nous l’avons dit précédemment, l’IL-8 est la principale
molécule chimio-attractante des neutrophiles, et cette sous-population de leucocytes
correspond à plus de 85% des cellules contenues dans le liquide synovial des patients
atteints de PR [158]. D’ailleurs en bloquant l’IL-8, on prévient le recrutement des
neutrophiles et des lésions tissulaires dans un modèle animal d’arthrite [159]. Les FLS
sécrètent des cytokines en réponse à de nombreux stimuli tels que l’hypoxie ou l’IL-1β
[160]. Dans une étude comparant les liquides synoviaux de plusieurs maladies
inflammatoires, ceux provenant de patients atteints de PR se sont révélés être les plus
faibles en oxygène. Certains cas allant jusqu’à une pression en O2 de 0 mm Hg [161].
L’hypoxie associée au contexte inflammatoire de la PR a de nombreux effets sur tout
l’environnement synovial. Par exemple, celle-ci est capable d’augmenter la production de
TNF et d’IL-1, deux cytokines majeures dans la pathologie de la PR [162].
L’environnement hypoxique du synovium arthritique modifie l’activité transcriptionnelle
des FLS, en partie à cause de la stabilisation du facteur résultant de l’hypoxie : HIF-1
[163]. On a précédemment démontré que les FLS arthritiques synthétisaient et sécrétaient
de l’IL-8 en réponse au S1P [60]. Comme l’hypoxie régule la production de S1P [164,
165], et que celui-ci est impliqué dans la réaction inflammatoire de la PR, on a recherché le
potentiel synergique entre le S1P et l’environnement hypoxique sur la sécrétion d’IL-8 chez
des FLS normaux et chez des FLS provenant de patients atteints de PR.

32
Chapitre 2 : TNF-α priming enhances in vivo LPA-mediated recruitment of leukocytes through LPA1 and LPA3: implication of CXCR2 ligand chemokines
Chenqi Zhao*, Anne Sardella*, Jerold Chun†, Patrice E. Poubelle*, Maria J. Fernandes*, and
Sylvain G. Bourgoin*
* Rheumatology and Immunology, CHUL Research Center and Faculty of Medicine, Laval
University, Québec, QC, Canada G1V 4G2; † The Scripps Research Institute, Department
of Molecular Biology, La Jolla, CA, USA 92037
Running title: LPA recruits leukocytes to inflammatory sites in vivo
Corresponding author:
Sylvain G. Bourgoin, PhD
Centre de Recherche en Rhumatologie et Immunologie, Local T1-49
Centre de Recherche du CHUQ-CHUL
2705 Boul. Laurier, Québec (QC), Canada, G1V 4G2
Phone: (418) 656-4141, ext. 46136
Fax: (418) 654-2765
Electronic address: [email protected]

33
2.1. Résumé en français La PR est une maladie inflammatoire chronique caractérisée par l'infiltration massive de
leucocytes dans les articulations concernées (incluant le synovium) et pouvant conduire à la
destruction du cartilage et des os. Le rôle du LPA dans la PR a récemment émergé dans la
littérature. En effet, le LPA est retrouvé en faible concentration dans le sérum et dans les
liquides biologiques, mais en concentrations élevées aux sites inflammatoire et au niveau
des tumeurs. Ce lysophospholipide provoque de nombreuses réponses cellulaires en se liant
à ses récepteurs spécifiques couplés aux protéines G (GPCRs), à savoir LPA1-6. Bien que le
LPA soit connu comme un chimio-attractant pour les cellules inflammatoires humaines in
vitro, son rôle in vivo reste encore largement inexploré. Dans l’étude suivante, nous avons
utilisé le modèle de la poche d’air chez la souris pour étudier le rôle du LPA dans le
recrutement des leucocytes in vivo, en utilisant d’une part, des agonistes/antagonistes
spécifiques des récepteurs aux LPA et d’autre part, des souris déficientes en récepteurs
LPA3. Nous avons montré que 1) l’injection de LPA dans la poche d’air déclenchait en
effet un recrutement de leucocytes et que les récepteurs LPA1 et LPA3 intervenaient tous
les deux dans ce phénomène; 2) le LPA stimulait le relargage de nombreuses cytokines pro-
inflammatoire telles que KC (chimiokines dérivées des kératinocytes) et IP-10 (protéine
inductible par l’interféron gamma) dans la poche d’air; 3) le TNF-α, injecté 16h avant le
LPA dans la poche d’air, était capable d’augmenter considérablement les effets induits par
le LPA; et 4) le recrutement de leucocytes provoqué par l’injection de LPA dans la poche
d’air était significativement réduit lorsque l’on bloquait les CXCR2 (récepteurs aux
chimiokines de type CXC tel que KC). Nous avons suggéré que le LPA, via ses récepteurs
LPA1 et LPA3, pourrait jouer un rôle important dans le maintient et/ou le déclenchement de

34
l’infiltration des cellules inflammatoires dans les maladies auto-immunes présentant une
inflammation chronique telle que la PR.

35
2.2. Abstract Rheumatoid arthritis (RA) is a chronic inflammatory disease characterized by extensive
leukocyte infiltration into the affected joint space including the synovium, synovial fluid,
and periarticular tissues, eventually leading to joint destruction. The role of
lysophosphatidic acid (LPA) in RA has recently emerged. LPA is a small, simple, naturally
arising bioactive lysophospholipid, present at low concentrations in serum and biological
fluids but in high concentrations at sites of inflammation and in tumors. LPA evokes a
variety of cellular responses via binding to and activation of its specific G protein-coupled
receptors (GPCRs), namely LPA1-6. Even though LPA is a chemoattractant for human
inflammatory cells in vitro, such a role for LPA in vivo remains largely unexplored. In the
present study we used the murine air pouch model of inflammation to study LPA-mediated
leukocyte recruitment in vivo, using selective LPA receptor agonist/antagonist and LPA3
deficient mice. We report that 1) LPA injection into the air pouch effectively induced
leukocyte recruitment and that both LPA1 and LPA3 were involved in this process; 2) LPA
stimulated the release of multiple proinflammatory cytokines/chemokines, particularly
CXCL1 chemokine keratinocyte-derived chemokine (KC) and interferon-inducible protein-
10 (IP-10) in the air pouch; 3) TNF-α injected into the air pouch prior to LPA strongly
potentiated LPA-mediated secretion of cytokines/chemokines including KC, IL-6 and IP-10
which preceded the enhanced leukocyte influx; and 4) blocking CXCR2 significantly
reduced the degree of leukocyte infiltration. We suggest that LPA, via LPA1 and LPA3
receptors, may play a significant role in inducing and/or sustaining the massive infiltration
of inflammatory cells in chronic inflammatory autoimmune diseases such as RA.

36
2.3. Introduction Rheumatoid arthritis (RA) is a chronic inflammatory disease characterized by extensive
leukocyte infiltration into the affected joint space including the synovium, synovial fluid,
and periarticular tissues, eventually leading to joint destruction [166]. A normal healthy
joint contains a virtual space and a nominal amount of synovial fluid that is acellular [167].
In RA, a typical knee joint may hold in excess of 50 ml of synovial fluid containing up to 2
x 109 infiltrating cells. The infiltrated cells in RA joints consist of macrophages, T cells, B
cells, mast cells, dendritic cells, natural killer cells and neutrophils [168]. Neutrophil
migration into the synovial tissue coincides with the onset of clinical disease or disease
severity [169-171]. Once recruited into the joint, leukocytes are exposed to a wide variety
of local proinflammatory cytokines such as TNF-α, IL-1β, GM-CSF and IL-6 [172]. In this
way, infiltration of synovial tissues by activated leukocytes together with inflammatory
mediators generated by both synoviocytes and infiltrated cells trigger synoviocyte
proliferation to form a marked expansion of the synovium called the pannus. The pannus
invades the surrounding joint tissues and leads to progressive cartilage and bone erosion
(reviewed in [173]).
Chemokines are small chemoattractant peptides which play key roles in the
accumulation of inflammatory cells at the site of inflammation. Some chemokines,
particularly CXC chemokines containing the ELR motif (including CXCL1, 2, 3, 5, 6, 7,
and 8) are crucial for neutrophil recruitment via binding to and activation of their receptors
CXCR1 or CXCR2 [174, 175]. In the mouse, IL-8/CXCL8 and a functional homolog of
CXCR1 are absent and keratinocyte-derived chemokine (KC, Groα, CXCL1) and MIP-2
appear to be the major ELR CXC chemokines expressed at sites of tissue inflammation

37
following injury and/or infection [176-180]. It is assumed that neutrophil migration in this
species is mediated mainly by KC and MIP-2 via binding to their sole receptor CXCR2
[181, 182]. In RA, ELR CXC chemokines such as IL-8/CXCL8, GRO-α/CXCL1, and
ENA-78/CXCL5 have been detected in high quantities in synovial fluid, synovial tissue,
and sera (reviewed in [183]).
Lysophosphatidic acid (LPA) induces the synthesis of cytokines and chemokines,
the expression of cyclooxygenase-2 (COX-2), and the production of PGE2 in synoviocytes
isolated from patients with RA [60, 123]. LPA is present in micromolar concentrations in
serum and biological fluids and in higher concentrations at sites of inflammation and tumor
[153]. LPA evokes a variety of cellular responses via binding to and activation of its
specific G protein-coupled receptors (GPCRs) [184]. So far, six GPCRs have been
identified as specific LPA receptors, namely, LPA1-6. Two additional GPCRs, GPR87
[185] and P2Y10 [186], may be activated by LPA. LPA has been implicated in various
physiological (cell growth, differentiation, migration, and survival) and pathological
(angiogenesis, cancer, and autoimmunity) situations. Elevated levels of LPA, LPA
synthesizing enzyme autotaxin (ATX), as well as LPA precursor lysophosphatidylcholine
(LPC), have been detected in synovial fluid from RA patients [123]. Moreover, LPA, either
exogenously applied or contained in synovial fluids of RA patients, stimulated synoviocyte
production of chemoattractive cytokines/chemokines (e.g., IL-8 and IL-6) [60], and other
inflammatory mediators (e.g., COX-2 and PGE2) [123]. Interestingly, the expression of
three LPA receptors, LPA1-3, has been described in synoviocytes [60, 123]. LPA has been
shown to be a chemoattractant for human inflammatory cells in in vitro studies. Chettibi et
al. showed that LPA was a powerful stimulator of human neutrophil polarisation and
motility [187]. Similarly, Fischer et al. described chemoattractive effects of LPA toward

38
human PMNs under agarose [154]. LPA also causes an increase in binding of monocytes
and neutrophils to human aortic endothelial cells [188]. Emerging evidence suggests a role
for ATX-LPA in facilitating lymphocyte migration into lymphoid and chronically inflamed
non-lymphoid tissues [189, 190]. The study of Kanda et al. demonstrated that LPA, locally
generated by ATX in high endothelial venules, facilitates lymphocyte transendothelial
migration via stimulating chemokinesis rather than chemotaxis [189]. The role of LPA in
the recruitment of leukocytes to inflammatory sites in vivo, however, remains elusive.
In the present study, we employed an in vivo experimental model of inflammation,
the murine air pouch model [155, 156, 191], to evaluate the effect of LPA on leukocyte
recruitment in vivo. To assess the LPA receptor dependency and the mechanisms of LPA-
induced leukocyte recruitment in vivo, we used both a pharmacological (LPA receptor
agonists/antagonists) and a genetic (LPA3-/- mice) approach. We demonstrate that both
LPA1 and LPA3 mediate the recruitment of leukocytes into the air pouch through a
mechanism that is dependent on LPA-mediated chemokine synthesis and exacerbated by
the proinflammatory cytokine TNF-α.

39
2.4. Materials and Methods Reagents
1-Oleoyl-sn-glycero-3-phosphate (LPA) was purchased from Sigma (St. Louis, MO, USA).
The LPA1/3 specific receptor antagonist (S)-phosphoric acid mono- (2-octadec-9-
enoylamino-3- [4-(pyridine-2-ylmethoxy)-phenyl]-propyl) ester (VPC32183) was obtained
from Avanti Polar Lipid Inc (Alabaster, AL, USA). The specific LPA2 agonist,
dodecylphosphate, and LPA3 agonist, L-sn-1-O-oleoyl-2-methyl-glyceryl-3-
phosphothionate (2S-OMPT), were obtained from Biomol (Plymouth Meeting, PA, USA)
and Echelon Biosciences Inc (Salt Lake City, UT, USA), respectively. Murine TNF-α was
from PeproTech Inc (Rocky Hill, NJ, USA). CXCR2 antagonist SB225002 was from
Calbiochem (San Diego, CA, USA). KC ELISA kit and Proteome ProfilerTM Mouse
Cytokine Array Panel A were purchased from R&D Systems Inc. (Minneapolis, MN,
USA). The human Cytokine/Chemokine Luminex Multiplex Immunoassay kit was from
Millipore Corporation (St. Charles, MO, USA).
Mice
LPA3 receptor knock out (KO) mice (LPA3-/-) were originally generated in the research
institute of one of the authors (Dr. Jerold Chun, The Scripps Research Institute, La Jolla,
CA, USA). Mice were subjected to genotyping for LPA3 alleles as described previously
[192, 193]. Female Balb/c mice 6-8 weeks old (from Charles River, St.-Colomban, Canada)
were used as background controls. All animal experimental procedures were approved by
the Animal Care Committee at Centre de Recherche du CHUL (CHUQ), Université Laval
and conformed to National Institutes of Health (NIH) guidelines and public law.

40
The air pouch model
Air pouches were raised on the dorsum of mice by subcutaneous injection of 3 ml sterile air
on days 0 and 3 as previously described [155]. Before the injection of air, mice were briefly
anesthetized with isoflurane. On day 7, LPA or LPA receptor agonist/antagonist in 1 ml of
the diluent (PBS + 0.1 % BSA) were injected into air pouches. In some experiments, TNF-
α was injected into air pouches 16 h prior to stimulation with LPA or LPA receptor agonist.
To test a systemic effect of LPA receptor or chemokine receptor CXCR2 antagonism on
LPA-induced leukocyte recruitment, LPA1/3 antagonist VPC32183 and CXCR2 antagonist
SB225002 were intravenously injected 15 min prior to LPA injection into the air pouch. At
specific times, mice were killed by asphyxiation using CO2. The air pouch exudates were
collected by washing once with 1 ml of PBS-5 mM EDTA, and then twice with 2 ml of
PBS-5 mM EDTA. After 5 min centrifugation at 1200 rpm, cell pellets were resuspended,
stained with acetic blue and leukocytes were counted. Fifty to seventy-five thousand cells
were centrifuged onto microscope slides at 500 rpm for 5 min using a cytospin centrifuge.
The slides were then air dried and stained with Diff-Quik (Sigma, St. Louis, MO, USA) to
allow quantification of leukocyte subpopulations. For the purpose of cytokine/chemokine
assays, air pouches were rinced twice with 0.5 ml of cold PBS. The exudates were then
centrifuged at 1200 rpm for 5 min at 4 C and supernatants were kept at -80C for later
cytokine/chemokine assays.

41
Assessment of cytokine/chemokine secretion in the air pouch
Proteome ProfilerTM Mouse Antibody Array Panel A. The air pouches of WT mice
(three mice per treatment group) were injected with TNF-α or its diluent (H2O) 16 h prior
to LPA injection. Expression of multiple cytokines/chemokines in response to LPA, TNF-
α, and TNF-α/LPA were assayed using the Proteome ProfilerTM Mouse Antibody Array
Panel A, following the recommended protocol from R&D Systems. The air pouch exudates
(1 ml/mouse) from each treatment group were combined, mixed with reconstituted
Cytokine Array Panel A Detection Antibody Cocktail, added to the array membranes, and
incubated at 4C overnight. The array was then incubated with streptavidin-horseradish
peroxidase followed by chemiluminescent detection. Each pair of duplicate spots in the
film represents a cytokine/chemokine.
KC ELISA. The air pouches of WT mice (five mice per treatment group) were treated
without or with TNF-α for 16 h prior to LPA injection. Aliquots (100 l) of the air pouch
exudates (five mice per group) obtained at different time points after LPA injection were
analyzed for the level of KC by ELISA according to the manufacturer's instructions (R&D
Systems). Samples were monitored in duplicate. Optical densities were determined using a
SoftMaxPro40 plate reader at 450 nm. The results were compared to a standard curve that
was generated using known concentrations (pg/ml) of KC and were expressed in pg/ml.
Luminex Multiplex Immunoassay. The air pouches of WT mice (five mice per treatment
group) were treated without or with TNF-α for 16 h prior to LPA injection. Inflammatory
chemokines in the air pouch lavages were quantified using a Luminex multiplex

42
immunoassay according to the manufacturer’s instructions (Millipore Corporation). The
mouse cytokine/chemokine multiplex immunoassay was designed for simultaneous
measurement of mouse IL-1β, IL-6, KC, MIP-2, and interferon-inducible protein-10 (IP-
10).
Statistical analysis
Unless otherwise stated, experiments were performed with 5-6 mice/treatment group and
results presented are expressed as mean SE or as representative studies. All statistical
analyses were performed using Prism 4.0 software. Statistical significance of the difference
between non-treated and treated samples, or between WT and KO mice was determined by
t-test (two-tailed p value). For the dose response and time course studies, statistical
significance between control and LPA or LPA agonists (dose response experiments) and
between samples treated at 0 h with those treated at indicated time points (time course
experiments) was determined by ANOVA, Dunnett’s Multiple Comparison Test. P values
less than 0.05 were considered statistically significant.

43
2.5. Results LPA recruits leukocytes to the air pouch in a dose- and time-dependent manner
To examine whether LPA is a chemoattractant for leukocytes in vivo, 1, 3, or 6 g of LPA
was injected into air pouches raised in WT mice. The choice of concentrations of LPA was
based on the results of a previous in vitro study [36]. Leukocyte accumulation in the air
pouch was measured 6 h later. Injection of LPA resulted in a dose-dependent accumulation
of leukocytes in air pouches. Significantly increased leukocyte counts were detected in air
pouches containing 3 g and 6 g of LPA (p<0.01 for LPA vs. control) (Fig. 12A). A
concentration of 3 μg LPA was used for all further experiments. A time course experiment
with 3 μg LPA revealed that an increase in the number of migrated leukocytes started to be
detectable 2 h post-LPA injection, reached the maximal level after 6 h and declined
thereafter (Fig. 12B). Staining of the recruited leukocytes with Diff-Quik revealed that
neutrophils were the predominant subpopulation of LPA-recruited leukocytes at 6 h post-
LPA injection (72.90 ± 0.03 % of the total leukocytes) (data not shown).
LPA1 and LPA3, but not LPA2, are involved in LPA-induced leukocyte recruitment into
the air pouch
To examine the potential involvement of LPA receptor(s) in LPA-induced leukocyte
recruitment, a genetic (using LPA3 receptor deficient mice) and pharmacological (using
LPA receptor agonists/antagonists) approach was used, either alone or in combination.
Because no difference was observed between WT and LPA3+/- mice regarding the number
of leukocytes recruited to the air pouch in response to LPA (p = 0.1186 for WT vs. LPA3+/-
for control group, and p = 0.5034 for WT vs. LPA3+/- for LPA group, Fig. 13A (upper

44
panel)) and OMPT (p = 0.9803 for WT vs. LPA3+/- for control group, and p = 0.9493 for
WT vs. LPA3+/- for OMPT group, Fig. 13D), either WT or LPA3
+/- mice were used as
controls throughout the study.
The number of leukocytes recruited into air pouches in response to LPA increased
in a time-dependent manner (Fig. 12B). A significant accumulation of leukocytes in the air
pouches of WT or LPA3+/- mice was observed 6 h after the injection of LPA (Figs. 13A,
upper and lower panels). This effect, however, was attenuated in LPA3-/- mice by 28 - 55 %
(Figs. 13A, p<0.05). When LPA was injected into air pouches together with the LPA1/3
receptor antagonist VPC32183 (10 μM), LPA-induced leukocyte recruitment was almost
completely blocked in both LPA3+/- (93 % decrease) (p<0.01) and in LPA3
-/- mice (98 %
decrease) (p<0.05) (Fig. 13A, lower panel). Similar to the air pouch administration of
VPC32183, i.v. injection (0.5 mg/kg) of this compound also significantly attenuated
leukocyte recruitment to air pouches by LPA in both WT (70 % decrease, p<0.05) and
LPA3-/- mice (86 % decrease, p<0.05) (Fig. 13B). To further confirm the role of the LPA3
receptor in LPA-mediated leukocyte recruitment, the LPA3 receptor agonist OMPT (1, 3,
and 6 g/pouch) was also injected into the air pouches as of WT mice (Fig. 13C). The
choice of concentrations of OMPT was based on the results from a previous in vitro study
[36]. A significant increase in leukocyte counts was detected at 1 g/pouch of OMPT and
the maximum amount of migrated cells was observed at 3 g/pouch of OMPT (p<0.01 for
OMPT 3 g/pouch vs. control group). On the other hand, the LPA2 receptor agonist
dodecylphosphate had no effect on leukocyte recruitment according to a dose-response
experiment using concentrations ranging from 1-6 g/pouch (data not shown). At 3
g/pouch of OMPT a similar accumulation of leukocytes was observed in the air pouches

45
of WT and LPA3+/- mice (p<0.01 for OMPT vs. control). In contrast to leukocytes recruited
by LPA in LPA3+/- mice where LPA-induced leukocyte recruitment was only partially
attenuated (Figs. 13A, 13B), leukocyte recruitment by OMPT was completely blocked in
LPA3-/- mice (Fig. 13D). The data suggest that both LPA1 and LPA3 are involved in LPA-
mediated leukocyte recruitment in vivo.
TNF-α potentiates LPA- and OMPT-induced leukocyte recruitment into the air pouch
To determine the impact of a proinflammatory environment on LPA- or OMPT-mediated
leukocyte recruitment, air pouches raised on WT mice were injected with 50 ng/pouch of
TNF-α 16 h prior to injecting LPA into the air pouches. As shown in Fig. 14A, TNF-α itself
had no significant effect on leukocyte accumulation compared to non-treated control. TNF-
α injection into the murine air pouch resulted in a transient recruitment of leukocytes, with
maximal accumulation at 2-4 h post-injection and returned to baseline by 8 h post-injection
[194]. TNF-α-recruited leukocytes in the air pouch might have gone through the apoptotic
process [195, 196] after 16 h of treatment. Pretreatment with TNF-α, however, strongly
enhanced LPA-induced leukocyte recruitment. The number of leukocytes recruited in TNF-
α-pretreated air pouches by LPA was 2.47 fold higher than that in unprimed air pouches
(p<0.01). We then compared the effect of TNF-α-priming on LPA- and OMPT-induced
leukocyte recruitment between LPA3+/- and LPA3
-/- mice (Figs. 14B, 14C). In LPA3-/- mice,
LPA- and OMPT-mediated leukocyte accumulation in TNF-α-primed air pouches was
reduced by 25 % (Fig. 14B, p<0.05) and 76 % (Fig. 14C, p<0.01) respectively compared to
LPA3+/- mice.

46
Secretion of multiple cytokines/chemokines in response to LPA precedes leukocyte
recruitment
Since LPA is a neutrophil chemoattractant [154, 187, 188] and it induces
cytokine/chemokine synthesis [60], the following experiments were designed to analyze
whether or not cytokines or chemokines secreted in air pouches play a role in the migration
of leukocytes toward LPA in TNF-α-pretreated air pouches. All experiments in this section
were performed with WT mice. Since we observed that neutrophils are the predominant
leukocytes in exudates of air pouches injected with LPA and TNF-α and that neutrophil
migration in the mouse is mediated mainly by KC and MIP-2 via binding to their sole
receptor CXCR2 [181, 182], the involvement of these two chemokines in LPA-mediated
leukocyte recruitment was first examined. As shown in Fig. 15, intravenous injection of the
CXCR2 antagonist SB225002 15 min prior to LPA administration into the air pouch
significantly decreased LPA-mediated leukocyte recruitment, in a dose-dependent manner,
in both TNF-α-pretreated (Fig. 15B) and unpretreated air pouches (Fig. 15A). In TNF-α-
unpretreated air pouches, SB225002 administration at 0.3 mg/kg, the highest concentration
tested, decreased infiltration of leukocytes induced by LPA by 81.3 % (Fig. 15A, p<0.05).
Similarly, in TNF-α-pretreated air pouch, SB225002 inhibited LPA-mediated leukocyte
recruitment (up to 61 %) in a concentration-dependent manner (Fig. 15B, p<0.05). These
data suggest that the CXCL1 chemokines, KC and/or MIP-2, play a dominant role in LPA-
induced neutrophil recruitment in air pouches.
A qualitative Mouse Cytokine/Chemokine Antibody Array assay, in which a broad
panel of cytokines and chemokines expression in air pouch exudates fluid can be examined,
was then performed. Expression of five cytokines/chemokines was induced by LPA
compared to non-treated control, including BLC (CXCL13/BCA-1), IL-1β, IL-6, IL-16 and

47
MIP-2 (Fig. 16A). The expression of TREM-1 (triggering receptor expressed on myeloid
cells 1), an activating receptor involved in inflammatory disease, was also induced by LPA
(Fig. 16A). When air pouches were primed with TNF-α, LPA induced a broader and more
intensive expression of inflammation markers. In addition to BLC (CXCL13/BCA-1), IL-
1β, IL-6, IL-16, MIP-2, and TREM-1, it also enhanced the expression of I-309
(CCL1/TCA-3), IL-1ra, IP-10, and KC (Fig. 16B).
The next series of experiments was performed to accurately quantify the expression
of cytokines and chemokines. Significant amounts of KC secretion were detected in TNF-
α-primed air pouches (Fig. 17). In TNF-α-primed air pouches, increased KC secretion was
seen starting at 1 h, reached maximum at 2 h after LPA injection (1719 ± 542.20 pg/ml,
p<0.001 for 2 h vs. non-treated control), and declined thereafter (Fig. 17). Thus, the peak of
KC secretion preceded that of neutrophil recruitment in the air pouch.
A more quantitative assay, the Luminex technology, was performed to analyze the
secretion of multiple cytokines/chemokines induced by LPA and OMPT with or without
TNF-α-priming in the air pouch (Fig. 18). Among the 5 cytokines/chemokines tested, MIP-
2 and IL-1β were not detected in response to LPA or OMPT in control and TNF-α-primed
air pouches, suggesting that the amounts of MIP-2 and IL-1β in the air pouch lavages were
below threshold detection limit of Luminex assay. In contrast, significant amounts of KC,
IL-6, and, to a lesser degree, IP-10 were stimulated by LPA and OMPT in TNF-α-primed
air pouches. The release of KC (Fig. 18A), IL-6 (Fig. 18B) and IP-10 (Fig. 18C) in
response to LPA and OMPT was strongly enhanced (in case of KC and IL-6) by TNF-α or
additive (in case of IP-10) to that of TNF-α. Particularly, TNF-α priming alone induced a
small release of KC (230.6 ± 17.08 pg/ml) when monitored after 16 h, which was
comparable to that induced by LPA (197.3 ± 17.75 pg/ml) and OMPT (371.1 ± 31.83 for

48
OMPT). Priming with TNF-α for 16 h, however, increased LPA- or OMPT-induced KC
release 14.04- (p<0.001) and 5.18- (p<0.01) fold, respectively (Fig. 18A). Similarly, TNF-
α, LPA and OMPT alone did not induce significant amount of IL-6 secretion as compared
to non-treated control. However, LPA- and OMPT-mediated IL-6 secretion increased
10.65- (p<0.01) and 6.51- (p<0.05) fold, respectively, in TNF-α-primed air pouches as
compared to that in unprimed air pouches (Fig. 18B).

49
2.6. Discussion Although leukocytes, particularly neutrophils, have been implicated in the onset of clinical
RA and in the regulation of disease activity and severity [197, 198], factors that trigger
leukocyte infiltration are still poorly understood. In the present study we investigated the
role of LPA in leukocyte migration into an inflammatory synovial-like environment. We
observed that LPA induced the recruitment of a mixed inflammatory leukocyte population,
mainly neutrophils, to murine air pouches in a LPA1 and LPA3 receptor-dependent manner.
Moreover, LPA-induced leukocyte accumulation in the air pouch was dependent on the
expression of multiple proinflammatory cytokines/chemokines, particularly KC, since
systemic antagonism of the KC receptor CXCR2 drastically reduced the degree of
leukocyte infiltration. Interestingly, significantly elevated concentrations of these
chemokines were detected in the exudates of TNF-α-primed air pouches, which preceded
an enhanced recruitment of leukocytes in response to LPA. To our knowledge, this is the
first study to characterize the mechanism by which LPA may recruit leukocytes into a
synovial-like environment.
Our findings in the present study clearly demonstrate that LPA is an efficient
stimulator of leukocyte recruitment and confirme several in vitro observations that LPA
may be a chemoattractant for human inflammatory cells. Chettibi et al. demonstrated that
LPA stimulates neutrophil polarisation and motility [187]. Fischer et al. showed that LPA
induced directed migration of PMNs [154]. Our previous in vitro study also demonstrated
that LPA causes an increase in the secretion of cytokines/chemokines, including IL-8, a
potent neutrophil chemoattractant, by RA synovial fibroblasts [60]. Our data also
corroborate another in vivo study in the guinea pig model of airway inflammation, showing

50
that LPA enhances the infiltration of eosinophils and neutrophils into the pulmonary airway
[199].
Since LPA induces the recruitment of leukocytes in vivo, the identification of LPA
receptors involved in this process is of therapeutic interest for diseases such as RA. We
provide direct evidence that LPA-induced leukocyte recruitment is mimicked by the
specific LPA3 agonist OMPT in WT mice, indicating the involvement of this receptor in
the process. This effect, however, is only partially attenuated in LPA3-/- mice, suggesting
receptor(s) other than LPA3 might be involved. When LPA was co-injected with the LPA1/3
receptor antagonist VPC32183 into the air pouch, leukocyte accumulation in the lumen was
almost totally abrogated in both LPA3+/- (mice of this genotype showed similar effects to
WT mice in response to stimulation with LPA) and in LPA3-/- mice. These findings clearly
demonstrate that in addition to LPA3, LPA1 is also involved in LPA-mediated leukocyte
recruitment. We thus suggest that LPA induces leukocyte recruitment into a synovial-like
environment.
We previously reported that LPA causes an increase in the secretion of
cytokines/chemokines, including IL-8, in RA synovial fibroblasts in vitro [60]. Since ATX,
the enzyme that produces LPA, and lysophosphatidylcholine LPC, the substrate of ATX,
are abundant in RA synovial fluids [123], it is reasonable to hypothesize that LPA in
synovial fluid of RA patients may induce the recruitment of inflammatory cells either
directly or indirectly by stimulating inflammatory cytokine/chemokine secretion in the RA
synovium.
TNF-α is a key mediator of the pathogenesis of RA [200]. We, therefore, also
examined the effect of LPA on leukocyte recruitment into murine air pouches injected with
TNF-α. TNF-α-pretreatment strongly enhanced LPA-induced leukocyte recruitment. The

51
enhancing effect of TNF-α on leukocyte recruitment was also seen with LPA3 receptor
agonist OMPT. Since TNF-α has been shown to upregulate LPA3 receptor mRNA
expression, which correlates with the enhancement of LPA-mediated cytokine/chemokine
secretion by RA synoviocytes [60], the potentiating effect of TNF-α-priming in a synovial-
like cavity on LPA-mediated leukocyte recruitment makes this lysophospholipid even more
relevant to RA pathology.
To further characterize the mechanism through which LPA recruits leukocytes in
vivo, we therefore evaluated the participation of CXCR2 ligands in LPA-mediated
leukocyte infiltration in TNF-α-primed and unprimed air pouches. The interaction between
CXCR2 and its cognate chemokine ligands, particularly KC, MIP-2, and IL-8, appears to
play a critical role in controlling the recruitment of leukocytes into sites of acute
inflammation in various model systems (reviewed in [57]). Our data shows that
antagonizing CXCR2 significantly reduced, in a concentration-dependent manner, LPA-
mediated recruitment of leukocytes into both TNF-α-pretreated and unpretreated air
pouches. Moreover, we provide direct evidence that LPA-mediated leukocyte recruitment
in TNF-α-pretreated air pouches preceded the peak of KC secretion, since the synthesis of
this chemokine peaked 2-4h prior to leukocyte accumulation. This suggests that KC is
responsible for the initiation of the leukocyte recruitment to the air pouch induced by LPA,
a conclusion that is supported by other reports. Fujiwara et al., demonstrated that CXCL1
contributes to neutrophil recruitment in monosodium urate crystal-induced arthritis in
rabbits [201]. Similarly, Koch et al. demonstrated in their study that an increased
expression of CXCL1 in the synovial fluid of RA patients correlated with the accumulation
of neutrophils [202].

52 Since CXCR2 antagonism did not completely block LPA-induced leukocyte
recruitment in TNF-α-primed air pouches, our data suggest that other chemokines or
inflammatory mediators could be involved at the beginning of the inflammatory response.
In our quantitative chemokine analysis, another CXCR2 ligand and neutrophil
chemoattractant MIP-2 was not detectable in the air pouch by the time KC was released.
KC and MIP-2 genes might each exhibit a distinct temporal pattern of expression, or kinetic
profile, following LPA stimulation, due to a cell type-specific bias for individual
chemokine gene expression. Indeed, Armstrong et al. recently reported that KC and MIP-2
are expressed in nonoverlapping cell populations at sites of surgical injury and suggested
that this cell type-specific pattern of chemokine expression is independent of the stimulus
used but rather an inherent characteristic of the different cell types (i.e., stromal,
endothelial, and epithelial cells favor KC, and myeloid leukocytes favor MIP-2) [203]. A
similar temporal pattern of KC and MIP-2 expression has also been demonstrated by other
researchers and in different models [204-206]. We detected the production of
proinflammatory cytokine IL-6 and another CXC chemokine IP-10/CXCL10 by LPA in
TNF-α-primed air pouches. The secretion of these two cytokines/chemokines was also
observed 4 h prior to leukocyte accumulation in the air pouch, suggesting their involvement
in the beginning of the inflammatory response. Both IL-6 and IP-10 are relevant to RA
pathophysiology and pathogenesis. High levels of IL-6 can be detected in synovial fluid
and serum in RA patients [207]. IL-6 has been reported to stimulate the release of IL-8 and
MCP-1 [208] and expression of adhesion molecules [209], resulting in recruitment of
leukocytes to inflammatory sites [210]. Similarly, IP-10 is up-regulated in RA synovial
tissue compared with OA synovial tissue [211, 212], and IP-10 serum levels in rheumatoid
factor-positive individuals were higher than those in control (OA or trauma) patients [212].

53
Interestingly, IP-10 may recruit Th1 cells into RA synovial tissue [211]. Thus, we suggest
that LPA binds to LPA1 and LPA3 inducing the release of CXCR2 ligands (e.g., KC) and
other chemoattractant mediators (e.g., IL-6, IP-10, CXCL13, and TREM-1) which then
mediated leukocyte migration into the air pouch. Indeed, LPA induces a global expression
of genes in mouse fibroblasts, including transcriptional factors, cytokines, chemokines, and
metalloproteinases [213], in order to commit fibroblasts to create an inflammatory
microenvironment that favors leukocyte accumulation. Further investigation is required to
determine which cell type in the air pouch lining tissue contributes to the expression of
these cytokines/chemokines.
In our qualitative chemokine analysis, we also observed that LPA, with or without
TNF-α pretreatment triggered the expression of CXCL13 and TREM-1. CXCL13 is a
homeostatic B cell chemoattractant [214] that is required for the normal organization of B
cell follicles in secondary lymphoid organs [215] and to recruit B cells in extralymphoid
sites when overexpressed [216]. CXCL13 messenger RNA and protein expression and
spontaneous CXCL13 secretion were detected in RA synovial fluid T cells [217] and it was
suggested that CXCL13 tissue expression may be critically involved in the process of
ectopic lymphoid neogenesis, a phenomenon seen in 25-50 % of RA patients, via
recruitment of B cells into the synovium [218]. TREM-1 is an immunoglobulin-like cell
surface receptor mainly expressed on neutrophils and CD14 monocytes [219]. TREM-1
expression is increased during both acute [220, 221] and chronic inflammation [222]. Very
recently, Kuai et al. reported that TREM-1 expression is increased in the synovium of RA
patients [223]. The expression of an eosinophil recruiting chemokine CCL1 (I-309), which
has been indicated in inflammatory diseases such as atopic dermatitis [224] and asthma

54
[225], was also seen to be upregulated by LPA in TNF-α-primed air pouches. Quantitative
analysis of these chemokines requires further investigation.
In summary, our study provides a definitive link between LPA and leukocyte
accumulation in a murine synovial-like inflammatory site in vivo. Moreover, LPA-induced
leukocytes recruitment is mediated by the LPA1 and LPA3 receptors that cause an increase
in the secretion of CXCL1 chemokines such as KC, and other proinflammatory cytokines
including IL-6 and IP-10. Since LPA is present in synovial fluids of RA patients, we
suggest that inflammatory cell accumulation in the RA synovium may be attributable in
parts to LPA. Our study contributes to a better understanding of the mechanisms involved
in promoting RA inflammation and implies that both LPA and CXCR2 receptors may be
effective targets for therapeutic intervention in RA.

55
2.7. Figure
0 1 3 6
0.0
0.5
1.0
1.5
LPA (g/pouch)
**
**A
NS
Leu
kocy
tes/
po
uch
(x1
06 )
0 2 4 6 8
0.0
0.5
1.0
1.5
Time (hours)
*
NSB
NS
NS
Leu
kocy
tes/
po
uch
(x1
06 )
Fig. 12. Effect of LPA on leukocyte recruitment into the air pouch. Air pouches were raised in the dorsal skin of WT mice as described in Materials and Methods. (A) On day 7, 1 ml of PBS containing various amounts of LPA (0, 1, 3, and 6 μg) was injected into the air pouches of WT mice. Six hours post-injection, the air pouch was rinsed with PBS and the leukocytes in the air pouch exudate counted. (B) One milliliter of PBS containing 3 μg LPA was injected into the air pouches of WT mice. At various time points (0, 2, 4, 6, and 8 h) air pouch exudates were collected and leukocytes were counted. Data represent the mean ± SE of at least six mice per group. * p < 0.05; ** p < 0.01.

56
0.0
0.5
1.0
1.5
2.0LPA3
+/+
LPA3+/-NS
NS
CTL LPA
A
Leu
kocy
tes/
po
uch
(x1
06 )
0.0
0.5
1.0
1.5
LPA3+/-
LPA3-/-
CTL LPA LPA+VPC 20MLPA+VPC 10M
** * (46 % decrease)
** (93 % decrease)
* (98 % decrease)
Leu
kocy
tes/
po
uch
(x1
06 )

57
0.0
0.5
1.0
1.5
2.0
LPA3+/+
LPA3-/-
CTL LPA LPA+VPC (0.5 mg/kg)
* (72 % decrease)
* (70 % decrease)
* (86 % decrease)
B
Leu
kocy
tes/
po
uch
(x1
06 )
0 1 3 6
0.0
0.5
1.0
1.5
OMPT (g/pouch)
Leu
kocy
tes/
po
uch
(x1
06 )
C **
**
**

58
0.0
0.5
1.0
1.5
2.0
2.5
LPA3+/+
LPA3+/-
LPA3-/-
CTL OMPT
*
**D
Leu
kocy
tes/
po
uch
(x1
06 )
Fig. 13. Involvement of LPA receptor(s) in LPA-induced leukocyte recruitment into the air pouch. One ml of PBS containing 3 μg LPA (A) was injected into the air pouches of WT or LPA3
-/- mice in the absence/presence of the LPA1/3 receptor antagonist VPC32183 (10 μM & 20 μM (A, right panel) or 0.5 mg/kg i.v. (B)). The LPA diluent solution (PBS + 0.1 % BSA) was used as a control (CTL). Leukocyte counts were performed 6 hours post-LPA injection. (C) One ml of PBS containing LPA3 receptor agonist OMPT (0, 1, 3, and 6 μg) was injected into the air pouch of WT mice. The OMPT diluent solution (H2O) was used as a control (CTL). Six hours after LPA or OMPT injection, the air pouches were rinced and the leukocytes in the air pouch exudates counted. (D) One milliliter of PBS containing 3 μg of OMPT was injected into the air pouches of WT (white columns), LPA3
+/- (grey columns), and LPA3-/- mice (dark columns) as
described in (C). Data represent the mean ± SE of at least five mice per group. * p < 0.05; ** p < 0.01.

59
control TNF- LPA TNF-+LPA
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
NS
** (1.47-fold increase)A
*
***
Leu
kocy
tes/
po
uch
(x1
06 )
0.00.51.01.52.02.53.03.54.04.55.05.5
LPA3+/-
LPA3-/-
CTL TNF- LPA+TNF-LPA
(55 % decrease) *
* (25 % decrease)* (25 % decrease)* (25 % decrease)B
Leu
kocy
tes/
po
uch
(x1
06 )

60
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0
4.5
LPA3+/-
LPA3-/-
CTL TNF- OMPT+TNF-OMPT
(67 % decrease) *
** (76 % decrease)C
Leu
kocy
tes/
po
uch
(x1
06 )
Fig. 14. Effect of TNF-α pre-treatment on LPA- and OMPT-induced leukocyte recruitment into the air pouch. At day 6 of air pouch establishment, 50 ng of TNF-α in 1 ml PBS was injected into the air pouch 16 h prior to LPA (A, B) and OMPT (C) air pouch injection in WT (A) or LPA3
+/- and LPA3
-/- (B, C). LPA diluent solution (PBS + 0.1 % BSA) was used as a control. At 6 h after LPA or OMPT injection, the air pouch was rinsed and the leukocytes in the air pouch exudates counted. Data represent the mean ± SE of five mice per group. * p < 0.05; ** p < 0.01.

61
0.0
0.5
1.0
1.5
LPA (3 g/pouch)
SB225002 (mg/kg)
** (81.3% decrease)A
-
-
+
-
+
0.01
+
0.03
+
0.1
+
0.3
*NS
NS
Leu
kocy
tes/
po
uch
(x1
06 )
0.0
1.0
2.0
3.0
4.0
LPA (3 g/pouch)SB225002 (mg/kg)TNF- (50 ng/pouch)
** (61 % decrease)B
-
--
-
-+
+
--
+
-+
+
0.01+
+
0.03
+
+
0.1+
+
0.3+
**NS
NS
Leu
kocy
tes/
po
uch
(x1
06 )
Fig. 15. Effect of CXCR2 antagonist on LPA-mediated leukocyte recruitment. One milliliter of PBS containing 3 μg LPA was injected into TNF-α-unprimed (A) and TNF-α-primed (B) air pouches with or without prior intravenous injection of CXCR2 antagonist SB225002 for 15 min at indicated doses in WT mice. LPA diluent solution (PBS + 0.1 % BSA) was used as a control. At 6 h after LPA injection, the air pouch was rinsed and the leukocytes in the air pouch exudates counted. Data represent the mean ± SE of three to five mice per group. * p < 0.05.

62

63
BL
C (
CX
CL
13/
BC
A-1
)C
5a
I-3
09
(C
CL
1/T
CA
-3)
sIC
AM
-1 IF
N-
IL-1
IL-1
raIL
-6IL
-16
IL-1
7IL
-27
IP-1
0 (C
XC
L10
/CR
G-2
)K
CM
-CS
FJ
E (
CC
L2
/MC
P-1
)M
CP
-5 (
CC
L1
2)M
IG (
CX
CL
9)
MIP
-2R
AN
TE
SS
DF
-1T
IMP
-1T
RE
M-1
0
200
400
600
800
1000NT
LPA
16A
Pix
el d
en
sity
(% o
f co
ntr
ol)

64

65
BL
C (
CX
CL
13/
BC
A-1
)
C5
a
I-3
09 (
CC
L1/
TC
A-3
)
sIC
AM
-1 IF
N-
IL-1
IL-1
ra
IL-6
IL-1
6
IL-1
7
IL-2
7
IP-1
0 (C
XC
L1
0/C
RG
-2)
KC
M-C
SF
JE
(C
CL
2/M
CP
-1)
MC
P-5
(C
CL
12)
MIG
(C
XC
L9
)
MIP
-2
RA
NT
ES
SD
F-1
TIM
P-1
TR
EM
-1
0
500
1000
15005000
5500
6000
6500TNF-TNF-/LPA
Pix
el d
en
sity
(%
of c
on
tro
l)
16B
Fig. 16. Effect of LPA and TNF-α pretreatment on the secretion of cytokines/chemokines in air pouches. LPA (3 μg) in 1 ml PBS was injected into the air pouch with (B) or without (A) TNF-α pre-treatment for 16 h in WT mice. At 2 h after LPA injection, air pouch exudates were collected and cytokine/chemokine secretion was analyzed using Proteome ProfilerTM Mouse Antibody Array Panel A as described in Material and Methods. Circled pairs of duplicate spots represent the cytokine/chemokine secretion whose was upregulated. The graphs represent the pixel density of the spots (% of control). The control used for figure 16A was NT and the control used for figure 16B was TNF-α.

66
- - 1 2 3 4
0
100
2001000
2000
3000
Time (hours)TNF- (50ng/pouch) - + + + + +
***
*
LPA
KC
(p
g/m
l)
Fig. 17. Effect of LPA on KC secretion in the air pouch. LPA (3 μg) in 1 ml PBS was injected into the air pouch with or without TNF-α pre-treatment for 16 h in WT mice. At the indicated time points, air pouch exudates were collected and KC secretion was analyzed by ELISA. Data represent the mean ± SE of five mice per group. *** p < 0.001.

67
0
500
1000
1500
2000
2500
A *** (14.04-fold increase)
** (5.18-fold increase)
***
***
***
TNF- (50 ng/pouch)
LPA (3 g/pouch)
-
-
+
-
-
+
+
+
-
-
+
-
OMPT (3 g/pouch) - - - - + +
KC
(p
g/m
l)
0
250
500
750
1000
1250
1500
1750** (10.65-fold increase)
* (6.51-fold increase)
B
NS
NS
NS
TNF- (50 ng/pouch)
LPA (3 g/pouch)
-
-
+
-
-
+
+
+
-
-
+
-
- - - - + +OMPT (3 g/pouch)
IL-6
(p
g/m
l)

68
0
100
200
300
C
* (1.91-fold increase)
* (4.01-fold increase)
*
NS
NS
TNF- (50 ng/pouch)
LPA (3 g/pouch)
-
-
+
-
-
+
+
+
-
-
+
-
- - - - + +OMPT (3 g/pouch)
IP-1
0 (p
g/m
l)
Fig. 18. Effect of LPA and OMPT on the secretion of cytokines/chemokines in the air pouch. LPA (3 μg) in 1 ml PBS was injected into the air pouch with or without TNF-α pre-treatment for in WT mice. Two hours after LPA injection, air pouch exudates were collected and multiple cytokine/chemokine secretion was analyzed using Luminex Multiplex Immunoassay as described in Materials and Methods. (A) KC secretion. (B) IL-6 secretion. (C) IP-10 secretion. Data represent the mean ± SE of five mice per group. * p < 0.05; ** p < 0.01; *** p < 0.001.

69
2.8. Acknowledgments We thank Dr. Philippe A. Tessier (Centre de Recherche en Infectiologie, Centre de
Recherche du CHUQ-CHUL) for his technical assistance and helpful discussions. We are
grateful to Ms. Lynn Davis for her editorial assistance. We would like to express thanks for
the use of the Bio-imaging Platform of the Centre de Recherche en Infectiologie, Centre de
Recherche du CHUQ-CHUL, for Luminex analysis.

70
2.9. Footnotes 1. This project is supported by research grants from the Arthritis Society of Canada
and the Canadian Arthritis Network (S.G.B). C.Z. is the recipient of the Canadian
Arthritis Network Post-Doctoral Fellowship Award. A.S. is the recipient of the
Canadian Arthritis Network Master Student Trainee Award.
2. C.Z. and A.S. contributed equally to this project. This work was performed by A.S.
in partial fulfillment of her MSc degree at the Faculty of Medicine, Laval
University.
3. Abbreviations: LPA, lysophosphatidic acid; GPCRs, G protein-coupled receptors;
RA, rheumatoid arthritis; MMPs, metalloproteinases; COX2, cyclooxygenase-2;
ATX, autotaxin; LPC, lysophosphatidylcholine, KC, keratinocyte-derived
chemokine; KO, knock out; IP-10, interferon-inducible protein-10.
4. Corresponding author:
Sylvain G. Bourgoin, PhD
Centre de Recherche en Rhumatologie et Immunologie, Local T1-49
Centre de Recherche du CHUQ-CHUL
2705 Boul. Laurier, Québec (QC), Canada, G1V 4G2
Phone: (418) 656-4141, ext. 46136
Fax: (418) 654-2765
Electronic address: [email protected]


72
Chapitre 3 : Hypoxic-like stress-mediated IL-8 production is S1P receptor independent in rheumatoid fibroblast-like synoviocytes
Chenqi Zhao, Anne Sardella, John A. Di Battista†, Patrice E. Poubelle, Maria J.
Fernandes and Sylvain G. Bourgoin
Centre de Recherche en Rhumatologie et Immunologie, Centre de Recherche du CHUQ-
CHUL, Départements d’Anatomie-Physiologie et Médecine, Faculté de Médecine,
Université Laval, Québec, Canada; †Département de Rhumatologie et Immunologie, Centre
Universitaire McGill, Montréal, Québec, Canada.
Correspondence to Maria J. Fernandes, Centre de Recherche en Rhumatologie et
Immunologie, Local T1-49, Centre de Recherche du CHUQ-CHUL, 2705 Boul. Laurier,
Québec (QC), Canada, G1V 4G2. Phone: (418) 656-4141, ext. 46106. Fax: (418) 654-276.
E-mail: [email protected] or Sylvain G. Bourgoin, Centre de Recherche en
Rhumatologie et Immunologie, Local T1-49, Centre de Recherche du CHUQ-CHUL, 2705
Boul. Laurier, Québec (QC), Canada, G1V 4G2. Phone: (418) 656-4141, ext. 46136. Fax:
(418) 654-2765. E-mail: [email protected].

73
3.1. Résumé en français Dans les articulations touchées par la PR, les FLS sécrètent des cytokines en réponse à de
nombreux stimuli tels que l’hypoxie. On a précédemment démontré que les FLS de PR
synthétisaient et sécrétaient de l’IL-8 en réponse au S1P. Comme l’hypoxie régule la
production de S1P, et que celui-ci est impliqué dans la réaction inflammatoire de la PR, on
a recherché le potentiel synergique entre le S1P et l’environnement hypoxique sur la
sécrétion d’IL-8 chez des FLS normaux et chez des FLS provenant de patients atteints de
PR. Nous avons démontré que 1) en conditions hypoxiques (induites par le CoCl2), les FLS
de patients atteints de PR sécrétaient moins d’IL-8 que les FLS normaux; 2) le S1P
synergisait avec le CoCl2 (qui mime l’hypoxie) pour augmenter la production d’IL-8 chez
les deux types de FLS; 3) l’hypoxie au CoCl2 augmentait la sécrétion d’IL-8 via les
récepteurs S1P2 et S1P3 chez les FLS normaux mais pas chez les FLS de PR; et 4)
l’expression de l’ARNm de la sphingosine kinase 1, enzyme qui produit le S1P, est
augmentée en présence de CoCl2 chez les FLS normaux mais pas chez les FLS provenant
de patients atteints de PR. Nous avons suggéré que les FLS provenant de PR étaient moins
sensibles à la sécrétion d’IL-8 due au CoCl2 que les FLS normaux. Le stress hypoxique
mimé par le CoCl2 stimule la production d’IL-8 de manière dépendante aux récepteurs du
S1P.

74
3.2. Abstract Objectives. Fibroblast-like synoviocytes (FLS) are a major source of cytokines in
rheumatoid arthritis (RA) joint. The modulation of the secretion of interleukin 8 (IL-8) by
FLS in response to hypoxia mimetic and/or sphingosine-1-phosphate (S1P) was
investigated.
Methods. FLS from RA and healthy donors were cultured under hypoxic-like or normoxic
conditions with or without S1P. IL-8 production was quantified using ELISA.
Results. Normal and RA FLS under normoxia released similar amounts of IL-8 in response
to S1P. In contrast, RA FLS incubated with cobalt chloride (CoCl2) released less IL-8 than
did normal FLS. The S1P2 and S1P3 receptor antagonists, JTE-013 and CAY10444,
reduced CoCl2-induced IL-8 production in normal but not in RA FLS. CoCl2 synergized
with S1P to enhance IL-8 production by both types of FLS. This synergistic effect of the
hypoxia mimetic and S1P on IL-8 production was inhibited by S1P receptor antagonists.
Conclusions. RA FLS are less prone to CoCl2-induced IL-8 secretion than normal FLS.
Functionally, hypoxic-like stress stimulates IL-8 production in a S1P receptor-dependent
manner in normal but not RA FLS. The results suggest that the sphingolipid pathway,
responsible for the production and/or secretion of S1P, is dysregulated in RA FLS.
KEY WORDS: Rheumatoid arthritis, synoviocyte, hypoxia, S1P, IL-8.

75
3.3. Introduction Rheumatoid arthritis (RA) is a chronic inflammatory disease characterized by profound
cellular and functional changes in the synovial joint. In the RA synovium, fibroblast-like
synoviocytes (FLS) undergo significant changes that render these cells potent effectors of
the inflammatory reaction [157]. FLS contribute to the recruitment of leukocytes through
the synthesis of chemotactic molecules such as IL-8 [157]. IL-8 is at the apex of the
hierarchy of chemoattractant molecules in arthritis since anti-IL-8 blocking antibodies
prevent neutrophil recruitment and tissue damage in an animal model of arthritis [159].
FLS secrete cytokines in response to a variety of stimuli including hypoxic stress
and growth factors like TGF-. Hypoxia is known to synergize with other stimuli to
up-regulate the secretion of various cytokines by FLS. Hypoxia, for instance, enhances
TGF--mediated release of VEGF by FLS .The hypoxic environment of the RA
synovium significantly modifies the transcriptional activity of FLS due, in part, to the
stabilization of hypoxia-inducible factors (HIFs) [163]. Hypoxic stress regulates
sphingosine kinase-1 expression and activity as well as the production of sphingosine-1-
phosphate (S1P) [164, 227].
Sphingosine-1-Phosphate, a bioactive sphingolipid metabolite, regulates a variety of
cellular processes via binding to five G protein-coupled cell surface receptors designated as
S1P1-5. Generally, the S1P content in cells is low and is tightly regulated by the
equilibrium between its synthesis, catalyzed by sphingosine kinases (SphKs), and its
degradation, catalyzed by S1P phosphatases (SPPs) and S1P lyase. Stimulation of SphK
may lead to a rapid and transient increase in intracellular S1P level. Once generated, S1P
can act either intracellularly as a second messenger or be released from cells to activate

76
ed in [228]).
surface S1P receptors to elicit paracrine or autocrine signaling cascades. The properties of
SphK/S1P signaling have led to extensive studies in cancer, immune and inflammatory
responses (as review
We previously reported that RA FLS synthesize and secrete IL-8 in response to S1P
[36]. Since hypoxic stress regulates the production of S1P, the potential for synergy
between the S1P pathway and hypoxic-like environment on the secretion of IL-8 by normal
and RA FLS was investigated.

77
3.4. Materials and methods Cell culture
Human primary FLS were obtained from 4 non-diseased and 4 RA articular synovia by
sequential enzymatic digestion as described previously [229]. RA patients undergoing
arthroplasty were diagnosed based on the criteria developed by the American College of
Rheumatology Diagnostic Subcommittee for RA. After sequential digestion, released cells
were transferred to culture flasks for propagation in DMEM supplemented with 10 % fetal
bovine serum, penicillin (100IU), and streptomycin (100 M). Cells were maintained under
Cells were used between passages 4 to 12.
IL-8 ELISA
FLS (5104 cells) were plated in 24-well plates and serum starved for 24 h prior to
stimulation with 200 µM cobalt chloride (CoCl2) (Sigma, St. Louis, MO) or 5 µM S1P
(Biomol, Plymouth Meeting, PA) in serum-free medium for 24 h. Where indicated, cells
were pre-treated for 30 min with 5 µM of the selective S1P2 or S1P3 receptor antagonists
JTE-013 and CAY10444 (Cayman Chemical, Ann Arbor, MI), respectively, in serum-free
medium prior to stimulation with CoCl2 (200 μM) or S1P (5 μM). Cell culture supernatants
were collected and IL-8 was quantified using ELISA (BioSource International Inc.,
Camarillo, CA). The detection limit was 15 pg/ml. Data are expressed either as pg/ml or as
the percentage of IL-8 secreted relative to the appropriate controls (normal or RA FLS
stimulated with S1P).

78
Semi-quantitative RT-PCR analysis
FLS (5105 cells) were plated in 6-well plates and serum starved for 24 h prior to
stimulation with 200 µM CoCl2 in serum-free medium for various times. At indicated time
points, the cell culture supernatants were removed and cells were washed once with cold
PBS. Total cellular RNA was extracted using Trizol reagent according to the
manufacturer’s instructions (Invitrogen, Burlington, ON, Canada). RNA (1 µg) was
reverse-transcribed using random priming and the Superscript II Reverse Transcriptase
system (Invitrogen). Semi-quantitative PCR reaction was performed using hSphK1 primers
(502 bp product), forward primers, 5’-GGC-AAC-GCG-CTG-GCA-GCT-TC-3’, reverse
primers, 5’-CAG-CGG-CCC-ATG-GGT-GC-3’; hSphK2 (306 bp product), forward
primers, 5’-GCC-TAC-TTC-TGC-ATC-TAC-ACC-TAC-3’, reverse primers, 5’-ATG-
AGG-TTG-AAG-GAC-AGC-CC-3’. PCR conditions were as follows: 3 min at 94°C
followed by 35 cycles with 15 s at 94°C, 30 s at 55°C, 1 min at 68°C, and 7 min at 68°C.
The ribosomal protein RPLP0 mRNA was used as an internal PCR control. RPLP0 (248 bp
product) primer sequences are as follows: forward primers, 5’-GTT-GTA-GAT-GCT-
GCC-ATT-G-3’, reverse primers, 5’-CCA-TGT-GAA-GTC-ACT-GTG-C-3’. The PCR
products were subjected to electrophoresis on a 2 % agarose gel and visualized by ethidium
bromide staining. Densitometry analysis was used for band quantification using the
software Alphalmager 2000. SphK1 and SphK2 genes were normalized to expression of
RPLP0 as an endogenous control.

79
Statistical analysis
Experiments were performed three times for each donor and results presented are expressed
as means SE. Statistical significance was assessed by 1-way ANOVA followed by
Dunnett’s Multiple Comparison post test (GraphPad Software Inc.). Significance was set at
P < 0.05.

80
3.5. Results Hypoxic-like conditions differentially regulate IL-8 secretion in normal and RA FLS.
Hypoxia induces IL-8 mRNA synthesis in RA FLS [163]. Whether the observed increase in
IL-8 mRNA correlates with an increase in IL-8 protein remains unknown. We, therefore,
compared the effect of normoxic and hypoxic-like conditions on IL-8 secretion by normal
and RA FLS. To mimic hypoxia, FLS were incubated with cobalt chloride (CoCl2), a
chemical inducer of hypoxia-inducible factor-1 (HIF-1) [230]. Under normoxic conditions,
small amounts of IL-8 were secreted by both normal and RA FLS (< 3 pg/ml). When
incubated with CoCl2 (Fig. 19A), normal FLS released significantly larger amounts of IL-8
(660 ± 137 pg/ml) compared to RA FLS (126 ± 27 pg/ml).
Hypoxic-like conditions synergize with S1P to enhance IL-8 production by normal
and RA FLS. Hypoxia is known to synergize with other stimuli to enhance the release of
cytokines by FLS [226]. We previously reported that S1P stimulates IL-8 production in RA
FLS [36]. To determine whether hypoxia and S1P have additive or synergistic effects on
IL-8 production by FLS, cells were cultured in the presence or absence of S1P under
normoxic or hypoxic-like conditions. We initially compared the secretion of IL-8 by
normal and RA FLS in response to S1P alone under normoxic conditions. As shown in Fig.
19B, similar amounts of IL-8 were produced by normal FLS (171 ± 31 pg/ml) and RA FLS
(146 ± 13 pg/ml) stimulated with S1P under normoxic conditions. Under hypoxic-like
conditions in the presence of S1P, IL-8 secretion was significantly enhanced in both normal
and RA FLS (1159 ± 141 pg/ml and 683 ± 68 pg/ml, respectively) when compared to
normoxic conditions (Fig. 19B). The effect of the hypoxia mimetic and S1P in combination
on IL-8 production was significantly superior to the arithmetic sum of the effect of S1P and

81
ctively).
hypoxia alone both in normal (1159 ± 141 pg/ml vs. 171 ± 32 pg/ml + 683 ± 27 pg/ml; P <
0.001) and RA FLS (683 ± 68 pg/ml vs. 146 ± 13 pg/ml + 125 ± 27 pg/ml; P < 0.001). The
data indicate that the hypoxia mimetic and S1P have a synergistic effect on IL-8 production
in both normal and RA FLS.
Hypoxic-like conditions enhance IL-8 secretion through S1P2 and S1P3 receptors in
normal but not RA FLS. S1P regulates a variety of cellular processes through binding to
G protein-coupled receptors, S1P1-5 [231]. We previously reported a role for S1P2 and
S1P3 in S1P-mediated IL-8 secretion in RA FLS [36]. Since hypoxia synergizes with S1P
in the production of IL-8 by FLS, the effect of the S1P2 antagonist JTE-013 and of the
S1P3 antagonist CAY10444 on S1P- and CoCl2-mediated IL-8 production in normal and
RA FLS was investigated. CAY10444 and JTE-013 reduced S1P-mediated release of IL-8
by 51 ± 5 % (P < 0.01) and 80 ± 5 % (P < 0.01) in normal FLS and by 64 ± 8 % (P < 0.01)
and 93 ± 0.6 % (P < 0.01) in RA FLS cultured under normoxic conditions, respectively
(Fig. 20A). Interestingly, CAY10444 and JTE-013 significantly reduced CoCl2-mediated
secretion of IL-8 by 59 ± 7 % and 27 ± 7 % (P < 0.001) in normal FLS but had no
inhibitory effect on CoCl2-induced IL-8 production in RA FLS (Fig. 20B). When cells
were incubated with S1P and CoCl2 in combination (Fig. 20C), the S1P3 and S1P2
antagonists reduced IL-8 secretion in normal (P < 0.01 and P < 0.01, respectively) and RA
FLS (P < 0.05 and P < 0.01, respe
Hypoxic-like conditions upregulate SphK1 mRNA expression in normal but not RA
FLS. Hypoxic stress regulates sphingosine kinase-1 expression and activity as well as the
production of sphingosine-1-phosphate (S1P) in different cell types [164, 227]. We report
CoCl2 (which mimic the hypoxic stress) upregulates sphingosine kinase-1 mRNA

82
expression in normal (Fig 21A) but not in RA FLS (21B). Sphingosine kinase-2 expression
was not up-regulated in both normal and RA FLS (data not shown).

83
3.6. Discussion FLS exhibit altered phenotypes in RA. Molecular changes in signaling cascades, apoptosis
responses and in the expression of adhesion molecules as well as matrix-degrading
enzymes have been reported for RA FLS [157]. The identification and characterization of
additional distinct characteristics of RA FLS will further our understanding of the
pathogenesis of this disease. Herein, we report a novel characteristic of RA FLS that
distinguishes these cells from normal FLS. RA FLS are less prone to release IL-8 in
response to the hypoxia mimetic CoCl2 in comparison to their normal counterparts. This
difference may be due to the signaling pathways employed by normal and RA FLS to
induce the release of IL-8.
The regulation of the release of IL-8, a potent chemokine, by stimuli in the
rheumatoid synovium remains poorly characterized. We report that the hypoxia mimetic
CoCl2 stimulates IL-8 production in RA FLS. This observation not only corroborates the
report that hypoxia induces the synthesis of IL-8 mRNA by RA FLS [163], but also
indicates that the hypoxic-like environment induced by CoCl2 mimics the effect of hypoxia
on FLS. Remarkably, the quantity of IL-8 released by normal FLS incubated with CoCl2 is
superior to that of RA FLS, suggestive that molecular pathways involved in IL-8
production are altered in RA FLS. Whether this difference between normal and RA FLS is
acquired during the early stages of RA or the later stages of the disease remains to be
determined.
To further characterize the difference in the release of IL-8 by FLS in response to
CoCl2, the S1P signaling pathway was investigated since both CoCl2 and hypoxia induce
the activation of sphingosine kinase-1 [164]. In normal FLS, IL-8 secretion induced by the
hypoxia mimetic CoCl2 is due, in part, to activation of the S1P2 and S1P3 receptors. The

84
data suggest that normal FLS synthesize and release S1P in response to hypoxia. In
contrast, S1P receptors are not involved in CoCl2-induced IL-8 production in RA FLS,
indicative that either these cells do not synthesize S1P or that the turnover of S1P is higher
in the RA FLS under our experimental conditions. Regarding the latter, the expression and
activity of the S1P phosphatase SPP2, a S1P-degrading enzyme, was found to be up-
regulated by inflammatory stimuli in various cells (e.g., neutrophils, endothelial cells)
[232]. Notably, up-regulation of SPP2 was detected in samples of skin lesions from patients
with psoriasis, a chronic inflammatory skin disease [232].
The reduced ability of RA FLS to release large quantities of IL-8 in a hypoxic
environment when compared to those produced by normal FLS is overcome by the addition
of S1P. The synergistic effect of hypoxia and S1P on IL-8 production confirms the
presence of functional S1P receptors in RA FLS. Our data suggest that a step in the
sphingolipid pathways responsible for the production and/or release of S1P under hypoxic-
like conditions is dysfunctional in FLS isolated from RA patients.

85
3.7. Funding This work was supported by research grants provided by Canadian Institutes of Health
Research and the Arthritis Society of Canada (to S.G.B.) and grants from Canadian
Arthritis Network Post-Doctoral Fellowship Award (to C.Z.) and Master Student Award (to
A.S.).

86
3.8. Acknowledgements We thank Ms. Lynn Davis for her editorial assistance.

87
3.9. Disclosure statement The authors have declared no conflicts of interest.

88
3.10. Figures A
- + - +0
100
200
300
400
500
600
700
800
900
CoCl2 (200 M)
normal donor RA donor
**
B
0
200
400
600
800
1000
1200
1400
S1P (5 M) - + + - + +CoCl2 (200 M) - - + - - +
normal donor RA donor
**
**
Figure 19. CoCl2 and S1P, either alone or in combination, stimulates IL-8 secretion in normal and RA FLS. Human primary FLS from normal (n = 4) and RA (n = 4) donors were incubated with 200 µM CoCl2, 5 µM S1P, or CoCl2 and S1P in combination. The amounts of IL-8 released in the supernatants were monitored after 24 h of stimulation. The data are the means ± SE from four experiments (4 different donors) performed in triplicate (3 individual experiments). For statistical comparative analyses, we compared RA and normal FLS treated with CoCl2 (A), and RA or normal FLS treated with S1P and CoCl2 with cells stimulated with S1P alone (B). ** p<0.01.

89
A
0
25
50
75
100
S1P (5 M) - + + + - + + +CAY10444 (5M) - - + - - - + -JTE-013 (5 M) - - - + - - - +
Normoxia
normal donor RA donor
**
**
**
**
B
Hypoxia
0
100
200
300
400
500
600
700
800
CoCl2 (200 M) - + + + - + + + CAY10444 (5M) - - + - - - + -JTE-013 (5 - - - + - - - +
normal donor RA donor
***
***

90
C
Hypoxia
0
100
200
300
400
500
600
700
800
900
CoCl2 (200 M) - - + + + - - + + + S1P (5 M) - + + + + - + + + +CAY10444 (5M) - - - + - - - - + -JTE-013 (5 - - - - + - - - - +
normal donor RA donor
*
**
**
**
Figure 20. Differential involvement of S1P receptors in CoCl2-mediated IL-8 secretion by normal and RA FLS. Human primary FLS from normal (n = 4) and RA (n = 4) donors were incubated with 5 µM S1P alone (A), 200 µM CoCl2 alone (B), or S1P and CoCl2 in combination (C). Where indicated, cells were pre-treated with S1P3 antagonist CAY10444 (5 µM) or S1P2 antagonist JTE-013 (5 µM) for 30 min before stimulation with S1P and/or CoCl2. The amounts of IL-8 released in the supernatants were monitored after 24 h stimulation. Data are expressed as percentage of S1P-induced IL-8 production in normal or RA FLS (A, C), or in pg/ml (B). The data are the means ± SE from four experiments (4 different donors) performed in triplicate (3 individual experiments). For statistical comparative analyses, the samples stimulated with S1P alone (A), CoCl2 alone (B), and S1P in combination with CoCl2 (C) were compared to those stimulated in the presence of CAY10444 or JTE-013. * p<0.05; ** p<0.01; *** p<0.001.

91
A
0 0.25 0.5 1 2 4 8
0
100
200
300
400
**
CoCl2, 200 M (hour)
normal donor
SP
hK
1/R
PL
P0
Ra
tio
(m
RN
A,
% o
f C
TL
)
B
0 0.25 0.5 1 2 4 8
0
50
100
150
200
CoCl2, 200 M (hour)
RA donor
NS
SP
hK
1/R
PL
P0
Ra
tio
(m
RN
A,
% o
f C
TL
)
Figure 21. CoCl2 stimulates Sphingosine kinase 1 mRNA expression in normal but not in RA FLS. Human primary FLS from normal (n=4) and RA (n=4) donors were incubated with CoCl2 at different time points (0, 15 min, 30 min, 1h, 2h, 4h, 8h). Sphingosine kinase-1 expression was examined at the mRNA level using RT-PCR. The data are the means ± SE from four experiments (4 different donors) performed in triplicate (3 individual experiments). For statistical comparative analysis, we compared non-treated FLS of normal (A) and RA (B) donors with FLS treated with CoCl2 at the time points indicated. RPLP0 was used as the Housekeeping gene to compare with SphK1 mRNA expression. ** p<0.01.

92
Chapitre 4 : Interprétation des résultats
4.1. Discussion Dans cette étude in vivo, nous avons réussi à démontrer que le LPA possédait des propriétés
chimio-attractantes pour les leucocytes, en recrutant ces derniers dans la poche d’air, et que
les récepteurs LPA1 et LPA3 étaient impliqués dans ce recrutement. On a aussi observé par
coloration Diff-Quick que la majorité des leucocytes recrutés étaient des neutrophiles
(72.90 ± 0.03 % des leucocytes totaux) (données non montrées). Ensuite, nous avons
prouvé que le recrutement des leucocytes induit par le LPA nécessitait la synthèse de
nombreuses cytokines/chimiokines, notamment KC, en utilisant l’antagoniste des
récepteurs CXCR2 (récepteurs sur lesquels KC est capable de se fixer), qui diminue
drastiquement le nombre de leucocytes recrutés par le LPA. D’après nos connaissances
actuelles, notre étude est la première qui caractérise d’une part les mécanismes par lesquels
le LPA pourrait recruter des leucocytes dans un environnement, qui par sa structure,
ressemble à celui du synovium, et d’autre part, qui identifie les récepteurs impliqués.
De nombreuses études suggèrent que le LPA est un chimioattractant pour les cellules
inflammatoires. Une équipe de recherche a par exemple démontré que le LPA pouvait agir
in vitro, comme un agent chimio-attractant sur les neutrophiles humains [154]. Chez le
cochon d’Inde, il a été démontré que le LPA pouvait augmenter l’infiltration et l’activation
des éosinophiles et des neutrophiles dans les voies aériennes pulmonaires [199]. De plus,
nos précédentes études in vitro ont démontré que le LPA induisait la sécrétion d’IL-8,
principale chimiokine impliquée dans le recrutement de neutrophiles [233], chez des
synoviocytes provenant de patients atteints de PR [60]. Une autre étude suggère que
l’autotaxine extracellulaire, l’enzyme productrice de LPA, exacerberait l’inflammation en
augmentant l’expression des intégrines à la surface membranaire des neutrophiles via LTB4
pour augmenter la chimiotaxie [234]. En effet, il est important de signaler que notre équipe
de recherche a mis en évidence la présence de cette enzyme, appelée aussi
lysophospholipase D, dans le liquide synovial de patients atteints de PR [60], et que des
quantités importantes de LPA et de son substrat, le LPC, ont été détectées dans les fluides
synoviaux de patients arthritiques [123]. Il est donc raisonnable d’imaginer que le LPA

93
présent dans les liquides synoviaux de patients arthritiques pourrait stimuler un recrutement
des cellules inflammatoires, de manière directe sur les leucocytes ou de manière indirecte
en provoquant la sécrétion de cytokines/chimiokines inflammatoires, par les cellules du
pannus synovial.
Dans nos précédentes études, on avait démontré que le LPA stimulait la sécrétion d’IL-8
par les fibroblastes synoviaux de patients atteints de PR et que les récepteurs LPA3 étaient
impliqués dans ce phénomène [60]. Il est donc raisonnable de penser que le récepteur LPA3
pourrait avoir un rôle à jouer dans le recrutement des leucocytes provoqué par le LPA, via
la synthèse de cytokines chimioattractantes pour les neutrophiles. Afin de confirmer cette
première hypothèse et de déterminer quels sont les récepteurs du LPA réellement impliqués
dans ce processus de migration des leucocytes au site inflammatoire de la poche d’air, nous
avons utilisé un agoniste du LPA, l’OMPT, qui, en se fixant spécifiquement sur les
récepteurs LPA3, provoque un recrutement significatif des leucocytes. Cependant, en
utilisant des souris K.O. pour le récepteur LPA3, et en éliminant ainsi les réponses
fonctionnelles des récepteurs LPA3, on n’inhibe pas complètement l’infiltration des
leucocytes dans la poche d’air (infiltration leucocytaire due au LPA). Ceci suggère qu’un
autre récepteur était impliqué dans ce phénomène. En utilisant un agoniste du récepteur
LPA2, on n’obtient pas de recrutement significatif de leucocytes par rapport au contrôle
(résultats non montrés), ce qui écarte l’implication du récepteur LPA2 dans ce mécanisme.
Par contre, en bloquant les récepteurs LPA1 et LPA3 en même temps, on obtient une
diminution de 98% du recrutement de leucocytes induit par le LPA dans les souris sauvages
et LPA3 -/-, ce qui suggère que le recrutement des leucocytes due au LPA est dépendant des
deux récepteurs LPA1 et LPA3. Ces deux récepteurs ont été impliqués dans la chimiotaxie
des cellules endothéliales via la production indirecte d’IL-8 [235]. Notre étude ne permet
cependant pas d’identifier clairement les cellules de la poche d’air qui produisent les
chimiokines impliquées dans le recrutement de leucocytes déclenché par le LPA. De plus, il
a été démontré qu’en diminuant l’expression des récepteurs LPA2 et LPA3, on inhibait la
production d’IL-8 dans les cellules ovariennes cancéreuses [236]. Le récepteur LPA1 a
aussi été démontré comme étant capable de stimuler la prolifération, la migration, et
l’adhésion des cellules cancéreuses du colon ainsi que la production de l’IL-8, tout en

94
excluant l’implication du récepteur LPA2 [237]. Il semble donc que les récepteurs LPA qui
contribuent à la synthèse d’IL-8 varient selon le type cellulaire.
Le TNF-α est une molécule clé dans le déclenchement et la chronicité des réponses
inflammatoires qui se produisent dans le synovium de la PR [238]. La surexpression du
TNF-α chez la souris déclenche le développement d’une polyarthrite érosive avec de
nombreuses caractéristiques observées chez les patients atteints de PR [200]. Nous avons
donc évalué le rôle potentiel que le TNF-α pouvait jouer dans le recrutement des leucocytes
en réponse au LPA dans notre modèle in vivo de la poche d’air. Dans cette optique, nous
avons prétraités les poches d’air avec du TNF-α, 16h avant l’injection de LPA, afin de créer
un microenvironnement inflammatoire. Cependant, 16h après l’injection de TNF-α, on
n’observe aucun changement, par rapport au contrôle, quant au nombre de leucocytes
recrutés. Ces observations n’excluent pas que le TNF-α puisse provoquer un recrutement de
leucocytes dans la poche d’air. D’ailleurs, le TNF-α est connu pour être un chimioattractant
pour les neutrophiles [239]. Dans un modèle murin de poche d’air, l’injection de TNF-α
entraine un recrutement de leucocytes qui est maximal à 4h post-injection et retourne à son
niveau de base après 8h [194]. Ceci explique l’absence de leucocytes à 16h post-injection
dans nos expériences avec le TNF-α seul. Par contre, le prétraitement au TNF-α a permis
d’augmenter considérablement les effets due au LPA, et à l’OMPT (agoniste du LPA
spécifique aux récepteurs LPA3). Dans nos précédentes études in vitro, nous avons
démontré que le TNF-α était capable d’augmenter l’expression des récepteurs LPA3, au
niveau ARNm, chez des synoviocytes de type B provenant de patients atteints de PR, et que
le LPA stimulait la synthèse de cytokines/chimiokines via les récepteurs LPA3 (sans
exclure le récepteur LPA1) [60]. De plus, une étude a démontré la présence de cellules
ressemblant aux FLS dans la poche d’air [156]. Nos résultats, qui montrent qu’un
prétraitement au TNF-α augmente considérablement les effets provoqués par le LPA et
l’OMPT, suggèrent que le TNF-α aurait pu augmenter l’expression des récepteurs LPA3 sur
certaines cellules présentes dans la poche d’air (potentiellement les FLS) et ainsi, pu
permettre au LPA de se fixer sur un plus grand nombre de récepteurs, entrainant par
conséquent une augmentation du recrutement des leucocytes. Cette hypothèse n’exclue pas
d’autres possibilités telles que la modulation par le TNF-α des voies de signalisation qui ont
été impliquées dans la production d’IL-8 en réponse au LPA.

95
Puisque le LPA est un chimioattractant pour les neutrophiles [199] et qu’il est aussi capable
d’activer la synthèse de chimiokines telles que l’IL-8 [60], nous avons voulu investiguer la
participation de médiateurs chimiotactiques pour les leucocytes tels que les chimiokines,
pour essayer d’expliquer le mécanisme par lequel le LPA recrute les leucocytes dans la
poche d’air. L’IL-8, et ses analogues KC ou MIP-2 chez la souris, via leur liaison aux
récepteurs CXCR2 jouent un rôle principal dans le recrutement des leucocytes aux sites
d’inflammation aiguë dans de nombreux modèles animaux [57]. Nous avons
spécifiquement évalué l’implication de ces ligands du CXCR2 dans l’infiltration des
leucocytes dans les poches d’air (suite à la stimulation au LPA), prétraitées ou non au TNF-
α. En effet, nos résultats montrent qu’en bloquant les récepteurs CXCR2, le nombre de
leucocytes recrutés dans la poche d’air après injection de LPA, est significativement réduit
(81.3%) et ceci d’une manière dépendante de la dose. Pour confirmer cette observation,
nous avons mesuré la présence de KC dans la poche d’air, qui a révélé que le pic de
sécrétion de KC survient environ 2h à 4h avant l’accumulation maximale de leucocytes
dans la poche d’air. De plus, lors de l’étude quantitative, KC a été détectée en quantité très
importante dans les poches d’air ayant été prétraitées au TNF-α, où l’on peut observer un
net effet synergique entre le LPA et le TNF-α. Ces observations suggèrent que KC pourrait
être responsable de l’initiation du recrutement des leucocytes due au LPA et que le TNF-α
potentialise cet effet via la sécrétion de KC.
Lorsqu’on bloque les récepteurs CXCR2 avant l’injection de LPA, on arrive à diminuer à
plus de 80% le recrutement des leucocytes dans la poche d’air. Dans ces conditions
expérimentales, les chimiokines MIP-2 et KC sont incapables de se fixer sur leurs
récepteurs CXCR2 car ceux-ci sont bloqués par l’antagoniste. Ceci suggère un rôle très
probable de KC, mais aussi potentiellement de MIP-2, dans le recrutement des leucocytes
dans la poche d’air induit par le LPA. D’ailleurs, il a été démontré que l’inflammation
chronique des articulations dépendait de nombreuses chimiokines pro-inflammatoires dont
principalement MIP-2 [240], qui est aussi responsable du recrutement des leucocytes chez
la souris [241]. À la suite de l’injection de LPA, nous avons réussi à détecté la présence de
MIP-2 dans les lavages des poches d’air. Bien que l’antagoniste bloque la majorité des
effets due au LPA, celui-ci ne les supprime pas complètement. Ceci nous a donc poussés à
rechercher l’éventuelle présence d’autres cytokines/chimiokines qui pourraient, elles aussi,

96
être impliquées dans le recrutement des leucocytes dans la poche d’air, à la suite de
l’injection de LPA.
Parmi les cytokines/chimiokines détectées dans nos études qualitatives et quantitatives, on
observe que l’IL-6 est présente en quantité importante dans les lavages de poches d’air
traitées au LPA et que cette quantité double lors d’un prétraitement au TNF-α. En effet,
l’IL-6 a un rôle prépondérant dans la PR car on la retrouve en quantité importante dans les
liquides synoviaux de patients atteints de PR [207] et les anti-IL-6 constituent déjà un
traitement existant contre la PR [242]. De plus, l’IL-6 va permettre, lors d’une
inflammation aiguë, d’amorcer la réponse immunitaire, et lors d’une inflammation
chronique, d’augmenter l’infiltration des cellules mononuclées comme les monocytes pour
amplifier l’inflammation [243]. Nos résultats suggèrent donc que l’IL-6 pourrait aussi
participer au recrutement de certaines populations de leucocytes dans la poche d’air à la
suite de l’action du LPA.
Le LPA a aussi provoqué l’expression de TREM-1 (« triggering receptor expressed on
myeloid cells 1 »), un récepteur impliqué dans les maladies inflammatoires et capables
d’induire la production d’IL-8 par les neutrophiles (résumé dans [244]). Très récemment,
Kuai et al. ont reporté que l’expression de TREM-1 était augmentée dans le synovium de
patients atteints de PR et que l’activation de TREM-1 conduisait à la production de
nombreuses cytokines pro-inflammatoires, notamment TNF-α, IL-1β et IL-8 [223].
Notre étude qualitative des cytokines présentes dans les lavages de poches d’air montre la
présence de l’IL-1β et de l’IL-1ra (« IL-1 receptor antagonist »), autant chez les souris
traitées au LPA seul que chez les souris ayant subit le prétraitement au TNF-α avant
l’injection de LPA. Ceci suggère une fois de plus le rôle possible du LPA dans
l’inflammation chronique de la PR. En effet, ces deux molécules ont des rôles importants
dans la PR, quasiment aussi importants que le TNF-α, car elles déterminent la sévérité de la
maladie [245]. Wong et al (1997) ont par ailleurs proposé une action neurorégulatrice de
l'IL-1β lors de l'inflammation systémique [246].
L’IL-16 (« lymphocyte chemoattractant factor »), qui est un chimioattractant des
lymphocytes et des éosinophiles [247], a aussi été détectée dans les lavages des poches

97
d’air traitées au LPA. Ceci suggère que l’IL-16 serait lui aussi potentiellement responsable,
de la même manière que KC et MIP-2, du recrutement des leucocytes dans la poche d’air,
amorcé par le LPA. De plus, les fibroblastes humains expriment l’IL-16 [248], ce qui sous-
entend que les fibroblastes qui tapissent la poche d’air pourraient synthétiser cette cytokine
en réponse au LPA. D’après nos connaissances, c’est la première étude qui suggère que le
LPA induit in vivo la sécrétion d’IL-16.
BLC (« B lymphocyte chemoattractant »)/CXCL13, dont la présence est détectée dans les
exsudats des poches d’air prétraitées au TNF-α puis traitées au LPA, a été récemment
proposé comme un nouveau marqueur potentiel dans la pathologie de la PR [249]. Cette
chimiokine est exprimée par les cellules dendritiques [250], qui ont un rôle indispensable
dans la présentation antigénique d’une manière générale, et par conséquent, dans la
pathologie de la PR. En effet, ces cellules sont présentes dans les liquides synoviaux [251]
et dans le synovium [252] des patients atteints de PR. Le milieu riche en cytokines de la PR
promeut la différenciation des cellules dendritiques et leurs fonctions qui conduisent à la
présentation antigénique aux lymphocytes T, responsable de l’auto-immunité [253]. Notre
étude ne permet pas d’identifier la présence de cellules dendritiques à l’intérieur de la
poche d’air. Cependant, ces résultats suggèrent que le LPA, dans un microenvironnement
pro-inflammatoire activé par le TNF-α, serait capable de stimuler la sécrétion de BLC et
contribuer ainsi à augmenter ou prolonger l’inflammation via l’activation des cellules
dendritiques.
IP-10/CXCL10 (« Interferon-gamma inducible protein-10 ») apparait dans la poche d’air
lorsque celle-ci subit un prétraitement au TNF-α 16h avant l’injection de LPA, et ce, aussi
bien dans l’analyse qualitative que quantitative. Cette chimiokine a été retrouvée en
quantité plus importante dans les liquides synoviaux de patients atteints de PR comparé à
ceux atteints d’ostéoarthrite (OA) [211, 212]. Les niveaux d’IP-10 retrouvés dans le serum
de patients positifs pour le facteur rhumatoïde (RF), qui est un marqueur de la PR, étaient
plus élevés que ceux des patients atteints d’OA [212]. L’IP-10 est aussi capable de recruter
les cellules Th1 dans les tissus synoviaux [211]. Pour toutes ces raisons, nous avons
suggéré que le LPA agirait sur ses récepteurs LPA1 et LPA3 pour relarguer des ligands des
CXCR2 (tels que KC et MIP-2) ou d’autres médiateurs pro-inflammatoires (IL-6, IP-10,

98
TREM-1 et CXCL13), qui eux aussi vont stimuler la migration des leucocytes dans la
poche d’air, bien que nos résultats ne permettent pas de déduire quel type cellulaire dans la
poche d’air contribue à l’expression de ces cytokines/chimiokines. Le LPA est aussi
capable d’induire l’expression d’une variété de gènes dans les fibroblastes de souris,
incluant des facteurs de transcription, des cytokines, des intégrines et des chimiokines
[213], contribuant à la mise en place d’un microenvironnement inflammatoire qui favorise
le recrutement des leucocytes.
Un autre aspect de mon travail de maîtrise était d’étudier l’effet de l’hypoxie sur des FLS
provenant de patients atteints de PR. Depuis de nombreuses années, on reconnait
l’importance de l’hypoxie dans la PR puisque le milieu hypoxique de la membrane et du
liquide synovial amplifie le phénomène inflammatoire [254]. En effet, les liquides
synoviaux provenant de patients atteints de PR sont connus pour avoir un taux très faible en
oxygène [161]. L’hypoxie peut activer la synthèse d’une variété de cytokines pro-
inflammatoires, incluant l’IL-8 [255]. Lorsque l’on provoque une hypoxie chimique au
CoCl2 sur des FLS de patients atteints de PR ou des FLS normaux, ceux-ci sécrètent de
l’IL-8. Étonnamment, la quantité d’IL-8 sécrétée par les FLS de PR est inférieure à celle
des FLS normaux. Ceci suggère que la voie moléculaire impliquée dans le relargage d’IL-8
est altérée chez les FLS de PR et par conséquent, que le phénotype des FLS de PR est
différent des synoviocytes normaux. D’importants changements morphologiques des
cellules de la membrane synoviale ont déjà été démontrés dans le cas de PR [256], ce qui
expliquerait pourquoi les FLS normaux et les FLS de la PR présentent des différences.
Afin de comprendre les différences observées entre les FLS normaux et les FLS de PR en
réponse à l’hypoxie au CoCl2, nous avons étudié la voie de signalisation du S1P puisque
l’hypoxie active la sphingosine kinase-1 [164]. Dans les FLS normaux, la sécrétion d’IL-8
due au CoCl2 est dépendante des récepteurs S1P2 et S1P3. Ceci suggère que les FLS
normaux synthétisent et relarguent du S1P en réponse à l’hypoxie. Cependant, la sécrétion
d’IL-8 provoquée par le CoCl2 chez des FLS de PR n’implique pas les récepteurs au S1P,
ce qui indique soit que le S1P n’est plus synthétisé par les cellules suite à la stimulation
hypoxique, soit que certaines enzymes dégradent le S1P synthétisé et empêche le S1P
d’agir. Suite à ces observations, nous avons regardé l’expression de l’ARNm des

99
sphingosines kinases 1 et 2 chez les FLS normaux et FLS arthritiques afin de démontrer
une éventuelle différence d’expression qui nous permettrait de confirmer notre hypothèse et
de l’expliquer. Nous avons démontré que l’expression de l’ARNm de la sphingosine kinase
1 était augmentée dans les FLS normaux mais pas dans les FLS de PR, suggérant que le
S1P ne serait probablement pas produit dans les FLS de PR. En ce qui concerne la
sphingosine kinase-2, nous n’avons détecté aucune différence entre les deux types de
synoviocytes (résultats non montrés), ce qui n’est pas surprenant car une étude a démontré
que seule l’expression de la sphingosine kinase-1 était augmentée lors d’une hypoxie [164].
L’incapacité des FLS de PR à libérer de grandes quantités d'IL-8 dans un environnement
hypoxique par rapport à celles produites par les FLS normaux est modifiée par l'ajout de
S1P. En effet, on observe un effet synergique entre l’hypoxie au CoCl2 et le S1P sur la
sécrétion d’IL-8 par les FLS, révélant la présence de récepteurs au S1P fonctionnels dans
les FLS de PR. Nos résultats suggèrent qu’une étape dans la synthèse et/ou la sécrétion de
S1P dans des conditions hypoxiques est dysfonctionnelle dans les FLS de PR.
4.2. Perspectives Dans la première partie de mon travail de maîtrise, nous avons donc réussi à démontrer que
le LPA agissait sur ses récepteurs LPA1 et LPA3 pour relarguer des ligands des CXCR2
capables de recruter les leucocytes vers un site inflammatoire. Dans un premier temps, nous
pourrions effectuer une mesure quantitative de l’IL-16, CXCL13, et TREM-1, qui sont des
cytokines importantes dans la PR et que nous avons détecté qualitativement dans les
lavages des poches d’air. Pour confirmer nos observations, nous pourrions par la suite
étudier les effets du LPA dans des souris K.O. pour LPA1 [257] et CXCR2 [258]. Aussi,
afin de vérifier si l’on arrive à diminuer une inflammation généralisée, on pourrait utiliser
des antagonistes des récepteurs LPA1/LPA3 et CXCR2 dans un modèle d’arthrite. Enfin,
dans le but de soutenir notre hypothèse, nous pourrions tenter d’identifier précisément les
cellules mise en jeu dans le recrutement des leucocytes induit par le LPA. Parallèlement, il
serait intéressant de mieux cerner les voies de signalisations utilisées par les récepteurs au
LPA, ainsi que de comprendre les conséquences du prétraitement au TNF-α sur les cellules,
afin de cibler spécifiquement les voies de signalisation à bloquer.

100
En ce qui concerne notre étude sur le S1P, comme nous avons récemment observé des
différences d’expression de la sphingosine kinase-1 chez les FLS de la PR et les FLS
normaux, nous pourrions essayer de détecter la présence de cette enzyme au niveau
protéique, suite à la stimulation au CoCl2 en utilisant un anticorps anti-sphingosine kinase-
1. De plus, nous pourrions confirmer que le CoCl2 reproduit bien les conditions de
l’hypoxie, en répétant nos expériences dans des conditions plus physiologiques pour
produire une hypoxie ambiante réelle (par exemple une chambre à hypoxie), c’est-à-dire en
manipulant les cellules dans un environnement pauvre en oxygène. Afin de mieux
comprendre les mécanismes qui diffèrent entre les FLS normaux et les FLS de PR, il serait
aussi intéressant de regarder l’expression de SPP2, une sphingosine phosphate phosphatase
qui dégrade le S1P et dont l’expression est induite pendant la réponse inflammatoire [232].
Enfin, nous pourrions mesurer la quantité de S1P sécrété par des FLS normaux ou de PR
dans un environnement hypoxique.
4.3. Conclusions Nos expériences ont souligné l’importance, au moins in vitro, des lysophospholipides LPA
et S1P dans la pathologie de la PR. Bloquer les fonctions des récepteurs au LPA, au S1P de
façon sélective, ou les enzymes qui produisent ces lysophospholipides (sphingosine kinase-
1, autotaxine) pourrait être envisagé pour traiter les maladies auto-immunes telles la
polyarthrite rhumatoïde. Depuis les dix dernières années, on ne cesse d’accroître nos
connaissances concernant ces importants lysophospholipides, dans le but de pouvoir un
jour proposer encore un nouveau traitement qui permettrait de diminuer l’inflammation, et
par conséquent, les douleurs des patients atteints de PR. Non seulement l’inflammation de
la PR est concernée par l’implication de ces lysophospholipides, mais ceux-ci contribuent
tout autant aux inflammations associées aux cancers et au développement des tumeurs et
des métastases [87]. Puisque certaines cibles thérapeutiques sont communes à la PR et aux
cancers, peut-être que ces recherches pourraient, par la même occasion, contribuer à
soigner les patients cancéreux ?

101
Bibliographie
1. Ford, D.K. and M. Schulzer, Synovial lymphocytes indicate "bacterial" agents may cause some cases of rheumatoid arthritis. J Rheumatol, 1994. 21(8): p. 1447-9.
2. Thomas, R., M. Turner, and A.P. Cope, High avidity autoreactive T cells with a low signalling capacity through the T-cell receptor: central to rheumatoid arthritis pathogenesis? Arthritis Res Ther, 2008. 10(4): p. 210.
3. Rannou, F., et al., Cartilage breakdown in rheumatoid arthritis. Joint Bone Spine, 2006. 73(1): p. 29-36.
4. Mor, A., S.B. Abramson, and M.H. Pillinger, The fibroblast-like synovial cell in rheumatoid arthritis: a key player in inflammation and joint destruction. Clin Immunol, 2005. 115(2): p. 118-28.
5. Doan, T. and E. Massarotti, Rheumatoid arthritis: an overview of new and emerging therapies. J Clin Pharmacol, 2005. 45(7): p. 751-62.
6. Venkateshan, S.P., et al., Efficacy of biologicals in the treatment of rheumatoid arthritis. a meta-analysis. Pharmacology, 2009. 83(1): p. 1-9.
7. Bowes, J. and A. Barton, Recent advances in the genetics of RA susceptibility. Rheumatology (Oxford), 2008. 47(4): p. 399-402.
8. Strietholt, S., et al., Epigenetic modifications in rheumatoid arthritis. Arthritis Res Ther, 2008. 10(5): p. 219.
9. Edwards, C.J. and C. Cooper, Early environmental factors and rheumatoid arthritis. Clin Exp Immunol, 2006. 143(1): p. 1-5.
10. Kobayashi, S., et al., Molecular aspects of rheumatoid arthritis: role of environmental factors. FEBS J, 2008. 275(18): p. 4456-62.
11. Cutolo, M., et al., Androgens and estrogens modulate the immune and inflammatory responses in rheumatoid arthritis. Ann N Y Acad Sci, 2002. 966: p. 131-42.
12. Da Silva, J.A. and G.M. Hall, The effects of gender and sex hormones on outcome in rheumatoid arthritis. Baillieres Clin Rheumatol, 1992. 6(1): p. 196-219.
13. Gerosa, M., et al., Rheumatoid arthritis: a female challenge. Womens Health (Lond Engl), 2008. 4(2): p. 195-201.
14. Thierry, C.B.I.R., Etude phénotypique, génotypique et fonctionnelle de lymphocytes T à récepteur gamma/delta dans la polyarthrite rhumatoïde. Analyse comparative avec les lymphocytes T alpha/beta = Phenotypic, genotypic and functional study of lymphocytes T gamma/delta in rheumatoid arthritis. Comparative analysis with lymphocytes T alpha/beta. 1992.
15. Puttick, A.H., et al., Genes associated with rheumatoid arthritis and mild inflammatory arthritis. II. Association of HLA with complement C3 and immunoglobulin Gm allotypes. Ann Rheum Dis, 1990. 49(4): p. 225-8.
16. Zhang, H., et al., Elimination of rheumatoid synovium in situ using a Fas ligand 'gene scalpel'. Arthritis Res Ther, 2005. 7(6): p. R1235-43.
17. de Vries, N. and P.P. Tak, The response to anti-TNF-alpha treatment: gene regulation at the bedside. Rheumatology (Oxford), 2005. 44(6): p. 705-7.
18. Meyer, J.M., et al., HLA-DRB1 genotype influences risk for and severity of rheumatoid arthritis. J Rheumatol, 1999. 26(5): p. 1024-34.
19. Wordsworth, B.P. and J.I. Bell, The immunogenetics of rheumatoid arthritis. Springer Semin Immunopathol, 1992. 14(1): p. 59-78.

102
20. Brenner, M., et al., The non-MHC quantitative trait locus Cia5 contains three major arthritis genes that differentially regulate disease severity, pannus formation, and joint damage in collagen- and pristane-induced arthritis. J Immunol, 2005. 174(12): p. 7894-903.
21. Laragione, T., et al., Cia5d regulates a new fibroblast-like synoviocyte invasion-associated gene expression signature. Arthritis Res Ther, 2008. 10(4): p. R92.
22. Laragione, T., et al., The arthritis severity locus Cia5d is a novel genetic regulator of the invasive properties of synovial fibroblasts. Arthritis Rheum, 2008. 58(8): p. 2296-306.
23. Barland, P., A.B. Novikoff, and D. Hamerman, Electron microscopy of the human synovial membrane. J Cell Biol, 1962. 14: p. 207-20.
24. Hamilton, J.A. and J. Slywka, Stimulation of human synovial fibroblast plasminogen activator production by mononuclear cell supernatants. J Immunol, 1981. 126(3): p. 851-5.
25. Iwanaga, T., et al., Morphology and functional roles of synoviocytes in the joint. Arch Histol Cytol, 2000. 63(1): p. 17-31.
26. Noss, E.H. and M.B. Brenner, The role and therapeutic implications of fibroblast-like synoviocytes in inflammation and cartilage erosion in rheumatoid arthritis. Immunol Rev, 2008. 223: p. 252-70.
27. Kramer, I., et al., Rheumatoid arthritis: targeting the proliferative fibroblasts. Prog Cell Cycle Res, 2003. 5: p. 59-70.
28. Reparon-Schuijt, C.C., et al., Regulation of synovial B cell survival in rheumatoid arthritis by vascular cell adhesion molecule 1 (CD106) expressed on fibroblast-like synoviocytes. Arthritis Rheum, 2000. 43(5): p. 1115-21.
29. Bombara, M.P., et al., Cell contact between T cells and synovial fibroblasts causes induction of adhesion molecules and cytokines. J Leukoc Biol, 1993. 54(5): p. 399-406.
30. Tran, C.N., et al., Molecular interactions between T cells and fibroblast-like synoviocytes: role of membrane tumor necrosis factor-alpha on cytokine-activated T cells. Am J Pathol, 2007. 171(5): p. 1588-98.
31. Blue, M.L., et al., Interacting monocytes and synoviocytes induce adhesion molecules by a cytokine-regulated process. Lymphokine Cytokine Res, 1993. 12(4): p. 213-8.
32. Hanyuda, M., et al., Activated leucocytes express and secrete macrophage inflammatory protein-1alpha upon interaction with synovial fibroblasts of rheumatoid arthritis via a beta2-integrin/ICAM-1 mechanism. Rheumatology (Oxford), 2003. 42(11): p. 1390-7.
33. Filer, A., et al., Differential survival of leukocyte subsets mediated by synovial, bone marrow, and skin fibroblasts: site-specific versus activation-dependent survival of T cells and neutrophils. Arthritis Rheum, 2006. 54(7): p. 2096-108.
34. Bradfield, P.F., et al., Rheumatoid fibroblast-like synoviocytes overexpress the chemokine stromal cell-derived factor 1 (CXCL12), which supports distinct patterns and rates of CD4+ and CD8+ T cell migration within synovial tissue. Arthritis Rheum, 2003. 48(9): p. 2472-82.
35. Kasama, T., et al., Vascular endothelial growth factor expression by activated synovial leukocytes in rheumatoid arthritis: critical involvement of the interaction with synovial fibroblasts. Arthritis Rheum, 2001. 44(11): p. 2512-24.

103
36. Zhao, C., et al., Specific and overlapping sphingosine-1-phosphate receptor functions in human synoviocytes: impact of TNF-alpha. J Lipid Res, 2008. 49(11): p. 2323-37.
37. Villiger, P.M., R. Terkeltaub, and M. Lotz, Production of monocyte chemoattractant protein-1 by inflamed synovial tissue and cultured synoviocytes. J Immunol, 1992. 149(2): p. 722-7.
38. Georganas, C., et al., Regulation of IL-6 and IL-8 expression in rheumatoid arthritis synovial fibroblasts: the dominant role for NF-kappa B but not C/EBP beta or c-Jun. J Immunol, 2000. 165(12): p. 7199-206.
39. Hashida, R., et al., Stimulation of prostaglandin synthesis in cultured rat synovial cells by a factor derived from polymorphonuclear leukocytes. Prostaglandins, 1984. 27(5): p. 697-709.
40. Buckley, C.D., Michael Mason prize essay 2003. Why do leucocytes accumulate within chronically inflamed joints? Rheumatology (Oxford), 2003. 42(12): p. 1433-44.
41. Halbwachs-Mecarelli, L., Neutrophiles dans l’hypersensibilité. Revue Française d'Allergologie et d'Immunologie Clinique, 2004. 45(1): p. 68-73.
42. Ethuin, F., S. Chollet-Martin, and L. Jacob, [A new role for neutrophils during sepsis: target and source of interleukin-12]. Ann Fr Anesth Reanim, 2004. 23(2): p. 132-7.
43. Kobayashi, Y., The role of chemokines in neutrophil biology. Front Biosci, 2008. 13: p. 2400-7.
44. Mohamed Bouaouina, L.H.-M., Activation des intégrines et adhérence des leucocytes et des plaquettes. Hématologie, 2006. 12: p. 21-33.
45. Chakravarti, A., I. Allaeys, and P.E. Poubelle, [Neutrophils and immunity: is it innate or acquired?]. Med Sci (Paris), 2007. 23(10): p. 862-7.
46. Weissmann, G. and H. Korchak, Rheumatoid arthritis. The role of neutrophil activation. Inflammation, 1984. 8 Suppl: p. S3-14.
47. Mohr, W., [Polymorphonuclear granulocytes in rheumatic tissue destruction VIII. Considerations on the inflammatory cartilage destruction in chronic arthritides in comparison with liver injuries by PMN's]. Z Rheumatol, 2003. 62(6): p. 539-46; discussion 547.
48. Brown, K.A., The polymorphonuclear cell in rheumatoid arthritis. Br J Rheumatol, 1988. 27(2): p. 150-5.
49. Cross, A., et al., Neutrophil apoptosis in rheumatoid arthritis is regulated by local oxygen tensions within joints. J Leukoc Biol, 2006. 80(3): p. 521-8.
50. Quayle, J.A., et al., Interleukin-1 expression by neutrophils in rheumatoid arthritis. Ann Rheum Dis, 1995. 54(11): p. 930-3.
51. Ottonello, L., et al., Synovial fluid from patients with rheumatoid arthritis inhibits neutrophil apoptosis: role of adenosine and proinflammatory cytokines. Rheumatology (Oxford), 2002. 41(11): p. 1249-60.
52. Chakravarti, A., et al., Surface RANKL of Toll-like receptor 4-stimulated human neutrophils activates osteoclastic bone resorption. Blood, 2009. 114(8): p. 1633-44.
53. Lee, A.N., C.E. Beck, and M. Hall, Rheumatoid factor and anti-CCP autoantibodies in rheumatoid arthritis: a review. Clin Lab Sci, 2008. 21(1): p. 15-8.

104
54. Miossec, P., Dynamic interactions between T cells and dendritic cells and their derived cytokines/chemokines in the rheumatoid synovium. Arthritis Res Ther, 2008. 10 Suppl 1: p. S2.
55. Kaplan, C. and A. Finnegan, Osteoclasts, pro-inflammatory cytokines, RANK-L and bone remodeling in rheumatoid arthritis. Front Biosci, 2003. 8: p. d1018-29.
56. De Gendt, C.M., et al., Relationship between interleukin-8 and neutrophil adhesion molecules in rheumatoid arthritis. Rheumatol Int, 1996. 16(4): p. 169-73.
57. Kobayashi, Y., Neutrophil infiltration and chemokines. Crit Rev Immunol, 2006. 26(4): p. 307-16.
58. Alvaro-Gracia, J.M., N.J. Zvaifler, and G.S. Firestein, Cytokines in chronic inflammatory arthritis. V. Mutual antagonism between interferon-gamma and tumor necrosis factor-alpha on HLA-DR expression, proliferation, collagenase production, and granulocyte macrophage colony-stimulating factor production by rheumatoid arthritis synoviocytes. J Clin Invest, 1990. 86(6): p. 1790-8.
59. Grimbacher, B., et al., TNF-alpha induces the transcription factor Egr-1, pro-inflammatory cytokines and cell proliferation in human skin fibroblasts and synovial lining cells. Rheumatol Int, 1998. 17(5): p. 185-92.
60. Zhao, C., et al., Regulation of lysophosphatidic acid receptor expression and function in human synoviocytes: implications for rheumatoid arthritis? Mol Pharmacol, 2008. 73(2): p. 587-600.
61. Chen, Q., et al., Tumor necrosis factor-alpha protects synovial cells from nitric oxide induced apoptosis through phosphoinositide 3-kinase Akt signal transduction. J Rheumatol, 2006. 33(6): p. 1061-8.
62. Kowanko, I.C. and A. Ferrante, Adhesion and TNF priming in neutrophil-mediated cartilage damage. Clin Immunol Immunopathol, 1996. 79(1): p. 36-42.
63. Lokuta, M.A. and A. Huttenlocher, TNF-alpha promotes a stop signal that inhibits neutrophil polarization and migration via a p38 MAPK pathway. J Leukoc Biol, 2005. 78(1): p. 210-9.
64. Assi, L.K., et al., Tumor necrosis factor alpha activates release of B lymphocyte stimulator by neutrophils infiltrating the rheumatoid joint. Arthritis Rheum, 2007. 56(6): p. 1776-86.
65. Choy, E.H. and G.S. Panayi, Cytokine pathways and joint inflammation in rheumatoid arthritis. N Engl J Med, 2001. 344(12): p. 907-16.
66. MacNaul, K.L., et al., Discoordinate expression of stromelysin, collagenase, and tissue inhibitor of metalloproteinases-1 in rheumatoid human synovial fibroblasts. Synergistic effects of interleukin-1 and tumor necrosis factor-alpha on stromelysin expression. J Biol Chem, 1990. 265(28): p. 17238-45.
67. Privat, S.J., A. Oo, and L.D. Waterbury, Production of interleukin-6, but not interleukin-8, induced by TNF-alpha or IL-1 beta in human fibroblast-like synoviocyte increases over cell passage. Proc West Pharmacol Soc, 2001. 44: p. 9-13.
68. Choi, H.M., et al., Differential effect of IL-1beta and TNF-alpha on the production of IL-6, IL-8 and PGE(2) in fibroblast-like synoviocytes and THP-1 macrophages. Rheumatol Int, 2009.
69. Nawroth, P.P., et al., Tumor necrosis factor/cachectin interacts with endothelial cell receptors to induce release of interleukin 1. J Exp Med, 1986. 163(6): p. 1363-75.

105
70. Haworth, C., et al., Expression of granulocyte-macrophage colony-stimulating factor in rheumatoid arthritis: regulation by tumor necrosis factor-alpha. Eur J Immunol, 1991. 21(10): p. 2575-9.
71. Koenders, M.I., L.A. Joosten, and W.B. van den Berg, Potential new targets in arthritis therapy: interleukin (IL)-17 and its relation to tumour necrosis factor and IL-1 in experimental arthritis. Ann Rheum Dis, 2006. 65 Suppl 3: p. iii29-33.
72. Rathanaswami, P., et al., Synergistic effect of interleukin-1 beta and tumor necrosis factor alpha on interleukin-8 gene expression in synovial fibroblasts. Evidence that interleukin-8 is the major neutrophil-activating chemokine released in response to monokine activation. Arthritis Rheum, 1993. 36(9): p. 1295-304.
73. Clark, I.A., How TNF was recognized as a key mechanism of disease. Cytokine Growth Factor Rev, 2007. 18(3-4): p. 335-43.
74. Feldmann, M. and R.N. Maini, Lasker Clinical Medical Research Award. TNF defined as a therapeutic target for rheumatoid arthritis and other autoimmune diseases. Nat Med, 2003. 9(10): p. 1245-50.
75. Kountz, D.S. and J.M. Von Feldt, Management of rheumatoid arthritis: A primary care perspective. J Fam Pract, 2007. 56(10 Suppl A): p. 59A-73A; quiz 74A.
76. Bongartz, T., et al., Anti-TNF antibody therapy in rheumatoid arthritis and the risk of serious infections and malignancies: systematic review and meta-analysis of rare harmful effects in randomized controlled trials. JAMA, 2006. 295(19): p. 2275-85.
77. Wolfe, F., et al., Tuberculosis infection in patients with rheumatoid arthritis and the effect of infliximab therapy. Arthritis Rheum, 2004. 50(2): p. 372-9.
78. Hochberg, M.C., et al., Comparison of the efficacy of the tumour necrosis factor alpha blocking agents adalimumab, etanercept, and infliximab when added to methotrexate in patients with active rheumatoid arthritis. Ann Rheum Dis, 2003. 62 Suppl 2: p. ii13-6.
79. Combe, B., Early rheumatoid arthritis: strategies for prevention and management. Best Pract Res Clin Rheumatol, 2007. 21(1): p. 27-42.
80. Gaffo, A., K.G. Saag, and J.R. Curtis, Treatment of rheumatoid arthritis. Am J Health Syst Pharm, 2006. 63(24): p. 2451-65.
81. Jalink, K., P.L. Hordijk, and W.H. Moolenaar, Growth factor-like effects of lysophosphatidic acid, a novel lipid mediator. Biochim Biophys Acta, 1994. 1198(2-3): p. 185-96.
82. Peyruchaud, O., Novel implications for lysophospholipids, lysophosphatidic acid and sphingosine 1-phosphate, as drug targets in cancer. Anticancer Agents Med Chem, 2009. 9(4): p. 381-91.
83. Rivera-Lopez, C.M., A.L. Tucker, and K.R. Lynch, Lysophosphatidic acid (LPA) and angiogenesis. Angiogenesis, 2008. 11(3): p. 301-10.
84. Lee, O.H., et al., Sphingosine 1-phosphate induces angiogenesis: its angiogenic action and signaling mechanism in human umbilical vein endothelial cells. Biochem Biophys Res Commun, 1999. 264(3): p. 743-50.
85. Claria, J., Regulation of cell proliferation and apoptosis by bioactive lipid mediators. Recent Pat Anticancer Drug Discov, 2006. 1(3): p. 369-82.
86. Dyatlovitskaya, E.V. and A.G. Kandyba, Role of biologically active sphingolipids in tumor growth. Biochemistry (Mosc), 2006. 71(1): p. 10-7.
87. Murph, M. and G.B. Mills, Targeting the lipids LPA and S1P and their signalling pathways to inhibit tumour progression. Expert Rev Mol Med, 2007. 9(28): p. 1-18.

106
88. Chun, J., et al., International Union of Pharmacology. XXXIV. Lysophospholipid receptor nomenclature. Pharmacol Rev, 2002. 54(2): p. 265-9.
89. Ishii, S., K. Noguchi, and K. Yanagida, Non-Edg family lysophosphatidic acid (LPA) receptors. Prostaglandins Other Lipid Mediat, 2009.
90. Tokumura, A., et al., Stimulatory effect of lysophosphatidic acids on uterine smooth muscles of non-pregant rats. Arch Int Pharmacodyn Ther, 1980. 245(1): p. 74-83.
91. Tokumura, A., et al., Lysophosphatidic acid-induced aggregation of human and feline platelets: structure-activity relationship. Biochem Biophys Res Commun, 1981. 99(2): p. 391-8.
92. van Corven, E.J., et al., Lysophosphatidate-induced cell proliferation: identification and dissection of signaling pathways mediated by G proteins. Cell, 1989. 59(1): p. 45-54.
93. Moolenaar, W.H., et al., Lysophosphatidic acid: a novel phospholipid with hormone- and growth factor-like activities. Cold Spring Harb Symp Quant Biol, 1992. 57: p. 163-7.
94. Panetti, T.S., Differential effects of sphingosine 1-phosphate and lysophosphatidic acid on endothelial cells. Biochim Biophys Acta, 2002. 1582(1-3): p. 190-6.
95. Ye, X., et al., Lysophosphatidic acid as a novel cell survival/apoptotic factor. Biochim Biophys Acta, 2002. 1585(2-3): p. 108-13.
96. Fuchs, B., et al., The phosphatidylcholine/lysophosphatidylcholine ratio in human plasma is an indicator of the severity of rheumatoid arthritis: investigations by 31P NMR and MALDI-TOF MS. Clin Biochem, 2005. 38(10): p. 925-33.
97. Mills, G.B. and W.H. Moolenaar, The emerging role of lysophosphatidic acid in cancer. Nat Rev Cancer, 2003. 3(8): p. 582-91.
98. Pilquil, C., et al., Lipid phosphate phosphatase-1 dephosphorylates exogenous lysophosphatidate and thereby attenuates its effects on cell signalling. Prostaglandins Other Lipid Mediat, 2001. 64(1-4): p. 83-92.
99. Tomsig, J.L., et al., Lipid phosphate phosphohydrolase type 1 (LPP1) degrades extracellular lysophosphatidic acid in vivo. Biochem J, 2009. 419(3): p. 611-8.
100. Moolenaar, W.H. and E.J. van Corven, Growth factor-like action of lysophosphatidic acid: mitogenic signalling mediated by G proteins. Ciba Found Symp, 1990. 150: p. 99-106; discussion 106-11.
101. Yanagida, K., et al., Identification and characterization of a novel lysophosphatidic acid receptor, p2y5/LPA6. J Biol Chem, 2009. 284(26): p. 17731-41.
102. An, S., et al., Characterization of a novel subtype of human G protein-coupled receptor for lysophosphatidic acid. J Biol Chem, 1998. 273(14): p. 7906-10.
103. Bandoh, K., et al., Molecular cloning and characterization of a novel human G-protein-coupled receptor, EDG7, for lysophosphatidic acid. J Biol Chem, 1999. 274(39): p. 27776-85.
104. Hecht, J.H., et al., Ventricular zone gene-1 (vzg-1) encodes a lysophosphatidic acid receptor expressed in neurogenic regions of the developing cerebral cortex. J Cell Biol, 1996. 135(4): p. 1071-83.
105. An, S., et al., Molecular cloning of the human Edg2 protein and its identification as a functional cellular receptor for lysophosphatidic acid. Biochem Biophys Res Commun, 1997. 231(3): p. 619-22.
106. Estivill-Torrus, G., et al., Absence of LPA1 signaling results in defective cortical development. Cereb Cortex, 2008. 18(4): p. 938-50.

107
107. Ohuchi, H., et al., Expression patterns of the lysophospholipid receptor genes during mouse early development. Dev Dyn, 2008. 237(11): p. 3280-94.
108. Sautin, Y.Y., et al., Hepatic oval (stem) cell expression of endothelial differentiation gene receptors for lysophosphatidic acid in mouse chronic liver injury. J Hematother Stem Cell Res, 2002. 11(4): p. 643-9.
109. Pradere, J.P., et al., LPA1 receptor activation promotes renal interstitial fibrosis. J Am Soc Nephrol, 2007. 18(12): p. 3110-8.
110. Guo, R., et al., Expression and function of lysophosphatidic acid LPA1 receptor in prostate cancer cells. Endocrinology, 2006. 147(10): p. 4883-92.
111. Moller, T., et al., Expression and function of lysophosphatidic acid receptors in cultured rodent microglial cells. J Biol Chem, 2001. 276(28): p. 25946-52.
112. Gobeil, F., Jr., et al., Modulation of pro-inflammatory gene expression by nuclear lysophosphatidic acid receptor type-1. J Biol Chem, 2003. 278(40): p. 38875-83.
113. Hama, K., et al., Lysophosphatidic acid and autotaxin stimulate cell motility of neoplastic and non-neoplastic cells through LPA1. J Biol Chem, 2004. 279(17): p. 17634-9.
114. Takuwa, Y., N. Takuwa, and N. Sugimoto, The Edg family G protein-coupled receptors for lysophospholipids: their signaling properties and biological activities. J Biochem, 2002. 131(6): p. 767-71.
115. S. M. Harrisond, C.R., G. Browne, J. T. Brownb, J. E. Cluderaya, B. Crookb, C. H. Daviesb, L. A. Dawsona, E. Graud, C. Heidbredera, P. Hemmatia, G. Hervieub, A. Howarthd, Z. A. Hughesa, A. J. Hunterb, J. Latchamc, S. Pickeringd, P. Pughb, D. C. Rogersb, C. S. Shilliama and P. R. Maycox, LPA1 receptor-deficient mice have phenotypic changes observed in psychiatric disease. Molecular and cellular neurosciences, 2003. 24(4): p. 1170-1179.
116. Im, D.S., et al., Molecular cloning and characterization of a lysophosphatidic acid receptor, Edg-7, expressed in prostate. Mol Pharmacol, 2000. 57(4): p. 753-9.
117. Hama, K., et al., Embryo spacing and implantation timing are differentially regulated by LPA3-mediated lysophosphatidic acid signaling in mice. Biol Reprod, 2007. 77(6): p. 954-9.
118. Shan, T.L., et al., Full-length coding sequences, polymorphism and chromosomal localizations of the porcine EDG4 and EDG7 genes. Mol Biol Rep, 2009. 36(4): p. 751-6.
119. Eichholtz, T., et al., The bioactive phospholipid lysophosphatidic acid is released from activated platelets. Biochem J, 1993. 291 ( Pt 3): p. 677-80.
120. Hosogaya, S., et al., Measurement of plasma lysophosphatidic acid concentration in healthy subjects: strong correlation with lysophospholipase D activity. Ann Clin Biochem, 2008. 45(Pt 4): p. 364-8.
121. Sugiura, T., et al., Lysophosphatidic acid, a growth factor-like lipid, in the saliva. J Lipid Res, 2002. 43(12): p. 2049-55.
122. Xu, Y., et al., Lysophosphatidic acid as a potential biomarker for ovarian and other gynecologic cancers. JAMA, 1998. 280(8): p. 719-23.
123. Nochi, H., et al., Stimulatory role of lysophosphatidic acid in cyclooxygenase-2 induction by synovial fluid of patients with rheumatoid arthritis in fibroblast-like synovial cells. J Immunol, 2008. 181(7): p. 5111-9.
124. Zhang, H., et al., Sphingosine-1-phosphate, a novel lipid, involved in cellular proliferation. J Cell Biol, 1991. 114(1): p. 155-67.

108
125. Spiegel, S., et al., Sphingosine-1-phosphate, a novel second messenger involved in cell growth regulation and signal transduction, affects growth and invasiveness of human breast cancer cells. Breast Cancer Res Treat, 1994. 31(2-3): p. 337-48.
126. Kitano, M., et al., Sphingosine 1-phosphate/sphingosine 1-phosphate receptor 1 signaling in rheumatoid synovium: regulation of synovial proliferation and inflammatory gene expression. Arthritis Rheum, 2006. 54(3): p. 742-53.
127. Rivera, J., R.L. Proia, and A. Olivera, The alliance of sphingosine-1-phosphate and its receptors in immunity. Nat Rev Immunol, 2008. 8(10): p. 753-63.
128. Pyne, S., K.C. Kong, and P.I. Darroch, Lysophosphatidic acid and sphingosine 1-phosphate biology: the role of lipid phosphate phosphatases. Semin Cell Dev Biol, 2004. 15(5): p. 491-501.
129. Mandala, S.M., et al., Molecular cloning and characterization of a lipid phosphohydrolase that degrades sphingosine-1- phosphate and induces cell death. Proc Natl Acad Sci U S A, 2000. 97(14): p. 7859-64.
130. Oskouian, B., et al., Regulation of sphingosine-1-phosphate lyase gene expression by members of the GATA family of transcription factors. J Biol Chem, 2005. 280(18): p. 18403-10.
131. Spiegel, S. and S. Milstien, Sphingosine-1-phosphate: an enigmatic signalling lipid. Nat Rev Mol Cell Biol, 2003. 4(5): p. 397-407.
132. Kohama, T., et al., Molecular cloning and functional characterization of murine sphingosine kinase. J Biol Chem, 1998. 273(37): p. 23722-8.
133. Liu, H., et al., Molecular cloning and functional characterization of a novel mammalian sphingosine kinase type 2 isoform. J Biol Chem, 2000. 275(26): p. 19513-20.
134. Lai, W.Q., et al., Anti-inflammatory effects of sphingosine kinase modulation in inflammatory arthritis. J Immunol, 2008. 181(11): p. 8010-7.
135. Limaye, V., et al., Chronic increases in sphingosine kinase-1 activity induce a pro-inflammatory, pro-angiogenic phenotype in endothelial cells. Cell Mol Biol Lett, 2009. 14(3): p. 424-41.
136. Lai, W.Q., et al., Distinct roles of sphingosine kinase 1 and 2 in murine collagen-induced arthritis. J Immunol, 2009. 183(3): p. 2097-103.
137. Maceyka, M., et al., SphK1 and SphK2, sphingosine kinase isoenzymes with opposing functions in sphingolipid metabolism. J Biol Chem, 2005. 280(44): p. 37118-29.
138. Kamada, K., et al., Expression of sphingosine kinase 2 in synovial fibroblasts of rheumatoid arthritis contributing to apoptosis by a sphingosine analogue, FTY720. Pathol Int, 2009. 59(6): p. 382-9.
139. Liu, H., et al., Sphingosine kinase type 2 is a putative BH3-only protein that induces apoptosis. J Biol Chem, 2003. 278(41): p. 40330-6.
140. Sanchez, T. and T. Hla, Structural and functional characteristics of S1P receptors. J Cell Biochem, 2004. 92(5): p. 913-22.
141. Okamoto, H., et al., EDG1 is a functional sphingosine-1-phosphate receptor that is linked via a Gi/o to multiple signaling pathways, including phospholipase C activation, Ca2+ mobilization, Ras-mitogen-activated protein kinase activation, and adenylate cyclase inhibition. J Biol Chem, 1998. 273(42): p. 27104-10.
142. Lee, M.J., et al., Vascular endothelial cell adherens junction assembly and morphogenesis induced by sphingosine-1-phosphate. Cell, 1999. 99(3): p. 301-12.

109
143. Yonesu, K., et al., Involvement of sphingosine-1-phosphate and S1P1 in angiogenesis: analyses using a new S1P1 antagonist of non-sphingosine-1-phosphate analog. Biochem Pharmacol, 2009. 77(6): p. 1011-20.
144. Sugimoto, N., et al., Inhibitory and stimulatory regulation of Rac and cell motility by the G12/13-Rho and Gi pathways integrated downstream of a single G protein-coupled sphingosine-1-phosphate receptor isoform. Mol Cell Biol, 2003. 23(5): p. 1534-45.
145. Goparaju, S.K., et al., The S1P2 receptor negatively regulates platelet-derived growth factor-induced motility and proliferation. Mol Cell Biol, 2005. 25(10): p. 4237-49.
146. Inoki, I., et al., Negative regulation of endothelial morphogenesis and angiogenesis by S1P2 receptor. Biochem Biophys Res Commun, 2006. 346(1): p. 293-300.
147. Waeber, C., N. Blondeau, and S. Salomone, Vascular sphingosine-1-phosphate S1P1 and S1P3 receptors. Drug News Perspect, 2004. 17(6): p. 365-82.
148. Kohno, T., et al., Sphingosine 1-phosphate promotes cell migration through the activation of Cdc42 in Edg-6/S1P4-expressing cells. Genes Cells, 2003. 8(8): p. 685-97.
149. Niedernberg, A., et al., Comparative analysis of human and rat S1P(5) (edg8): differential expression profiles and sensitivities to antagonists. Biochem Pharmacol, 2002. 64(8): p. 1243-50.
150. Toman, R.E. and S. Spiegel, Lysophospholipid receptors in the nervous system. Neurochem Res, 2002. 27(7-8): p. 619-27.
151. Novgorodov, A.S., et al., Activation of sphingosine-1-phosphate receptor S1P5 inhibits oligodendrocyte progenitor migration. FASEB J, 2007. 21(7): p. 1503-14.
152. Jaillard, C., et al., Edg8/S1P5: an oligodendroglial receptor with dual function on process retraction and cell survival. J Neurosci, 2005. 25(6): p. 1459-69.
153. Ishii, I., et al., Lysophospholipid receptors: signaling and biology. Annu Rev Biochem, 2004. 73: p. 321-54.
154. Fischer, L.G., et al., Local anesthetics attenuate lysophosphatidic acid-induced priming in human neutrophils. Anesth Analg, 2001. 92(4): p. 1041-7.
155. Willoughby, D.A., et al., The use of the air pouch to study experimental synovitis and cartilage breakdown. Biomed Pharmacother, 1986. 40(2): p. 45-9.
156. Edwards, J.C., A.D. Sedgwick, and D.A. Willoughby, The formation of a structure with the features of synovial lining by subcutaneous injection of air: an in vivo tissue culture system. J Pathol, 1981. 134(2): p. 147-56.
157. Huber, L.C., et al., Synovial fibroblasts: key players in rheumatoid arthritis. Rheumatology (Oxford), 2006. 45(6): p. 669-75.
158. Krey, P.R. and D.A. Bailen, Synovial fluid leukocytosis. A study of extremes. Am J Med, 1979. 67(3): p. 436-42.
159. Nishimura, A., et al., Attenuation of monosodium urate crystal-induced arthritis in rabbits by a neutralizing antibody against interleukin-8. J Leukoc Biol, 1997. 62(4): p. 444-9.
160. Jackson, J.R., et al., Expression of vascular endothelial growth factor in synovial fibroblasts is induced by hypoxia and interleukin 1beta. J Rheumatol, 1997. 24(7): p. 1253-9.
161. Lund-Olesen, K., Oxygen tension in synovial fluids. Arthritis Rheum, 1970. 13(6): p. 769-76.

110
162. Ghezzi, P., et al., Hypoxia increases production of interleukin-1 and tumor necrosis factor by human mononuclear cells. Cytokine, 1991. 3(3): p. 189-94.
163. Ahn, J.K., et al., Role of hypoxia-inducible factor-1alpha in hypoxia-induced expressions of IL-8, MMP-1 and MMP-3 in rheumatoid fibroblast-like synoviocytes. Rheumatology (Oxford), 2008. 47(6): p. 834-9.
164. Anelli, V., et al., Sphingosine kinase 1 is up-regulated during hypoxia in U87MG glioma cells. Role of hypoxia-inducible factors 1 and 2. J Biol Chem, 2008. 283(6): p. 3365-75.
165. Schnitzer, S.E., et al., Hypoxia enhances sphingosine kinase 2 activity and provokes sphingosine-1-phosphate-mediated chemoresistance in A549 lung cancer cells. Mol Cancer Res, 2009. 7(3): p. 393-401.
166. Firestein, G.S., Evolving concepts of rheumatoid arthritis. Nature, 2003. 423(6937): p. 356-61.
167. Baker, M.S., S.P. Green, and D.A. Lowther, Changes in the viscosity of hyaluronic acid after exposure to a myeloperoxidase-derived oxidant. Arthritis Rheum, 1989. 32(4): p. 461-7.
168. Tak, P.P. and B. Bresnihan, The pathogenesis and prevention of joint damage in rheumatoid arthritis: advances from synovial biopsy and tissue analysis. Arthritis Rheum, 2000. 43(12): p. 2619-33.
169. Cross, A., et al., Neutrophil gene expression in rheumatoid arthritis. Pathophysiology, 2005. 12(3): p. 191-202.
170. Issekutz, A.C. and T.B. Issekutz, Quantitation and kinetics of polymorphonuclear leukocyte and lymphocyte accumulation in joints during adjuvant arthritis in the rat. Lab Invest, 1991. 64(5): p. 656-63.
171. Wipke, B.T. and P.M. Allen, Essential role of neutrophils in the initiation and progression of a murine model of rheumatoid arthritis. J Immunol, 2001. 167(3): p. 1601-8.
172. Feldmann, M., et al., Cytokine production in the rheumatoid joint: implications for treatment. Ann Rheum Dis, 1990. 49 Suppl 1: p. 480-6.
173. Feldmann, M., F.M. Brennan, and R.N. Maini, Rheumatoid arthritis. Cell, 1996. 85(3): p. 307-10.
174. Bacon, K.B. and J.J. Oppenheim, Chemokines in disease models and pathogenesis. Cytokine Growth Factor Rev, 1998. 9(2): p. 167-73.
175. Rollins, B.J., Chemokines. Blood, 1997. 90(3): p. 909-28. 176. Bozic, C.R., et al., Expression and biologic characterization of the murine
chemokine KC. J Immunol, 1995. 154(11): p. 6048-57. 177. Fahey, T.J., 3rd, et al., Cytokine production in a model of wound healing: the
appearance of MIP-1, MIP-2, cachectin/TNF and IL-1. Cytokine, 1990. 2(2): p. 92-9.
178. Huang, S., et al., Expression of macrophage inflammatory protein-2 and KC mRNA in pulmonary inflammation. Am J Pathol, 1992. 141(4): p. 981-8.
179. Rovai, L.E., H.R. Herschman, and J.B. Smith, The murine neutrophil-chemoattractant chemokines LIX, KC, and MIP-2 have distinct induction kinetics, tissue distributions, and tissue-specific sensitivities to glucocorticoid regulation in endotoxemia. J Leukoc Biol, 1998. 64(4): p. 494-502.
180. Safirstein, R., et al., Expression of cytokine-like genes JE and KC is increased during renal ischemia. Am J Physiol, 1991. 261(6 Pt 2): p. F1095-101.

111
181. Bozic, C.R., et al., The murine interleukin 8 type B receptor homologue and its ligands. Expression and biological characterization. J Biol Chem, 1994. 269(47): p. 29355-8.
182. Lee, J., et al., Chemokine binding and activities mediated by the mouse IL-8 receptor. J Immunol, 1995. 155(4): p. 2158-64.
183. Koch, A.E., Chemokines and their receptors in rheumatoid arthritis: future targets? Arthritis Rheum, 2005. 52(3): p. 710-21.
184. Moolenaar, W.H., Lysophosphatidic acid signalling. Curr Opin Cell Biol, 1995. 7(2): p. 203-10.
185. Tabata, K., et al., The orphan GPCR GPR87 was deorphanized and shown to be a lysophosphatidic acid receptor. Biochem Biophys Res Commun, 2007. 363(3): p. 861-6.
186. Murakami, M., et al., Identification of the orphan GPCR, P2Y(10) receptor as the sphingosine-1-phosphate and lysophosphatidic acid receptor. Biochem Biophys Res Commun, 2008. 371(4): p. 707-12.
187. Chettibi, S., et al., Effect of lysophosphatidic acid on motility, polarisation and metabolic burst of human neutrophils. FEMS Immunol Med Microbiol, 1994. 8(3): p. 271-81.
188. Rizza, C., et al., Lysophosphatidic acid as a regulator of endothelial/leukocyte interaction. Lab Invest, 1999. 79(10): p. 1227-35.
189. Kanda, H., et al., Autotaxin, an ectoenzyme that produces lysophosphatidic acid, promotes the entry of lymphocytes into secondary lymphoid organs. Nat Immunol, 2008. 9(4): p. 415-23.
190. Nakasaki, T., et al., Involvement of the lysophosphatidic acid-generating enzyme autotaxin in lymphocyte-endothelial cell interactions. Am J Pathol, 2008. 173(5): p. 1566-76.
191. Delano, D.L., et al., Genetically based resistance to the antiinflammatory effects of methotrexate in the air-pouch model of acute inflammation. Arthritis Rheum, 2005. 52(8): p. 2567-75.
192. Contos, J.J., et al., Requirement for the lpA1 lysophosphatidic acid receptor gene in normal suckling behavior. Proc Natl Acad Sci U S A, 2000. 97(24): p. 13384-9.
193. Ye, X., et al., LPA3-mediated lysophosphatidic acid signalling in embryo implantation and spacing. Nature, 2005. 435(7038): p. 104-8.
194. Tessier, P.A., et al., Chemokine networks in vivo: involvement of C-X-C and C-C chemokines in neutrophil extravasation in vivo in response to TNF-alpha. J Immunol, 1997. 159(7): p. 3595-602.
195. Akgul, C. and S.W. Edwards, Regulation of neutrophil apoptosis via death receptors. Cell Mol Life Sci, 2003. 60(11): p. 2402-8.
196. Akgul, C., D.A. Moulding, and S.W. Edwards, Molecular control of neutrophil apoptosis. FEBS Lett, 2001. 487(3): p. 318-22.
197. Edwards, S.W. and M.B. Hallett, Seeing the wood for the trees: the forgotten role of neutrophils in rheumatoid arthritis. Immunol Today, 1997. 18(7): p. 320-4.
198. Hampton, M.B., A.J. Kettle, and C.C. Winterbourn, Inside the neutrophil phagosome: oxidants, myeloperoxidase, and bacterial killing. Blood, 1998. 92(9): p. 3007-17.

112
199. Hashimoto, T., et al., Lysophosphatidic acid enhances in vivo infiltration and activation of guinea pig eosinophils and neutrophils via a Rho/Rho-associated protein kinase-mediated pathway. J Pharmacol Sci, 2003. 91(1): p. 8-14.
200. Li, P. and E.M. Schwarz, The TNF-alpha transgenic mouse model of inflammatory arthritis. Springer Semin Immunopathol, 2003. 25(1): p. 19-33.
201. Fujiwara, K., et al., Involvement of CXC chemokine growth-related oncogene-alpha in monosodium urate crystal-induced arthritis in rabbits. Lab Invest, 2002. 82(10): p. 1297-304.
202. Koch, A.E., et al., Growth-related gene product alpha. A chemotactic cytokine for neutrophils in rheumatoid arthritis. J Immunol, 1995. 155(7): p. 3660-6.
203. Armstrong, D.A., et al., Neutrophil chemoattractant genes KC and MIP-2 are expressed in different cell populations at sites of surgical injury. J Leukoc Biol, 2004. 75(4): p. 641-8.
204. Hall, L.R., et al., CXC chemokine receptor 2 but not C-C chemokine receptor 1 expression is essential for neutrophil recruitment to the cornea in helminth-mediated keratitis (river blindness). J Immunol, 2001. 166(6): p. 4035-41.
205. Kielian, T., B. Barry, and W.F. Hickey, CXC chemokine receptor-2 ligands are required for neutrophil-mediated host defense in experimental brain abscesses. J Immunol, 2001. 166(7): p. 4634-43.
206. Tateda, K., et al., Chemokine-dependent neutrophil recruitment in a murine model of Legionella pneumonia: potential role of neutrophils as immunoregulatory cells. Infect Immun, 2001. 69(4): p. 2017-24.
207. Houssiau, F.A., et al., Interleukin-6 in synovial fluid and serum of patients with rheumatoid arthritis and other inflammatory arthritides. Arthritis Rheum, 1988. 31(6): p. 784-8.
208. Rattazzi, M., et al., C-reactive protein and interleukin-6 in vascular disease: culprits or passive bystanders? J Hypertens, 2003. 21(10): p. 1787-803.
209. Dessein, P.H., B.I. Joffe, and S. Singh, Biomarkers of endothelial dysfunction, cardiovascular risk factors and atherosclerosis in rheumatoid arthritis. Arthritis Res Ther, 2005. 7(3): p. R634-43.
210. Lipsky, P.E., Interleukin-6 and rheumatic diseases. Arthritis Res Ther, 2006. 8 Suppl 2: p. S4.
211. Hanaoka, R., et al., A novel mechanism for the regulation of IFN-gamma inducible protein-10 expression in rheumatoid arthritis. Arthritis Res Ther, 2003. 5(2): p. R74-81.
212. Patel, D.D., J.P. Zachariah, and L.P. Whichard, CXCR3 and CCR5 ligands in rheumatoid arthritis synovium. Clin Immunol, 2001. 98(1): p. 39-45.
213. Stortelers, C., R. Kerkhoven, and W.H. Moolenaar, Multiple actions of lysophosphatidic acid on fibroblasts revealed by transcriptional profiling. BMC Genomics, 2008. 9: p. 387.
214. Legler, D.F., et al., B cell-attracting chemokine 1, a human CXC chemokine expressed in lymphoid tissues, selectively attracts B lymphocytes via BLR1/CXCR5. J Exp Med, 1998. 187(4): p. 655-60.
215. Ansel, K.M., et al., A chemokine-driven positive feedback loop organizes lymphoid follicles. Nature, 2000. 406(6793): p. 309-14.

113
216. Luther, S.A., et al., BLC expression in pancreatic islets causes B cell recruitment and lymphotoxin-dependent lymphoid neogenesis. Immunity, 2000. 12(5): p. 471-81.
217. Manzo, A., et al., Mature antigen-experienced T helper cells synthesize and secrete the B cell chemoattractant CXCL13 in the inflammatory environment of the rheumatoid joint. Arthritis Rheum, 2008. 58(11): p. 3377-87.
218. Shi, K., et al., Lymphoid chemokine B cell-attracting chemokine-1 (CXCL13) is expressed in germinal center of ectopic lymphoid follicles within the synovium of chronic arthritis patients. J Immunol, 2001. 166(1): p. 650-5.
219. Colonna, M., H. Nakajima, and M. Cella, A family of inhibitory and activating Ig-like receptors that modulate function of lymphoid and myeloid cells. Semin Immunol, 2000. 12(2): p. 121-7.
220. Bleharski, J.R., et al., A role for triggering receptor expressed on myeloid cells-1 in host defense during the early-induced and adaptive phases of the immune response. J Immunol, 2003. 170(7): p. 3812-8.
221. Bouchon, A., et al., TREM-1 amplifies inflammation and is a crucial mediator of septic shock. Nature, 2001. 410(6832): p. 1103-7.
222. Nochi, H., et al., Modulation of hepatic granulomatous responses by transgene expression of DAP12 or TREM-1-Ig molecules. Am J Pathol, 2003. 162(4): p. 1191-201.
223. Kuai, J., et al., TREM-1 expression is increased in the synovium of rheumatoid arthritis patients and induces the expression of pro-inflammatory cytokines. Rheumatology (Oxford), 2009. 48(11): p. 1352-8.
224. Gombert, M., et al., CCL1-CCR8 interactions: an axis mediating the recruitment of T cells and Langerhans-type dendritic cells to sites of atopic skin inflammation. J Immunol, 2005. 174(8): p. 5082-91.
225. Di Sciascio, M.B., et al., Eosinophil recruiting chemokines are down-regulated in peripheral blood mononuclear cells of allergic patients treated with deflazacort or desloratadine. Int J Immunopathol Pharmacol, 2007. 20(4): p. 745-51.
226. Berse, B., et al., Hypoxia augments cytokine (transforming growth factor-beta (TGF-beta) and IL-1)-induced vascular endothelial growth factor secretion by human synovial fibroblasts. Clin Exp Immunol, 1999. 115(1): p. 176-82.
227. Yun, J.K. and M. Kester, Regulatory role of sphingomyelin metabolites in hypoxia-induced vascular smooth muscle cell proliferation. Arch Biochem Biophys, 2002. 408(1): p. 78-86.
228. Takabe, K., et al., "Inside-out" signaling of sphingosine-1-phosphate: therapeutic targets. Pharmacol Rev, 2008. 60(2): p. 181-95.
229. Faour, W.H., et al., Prostaglandin E(2) regulates the level and stability of cyclooxygenase-2 mRNA through activation of p38 mitogen-activated protein kinase in interleukin-1 beta-treated human synovial fibroblasts. J Biol Chem, 2001. 276(34): p. 31720-31.
230. Yuan, Y., et al., Cobalt inhibits the interaction between hypoxia-inducible factor-alpha and von Hippel-Lindau protein by direct binding to hypoxia-inducible factor-alpha. J Biol Chem, 2003. 278(18): p. 15911-6.
231. Kluk, M.J. and T. Hla, Signaling of sphingosine-1-phosphate via the S1P/EDG-family of G-protein-coupled receptors. Biochim Biophys Acta, 2002. 1582(1-3): p. 72-80.

114
232. Mechtcheriakova, D., et al., Sphingosine 1-phosphate phosphatase 2 is induced during inflammatory responses. Cell Signal, 2007. 19(4): p. 748-60.
233. Baggiolini, M. and I. Clark-Lewis, Interleukin-8, a chemotactic and inflammatory cytokine. FEBS Lett, 1992. 307(1): p. 97-101.
234. Kannan, S., Neutrophil chemotaxis: autotaxin induced attenuation of purine/pyrimidine signaling precede the overexpression of Integrin(s). Med Hypotheses, 2005. 65(1): p. 197-8.
235. Lin, C.I., et al., Lysophosphatidic acid regulates inflammation-related genes in human endothelial cells through LPA1 and LPA3. Biochem Biophys Res Commun, 2007. 363(4): p. 1001-8.
236. Yu, S., et al., Lysophosphatidic acid receptors determine tumorigenicity and aggressiveness of ovarian cancer cells. J Natl Cancer Inst, 2008. 100(22): p. 1630-42.
237. Shida, D., et al., Lysophosphatidic acid (LPA) enhances the metastatic potential of human colon carcinoma DLD1 cells through LPA1. Cancer Res, 2003. 63(7): p. 1706-11.
238. Matsuno, H., et al., The role of TNF-alpha in the pathogenesis of inflammation and joint destruction in rheumatoid arthritis (RA): a study using a human RA/SCID mouse chimera. Rheumatology (Oxford), 2002. 41(3): p. 329-37.
239. Zhang, Y., B.F. Ramos, and B.A. Jakschik, Neutrophil recruitment by tumor necrosis factor from mast cells in immune complex peritonitis. Science, 1992. 258(5090): p. 1957-9.
240. Kasama, T., et al., Interleukin-10 expression and chemokine regulation during the evolution of murine type II collagen-induced arthritis. J Clin Invest, 1995. 95(6): p. 2868-76.
241. Ohtsuka, Y., et al., MIP-2 secreted by epithelial cells increases neutrophil and lymphocyte recruitment in the mouse intestine. Gut, 2001. 49(4): p. 526-33.
242. Nishimoto, N., Interleukin-6 in rheumatoid arthritis. Curr Opin Rheumatol, 2006. 18(3): p. 277-81.
243. Kaplanski, G., et al., IL-6: a regulator of the transition from neutrophil to monocyte recruitment during inflammation. Trends Immunol, 2003. 24(1): p. 25-9.
244. Tessarz, A.S. and A. Cerwenka, The TREM-1/DAP12 pathway. Immunol Lett, 2008. 116(2): p. 111-6.
245. Buchs, N., et al., IL-1B and IL-1Ra gene polymorphisms and disease severity in rheumatoid arthritis: interaction with their plasma levels. Genes Immun, 2001. 2(4): p. 222-8.
246. Wong, M.L., et al., Interleukin (IL) 1beta, IL-1 receptor antagonist, IL-10, and IL-13 gene expression in the central nervous system and anterior pituitary during systemic inflammation: pathophysiological implications. Proc Natl Acad Sci U S A, 1997. 94(1): p. 227-32.
247. Ferland, C., et al., IL-16 activates plasminogen-plasmin system and promotes human eosinophil migration into extracellular matrix via CCR3-chemokine-mediated signaling and by modulating CD4 eosinophil expression. J Immunol, 2004. 173(7): p. 4417-24.
248. Sciaky, D., et al., Cultured human fibroblasts express constitutive IL-16 mRNA: cytokine induction of active IL-16 protein synthesis through a caspase-3-dependent mechanism. J Immunol, 2000. 164(7): p. 3806-14.

115
249. Rioja, I., et al., Potential novel biomarkers of disease activity in rheumatoid arthritis patients: CXCL13, CCL23, transforming growth factor alpha, tumor necrosis factor receptor superfamily member 9, and macrophage colony-stimulating factor. Arthritis Rheum, 2008. 58(8): p. 2257-67.
250. Vissers, J.L., et al., BLC (CXCL13) is expressed by different dendritic cell subsets in vitro and in vivo. Eur J Immunol, 2001. 31(5): p. 1544-9.
251. de Vere Tyndall, A., et al., Veiled (dendritic) cells in synovial fluid. Lancet, 1983. 1(8322): p. 472-3.
252. Cavanagh, L.L., et al., Rheumatoid arthritis synovium contains plasmacytoid dendritic cells. Arthritis Res Ther, 2005. 7(2): p. R230-40.
253. Sarkar, S. and D.A. Fox, Dendritic cells in rheumatoid arthritis. Front Biosci, 2005. 10: p. 656-65.
254. Taylor, P.C. and B. Sivakumar, Hypoxia and angiogenesis in rheumatoid arthritis. Curr Opin Rheumatol, 2005. 17(3): p. 293-8.
255. Tamm, M., et al., Hypoxia-induced interleukin-6 and interleukin-8 production is mediated by platelet-activating factor and platelet-derived growth factor in primary human lung cells. Am J Respir Cell Mol Biol, 1998. 19(4): p. 653-61.
256. Li, X. and S.S. Makarov, An essential role of NF-kappaB in the "tumor-like" phenotype of arthritic synoviocytes. Proc Natl Acad Sci U S A, 2006. 103(46): p. 17432-7.
257. Tager, A.M., et al., The lysophosphatidic acid receptor LPA1 links pulmonary fibrosis to lung injury by mediating fibroblast recruitment and vascular leak. Nat Med, 2008. 14(1): p. 45-54.
258. Devalaraja, R.M., et al., Delayed wound healing in CXCR2 knockout mice. J Invest Dermatol, 2000. 115(2): p. 234-44.
259. Oshima, M., P. Deitiker, and M.Z. Atassi, Targeting the antigen-binding site of HLA-restricting alleles in treatment of autoimmune disease. Crit Rev Immunol, 2007. 27(3): p. 271-88.

Annexes
Annexe 1. Répartition des gènes HLA sur le chromosome 6 humain
[259]

117
Annexe 2. Expression des HLA à la surface de la membrane cellulaire
(John M. Berestecky, Ph.D., University of Hawaii, 2007)

118
Annexe 3. Hématopoïèse
CFU-GEMM : Colony Forming Unit-Granulocyte-Erythroid-Makrophage-Megakaryocyte. CFU-GM: Colony Forming Unit- Granulocyte Makrophage; CFU-Mk: Colony Forming Unit- Megakaryocyte; CFU-E: Colony Forming Unit- Erythroid; CFU-G: Colony Forming Unit-Granulocyte; CFU-M: Colony Forming Unit-Makrophage; CFU-Eo: Colony Forming Unit-Eosinophil; CFU-Mast: Colony Forming Unit- Mast cell (Vial J.P. et Praloran V. 2005)

119
Annexe 4. Récepteurs aux chimiokines chez les leucocytes humains et leurs ligands spécifiques.
Thymocytes + + + + +
CD34+ cell + +
NK cell + + + + +
B lymphocyte + + + + + + + +
T lymphocyte + + + + + + + + + + + + + + +
Immature DC + + + + +
Mature DC + + +
Red blood cell +
Macrophage + + + + + + + +
Basophil + +
Eosinophil + + +
Neutrophil + + + + +
Platelets +
CXCR1 CXCR2 CXCR3 CXCR4 CXCR5 CCR1 CCR2 CCR3 CCR4 CCR5 CCR6 CCR7 CCR8 CCR9 XCR1 CX3CR1 Duffy D6
CXCL1/Gro +++ +++
CXCL2/Gro +++
CXCL3/Gro +++
CXCL4/PF-4
CXCL5/ENA-78 + +++
CXCL6/GCP-2 ++
CXCL7/NAP-2 +++ +++
CXCL8/IL-8 +++ +++ +++
CXCL9/Mig +++
CXCL10/ IP-10 +++

120
CXCL11/I-TAC +++
CXCL12/SDF-1 +++
CXCL13/BCA1 +++
CXCL14/BRAK
CCL1/I-309 +++
CCL2/MCP-1 + +++ +++ ++
CCL3/MIP-1 +++ +++
CCL4/MIP-1 + +++ + +++
CCL5/RANTES +++ ++ +++ +++ +++
CCL7/MCP-3 +++ +++ ++ + +
CCL8/MCP-2 ++ +++ ++ +++ +++
CCL11/eotaxin + +++ + +
CCL13/MCP-4 +++ +++ +++ + ++
CCL14/HCC-1 +++ +++
CCL15/Lkn-1 +++ +++
CCL16/LEC
CCL17/TARC +++ +
CCL18/PARC
CCL19/ELC +++
CCL20/LARC +++
CCL21/SLC + +++
CCL22/MDC +++
CCL23/MPIF-1 +++
CCL24/MPIF-2 +++
CCL25/TECK +++
CCL26/eotaxin-3 ++
XCL1/lymphotactin +++
XCL2/lymphotactin +++

121
Jour 0 Jour 6 Jour 7
1ère injection sous-cutanée d’air stérile
FORMATION DE LA POCHE
Injection +/- TNF- 16h avant le traitement au
LPA ou OMPT
Injection LPA ou OMPT et collecte des cellules à 2h, 4h, 6h ou 8h post-injection
Jour 3
2ème injection d’air stérile
MAINTIENT DE LA STRUCTURE DE LA POCHE
• Jour 0: L’injection sous-cutanée d’air stérile sur le dos dela souris induit la perte de contact des cellules entre elles,qui provoque une inflammation aigüe.• Jour 3: Après 3 jours, la poche formée commence àcicatriser. Il est donc nécessaire d’effectuer une deuxièmeinjection d’air, afin de la maintenir.• Jour 6 et 7: Six jours après la première injection sous-cutanée d’air stérile, une structure qui ressembleétroitement à la membrane synoviale est formée. Onretrouve notamment des types cellulaires « fibroblast-like »et « macrophage-like » de morphologie comparable auxsynoviocytes de type A et B retrouvé dans la membranesynoviale des patients atteints de PR (Willoughby et al.,1981).
POCHE D’AIR PRÊTE APRÈS 6 JOURS
Ce modèle a l’avantage d'éviter les difficultés techniques deprélèvement dans un genou. Administrer directement un stimulusinflammatoire à une articulation du genou et collecter des exsudatsinflammatoires par la suite est techniquement très difficile à exécutersans causer de dommages menant à l'inflammation.Ici, on peut facilement prélever de plus grand volume d’exsudatinflammatoire suite au stimulus que nous voulons étudier (LPA ouOMPT).
Annexe 5. Schéma expérimental du modèle de la poche d’air.