Embed Size (px)
Citation preview

Int. J. Insect MorphoL & Embryol., Vol. 9, pp. 25 to 40. 0020-7322/80/0201-0025502.00/0 © Pergamon Press Ltd. 1980. Printed in Great Britain.
ULTRASTRUCTURE DE L'EPITHELIUM FOLLICULAIRE ET DES ENVELOPPES DE L'OVOCYTE CHEZ
LEPISMACHILIS TA RGIONII (GRASSI) (THYSANURA : MACHILIDAE)
JACQUES BITSCH
Universit6 Paul Sabatier, 118 route de Narbonne 31077 Toulouse Cedex, France
(Accepted 17 September 1979)
Abstract--During oogenesis in Lepismachilis targionff, the follicular epithelium under- goes important transformations, synchronous with those of the oocyte. Three successive phases can be distinguished: (1) During previtellogenesis and primary vitellogenesis, a monolayered epithelium is formed around the oocyte; the cells then become more numerous through repeated mitoses: (2) From the beginning of secondary vitellogenesis, the follicular ceils acquire new characteristics. Phenomena of the endomitosis type lead to the formation of polyploid, highly lobate nuclei, while the cytoplasm becomes richer in organelles, indicating intense metabolic activity. The secretion reaches the perioocytic space, where the so-called previtelline envelope is deposited. The oocyte draws yolk precursors from this secretion, which probably contains nutrients supplied by the haemolymph through the intercellular spaces. At the end of vitellogenesis this secretion becomes very dense and forms the vitelline envelope: (3) Once vitellogene~is is over, the follicular cells become fiat and present new cytoplasmic changes, with the formation of numerous vesicles of various kinds. Junctions of the adhaerens type, which were absent during secondary vitellogenesis, become well developed. Finally the five different layers of the chorion are deposited.
Index descriptors (in addition to those in title): Cellular differentiation, chorion, inter- cellular spaces, endomitosis, vitelline envelope, yolk precursors.
I N T R O D U C T I O N
PARMI LES lnsectes, les Thysanoures constituent un groupe original, qui pr6sente h la fois des caract6res de Pt6rygotes et des caract6res rappelant ceux de certains Crustac~s. Ceci est vrai en particulier pour leur d6veloppement post-embryonnaire et pour leur reproduction. Rappelons que chez les Thysanoures l 'ovaire est de type panoistique; le fonctionnement ovarien est soumis ~ des cycles alternant avec les cycles de mue; les phases terminales de la maturat ion ovarienne ont lieu au d6but de chaque stade d' intermue, la ponte se situant vers le milieu des stades. La f6condation, r6alis6e dans les tout premiers jours de chaque stade, est indispensable au complet d6veloppement des ovocytes (Watson, 1964).
L'ovogen6se des L~pismes a fait l 'objet d 'observat ions anciennes, comme celles de Gross (1903) et de Perrot (1933), puis plus r6centes comme celles de Woodland (1958) et de Rohdendorf(1968). Une courte note, due h Cone et Scalzi (1967), signale quelques caract6- ristiques ultrastructurales des ovocytes et des cellules folliculaires de Lepisma saccharina. R~cemment deux articles ont &6 consacr6s ~t l 'ovogen~se de Thermobia domestica; Fun concerne des recherches cytochimiques (Wojewoda, 1976), l 'autre porte sur le d6veloppe- ment embryonnaire des gonades (Klag, 1977).
Dans la famille des Machilidae, nous avons donn6 les caract6ristiques histologiques de l 'ovogen6se de Machilis burgundiae (Bitsch, 1968). Dans une note consacr6e h l '&ude cytologique des enclaves vitellines, Cantacuz6ne et Martoja (1972) ont mis en 6vidence des
25

26 JACQUES BITSCH
faits originaux concernant la vitellogen~se de Petrobius maritimus. Nous nous proposons de compl6ter l'6tude ultrastructurale de l'ovogen6se dans le cas de Lepismachilis targionii. Ce travail fait l 'objet de deux articles: la pr6sente note est consacr6e h l'6pith61ium folliculaire et aux enveloppes de l 'ovocyte; un second article portera sur les cellules germinales et sur la vitellogen~se. Par ailleurs nous avons d6crit r6cemment la structure et I'origine des enveloppes externes de l 'ovariole (Bitsch, 1978).
MATERIEL ET TECHNIQUES
Les Lepismachilis targionii ont 6t6 captur6s dans la liti/~re de feuilles mortes, en for6t de Bouconne, aux environs de Toulouse, puis 61ev6s quelque temps au laboratoire. Les femelles ont 6t6 diss6qu6es dans du liquide de Ringer avant fixation des ovaires. Les techniques histologiques et histochimiques utilis6es sont celles mentionn6es dans un pr6c6.dent article (Bitsch, 1968). Pour la microscopie 61ectronique les pi6ces ont 6t6 fix6es pendant 16 18 hr h 4°C dans le m61ange paraformald6hyde b. 2%-glutarald6hyde h 3%, puis postfix6es pendant 50 rain ~i 4°C dans une solution h 1% de t6troxyde d'osmium; enfin certaines pi~ces ont 6t6 color6es pendant 1 hr 5. l'acetate d'uranyle. Un m61ange 6pon-araldite a servi de milieu d'inclusion. Des coupes semi-fines, colore~es au bleu de toluidine borat6, ont 6t6 systematiquement pratiqu6es avant confection des coupes ultra-fines. Ces derni6res ont 6t6 contrast6es h l'ac6tate d'uranyle et au citrate de plomb.
OBSERVATIONS
Pr6sentes dans le germarium sous forme de cellules pr6folliculaires, les cellules somatiques de l'ovariole s'organisent en un 6pith61ium unistratifi6 autour des ovocytes en d6but de croissance. Puis au cours de l'ovogen6se les cellules folliculaires subissent de profondes transformations, qui sont en rapport avec l'6volution des ovocytes le long de l'ovariole. Pour d6crire en d6tail ces transformations cytologiques, il est commode de se r6f6rer aux cinq phases suivantes de l'ovogen~se (qui seront d6crites plus en d6tail dans un prochain article): (a) Phase de diff6renciation des ovocytes. Elle se d6roule dans le germarium qui contient des ovogonies et des ovocytes en ddbut de prophase m6iotique. (b) Phase de pr6vitellogen6se. Les ovocytes commencent leur accroissement; d 'abord plac6s de faqon irr6guli6re, ils deviennent ensuite r6guli~rement empil6s. (c) Phase de vitellogen6se endog~ne (ou primaire). Des globules vitellins se forment ~t l'int6rieur de citero_es ergastoplasmiques dilat6es. (d) Phase de vitellogen6se exog6ne (ou secondaire). Le d6p6t d'une grande quantit6 de vitellus dans l 'ovocyte correspond h l ' incorporation, par micropinocytose, de prot6ines d'origine extraovarienne. (e) Phase de d6p6t des enveloppes de l 'ovocyte: enveloppe vitelline et chorion.
1. Les cellules pr~folliculaires du germarium et la mise en place de I'epithelium !"olliculaire autour des ovocytes en prOvitellogenOse
A l'int6rieur du germarium, les cellules pr~folliculaires sont intercal6es entre les ovogonies et les jeunes ovocytes. Elles se distinguent des cellules germinales par leur noyau un peu plus allong6, par leur cytoplasme clair pauvre en organites et surtout par leur forme: elles 6mettent des prolongements cytoplasmiques qui se glissent entre les cellules germinales, ainsi qu'entre ces derni~res et la mince lame basale (tunica propria) enveloppant l 'apex de l'ovariole.
D'autres cellules somatiques, fortement aplaties, se groupent pour former des ensembles dispos6s transversalement ~t la base du germarium, puis entre les follicules successifs. Ces cellules interfolliculaires ~t cytoplasme clair pauvre en organites ne montrent pas de carac- t6res ultrastructuraux particuliers.
Autour des ovocytes en d6but de pr6vitellogen~se les cellulesfolliculaires commencent se disposer de fa9on r6guli~re pour former un mince 6pith61ium unistratifi6. Au stade tr6s

Ultrastructure de l'Epithelium Folliculaire 27
m . . . . . . . . . . . . . . . . - ' " " " . . . . . . . . . . " ' ~ - 4 ,
• i f I i " •
FIGS. 1-4. Sch6mas montrant les changements de l'6pith61ium folliculaire autour d'un ovocyte b. diff6rents stades du cycle reproducteur. L'6chelle des figures est indiqu~e par les segments, qui repr6sentent 3 p.m. 1. Stade de pr6vitellogen6se. 2. D6but de vitellogen6se endog6ne. 3. Fin de vitellogen6se endog6ne. 4. Stade de vitellogen6se exog6ne, b = lame basale; e = espace p6riovo- cytaire; ee = 6pith61ium externe de l'ovariole; ef = 6pith61ium folliculaire; m = muscle; ma =
macula adhaerens; o = ovocyte; tp = tunica propria.
jeune repr6sent6 sur la Fig. 1, les cellules foll iculaires sont peu nombreuses et tr6s aplat ies ; leur 6paisseur ne d@asse pas 1 lam, sauf / t l ' endro i t des noyaux, pou r t an t eux-m~mes tr6s aplat is . Le cy top lasme est encore tr6s clair, pauvre en organites. D ' a b o r d 6troi tement appliqu6es contre l 'ovocyte, les cellules foll iculaires s 'en 6cartent bient6t , cr6ant un mince espace p6riovocytaire, dans lequel se placent les premi6res microvillosit6s ovocytaires , courtes et peu nombreuses.
2. L'#pithdlium folliculaire autour des ovoeytes en vitellogen~se endogkne Un 6pith61ium cubique, don t la hau teur a t te int 5-7 lain se const i tue au tour des ovocytes
of] appara issen t les premiers globules vitellins (Figs. 2, 3, 7). Une augmenta t ion impor tan te du nombre des cellules foll iculaires r6sulte de mitoses banales, don t le fuseau est en g6n6ral parall~le /t la surface du follicule. Dans le cy top lasme encore peu a b o n d a n t on note un accroissement du nombre des r ibosomes libres, de I 'ergastoplasme, des mi tochondr ies

2 8 J A C Q U E S B I T S C H
t p. b m .~..<_~+.../m
. . . . . . . . . . = - . . . . : . . ~ _ . . . ~ . . . . / . . ~ ~ ~ - - - r - E - = - - ~ : = ~ =
. ~ . . . . . . . ~ . . . . . . . . . , . ~ ~ . ~ . . ~ 7 ~ 7 . . ~ . ~ . • , . . . . 6 ~ . . . • . . . . . . i 1 1 ! 9 . ~ . ~ Y . J - - / ~ ~ " ~
0 " . . 1 " : ~ . " ' ' - Y ( ~ " : : ; " : ' . ' ~ : " " • . . . . . " . ' " . . . . " " "
Fi~s. 5 et 6. Schimas de l'ipithilium folliculaire lors de la formation du chorion. Les segments reprisentent 3 p.m. 5. Dibut du d~pit du chorion (ch) au-dessus de l'enveloppe vitelline (ev). 6. Fin du dip6t du chorion. Les diffirentes couches du chorion sont numiroties de 1-5. Autres
abriviations comme sur les Figs. 1-4.
sphiriques ou peu allongies; les complexes golgiens sont assez diveloppis, associis ~ des visicules claires. Les noyaux, de forme ovoide, mesurent environ 5 × 10 ~tm; ils contiennent un gros nucl6ole central, de 1-1.5 ilm de diamitre. Pris de l 'apex des cellules folliculaires les membranes latirales pr6sentent des jonctions de type macula adhaerens (comme ces jonctions ne paraissent pas se continuer sur tout le pourtour des cellules, nous pensons qu'il s'agit de maculae et non de zomdae).
3. L'~pith~lium folliculair autour des ovocytes en vitellogenOse exog~ne Apris la p6riode de multiplication, les cellules folliculaires augmentent de taille.
L ' ipi thi l ium folliculaire devient columnaire, la hauteur des cellules passe de 10-20 ~tm; par la suite les cellules reprennent une forme cubique (Fig. 4), en rapport avec l ' inorme accrois- sement de l 'ovocyte voisin. Les cytoplasmes montent des signes d'une intense activit6 m&abolique, tandis que les noyaux augmentent de taille et prennent une forme tris lobie. l,es espaces entre les cellules folliculaires demeurent presque toujours 6troits (de l 'ordre de 150 •) tout au long de la vitellogenise, mais les jonctions de type macula adhaerens ne sont que rarement visibles.
(a) Modifications nttcl~aires. Dans l ' ipi thi l ium qui entoure les ovocytes de grande taille, les noyaux se prisentent sous forme de masses d'environ 15 sur 25 lain, faites de plusieurs lobes pourvus chacun d'un nucl6ole (Fig. 11). Les observations faites en microscopie 61ectronique suggirent que la formation de ces noyaux lobis comporte la succession des 6tapes suivantes. On note d 'abord une condensation de la chromatine en figures chromo- somiques, suivie de la disparition de l 'enveloppe nucliaire (Fig. 9). I1 n 'a jamais 6t6 possible de diceler la prisence d 'un fuseau. Les chromosomes se dispersent ensuite dans le cyto- plasme, sans montrer une figure caractiristique de mitose. Dis le moment o~ les chromo-
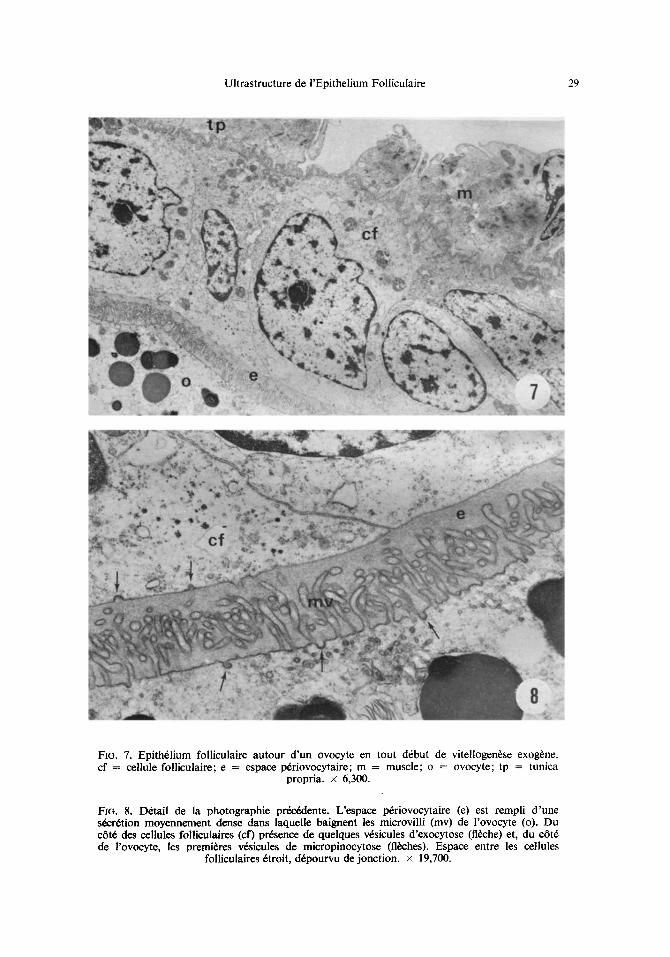
Ultrastructure de I'Epithelium Folliculaire 29
FIG. 7. Epith61ium folliculaire autour d 'un ovocyte en tout d6but de vitellogen6se exog~ne. cf = cellule folliculaire; e = espace p6riovocytaire; m = muscle; o = ovocyte; tp = tunica
propria, x 6,300.
Fla. 8. D6tail de la photographie pr6c6dente. L'espace p6riovocytaire (e) est rempli d 'une s6cr6tion moyennement dense dans laquelle baignent les microvilli (mv) de l'ovocyte (o). Du c6t6 des cellules folliculaires (cf) pr6sence de quelques v6sicules d'exocytose (fl~che) et, du c6t6 de rovocyte, les premi6res v6sicules de micropinocytose (fl~hes). Espace entre les cellules
folliculaires 6troit, d6pourvu dejonction. × 19,700.

30 JACQUES BITSCH
somes sont 6paissis, on remarque l'existence, dans le cytoplasme, de nombreuses cavit6s ergastoplasmiques, qui tendent h cerner les masses chromosomiennes et ~t reconstituer une enveloppe nucl6aire. Un noyau tr6s lob6 se reforme (Fig. 10), au sein duquel les images chromosomiennes disparaissent peu ~t peu au profit de mottes de chromatine dense. On retrouve alors l'aspect d'un noyau interphasique.
Ainsi l'augmentation de taille et la lobation accentu6e des noyaux s'effectuent par un ph6nom6ne d6riv6 d'une endomitose, avec d6roulement d'une prophase allant jusqu'~ la disparition de l'enveloppe nucl6aire, sans formation de fuseau ni figures de s6paration des masses chromatidiennes en deux lots, ni cytodi6r6se. I1 faut 6galement noter que ces ph6nom6nes nucl6aires se d6roulent dans des cellules d6jh diff6renci6es, riches en organites.
(b) Modifications cytoplasmiques. Le cytoplasme des cellules folliculaires s'enrichit en ribosomes libres, en ergastoplasme et en mitochondries. I1 pr6sente souvent une polarit6, tenant h ce que le reticulum endoplasmique, pouvant alors former des empilements assez r6guliers, se iocalise principalement dans la partie basale des cellules et dans la r6gion p6rinucl6aire. C'est 6galement dans ces r6gions qu'on observe des complexes golgiens plus ou moins d6velopp6s. De plus on voit apparaitre des gouttelettes lipidiques, associ6es au reticulum endoplasmique (Fig. 15). D'abord petites et espac6es, ces gouttelettes augmentent ensuite de taille. Les plus grosses, de forme ovale, mesurent 1.5 sur 2 I~m environ; plusieurs d'entre elles peuvent s'accoler pour former de volumineux areas.
(c) Enveloppe prdvitelline et enveloppe vitelline. L'espace p6riovocytaire qui s'est constitu6 lors de la phase cte viteilogen6se endog6ne, persiste pendant toute la vitellogen6se exog6ne. 11 mesure de 1-1.5 lam de hauteur; il est en partie occup6 par les nombreuses microvillosit6s de l'ovocyte, qui s'allongent peu ~t peu et parfois se ramifient, tandis que les Cellules follicu- laires n'6mettent ~t leur apex qu'un petit nombre de digitations. L'espace libre est enti6rement et uniform6ment rempli d'une substance tr6s finement grenue. D'abord assez claire aux 61ectrons (Fig. 8), cette substance devient progressivement plus dense. On peut nommer enveloppe prdvitelline la couche ainsi form6e, dans laquelle sont engag6es les microvillosit6s ovocytaires et ~t laquelle se substituera, en find e vitellogen6se, l'enveloppe viteUine propre- ment dite. Dans notre 6tude de 1968, nous avions utilis6 de mani6re globale le terme de membrane vitelline et signal6 sa richesse en polysaccharides (elle est fortement APS ÷) et en prot6ines.
L'origine des constituants de l'enveloppe pr6vitelline de Lepismachilis est difficile 6tablir. Aucune image ultrastructurale ne sugg6re la participation du cortex ovocytaire son 61aboration. Au contraire certaines observations plaident en faveur du r61e des cellules folliculaires. Au stade de la formation de l'enveloppe pr6vitelline, le cytoplasme de ces cellules renferme quelques v6sicules de 0.1-0.4 lam de diam6tre, remplies d'un mat6riel assez dense aux 61ectrons et finement granulaire. Les espaces compris entre les cellules folliculaires sont un peu dilat6s (ils mesurent en moyenne 200 A) et s'61argissent m~me par endroits pour donner de v6ritables poches; ces espaces sont remplis d'une substance finement granulaire, dense aux 61ectrons (Fig. 12). (~t et lh des v6sicules rev&ues, contenant une substance semblable, s'ouvrent dans les espaces intercellulaires (Figs. 13, 14). Des v6sicules de m~me type sont observ6es le long de la membrane plasmique apicale des cellules folliculaires: elles s'ouvrent alors directement dans l'espace p6riovocytaire (Fig. 8).
Vers la fin de la phase de vitellogen6se se produit une densification de la substance occupant l'espace p6riovocytaire, tandis que disparaissent les microvillosit6s de l'ovocyte. Ainsi se constitue une couche granulaire, assez homog6ne, d'une 6paisseur d ' l ~tm environ, qui repr6sente l'enveloppe vitelline proprement dite (Fig. 19). I1 n'est pas rare d'observer
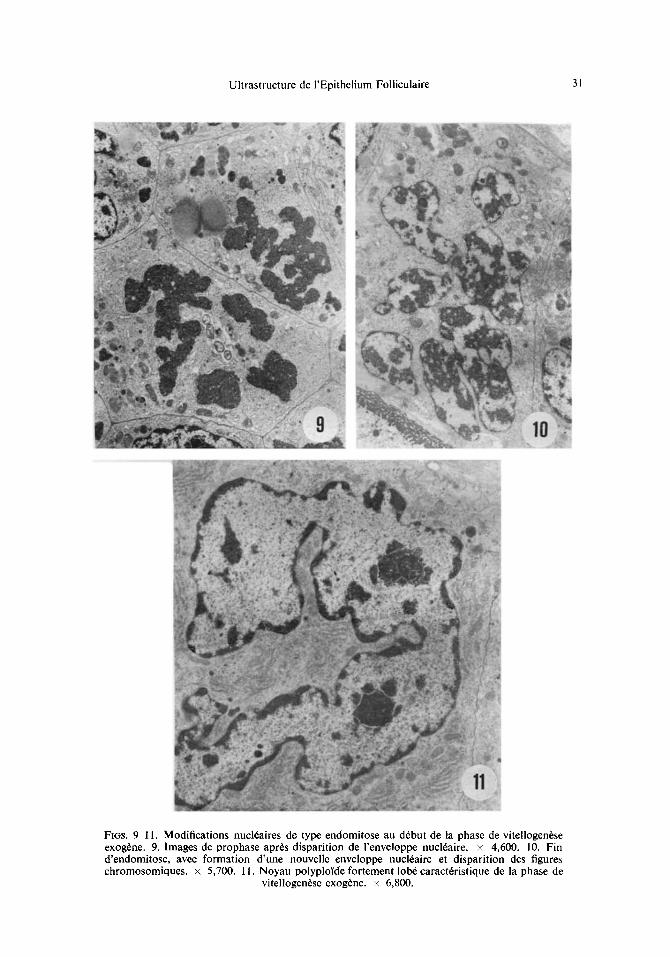
Ultrastructure de I'Epithelium Folliculaire 31
Fl6s. 9-11. Modifications nucl6aires de type endomitose au d6but de la phase de vitellogenese exog6ne. 9. Images de prophase apr6s disparition de l'enveloppe nucl6aire, x 4,600. 10. Fin d'endomitose, avec formation d'une nouvelle enveloppe nucl6aire et disparition des figures chromosomiques. × 5,700. 11. Noyau polyplol'de fortement lob6 caract6ristique de la phase de
vitellogen6se exog6ne, x 6,800.

32 JACQUES BITSCH
dans cette enveloppe des traces de cavit6s qui 6taient pr6c6demment occup6es par des microvillosit6s.
Dans l'6pith61ium folliculaire entourant les ovocytes en cours de vitellogen6se exog6ne, on observe parfois des cellules diss6min6es qui apparaissent plus sombres que les autres. Ces cellules particuli6res sont g6n6ralement 6tir6es dans le sens de la hauteur. Etles se distinguent des cellules voisines par la densit6 du hyaloplasme, par le nombre 61ev6 des ribosomes libres, par un gonflement des saccules ergastoplasmiques et de l'espace p6rinucl6aire, par l'abondance des mitochondries 5. matrice moyennement dense, enfin par la pr6sence de noyaux lob6s tr6s denses aux 61ectrons. En outre ces cellules renferment des inclusions lipidiques et souvent des lysosomes. En fait on trouve des stades interm6diaires entre les cellules folliculaires banales et des cellules tr6s 6tir6es et tr6s denses; ces derni6res r6sultent probablement de ph6nom6nes de d6g6n6rescence.
4. L'kpithOlium folliculaire lors du d~pot du chorion Pendant la formation du chorion, l'6pith61ium folliculaire s'6tire et s'amincit peu ~t peu,
pour ne plus mesurer que 3-5 ~tm d'6paisseur iorsque le chorion est achev6 (Figs. 5, 6, 16). Les noyaux demeurent plurilob6s, riches en chromatine, mais ils subissent un aplatissement prononc6. Le cytoplasme, encore riche en ribosomes libres et en complexes golgiens, pr6sente une modification des structures ergastoplasrniques: le syst6me de lames aplaties est remplac6 par un ensemble de v6sicules dilat6es, dont la membrane tend h perdre ses ribosomes (Fig. 17). Apparaissent 6galement d'autres v6sicules de tailles et de contenus vari6s; les plus abondantes sont petites, rev&ues ou non, /~ contenu dense ou clair. A c e stade un remaniement des contacts cellulaires se marque par le d6veloppement des jonctions de type zonula adhaerens (Fig. 18). La membrane apicale des cellules folliculaires forme des microvillosit6s dispos~es assez irr6guli~rement.
La structure du chorion a 6t6 observ6e principalement autour d'ovocytes mOrs contenus dans l'ovariole; b~ ce stade le chorion atteint environ sept lam d'6paisseur et montre cinq couches superpos6es (Fig. 19): (i) Couche 1. Mince assise fibrillaire d'environ 0.2-0.4 lam; moyennement dense aux 61ectrons, cette couche repose directement sur l'enveloppe vitelline. Elle renferme des cristaux en forme de baguettes ou de plaques claires, d'environ 0.2-0.3 ~tm de long pour une 6paisseur de 0.01 lam. Sur les coupes, ces cristaux ne forment pas une couche continue, ils se disposent plus ou moins parall61ement h la surface de l'enveloppe vitelline et apparaissent souvent group6s par 3 ou 4 (Fig. 21). I1 est probable que ces formations repr6sentent les cristaux de cire signal6s par les auteurs chez d'autres insectes. On observe parfois des cristaux identiques au sein m6me de l'enveloppe vitelline (Fig. 22). (ii) Couche 2. Son 6paisseur est d'environ 1-1.5 lam. Tr6s peu dense aux 61ectrons, elle est en continuit6 avec la couche pr6c6dente, mais son r6seau de fibrilles est beaucoup plus l~tche. Cette couche est fragile, c'est ~t son niveau que se cr6ent presque toujours des d6chirures sur les coupes. (iii) Couche 3. Dense aux 61ectrons, cette couche d'une 6paisseur inf6rieure 0.05 lam est finement granulaire. (iv) Couche 4. C'est la couche la plus 6paisse du chorion, atteignant 4 lam. Elle se compose de fibrilles moyennement denses aux 61ectrons, parail61es entre elles, mais pr6sentant une disposition h61icoidale plus ou moins r6guli6re, qui rappelle celle d'un endocuticule. (v) Couche 5. D'une 6paisseur d'environ 1.5 ~tm, elle se compose d'une matrice tr6s claire aux 61ectrons, finement granuleuse, dans laquelle sont diss6min6s des amas tr6s denses, de taille vari6e; ces areas tendent ~t s'aligner dans la zone de contact avec la partie superficielle de la couche 4. Le contact entre les deux couches se fait par une intrication de leur mat6riel respectif (Fig. 20)
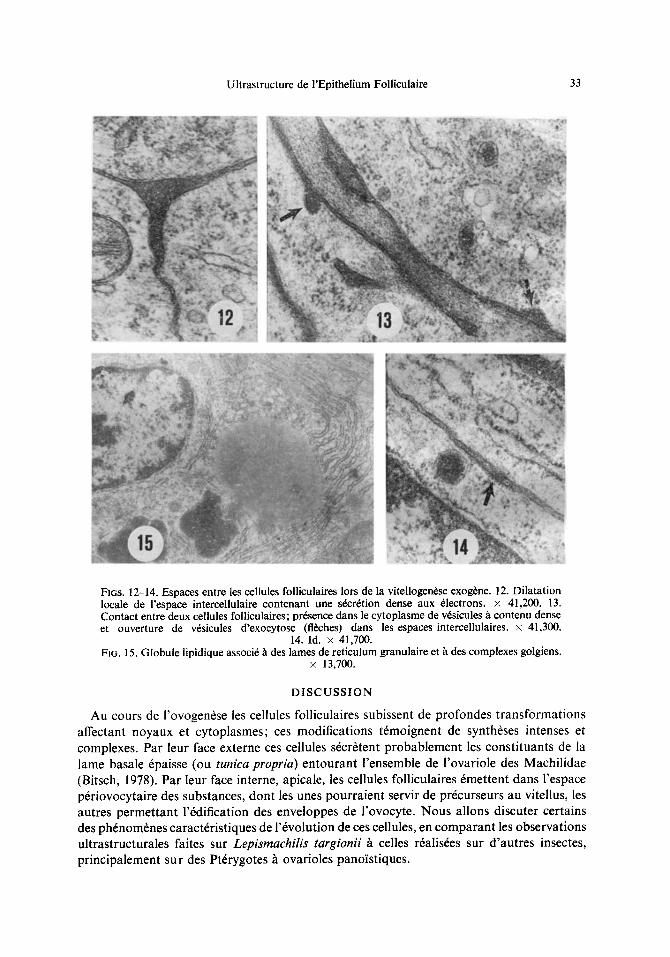
Ultrastructure de rEpithelium Folliculaire 33
Fi6s. 12-14. Espaces entre les cellules foiliculaires lors de la vitellogen6se exog6ne. 12. Dilatation locale de l'espace intercellulaire contenant une s6cr~tion dense aux 61ectrons. x 41,200. 13. Contact entre deux cellules folliculaires; pr6sence dans le cytoplasme de v6sicules h contenu dense et ouverture de v6sicules d'exocytose (fl~hes) dans les espaces intercellulaires, x 41,300.
14. Id. x 41,700. FIG. 15. Globule lipidique associ6 h des lames de reticulum granulaire et ~. des complexes golgiens.
x 13,700.
DISCUSSION
Au cours de l 'ovogen6se les cellules folliculaires subissent de profondes t ransformations affectant noyaux et cytoplasmes; ces modifications t6moignent de synth6ses intenses et complexes. Par leur face externe ces cellules s6cr~tent probablement les constituants de la lame basale 6paisse (ou tunica propria) entourant l 'ensemble de l 'ovariole des Machilidae (Bitsch, 1978). Par leur face interne, apicale, les cellules folliculaires ~mettent dans l 'espace p6riovocytaire des substances, dont les unes pourraient servir de pr6curseurs au vitellus, les autres permettant l '6dification des enveloppes de l 'ovocyte. Nous allons discuter certains des ph6nom+nes caract6ristiques de l '6volution de ces cellules, en comparan t les observations ultrastructurales faites sur Lepisrnachilis targionii ~ celles r6alis6es sur d 'autres insectes, principalement sur des Pt6rygotes h ovarioles panoistiques.
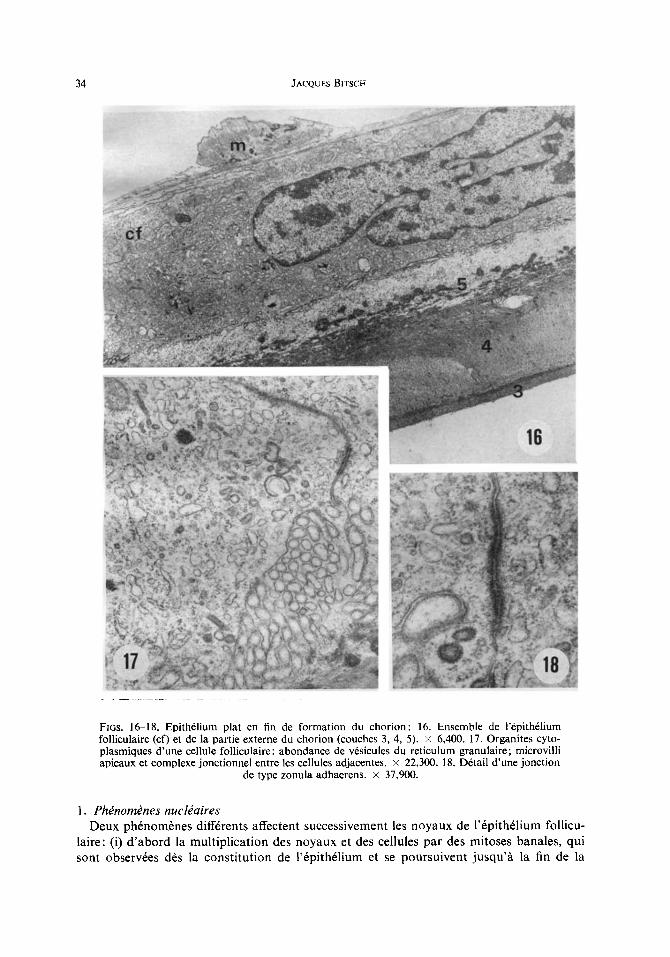
34 JACQUES BITSCi4
FIGS. 16-18. Epith61ium plat en fin de formation du chorion: 16. Ensemble de 1'6pith61ium follicu|aire (cf) et de la partie externe du chorion (couches 3, 4, 5). × 6,400. 17. Organites cyto- plasmiques d'une cellule folliculaire: abondance de v6sicules du reticulum granulaire; microvilli apicaux et complexe jonctionnel entre les cellules adjacentes, x 22,300. 18. D6tail d'une jonction
de type zonula adhaerens, x 37,900.
1. Ph~nom~nes nucl~aires Deux ph6nom6nes diff6rents affectent successivement les noyaux de l '6pith61ium follicu-
laire: (i) d ' a b o r d la mul t ip l ica t ion des noyaux et des cellules pa r des mitoses banales , qui sont observ6es d6s la const i tu t ion de 1'6pith61ium et se poursu ivent jusqu'~t la fin de la
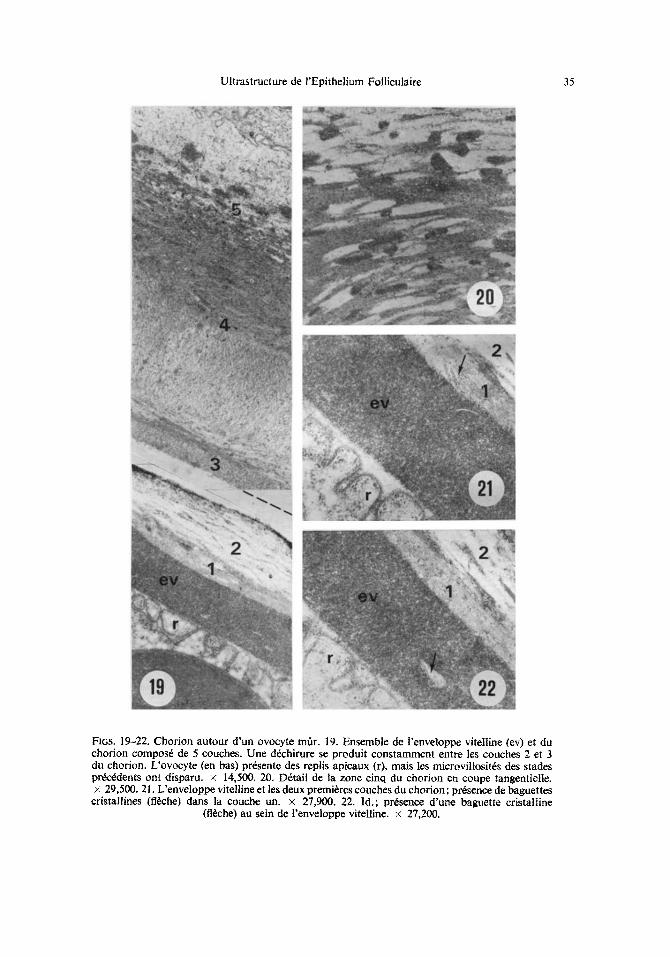
Ultrastructure de rEpithelium Folliculaire 35
Fins. 19-22. Chorion autour d'un ovocyte m~r. 19. Ensemble de l'enveloppe vitelline (ev) et du chorion compos6 de 5 couches. Une d6chirure se produit constamment entre les couches 2 et 3 du chorion. L'ovocyte (en bas) pr6sente des replis apicaux (r), mais les microvillosit6s des stades prekz6dents ont disparu, x 14,500. 20. D6tail de la zone cinq du chorion en coupe tangentielle. × 29,500.21. L'enveloppe vitelline et les deux premieres couches du chorion; pr6sence de baguettes
cristallines (ft~ehe) dans la couche un. x 27,900. 22. Id.; presence d'une baguette cristalline (fl~che) au sein de l'enveloppe vitelline, x 27,200.

36 JACQUES BITSCH
vitellogen6se endog6ne; (ii) puis la formation de noyaux de grande taille, fortement lob6s. ~t la suite de phgnom6nes de type endomitose; cette seconde phase intervient au d6but de la vitellogen6se exog6ne.
La succession de deux phases dans les transformations nucl6aires affectant les cellules foiliculaires a 6t6 mentionn6e chez plusieurs insectes, mais la seconde phase, g6n6ralement moins marqu6e que chez Lepismachilis, a 6t6 diversement interpr6t6e. Pour certains auteurs cette phase correspond h des mitoses aboutissant h des cellules bi- ou multinucl66s (Gross, 1903 chez Lepisma; von Kraft, 1960 chez l'Orthopt6re Tachycines; Beams et Kessel, 1969; Tembhare et Thakare, 1975 chez les Odonates). Dans son 6tude en microscopie optique sur l'ovogen6se de Periplaneta, Bonhag (1959) indique que les cellules folliculaires entourant les ovocytes charg6s de vitellus renferment plusieurs noyaux group6s en une masse compacte; mais ces agr6gats proviendraient de v6ritables mitoses et non d'amitoses. Dans d'autres cas les auteurs invoquent ou d6montrent des ph6nom6nes d'endomitose pour la formation de noyaux de grande taille. Ainsi chez Habrobracon King et Cassidy (1973) apportent la preuve, par une technique d'incorporation de thymidine tritige, que la croissance nucl6aire dans les cellules folliculaires r6sulte d'une r6plication endomitotique de I'A.D.N.
Chez Machilis des observations histologiques nous avaient conduit h d6crire, dans les cellules folliculaires entourant les ovocytes charg6s de vitellus, l'existence de masses plurinucl66es conduisant ~t des agr6gats et finalement h une fusion des noyaux (Bittch, 1968). Les 6tudes faites en microscopie 61ectronique sur Lepismachilis ne confirment pas cette conception. Elles montrent des images de prophase (condensation des chromosomes, disparition de l'enveloppe nucl6aire), mais sans formation de fuseau, ni disjonction anaphasique des chromatides. Le noyau qui se reconstitue ensuite apparaft de grande taille et fortement lob6. Ces images peuvent ~tre interpr~tges comme correspondant ~ un type d6riv6 d'une endomitose, conduisant b~ des noyaux polyploides. Des conclusions analogues ont 6t6 tir6es par Schmidt (1959) chez Lepisma saccharina. Cet auteur, sur la base d'observa- tions histologiques et de mesures volum6triques, admet que la plupart des noyaux des cellules folliculaires subissent des endomitoses conduisant ~t une t6tra- ou ~ une octoploidie, mais que se d6roulent aussi des mitoses diploides ~ octoploides, le plus souvent t6traploides.
2. R6les eventuels de l'~pith~lium folliculaire dans la vitellogenOse et dans la s~cr~tion de st~ro~des
Pendant toute la phase de vitellogen6se exog~ne l'activit6 de synth6se des cellules folliculaires est tr6s grande. L'augmentation du volume cytoplasmique et l'abondance de divers organites (ribosomes libres, empilements de saccules ergastoplasmiques, complexes golgiens, mitochondries) t6moigne de nombreuses syntheses prot6iques. De plus de volumi- neuses gouttelettes lipidiques se forment au contact 6troit des saccules du reticulum endo- plasmique et des v~sicules golgiennes. La polarit~ des cellules folliculaires rend compte de ce que la majeure partie des produits 61abor6s sont rejet6s dans l'espace p6riovocytaire.
Des fairs analogues sont connus chez de nombreux insectes. En ce qui concerne le m6tabolisme lipidique, divers auteurs ont mentionn6 la formation de "concentric whorls" du reticulum endoplasmique entourant les gouttelettes lipidiques synth6tis6es par les cellules folliculaires (Cummings et al., 1971 chez Drosophila; Liu et al., 1975 chez Simulium). Chez Drosophila, King et Koch (1963) ont suppos6 que les gouttelettes lipidiques des cellules folliculaires servaient ~t la fabrication de la membrane vitelline, qui contient en outre des prot6ines et des polysaccharides.
En ce qui concerne les synth6ses prot6iques, la question est de savoir si les cellules

Ultrastructure de l'Epithelium Folliculaire 37
folliculaires participent/t l'61aboration de pr6curseurs du vitellus, ou si elles servent seule- ment h l'6dification des enveloppes de l'ovocyte. Chez divers Pt6rygotes il a 6t6 d6montr6 que la majeure partie des prot6ines incorpor6es par l'ovocyte a une origine extra-ovarienne et que ces mol6cules transitent par les espaces intercellulaires dilat6s (Roth et Porter, 1964; Huebner et Anderson, 1972; Cruickshank, 1972; Cummings, 1972). Toutefois il est possible de mettre 6galement en 6vidence une participation des cellules folliculaires aux synth6ses de pr6curseurs du vitellus (Anderson et Telfer, 1969, 1970; Telfer et Smith, 1970; Cruickshank, 1971, 1972). Chez Leptinotarsa De Loof et Lagasse (1970) sugg6rent que des prot6ines fabriqu6es par les cellules folliculaires pourraient jouer le r61e d'enzymes n6cessaires I'incorporation des pr6curseurs du vitellus.
Chez Lepismachilis targionii les espaces entre les cellules folliculaires demeurent g6n6rale- ment non dilat6s pendant la vitellogen6se. Cependant on a observ6 dans quelques cas des dilations de ces espaces, avec formation de poches; ces espaces dilat6s sont alors remplis d'un mat6riel dense semblable ~t celui qui constitue l'enveloppe pr6vitelline et apparemment 6mis par exocytose h partir des cellules folliculaires elles-m6mes. De plus il appara~t au cours de I'ovogen6se une variation dans le d6veloppement des jonctions de type adhaerens, situ6es pr6s de l'apex des cellules folliculaires. Darts le jeune 6pith61ium les jonctions sont pr6sentes sous forme de courtes macules; pendant la vitellogen6se exog6ne, alors que le volume des cellules s'accrolt, elles deviennent trbs rares; enfin d6s |e d6but de la choriono- genbse des jonctions sont de nouveau visibles, elles prennent un assez grand d6veloppement et tendent vers le type zonula adhaerens. Ainsi pendant toute la vitellogen6se exog6ne les espaces intercellulaires, s'ils sont peu dilat6s, ne sont pas ferm6s; ils peuvent 6tre le si6ge de transport de m6tabolites provenant de l'h6molymphe et des cellules folliculaires, puis gagnant l'espace p6riovocytaire off l'ovocyte puise les pr6curseurs du vitellus. Nous pensons donc qu'un r61e des cellules folliculaires dans la vitellogen6se doit ~tre envisag6 chez Lepismachilis, mais il est 6vident que d'autres techniques seraient n6cessaires pour confirmer cette opinion.
Parmi les synth6ses que r6alisent les cellules folliculaires, on peut 6galement envisager celles de st6roides du groupe des ecdysones. Des travaux r6cents ont montr6 en effet que l'ovaire et les oeufs de divers insectes (comme Aedes, Belli¢ositermes, Locusta, Schistocerca, Bombyx et Galleria) renfermaient de telles substances. Dans quelques cas, comme chez Locusta migratoria (Lagueux et al., 1977), la preuve a 6t6 donn6e que l'6pith61ium folliculaire est le site de biosynth6se de ces ecdysones (Lagueux et al., 1977; Goltzene et al., 1978). 11 pourrait en &re de m~me chez les Thysanoures.
3. Formation des enveloppes de l'oeuf On dispose de peu de donn6es pr6cises sur les enveloppes de l'oeuf des Aptdrygotes.
Dans le cas des Protoures, Collemboles et Diploures Campodeidae, l'6pith6lium folliculaire est g6n6ralement tr6s mince (Pal6vody, 1971, 1976; Matsuzaki, 1973; Jura, 1975; Asaba et Ando, 1978); il existe une seule enveloppe, parfois nommde chorion, mais qui correspond plut6t h une enveloppe vitelline et qui, au moins chez Folsomia, tire son origine de sub- stances 6raises par l'ovocyte (Pal6vody, 1971, 1976). Chez les Thysanoures, comme c'est ]a r6gle chez les Pt6rygotes, deux enveloppes sont pr6sentes (enveloppe vitelline et chorion).
(a) Enveloppe prdvitelline et enveloppe vitelline. Le mode de formation de l'enveloppe vitelline varie selon les insectes; en rbgle g6n6rale, les auteurs notent d'abord l'apparition, dans l'espace p6riovocytaire, de plaques ou de grains s6par6s, tr~s denses, puis leur fusion

38 JACQUES BITSCH
pour constituer une enveloppe continue (King et Koch, 1963; Mathew et Rai, 1975; Bassemir, 1977).
Chez Lepismachilis au contraire on n'observe pas l'6mission de grains denses; l'espace p6riovocytaire se remplit d'embl6e d'une s6cr6tion qui enrobe les microbillosit6s ovocytaires et qui persiste pendant toute la dur6e de la vitellogen6se; nous la qualifions d'enveloppe prdvitelline, pour reprendre une expression d6js. utilis6e par d'autres auteurs. Loin de constituer une barri6re physiologique, elle se pr6sente comme une couche dans laquelle l'ovocyte pulse les m6tabolites n6cessaires 5. la formation du vitellus. Faite d'un mat6riel homog~ne, finement granulaire, sa densit6 aux 61ectrons s'accroit peu 5. peu, pour atteindre un maximum 5. la fin de la vitellogen6se. A ce moment les microvillosit6s ovocytaires disparaissent et il se constitue une couche dense, l'enveloppe vitelline proprement dite.
La question de l'origine des substances qui servent 5. l'6dification de l'enveloppe vitelline des insectes a 6t6 tr6s controvers6e. Pourtant la plupart des auteurs r6cents admettent que cette enveloppe est fabriqu6e par les cellules folliculaires, chez des insectes aussi diff6rents que Drosophila (King et Koch, 1963; Cummings et al., 1971), Gryllus (Favard-Sereno, 1966, 1972), Aeschna (Beams et Kassel, 1969). Ephestia (Cummings, 1972), Habrobracon (King et Cassidy, 1973), Aedes (Mathew et Rai, 1975) ou Simulium (Liu et al., 1975). Seuls quelques auteurs supposent que le cortex ovocytaire participe 6galement 5. l'61aboration de la membrane vitelline, comme chez Bombus (Hopkins et King, 1966), Anagasta (~ Ephestia) (Cruickshank, 1971, 1972). Chez Locusta migratoria Bassemir (1977) constate que les cellules folliculaires 6mettent, dans les espaces intercellulaires, des pr6curseurs qui deviennent des corps denses dans l'espace p6riovocytaire, puis une seconde s6cr6tion plus fine qui cimente les corps denses; toutefois rauteur n'exclut pas la possibilit6 que l'ovocyte s6cr6te un facteur qui condenserait les substances pr6sentes dans l'espace p6riovocytaire.
Chez Lepismachilis les observations ultrastructurales montrent la participation des cellules folliculaires 5. l'61aboration des pr6curseurs de l'enveloppe vitelline, alors qu'aucune image cytologique ne t6moigne de l'6mission de s6cr6tions par le cortex de l'ovocyte.
(b) Chorion. La distinction classique entre un endo- et un exochorion, facile 5. faire sur les coupes histologiques, apparait moins nette en ultrastructure chez Lepismachilis. En fait les couches un et deux du chorion, fines et solidaires de l'enveloppe vitelline, se confondent, en histologie, pour donner la "membrane vitelline" ; les couches trois et quatre correspondent sans doute 5. l'endochorion et la couche cinq 5. l'exochorion non encore achev6.
La microscopie 61ectronique a r6v616 la pr6sence de formations cristallines, en forme de baguettes ou de plaques, dans la partie du chorion qui repose imm6diatement sur l'enveloppe vitelline. De telles formations, consid~r6es comme des cires cristaUis6es, ont 6t6 signal6es chez divers insectes (Beament, 1946; King et Koch, 1963; Teller et Smith, 1970; Cruickshank, 1972). Elles forment un rev~tement imperm6able.
D'apr6s certains auteurs cette couche de cire se forme apr6s le d6p6t du chorion, de sorte que son origine probable serait i'ovocyte (Furneaux et MacKay, 1976). Ainsi les cires devraient traverser l'enveloppe vitelline par des canalicules analogues aux "wax canal filaments" de la cuticule (Furneaux et MacKay, 1976). Chez Lepismachilis les baguettes de cire se mettent en place d6s le b6but de la formation du chorion; cependant cette observation n'est pas contradictoire avec une origine ovocytaire. D'ailleurs il a 6t6 possible de voir quelques cristaux de cire 5. l'int6rieur de l'enveloppe vitelline, dans les cavit6s laiss6es libres par la disparition des microvillosit6s ovocytaires.
La structure h61icoidale des fibrilles constituant la couche principale du chorion (couche quatre) de Lepismachilis rapelle celle d6crite chez d'autres insectes (Teller et Smith, 1970;

Ultrastructure de l'Epithelium Folliculaire 39
Smith et al., 1971; Barbier et Chauvin, 1974; Furneaux et MacKay , 1976). Une telle
structure 6voque de fa~on frappante l 'archi tecture h61icoidale caract&ist ique de l 'endo-
cuticule des insectes (Bouligand, 1965, 1972).
Enfin en ce qui concerne la couche 5, sa structure h6t6rog6ne, d6crite chez Lepismachil is
autour d 'ovocytes mfirs situ~s dans l 'ovariole, laisse penser qu'el le n 'a pas encore atteint sa
const i tut ion d6finitive. Mais il ne nous a pas 6t6 possible de v6rifier ce fait sur des oeufs
f ra ichement pondus.
L 'ensemble des observat ions fakes sur Lepismachi l is targionii mont ren t la diversit6 des
syntheses dont est capable l'6pith61ium folliculaire et confirment l ' impor tance de ce tissu
dans le cas des ovarioles panoistiques.
B I B L I O G R A P H I E
ANDERSON, L. M. and W. H. TELFER. 1969. A follicle cell contribution to the yolk spheres of moth oocytes. Tissue Cell 1 : 633-44.
ANDERSON, L. M. and W. H. TELFER. 1970. Trypan blue inhibition of yolk deposition-a clue to follicle cell function in cecropia moth. J. EmbryoL Exp. MorphoL 23: 35-52.
ASABA, H. and H. Ayoo. 1978. Ovarian structure and oogenesis in Lepidocampa weberi Oudemans (Diplura: Campodeidae). Int. J. Insect MorphoL EmbryoL 7: 405-14.
BARB1ER, R. and G. CHAUVKN. 1974. Ultrastructure et r61e des a6ropyles et des enveloppes de l'oeuf de Galleria mellonella. J. Insect PhysioL 20: 809-20.
BASSEMIR, U. 1977. Ultrastructural differentiation in the developing follicle cortex of Locusta migratoria, with special reference to vitelline membrane formation. Cell Tissue Res. 185: 247-62.
BEAMENT, J. W. L. 1946. The formation and structure of the chorion of the egg in an Hemipteran, Rhodnius prolixus. Q. J. Microsc. Sci. 87: 393-439.
BEAMS, H. W. and R. G. KESSEL. 1969. Synthesis and deposition of oocyte envelopes (vitelline membrane, chorion) and the uptake of yolk in the dragonfly Aeschna (Odonata, Aeschnidae). J. Cell Sci. 4: 241-64.
BITSCH, J. 1968. Donnees histologiques sur l'oogen6se des Machilis (Insecta, Thysanura). Ann. Sci. Nat. ZooL 10: 267-90.
BITSCH, J. 1978. Structure de la gaine conjonctivo-musculaire entourant les ovarioles de Lepismachilis targionii (Grassi) (Thysanura: Machilidae). R61e possible de granulocytes ~. son contact. Int. J. Insect Morphol. Embryol. 7: 347-58.
BONHAG, P. F. 1959. Histological and histochemical studies on the ovary of the American cockroach Periplaneta americana (L.). Univ. Calif. PubL Entomol. 16: 81-124.
BOULIGAND, Y. 1965. Sur une architecture torsad~e r6pandue dans de nombreuses cuticules d'Arthropodes. C. R. Acad. Sci. Paris 261D: 3665-68.
BOULIGANO, Y. 1972. Twisted fibrous arrangements in biological materials and cholesteric mesophases. Tissue Cell4: 189-217.
CANTACUZENE, A. M. et R. MARTOJA. 1972. Origine des enclaves vitellines de l'oocyte d'un Insecte Thysan- oure, Petrobius maritimus. C. R. Acad. Sci. Paris 274D: 1723-26.
CONE, M. V. and H. A. SCALZL 1967. An ultrastructural study of oogenesis in the silverfish Lepisma sp. (Thysanura). J. Cell Biol. 35: 163A.
CRtJlCKSHANK, W. J. 1971. Follicle cell protein synthesis in moth oocytes. J. Insect PhysioL 17: 217-32. CRUICKSHANK, W. J. 1972. Ultrastructural modifications in the follicle cells and egg membranes during
development of flour moth oocytes. J. Insect PhysioL 18: 485-98. CUMMINOS, M. R. 1972. Formation of the vitelline membrane and chorion in developing oocytes of Ephestia
kuehn&lla (Lep., Phycitidae). Z. Zellforsch. Mikrosk. Anat. 127: 175-88. CUM~amGS, M. R., N. M. BROWN and R. C. KING. 1971. The cytology of the vitellogenic stages of oogenesis
in Drosophila melanogaster (Dipt., Drosophilidae). Ill. Formation of the vitelline membrane. Z. Zellforsch. Mikrosk. Anat. 118: 482-92.
DE LOOF, A. and A. LAOASSE. 1970. The ultrastructure of the follicle cells of the ovary of the Colorado beetle in relation to yolk formation. J. Insect PhysioL 16:211-20.
FAVARO-SERENO, C. 1966. Rfile de l'appareil de Golgi dans la s6cr6tion du chorion de l'oeuf chez le Grillon (Insecte, Orthopt~re). Pith Int. Congr. Electron Microsc. (Kyoto) 2: 553-54.
FAVARD-SERE~O, C. 1972. Ovogen6se d'un lnsecte Orthopt~re: le Grillon. Etude ultrastructurale. Th6se Univ. Paris VI.
FtJRNEAUX, P. J. S. and A. L. MACKAY. 1976. The composition, structure and formation of the chorion and the vitelline membrane of the eggshell, pp. 157-76. In H. R. Hepburn (ed.) The lnsect Integument, Elsevier Sci. Publ. Comp., Amsterdam, Netherlands.

40 JACQUES BITSCH
GOLTZENE, F., M. LAGUEUX, M. CHARLET and J. A. HOFMANN. 1978. The follicle cell epithelium of maturing ovaries of Locusta rnigratoria. A new biosynthetic tissue for ecdysone. Hoppe-Seyler's Z. Physiol, Chem. 359: 1427-34.
GROSS, J. 1903. Untersuchungen Ober die Histologie des Insecktenovariums. Zool. Jahrb. Anat. 18:71-186. HOPKINS, C, R. and R. C. KING. 1966. An electron microscopical and histochemical study of the oocyte
periphery in Bombus terrestris during vitellogenesis. J. Cell Sci. 1: 201-16. HUEBNER, E. and E. ANDERSON. 1972. A cytological study of the ovary of Rhodnius pro/ixus (Hem., Het.,
Reduviidae). I. The ontogeny of the follicular epithelium. J. MorphoL 136: 459-94. JURA, C. 1975. Ovaries structure in Acerentomon dispar Stach (Protura). Acta Biol. Cracov. Zool. 18: 55-65. KING, R. C. and J. D. CASSIDY. 1973. Ovarian development in Habrobracon juglandis (Ashmead) (Hyme-
noptera: Braconidae). II. Observations on growth and differentiation of component cells of egg chamber and their bearing upon interpretation of radiosensitivity data from Habrobracon and Drosophila. Int. J. Insect Morphol. Embryol. 2: 117-36.
KING, R. C. and E. A. KOCH. 1963. Studies on ovarian follicle cells of Drosophila. O. J. Microsc. Sci. 104: 297-320.
KLAG, J. 1977. Light and electron microscopic study of embryonic development of gonads in Therrnobia domestica (Packard) (Thysanura) with special reference to intercellular contacts. Acta Biol. Craeov. ZooL 20: 229-36.
KRAFT, A. von. 1960. Entwicklungsgeschichtliche und histochemische Untersuchungen zur Oogenese von Tachycines (Insecta, Saltatoria). II. Mitteilung: Die Entwicklung der Oocyten. Zool. Jahrb. Anat. 78: 485-558.
LAGUEUX, M., M. HIRN and J. A. HOFFMANN. 1977. Ecdysone during ovarian development in Locusta migratoria. J. Insect PhysioL 23: 109-19.
LIu, T. P., J. J. DARLEY and D. M. DAVIES. 1975. Differentiation of ovariolar follicular cells and formation of previtelline-membrane substance in Simuliurn vittaturn Zetterstedt (Diptera: Simuliidae). Int. J. Insect MorphoL ErnbryoL 4: 331-40.
MATHEW, G. and K. S. RAI. I975. Structure and formation of egg membranes in Aedes aegypti (L.) (Diptera: Culicidae). Int. J. Insect MorphoL EmbryoL 4: 369-80.
MATSUZAKI, M. 1973. Oogenesis in the springtail, Tomocerus minutus Tullberg (Collembola: Tomoceridae). Int. J. Insect Morphol. Embryol. 2: 335-49.
PALEVODY, CI. 1971. L'ovogen6se chez les Collemboles. Structure et 6volution de l'ovaire. C. R. Acad. Sc. Paris 272D: 3165-68.
PALEVODY, CI. 1976. L'ovogen6se chez les Collemboles Isotomides. Cytologic et approche physiologique. Th6se Universit6 Toulouse.
PERROT, J. L. 1933. La spermatogen6se et l'ovogen6se de Lepisrna (Thermobia) dornestica. H6t6ropycnose dans un sexe homogam6tique. Z. Wiss. Biol. Abt. Zellforsch. 18: 573-92.
ROHDENDORE, E. B. 1968. The fundamental schema of oogenesis in the firebrat Lepisrnodes inquilinus (Thysanura, Lepismatidae) and its periodisation in connection with imaginal moulting cycles. Acta Entomol. Bohemoslov. 65: 341-48.
ROTH, T. F. and K. R. PORTER. 1964. Yolk protein uptake in the oocyte of the mosquito Aedes aegypti L. J. Cell Biol. 20: 313-32.
SCHMIDT, E. M. 1959. Untersuchungen zur Cytologie des Wachstums in der postembryonalen Entwicklung von Lepisma saceharina L. Zool. Jahrb. Anat. 77: 357-410.
SMITH, D. S., W. H. TELFER and A. C. NEVILLE. 1971. Fine structure of the chorion of a moth, Hyalophora eecropia (Lep. Saturniidae). Tissue Cell 3: 477-98.
TELFER, W. H. and D. S. SMITH. 1970. Aspects of egg formation, pp. 117-34. In A. C. Neville (ed.) Insect Ultrastructure. Symp. R. Entornol. Soc. Lond. no. 5, Blackwell Sci. Publ., Oxford, England.
TEM~HARE, D. B. and V. K. TIqAKARE. 1975. The histological and histochemical studies on the ovary in relation to vitellogenesis in the dragonfly, Orthetrurn chrysis Selys (Libellulidae: Odonata.). Z. Mikrosk.-Anat. Forsch. 89: 108-27.
WATSON, J. A. L. 1964. Moulting and reproduction in the adult firebrat, Therrnobia domestica (Packard) (Thysanura, Lepismatidae). II. The reproductive cycles. J. Insect Physiol. 10: 399-408.
WOJEWODA, m. 1976. Cytochemical and cytophotometric investigations into the ovary growth and oogenesis in Thermobia dornestica (Packard). Zool. Pol. 25: 47-80.
WOODLAND, J. T. 1958. Oogenesis and fertilization in Thermobia dornestica (Packard). Psyche 65:11-25.









![Synthèse - mced.fr · de la croissance folliculaire humaine n’ont pas changé de façon substantielle au cours des dernières années [1], des avancées significa-tives dans la](https://img.pdfslide.fr/doc/110x75/5b9c4bcc09d3f2f6368c5f37/synthese-mcedfr-de-la-croissance-folliculaire-humaine-nont-pas-change.jpg)