Embed Size (px)
Citation preview

MISES AU POINT
Valeur semiologique des apolipoprot( ines
J . C . F R U C H A R T *
Semiological value of apolipoproteins.
Mots cl~s : apolipoprot6ines m6tabolisme lipidique.
Rev. Mdd. interne, 1985, 6, 41-48.
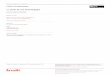
L'importance de la partie prot6ique des lipo- prot6ines, les apolipoprot6ines, dans le t r ans port des lipides, les interactions lipoprot6ines- r6cepteurs ainsi que dans la r6gulation de l'ac- tivit6 des enzymes impliqu6es dans le m6tabo- lisme des particules lipoprot6iques, a 6t6 r6- cemment reconnue (1, 7) (fig. 1).
Sur la base de la classification d'Alaupovic, douze apolipoprot6ines peuvent 6tre individua- lis6es au sein des lipoprot6ines (tableau I).
- - L ' a p o l i p o p r o t ~ i n e A-I (apo A-I), apopro- t6ine majeure des lipoprot6ines de haute den- sit6 (HDL), a une masse mol6culaire de 28331, et sa s6quence en amino-acides est connue. Elle existe dans le plasma sous diff6rentes formes s6parables en iso61ectrofocalisation (apo A-I Milano, Munster, Marburg, etc.). L'apo A-I est un activateur de la 16cithine cho- lest6rol acyltransf6rase (LCAT), enzyme qui es- t6rifie le cholest6rol libre des HDL et participe
la transformation des H D L 3 en HDL2. L'apo A-I est synth6tis6e et secr6t6e sous forme d ' H D L par le foie, mais provient aussi de Fin- testin o~ elle est incorpor6e dans les chylomi-
crons. De r6centes 6tudes sugg6rent que l 'apo A-I est synth6tis6e sous forme de propeptide cliv6 intracellulairement avant d'etre s6cr6t6 (5, 15). Le catabolisme de ces chylomicrons sous l'action de la lipoprot6ine-lipase aboutit & la li- b6ration de constituants de surface qui servent de pr6curseurs 5 au moins une partie des H D L plasmatiques.
Les valeurs physiologiques habituelles dans le s6rum des sujets adultes sont comprises en- tre 1,14 et 1,62 g/l. La concentration s6rique en apo A-I augmente dans les hyper-alphalipo- prot6in6mies familiales (gage de long6vit6) (36), au cours de l'exercice physique (29) et de la prise mod6r6e d'alcool (37). Elle diminue en cas d'hypertriglyc6rid6mie (33) et dans les af- fections h6patiques graves (h6patites, cirrhoses, cancers), du fait d 'un d6faut de synth6se (25). Au cours de la maladie de Tangier, qui se ma- nifeste cliniquement par une spl6mom6galie et une neuropathie, l 'apo A-I est quasiment ab- sente du s6rum, alors qu'on y retrouve la pro- apo A-I ; il est probable qu'il existe darts cette affection un d6ficit en enzyme qui assure le cli- vage de la pro-apo A-I ou une anomalie struc- turale qui emp6che le transport de la pro-apo A-I ~t son site de clivage (6).
Plusieurs affections, s 'accompagnant d 'une tr6s forte diminution voire d'une disparition de l 'apo A-I s6rique, ont 6t6 d6crites r6cemment : d6ficit en HDL, d6ficit familial en .apoprot6ine A-I et C-III (34), absence d 'apo A-I, ~fish-eye disease>> (39).
* Ddpartement d'Etude des lipides-lipoprotdines et normo- lipoprotdindmiants, facultO de pharmacie, 3, rue du Profes- seur-Laguesse, 59045 Lille Cedex.
Tirds ?t part : J.C. Fruchart, m~me adresse.
Abrdviations utilisdes: H D L = l ipoprot6ines de h a u t e densit6, L D L = l ipoprot6 ines de basse densi t6, V L D L = l ipoprot6 ines de tr6s bas se densit6, I D L = lipo- prot6 ines de densi t6 in term6dia i re (1,006 & 1,019), L C A T = cholest6rol acyl t ransf6rase .

42 J. C. Fruchart La Revue de Mddecine interne Janvier 1985
<22 Intestin
A-T
E ~ B 4 8 LPL
s Cs, A-T J
//~ B 100
Foie
.... v Foie E B48
Fig. I
. . . . .
B100 } r~cepteurs [ ] dufoieetdes
I autres tissus
B 100 ~.._
Apolipoprot6ines et m6tabolisme des lipoprot6ines. AI = apolipoprot6ine AI, B48 et B 100 = sous-types d 'apol ipo- prot6ine B, C ' s = apolipoprot6ine C, E = apolipoprot6ine E, LPL = lipoprot6ine lipase.
Le r61e jou6 par l'apo A-I en tant que facteur protecteur vis-a-vis de l'ath6roscl6rose est en- core discutS. Le d6ficit en apo A-I dans la ma- ladie de Tangier n'est pas ath6rog6ne alors qu'il l'est fortement lorsqu'il existe un d6ficit associ6 en apo C-Ill (39). De nombreuses 6tudes 6pid6miologiques r6trospectives r6ali- s6es sur des patients coronarographi6s mon- trent d'6troites relations entre la diminution de la concentration s6rique en apo A-I et l'ath6- roscl6rose coronarienne (35, 30).
- - L'apol ipoprot~ine A-II (apo A-II) est une prot6ine de structure des HDL form6e de deux chalnes prot6iques identiques de 77 amino- acides unies par un pont disulfure ; elle est synth6tis6e sous forme de pro-apo A-II par l'intestin et le foie (19). L'apo A-II est un acti- vateur de la triglyc6ride-lipase h6patique (22).
Les valeurs physiologiques habituelles dans le s6rum des sujets adultes sont comprises en- tre 0,27 et 0,71 g/l.
La concentration s6rique e n apo A-II n'est pas sensible aux glucides, mais augmente nette-
ment chez les buveurs excessifs (37). En cas d'atteinte de la cellule h6patique (h6patite, cir- rhoses, cancers), elle chute rapidement et pro- portionnellement h la gravit6 de l'affection en cause (25). Bien que certains auteurs aient rap- port6 une baisse de la concentration s6rique en apo A-II dans l'ath6roscl6rose coronarienne, des 6tudes r6trospectives r6centes r6alis6es sur plusieurs milliers de sujets coronarographi6s ne confirment pas la valeur discriminante de ce param6tre pour 6valuer le risque ath6rog6ne (25).
--L'apolipoprot~ine A - I V (apo A - I V ) de masse mol6culaire 46000 est synth6tis6e dans l'intestin, et secr6t6e sous forme de chylomi- crons. Des variants structuraux ont 6t6 d6crits. On ne connait pas encore leur signification cli- nique.
- - L'apol ipoprot~ine B (apo B) est la pro- t6ine structurale des chylomicrons, VLDL, LDL et Lp (a). Elle est synth6tis6e dans l'intes- tin et le foie off elle joue un r61e important

Tome VI Valeur sdmiologique des apolipoprotdines 43 Num&o 1
Tableau I
PROPRII~TI~S ET FONCTIONS DES APOPROTEINES
APOLIPO- MASSE MOL13CULAIRE SITE DE FONCTION PROTI~INE X 10 - 3 SYNTHI~SE
A-I 28,3 intestin cofacteur de la LCAT foie
A-II 17 intestin prot6ine de structure des HDL
A-IV 46 intestin non connu
B 100 549 foie synth6se et transport des VLDL et des LDL
B48 265 intestin synthSse et transport foie des chylomicrons et des
VLDL
C-1 6,5 foie activateur de la LCAT
C-II 8,9 foie activateur de la LPL intestin
C-III 9 foie inhibiteur de la LPL 0,1,2 intestin
D 20 9 r61e dans le m6tabolisme des esters de cholest6rol
E 39 foie permet la fixation des ~ remnants ~ des VLDL et des chylomicrons sur les r6cepteurs cellulaires h6patiques
F 30 foie ?
H ? activateur de la LPL
Lp (a +) 250 foie .9
clans la s6cr6tion et le transport de ces lipopro- t6ines. Sa structure est encore real connue mais en utilisant l'61ectrophorSse en gel de polyacry- lamide en pr6sence de dodecylsulfate de so- dium, Kane et coll. (24) ont individualis6 deux sous-types, l 'apo B100 de masse mol6culaire 340000 et l'apo B48 dont la masse mol6culaire est environ 48 p. 100 de celle de l'apo B100.
L'apo B48, prot6ine de structure des chylo- microns, est synth6tis6e exclusivement dans l'intestin. Aprbs hydrolyse des triglyc6rides des chylomicrons par la lipoprot6ine-lipase, l 'apo B48 reste ~t la surface des particules r6siduelles (~remnants)0 qui sont ensuite capt6es par le foie apr6s fixation sur des r6cepteurs sp6cifi- ques (apo E r6cepteurs). Sa dur6e de vie brSve
(10 minutes) en fait un excellent marqueur du degr6 de captation h6patique de ces particules r6siduelles qui sont bien connues pour leur pouvoir ath6rog6ne.
L'apo B100 pr6domine dans les LDL, IDL et VLDL, mais est aussi pr6sente dans les chylo- microns. Au niveau du foie, la synth6se de l'apo B100 est induite par les acides gras libres provenant de la lipolyse extra-h6patique ou d 'un syst6me dans l'h6patocyte. L'apo B100 est incorpor6e essentiellement dans les VLDL qui seront ensuite catabolis6es en IDL puis en LDL ; lors de cette transformation l'apo B100 reste g la surface de ces lipoprot6ines. Une par- tie de l 'apo B 100 peut ~tre s6cr6t6e directement sous forme de LDL, en particulier en cas

44 J. C. Fruchart La Revue de Mddecine interne Janvier 1985
d'hyperlip6mie de type lI. L'apo B100 des LDL interagit avec des r6cepteurs sp6cifiques (apo B, E r6cepteurs) qu'on trouve dans des cellules extra-h6patiques comme les fibro- blastes, les myocytes etc. et avec des r6cepteurs h6patiques (7) ; apr6s internalisation, les LDL sont d6grad6es et l 'apo B est prot6olys6e. Les valeurs physiologiques habituelles pour l 'apo BI00 dans le s6rum de sujets adultes sont com- prises entre 0,7 et 1,3 g/1. La concentration s6- rique en apo B48 est normalement tr6s faible et n6gligeable par rapport fi celle de l 'apo BI00.
On observe une augmentation de la concen- tration s6rique en apo B si le catabolisme des lipoprot6ines riches en apo B e s t emp6ch6 ou s'il existe une surproduction de ces lipopro- t6ines d6passant les capacit6s d'6puration de l'organisme.
Les anomalies du catabolisme des lipopro- t~ines riches en apo B sont de trois types :
D6faut de lipolyse des lipoproteines riches en triglyc6rides par d6ficit en lipoprot6ine- lipase ou en activateurs de l'enzyme (apo C-II, apo H), par hypersynth6se ou anomalies du ca- tabolisme d'inhibiteurs de l'enzyme (apo C- III). On observe ces anomalies dans les hyper- lipoprot6in6mies de type I, IV et V de la classi- fication internationale, dans le diab6te insuli- noprive et l 'insuffisance r6nale (2, 9). Au cours de nombreuses affections h6patiques, l 'apo B s'accumule dans des LDL riches en triglyc6- rides du fait d'une diminution de l'activit6 de la triglyc6ride-lipase h6patique associ6e fi un d6ficit en LCAT et en prot6ine assurant le transfert du cholest6rol entre les lipoprot6ines.
D6faut de conversion des VLDL en LDL du fait d 'une anomalie des apoprot6ines per- mettant l'interaction avec les r6cepteurs (apo E). Ce d6faut est responsable de l 'hyperlipo- prot6in+mie de type III de la classification in- ternationale (31, 40).
Defaut de catabolisme des LDL du fait d'une absence ou d'une diminution de synthbse des r6cepteurs, d'une anomalie structurale res- ponsable d 'un d6faut de reconnaissance entre la lipoprot6ine et le r6cepteur ou d'une mau- vaise internalisation. Ce d6faut est responsable de l 'hypercholest6rol6mie essentielle (type II A homozygote ou h6t6rozygote de la classifica- tion internationale) (7).
L'hyperproduction d 'apo B se retouve dans une grande vari6t6 d'hyperlipoprot6in6mies primitives (types II A, II B, III, IV de la classi- fication internationale) et secondaires (diab6te non insulinoprive, syndrome n6phrotique, ob6- sit6), mais les m6canismes ~t l'origine de cette
Tableau II
VALEURS PHYSIOLOGIQUE HABITUELLES CHEZ L'ADULTE
Apo A 1 1,14 h 1,62 g/l Apo A II 0,27 A 0,71 g/1 Apo B 100 0,7 ~t 1,3 g/1 Apo B 48 taux n6gligeable Apo C I 50 A 110 rag/1 Apo C II 10/t 67 mg/1 Apo C III 80 fi 150 mg/1 Apo E 30 fi 50 mg/1
hyperproduction sont encore mal connus. De nombreuses 6tudes 6pid6miologiques r6tros- pectives, r6alis6es en particulier chez des sujets coronarographi6s, ont mis en 6vidence une 6troite corr61ation entre le risque ath6rog6ne et la concentration s6rique en apo B totale ou en LDL apo B e t ce, m~me chez les sujets normoli- p6miques (14, 28, 35). Darts ce dernier cas, l 'hyperproduction d 'apo B entraine la synth6se de particules lipoprot6iniques ath6rog6nes, re- trouv6es en ultracentrifugation dans la zone des LDL mais de composition anormale (aug- mentation du rapport apo B/cholest6rol). Sni- derman et coll. ont r6cemment propos6 d'indi- vidualiser cette affection sous le nom d'hyper- apo-b6talipoprot6in6mie (28).
La concentration en apo B s6rique est tr6s di- minu6e voire nulle au cours d'une maladie rare caract6ris6e cliniquement par une r6tinite, une acanthocytose et des symptomes neurologi- ques, l'ab6talipoprot6in6mie. Aucune lipopro- t6ine contenant de l 'apo B n'est s6cr6t6e [chylo- microns, VLDL, LDL, Lp (a)].
Une variante de cette affection a 6t6 d6crite au cours de laquelle seuls les chylomicrons et l 'apo B48 sont normalement synth6tis6s et s6- cr6t6s. Les manifestations cliniques se limitent alors ~t une atteinte neurologique tr6s mod6r6e (39).
- - Les apo l ipoprot~ ines C- l , C-II , C-Ill (apo C-I, apo C-II, apo C-III) sont des peptides de faible masse mol6culaire qu'on trouve essen- tiellement dans les chylomicrons, les VLDL et les HDL, et qu'on peut s6parer par chromato- graphie d'6change ionique ou 61ectrophor~se en gel de polyacrylamide en milieu urge. L'iso- 61ectrofocalisation analytique diff~rencie au moins trois fractions d 'apo C-III (apo C-III0, apo C-III1, apo C-III2) en fonction de leur point iso61ectrique ; ces sous-unit6s qui ne dif- ferent que par leur contenu en acide sialique

Tome VI Valeur sdmiologique des apolipoprotdines 45 Numdro 1
peuvent ensuite etre quantifi6es par densitom6- trie (8). La structure de ces diff6rentes apopro- t6ines est totalement 61ucid6e :
-- L'apo C-I a une masse mol6culaire de 6550 et contient 57 amino-acides. Sa synth6se totale a 6t6 r6alis6e. Le peptide natif ou synth6- tique active la 16cithine-cholest6rol-acyltransf6- rase mais la signification physiologique de cette activation comparativement & celle de l 'apo A-I n'est pas connue.
-- L'apo C-II a une masse mol6culaire de 8900 et contient 78 amino-acides. C'est un co- facteur sp6cifique de la lipoprot6ine-lipase qui est transf6r6 des HDL2 aux lipoprot6ines riches en triglyc6rides afin d'acc616rer les pro- cessus de lipolyse. L'apo C-II poss6de d'autre part une affinit6 particuli6re pour les phospho- lipides. Des travaux r6cents r6alis6s & l'aide de peptides synth6tiques ont permis d'identifier trois sites diff6rents dans la mol6cule (11) :
un site de liaison aux phospholipides (r6gion 43-51) ;
un site de liaison ~ la lipoprot6ine-lipase (r6gion 75-78) ;
un site d'activation de la lipoprot6ine-li- pase (r6gion 55-65).
-- Les apo C-III de masse mol6culaire 9000 contiennent 79 amino-acides et portent 2, 1 ou 0 r6sidus d'acide sialique. Dans certaines conditions exp6rimentales, l 'apo C-III peut in- hiber la lipoprot6ine-lipase, mais on ne connait pas encore son mode d'action dans la r6gula° tion in vivo de la lipolyse.
Le m6tabolisme des apoprot6ines C-I, C-II et C-III est encore mal connu. Synth6tis6es principalement dans le foie et l'intestin, elles so.nt s6cr6t6es dans le sang o/t elles sont en 6quilibre entre les lipoprot6ines riches en tri- glyc6rides et les HDL. Les chylomicrons et les VLDL natifs regoivent un compl6ment d'apo- prot6ines C-II et C-III fourni par les H D L ; lors de la lipolyse, par la lipoprot6ine-lipase, les apoprot6ines C-II et C-III quittent l'6difice et se retrouvent en grande pattie dans les HDL. Un enrichissement en apoprot6ines c - i I et C-III des particules r6siduelles inhibe leur cap- tation par le foie et l 'hydrolyse des triglyc6rides restants par la triglyc6ride-lipase h6patique.
Les valeurs physiologiques habituelles dans le s6rum des sujets adultes sont respectivement pour les apo C-I, C-II, C-III comprises entre 50 et 110 mg/1, 10 et 67 mg/1 et 80 et 150 mg/1.
La concentration s6rique en apo C-II et apo C-III s'616ve dans les hyperlipoprot6in6mies de type III, IV et V de la classification internatio- nale. Une augmentation isol6e de l 'apo C-III
est observ6e au cours du diab6te non insulino- d6pendant ainsi qu'au cours de l 'insuffisance r6nale (2). Darts ce dernier cas une anomalie du catabolisme de l 'apo C-III, qui se fait en grande partie au niveau r6nal, pourrait expli- quer, du fait de l'action inhibitrice au niveau de la lipolyse, l 'accumulation de particules r6- siduelles riches en triglyc6rides observ6e dans cette affection (8, 20).
En utilisant un dosage immunologique de l 'apo C-III dans les lipoprot6ines pr6cipitables et non pr6cipitables par l'h6parine en pr6sence d'ions mangan6se, Alaupovic et coll. ont d6fini un rapport C-III-R (C-III des lipoprotbines non pr6cipitables assimil6es aux H D L / C - I I I des lipoprot6ines non pr6cipitables assimil6es aux lipoprot6ines de basse densit6) qui rend compte de l'efficacit6 des processus d'6pura- tion des triglyc6rides (1). Si le catabolisme des lipoprot6ines de basse densit6 se fait mal, l 'apo C-III ne peut retourner darts les H D L et le rap- port diminue. Ce rapport qui est normalement de 8 peut descendre ~t 0,05 au cours de cer- taines hypertriglyc6rid6mies majeures. I1 serait un param6tre discriminant pour le d6pistage de l'ath6roscl6rose coronarienne (1).
On a d6crit r6cemment quelques cas de d6f i cit g6n6tique en apo C-II qui se traduisent par une hypertriglyc6rid6mie majeure avec chylo- micron6mie, pouvant induire une pancr6atite aigue (4). I1 importe de rechercher ce d6ficit car les manifestations cliniques et biologiques r6- gressent spontan6ment en injectant au patient du plasma normal ou de l 'apo C-II synth6ti- clue.
Dans certains cas d'hyperlipoprot6in6mies de type IV ou V de la classification internatio- nale ou de dyslipoprot6in6mies secondaires (insuffisance r6nale, diabbte non insulino- d6pendant) le rapport apo C- I I /apo C-II1 des VLDL, qu'on peut 6valuer facilement par iso- 61ectrofocalisation analytique, est significative- ment diminu6 (8, 16). Cette modification et le d6ficit des processus de lipolyse qui en r6sulte pourraient 8tre responsables des accidents art6- riopathiques qui surviennent au long cours chez ces patients.
- - L ' a p o l i p o p r o t 6 i n e D (apo D) est une gly- coprot6ine trouv6e essentiellement dans les H D L qui pourrait jouer le r61e dans la synth6se et le transfert des esters de cholest6rol aux lipo- prot~ines. On ne connait pas encore la valeur s6miologique de cette apoprot6ine.
- - Les apoprot6ines E (apo E) repr6sentent l0 & 20 p. 100 de la pattie prot6ique des

46 J. C. Fruehart La Revue de Mddecine interne Janvier 1985
VLDL et des chylomicrons, mais sont pr6sentes aussi/t un degr6 moindre dans toutes les autres lipoprot6ines (en particulier dans les HDL1 et les particules r6siduelles ou ~<remnants)~ for- m6es lors du catabolisme des lipoprot6ines riches en triglyc6rides).
Elles oat une masse mol6culaire de 34145 et comportent 299 amino-acides (dont 35 argi- nine = ~arginine rich peptide) 0. L'iso61ectro- focalisation sdpare plusieurs formes d 'apo E dont 3 principales E-2, E-3, E-4 dont la s6- quence primaire eat connue et qui diff6rent par leur contenu en cyst6ine et en arginine.
E-2 : cyst6ine en 112 et 158, E-3 : cyst6ine en 112 et arginine en 158, E-4 : arginine en 112 et ~58.
En 1977, Utermann a d6couvert qu'il existait un polymorphisme g6n6tique de l 'apo E (41). Six ph6notypes diff6rents peuvent 6tre indivi- dualis6s en appliquant l'iso~lectrofocalisation analytique en une ou deux dimensions aux apo VLDL. Le polymorphisme est sous le contr61e de trois all61es codominants apo E-2, E-3 et apo E-4 conduisant /t trois ph6notypes homo- zygotes E-2/E-2, E-2/E-4 et E-4/E-4 et/~ trois ph6notypes h6t6rozygotes E-2/E-3, E-2/E-4 et E-3/E-4.
Plus r6cemment, deux autres variants d 'apo E-2 ont 6t6 d6crits. Ils se distinguent respective- ment de l 'apo E-3 par le remplacement de l'ar- ginine en 145 par une cyst6ine et de la lysine en 146 par une glutamine (38).
Les apo E ont une origine essentiellement h6.patique et sont s6cr6t6es sous forme de H D L nalssantes disco[dales. Elles passent ensuite sur les lipoprot6ines riches en triglyc6rides (chylomicrons et VLDL) et subsistent g la sur- face des particules r6siduelles ou ~remnants~ form6es lors de la lipolyse des triglyc6rides par la lipoprot6ine lipase. C'est grfice /~ l'affinit6 des apo E pour les r6cepteurs cellulaires que les ~remnants)) pourront 6tre capt6s et d6gra- d6s (21). Deux types de r6cepteurs reconnais- sent l 'apo E :
-- le premier est le classique LDL (B/E) r6- cepteur trouv6 dans les cellules p6riph6riques et aussi dans le foie,
- - le second, exclusivement h6patique, est connu sous le nom d'apo E-r6cepteur ou rem- nant-r6cepteur. La pr6sence d'une arginine en position 158 est essentielle pour que l 'apo E des ~remnants)) puisse se fixer sur ce r6cep- teur.
Les valeurs physiologiques habituelles dans le s6rum de sujets adultes sont pour l 'apo E comprises entre 30 et 50 mg/1.
On observe une augmentation nette de la concentration s6rique en apo E dans l'hyperli- poprot6in6mie familiale de type III qui s'ac- compagne de xanthomes palmaires et 6ruptifs et d'ath6roscl6rose pr6coce, coronarienne ou p6riph6rique. Alors que chez les sujets nor- maux, l 'apo E repr6sente approximativement 15 p. 100 de la totalit6 des apo VLDL, chez les sujets porteurs de cette dyslipoprot6in6mie tr6s ath6rog6ne, elle peut repr6senter 57 p. 100 de ces m6mes apoprot6ines. L'analyse par iso61ec- trofocalisation montre en outre que cette affec- tion atteint toujours des patients de g6notype E-2/E-2 (homozygotie E-2). Ce g6notype n'est retouv6 que dans 1 p. 100 de la population et seulement 2 p. 100 de ces patients d6velop- pent des signes cliniques d'hyperlipoprot6in6- mie de type III (40).
On ne sait pas encore parfaitement pourquoi dans 2 p. 100 des cas, u n homozygote E-2 d6- veloppe une hyperlip6mie (association de deux tares .9) et si les homozygotes E-2 sans hyperli- p6mie sont menac6s d'ath6roscl6rose.
Tr6s r6cemment Ghiselli et coll. ont sugg6re que le ph6notype E-4/E-4 6tait fr6quent chez les sujets atteints d'hyperlipoprot6in6mie fami- liale de type V mais l'existence et la significa- tion d'une telle association sont encore discu- tbes (18).
Quelques cas de d6ficits complets en apo E ont 6t6 d6crits. Ils se manifestent cliniquement comme une dyslipoprot6in6mie de type III avec ath6roscl6rose pr6matur6e et pr6sence de xanthomes tub6ro-6ruptifs (39).
-- L'apoproteine F (apo F) trouv6e dans les HDL est une apoprot6ine mineure de pHI bas dont la fonction et la valeur s6miologique sont inconnues.
- - L ' a p o p r o t ~ i n e H (apo H) qui poss6de une affinit6 marqu6e pour les chylomicrons est un activateur de la lipoprot6ine-lipase et son d6fi- cit g6n6tique peut induire une hyperchylomi- cron6mie.
- - L ' a p o p r o t 6 i n e Lp ( a + ) [apo Lp (a +)] est sp6cifique de la Lp (a + ) lipoprot6ine particu- li6re de densite comprise entre celle des LDL et des HDL et de migration pr6-b6ta en gel d'aga- rose. L'apo Lp ( a + ) existe chez tous les indivi- dus mais ~t des concentrations tr6s variables. Son m6tabolisme ne semble pas faire intervenir

Tome VI Valeur sdmiologique des apolipoprotdines 47 Numdro 1
une fixation de la Lp (a + ) sur les apo B, E r6- cepteurs, et est encore mal connu. Une concen- tration s6rique 61ev6e en apo Lp ( a + ) pr6dis- poserait/l l'ath6roscl6rose (23).
A la faveur de certains progr6s techniques et ~t la lumi6re de r6cents travaux, nos connais- sances sur les apoprot6ines ont beaucoup pro- gress6 et il serait souhaitable de revoir la classi- fication des dyslipoprot6in6rnies en tenant compte non seulement des lipides mais aussi de la pattie prot6ique des lipoprot6ines. La d6- termination du profil apoprot6inique peut ac- tuellement 6tre utilis6e comme crit6re de dia- gnostic e t /ou comme m6thode tr6s sensible de contr61e des th6rapeutiques normolip6miantes.
Dans un proche avenir, il serait souhaitable d'analyser les particules lipoprot6iques au ni- veau mol6culaire. Des travaux r6cents ont montr6 que ces particules existent dans le sang sous forme de lipides associ6s /l une, deux, trois ou quatre apoprot6ines (3, 10, 12, 13, 27, 26, 32). La classification particulaire des lipo- prot6ines, bas6e sur leur composition en apo- prot6ines, devrait supplanter celles pr6c6dem- ment utilis6es. L'introduction d'anticorps monoclonaux (27) et oligoclonaux (17) permet- tra sans aucun doute de s6parer et de quantifier les particules lipoprot6iques et de progresser ainsi consid6rablement dans la compr6hension des dyslipoprot6in6mies.
BIBLIOGRAPHIE
1. ALAUPOV1C P.: The role of apolipoproteins in lipid transport processes. Ric. Clin. Lab., 1982, 12, 3-21.
2. ALAUPOVIC P., MAC CONATttY W., FESMIRE J. : Apolipo- proteiri profiles of dyslipoproteinemic states. In : K. Lippel Eds. Proceedings of the Workshop on apolipopro- tein quantification, NIH Publication n ° 83-1266, 1983, p. 27-53.
3. ArMEH R.F., SHE~'rIERD J., PACKARD C.J. : Subpopula- tions of apolipoprotein A-I in human high density li- poproteins. Their metabolic properties and respons to drug therapy. Bioehim. Biophys. Aeta, t983, 751, 175-188.
4. BRECKINRIDGE W.C., LIIq'LE A., STEINER G., CHOW A., POAPST M. : Hypertrilgyceridemia associated with a de- ficiency of apolipoprotein C-II. N. Engl. J. Med., 1978, 298, 1265-1273.
5. BREWER H.B., FAIRWELL Jr T., KAY L., MENG M., RO- NAN R., LAW S., LIGHT J.A. : Human sequence proapo A-I: isolation and aminoterminal sequence. Biochem. Biophys. Res. Commun. 1983, 113/2, 626-632.
6. BREWER H.B., FAIRWELL J.T., MENG M., KAY L., RO- NAN R.: Human proapo A-I Tangier: isolation of proapo A-I Tangier and aminoacid sequence of the propeptide. Bioehem. Biophys. Res. Commun. 1983, 113, 934-940.
7. BROWN M.S., GOLDSTEIN J.L. : Lipoproteins receptors in the liver. J. Clin. Invest., 1983, 72, 743-747.
8. BUGUGNAN1 M.J., KOEF1GAN M., KORA I., OUVRY D., CLAYEY V., FRUCrlART J.C. : Rapid assessment of the distribution of apolipoproteins C-II and C-III subspe- cies in triglyceride rich lipoproteins by isolelectric fo- cusing. Clin. Chem., 1984, 30/3, 349-351.
9. CARLSON L.A., BALLANI~fNE D. : Changing relative pro- portions of apolipoprotein C-II and C-III of very low density lipoproteins in hypertriglyceridemia. Atheros- clerosis, 1976, 23, 563-568.
10. CASTRO G.R., FIELDING C.J. : Evidence for the distribu- tion of apolipoprotein E between lipoprotein classes in human normocholesterolemic plasma and for the ori- gin of unassociated apolipoprotein E (LpE). J. Lipid Res., 1984, 25, 53-67.
11. CATAPANO A.L. : Apo C-II and LPL activity. Ric. Clin. Lab., 1982, 12, 35-40.
12. CHEUNG M.C., ALBERS J.J. : Distribution of high den- sity lipoprotein particles with different apoprotein composition : particles with A-I and A-II and particles with A-I but no A-II. d. Lipid Res., 1982, 23, 747-753.
13. CHUNG HON G.B., SEGREST J.P. : Resistance of a very low density lipoprotein subpopulation from familial dysbetatipoproteinemia to in vitro lipolytic conversion to the low density lipoprotein density fraction. J. Lipid Res., 1983, 24, 1148-1159.
14. DOUSTE-BLAZY P., SCHEIDEGGER B., CAMARE R., MAR- CEL Y.L., MARCO J., BERNADET P. : Lipoprotein and apoprotein values in coronary angiography normocho- lesterolemic male patients. In : J.L. De Gennes et al. Eds. Latent Dyslipoproteinemias and dtherosclerosis, New-York, 1984, p. 179-186, Raven Press.
15. EDELSTEIN C., GORDON J.I., TOSCAS K., SIMS H.F., STRAUSS A.W., SCANU A.M. : In vitro conversion of proapoprotein A-I to apoprotein A-I. J. BioL Chem., 1983, 258, 11430-11433.
16. ERKELENS D.W., MOCKING J.A.J. : The C- I I /C- I I I ratio of transferable apolipoprotein in primary and secon- dary hypertriglyceridemia. Clin. Chim. Acta, 1982, 121, 59-65.
17. GESQUIERE J.C., CACHERA C., DELPIERRE C., KODJO E.; TARTAR A., FRUCHART J.C. : Production of oligoclonal antibodies against apolipoproteins C-II and E. Clin. Chem., 1984, in press.
18. GHISELLI G,, SCHAEFER E.J., GASCON P., BREWER H.B. Jr. : Increased prevalence of apolipoprotein E-4 in type V hyperlipoproteinemia. J. Clin. Invest., 1982, 70, 474-479.
19. GORDON J.I., BUDELIER K.A., SIMS H,F., EDELSTE~N C., SCANU A.M. , STRAUSS A.W. : Biosynthesis of human preproapolipoprotein A-IL J. Biol. Chem., 1983, 258, 14054-14059.
20. HOLDSWORTH G., STOCKS J., DODSON P., GALTON D.J. : An abnormal triglyceride-rich lipoprotein containing excess sialytated apolipoprotein C-III. J. Clin. Invest., 1982, 69, 932-939.

48 J .C. Fruchart La Revuede Mddeeine interne Janvier 1985
21. INNERARITY T.L., FRIEDLANDER E.J., RALL S.C., WEIS- GRABER K.H. , MAHLEY R.W. : The receptor binding do- main of human apolipoprotein E binding of apolipo- protein E fragments. J. BioL Chem., 1983, 258, 12341-12347.
22. JAHN C.E., OSBORNE J.C., SCHAEFER E.J., BREWER H.B. : Activation of the enzyme activity of hepatic lipase by apolipoprotein A-II. Cur. J. Biochem., 1983, 131, 25-29.
23. JAILLARD J., SEZILLE G., DEWAILLY P., FRUCHART J.C. : D6tection d'une lipoprot6ine particuli6re; la Lp(a). Son incidence darts l'ath6roscl6rose avec ou sans hyperlip6mie associ6e. Ann. Mdd. Interne, 1977, 128, 739-744.
24. I~NE J.P. : Apolipoprotein B : structural and metabolic heterogeneity. Ann. Rev. Physiol., 1983, 45, 637-650.
25. KORA 1., VANHOUTTE G., PARRA H., BERMANN M.C., CACHERA C., CLAVEY V., PUCHOIS P., FRUCHART J.C. : Dosage de l'apolipoprot6ine A-II s6rique par 61ec- troimrnunodiffusion sur plaques pr6tes ~ l'emploi. Ap- plications cliniques. Ann. BioL Clin., 1984, in press.
26. KOREN E., SOLTER D., KNIGHT C., ALAUPOVIC P. : Isola- tion of apolipoprotein B containing lipoproteins by the use of monoclonal antibodies. Circulation, 1983, 68: 4, II, p. 111-217.
27. KOREN E., MACCONATHY W.J., ALAUPOVIC P. : Isola- tion and characterization of apolipoprotein F contai- ning lipoproteins from human plasma. Biochem., 1982, 21, 5347-5351.
28. KWITEROVICH P., SNIDERMAN A. : Clinical and bioche- mical characteristics of dyslipoproteinemias associated with an excess of apoprotein B or A-I. In : K. Lippel Eds. Proceedings of the Workshop on apolipoprotein quantification, NIH Publication no83-1266, 1983~ p. 71-100.
29. LOPES A,, VIAL R., BALART L., ARROYAVE G. : Effect of exercise and physical fitness on serum lipids and lipo- protein. Atherosclerosis, 1974, 20, 1-16.
30. MACIEJKO J.J., HOLMES D.R., KOTTKE B.A., ZINMEISTER A.R., DINH D.M., MAO S.J.T. : Apolipoprotein A-I as a marker of angiographically assessed coronary artery disease. N. Engl. J. Med., 1983, 31)9, 385-389.
31. MAHLEY R.W. : Apolipoprotein E and cholesterol me- tabolism. Klin. Wochenschr., 1983, 61,225-232.
32. NESTEL P., BILLINGTON T., TADA N., NUGENT P., FIDGE N. : Heterogeneity of very low density lipoprotein me- tabolism in hyperlipemic subjects. Metabolism, 1983, 32, 810-817.
33. NIKKILA E.A',: Metabolic regulation of plasma high density lipoprotein concentrations. Cur. d. Clin. Invest., 1978,8, 111-123.
34. NORUM R.A., LAKIER J.Bo, GOLDSTEIN S., ALAUPOV1C P. : Familial deficiency of apolipoproteins A-I and C- III and precocious coronary artery disease. N. Engl. J. Med., 1982, 306, 1513-1519.
35. PARRA H., FIEVET C., BONIFACE B., BERTRAND M., DU- THILLEUL P., FRUCHART J.C. : Lipoproteins, apotipopro- teins and coronary artery disease assessed by coronary arteriography. In: J.L. De Gennes et al Eds. Latent Dyslipoproteinemias and Atherosclerosis, New-York, 1981, p. 187-197, Raven Press.
36. PATSCH W., KUISK I., GLUECK C., SCHONFELD G. : t ipo- proteins in familial hyperalphalipoproteinemia. Arte- riosclerosis, 1981, 1, 156-161.
37. PUCHOIS P., FONTAN M., GENTILINI J.L., GELEZ P., FRU- CHART J.C. : Serum apolipoprotein A-II, a biochemical indicator of alcohol abuse. Clin. Chem. Acta, 1984, in press.
38. RALL S.C. Jr., WEISGRABER K.H., INNERARITY T.L., MAHLEY R.W. : Structural basis for receptor binding heterogeneity of apolipoprotein E from type III hyper- lipoproteinemic subjects. Proc. Natl. Acad. Sci., USA, 1982, 79, 4696-4700.
39. SCHAEFER E.J. : Disorders of lipoprotein metabolism associated with apolipoprotein deficiencies. In: K. Lippel Eds. Proceedings of the Workshop on apolipopro- tein quantification, NIH Publication n ° 83-1266, 1983, p. 54-70.
40. UTERMANN G., JAESCHKE M., MANGEL J. : Familial hyperlipoproteinemia type III : deficiency of a specific apolipoprotein (apo E-III) in the very low density lipo- proteins. FEBS Lett., 1975, 55, 352-355.
41. UTERMANN G., HEES M., STEINMETZ A. : Polymorphism of apolipoprotein E and occurence of dysbetalipopro- teinemia in man. Nature, 1977, 269, 604-607.