Embed Size (px)
Citation preview

Correspondances en Métabolismes Hormones Diabètes et Nutrition - Vol. XVII - n° 7 - septembre 2013208208
d o s s i e r t h é m a t i q u e
Surrénales : nouvelles approches
dddd oodd odddddddddddd s ss ss sss ssss sss s iiii ei ei ei ee rrr t ht ht ht ht h ééé mmmé méé mééé m a ta tttt i qqi qii q u eu euuu
SSuurrénnaalleess : nnnoooouuuuvvvveelllleess aapprocchhes
Anomalies génétiques dans les adénomes de Conn et l’hyper-aldostéronisme familial : implications pour la prise en charge et le traitementGenetic abnormalities in Conn’s adenoma and familial hyper-aldosteronism: implications for diagnosis and treatmentMaria-Christina Zennaro1, 2, 3, Sheerazed Boulkroun1, 3
po
int
s f
or
ts
Hig
hl
igh
ts
» Ces deux dernières années ont été marquées par des avancées formidables dans nos connaissances des bases génétiques des adénomes de Conn et de l’hyperaldostéronisme familial.
» Grâce à des techniques de séquençage à haut débit de tous les exons codants du génome et l’analyse de cohortes de centaines de patients par plusieurs consortiums, 3 nouveaux gènes ont été identifi és, dont les mutations somatiques sont retrouvées dans environ la moitié des adénomes de Conn.
» Des mutations constitutionnelles similaires sont transmises de façon autosomique dominante dans l’hyperaldostéronisme familial de type 3.
» L’ensemble de ces mutations aff ecte le maintien de l’équilibre ionique intracellulaire, ce qui peut être considéré maintenant comme le mécanisme principal de dérégulation de la production d’aldostérone dans la surrénale.
» Ces découvertes éclaircissent notre compréhension des mécanismes pathogéniques de la maladie et pourraient ouvrir la voie au développement de nouveaux traitements.
Mot-clés : Aldostérone – Surrénale – Adénome – Canal potassique – ATPase.
In the past two years, significant progress has been made in the understanding of the genetic bases of aldosterone producing adenoma and familial hyperaldosteronism.
Whole exome sequencing and analysis of a large number of patients by different consortia have identified somatic mutations in three new genes in approximately 50% of aldosterone producing adenomas.
Similar germline mutations are transmitted as an autosomal dominant trait in familial hyperaldosteronism type 3.
All these mutations affect the maintenance of intracellular ionic homeostasis, which can be considered the principal mechanism leading to dysregulated aldosterone production in the adrenal gland.
These discoveries increase our understanding on the pathogenic mechanisms of the disease and may open new perspectives for the development of new treatment options.
Keywords : Aldosterone – Adrenal – Adenoma – Potassium channel – ATPase.
L’ hyperaldostéronisme primaire (HAP) est la conséquence d’une production exces-sive d’aldostérone par la surrénale, res-
ponsable d’hypertension artérielle (HTA) souvent résistante et accompagnée d’hypokaliémie parfois sévère. Il s’agit de la forme la plus commune d’HTA secondaire. La prévalence de l’HAP chez les sujets hypertendus est estimée à environ 10 % des patients référés dans des centres spécialisés et jusqu’à 7 % dans des études de population (1, 2). Cependant, la
prévalence de la maladie augmente avec la sévérité de l’HTA et atteint jusqu’à 20 % chez les patients avec HTA résistante (3, 4). Le diagnostic précoce d’HAP a un impact très important sur l’issue théra-peutique et la survie, étant donné les effets cardio-vasculaires délétères de l’excès d’aldostérone et la possibilité d’un traitement étiologique chirurgical (5, 6). Les patients avec HAP présentent des formes plus sévères d’hypertrophie ventriculaire gauche et d’altération de la fonction diastolique que les
1 Inserm, UM970, Paris-
Centre de recherche cardiovasculaire (PARCC).
2 Université Paris-Descartes
Sorbonne Paris cité.3 AP-HP, service de géné-
tique, hôpital européen Georges-Pompidou, Paris.
Anomalies génétiques dans les adénomes de Conn et l’hyper aldostéronisme familial : implications pour la prise en charge et le traitement
patients avec HTA essentielle et plus d’événements cardiaques, notamment infarctus du myocarde, fibrillation auri culaire et accidents vasculaires céré-braux (7-9). Parmi les différentes formes d’HAP, les adénomes produisant de l’aldostérone (APA, ou adénomes de Conn) et l’hyperplasie bilatérale des surrénales (BAH) représentent ensemble environ 95 % des cas. Selon la population étudiée et les procédures diagnostiques utilisées, la proportion de patients avec APA peut atteindre 60 % (10, 11).Dans la glande surrénale, l’aldostérone est synthétisée au niveau de la zone glomérulée du cortex par une série d’étapes enzymatiques, alors que le cortisol l’est au niveau de la zone fasciculée (12). Les étapes fi nales de la biosynthèse de l’aldostérone sont catalysées par une enzyme, l’aldostérone synthase (codée par le gène CYP11B2), alors que la 11β-hydroxylase (codée par CYP11B1) est responsable des dernières étapes de la biosynthèse du cortisol. Ces enzymes sont très homologues et leurs gènes sont localisés en tandem sur le chromosome 8q21-q22.
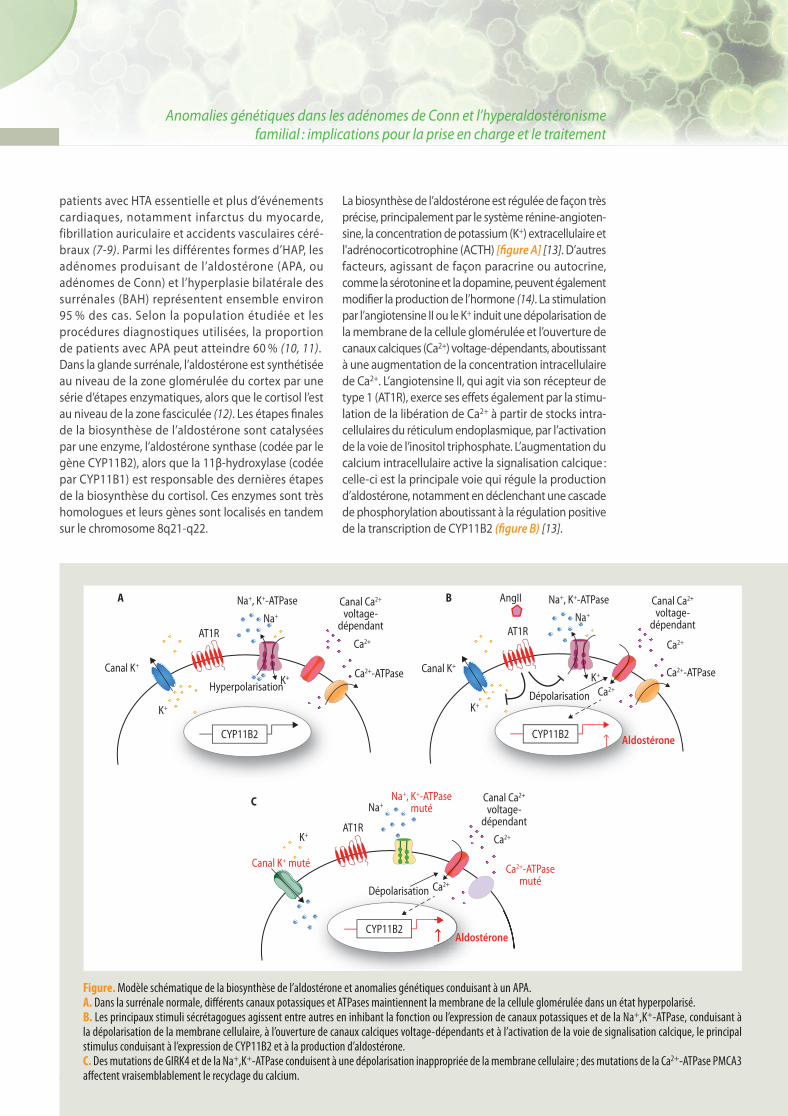
La biosynthèse de l’aldostérone est régulée de façon très précise, principalement par le système rénine-angioten-sine, la concentration de potassium (K+) extracellulaire et l'adrénocorticotrophine (ACTH) [fi gure A] [13]. D’autres facteurs, agissant de façon paracrine ou autocrine, comme la sérotonine et la dopamine, peuvent également modifi er la production de l’hormone (14). La stimulation par l’angiotensine II ou le K+ induit une dépolarisation de la membrane de la cellule glomérulée et l’ouverture de canaux calciques (Ca2+) voltage-dépendants, aboutissant à une augmentation de la concentration intracellulaire de Ca2+. L’angiotensine II, qui agit via son récepteur de type 1 (AT1R), exerce ses eff ets également par la stimu-lation de la libération de Ca2+ à partir de stocks intra-cellulaires du réticulum endoplasmique, par l’activation de la voie de l’inositol triphosphate. L’augmentation du calcium intracellulaire active la signalisation calcique : celle-ci est la principale voie qui régule la production d’aldostérone, notamment en déclenchant une cascade de phosphorylation aboutissant à la régulation positive de la transcription de CYP11B2 (fi gure B) [13].
Figure. Modèle schématique de la biosynthèse de l’aldostérone et anomalies génétiques conduisant à un APA. A. Dans la surrénale normale, diff érents canaux potassiques et ATPases maintiennent la membrane de la cellule glomérulée dans un état hyperpolarisé. B. Les principaux stimuli sécrétagogues agissent entre autres en inhibant la fonction ou l’expression de canaux potassiques et de la Na+,K+-ATPase, conduisant à la dépolarisation de la membrane cellulaire, à l’ouverture de canaux calciques voltage-dépendants et à l’activation de la voie de signalisation calcique, le principal stimulus conduisant à l’expression de CYP11B2 et à la production d’aldostérone. C. Des mutations de GIRK4 et de la Na+,K+-ATPase conduisent à une dépolarisation inappropriée de la membrane cellulaire ; des mutations de la Ca2+-ATPase PMCA3 aff ectent vraisemblablement le recyclage du calcium.
Canal K+
Canal K+ muté
Canal K+
AT1R AT1R
AT1R
AngIINa+, K+-ATPase
Na+
Na+
Na+, K+-ATPase
Na+, K+-ATPasemuté
Na+
K+ K+
K+
K+K+
CYP11B2 CYP11B2
CYP11B2
Aldostérone
Aldostérone
Canal Ca2+
voltage-dépendant
Canal Ca2+
voltage-dépendant
Ca2+-ATPase
Ca2+-ATPasemuté
Canal Ca2+
voltage-dépendant
Ca2+-ATPaseHyperpolarisation
Dépolarisation
Dépolarisation
A B
C
Ca2+
Ca2+
Ca2+ Ca2+
Ca2+

Correspondances en Métabolismes Hormones Diabètes et Nutrition - Vol. XVII - n° 7 - septembre 2013210210
d o s s i e r t h é m a t i q u e
Surrénales : nouvelles approches
dddd oodd odddddddddddd s ss ss sss ssss sss s iiii ei ei ei ee rrr t ht ht ht ht h ééé mmmé méé mééé m a ta tttt i qqi qii q u eu euuu
SSuurrénnaalleess : nnnoooouuuuvvvveelllleess aapprocchhes
Formes familiales d’hyperaldostéronisme primaire
Bien que la plupart des cas d’HAP soient sporadiques, 1 à 6 % sont des formes familiales. À ce jour, 3 formes dif-férentes transmises de façon autosomique dominante ont été décrites, et, pour 2 d’entre elles (FH-I et FH-III), le défaut génétique sous-jacent élucidé : l’hyperaldos-téronisme familial de type I (FH-I), de type II (FH-II) et de type III (FH-III).
Hyperaldostéronisme familial de type IEn 1966, le cas d’un père et de son fi ls présentant une HTA associée à une hypokaliémie et un tableau bio-logique d’hyperaldostéronisme primaire a été décrit (15). De façon remarquable, toutes les anomalies étaient réversibles par un traitement avec dexaméthasone. Le FH-I, que l’on appelle également hyperaldostéronisme suppressible par les glucocorticoïdes (glucocorticoid suppressible aldosteronism [GRA]), est transmis de façon autosomique dominante et il se caractérise par une HTA précoce et sévère, le plus souvent avant l’âge de 20 ans (16). Les patients présentent des tableaux d’HAP de sévérité variable, avec, dans certains cas, des adénomes, et une production importante de stéroïdes hybrides, 18-hydroxycortisol et 18-oxocortisol, détectables dans les urines (14). Le FH-I est causé par un gène hybride résultant d’un enjambement inégal entre les gènes adja-cents CYP11B1 et CYP11B2, dans lequel les séquences codant pour l’aldostérone synthase sont placées sous le contrôle des séquences régulatrices de CYP11B1. Ainsi, ce gène chimérique place la synthèse d’aldostérone sous le contrôle de l’ACTH au lieu de l’angiotensine II, avec une expression qui s’étend à travers l’ensemble du cortex surrénalien (17, 18). De la régulation de la production d’aldostérone par l’ACTH résulte également une production rythmique circadienne d’aldostérone, similaire à celle du cortisol (19). Le traitement par des glucocorticoïdes exogènes, qui diminuent la production d’ACTH, réduit de façon effi cace l’hyperaldostéronisme et corrige le tableau clinique. Il est important d’utiliser la dose minimale effi cace de glucocorticoïdes, permettant de normaliser la pression artérielle et la kaliémie sans nécessairement inhiber complètement l’ACTH, afi n de ne pas supprimer la régulation circadienne du cortisol. En eff et, une dose trop élevée peut induire un syndrome de Cushing iatrogénique et réduire la croissance linéaire chez l’enfant (20). Il a été démontré qu’un traitement à des doses faibles de glucocorticoïdes (dexaméthasone [0,125-0,25 mg/j] ou prednisone [2,5 ou 5 mg/j]) pouvait contrôler l’hypertension de façon prolongée pendant plusieurs années, tout en maintenant des paramètres
échocardiographiques normaux (19). L’ajout d’un inhi-biteur minéralocorticoïde peut être envisagé dans les cas où le contrôle tensionnel est insatisfaisant. Chez les enfants, l’utilisation d’éplérénone est préférable afi n d’éviter les eff ets collatéraux des glucocorticoïdes (retard de croissance) ou de la spironolactone (eff ets antiandrogéniques) [20].La prévalence du FH-I est estimée entre 0,66 % et 1 % (18, 21), mais peut être bien plus élevée et atteindre 3,1 % dans des cohortes pédiatriques hypertendues (22). Selon les recommandations de l’Endocrine Society, le dépistage génétique du FH-I doit être envisagé chez des patients jeunes avec une HTA sévère ou résistante et une histoire familiale d’HTA précoce ou d’AVC hémorra-gique prématuré (20). Cependant, il existe une certaine variabilité intra- et interfamiliale du phénotype du FH-I, et la sévérité de l’HTA peut être variable au sein d’une même famille (21). Cette variabilité dépend d’autres facteurs héréditaires impliqués dans la régulation de la pression artérielle et de facteurs environnementaux, comme le régime sodé. Ainsi, l’histoire familiale de FH-I ne révèle pas de façon systématique une HTA sévère chez les apparentés de premier degré. Le test génétique de dépistage est réalisé par Southern blot ou long-range PCR, les 2 techniques sensibles et spécifi ques pour établir le diagnostic de FH-I. Ainsi, l’analyse génétique permet de se substituer à la mesure des taux urinaires de 18-oxo-cortisol et de 18-hydroxycortisol ainsi qu’à la nécessité de réaliser un test de suppression à la dexaméthasone, les 2 tests pouvant conduire à des diagnostics erronés (20).
Hyperaldostéronisme familial de type IIUne deuxième forme d’hyperaldostéronisme familial, non suppressible par les glucocorticoïdes, a été décrite en 1991 (23). Comme le FH-I, cette forme est transmise de façon autosomique dominante. Les patients avec FH-II présentent une réponse variable de l’aldostérone au test postural et à l’angiotensine II, et, au sein de la même famille, il est fréquent de retrouver diff érents sous-types d’HAP (APA ou BAH). En eff et, les patients avec FH-II ne se distinguent en rien de patients atteints d’HAP sporadique, et le diagnostic de FH-II repose exclu-sivement sur la présence de 2 ou plusieurs membres atteints dans une même famille. Une certaine variabi-lité phénotypique est également caractéristique de la maladie (21, 24). Parmi les formes familiales, le FH-II est la forme la plus fréquente, avec une prévalence esti-mée autour de 2,8 % à 6 % dans des populations adultes avec HAP (21, 24, 25). Le défaut génétique sous-jacent est encore inconnu, bien qu’un locus génétique ait été identifi é sur le chromosome 7p22 dans un certain nombre de familles atteintes de FH-II (26). Cependant,

Correspondances en Métabolismes Hormones Diabètes et Nutrition - Vol. XVII - n° 7 - septembre 2013 211211
Anomalies génétiques dans les adénomes de Conn et l’hyper aldostéronisme familial : implications pour la prise en charge et le traitement
le séquençage de gènes candidats localisés dans cette région, notamment les gènes codant pour la fascine 1 (FSCN1) et la sous-unité régulatrice β de la protéine kinase de type I cAMP-dépendante (PRKAR1B) n’a pas mis en évidence de mutation causale (24). Par ailleurs, aucune anomalie génétique n’a été identifi ée dans les gènes codant pour l’aldostérone synthase (CYP11B2), pour le récepteur à l’angiotensine II de type 1 (AT1R) ni dans le gène suppresseur de tumeur (27). Il est à noter que des mutations somatiques de KCNJ5 (voir p. 212) ont été décrites récemment dans des adénomes de Conn chez des patients avec un hyperaldostéronisme familial non suppressible par les glucocorticoïdes classés comme atteints de FH-II (28). Cela évoque la possibilité que, dans un certain nombre de cas, le FH-II soit dû à une agrégation familiale d’HAP sporadique, étant donné la fréquence élevée de la maladie parmi les sujets hyper-tendus. Par ailleurs, le diagnostic génétique a permis de reclasser un certain nombre de familles FH-II en FH-III, ce qui révèle que cette dernière forme d’HAP familiale a une hétérogénéité phénotypique insoupçonnée (28).
Hyperaldostéronisme familial de type IIIPlus récemment, une nouvelle forme d’HAP a été décrite (29). Les membres atteints de la famille princeps présen-taient une HTA sévère, d’apparition très précoce chez l’enfant, et résistante au traitement, accompagnée d’une hypokaliémie profonde. On retrouvait également des taux élevés de stéroïdes hybrides, 18-oxocortisol et 18-hydroxycortisol, dans les urines, mais la production d’aldostérone n’était pas supprimée par la dexamétha-sone. L’hyperaldostéronisme était dû à une hyper plasie bilatérale massive des surrénales, nécessitant une sur-rénalectomie bilatérale pour contrôler la pression arté-rielle (29). L’origine génétique du FH-III a été attribuée très récemment à la présence d’une mutation gain de fonction du gène KCNJ5, qui code pour le canal potas-sique GIRK4 (G protein-activated inward rectifi er potassium channel 4) [30]. Cette mutation p.T158A, qui remplace la thréonine en position 158 par une alanine, est loca-lisée juste au-dessus du fi ltre de sélectivité du canal et conduit à une perte de la sélectivité ionique au K+ avec une conductance accrue pour le Na+. Étant donné que le potentiel de membrane de la cellule glomérulée est voisin du potentiel du repos du potassium, l’augmenta-tion de la conductance au sodium conduit à la dépolari-sation de la membrane cellulaire, l’ouverture de canaux calciques voltage-dépendants et l’activation de la signa-lisation calcique, aboutissant à une augmentation de la production d’aldostérone (fi gure C, p. 209) [13, 31]. Par la suite, d’autres mutations et un certain degré de variabilité phénotypique ont été décrits dans le FH-III. Notamment,
une mutation constitutionnelle héréditaire, p.G151R, a été identifi ée dans 2 familles avec une forme précoce et sévère d’hyperaldostéronisme et une hyperplasie bilaté-rale massive des surrénales (32), et un tableau clinique similaire était également associé à la mutation p.I157S (33). À l’inverse, une autre mutation constitutionnelle, p.G151E, était associée à un hyperaldostéronisme familial modéré dans 3 familles (28, 32). De façon intéressante, toutes ces mutations sont localisées dans la même région autour du fi ltre de sélectivité du canal GIRK4, conduisant à des modifi cations fonctionnelles similaires de l’activité du canal. Cependant, les patients porteurs de la muta-tion p.G151E présentent des symptômes modérés par rapport aux patients porteurs des autres mutations, avec une HTA et une hypokaliémie facilement contrôlables par un traitement médical et sans signe d’hyperplasie des surrénales à l’imagerie. Cette présentation clinique, qui ressemble plutôt à un FH-II, a mis en évidence une hétérogénéité phénotypique du FH-III. Des investigations génétiques supplémentaires dans des familles atteintes pourront révéler toute l’étendue du spectre clinique de cette forme d’HAP familial.
Canaux potassiques et ATPases : la nouvelle biologie de l’HAP sporadique
Le maintien du potentiel de membrane de la cellule glomérulée est un mécanisme central de la régulation de la production d’aldostérone (fi gure A) [13]. Dans les cellules de la zone glomérulée, la conductance principale est celle du potassium, étant donné que de nombreux canaux potassiques sont exprimés dans ces cellules. Ainsi, le potentiel de membrane suit de près le potentiel d’équilibre du K+ dans un large intervalle de concentra-tions extracellulaires de K+. Parmi d’autres canaux, le rôle physiologique des canaux TASK (TWIK-related acid-sensitive potassium channel) a été mis en évidence dans diff érents modèles murins d’inactivation, qui présentent tous des anomalies de la régulation de la production d’aldostérone, avec des tableaux d’hyperaldostéronisme variables (34-36). Le gradient de concentration du K+
entre l’espace intracellulaire et extracellulaire, requis pour l’établissement du potentiel de membrane, est généré par l’activité de la pompe Na+-K+ ATPase, qui transporte 2 ions K+ vers l’intérieur de la cellule contre 3 ions Na+ vers l’extérieur. Des mutations dans diff érents gènes codant pour des protéines impliquées dans la régulation du potentiel de membrane des cellules de la zone glomérulée et l’homéostasie ionique ont récem-ment été identifi ées dans l’ADN tumoral (somatique) de patients avec adénome de Conn sporadique.

Correspondances en Métabolismes Hormones Diabètes et Nutrition - Vol. XVII - n° 7 - septembre 2013212212
d o s s i e r t h é m a t i q u e
Surrénales : nouvelles approches
dddd oodd odddddddddddd s ss ss sss ssss sss s iiii ei ei ei ee rrr t ht ht ht ht h ééé mmmé méé mééé m a ta tttt i qqi qii q u eu euuu
SSuurrénnaalleess : nnnoooouuuuvvvveelllleess aapprocchhes
Au-delà des mutations constitutionnelles dans le FH-III, des mutations somatiques récurrentes de KCNJ5 ont été identifi ées dans une proportion considérable d’APA sporadiques (30, 37). Comme pour les mutations constitutionnelles, les mutations p.G151R et p.L168R se trouvent à proximité ou à l'intérieur du fi ltre de sélec-tivité de GIRK4 ; elles se traduisent par une perte de la sélectivité ionique du canal, avec une conductance au sodium augmentée entraînant une dépolarisation mem-branaire (fi gure C) [13]. Ces mutations sont retrouvées dans 34 à 47 % des APA dans les pays occidentaux (37-40), et jusqu’à 65 % chez des patients japonais (41). Une étude européenne, portant sur 380 patients avec APA et 174 avec BAH, recrutés par le réseau français COMETE (Cortico- et MÉdullo-surrénale, les Tumeurs Endocrines) et des centres de référence du réseau ENS@T (European Network for the Study of Adrenal Tumors), a montré que des mutations somatiques de KCNJ5 étaient signifi cativement plus fréquentes chez les femmes et les sujets jeunes et étaient associées à des taux d’aldostérone préopéra-toires plus élevés, mais pas à la réponse thérapeutique postchirurgicale (37). Bien que des mutations hérédi-taires de KCNJ5 soient responsables de FH-III, aucune des mutations récurrentes n’a été retrouvée sur l’ADN constitutionnel de sujets avec BAH. Il est également à noter que la présence de ces mutations est associée à des taux plus élevés d’aldostérone au cathétérisme des veines surrénaliennes, ce qui pourrait introduire un biais diagnostique et conduire à en surestimer la fréquence, dans la mesure où ces patients seraient plus aisément sélectionnés pour une intervention chirurgicale (42).Début 2013, des mutations somatiques de 2 nouveaux gènes, ATP1A1 et ATP2B3, codant respectivement pour la sous-unité α1 de la Na+-K+ ATPase et la pompe calcique membranaire PMCA3, ont été identifi ées dans les APA (13, 43). Comme pour GIRK4, les mutations sont retrou-vées dans des régions très conservées impliquées dans l’interaction avec les cations transportés, le potassium pour la Na+-K+ ATPase et le calcium pour PMCA3, ce qui souligne le rôle fondamental de l’équilibre ionique intra-cellulaire et du maintien du potentiel de membrane dans la pathogenèse de l’APA. Deux mutations du gène ATP1A1 ont été identifi ées : une mutation p.Leu104Arg et une substitution p.Val332Gly. Une petite délétion, p.Phe100_Leu104del, chevauchant la Leu104, a éga-lement été mise en évidence. Diff érentes délétions en phase chevauchantes ont été identifi ées dans le gène ATP2B3 ; celles-ci ont comme conséquence la délétion de 2 acides aminés (p.Leu425_Val426del ou p.Val426_pVal-427del). Les deux ATPases sont fortement exprimées dans le cortex de la surrénale. Des études in vitro ont montré que les mutations d’ATP1A1 diminuent de façon signifi -
cative l’activité physiologique de la Na+-K+ ATPase ainsi que l’affi nité de la pompe pour le potassium. Des mesures électrophysiologiques réalisées sur des cultures primaires de cellules dérivées d’adénome de Conn ont également montré que les cellules porteuses de mutations de la Na+-K+ ATPase présentent une dépolarisation inappro-priée de la membrane cellulaire. Ainsi, les mutations de la Na+-K+ ATPase conduisent, comme les mutations aff ectant GIRK4, à une dépolarisation inappropriée de la membrane cellulaire responsable d’une augmentation de l’infl ux de calcium dans la cellule ; les mutations de la Ca2+ ATPase PMCA3, quant à elles, aff ectent vraisem-blablement le recyclage du calcium. L’augmentation de la signalisation calcique dans la cellule conduit à la production autonome d’aldostérone (fi gure C) [13].L’étude de 309 échantillons a permis également de démontrer que des mutations somatiques de ces ATPases sont retrouvées dans environ 7 % des APA, exclu-sivement dans des tumeurs ne portant pas de mutations de KCNJ5. À l’inverse des mutations de KCNJ5, elles sont plus fréquentes chez l’homme que chez la femme. Elles sont également associées à des taux préopératoires d’aldostérone plus élevés et à une kaliémie plus basse (43). Ainsi, la découverte de l’implication de ces deux nouveaux gènes dans la pathogenèse de l’APA permet désormais de connaître l’origine de la maladie dans près de 50 % des cas d’adénome de Conn sporadique. Elle off re également la possibilité d’établir un modèle physio-pathologique du développement de l’adénome de Conn (fi gure) [13], associé avec des prévalences diff érentes selon le sexe et un tableau clinique plus sévère d’HAP.
Conclusion
Un diagnostic rapide et effi cace d’HAP, ainsi que l’iden-tifi cation des sous-types cliniques et génétiques, est essentiel pour la mise en place d’un traitement effi cace visant à prévenir les eff ets cardiovasculaires adverses de l’aldostérone. Les formes familiales d’HAP représentent 1 à 6 % des cas. Le FH-I doit être considéré chez les patients jeunes ayant un HAP et une histoire familiale d’hypertension, d’HAP ou d’AVC précoces ; le FH-III doit être suspecté chez le sujet jeune, particulièrement l’enfant, chez qui a été exclu le diagnostic de FH-I, pré-sentant un hyperaldostéronisme et une hypokaliémie sévères, et une HTA résistante au traitement médical. Dans les 2 cas, le diagnostic génétique est sensible et spécifi que, et peut guider de façon effi cace la prise en charge thérapeutique. À l’inverse, le FH-II ne peut être distingué d’un HAP sporadique, et aucun test géné-tique n’est disponible pour l’instant. La recherche de

Correspondances en Métabolismes Hormones Diabètes et Nutrition - Vol. XVII - n° 7 - septembre 2013 213213
Anomalies génétiques dans les adénomes de Conn et l’hyper aldostéronisme familial : implications pour la prise en charge et le traitement
mutations somatiques de KCNJ5, d’ATP1A1 et d’ATP2B3 dans les adénomes de Conn pourrait avoir un bénéfi ce clinique pour le patient dans le futur, une fois que des études prospectives auront évalué l’association de ces mutations avec des paramètres liés à la réponse thérapeutique postchirurgicale. L’identifi cation d’un autre gène dans l’HAP sporadique et familial vient d’être annoncée au congrès de 2013 de l’Endocrine Society par 2 groupes diff érents ; ce travail est sous presse au moment de la rédaction de cet article. Il est envisa-geable que l’identifi cation des anomalies génétiques responsables d’HAP aboutissent au développement de nouvelles approches thérapeutiques, qui pourraient apporter un bénéfi ce à une proportion considérable de patients hypertendus. ■
1. Plouin PF, Amar L, Chatellier G. Trends in the prevalence of primary aldosteronism, aldosterone-producing adenomas, and surgically correctable aldosterone-dependent hypertension. Nephrol Dial Transplant 2004;19(4):774-7.
2. Hannemann A, Bidlingmaier M, Friedrich N et al. Screening for primary aldosteronism in hypertensive subjects: results from two German epidemiological studies. Eur J Endocrinol 2012;167(1):7-15.
3. Douma S, Petidis K, Doumas M et al. Prevalence of primary hyperaldosteronism in resistant hypertension: a retrospective observational study. Lancet 2008;371(9628):1921-6.
4 . Ca l h o u n D A , N i s h i z a ka M K , Za m a n M A e t a l . Hyperaldosteronism among black and white subjects with resistant hypertension. Hypertension 2002;40(6):892-6.
5. Rossi GP, Bolognesi M, Rizzoni D et al. Vascular remodeling and duration of hypertension predict outcome of adrena-lectomy in primary aldosteronism patients. Hypertension 2008;51(5):1366-71.
6. Letavernier E, Peyrard S, Amar L et al. Blood pressure out-come of adrenalectomy in patients with primary hyperaldo-steronism with or without unilateral adenoma. J Hypertens 2008;26(9):1816-23.
7. Milliez P, Girerd X, Plouin PF et al. Evidence for an increased rate of cardiovascular events in patients with primary aldo-steronism. J Am Coll Cardiol 2005;45(8):1243-8.
8. Rossi GP, Sechi LA, Giacchetti G et al. Primary aldostero-nism: cardiovascular, renal and metabolic implications. Trends Endocrinol Metab 2008;19(3):88-90.
9. Savard S, Amar L, Plouin PF et al. Cardiovascular compli-cations associated with primary aldosteronism: a controlled cross-sectional study. Hypertension 2013. [Epub ahead of print]
10. Rossi GP, Bernini G, Caliumi C et al. A prospective study of the prevalence of primary aldosteronism in 1,125 hypertensive patients. J Am Coll Cardiol 2006;48(11):2293-300.
11. Kupers EM, Amar L, Raynaud A et al. A clinical prediction score to diagnose unilateral primary aldosteronism. J Clin Endocrinol Metab 2012;97(10):3530-7.
12. Connell JM, Davies E. The new biology of aldosterone. J Endocrinol 2005;186(1):1-20.
13. Zennaro MC, Boulkroun S. Correspondances en Métabolismes Hormones Diabètes et Nutrition vol. XVII-2013; 17(3):39.
14. Quinn SJ, Williams GH. Regulation of aldosterone secretion. Annu Rev Physiol 1988;50:409-26.
15. Sutherland DJ, Ruse JL, Laidlaw JC. Hypertension, increased aldosterone secretion and low plasma renin activity relieved by dexamethasone. Can Med Assoc J 1966;95(22):1109-19.
16. New M. Hypertension of childhood with suppressed renin. Endocr Rev 1980;1(4):421-30.
17. Lifton RP, Dluhy RG, Powers M et al. A chimaeric 11β-hydroxylase/aldosterone synthase gene causes gluco-corticoid-remediable aldosteronism and human hypertension. Nature 1992;355(6357):262-5.
18. Pascoe L, Jeunemaitre X , Lebrethon MC et al. Glucocorticoid-suppressible hyperaldosteronism and adre-nal tumors occurring in a single French pedigree. J Clin Invest 1995;96(5):2236-46.
19. Stowasser M, Bachmann AW, Huggard PR et al. Treatment of familial hyperaldosteronism type I: only partial suppression of adrenocorticotropin required to correct hypertension. J Clin Endocrinol Metab 2000;85(9):3313-8.
20. Funder JW, Carey RM, Fardella C et al. Case detection, diagnosis, and treatment of patients with primary aldoste-ronism: an endocrine society clinical practice guideline. J Clin Endocrinol Metab 2008;93(9):3266-81.
21. Mulatero P, Tizzani D, Viola A et al. Prevalence and cha-racteristics of familial hyperaldosteronism: the PATOGEN study (Primary Aldosteronism in TOrino-GENetic forms). Hypertension 2011;58(5):797-803.
22. Aglony M, Martinez-Aguayo A, Carvajal CA et al. Frequency of familial hyperaldosteronism type 1 in a hypertensive pediatric population: clinical and biochemical presentation. Hypertension 2011;57(6):1117-21.
23. Stowasser M, Gordon RD, Tunny TJ et al. Familial hyperal-dosteronism type II: fi ve families with a new variety of primary aldosteronism. Clin Exp Pharmacol Physiol 1992;19(5):319-322.
24. Médeau V, Assié G, Zennaro MC, Clauser E, Plouin PF, Jeunemaitre X. Aspect familial de l’hyperaldostéronisme primaire : analyse de familles compatibles avec un hyper-aldostéronisme primaire de type 2. Ann Endocrinol (Paris) 2005;66(3):240-6.
25. Stowasser M, Gordon RD. Familial hyperaldosteronism. J Steroid Biochem Mol Biol 2001;78(3):215-29.
26. Laff erty AR, Torpy DJ, Stowasser M et al. A novel genetic locus for low renin hypertension: familial hyperaldostero-nism type II maps to chromosome 7 (7p22). J Med Genet 2000;37(11):831-5.
27. Stowasser M, Gordon RD. Primary aldosteronism: from genesis to genetics. Trends Endocrinol Metab 2003;14(7):310-7.
28. Mulatero P, Tauber P, Zennaro MC et al. KCNJ5 mutations in European families with nonglucocorticoid remediable familial hyperaldosteronism. Hypertension 2012;59(2):235-40.
29. Geller DS, Zhang J, Wisgerhof MV et al. A novel form of human mendelian hypertension featuring nonglucocor-ticoid-remediable aldosteronism. J Clin Endocrinol Metab 2008;93(8):3117-23.
30. Choi M, Scholl UI, Yue P et al. K+ channel mutations in adrenal aldosterone-producing adenomas and hereditary hypertension. Science 2011;331(6018):768-72.
31. Oki K, Plonczynski MW, Luis Lam M et al. Potassium channel mutant KCNJ5 T158A expression in HAC-15 cells increases aldosterone synthesis. Endocrinology 2012;153(4):1774-82.
32. Scholl UI, Nelson-Williams C, Yue P et al. Hypertension with or without adrenal hyperplasia due to diff erent inherited mutations in the potassium channel KCNJ5. Proc Natl Acad Sci U S A 2012;109(7):2533-8.
33. Charmandari E, Sertedaki A, Kino T et al. A novel point mutation in the KCNJ5 gene causing primary hyperaldoste-ronism and early-onset autosomal dominant hypertension. J Clin Endocrinol Metab 2012;97(8):E1532-9.
34. Heitzmann D, Derand R, Jungbauer S et al. Invalidation of TASK1 potassium channels disrupts adrenal gland zonation and mineralocorticoid homeostasis. Embo J 2008;27(1):179-87.
35. Davies LA, Hu C, Guagliardo NA et al. TASK channel deletion in mice causes primary hyperaldosteronism. Proc Natl Acad Sci USA 2008;105(6):2203-8.
36. Penton D, Bandulik S, Schweda F et al. Task3 potassium channel gene invalidation causes low renin and salt-sensitive arterial hypertension. Endocrinology 2012;153(10):4740-8.
37. Boulkroun S, Beuschlein F, Rossi GP et al. Prevalence, clini-cal, and molecular correlates of KCNJ5 mutations in primary aldosteronism. Hypertension 2012;59(3):592-8.
38. Azizan EA, Murthy M, Stowasser M et al. Somatic muta-tions aff ecting the selectivity fi lter of KCNJ5 are frequent in 2 large unselected collections of adrenal aldosteronomas. Hypertension 2012;59(3):587-91.
39. Akerstrom T, Crona J, Delgado Verdugo A et al. Comprehensive re-sequencing of adrenal aldosterone pro-ducing lesions reveal three somatic mutations near the KCNJ5 potassium channel selectivity fi lter. PLoS One 2012;7(7):e41926.
40. Monticone S, Hattangady NG, Nishimoto K et al. Eff ect of KCNJ5 mutations on gene expression in aldosterone-producing adenomas and adrenocortical cells. J Clin Endocrinol Metab 2012;97(8):E1567-72.
41. Taguchi R, Yamada M, Nakajima Y et al. Expression and mutations of KCNJ5 mRNA in Japanese patients with aldosterone-producing adenomas. J Clin Endocrinol Metab 2012;97(4):1311-9.
42. Seccia TM, Mantero F, Letizia C et al. Somatic mutations in the KCNJ5 gene raise the lateralization index: implications for the diagnosis of primary aldosteronism by adrenal vein sampling. J Clin Endocrinol Metab 2012;97(12):E2307-13.
43. Beuschlein F, Boulkroun S, Osswald A et al. Somatic mutations in ATP1A1 and ATP2B3 lead to aldosterone- producing adenomas and secondary hypertension. Nat Genet 2013;45(4):440-4.
R é f é r e n c e s
Information de dernière minuteAprès la rédaction de cet article, des mutations somatiques du gène CACNA1D, codant pour Cav1.3, la sous-unité α1 du canal calcique voltage-dépendant de type L, se sont avérées être impliquées dans environ 10 % des adénomes de Conn. Des muta-tions constitutionnelles similaires ont été identifi ées dans une nouvelle forme d’hyperal dostéronisme associée à un syndrome neurologique complexe avec crises épileptiques chez l’enfant (1, 2). Ces mutations modifi ent les propriétés des canaux calciques L, favorisant l’infl ux de calcium dans la cellule à des voltages plus bas et une activation inappropriée de la voie de signalisation calcique.
1. Scholl UI, Goh G, Stölting G et al. Somatic and germline CACNA1D calcium channel mutations in aldosterone-producing adenomas and primary aldosteronism. Nat Genet 2013;doi:10.1038/ng.2695. [Epub ahead of print].
2. Azizan EA, Poulsen H, Tuluc P et al. Somatic mutations in ATP1A1 and CACNA1D underlie a common subtype of adrenal hypertension. Nat Genet 2013;doi: 10.1038/ng.2716. [Epub ahead of print].
Maria-Christina Zennaro déclare ne pas avoir de liens d’intérêts.













![PROLFIL CLINIQUE ET ETIOLOGIQUE DE … · Mohammed VI par les adénomes à prolactine. BIBLIOGRAPHIE: [1].Brue T, Delemer B. Diagnostic et prise en charge des hyperprolactinémies-Consensus](https://img.pdfslide.fr/doc/110x75/608450e2ff7d526a8d044ee9/prolfil-clinique-et-etiologique-de-mohammed-vi-par-les-adnomes-prolactine.jpg)