Embed Size (px)
Citation preview

REVUE
Cytokines pro-inflammatoires, prostaglandineset chondrocyte : mécanismes d’activationintracellulaire
Francis BerenbaumService de rhumatologie et UPRES-A CNRS 7079, université Pierre et Marie Curie, Paris VI, hôpital Saint-Antoine, 182,rue du Faubourg Saint-Antoine, 75012 Paris, France
chondrocyte / cytokines / MPA kinases / NF-jB / prostaglandines / signalisation intracellulaire
chondrocyte / cytokines / intracellular signal / MPA kinases / NF-jB / prostagladins
Le chondrocyte articulaire représente le seul type cellu-laire présent dans la matrice cartilagineuse, tissu nonvascularisé et non innervé. Il s’agit d’une cellule spécia-lisée, hautement différenciée, issue de la lignée mésen-chymateuse et susceptible de se dédifférencier en unecellule fibroblastique. Son rôle physiologique consistepour l’essentiel à maintenir un équilibre entre synthèseet dégradation de la matrice, grâce à la synthèse deprotéoglycanes et de collagène de type II d’une part, etde diverses protéases d’autre part.
Malgré la faible activité métabolique constatée enpériode normale, le chondrocyte est capable de synthè-ses nombreuses et variées si un stress survient, qu’il soitde nature mécanique, inflammatoire, infectieux oumétabolique. Parmi ces synthèses, on retrouve des cyto-kines et des dérivés de l’acide arachidonique (ou icosa-noïdes) qui vont avoir comme rôle de modulerl’activation chondrocytaire, par voie autocrine, para-crine ou directement par voie intracellulaire. De plus,d’autres cellules présentes dans l’articulation, commepar exemple les synoviocytes et les cellules endothélialesdu tissu synovial, vont également participer à la régula-tion chondrocytaire en synthétisant elles aussi des cyto-kines et des icosanoïdes, dans la cavité articulaire. Grâceà leur diffusion passive à travers la matrice cartilagi-neuse, ces médiateurs vont pouvoir atteindre les chon-drocytes. Enfin, les capacités de rétention de cytokines
par la matrice elle-même peuvent entraîner un effet« réservoir » du cartilage provoquant une nouvellesource de médiateurs par relarguage à proximité deschondrocytes lors de la dégradation matricielle.
Ainsi, le chondrocyte en situation pathologique seretrouve soumis à une « soupe » de médiateurs lipidi-ques et protidiques présents dans son environnementpéricellulaire. Ces médiateurs vont entrer en contactavec des récepteurs spécifiques présents à la surface deschondrocytes. La liaison du ligand à son récepteur vaentraîner l’activation de différentes cascades intracellu-laires de phosphorylation. Ces phosphorylations vontprovoquer la liaison de facteurs transcriptionnels surdes promoteurs de gènes comportant des séquences deliaison spécifiques pour ces facteurs, entraînant soit uneactivation soit une répression transcriptionnelle d’unensemble de gènes dits inductibles. De plus, ces phos-phorylations en cascade peuvent moduler les étapespost-transcriptionnelles et traductionnelles de ces gènesselon des séquences mal connues encore aujourd’hui.
CYTOKINES PRO-INFLAMMATOIRESET CHONDROCYTE
Parmi l’ensemble des cytokines pro-inflammatoiresconnues à ce jour, seules quelques-unes semblent jouerun rôle important dans l’activation chondrocytaire. Ils’agit de l’IL-1 surtout, et du TNF-α qui peuvent, à leur
Rev Rhum [Ed Fr] 2000 ; 67 : 677-80Proinflammatory cytokines, prostaglandins, and the chondrocyte: mechanisms of intracellular activation –Joint Bone Spine 2000 ; 67 (in press)© 2000 Éditions scientifiques et médicales Elsevier SAS. Tous droits réservésS1169833000000259/REV

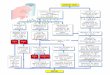
tour, provoquer la synthèse d’IL-6, d’IL-8, de LIF(leukemia inhibitory factor) et d’IL-17. Le chondrocytearticulaire arthrosique surexprime le récepteur à l’IL-1de type I au détriment du type II et surexprime lerécepteur au TNF-α p55TNF-R au détriment dup75TNF-R [1]. La liaison de l’IL-1 sur son récepteurprovoque l’auto-activation du récepteur en recrutantdeux protéines, la protéine accessoire du récepteur del’IL-1 ou IL-1RacP et MyD88, formant ainsi un com-plexe capable d’être phosphorylé par IRAK (IL-1 recep-tor associated kinase), protéine pouvant ensuite interagiravec un autre facteur dénommé TRAF-6 (TNF-receptorassociated protein). La liaison du TNF-α à son récepteurprovoque le recrutement des protéines TRADD (type-Ireceptor-associated death domain protein), FADD,TRAF-2 et RIP (ring finger interacting protein). Lescomplexes multiprotéiques des récepteurs à l’IL-1 et duTNF-α peuvent alors activer TAK-1 [2], une kinasesituée au carrefour des deux grandes voies de signalisa-tion, la voie NF-jB et la voie des MAP kinases [3](figure 1).
NF-jB et chondrocyte
NF-jB est un facteur transcriptionnel existant sous laforme d’un homo- ou d’un hétérodimère d’une famillede cinq protéines dénommées p65, c-Rel, RelB, p50/p105 et p52/p100, le dimère le plus fréquent étantp50-p65 [4]. Il est présent dans le cytosol sous formeinactive, lié à une famille de protéines inhibitricesdénommée I-jB. La liaison de l’IL-1 ou du TNF à sonrécepteur aboutit à l’activation d’une cascade complexequi va provoquer la phosphorylation de I-jB. I-jB peutalors être ubiquitinée1, puis être dégradée dans le pro-téasome. NF-jB est ainsi libéré de I-jB et peut ensuiteêtre transloquée dans le noyau et activer la transcriptionde différents gènes possédant un élément de réponse
1 L’ubiquitine est une petite protéine qui une fois attachée de façoncovalente à sa cible, entraîne la reconnaissance de celle-ci par uneparticule protéolytique 26S appelée le protéasome, particule danslaquelle sont dégradées de nombreuses protéines cytosoliques
Figure 1. Rôle des voies de signalisation NF-jB et MAPK dans l’expression des gènes inductibles. TNF-R : récepteur au TNF-α ; IL-1R :récepteur à l’IL-1 ; FADD : Fas-associated death domain protein ; TRADD : TNF-R-associated death domain protein ; TRAF : TNF-receptorassociated factors ; IRAK : IL-1 receptor associated kinase ; NIK : Nck interacting kinase ; NF-jB : nuclear factor-jB ; IjB : inhibitor of NF-jB ;IKK : I-jB kinase ; AP-1 : activator protein-1 ; C/EBP : CCAAT/enhancer binding proteins ; TAK-1 : TGF-beta-activating kinase ; MAPK :mitogen-activated protein kinase ; MAPKK : MAPK kinase ; MAPKKK : MAPKK kinase ; MMP : matrix metalloproteinase ; COX-2 :cyclooxygénase-2 ; JNK : Jun amino-terminal kinase.
678 F. Berenbaum

pour NF-jB. Parmi ces gènes, on retrouve des cytoki-nes, des molécules d’adhésion, des protéines de la phaseaiguë de l’inflammation, I-jB, p50, c-rel, lacyclooxygénase-2 [5]. Dans les chondrocytes, cette voieest nécessaire pour l’expression de MMP-13 [6]. Ellemodule l’apoptose chondrocytaire [7]. On a remarquéégalement que l’induction de la iNOS par l’IL-17 néces-sitait l’activation de cette voie [8, 9]. Enfin, l’inhibitionde l’expression du facteur transcriptionnel impliquédans la différenciation chondrocytaire SOX-9 par l’IL-1passe par l’activation de la voie NF-jB. [10].
MAPK et chondrocyte
Parallèlement à la voie NF-jB, l’IL-1 et le TNF-α sontcapables d’activer une autre voie de signalisation dénom-mée voie des MAPK (mitogen-activated protein kinase).Les MAPK correspondent à une cascade de trois modu-les de protéine–kinases : les MAPK sont phosphoryléespar les MAPK kinases (MKK) qui sont elles-mêmesphosphorylées par les MKK kinases (MKKK). Ondénombre à ce jour 12 MAPKs, sept MKKs et 14MKKKs [11]. De plus, il existe quatre isoformes de lap38 MAPK dénommées αp38 MAPK, �p38 MAPK,δp38 MAPK et γp38 MAPK avec probablement desspécificités de substrat. À priori, les possibilités de com-binaison sont telles que le système paraît extraordinai-rement complexe. En fait, le type d’effecteur sélectionneune combinaison plutôt qu’une autre. Par exemple,l’IL-1 et le TNF-α activent préférentiellement les modu-les p44/42 MAPK (ou ERK1/2), Jun NH2-terminalkinases (ou JNK) et p38 MAPK. Ces trois modules vontaboutir à l’activation et/ou la synthèse de différentsfacteurs transcriptionnels tels que AP-1 ou C/EBP. Ilest démontré que les voies JNK et p38 MAPK sontspécifiquement activées par l’IL-1 dans les chondrocy-tes [12].
Les voies JNK [13] et p38 MAPK [14] interviennentdans l’activation de la cyclooxygénase-2 chondrocytaireet de la MMP-13 [15] alors que l’expression de MMP-1passe par les voies p38 et probablement ERK [15]. Lavoie p38 MAPK joue également un rôle dans l’expres-sion de la iNOS [16].
Récepteurs aux prostaglandines et chondrocyte
Les prostaglandines peuvent provoquer une activationcellulaire par l’intermédiaire de récepteurs présents à lasurface cellulaire, dénommés EP-1 à EP-4, classifica-tion fondée sur des différences de profil pharmacologi-que et de voies de signalisation [17]. Ils appartiennent à
la famille des récepteurs de type protéine G à septdomaines transmembranaires. L’activation de EP-1 aug-mente la concentration en calcium intracellulaire, l’acti-vation de EP-2/EP-4 stimule l’adénylate cyclase,l’activation de EP-3 inhibe l’adénylate cyclase. Le récep-teur de type EP-1 a été mis en évidence à la surface deschondrocytes épiphysaires [18].
Plus récemment, il a été montré que certains média-teurs lipidiques, et en particulier des dérivés de l’acidearachidonique comme le leucotriène B4, la ∆15-prostaglandine J2, la prostaglandine D2 et l’acide ara-chidonique lui-même, pouvaient se lier directement àune famille de facteurs transcriptionnels, appelés PPAR(peroxisome proliferator activator receptor) [19]. Les fac-teurs de transcription PPAR appartiennent à la super-famille des récepteurs nucléaires et existent sous troisisoformes : α,�/δ et γ. Le changement de conforma-tion de ce récepteur provoqué par la liaison du média-teur lipidique augmente l’affinité de ce récepteur(hétérodimérisé avec les récepteurs à l’acide cis-rétinoïques RXR) pour une séquence spécifique appeléePPRE (PPAR-responsive element) présente sur le promo-teur de certains gènes. Les chondrocytes articulairespossèdent les isoformes PPARα et PPARγ [20]. Leseffets réels en terme d’activation transcriptionnellevarient en fonction de l’isoforme de PPAR, du promo-teur et du type cellulaire.
Ainsi les prostaglandines peuvent agir soit par voieauto- ou paracrine par l’intermédiaire de récepteursmembranaires, soit directement par l’intermédiaire derécepteurs intranucléaires, sans nécessiter une synthèseextracellulaire.
CONCLUSION
Il y a moins de 30 ans, les premières cytokines étaientdécouvertes. Puis des dizaines d’autres ont été mises enévidence. On a craint alors que la complexité théoriquedu réseau cytokinique ne permettrait jamais de dégagerune cible plus importante qu’une autre en thérapeuti-que anti-inflammatoire. L’efficacité des traitements anti-TNF-α nous montrent qu’au delà de cette complexité,il était possible de hiérarchiser la place de ces cytokinesen fonction de leur action pro- ou anti-inflammatoire.De la même manière, nous connaissons désormais lesprincipales voies de signalisation impliquées dansl’expression des gènes de protéines pro-inflammatoireset prodégradatives. Une hiérarchisation de ces voies esten cours de réalisation, visant à rechercher des voiescommunes à plusieurs cytokines impliquées dans
Cytokines pro-inflammatoires, prostaglandines et chondrocyte 679

l’induction de l’expression de ces gènes. Actuellement,les voies NF-jB et p38 MAPK semblent être les ciblesles plus intéressantes. La mise en évidence de ces voiesde signalisation comporte des conséquences pratiquestrès prometteuses : elle a permis en effet la réalisation denouveaux médicaments actuellement en expérimenta-tion. Ils ont comme avantage à priori d’être plus effica-ces car une seule molécule devient capable de bloquerl’action intracellulaire de plusieurs cytokines d’un coup,empêchant ainsi d’éventuelles synergies d’action commepar exemple la synergie IL-1/TNF-α. Des études clini-ques actuellement en cours devraient confirmer pro-chainement l’intérêt de ces données théoriques.
REFERENCES
1 Martel-Pelletier J, Alaaeddine N, Pelletier JP. Cytokines andtheir role in the pathophysiology of osteoarthritis. Front Biosci1999 ; 4 : D694-703.
2 Ninomiya-Tsuji J, Kishimoto K, Hiyama A, Inoue J, Cao Z,Matsumoto K. The kinase TAK1 can activate the NIK-I kap-paB as well as the MAP kinase cascade in the IL-1 signallingpathway. Nature 1999 ; 398 : 252-6.
3 Firestein GS, Manning AM. Signal transduction and transcrip-tion factors in rheumatic disease. Arthritis Rheum 1999 ; 42 :609-21.
4 Baldwin AS. The NF-jB and IjB proteins: new discoveries andinsights. Annu Rev Immunol 1996 ; 14 : 649-81.
5 Barnes PJ, Karin M. Nuclear factor-kappaB: a pivotal transcrip-tion factor in chronic inflammatory diseases. N Engl J Med1997 ; 336 : 1066-71.
6 Mengshol JA, Vincenti MP, Coon CI, Barchowsky A, Brincke-rhoff CE. Interleukin-1 induction of collagenase 3 (matrixmetalloproteinase 13) gene expression in chondrocytes requiresp38, c-Jun N-terminal kinase, and nuclear factor kappaB: dif-ferential regulation of collagenase 1 and collagenase 3. ArthritisRheum 2000 ; 43 : 801-11.
7 Kuhn K, Hashimoto S, Lotz M. IL-1 beta protects humanchondrocytes from CD95-induced apoptosis. J Immunol2000 ; 164 : 2233-9.
8 Martel-Pelletier J, Mineau F, Jovanovic D, Di Battista JA,Pelletier JP. Mitogen-activated protein kinase and nuclear fac-tor kappaB together regulate interleukin-17-induced nitric oxideproduction in human osteoarthritic chondrocytes: possible roleof transactivating factor mitogen-activated protein kinase-activated proten kinase (MAPKAPK). Arthritis Rheum 1999 ;42 : 2399-409.
9 Shalom-Barak T, Quach J, Lotz M. Interleukin-17-inducedgene expression in articular chondrocytes is associated withactivation of mitogen-activated protein kinases and NF-kappaB.J Biol Chem 1998 ; 273 : 27467-73.
10 Murakami S, Lefebvre V, Crombrugghe B de. Potent inhibitionof the master chondrogenic factor Sox9 gene by interleukin-1and tumor necrosis factor-alpha. J Biol Chem 2000 ; 275 :3687-92.
11 Garrington TP, Johnson GL. Organization and regulation ofmitogen-activated protein kinase signaling pathways. Curr OpinCell Biol 1999 ; 11 : 211-8.
12 Yu Geng, Valbracht J, Lotz M. Selective activation of themitogen-activated protein kinase subgroups c-Jun NH2 termi-nal kinase and p38 by IL-1 and TNF in human articularchondrocytes. J Clin Invest 1996 ; 98 : 2425-30.
13 Miller C, Zhang M, He Y, Zhao J, Pelletier JP, Martel-Pelle-tier J, et al. Transcriptional induction of cyclooxygenase-2 geneby okadaic acid inhibition of phosphatase activity in humanchondrocytes: co-stimulation of AP-1 and CRE nuclear bin-ding proteins. J Cell Biochem 1998 ; 15 ; 69 : 392-413.
14 Berenbaum F, Thomas B, Crofford LJ, Biou H, Humbert L,Salvat C, et al. Regulation of the cyclooxygenase-2 promoter inarticular chondrocyte [abstract]. Arthritis Rheum 1999 ; 42.
15 Mengshol JA, Vincenti MP, Coon CI, Barchowsky A, Brincke-rhoff CE. Interleukin-1 induction of collagenase 3 (matrixmetalloproteinase 13) gene expression in chondrocytes requiresp38, c-Jun N-terminal kinase, and nuclear factor kappaB: dif-ferential regulation of collagenase 1 and collagenase 3. ArthritisRheum 2000 ; 43 : 801-11.
16 Badger AM, Cook MN, Lark MW, Newman-Tarr TM,Swift BA, Nelson AH, et al. SB203580 inhibits p38 mitogen-activated protein kinase, nitric oxide production, and induciblenitric oxide synthase in bovine cartilage-derived chondrocytes. JImmunol 1998 ; 161 : 467-73.
17 Coleman RA, Smith WL, Narumiya S. International union ofpharmacology classification of prostanoid receptors: properties,distribution, and structure of the receptors and their subtypes.Pharmacol Rev 1994 ; 46 : 205-29.
18 Del Toro F Jr, Sylvia VL, Schubkegel SR, Campos R,Dean DD, Boyan BD, et al. Characterization of prostaglandinE(2) receptors and their role in 24,25-(OH)(2)D(3)-mediatedeffects on resting zone chondrocytes. J Cell Physiol 2000 ; 182 :196-208.
19 Kersten S, Desvergne B, Wahli W. Roles of PPARs in healthand disease. Nature 2000 ; 405 : 421-4.
20 Bordji K, Grillasca JP, Gouze JN, Magdalou J, Schohn H,Keller JM, et al. Evidence for the presence of peroxisomeproliferator-activated receptor (PPAR) alpha and gamma andretinoid Z receptor in cartilage. PPARgamma activation modu-lates the effects of interleukin-1beta on rat chondrocytes. J BiolChem 2000 ; 275 : 12243-50.
680 F. Berenbaum