Embed Size (px)
Citation preview

European J. Biochem. 7 (1969) 626-530
DAficience en cytochrome oxydase chez les souches de Saccharomyces cerevisiae exigeant la thrkonine pour leur croissance
1. Relations g6n6tiques
Y. SURDIN et H. DE ROBICHON-SZULMAJSTER
Laboratoire d‘Enzymologie du Centre National de la Recherche Scientifique, Gif-sur-Yvette
T. M. LACHOWICZ et P. P. SLONIMSKI
Centre de G6n8tique MolBcnlaire du Centre National de la Recherche Scientifique, Gif-sur-Yvette
( R e y le 19 aoOt 1968)
During the study of the biosynthesis of threonine and methionine in Saccharomyces cerevisiae it appeared that the strains carrying the unliked mutated alleles thr, or thr3 were devoid of cytochrome oxidase activity. Genetic studies have shown that: (a) these strains are not petites; (b) the cytochrome oxidase deficiency segregates with the auxotrophic character for threonine and methionine; (c) reversion to prototrophy simultaneously leads to the recovery of cyto- chrome oxidase synthesis; (d) the presence of a suppressor for one heteroallele of the th2 locus leads also to t,he recovery of cytochrome oxidase synthesis.
Additional studies have shown that partial cytochrome oxidase deficiency is found in mutants blocked a t other steps in the threonine biosynthetic pathway. The multiplicity of unlinked genes giving such an effect has led to the hypothesis of a metabolic relationship between the two phenotypes.
Chez Saccharornyces cerevisiae un grand nombre de mutants B dkficience respiratoire sont connus. Cer- tains doivent leur deficience A la perte d’un facteur cytoplasmique [i], d’autres A des mutations affectant diffkrents genes chromosomiques [2,3].
Au cours d’etudes biochimiques concernant la chaine de biosynthkse de la methionine et de la threonine (Fig. 1) il est apparu que des souches por- tant les alleles mutes thrz ou thr, presentent Bgale- ment une d6ficience en cytochrome oxydase.
Le present article a pour but de clarifier la nature des relations existant entre ces diffkrents caractkres.
MATBRIEL E T MgTHODES Souches
Au cours de ce travail nous avonsutilisk differentes souches haploides de S. cerevisiae dont les noms et genotypes seront donnes au fur e t B mesure.
Abre‘viations non-usuelles. Semi-ald6hyde aspartique, ASA.
Enzymes. Cytochrome oxydase ou cytochrome c : 0, oxidoreductase (EC 1.9.3.1); succino cytochrome c r8duc- tase; aspartokinase ou ATP : L-aspartate 4-phosphotrans- ferase (EC 2.7.2.4) ; semi-ald8hyde aspartique d6shydro- ghnase ou L-aspartate-8-semi aldehyde : NADP oxidoreduc- tase (phosphorylante) (EC 1.2.1.11); homoshrine d6shydro- gknase ou L-homos8rine : NAD oxidor6dnctase (EC 1.1.1.3).
Milieux de culture Milieu complexe YPGA. Ce milieu contient pour
1 1 : extrait de l e m e Difco, 5 g ; Bacto-peptone Dico, 5 g; glucose, 30 g ; adenine, 100 mg.
Milieu synthdtique. Milieu minimum Go [4] dans lequel la source carbonke est le glucose.
Milieu synthdtique compl6mentd. On aj oute au milieu minimum Go selon les besoins : DL-thrhonine, 100 mg/l; DL-mhthionine, 20 mg/l; L-histidine, 100 mg/l; L-tryptophane, 20 mg/l; adenine, 20 mg/l; uracile, 20 mg/l.
Tous ces milieux peuvent &re solidifies par l’ad- dition de 30 g/l de bacto-agar.
Milieux de prdsporulation et de sporubtion. Ce sont ceux dbcrits par McClary et coll. [5].
Analyse gdndtique Les croisements, l’isolement des diploides et la
dissection des asques ont B t B effectues selon les tech- niques dkcrites par Mortimer e t Johnson [6] . Les phknotypes des spores ont Bt6 determines sur des milieux sklectifs. La nomenclature des h8t6roall8les aux loci thrz et thr, est celle de Clavillier [7].
Prdpration des extraits acellulaires Pour le dosage des enzymes respiratoires les
cellules, cultivees pendant 72 heures en milieu YPGA

Vol.7, No.4, 1960 Y . SURDIN, H. DE ROBICHON-SZULMAJSTER, T. M. LACROWICZ et P. P. SLONIMSKI 527
thr, thr, thr,
semi-ald6hyde I aspartyl acide . P
I P
I acide - 7
glutamique aspartique phosphate aspartique
It me‘thionine - c-- 0-ac6tyl =====T homoserine
t 11 - thr1
I
homoserine
mea homose‘rine
phosphate - thrq
threonine Fig. 1. Voie de biosyntMse de la thre’onine et de la me’thionine chez Saccharomyces cerevisiae
liquide en ahrobiose sont traitees comme le decrivent Sherman et Slonimski [3]. Pour le dosage des enzymes de la biosynthkse de la threonine et de la methionine, les cellules, cultivees sur milieu synthetique sont broyees comme le decrivent de Robichon-Szulmajster et Corrivaux [S].
Techniques biochimiques Observation spectroscopique des cytochromes sur
des cellules entikres congelees dans l’azote liquide [3] ; d6termination spectroscopique des cytochromes dans le spectrophotomdtre de Cary 15 sur des cellules entikres, non congelees ou congelees dans l’azote liquide [9].
Dosages enzymtiques La cytochrome c oxydase et la succino cyto-
chrome c reductase sont dosees selon Sherman et Slonimski [3] ; l’aspartokinase est dosee selon la methode de Stadtman et coll. [lo]; la L-aspartate- ,?I-semi-aldehyde deshydrogenase est dosee selon la methode dkcrite par Surdin [I I] ; l’homoskrine d6shy- drogenase est dosee dans les conditions decrites par Karassevith et de Robichon-Szulmajster [12].
Les proteines sont dosees soit par la methode de Lowry et coll. [13] soit par la methode du biuret [14].
RI~SULTATS L’Btude de l’intensit6 respiratoire de differents
mutants auxotrophes a montre qu’apr8s culture sur YPGA ou milieu synthetique les souches portant l’allkle mute thr, ont une valeur de &02 3 B 7 fois plus basse que celle des souches portant l’allkle THR,. Cette diminution de la vitesse de respiration concerne plusieurs substrats tels que l’dthanol, le pyruvate et le glucose. D’autre part, les diploides thr2/thr2 sont incapables de sporuler [7]. On sait que, chez la levure, la sporulation requiert un m6tabolisme aerobie intense. On pouvait penser que ces anomalies dans
le metabolisme respiratoire Btaient soit likes B la presence de la mutation thr2 ) soit fortuites.
l h d e des mutants thr, La cytochrome oxydase et la succino-cytochrome
c reductase ont Bt6 dosees chez quelques souches portant l’alldle thr, et chez quelques sBgr6gants issus d’un diploide THR2/thr2 ; parallklement, le spectre des cytochromes de ces souches a Bt6 6tudi6. Les resultats (Tableau 1) indiquent que le caractkre d’auxotrophie pour la threonine et la methionine (da B la presence de l’alldle mute thr2) entraine, dans tous les cas, une absence presque totale du spectre des cytochromes (a + a3) ainsi que de l’activit6 de la cytochrome c
Tableau I . Se‘gre‘gation de l’activitk de la cytochrome oxydase et de la succinocutochrome c re‘ductase chez un divloEle hkte’ro-
zygote thrJTHR, Les souches 18-27 et 22-13 proviennent de la collection de L. Clavillier. Gbnotypes: 18-27: a, ad3.&, tr,, me,; 22-13: a, url, thr2-? Les souches sont cultivbes en milieu YPGA e t prblevbes en phase stationnaire de la croissance aerobie. Les enzymes sont doses comme indiqub dans Materiel et Mbthodes. Les cytochromes (a + a3) ont 4th determines par observation spectroscopique sur des cellules intactes, con- gelbes dans I’azote liquide. Les activites de cytochrome oxydase ou de succino-cytochrome c reductase representent les vitesses d’oxydation ou de reduction du cytochrome c de cheval B 28’. Elles sont exprimbes en variation d‘absor- bance b 550 mp par minute e t par milligramme de protbines
Croissance sur Cyto- n-.A- Succino milieu sans chromr
thrbonineetsans ,,, , Souches
Parent 18-27 + ++ 0,790 0,042 Parent 22-13 - traces 0,004 0,068 Diplofde
18-27 X 22-13 + ++ 0,335 0,180 Tetrade 5:
spore A - traces 0,009 0,082 spore B + ++ 0,850 0,125 spore C - traces 0,007 0,095 spore D + ++ 0,380 0,075

528 Cytochrome oxydase et exigence en threonine chez S. cerevisiae. 1 European J. Biochem.
oxydase. Au contraire, l’activit6 de la succino-cyto- chrome c reductase ne reflete pas la presence ou l’absence de l’allble thr,. En plus de l’analyse de la tetrade dont les r6sultats sont port& dans le Tableau 1, 9 autres tetrades ont Bt6 Btudiees spectroscopique- ment. Dans tous les cas, les deux phenotypes, dkficience en cytochromes (a + a,) et exigence en threonine et methionine ont s6grt5gt5 ensemble. Notons que les souches thr, possedent le cytochrome b ce qui montre qu’il ne s’agit pas de ((petites)) cytoplasmiques. Ceci a BtB v6rifit5 d’autre part par Btalements sur des milieux contenant du glyc6rol ou du lactate, sources
Tableau 2. Relations entre la teneur en cytochromes ( a + ag) et la prksence des diffkrents allbles mutks a u locus T H R ,
La souche 4094-B provient de la collection de F. Sherman et les souches 19-11-, 21-7, 22-11, 23-3 proviennent de cello de L. Clavillier. L’ASA deshydrogenase est dosee b 28” selon la methode decrite dans Materiel et MBthodes. L’activite est exprimbe en nmoles de 8-aspartyl-phosphate formees par minute e t par milligramme de protbines. Lo spectre des cytochromes a BtB enregistre au spectrophotomMre Cary 15 sur les cellules intactes. La teneur en cytochromes (a + a3) est estimee en unites arbitraires (voir legende dc la Fig.2)
Croissanee sur Teneur en Activit6 ASA Souches Ghotypes sans cyto- mbthionine chromes dkshydro-
ni threonine (a + a.) g6nasique
4094-B a, adz, ur,, THR, + 35 77 Mutants
0 0,5 19-11 GC, ur,, thr,., - 0 Q,2 21-7 a, thr,., - 0 092 22-11 GC, thr,., - 0 022 23-3 a, thr,, -
19-11 R, a, ur, + 6 399 21-7 R, a + 21 4,2
Souches reverses
22-11 R, cc + 10 2,4 23-3 R, a + 6 4,o
carbon6es utilisables seulement par la voie respira- toire. Au moins 80 de la population de chaque clone thr, est capable d’utiliser ces deux substrats. D’autre part, l’analyse genetique de ces tetrades a montre que la segregation du gene thr, et de la deficience en cytochromes (a + a3) est independante des autres genes presents dans ce croisement (signe, ad,, trl ) ur,, me1).
On sait que le locus thr2 d6teImine la synthhse de 1’ASA deshydrogknase [15] ; comme le montre le Tableau 2, les 4 het6roalleles connus S ce locus presentent la double deficience en cytochromes (a + a3) et en ASA d6shydrog6nase. De plus, des reversions spontanees, selectionn6es B partir des quatre h6ttkoall8les mutes au locus thrz, ont conduit B la restauration d’une activite partielle pour 1’ASA deshydrogknase et d’un spectre normal pour les cytochromes (a + a,). I1 n’a pas 6t6 vkrifi6 si les reversions citees dans le Tableau 2 sont dues S des
mutations reverses au locus m6me ou B des mutations de suppresseurs. Par contre, l’analyse g6netique du croisement des souches 19-11 (porteuse de l’alkle thr,,) et 21-7 (porteuse de I’allhle thr2.2) a r6vel6 une situation genetique intkressante qui renforce la correlation entre la deficience en ASA d6shydrogenase et la faible activit6 de la cytochrome oxydase. La diplo’ide issu de ce croisement est capable de sporuler et n’exige plus la threonine et la methionine pour sa croissance. Cette observation pouvait &re interprktde comme &ant diie B une compl6mentation fonction- nelle entre les deux h6t6roall&les [7]. Cependant,
Tableau 3. Effet de la prksence d’un suppresseur spkcifique sur la synthese des cytochromes (a + ag) et de 1’ASA de‘shydro-
gknase Pour la determination des enzymes voir la Kgende du Ta- bleau 2. La nomenclature des suppresseurs est celle du Microbial Genetics Bulletin Oak Ridge National Laboratory,
Oak Ridge 1963
Souches Ghotypes
Parents 19-11 thr,.,, S+ 21-7 thr,.,, S
Diploide CC25 = 19-11 x21-7
TBtrade CC25 23A thr,.,, S+ CC25 23B thr,.,, S CC25 23C thr,.,, S CC25 23D thr,.,, s+
Teneur en Aetivite ASA cytochromes d6shydro-
(a + a,) genasique
14 23,7
0 0,80 7 67 7 35 2 1,6
l’analyse des segr6gants issus de ce croisement montre que cette interpretation ne peut plus 6tre retenue. Tout se passe comme si la souche 21-7 portait, en plus de l’allhle thr2-, un suppresseur dominant capable d’agir specifiquement sur l’allhle thr2, [16]. Lorsque ce suppresseur est pr6sent en m6me temps que l’alI6le sur lequel il agit (thr,,) on observe la restauration de l’activit6 de 1’ASA d6shydrogenase et l’apparition de quantites relativement Blevees de cytochromes (a + a,). Cette restauration n’a pas lieu lorsque le suppresseur est en presence de l’allhle thr2-, (Tableau 3).
&ude d’autres mutants thrLonine-m6thionine ex i g eu n t s
Le parallelisme entre la deficience en ASA d6s- hydrogenase et en cytochrome oxydase pouvait &re cause par la double auxotrophie en threonine et en methionine presentee par ces souches. Les spectres de mutants ayant le m6me phenotype nutritionnel gue les mutants thr, ont 6te examin6s (Fig.2). Les souches portant l’allkle mute thr, sont, comme celles portant l’all8le mute thr2) deficientes en cytochromes (a + a3). Au contraire, les souches portant les alleles
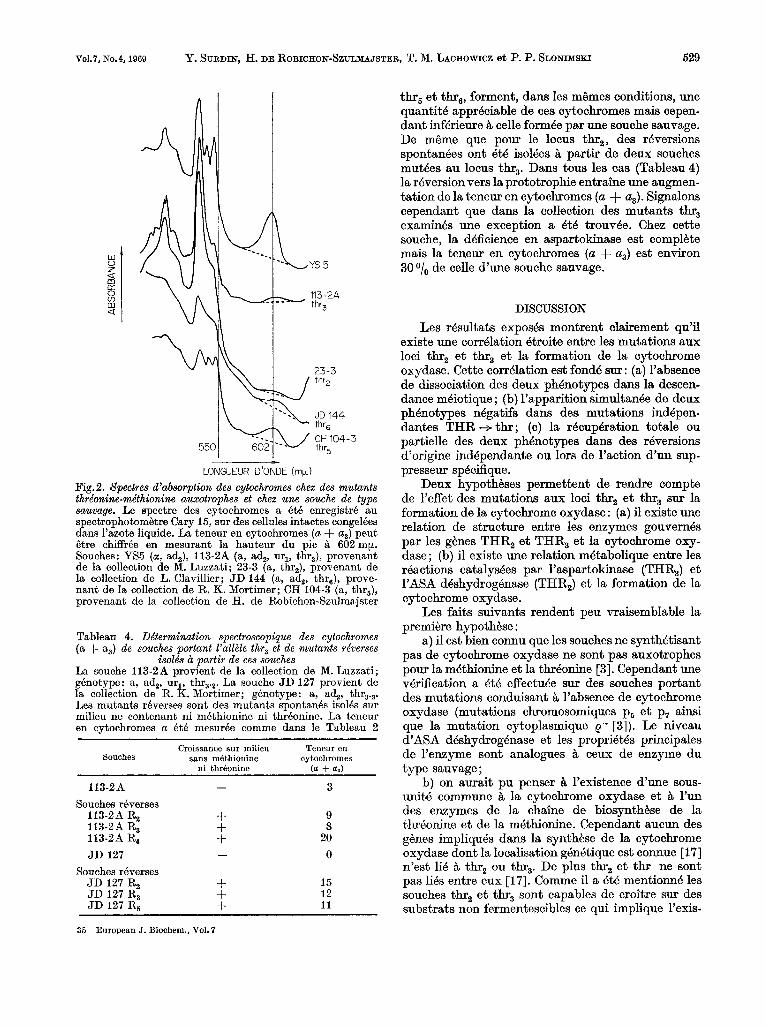
Vo1.7, No.4,1969 Y. SURDIN, H. DE ROBICHON-SZULMAJSTER, T. M. LACHOWICZ et P. P. SLONII~SKI 529
11 3 -2A thr,
J :;i3 \ JD 144
icO4-3
LONGUEUR D’ONDE (mp)
Fig. 2. Spectres d’absorption des cytochromes chez des mutants thrionine-me‘thionine auxotrophes et chez une souche de type sauvage. Le spectre des cytochromes a BtB enregistre au spectrophotom&tre Cary 15, sur des cellules intactes congelees dans l’azote liquide. La tencur en cytochromes ( a + a3) peut &re chiffree en mesurant la hauteur du pic L 602 mp. Souches: YS5 (a, a$), 113-2A (a, a$, ur,, thr3), provenant de la collection de M. Luzzati; 23-3 (a, thr2), provenant de la collection de L. Clavillier; J D 144 (a, ad,, thr,), prove- nant de la collection de R. K. Mortimer; CH 104-3 (a, thr5), provenant de la collection de H. de Robichon-Szulmajster
Tableau 4. De’termination spectroscopique des cytochromes (a + a3) de souches portant l’allkle thr, et de mutants rivverses
isole‘s 2c partir de ces souches La souche 113-2A provient de la collection de M. Luzzati; genotype: a, ad,, url, thr,.,. La souche JD 127 provient de la collection de R. K. Mortimer; gbnotype: a, a$, thr,.,. Les mutants reverses sont des mutants spontanes isolbs sup milieu ne contenant ni methionine ni threonine. La teneur en cytochromes a 6tk mesuree comme dans le Tableau 2
Souches sans methionine cytochromes Croissance 6ur milieu Teneur en
ni thrbonine (a + a,)
3 113-2A -
Souches reverses 113-2A R, + 113-2A R3 + 113-2A R, +
9 8
20 0 J D 127 -
J D 127 R, + 15 Souches reverses
J D 127 R, + 12 J D 127 R, + 11
thr, et the, foment, dans les m6mes conditions, une quantite appreciable de ces cytochromes mais cepen- dant infdrieure it celle formee par une souche sauvage. De m6me que pour le locus thr2, des reversions spontanbes ont B t B isolees it partir de deux souches mutees au locus thr3. Dans tous les cas (Tableau 4) la reversion vers la prototrophie entraine une augmen- tation de la teneur en cytochromes (a + a3). Signalons cependant que dans la collection des mutants thr, examines une exception a Bt6 trouvde. Chez cette souche, la deficience en aspartokinase est complete mais la teneur en cytochromes (a + a,) est environ 30 O l 0 de celle d’une souche sauvage.
DISCUSSION Les rksultats exposes montrent clairement qu’il
existe une correlation Btroite entre les mutations aux loci thr, et thr, et la formation d0 la cytochrome oxydase. Cette corrklation est fond6 sur: (a) l’absence de dissociation des deux phenotypes dans la descen- dance meiotique ; (b) l’apparition simultanhe de deux phenotypes negatifs dans des mutations ind6pen- dantes THR + thr; (c) la recuperation totale ou partielle des deux phenotypes dans des reversions d’origine independante ou lors de l’action d’un sup- presseur sphcifique.
Deux hypotheses permettent de rendre compte de l’effet des mutations aux loci thr2 et thr, sup la formation de la cytochrome oxydase : (a) il existe une relation de structure entre les enzymes gouvern6s par les genes THR, et THR, et la cytochrome oxy- dase; (b) il existe une relation metabolique entre les reactions catalyshes par l’aspartokinase (THR,) et 1’ASA d6shydrogenase (THR,) et la formation de la cytochrome oxydase.
Les faits suivants rendent peu vraisemblable la premiere hypothese :
a) il est bien connu que les souches ne synth6tisant pas de cytochrome oxydase ne sont pas auxotrophes pour la methionine et la threonine [3]. Cependant une verification a ht6 effectuee sur des souches portant des mutations conduisant b l’absence de cytochrome oxydase (mutations chromosomiques ps et p, ainsi que la mutation cytoplasmique e- [3]). Le niveau d’ASA dhshydrogbnase e t les proprihtes principales de l’enzyme sont analogues b ceux de enzyme du type sauvage ;
b) on aurait pu penser L l’existence d’une sous- unit6 commune B la cytochrome oxydase et B l’un des enzymes de la chaine de biosynthese de la threonine et de la mhthionine. Cependant aucun des genes impliques dans la synthbse de la cytochrome oxydase dont la localisation genetique est connue [17] n’est lie B thr, ou thr3. De plus thr2 et thr ne sont pas lies entre eux [17]. Comme il a At6 mentionnh les souches tbr2 et thr3 sont capables de croitre sur des substrats non fermentescibles ce qui implique l’exis-
~~
35 European J. Biochem., Vo1.7

530 Y. SURDIN et al. : Cytochrome oxydase et exigence en thrbonine chez 8. cerevisiae. 1 European J. Biochem.
tence d’un metabolisme respiratoire suffisant donc une synthhse possible de la cytochrome oxydase dans des conditions bien determinees.
L’hypothese la plus probable est done celle d’une consequence metabolique due b l’absence des alleles actifs THR, et THR,. L’intemption de la chaine de biosynthese de la mkthionine et de la threonine pour- rait interferer avec la synthhse de la cytochrome oxydase de deux manibres :
a) l’accumulation de produits intermediaires resultant de la presence de l’une ou l’autre des deux mutations, ou l’absence de formation d’un metabolite due B I’interruption de la chaine de biosynthese, pourrait entrainer l’incapacite de former l’une des composantes de la cytochrome oxydase (apoenzyme ou groupe prosthetique) ;
b) les causes precedentes pourraient agir non pas au niveau de la synthese d‘une composante specifique de la cytochrome oxydase, mais sup le fonctionement du systeme assurant la regulation de cette biosyn- these.
L’Btude plus approfondie des relations mCtaboli- ques entre la cytochrome oxydase et la biosynthkse de la threonine fait l’objet de la publication suivante.
Les auteurs sont heureux de remercier Madame H. Che- rest qui a effectu6 les croisements et I’isolement des s6grB- grants, et Madame D. Henri pour sa pr6cieuse collaboration technique au cours de ce travail. Ce travail a b6n6fici6 de l’aide de la DBlBgation GBnBrale B la Recherche Scientifique et Technique (ComitB de Biologie MolBculaire). T. M. Lacho- wicz Btait boursier des relations culturelles franco-polonaises (1960-1961).
BIBLIOGRAPHIE 1. Slonimski, P. P., et Ephrussi, B., Ann. Inst. Pusteur, 77
(1949) 47.
2. 3.
4.
5.
6.
7.
8.
9.
10.
11. 12.
13.
14. 15.
16.
17.
Sherman, F., Genetics, 48 (1963) 375. Sherman, F., et Slonimski, P. P., Biochim. Biophys.
Acta, 90 (1964) 1. Galzy, P., et Slonimski, P. P., Compt. Rend. 245 (1957)
2556. McClary, D. O., Nulty, W. L., et Miller, G. R., J. Bac-
teriol. 78 (1959) 362. Mortimer, R. K., et Johnson, J. R., J . Bucteriol. 78
(1959) 272. Clavillier, L., These de Doctorat de SpBcialitB, Univer-
sit6 de Paris, 1962. Robichon-Szulmajster, H. de, et Corrivaux, D., Bio-
chim. Biophye. Acta, 73 (1963) 248. Claisse, M., P6r6, G., et Slonimski, P. P., r6sultats
in6dits. Stadtman, E. R., Cohen, G. N., Le Bras, G., et de Ro-
bichon-Szulmajster, H., J. Biol. Chem. 236 (1961) 2033.
Surdin, Y., European J . Biochem. 2 (1967) 341. Karassevitch, Y., et de Robichon-Szulmajster, H., Bio-
Lowry, 0. H., Rosenbrough, N. J., Farr, A. L., et Ran-
Stickland, L. H., J . Gen. MicrobioZ. 5 (1951) 698. Robichon-Szulmajster, H. de, Surdin, Y., et Mortimer,
Cherest, H. et de Robichon-Szulmajster, H., rbsultats
Mortimer, R. K., et Hawthorne, D. C., Genetics, 53
chim. Biophys. Actu, 73 (1963) 414.
dall, R. J., J . Biol. Chem. 193 (1951) 265.
R. K., Genetics, 53 (1966) 609.
non publi6s.
(1966) 165.
Y. Surdin et H. de Robichon-Szulmajster Laboratoire d’Enzymologie du C.N.R.S. F-91 Gif-sur-Yvette, France
P. P. Slonimski Centre de GBnBtique Mol6culaire du C.N.R.S. F-91 Gif-sur-Yvette, France
Adresse actuelle de T. M. Lachowicz: Zaklad Microbiologii uniwersytetu Wroclaw, Pologne




















![EMBO Negative transcription of the Saccharomyces (CTT1) cAMP · of CTI] transcript are observed after derepression for 30 min (Figure 3). Further experiments (data not shown) demonstrated](https://img.pdfslide.fr/doc/110x75/604a582c497b5b7fba59e117/embo-negative-transcription-of-the-saccharomyces-ctt1-camp-of-cti-transcript.jpg)








