Embed Size (px)
Citation preview

Université d’Oran (Es-Senia)
Faculté des Sciences de la Nature et de la Vie
Département de Biotechnologie
Mémoire de MAGISTER
en Biotechnologie
Option : Intérêt des microorganismes en agriculture et en agroalimentaire
Présenté et soutenu publiquement par :
ZEMRI Rahma
Thème :
Pr. Aoues A. Université d’Oran Président
Pr. Kaid- Harche M. USTOMB (ex USTO) Examinateur
Pr. Hadjadj Aoul S. Université d’Oran Examinateur
Pr. Bellahcene M. C. U. Ain Temouchent Examinateur
Pr. Fortas Z. Université d’Oran Rapporteur
Mycorhization en conditions contrôlées du chêne
vert (Quercus ilex L.) par la truffe noire
(Tuber melanosporum Vitt.)
Soutenu le 02 juillet 2013 devant le jury composé de

حين ي ٱلر ـ حو بسن ٱهلل ٱلر
In the name of Allah, the Beneficent, the Merciful
لىي هي وظللا عليڪن ٱلغوام وأسلا عليكن ٱلوي وٱلس لى
كن ـ ت ها رزق ـ وها طيب
أفسهن يظلوىى ) اى كي ـ (٧٥ظلوىا ول
―And we caused the white cloud to overshadow you and sent down on
you the manna and the quails, (saying): Eat of the good things
wherewith we have provided you - they wronged us not, but they did
wrong themselves.‖ (57)


Dédicaces
Je dédie ce modeste travail
A mes parents
A mon mari
A ma sœur et mes deux frères

Remerciements Ce travail a été effectué au Laboratoire de Biologie et de Micro-
organismes et de Biotechnologie (LBMB), à l’Université d’ Oran (es-sénia).
A l’issue de ce travail, je remercie d’abord Dieu tout puissant de m’avoir donné la volonté, le courage et la patience pour achever ce modeste mémoire de Magister.
Je tiens à exprimer ma gratitude et mes sincères remerciements à mon Professeur FORTAS Zohra qui a dirigé ce travail. Je la remercie vivement pour sa rigueur scientifique durant toute ma formation, ses nombreux conseils, ses encouragements et sa disponibilité tout au long de ce travail. Elle mérite un remerciement spécial.
Ma profonde gratitude et mes remerciements vont à M. le Professeur Aoues A. de l’Université d’Oran qui me fait l’honneur de présider le jury malgré ses nombreuses charges pédagogiques et scientifiques.
J'exprime mes sincères remerciements à Mme le Professeur KAID-HARCHE M., directrice du Laboratoire de recherche en Biotechnologie à l’USTOM (ex USTO) qui me fait l’immense honneur de siéger dans mon jury et d’accepter d’examiner mon mémoire de Magister. Je la remercie vivement pour les techniques de cytologie végétale acquises dans son laboratoire.
Mes remerciements s’adressent également à M. le Professeur Bellahcène M., du Centre Universitaire d’Ain Temouchent d’avoir accepté d’examiner ce modeste travail.
J'exprime aussi mes vifs remerciements à M. le Professeur Hadjaj-Aoul S., responsable du Laboratoire d’Ecologie végétale à l’Université d’Oran, qui me fait l’immense honneur de siéger dans mon jury et d’accepter d’examiner mon mémoire de Magister.
Enfin, je n’oublierai pas de remercier tous les membres du Laboratoire LBMB pour leur soutien, leur aide et leur gentillesse en particulier Mme DIB-BELLAHOUEL S. et Mme BOUREGBA M.

introduction 1
Chapitre I : Analyse bibliographique
1- La truffe et la trufficulture 3
1-1-Histoire de la truffe 3
1-2–Répartition géographique des truffières dans le monde 3
1-2-1- Répartition géographique naturelle des truffières 3
1-2-2- Répartition géographique des cultures de truffes 5
1-3– Ecologie de la truffe noire 7
1-3-1- Climat 7
1-3-2- L‘altitude 7
1-3-3- Le sol truffier 7
1-4- Les plantes hôtes de la truffe noire 8
1-5- Taxonomie 8
2 -Quelques notions sur la truffe noire du Périgord (Tuber melanosporum Vitt.) 14
2-1-Comparaison entre la truffe et un champignon épigé 15
2.2- Les composants morphologiques du corps fructifère truffier 15
2-2-1- Le péridium 16
-Une cristallisation de surface avec polysaccharides 17
-Une cristallisation de surface par l‘anion d‘oxalate 17
2-2-2- La trame 17
2-2-2-1- Le tissu de soutien 18
2-2-2-2- Le tissu fertile : 18
2-3- Composition chimique de la truffe noire du Périgord 21
3- Les ectomycorhizes 24
Sommaire

3-1- Généralités
24
3-2- Les mycorhizes et le mycélium truffier 28
3-2-1 Les ectomycorhizes de la truffe 28
3-2-2- L‘organisation du mycélium truffier 28
3-2-2- 1- L‘organisation extra-racinaire du mycélium 28
3-2-2- 2 - L‘organisation du mycélium intra-racinaire 30
3-2-2- 3- La truffe et les stromas 30
3-2-3- Le mycélium truffier et la vie sur terre 31
3- 3- Production et commercialisation des plants mycorhizés par la truffe 34
4 – Caractéristiques botaniques de chêne vert 37
4—1 Taxonomie 37
4--2 - Description morphologie du chêne vert 38
4-3- L‘habitat et répartition 38
4-4- Utilité 40
Chapitre II : Matériel et méthodes
1-Matériel 42
1-1-Matériel fongique 42
1-2- Matériel végétal 42
1-3-Substrat hors-sol 43
2- Méthodes 44
2-1- Etudes morphologiques des ascocarpes 44
2-1-1- Examen macroscopique sous la loupe stéréoscopique 44
2-1-2- Examen macroscopique direct des ascocarpes 45
2-2- Etudes microscopiques de l‘ascocarpe 45
2-2-1- Etudes microscopiques du péridium 45
2-2-2- Etudes microscopiques de la glèbe 45
2-3- Réalisation des synthèses mycorhiziennes 46
2-3-1- Désinfection des glands de chêne vert 46
2-3-2- La stratification et germination des glands de chêne vert 46

2-3-3- Préparation et désinfection du substrat hors-sol 47
2-3-4- Préparation de l‘inoculum 47
2-3-5- Inoculation du substrat 48
2-3-6- Conditions de culture des plants 49
2-4- Les méthodes d‘étude des associations mycorhiziennes 49
2-4-1- Etude des mycorhizes 49
2-4-2– Etude de la croissance des plants 51
2-4-2-1- Estimation des paramètres de croissance des plants 51
2-4-2-2- Estimation du taux de mycorhization 51
2-4-2-3- Interprétation statistique 51
Chapitre III : Résultats et discussion
1- Etude morphologique des ascocarpes de Tuber melanosporum 52
1-1 Examens macroscopiques des ascocarpes 52
1-2 Examen du péridium de l‘ascocarpe sous la loupe stéréoscopique 53
1-3 Examens microscopiques de la glèbe des ascocarpes 57
2- Etude de la mycorhization des plants de chêne vert 61
2-1 Effet de la mycorhization sur la croissance aérienne des plants de chêne vert 61
2-2 Effet de la mycorhization sur la croissance racinaire des plants de chêne vert 66
2-3 Etude morphologique des mycorhizes 69
2-4 Etude microscopique de l‘association mycorhiziènne 72
conclusion 76
Références bibliographiques 73
Annexe 87

Liste des tableaux
Tableau 1 : Un aperçu sur le classement des arbres selon leur pouvoir mycorhizien avec le
Tuber melanosporum
9
Tableau 2 : Diverses appellations de Tuber melanosporum Vitt. 10
Tableau 3 : Différences entre la truffe et le champignon épigé (in CHEVALIER et al., 2008). 15
Tableau 4 : Principales caractéristiques morphologiques de quelques espèces de Tuber . 20
Tableau 5 : Composition chimique de deux types de truffe (HARKI et al., 2006 ; in WANG et
MARCONE, 2011).
21
Tableau 6 : Composition chimique de la truffe noire du Périgord pendant trois derniers stades
de sa maturité (HARKI et al., 2006).
22
Tableau 7 : Composition chimique en pourcentage des principaux composants au stade mature
de Tuber melanosporum (HARKI et al, 2006).
23
Tableau 8 : Familles et genres d‘arbres formant des ectomycorhizes (DRENOU et al., 2006 ;
BA et al., 2011).
26
Tableau 9 : Différence entre les deux phases végétative et reproductive (GRENTE et
DELMAS, 1974 ; GRENTE et CHEVALIER, 1980 ; CHEVALIER et
FROCHOT, 1997 ; DESSOLAS et al., 2008).
32
Tableau
10 :
Taxonomie du chêne vert (in HAICHOUR, 2009). 37
Tableau
11 :
Description morphologique du chêne vert (BELKHADER, 2003 ; NACEF et
DJERNOUMI, 2004 ; KREMER, 2005 ; ZITOUNI, 2010).
39
Tableau
12 :
Paramètres de croissance estimés pour l‘espèce végétale étudiée 51
Tableau
13 :
Comparaison de nos résultats sur les paramètres de l‘ascocarpe de T.
melanosporum avec ceux de la littérature
53
Tableau
14 :
Comparaison de nos résultats sur l‘aspect du péridium de T. melanosporum avec
ceux de la littérature.
54
Tableau
15 :
L‘évaluation statistique des paramètres de croissance des plants de chêne vert. 61
Tableau
16 :
L‘évaluation statistique des paramètres de développement de la partie racinaire des
plants de chêne vert.
67

Fig. 1 : Schéma explicatif des causes de la chute de la production de la truffe et de
la trufficulture (GRENTE et CHEVALIER, 1973 ; GRENTE et
DELMAS, 1974)
5
Fig. 2: Répartition des régions à production truffière naturelle ou cultivée dans le monde
(GRENTE et CHEVALIER, 1973 ; PEREZE et al., 2007 ;PRUETT et al., 2008 ;
LAQBAQBI et al., 2011 ; CORTES et al., 2011 ; SHANNON, 2011
6
Fig. 3 : Classification du genre Tuber selon JEANDROZ et al. (2008) et
BONITO et al. (2010).
11
Fig. 4 : L‘arbre phylogénique du genre Tuber (BONITO et al., 2010)
.
12
Fig. 5 : Une partie de l‘arbre phylogénique décrit par LAESSOE et HANSEN
(2007) montre le taxon des Tuberaceae.
13
Fig. 6: La position des Tuberaceae dans l‘ordre des Pézizales (LAESSOE et
HANSEN, 2007).
13
Fig. 7: Ascocarpes de la truffe noire du Périgord (A) (LAURENT, 2003) et fructification
souterraine de la truffe noire (B) (PARGNEY et MEUNIER, 2004).
14
Fig. 8 : Un ascocarpe de Tuber melanosporum de poids important (in CALLOT, 1999)(A) ; Aspect du péridium de Tuber melanosporum
(BENAC, 2006)(B)
16
Fig. 9 : Aspect de la glèbe (A et B) et du péridium de Tuber melanosporum
(BRUN, 2002).
17
Fig. 10 : Morphologie des ascospores de Tuber melanosporum
19
Fig. 11 : Type mycorhizien.
25
Fig. 12: La forme d‘une ectomycorhize
27
Fig. 13 : Mycorhizes de Tuber melanosporum (ROBIN et al., 2008).
28
Liste des figures

Fig. 14 : Forme de spinules chez des espèces de Tuber (PARGNEY et
MANCHABLON, 2003).
29
Fig. 15 : Coupe longitudinale dans une ectomycorhize de Truffe dont : M est le
manchon dense auteur de la racine ainsi que le réseau d‘ Hartig et
allongement de l‘assise corticale (DEXHEIMER, 1997).
30
Fig. 16: Aspect schématique d‘un stroma (PARGNEY et al., 2008).
31
Fig. 17 : Schéma explicatif des deux phases de la vie de la truffe (NB : les
proportions entre les différents organismes ne sont pas respectées),
(DESSOLAS et al., 2008)
32
Fig. 18 : Schéma simplifié du principe d‘une association symbiotique entre la truffe
et l‘arbre truffier.
33
Fig. 19 : Cycle de vie de Tuber melanosporum (KUES et MARTIN, 2011).
35
Fig. 20 : Schéma simplifié de mycélium truffier entre l‘ancienneté et la modernité
(GRENTE et DELMAS, 1974, CHEVALIER et GRENTE, 1980 ;
CHEVALIER, 1985 ; DESSOLAS et al., 2008).
36
Fig. 20 : Répartition du chêne vert (Quercus ilex) dans l‘hémisphère du nord
(BARBERO et LOISEL, 1980)
41
Fig. 20 : Distribution du chêne vert (Quercus ilex) en Algérie (HAICHOUR, 2009).
41

Planche I : Morphologie et aspect du péridium de l‘ascocarpe de la truffe noire 55
Planche II : Morphologie de l‘asque et de l‘ascospore de la truffe noire 59
Planche III : Croissance des plants de chêne vert inoculés par Tuber melanosporum, après 12 mois de culture hors sol en serre.
63
Planche I V : Effet de l‘inoculation de Tuber melanosporum sur la croissance
aérienne des plants de chêne vert.
65
Planche V : Effet de l‘inoculation de Tuber melanosporum sur le
développement racinaire des plants de chêne vert.
68
Planche VI : Morphologie des racines de plants de chêne vert inoculés par Tuber melanosporum observées sous la loupe binoculaire.
70
Planche VII : Association mycorhizienne entre Chêne vert et Tuber melanosporum, après 12 mois de culture hors sol en serre.
73
Liste des planches 40


Introduction :
Les truffes du genre Tuber (Ascomycètes, Pézizales) sont des champignons
souterrains qui se forment dans les couches superficielles du sol et qui sont connues
pour leur valeur gastronomique (MURAT et al., 2008 ; DESSOLAS et al., 2008 ;
THERVILLE, 2009 ; SOURZAT, 2011). L‘espèce la plus précieuse d‘entre elles est
la truffe noire du Périgord « Tuber melanosporum Vitt. » nommée aussi le diamant
noir de la cuisine par BRILLAT SAVARINE (GRENTE et CHEVALIER, 1973).
Elle a une valeur gastronomique et gustative importante par rapport aux autres
truffes. Elle est utilisée dans la cuisine européenne, sa bonne saveur la rend
indispensable dans les restaurants chics à cinq étoiles, elle est nommée aussi la truffe
noble car elle est présente sur les table des rois pour son intérêt gustatif important
(DESSOLAS et al., 2008).
Sa composition organique et inorganique est intéressante et différente des
autres espèces de Tuber , elle est riche en protéines et en sucres qui lui donnent une
valeur énergétique élevée ; elle contient des sels minéraux, une faible quantité de
lipides et de la mélanine qui est responsable de sa couleur noire (HARKI et al.,
1996 ; DIEZ et al., 2003 ; HARKI et al., 2006).
La truffe est un champignon mycorhizien, ne peut donc accomplir son cycle
qu‘avec une plante hôte (CHEVALIER et FROCHOT, 1997 ; DRENOU, 2006 ; BA
et al., 2011) en particulier des arbres dont les plus importants appartiennent aux
familles des Fagacées et Pinacées (CHEVALIER, 1985 ; CHEVALIER , 1996 ;
DOMINGUEZ et al., 2005 ; 2006 ; GARCIA-MONTERRO et al., 2007 ; 2009). La
mycorhization de ces arbres avec ce champignon est connue pour améliorer leur
vigueur et en particulier accroitre leur résistance à la sécheresse et améliorer
notablement leur taux de survie. Elle permet aussi la fructification de ce champignon
comestible.
Dans notre travail, nous nous sommes intéressés à ce précieux champignon
inconnu en Algérie et plus particulièrement la truffe noire, Tuber melanosporum Vitt.
car il peut s‘associer à diverses essences forestières (chêne, pin..) en établissant des
ectomycorhizes. Il peut donc être introduit dans notre pays par le biais de reboisements

artificiels de chênes et pins mycorhizés en pépinière. C‘est dans cette perspective que
nous avons entrepris notre recherche.
Notre étude porte sur la mycorhization contrôlée d‘une espèce forestière,
Quercus ilex L. (chêne vert) avec Tuber melanosporum Vitt., champignon hypogé
comestible récolté en France.
Cette essence forestière a été choisie pour sa dominante dans les forêts algériennes,
elle peut donc être mycorhizée par ce champignon puis introduite par le biais de
reboisements artificiels dans des terrains où les conditions écologiques sont favorables
au développement des deux partenaires de la symbiose.
L‘objectif de notre travail est de :
- Réaliser des synthèses mycorhiziennes en conditions gnotoxéniques, sur un
substrat hors sol désinfecté. Les plants issus de la germination des glands de chêne
sont cultivés en pots ouverts et inoculés avec des suspensions de spores de Tuber
melanosporum Vitt. selon la technique couramment utilisée à l‘INRA de
Clermont-Ferrand, pour la production des plants de chêne mycorhizés par la
truffe noire du Périgord (Tuber melanosporum Vitt.) ;
- Estimer statistiquement la croissance des plants par la mesure de la hauteur de la
tige, le nombre des feuilles et sa surface, le poids frais et sec des plants, ainsi que la
longueur de la racine principale, la longueur des racines secondaire, le poids frais
de la partie racinaire et le nombre des racines latérales des plants inoculés ou non
avec des suspensions sporales de Tuber melanosporum Vitt.
- Et enfin, mettre en évidence la présence des ectomycorhizes chez cette essence
forestière inoculée en conditions contrôlées en décrivant leur morphologie par
des observations macroscopiques et microscopiques et en estimant le taux
d‘infection mycorhizienne des plants inoculés par le champignon.

1- La truffe et la trufficulture
1-1-Histoire de la truffe :
Les truffes ont une vielle histoire, elles sont connues depuis l‘Antiquité dès 3500
av. J-C, et pour la première fois vers le 4 éme
siècle , le philosophe grec
THEOPHRASTE (372-287 av. J-C) mentionna dans un ouvrage « Histoire des
plantes » que les truffes sont des végétaux sans racines, engendrées par les pluis
automnales, par contre le moraliste PLUTARQUE (46-125 ap. J-C) considéra ces
végétaux comme une production issue de la fusion de l‘eau, de la terre et de la foudre
(PARGNEY, 1994 ).
Elles sont connues chez les Athénéens et les Romains, elles apparaissent sur leur table
somptueuse comme additif alimentaire, puis leur généralisation vers le 12 éme
siècle a
été faite par les romains. Vers le 13 ème
siècle, la truffe noire est localisée en France
d‘abord au Périgord ; elle se présente sur les tables les plus somptueuses jusqu‘au 16 ème
siècle et sa présence est obligatoire dans tous les festins d‘où sa dénomination de truffe
noble. Dès 1794, la truffe été exportée pour la première fois vers la Hollande et
l‘Allemagne pour être utilisée dans les hôtelleries et les restaurants (CHEVALIER et
al., 2008)
BRILLAT-SAVARINE l‘a appelé « Le diamant noir de la cuisine » (CHEVALIER et
GRENTE, 1980)
La renommée internationale de la truffe et son importance économique ont incité
VITTADINI à introduire les truffes dans le monde des champignons vers le 18 ème
siècle et vers le 19 ème
siècle son cycle vital a été étudié par les botanistes (PARGNEY,
1994 ; CHEVALIER et al., 2008)
Le Coran a aussi évoqué la truffe et son origine (in EL BAKARA, 57) que
Dieu a donnés aux israéliens au temps du prophète Moussa et son importance a été
décrite par le prophète Mohammed dans un hadith (in Fath el Bari).
1-2–Répartition géographique des truffières dans le monde :
1-2-1- Répartition géographique naturelle des truffières :

La production naturelle de la truffe noire se trouve dans la région
méditerranéenne en particulier en France, en Italie et en Espagne (BARCHFIELD,
2008).
En France : Tuber melanosporum se récolte dans des régions situées dans le Périgord,
Quercy, Angoumois, Poitou, Provence, Dauphiné, et aussi en Bourgogne. Cette espèce
de Tuber est aussi signalée de façon plus ponctuelle dans d‘autres régions françaises à
microclimat favorable « la Meuse ». De nombreux chercheurs ont remarqué que la
truffe noire s‘associe seulement avec le chêne dans le Périgord (CHEVALIER et
FROCHOT, 1997 ; PARGNEY, 1994 ; HALL et al., 2007 ; WANG et MARCONE,
2011).
La France utilise des méthodes pratiques pour la régénération naturelle ou assistée
afin d‘étendre les zones pédoclimatiques potentiellement propices à la trufficulture
(MOUSSU et LAURIAC, 2008)
En Espagne : les régions rurales productives de truffe noire du Périgord sont :
Catalonie, Aragon, Valencia, Navarre, Castille et Léon (OLIACH et al., 2011). De
nombreuses recherches (agricoles, chimiques…) ont été entrepris pour améliorer la
culture de ce champignon en Espagne (GARCIA-MONTERO et al., 2007 ; WANG et
MARCONE, 2011).
En Italie : elle est connue par la production de carpophores de Tuber melanosporum
dans des sites naturels avec Cistus monspeliensis et même dans des sites où la truffe
noire du Périgord est cultivée avec d‘autres espèces de Cistes tel que Cistus incanus
(FONTANA et GIOVANNETTI, 1979). De nombreux travaux sur la truffe ont été
effectués en 1972 par FONTANA en collaboration avec CHEVALIER de l‘INRA
de Clermont –Ferrand en France (FONTANA et al., 1972 ; GARCIA-MONTARO et
al., 2007, 2009)
En Grèce : des travaux sur la truffe ont été effectués en Xiromero dans des régions de
production naturelle de Tuber melanosporum (CHETON et MORAR, 2011).
En Palestine occupée : la truffe se développe naturellement dans les montagnes sous le
chêne (KUES et MARTIN, 2011). De nombreuses études sont effectuées pour
étendre la culture du Tuber melanosporum en l‘associant avec différentes espèces de
chênes et de noisetiers (PINKAS et al., 2000).

1-2-2- Répartition géographique des cultures de truffes
La truffe a connu une chute de production qui est due à des facteurs multiples
(CHEVALIER et GRENTE, 1973 ; GRENTE et DELMAS, 1974 ; SOURZAT,
2011) qui sont résumés dans la figure 1. Cette chute de production naturelle a incité les
chercheurs européens à développer la trufficulture grâce à des techniques de
productions de plants truffiers internationalement commercialisables (GRENTE et
CHEVALIER, 1973 ; GRENTE et DELMAS, 1974 ; REYNA, 2000 ; BENAC,
2006 ; CHEVALIER et al., 2008 ; SMITH et READ, 2008 ; SAVOIS et
LARGETEAU, 2011).
Fig. 1 : Schéma explicatif des causes de la chute de la production de la truffe et de la
trufficulture (GRENTE et CHEVALIER, 1973 ; GRENTE et DELMAS, 1974)
Ces méthodes de trufficulture ont été appliquées dans d‘autres pays du
monde comme les Etats Unis, la Nouvelle Zélande et l‘Australie (HALL et al., 2007 ;
PRUETT et al., 2008). En Nouvelle Zélande, les premiers ascocarpes de Tuber
melanosporum ont été récoltés en 1993 ce qui a considérablement développée la
trufficulture dans ce pays (CHEVALIER et FROCHOT, 1997 ; HALL et al., 2007 ;
GUERIN-LAGUETTE et al., 2011).

- Au Canada, les premiers essais d‘inoculation des plants par Tuber melanosporum
ont été effectués par les chercheurs de l‘Université de British Columbia, à
Vancouver et les premières truffes ont été récoltées 5 ans après plantation
(SHANNON, 2011).
- En Amérique latine, une plantation de plants mycorhizés par Tuber melanosporum
a été installée en 2003 au Chili (SANTELICES et al., 2004 ; PEREZE et al., 2007) et
en 2010 au sud de l‘Argentine exactement en Pantagonie (CORTES et al., 2011 ).
- Sur le continent africain, les premiers plants de chênes verts mycorhizés par Tuber
melanosporum et produits par Agri-truffe (France) ont été introduits en 2000 dans la
région d‘ Oujda , Maroc (LAQBAQBI et al., 2011).
De nouvelles truffières cultivées se trouvent en Serbie et Croatie (WANG et
MARCONE, 2011). La figure 2 indique les régions à production truffière naturelle ou
cultivée dans le monde.
Fig. 2: Répartition des régions à production truffière naturelle ou cultivée dans le
monde (GRENTE et CHEVALIER, 1973 ; PEREZE et al., 2007 ;PRUETT et al., 2008 ;
LAQBAQBI et al., 2011 ; CORTES et al., 2011 ; SHANNON, 2011)
1-3– Ecologie de la truffe noire :
Pour la trufficulture, l‘arbre et le champignon sont les deux composants de
base, leur développement est sous la dépendance des conditions physico-chimique du
N
E W
S

milieu, du climat et de l‘altitude. L‘écologie des truffières a beaucoup été étudiée par
les chercheurs de l‘INRA de Clermont –Ferrand (DELMAS et GRENTE, 1974 ;
CHEVALIER et GRENTE, 1980; DESSOLAS et al.,s.. ? 2008 ; SOURZAT, 2011)
1-3-1- Climat :
Selon PRADEL en 1914 dit que : « les truffes poussent sous de nombreux climats »
(CHEVALIER et al., 2008) ; Elles sont très dépendantes du climat et de la
pluviométrie, qui sont des facteurs dominants dans l‘activité du mycélium truffier, le
développement de mycorhizes et la formation des ascocarpes (PARGNEY, 1994 ;
CHEVALIER et al., 2008).
Pour la truffe noire du Périgord nécessite un climat méditerranéen, froid pluviaux en
hiver et chaud sec en été, plus précisément les régions méridionales au voisinage des
masses marines (GRENTE et DELMAS, 1974 ; CASERMEIRO et al., 2007)
Une bonne répartition entre la pluviométrie, la température, l‘insolation qui veut dire
une alternance entre ces trois paramètres, pour les pluies pas trop abondantes et au bon
moment (CHEVALIER et al., 2008). D‘après CALLOT et al., (1999), des études
pédologiques montrent que le fonctionnement hydrique des sols calcaires constitue le
facteur essentiel de la production de la truffe.
1-3-2- L‘altitude :
PRADEL (1914) a dit que : « altitudes moyennes, ni trop maigres, ni trop fertiles pour
la culture de la truffe » (CHEVALIER et al., 2008) la truffe noire nécessite une altitude
de 200 à 800 mètres si l‘exposition est ensoleillée, elle se détecte le sud-est et sud-ouest
de France (GRENTE et DELMAS, 1974).
1-3-3- le sol truffier :
Le sol constitue le support et le milieu nutritif de l‘arbre et de champignon, les zones
trufficoles sont des zones calcaires, bien drainées, sans discontinuités structurales et un
horizon profond à porosité biologique (GRENTE et DELMAS, 1974; PARGNEY,
1994).
les sols des truffières s‘étendent sur les vastes dépôts jurassiques tel que le Périgord en
France qui sont des terrains calcaires riches en calcium avec calcaire total de 8%
approximativement, et doit être moins rocheux (- 10%) et peu profond de 15 à 40 cm
pour faciliter le travail du sol, et un sous-sol friable permet au système radiculaire

pivotant de l‘arbre de pénétrer ; la structure de sol doit être bien équilibrée en
éléments minéraux et en matières organiques dont le rapport carbone –azote est voisin
de 10 (C/N :10)ce qui permet la formation de brulé par la truffe noire(GRENTE et
DELMAS, 1974; GARCIA-MONTERO et al., 2007 ; 2009 ; ROBIN et al., 2008)
Le pH du sol est alcalin, qui favorise la production de la truffe en général et plus
précisément la truffe noire du Périgord Tuber melanosporumVitt. (GRENTE et
DELMAS, 1974 ; ROBIN et al, 2008)
L‘activité biologique du sol de la méso-faune « Oribates, Collemboles, Acariens, les
vers de terre.. », est importante, ces derniers jouent un rôle fondamental dans la
décomposition et l‘aération du milieu, qui est indispensable à la croissance du
champignon, et aussi la formation des boulettes fécales par les vers de terre tout ceci
aide à la formation de l‘ascocarpe, ainsi que la flore bactérienne qui est présente dans
les zones trufficoles (GRENTE et DELMAS, 1974; PARGNEY, 1994).
1-4- Les plantes hôtes de la truffe noire:
La truffe noire est un champignon mycorhizien, qui ne peut donc accomplir son
cycle qu‘avec une plante-hôte, ce sont pour la plupart des essences forestières
(CHEVALIER, 1985 ; CHEVALIER et FROCHOT, 1997). Dans la nature, certaines
essences sont plus aptes que d‘autres à produire des truffes (CHEVALIER et
FROCHOT, 1997). Les espèces forestières les plus répandues sont Quercus, Corylus
et l‘Ostrya, elles associent avec Tuber melanosporum dans les régions
méditerranéennes en particulier en France, Italie, Espagne (tableau 1) (CHEVALIER
et GRENTE, 1980 ; CASERMEIRO et al., 2007).
Cependant, on trouve en seconde classe d‘autres plantes hôtes comme le Hêtre, le
Pin et le Charme (tableau 1) (LE TACON et GARBAY, 1986 ; CHEVALIER et
FROCHOT, 1997) et certaines plantes sub-ligneuses tels que les Cistes et les
hélianthèmes (CHEVALIER et GRENTE, 1980) En Italie, les Cistes (Cistus
monspeliensis) font des associations symbiotiques dans des sites naturels avec Tuber
melanosporum (GARCIA-MONTERO et al., 2007).
Le choix de l‘essence forestière doit être compatible avec les exigences écologiques de
l‘espèce de truffe, et on doit tenir compte dans la culture de certaines caractéristiques

comme la sensibilité aux contaminants des racines, la morphologie du système racinaire
et la longévité de l‘arbre (CHEVALIER et FROCHOT, 1997).
Tableau 1 : Un aperçu sur le classement des arbres selon leur pouvoir mycorhizien
avec le Tuber melanosporum.
Classement Plante hôte Références
La première classe
(Feuillux)
Quercus, Corylus
et l‘Ostrya
CHEVALIER et GRENTE, 1980 ;
CHEVALIER, 1996 ;
CHEVALIER et FROCHOT,
1997 ; CASERMEIRO et al., 2007.
La deuxième classe
(Résineux)
Hêtre, Pinus et
Charme
GARBAY et TACON, 1986 ;
CHEVALIER, 1996 ;
CHEVALIER et FROCHOT,
1997.
La troisième classe Cistes, Hélianthème. CHEVALIER et GRENTE, 1980 ;
WENKART et al., 2001 ;
GARCIA-MONTERO et al., 2007.
1-5- Taxonomie :
Les truffes sont des champignons hypogés ectomycorhiziens appartenant au
genre Tuber ; elles sont mondialement connues pour leur valeur culinaire (SHAW,
1995 ; WANG et MARCONE, 2011). L‘espèce de Tuber la plus noble est le Tuber
melanosporum Vitt. qui a diverses appellations (tableau 2).
Le genre Tuber est nommé pour la première fois par MICHELI (1729)
signifiant les truffes à corps fructifères. Ce taxon regroupent les espèces de truffes
européennes et les plus intéressantes en trufficulture pour leur valeur économique
importante et leur distribution géographique (URBAIN et al., 2004 ; LE TACON et
al., 2006 ; MELLO et al., 2006 ; LAESSOE et HANSEN, 2007 ; MURAT et al.,
2008).
La nomenclature scientifique de Tuber melanosporum a été proposée par
VITTADINI (1831) puis repris par TRAPPE (2001) (PARGNEY, 1994 ; HANSEN
et LAESSOE, 2007).

Le genre Tuber signifie « truffe » et melanosporum « spore noire » qui veut dire truffe
à spores noires « anciennement nommée « Tuber nigrum » (RIOUSSET et al., 2001 ;
MOUSSU et LAURIAC, 2008).
Des études de Biologie moléculaire ont permis de donner une première
classification phylogénétique des espèces de Tuber réparties en 5 groupes dont celui
de Melanosporum (JEANDROZ et al., 2008) (Fig. 3). Puis une seconde plus
complète effectuée par BONITO et al. (2010) avec 9 groupes incluant celui de
Melanosporum (Fig.4).
Tableau 2 : Diverses appellations de Tuber melanosporum Vitt.
Appellations Commentaire Références
La truffe noble Se trouve sur les tables somptueuses des rois d‘Europe
mentionnée par PRADEL (1914).
(DESSOLAS et al., 2008)
Le diamant noir Diamant noir de la cuisine nommé par BRILLAT
savarin
(GRENTE et CHEVALIER,
1973)
La truffe noire du
Périgord
Récoltée pour la première fois au Périgord sous le
chêne
(WANG et MARCONE,
2011)
La vraie truffe A cause de sa valeur gustative, très parfumée et
estimée par rapport aux autres espèces du genre
Tuber.
(MOUSSU et LAURIAC,
2008)
La truffe Représente l‘exemple le plus célèbre de truffes
hypogées.
(CHEVALIER et
FROCHOT, 1997)
La famille des Tubéracées (DUMORT, 1822) regroupe le genre Tuber et
d‘autre genres à espèces hypogées (URBAIN et al., 2004 ; LAESSOE et HANSEN,
2007). Dans la classification ancienne, la famille des Tubéracées appartenait à
l‘ordre des Tubérales (KORF, 1973 ; TRAPPE , 1971) puis elle a été ensuite
transférée dans l‘ordre des Pézizales par TRAPPE (1979). Cette famille regroupait
seulement deux genres (Tuber et Paradoxa) puis LAESSOE et HANSEN (2007)

ont rajouté d‘autres genres (RIOUSSET et al., 2001 ; LAESSOE et HANSEN, 2007)
(Figs. 4 et 5)
Fig. 3: Classification du genre Tuber selon JEANDROZ et al. (2008) et BONITO et al. (2010).

Fig. 4 : L‘arbre phylogénique du genre Tuber (BONITO et al., 2010) .

Fig. 5: Une partie de l‘arbre phylogénique décrit par LAESSOE et HANSEN (2007)
montre le taxon des Tuberaceae.
L‘ordre des Pézizales regroupe 6 familles plus les Tuberaceae, la figure 6
montre la disposition taxonomique des Tuberaceae dans les Pézizales. Le caractère
d‘identification dans l‘ordre des Pézizales est basé sur le nombre de noyaux par
spore, pour le Tuber melanosporum à 6-8 noyaux par spore (HANSEN, 2006 ;
LAESSOE et HANSEN, 2007).
Les Pézizales sont classées dans les Euascomycètes caractérisés par un thalle
filamenteux et dans la sous-classe des Discomycètes.
La répartition des espèces est basée sur la qualité de l‘ascocarpe et de l‘hyménium ; les
espèces dont les spores se forment dans l‘asque font partie des Ascomycètes
(Ascomycota) (ROUX et al., 1999 ; RIOUSSET et al., 2001 ; URBAIN et al.,2004 ;
HANSEN, 2006 ; BONITO et al., 2011).
Les premières études génétiques basées sur le noyau truffier ainsi que les
chromosomes ont été effectuées par BONFANTE (1973) sur certaines espèces de
Tuber dont Tuber melanosporum. De nombreux travaux ont ensuite été effectués
sur le génome de Tuber melanosporum (BERTAULT et al.,1998 ; PERCUDANI et
al. , 1999 ; MABRU et al. , 2004 ; MURAT et al. , 2004 ; WANG et al. , 2006 ;
RUBINI et al. , 2011). Certains chercheurs ont séquencé tout le génome de Tuber
melanosporum et ont montré que la séquence de cette espèce de Tuber est la plus

grande (125 mb) et la plus complexe que celle des autres espèces de Tuber
(MARTIN et al., 2010 ; RUBINI et al., 2011).
Fig. 6 : La position des Tuberaceae dans l‘ordre des Pézizales (LAESSOE et
HANSEN, 2007).
2 -Quelques notions sur la truffe noire du Périgord (Tuber melanosporum
Vitt.)
La truffe noire est le résultat d‘une fructification souterraine d‘un champignon ;
ce précieux tubercule appelé Tuber melanosporum Vitt. a des propriétés
organoleptiques et un intérêt économique important (Fig. 7). Ce champignon fait
l‘objet de nombreuses recherches dans différents domaines, génétique, chimique,
mycologique, agriculture, etc.
Pézizales

Fig. 7 : Ascocarpes de la truffe noire du Périgord (A) (LAURENT, 2003) et
fructification souterraine de la truffe noire (B) (PARGNEY et MEUNIER, 2004).
2-1-Comparaison entre la truffe et un champignon épigé :
La truffe noire nommé « la vraie truffe » appartient aux Ascomycètes et c‘est un
champignon hypogé (fructification souterraine ou ascocarpes) contrairement aux autres
champignons épigés appartenant aux Basidiomycètes dont les fructifications ou
carpophores sont aériennes (O‘DONNEL, 1997 in BONUSO, 2008).
Chez la truffe, la respiration se fait par absorption de l‘air par l‘intermédiaire des veines
aérifères qui constituent en partie sa trame et aboutissent au péridium, par contre, chez
le champignon épigé la respiration est normale comme les végétaux (PRADEL, 1914)
(Tableau 3).
La truffe est nommée aussi « modus vivendi » (CHEVALIER et al. ,2008). Selon
CHATIN (1892), elle possède le spongiole un mycélium court lui permettant
d‘absorber des nutriments lorsqu‘elle coupe ses connexions avec la racine de la plante
hôte (CALLOT et al., 1999).
Tableau 3 : Différences entre la truffe et le champignon épigé (MANCHABLON et
PARGNEY, 2003 ; CHEVALIER et al., 2008).
Caractères Tubercule truffier Champignon épigé
Espèce
souterraine aérienne
L‘élément foliacé
absence absence
Respiration
Absorbe l‘air par des veines Respire à l‘air libre
Famille
Ascomycète Basidiomycète
Exemple
Tuber melanosporum Amanita ponderosa
2.2- Les composants morphologiques du corps fructifère truffier :

Le poids du corps fructifère truffier ou ascocarpe que l‘on récolte est variable
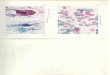
(Fig. 8 A) celui de Tuber melanosporum (CALLOT et al., 1999).
L‘ascocarpe est constitué par le péridium, la trame, les tissus de soutien et fertile
Fig. 8: (A) Un ascocarpe de Tuber melanosporum de poids important (CALLOT,
1999) ; (B) Aspect du péridium de Tuber melanosporum (BENAC, 2006).
2-2-1- Le péridium :
Le péridium ou écorce externe, est une enveloppe protectrice qui est formée par
des écailles pyramidales plus ou moins proéminentes ou de filaments agglomérés
(CHEVALIER et al., 2008), sa surface est rugueuse et parsemée d‘aspérités
verruqueuses. Il exprime certain type de couleur selon le stade de maturité de la truffe ;
celui de Tuber melanosporum a une couleur brun-noir (Fig.8B) (CHEVALIER et
GRENTE, 1980 ; BENAC, 2006). Le péridium est composé parfois par des veines
aérifères (in CHEVALIER et al., 2008).
TANG et al. (2008) ont constaté pour la première fois un phénomène de
fermentation lors du stockage des ascocarpes de Tuber melanosporum provoquant la
formation de biomasses d‘exo-polysaccharides (EPS) et d‘intra-polysaccharide (IPS)
sur le corps fructifère. Ce phénomène est encore mal optimisé (TANG et al., 2008).
PARGNEY et al. (2008) a observé un phénomène similaire sur la partie externe des
ascocarpes présents dans le sol. Ces ascocarpes sont recouverts de matériaux divers
provenant probablement du recyclage continuel des organismes de surface, de débris
d‘hyphes et de matières végétales, de divers cadavres d‘animaux et de micro-
organismes (bactéries, levures) et aussi de l‘accumulation des rejets de la truffe
(PARGNEY et al., 2008) telle que la cristallisation de surface. Ces mêmes auteurs ont
observé chez la truffe deux cas de cristallisation suivants :
-Une cristallisation de surface avec polysaccharides :

Ce sont des précipitations blanchâtres très riches en carbonate de calcium qui
s‘organisent en plaques et se développent entre les stries d‘accroissement des écailles
du péridium. Ces concentrations minérales sont en relation avec les polysaccharides
qui favorisent le développement des microorganismes « bactéries » ; ces plaques
carbonatées assurent une protection contre le dessèchement de la truffe par leur
superposition sur les porosités du péridium (PARGNEY et al., 2008 ; TANG et al.,
2008).
-Une cristallisation de surface par l‘anion d‘oxalate :
L‘anion oxalate qui est abondant dans le sol, et en présence de calcium secrété
par le champignon, il forme des précipitations d‘oxalate de calcium (PARGNEY et al.,
2008) ; chez la truffe, ce phénomène ne modifie pas le pH du milieu environnant
comme le cas des autres champignons (CALLOT et al., 1999, PARGNEY et al., 2008).
2-2-2- La trame :
La truffe contient un tissu qui est « la trame » à texture granuleuse et
hétérogène, appelée aussi « gléba ou la glèbe » ou « la chair » (Fig. 9). Cette dernière
est subdivisée en deux tissus complémentaires : le tissu de soutien et le tissu fertile
(CHEVALIER et GRENTE, 1973, 1980; CHEVALIER et al., 2008).
Fig. 9 : Aspect de la glèbe (A et B) et du péridium de Tuber melanosporum (BENAC,
2006).
2-2-2-1- Le tissu de soutien :
A
glèbe

Le tissu de soutien à une texture granuleuse, constitué par des filaments de plus
en plus agglomérés ayant l‘aspect des veines blanches (CHEVALIER et GRENTE,
1980 ; BENAC, 2006)
2-2-2-2- Le tissu fertile :
Le tissu fertile est une zone sombre appelée « hyménium », il est replié en
circonvolutions cérébrosides ce qui donne à la gléba cet aspect marbré. Mais une glèbe
toute blanche correspond à une truffe immature (BENAC, 2006 ; CHEVALIER et al.,
2008), L‘hyménium est constitué principalement par l‘asque et les ascospores.
L‘asque :
Les asques « sacs » *« thèques » ou « sporanges» ce sont les cellules terminales
des veines blanches. Selon RIOUSSET et al (2001), les Tuber possède des asques
globuleux, entourés par une membrane très mince. Ils contiennent un liquide
protoplasmique, composé de gouttelettes d‘huile et en même temps des petits corps
arrondis ou ovales, de couleur brun foncé et entouré par des épines qui sont les
caractéristiques de la spore. Cette cellule terminale subit des transformations, tantôt elle
apparait arrondie et dès lors c‘est une thèque, tantôt elle revêt la forme d‘un corps
ovulaire allongé et constitue la paraphyse. (GRENTE et CHEVALIER, 1980 ;
GRENTE et DELMAS, 1974 ; LAURENT, 2003 ; CHEVALIER et al., 2008).
L‘ascospore :
Les ascospores sont les corpuscules noires contenus dans l‘asque au nombre de
2 à 5, de quelques centièmes de millimètres de diamètre en forme « de ballon de
rugby » et qui sont généralement ellipsoïdes pour le Tuber melanosporum (Fig. 10 A)
et ornementées (Figs. 10 B et C). Les ascospores immatures sont toujours hyalines ;
leur couleur change selon le degré de maturité. Leur morphologie permet de distinguer
les espèces et leur ornementation, elle est importante en systématique (CHEVALIER
et GRENTE, 1980 ; RIOUSSET et al., 2001 ; CHEVALIER et al., 2008 ; ROBIN et
al., 2008).
PARGUEY-LEDUC (1992) a montré que le tissu externe de la spore de Tuber
melanosporum est de nature chitineuse. D‘autres chercheurs ont utilisé la spore pour

le séquençage de l‘ADN de Tuber melanosporum (MABRU et al., 2004).
Fig. 10 : Morphologie des ascospores de Tuber melanosporum.
(A) ; (B) : Ascospores de Tuber melanosporum de forme ovulaire contenues
dans des asques (ROBIN et al., 2008).
(C)Ascospores observées au microscope électronique à balayage montrant les
épines (CALLOT et al., 1999).
Il existe de nombreuses espèces de truffes intéressantes, une trentaine d‘espèces
sont répandues dans les zones de production en France, Italie, Espagne ; certaines
espèces sont utilisées pour la production de plants mycorhizés telles que la truffe noire
et la truffe de bourgogne et le reste pour la consommation humaine (CHEVALIER et
FROCHOT, 1997 ; LAURENTE, 2003) (Tableau 4).

Tableau 4 : Principales caractéristiques morphologiques de quelques espèces de Tuber.
2-3- Composition chimique de la truffe noire du Périgord
Les truffes reconnues comme un additif alimentaire ont une valeur nutritive importante
pour l‘être humain. Elles contiennent 75 % d‘eau, elles sont riches en protéines qui
représentent une source énergétique importante pour l‘homme (MACONNEL et
ESSELEN, 1947 ; WANG et MARCONE, 2011) en glucides ainsi qu‘en sels
minéraux par contre, le pourcentage des lipides est très minime dans un ascocarpe
mature (PINKAS et al., 2000 ; HARKI et al, 2006) (tableau 5).
Elles sont riches en acides aminés Pour l‘arôme et la flaveur de ce précieux tubercule
de Tuber melanosporum , le groupement sulfure se lie avec une large gamme d‘acides
aminés (DIAZ, 2003 ; HARKI et al., 2006). La dégradation de certains acides aminés
libère des groupements aldéhyde et alcools qui sont responsables de l‘arôme (DIAZ et
al., 2003 ; HARKI et al., 2006 ; WANG et MARCONE, 2011). La composition
nutritionnelle ou chimique de la truffe noire du Périgord, change durant le temps
(tableau 6).
Parmi les truffes reconnues dans le monde, la truffe noire diffère des truffes du désert
par sa morphologie, sa « couleur », son écologie et sa composition chimique. Les
truffes du désert sont plus riches en glucides et en sels minéraux que la truffe noire du
Périgord (tableau 5). HARKI et al . (2006) ont étudié la composition chimique de
Tuber melanosporum Vitt. pendant les trois derniers stades de sa maturité (tableau 6).

Tableau 5: Composition chimique de deux types de truffe (HARKI et al., 2006 ;
WANG et MARCONE, 2011).
Tableau 6 : Composition chimique de la truffe noire du Périgord pendant trois
derniers stades de sa maturité (HARKI et al., 2006).
Stade M1% Stade M2% Stade M3%
Protéines 27.6 31.7 29.7
Glucides 8 11 30.6
Lipides 7.8 5 5.4
Stérols 1.2 0.9 0.6
Mélanine 3.1 8.3 15.2
Sels minéraux 5 7.3 7.4
T. melanosporum est riche en glucose (tableau 6) et au dernier stade de
développement de l‘ascocarpe, la teneur en sucres est très élevée, elle représente
30.6% du poids sec total du corps fructifère (tableau 4) (HARKI et al., 2006). Selon
Truffe du désert (%) Truffe noire (%)
Protéines 20-27 29.7
Glucides 60 30.6
Lipides 3-7.5 5.4
Sels minéraux 7-13 7.4
Mélanine 0 15.2
Eau 70 75

LE TOURNEAU (1966) la composition des carbohydrates ou glucides est importante
pour le développement de l‘ascocarpe et pour la structure des spores qui nécessite une
quantité abondante de sucres (LE TACON et GARBAYE, 1986 ; HARKI et al.,
2006).
Les stérols représentent 1.2 % avant la maturation de l‘ascocarpe et 0.6% pendant la
maturation donc Tuber melanosporum est pauvre en stérols ainsi qu‘en lipides (tableau
6), l‘ergostérol et brassicastérol sont des composants essentiels de la membrane des
organismes eucaryotes, T. melanosporum comporte une quantité minime de
l‘ergostérol (HARKI et al., 2006), ce dernier a un effet bénéfique pour la santé comme
un anti-hyper-lipidique, anti-inflammatoire, anti-oxydatif et aussi a un effet inhibiteur
sur des champignons et bactéries (TANG et al., 2008).
La quantité de lipides est minime, elle représente 5.4% du poids sec dans
l‘ascocarpe mature; la quantité d‘acide linoléique est par contre importante (tableau
7), ce dernier est responsable de l‘arôme de la truffe noire dont la qualité différencie
les variétés de truffes (DIAZ et al., 2003 ; HARKI et al., 2006 ; WANG et
MARCONE, 2011). Selon TANG et al. (2008), les stérols produits lors de la
fermentation de surface de l‘ascocarpe par des espèces microbiennes comme les
bactéries libèrent aussi des composés volatils tels que les androstérols,
Tableau 7 : Composition chimique en pourcentage des principaux composants au
stade mature de Tuber melanosporum (HARKI et al, 2006).
de même certains acides gras et nucléosides formés, provoquent la germination des
ascospores de la truffe (TANG et al., 2008).
La coloration noire de T. melanosporum est due à la présence de la mélanine
(15%) (HARKI et al., 2006). La composition chimique des Tuber varie
quantitativement selon les espèces, l‘âge des ascocarpes, leur origine, le type de sol,
ainsi que le stade de développement (HARKI 2006).

3- Les ectomycorhizes :
3-1- Généralités :
Les mycorhizes sont des associations symbiotiques à bénéfice réciproque entre
un champignon et une plante. Il existe deux grands groupes de mycorhizes : les
ectomycorhizes et les endomycorhizes. Les ectendomycorhizes forment un groupe
intermédiaire (Fig. 1).
Chez les ectomycorhizes, les partenaires fongiques sont des Basidiomycètes
(Amanites, Bolets..) et /ou des Ascomycètes (Truffes). Elles se rencontrent chez 3 à 5%
des espèces végétales forestières et économiquement importantes (MEOTTO, 1996 ;
ROBIN et al., 2008) (tableau 8).
Ce type d‘association permet le maintien de la biodiversité végétale et fongique,
la régénération des fonctions et des cycles biogéochimiques qui touche indirectement
l‘écosystème et l‘amélioration de la nutrition minérale de l‘arbre ainsi que sa
protection contre les stress biotiques (agents pathogènes) (PLASSARD et al., 1997 ;
SELOSSE, 2006 ; DRENOU, 2006 ; BA et al., 2011 ; CAIRNEY, 2012).
A

Fig. 11: Type mycorhizien.
A : Structures de colonisation racinaire de la symbiose ectomycorhizienne (1) et
endomycorhizienne de type arbusculaire (2) (BONFANTE et GENRE, 2010).
B : Schéma d‘une coupe transversale montrant les principaux types d‘association
mycorhizienne entre des champignons du sol et des racines de végétaux
(a : racine sans mycorhize, b :endomycorhizes à vésicules et à arbuscules, c : endomycorhizes à
pelotons, d : ectendomycorhizes, e : ectomycorhizes chez les angiospermes, f : ectomycorhizes
chez les Gymnospermes) (DUHOUX et NICOLE, 2004).
Tableau 8 : Familles et genres d‘arbres formant des ectomycorhizes (DRENOU,
2006 ; BA et al., 2011).
Le champignon infecte la racine courte de ces arbres et donne des
ectomycorhizes qui peuvent être simples, racémeuses, coralloides et dichotomes (Figs.
12 A et B) (SELOSSE, 2000 ; BA et al., 2011). Le mycélium entoure la racine courte
formant le manteau et les hyphes s‘insinuent entre les cellules corticales formant le
Familles d‘arbres Exemples de genres
Pinacées
Fagacées
Bétulacées
Salicacées
Tiliacées
Myrtacées
Juglandacées
Pins, sapins, épicéas, mélèze, tsuga, pseudotsugas, cèdres.
Chênes, hêtres, châtaigniers.
Bouleaux, noisetiers, aulnes, charmes.
Saules, peupliers.
Tilleuls.
Eucalyptus.
Noyers, caryas, ptérocaryas.

réseau d‘Hartig ; il y a absence des poils absorbant sur la racine mycorhizée (Fig. 12
C) (PLASSARD et al., 1997 ; DESSOLAS et al., 2008). Le réseau d‘hyphes externe
ou extra matricielles (Fig.16, D, E) colonise le sol et infecte d‘autres racines
(STRULLU, 1991 ; DEXHEIMER, 1997 ; CAIRNEY, 2012).
L‘identification des mycorhizes est utile dans le cas d‘un champignon d‘intérêt
économique tel est le cas de Tuber utilisé pour la production des plants mycorhizés
en pépinière et des carpophores (CHEVALIER et GRENTE, 1973 et 1978 ;
CHEVALIER, 1996). L‘identification et la caractérisation des espèces fongiques et
des ectomycorhizes reposent sur des examens macroscopiques et microscopiques
(couleur des ectomycorhizes, structure de manteau fongique…) et des études
moléculaires (DIAZ et al., 2003 ; BONITO et al., 2011). Les connaissances sur la
morphologie et l‘anatomie des ectomycorhizes permet de connaitre la diversité
fongique ainsi que les populations de champignons mycorhizogènes et quantifier le
potentiel des sols (TORRES et HONRUBIA, 1994 ; DIAZ et al., 1996 ;
DOMINGUEZ et al., 2005) et vérifier la mycorhization contrôlée (CHEVALIER et
POLACSEK, 1973 ; PEREZ et al., 2007 ; MOUSAIN et al., 1994) finalement
connaitre la performance du champignon inoculé (TAGLIAFERRO, 1996 ;
LAQUBAQUBI et al., 2011).

3-3- Les mycorhizes de la truffe noire
:
3-2- Les mycorhizes et le mycélium truffier
3-2-1 Les ectomycorhizes de la truffe
D
b
A B
C
E D
D
Fig. 12: La forme d‘une ectomycorhize.
(A) Les différents types de mycorhize (SELOSSE, 2000)
(B) Forme dichotomique des mycorhizes de pin d‘Alep (SELOSSE et HELME-GUIZON, 2010).
(C) La différence entre une racine mycorhizée et une racine non mycorhizée (PLASSARD et al., 1997)
(D) La forme schématique d‘une ectomycorhize longitudinale (JANY, 1998)
(E) La forme schématique d‘une coupe transversale d‘une ectomycorhize (SELOSSE, 2000)
(F) La forme schématique d‘une coupe transversale d‘une éctomycorhize (SELOSSE, 2000)

3-2- les mycorhizes et le mycélium truffier :
3-2-1- les ectomycorhizes de la truffe :
Les mycorhizes de truffe sont de type « ectomycorhize » (PARGNEY, 1994) , leur
structure est complexe et comprend plusieurs zones (DEXHEIMER, 1997 ; DRENOU, 2006).
Elles ressemblent à des massues courtes et épaisses portées par les racines longues de la plante
hôte (Fig. 13) (GRENTE et CHEVALIER, 1973 ; 1980 ; PARGNEY, 1994) plus précisément
les racines secondaires (PARGNEY et al., 2008) avec différents aspects selon les plantes hôtes
associées (SELOSSE et HELME-GUIZON, 2010).
B
3-2-2- L‘organisation du mycélium truffier
3-2-2- 1- L‘organisation extra-racinaire du mycélium
Le mycélium en contact avec des racines permet d‘induire le développent des courtes
racines, le mycélium truffier produit l‘acide acétique -3-indole et l‘éthylène qui donne un
signal à la plante pour la formation des racines primaires et latérales (SPLIVALLO et al.,
2007, 2009). Le mycélium est subdivisé en hyphes, une partie des hyphes s‘organise auteur de
la racine initiée en formant une gaine protectrice appelée manteau (PARGNEY, 1994 ;
DRENOU, 2006), Les excroissances externes issues des hyphes du manteau qui s‘allongent et
pénètrent dans les particules du sol forment un mycélium frangeant (PARGNEY et al., 2008).
Le mycélium frangeant :
Nommé aussi hyphe émanant (AGERER, 2006), il caractérise les mycorhizes de truffe, sa
morphologie et sa structure varie selon les espèces de truffes (PARGNEY, 1994). Il est
subdivisé en deux organes importants pour le développement de la mycorhize les spinules et
B

les hyphes extra matriciels qui explorent le milieu environnemental, augmentent
considérablement les zones de contact entre le sol et le manteau fongique, ce sont toutefois
des structures temporaires qui se développent dans certaines conditions environnementales,
l‘humidité et le compactage du sol sont des facteurs déterminants dans l‘extension de ce
mycélium frangeant (PARGNEY et al., 2008).
Les spinules :
Dans le cas des Tuber, les cystides sont nommées les spinules (AGERER, 2006,
PEREZ et al., 2007). Ce sont des ornementations caractéristiques dont la morphologie varie en
fonction des espèces. Dans le cas de T. melanosporum, elles sont longues, pigmentées,
cloisonnées et ramifiées à angle droit (Fig. 14) (PARGNEY, 1994 ; PARGNEY et
MA NCHABLON, 2003 ; PARGNEY et al., 2008). Les ramifications des spinules ne sont pas
observables dans toutes les mycorhizes examinées (MEOTTO, 1996) ; elles ont une paroi
épaisse donnant la rigidité qui leur permet de résister aux contraintes externes du sol
(PARGNEY et al., 2008).
Fig. 14 : Forme de spinules chez des espèces de Tuber (PARGNEY et MANCHABLON, 2003).
Les hyphes extra matriciels :
Ces hyphes ont par contre, des parois plus fines et sont donc plus souples ; ils peuvent
s‘insinuer facilement dans les particules du sol (PARGNEY et al., 2008) et ont la capacité de
mobiliser des nutriments et minéraux (GRENTE et DELMAS,1974 ; PLASSARD et al.,
1997 ; AGERER, 2001 ; CAIRNEY, 2012).
3-2-2- 2 - L‘organisation du mycélium intra-racinaire :

Le reste des hyphes pénètrent dans la racine en s‘insinuant entre les cellules racinaires,
mais sans jamais pénétrer dans ces cellules ; elles forment le réseau de Hartig (Fig. 15). La
racine qui héberge le mycélium fongique se transforme morphologiquement, la morphogénèse
des cellules corticales sera modifiée ainsi que son allongement sera réduit. Ce contact entre
les hyphes et les cellules racinaires va augmenter les zones d‘échanges entre le mycélium et la
racine et faciliter les relations, la réciprocité d‘échanges aboutit à une symbiose entre les deux
organismes (PARGNEY, 1994 ; DEXHEIMER, 1997 ; PARGNEY et al., 2008 ; CAIRNEY,
2012).
Fig. 15 : Coupe longitudinale dans une ectomycorhize de Truffe dont : M est le manchon dense
auteur de la racine ainsi que le réseau d‘ Hartig et allongement de l‘assise corticale
(DEXHEIMER, 1997).
3-2-2- 3- La truffe et les stromas :
Il y a un autre type de relation mycélium-racine, ce sont des proliférations anarchiques
fongiques à la surface des racines longues d‘arbres mycorhizés formées par différentes espèces
de truffes (genre Tuber), elles se présentent sous l‘aspect de boursouflures, plus ou moins
juxtaposées. Certaines portions racinaires peuvent en porter, alors que les voisines en sont
dépourvues (PARGNEY et al., 2008) Les stromas sont constitués d‘hyphes agrégés, au début
leur surface est lisse, mais lorsqu‘ils prolifèrent, elle devient granuleuse (Fig. 16) (PARGNEY
et JALADE, 1995 ; GUINBERTEAU et COURTECUISSE, 1997), on trouve :
Les hyphes plus superficiels émergeant de la surface.
Les hyphes les plus profonds.

Fig. 16 : Aspect schématique d‘un stroma (PARGNEY et al., 2008).
3-2-3- Le mycélium truffier et la vie sur terre :
Le mycélium truffier peut effectuer deux phases : la phase végétative et la phase
reproductive qui sont importantes pour sa vie en terre qui aboutit à la fin à former la truffe
(Tableau 9 ; Fig. 17) (GRENTE et DELMAS, 1974 ; GRENTE et CHEVALIER, 1980 ;
CHEVALIER et FROCHOT, 1997 ; DESSOLAS et al., 2008).
Dans le cycle de vie de la truffe, différent facteurs, tels que les caractéristiques du sol,
le pH, les éléments nutritifs et les conditions climatiques, les caractéristiques de la plante hôte,
peuvent influencer le mycélium truffier ainsi que l‘installation de la mycorhization et
l‘obtention d‘un corps fructifère (GARCIA-MONTERO et al., 2007, 2009 ; KUES et
MARTIN, 2011).
Tableau 9 : Différence entre les deux phases végétative et reproductive (GRENTE et
DELMAS, 1974 ; GRENTE et CHEVALIER, 1980 ; CHEVALIER et FROCHOT, 1997 ;
DESSOLAS et al., 2008).
Fig. 17: Schéma explicatif des deux phases de la vie de la truffe (NB : les proportions entre
les différents organismes ne sont pas respectées), (DESSOLAS et al., 2008).
Le mycélium truffier et la phase végétative :
Le cycle commence par la germination de la spore libérée par la dégradation de
l‘ascocarpe truffier, ainsi que l‘intervention de la microfaune du sol, elle émet un tube
germinatif donnant un filament mycélien (mycélium truffier) haploïde MAT+ ou MAT- qui
s‘unissent aux extrémités racinaires (GRENTE et DELMAS, 1974 ; ROUQUEROL et

PAYRE, 1975 ; KUES et MARTIN ; 2011). Ce mycélium truffier (Fig. 19 B) peut être
propagé par des espèces animales (mouche truffigène, porc, chien, limaces et petits rongeurs)
(CALLOT et al., 1999; DESSOLAS et al., 2008).
Le mycélium truffier produit l‘acide acétique 3-indole et l‘éthylène qui est un signal pour les
plantes hôtes pour synthétiser des racines latérales et des racines courtes (Fig. 19 C)
(SPLIVALLO et al., 2007 ; 2009) Les hyphes forment le manteau pseudo parenchymateux
auteur de la racine courte (Fig. 19 D) (KUES et MARTIN, 2011), ils peuvent aussi formant des
stromas par l‘infection des racines longues (DESSOLAS et al., 2008). Ils colonisent les cellules
rhizodermiques, les cellules corticales en formant le réseau d‘Hartig (Fig. 20 E) (KUES et
MARTIN, 2011) et les espèces bactériennes facilitent cette association dans l‘interface
d‘échange entre les deux partenaires.
Le mycélium extra matriciel issu du manteau colonise le sol pour rechercher l‘eau et les
minéraux pour la plante et en contrepartie il reçoit les composés carbonés (Fig. 18).
Fig. 18: Schéma simplifié du principe d‘une association symbiotique entre la truffe et l‘arbre
truffier.
Le mycélium truffier et la phase reproductive :
Après la formation des mycorhizes (GRENTE et DELMAS, 1974 ; CHEVALIER et al.,
2008 ; KUES et MARTIN, 2011) il y a apparition d‘une zone claire autour de l‘arbre « le
brulé » résultat d‘une action phytotoxique vis à vis de certains microorganismes (PARGNEY,
1994 ; GARCIA-MONTERO et al., 2007)
Des enroulements mycéliens forment des pelotons, les filaments (MAT+) et (MAT-) après
reproduction sexuée conduisent à la formation du primordium ou jeune ascocarpe (Fig. 19
G, H) (GRENTE et DELMAS, 1974 ; MELLO et al., 2006 ; RUBINI et al., 2011 ; MARTIN
et al., 2010 ; KUES et MARTIN, 2011). Ce dernier coupe ses connexions avec la plante hôte
et se nourrit par les filaments mycéliens externes puis termine sa maturité par la constitution
verruqueuse de son péridium (GRENTE et DELMAS, 1974 ; CHEVALIER et al., 2008) La
formation d‘un ascocarpe de T. melanosporum s‘effectue dans les 6 à 8 mois (KUES et
MARTIN, 2011).

Les ascocarpes matures peuvent ensuite être récoltés par le chien ou le porc (Fig.19I)
(GRENTE et CHEVALIER, 1980 ; GRENTE et DELMAS, 1974 ; CHEVALIER et al.,
2008 ; MARTIN et KUES, 2011).
3- 3- Production et commercialisation des plants mycorhizés par la truffe
La mycorhization des plants nécessite la production d‘inoculum fongique. De
nombreuses études ont été effectuées pour produire du mycélium pour la trufficulture car il
est le point de départ du développement de la truffe et le trait d‘union entre la truffe et l‘arbre
producteur, ce qui explique pourquoi la truffe se développe à côté d‘un arbre précisément
dans le périmètre occupé par le système radiculaire (BENAC ,2006 ; DESSOLAS et al., 2008).
Or, l‘isolement du mycélium de Tuber est difficile à obtenir, on en connait aucun milieu
favorable à la germination des ascospores de Tuber. De nombreux essais de germination des
ascospores de T. melanosporum ont été effectués par certains chercheurs (GRENTE et al. ,
1972 ; ROUQUEROL et PAYRE , 1975a) ainsi que l‘obtention du mycélium (FONTANA ,
1968 ; ROUQUEROL et PAYRE ,1975b).
Le mycélium de T. melanosporum a aussi été isolé à partir de mycorhize (CHEVALIER,
1972 et 1973 ; BRUNEL, 1973) cependant la production de mycélium à grande échelle est
toujours inexistante.
L‘inoculation des arbres avec T. melanosporum peut s‘effectuer par des méthodes
anciennes qui consistent à placer au contact des radicelles de la plante hôte le mycélium
truffier (méthode de BOULANGER , 1899) ou la solution sporale ( méthode de BRESSY
in CHEVALIER et GRENTE, 1978) (Fig. 20).
Une autre méthode a été proposée par GRENTE (1972), elle consiste à introduire dans le
sol des plants déjà mycorhizés par la truffe (CHEVALIER et GRENTE, 1973). Ces plants
mycorhizés sont le résultat final des travaux des chercheurs de l‘INRA de Clermont Ferrand
(France) et de TURIN (Italie) pendant de nombreuses années et leur commercialisation
sous le nom « des plants mycorhizés par la truffe selon le procédé INRA-ANVAR. » est
assurée par la société ANVAR)(GRENTE et DELMAS, 1974, CHEVALIER et GRENTE,
1980 ; CHEVALIER, 1985 ; DESSOLAS et al., 2008).

Fig. 19: Cycle de vie de Tuber melanosporum (KUES et MARTIN, 2011).

Fig. 20: Schéma simplifié de mycélium truffier entre l‘ancienneté et la modernité (d‘après :
GRENTE et DELMAS, 1974, CHEVALIER et GRENTE, 1978 ; CHEVALIER et
GRENTE, 1980 ; CHEVALIER, 1985 ; DESSOLAS et al., 2008).
4 – Caractéristiques botaniques de chêne vert
4—1 Taxonomie
Le chêne vert est l‘arbre qui s‘associe avec la truffe noire au Périgord et qui a fait l‘objet
de nombreuses études à l‘INRA de Clermont Ferrant en 1970. Il est devenu essentiel en
trufficulture, en sylviculture et pour la commercialisation par des pépiniéristes. Connue aussi
sous l‘appellation chêne yeuse, il est dominant dans les régions méditerranéennes, son nom
scientifique est Quercus ilex L. et il appartient à la famille des Fagacées (Tableau 10)
(GRENTE et DELMAS, 1974 ; CHEVALIER et GRENTE, 1978 ; BARBERO et LOISEL,
1980 ; CHEVALIER, 1996 ; CHEVALIER et FROCHOT, 1997 ; KREMER, 2005 ; WANG
ET MARCONE, 2011). Son nom vernaculaire en l‘Algérie est « kerruch »ou « chajret al
bellut ».
Tableau 10 : Taxonomie du chêne vert (in HAICHOUR, 2009).
Taxon Nomination
Embranchement Spermaphyte
Sous-embranchement Angiospermes
Classe Dicotylédone
Ordre Fagales
Famille Fagacée
Genre Quercus
Espèce Quercus ilex

Quercus ilex ou chêne vert est l‘arbre qui s‘associe avec la truffe noire au Périgord et
qui a fait l‘objet de nombreuses études à l‘INRA de Clermont Ferrant en 1970. Il est devenu
essentiel en trufficulture, en sylviculture et pour la commercialisation par des pépiniéristes.
Connue aussi sous l‘appellation chêne yeuse, il est dominant dans les régions
méditerranéennes, son nom scientifique est Quercus ilex L. et il appartient à la famille des
Fagacées (Tableau 10) (GRENTE et DELMAS, 1974 ; CHEVALIER et GRENTE, 1978 ;
BARBERO et LOISEL, 1980 ; CHEVALIER, 1996 ; CHEVALIER et FROCHOT, 1997 ;
KREMER, 2005 ; WANG ET MARCONE, 2011). Son nom vernaculaire en l‘Algérie est
« kerruch »ou « chajret al bellut ».
4--2 - Description morphologie du chêne vert:
Le chêne est un espèce polymorphe, à diversité morphologique variable, difficile à
décrire avec précision, il s‘hybride facilement avec différents types du chêne, ce qui explique
que Quercus rotundifolia est une variété du Quercus ilex. Quercus rotundifolia est fréquent en
Afrique du nord (BARBERO et LOISEL, 1980, HAICHOUR, 2009). Ses caractéristiques
morphologiques sont regroupées dans le tableau 11.
4-3- L‘habitat et répartition :
Le chêne vert occupe une superficie mondiale de 200.000 ha des arbres forestiers les
plus importants dans la région méditerranéenne. Il est réparti dans tout le Bassin
méditerranéen (Fig. 21), de la Grande Bretagne jusqu'à l‘Himalaya ; les espèces les plus âgées
se rencontrent en Asie centrale (DEHMANI-MEGREROUCH, 2002).
Il est présent en Afrique du nord et à l‘ouest de la méditerranée, de la plaine à la montagne
et peut atteindre une altitude de 1100m jusqu‘à 2200m dans les montagnes de l‘Atlas
(BARBERO et LOISEL, 1980 ; BELKHADER, 2003 ; KERMER, 2005) ; Il constitue une
part importante des forêts algériennes avec une superficie de 108000 ha, répartie de la frontière
marocaine à la frontière tunisienne, du littoral à l‘atlas saharien (Fig. 22) (FERKA ZAZOU,
2006 ; ALLAIN, 2007 ; HAICHOUR, 2009). C‘est l‘une des principales espèces de feuillus
méditerranéennes (KERMER, 2005 ; ALLAIN, 2007)

Le chêne vert recouvre ou colonisent les régions tempérées aussi bien dans les zones littorales
et sub-littorales qu‘en montagne humide, il représente l‘essence dominante du midi-
méditerranéen (BARBERO et LOISEL, 1980).
Tableau 11: Description morphologique du chêne vert (BELKHADER, 2003 ; NACEF et
DJERNOUMI, 2004 ; KREMER, 2005 ; ZITOUNI, 2010).
Il pousse dans des sols riches en humus, lourds et souvent calcaires (CHEVALIER et
GRENTE, 1980 ; GRENTE et DELMAS, 1974 ; CHEVALIER et FROCHOT, 1997 ;
KREMER, 2005) ; ces deux paramètres sont les plus importants pour le développement de
Tuber melanosporum (CHEVALIER et FROCHOT, 1997.
Le chêne vert couvrait à l‘origine d‘importants espaces dans les régions méditerranéennes, il a
été trop exploité dès l‘Antiquité (KREMER, 2005).
4-4- Utilité :
- Côté économique : le bois du chêne vert est solide et compact, et très recherché en
construction navale ainsi qu‘en charpenterie et comme bois d‘œuvre, son écorce fournit un
tan d‘excellente qualité (BELKHADER, 2003 ; KERMER, 2005) et produit un charbon de la
premier qualité (ZITOUNI, 2010)

- Côté rural : les glands sont consommés et utilisés pour extraire une huile de saveur douce en
Algérie (BELKHADER, 2003).
- Côté thérapeutique : les feuilles, l‘écorce et les glands sont indiqués en thérapeutique comme
des calmants et l‘écorce est utilisée contre les gerçures et les dermatoses et en poudre
antihémorragique (BELKHADER, 2003 ; DJERNOUMI et NACEF, 2004 ; ALLAIN, 2007).
Fig. 21: Répartition du chêne vert (Quercus ilex) dans l‘hémisphère du nord (BARBERO et
LOISEL, 1980)
N

Fig. 22 : Distribution du chêne vert (Quercus ilex) en Algérie (HAICHOUR, 2009).

Chapitre II : Matériel et méthodes
1 – Matériel :
Nous avons utilisé trois catégories principales de matériel : fongique, végétal et un substrat de
culture qui sont représenté sur le schéma suivant.
1-1- Matériel fongique :
L‘espèce fongique utilisée dans nos expérimentations est la Truffe noire du
Périgord Tuber melanosporumVitt. .
Les échantillons d‘ascocarpes de Tuber melanosporumVitt. desséchés nous ont été fournis par
M. Chevalier de l‘Institut National de Recherche Agronomique (INRA) de Clermont Ferrand
(France).
1-2- Matériel végétal:
Nous avons utilisé une espèce végétale le chêne vert (Quercus ilex L.) :
Les glands de chêne sont ramassés 2011 sous un chêne de la forêt de M‘sila , ils sont mis
directement en stratification pour éviter leur dégradation.
Matériel
*Vermiculite
*Tourbe blonde
*Perlite
Substrat
Chêne vert
Essence végétale
Tuber
melanosporum
Champignon

1-3-Substrat hors-sol :
Le substrat hors- sol utilisé est composé d‘un mélange de trois substrats différents :
* Vermiculite
* Perlite
* Tourbe blonde
- la vermiculite :
C‘est un substrat d‘origine volcanique, est un minerai, expansé en lamelles marrons a une
forme en accordéon, brillant, inerte et neutre (pH 7), stérile, léger , aérée , hydrophile. Il est
un substrat absorbant et sans odeur (LALIBERTE, 2006). Ce minerai est d‘abord trié, calibré,
et tamisé puis il est chauffé à haute température 800 à 1000°C, sous l‘effet de la chaleur les
lamelles s‘exfolient, il a d‘autres usages que l‘horticulture (BERKOWIEZ, 2009).
- la perlite :
Lave extrudée ou plus précisément une roche volcanique siliceuse, la perlite est un substrat
minerai, se présente sous forme de granulés blancs expansés en cellules fermées blanches à
paroi dure, inerte et neutre (pH 6.8), stérile, aérée et hydrophile, il a une granulométrie de 0.1
à 0.6mm (BERKOWIEZ, 2009). Ce minerai est obtenu par un broyage puis chauffage auteur
de 1200°C, le traitement thermique provoque une expansion qui donne des billes vitrifiées 10 à
15 fois plus grosses que leur volume initial (BERKOWIEZ, 2009).
- la tourbe blonde :
La tourbe est un composé organique utilisée pour la confection de mélange d‘une culture
hors-sol, elle améliore la porosité et la rétention en eau, et enrichit le sol en matières organique
(BOURGEON, 2008).
Le substrat hors- sol est ensuite additionné de deux autres composants :
-Les billes d‘argile à calibre moyen, utilisé pour activer le système, pour obtenir un support très aéré
permettant d‘oxygéner les racines.
Substrat hors-sol inerte forme un milieu fertile pour faciliter le
développement de la plante et la truffe (CHEVALIER, 1984)

-La craie sous forme de carbonate de calcium (Ca Co3) en poudre blanche ; elle est indispensable car
les truffes nécessitent un substrat riche en calcaire ou au moins riche en calcium pour assurer le
développement (FORTAS et CHEVALIER, 1992 ; CHEVALIER et FROCHOT, 1997)
2- Méthodes :
Les différentes méthodes d‘étude effectuées sont schématisées ci-dessous.
2-2- Etudes morphologiques des ascocarpes :
2-2-1- Examen macroscopique direct des ascocarpes
On effectue un examen à l‘œil nu de l‘ascocarpe comme indiqué sur le schéma suivant
pour connaître quelques caractéristiques du champignon:
Méthodes
Microscopique
Macroscopique
Etudes
Morphologiques
Loupe stéréoscopique
Mycorhization des plants
2. Milieu hors-sol
1. Pré-germination des glands
3. Inoculation des plants
Etude macroscopique de l‘ascocarpe
Poids
Diamètre
Port aérifère
Odeur
Couleur du péridium
péridium
Ascocarpes
T. melanosporum
et
Mycorhizes

2-2-2- Examen macroscopique sous la loupe stéréoscopique:
Les observations de l‘ascocarpe sous la loupe stéréoscopique permettent de connaître les
caractéristiques du péridium et de la glèbe :
- l‘examen du péridium permet de connaître :
son aspect
sa couleur
-l‘examen de la glèbe permet de distinguer :
le tissu stérile
et le tissu fertile
2-2- Etudes microscopiques de l‘ascocarpe :
Les examens microscopiques photoniques permettent d‘observer la morphologie du
mycélium externe, du tissu stérile et du tissu fertile contenant les organes de reproduction
(asques et ascospores.
2-2-1- Etudes microscopiques du péridium :
- Examens microscopiques de l‘aspect du péridium
Cet examen microscopique est effectué sur un fragment de la partie supérieure du
péridium , prélevé à l‘aide d‘un scalpel et déposé entre lame et lamelle dans une goutte
d‘eau distillée.
-Examen microscopique des houppes mycéliennes :
Des filaments mycéliens visibles sous la loupe stéréoscopique sont prélevés sur la surface
du péridium et examinés au microscope photonique entre lame et lamelle dans une
goutte d‘eau distillée.
Péridium
Glèbe

2-2-2- Etudes microscopiques de la glèbe :
La glèbe ou chair est composée de deux types de tissu : stérile et fertile.
- le tissu stérile :
Un fragment de gléba constitué de tissu stérile est prélevé sous la loupe stéréoscopique à
l‘aide d‘un scalpel et examiné au microscope photonique entre lame et lamelle dans une
goutte d‘eau distillée.
- le tissu fertile :
Un fragment de gléba est prélevé à l‘aide d‘un scalpel dans le tissu fertile contenant les
asques et les ascospores et examiné au microscope photonique entre lame et lamelle dans
une goutte d‘eau distillée. Pour observer les ornementations de l‘ascospore, on remplace
la goutte d‘eau distillée par quelques de bleu de coton à 0.1%
Les dimensions des asques et des ascospores sont mesurées à l‘aide d‘un micromètre
oculaire étalonné muni d‘une échelle micrométrique.
2-3- Réalisation des synthèses mycorhiziennes :
2-3-1- Désinfection des glands de chêne vert
La désinfection des glands peut être effectuée avant ou après la stratification, il est
préférable de les désinfecter après la stratification (BONITO et al., 2011).
Dès leur récolte, les glands de chêne peuvent être désinfectés soit avec l‘hypochlorite de
sodium à 10% (PEREZ et al., 2007) ou avec l‘eau oxygénée(BONITO et al., 2011).
Dans notre travail, nous avons désinfecté les glands avec l‘hypochlorite de sodium à 10%
pendant 20 min. Les glands sont ensuite rincés deux fois à trois fois dan l‘eau distillée.
2-3-2- La stratification et germination des glands de chêne vert
Selon certains auteurs , une prégermination des glands dans de l‘eau distillée froide
permet après 24 h de sélectionner les glands à fort pouvoir germinatif (se déposant au fond
du bac utilisé) de ceux qui ont un pouvoir germinatif très faible qui flottent à la surface de l‘eau
ou peuvent être contaminés (PEREZ et al., 2007). Cette méthode est appliquée dans les
pépinières de la conservation des forêts de la Wilaya d‘Oran, elle permet de placer les glands
directement dans les pots.

Dans notre travail, la désinfection des glands a été poursuivie par une stratification qui consiste
à les mettre dans un plateau rempli de sable (autoclavé 1h à 120°C), l‘ humidifié avec l‘eau
distillée stérile puis les placer au réfrigérateur à 4°C en les recouvrant avec un morceau de
tissu en coton . Le sable doit être humidifié de nouveau avec de l‘eau distillée stérile pour
éviter le desséchement du sable.
Cette technique permet le développement du bourgeon plus précocement et la durée de la
stratification est d‘environ 70 jours (MULLER et al., 1994 ; BONITO et al., 2011).
2-3-3- Préparation et désinfection du substrat hors-sol :
Le substrat utilisé est celui mis au point par l‘INRA de Clermont Ferrand (France) pour la
production des plants truffiers (CHEVALIER, 1985)
La méthode de préparation du substrat hors -sol et sa désinfection sont indiquées sur la figure
suivante :
2-3-4- Préparation de l‘inoculum
L‘inoculum est préparé selon la technique utilisée par l‘INRA de Clermont-Ferrand
pour l‘obtention des plants truffiers (CHEVALIER et GRENTE, 1979). Cette technique a été
testée d‘abord par les italiens de l‘INPL et de l‘université de Turin (FONTANA et al., 1972)
Vermiculite
Perlite
Tourbe blonde
Stérilisation au four Pasteur (GUIRAUD, 1998, PEREZ et al., 2007)
Substrat stérile
La craie
Fertilisant
Le mélange

puis appliquée en France, à l‘INRA de Clermont Ferrand par CHEVALIER et GRENTE en
1973 ; GIRAUD, 1998 ; CHEVALIER et FROCHOT, 1997). Elle est la plus utilisée dans le
monde pour la production de plants mycorhizés par la truffe noire.
L‘inoculum est sous forme d‘une suspension sporale, les étapes de préparation sont les
suivantes :
1- Ascocarpes hydratés : l‘hydratation pendant 24h dans du l‘eau distillée stérile, une
quantité 2g d‘ascocarpe par litre de substrat.
2- Broyage des ascocarpes hydratés : le broyage des ascocarpes hydratés avec un mixeur
électrique jusqu‘à l‘obtention d‘une solution sporale homogène.
2-3-5- Inoculation du substrat
L‘inoculum préparé précédemment est ensuite mélangé au substrat hors-sol qui
contient de la perlite, de la vermiculite et de la tourbe blonde (v/v/v), de la craie (Ca CO3) et
un fertilisant. Après une bonne homogénéisation de tout ce mélange, le substrat est ensuite
rempli dans des pots ouvert en matière plastique de 400ml comme indiqué dans le tableau
suivant :
Nombre
de pots
Témoin
(sans
inoculum)
Inoculé
Pour le
chêne vert
1 0 15

Les glands germés sont directement semés dans leurs pots respectifs.
2-3-6- Conditions de culture des plants :
Les plants en pots sont élevés dans une serre non climatisée ; ils sont arrosés avec de l‘eau
distillée et une solution minérale.
- Arrosage avec l‘eau distillée :
L‘arrosage des plantules avec l‘eau distillé s‘effectue par système de capillarité selon la
période de culture en serre qui est non climatisée. Par exemple en été, on arrose les plants
deux fois par semaine et en hiver une seule fois par semaine parfois tous les 15 jours.
L‘arrosage des plants cultivés en pots ouvert sur substrat hors-sol est compliqué car il dépend
de la capacité de rétention en eau des particules du substrat.
-Arrosage avec une solution minérale :
L‘arrosage avec une solution minérale est favorable pour la survie des plants, car les jeunes
plants mycorhizés sont souvent chétifs et ont une reprise difficile, donc la solution minérale
permet à la fois la croissance de l‘arbre et un développement suffisant du champignon surtout
dans le cas d‘une culture hydroponique (CHEVALIER et DESMAS, 1977).
La solution minérale nécessite la présence de l‘azote (N), du phosphate (P) , du potassium (K) ,
du calcium (Ca), du magnésium (Mg), du fer (Fr) et une gamme d‘oligoéléments ; sa
composition varie selon les espèces végétales mises en culture
2-4- Les méthodes d‘étude des associations mycorhiziennes
2-4-1- Etude des mycorhizes
Après la culture des plants, on prélève au hasard quelques plants inoculés de chêne pour
déceler sous la loupe stéréoscopique la présence des ectomycorhizes sur les racines courtes.
Après l‘installation des mycorhizes, on effectue les études suivantes :
- Etudes macroscopiques des ectomycorhizes :

Le plant est délicatement enlevé de son pot pour ne pas briser les racines courtes. Le
système racinaire est soigneusement lavé à l‘eau distillée afin d‘éliminer les particules du
substrat de culture qui adhèrent aux racines courtes puis examiné sous la loupe stéréoscopique
pour décrire la morphologie des ectomycorhizes (couleur, type d‘ectomycorhize, …..) formées
par Tuber melanosporum.
- Etudes microscopiques des ectomycorhizes:
Cette étude microscopique est effectuée sur des coupes transversales à main levée des
ectomycorhizes prélevées sous la loupe stéréoscopique. Les étapes d‘obtention de ces coupes
sont indiquées dans le schéma suivant
2-4-2 – Etude de la croissance des plants
2-4-2 -1- Estimation des paramètres de croissance des plants
La croissance des plants mycorhizés et des plants témoins a été estimée par différents
paramètres indiqués dans le tableau 12.

Tableau 12 : Paramètres de croissance estimés pour l‘espèce végétale étudiée.
2-4-2-2- Estimation du taux de mycorhization
L‘estimation de l‘infection du système racinaire des plants par le champignon
ectomycorhizien a été effectuée selon la méthode utilisée à l‘INRA de Clermont Ferrand
(France) pour contrôler les plants truffiers élevés en serre avant leur commercialisation
(DOMINGUEZ et al., 2005).
La mycorhization des plants est exprimée par :
A % : représente le nombre des racines contaminées.
T% : représente le nombre des racines infectées par Tuber melanosporum.
N% : représente le nombre des racines non mycorhizées.
2-4-2 -3 Interprétation statistique :
Les résultats de la mesure de la croissance des plants sont soumis à des analyses de
variance et l‘écartype effectué par l‘EXEL et des comparaisons de moyennes à l‘aide de la loi
de Student avec une estimation de t selon DELMAS (2010)
M = A% + T%+N%


1-Etude morphologique des ascocarpes de Tuber melanosporum
1-1 Examens macroscopiques des ascocarpes :
Cette espèce de Tuber a une odeur forte et agréable même à l‘état congelé ou desséché.
Cette caractéristique importante chez Tuber melanosporum, rend l‘espèce pionnière pour les
études aromatiques à partir des années 80 (CHEVALIER et GRENTE, 1980 ; CALLOT et al.,
1999 ; SPLIVALLO et al., 2007 ; DESSOLAS et al., 2008), cette forte odeur est due à la
présence de stérols (PACIONI et al., 1990 ; HARKI et al., 1996). C‘est l‘espèce la plus
parfumée par rapport aux autres espèces de Tuber et aux truffes de désert ou Terfez qui ont
une odeur plus faible (RAVOLANIRINA, 1986 ; WANG et MARCONE, 2011). Cette odeur
attire les chiens dressés lors du cavage effectué par les chercheurs de la truffe (CHEVALIER,
1985, CALLOT et al., 1999, MANCHABLON et PARGNEY, 2003).
L‘ascocarpe a une forme sub-globuleuse (Planche I, Fig. A) et porte une boursoufflure
bien visible (Planche I, Fig. B).
Selon certains auteurs cette petite déformation caractéristique chez le genre Tuber provient de
la composition et la qualité du sol truffier qui modifie la forme de l‘ascocarpe; il a aussi une
influence sur la qualité de l‘ascocarpe (PARGNEY,1994 ; DESSOLAS et al., 2008). Les
travaux de GRENTE et CHEVALIER (1980) montrent un ascocarpe de Tuber melanosporum
portant plein de boursoufflures ou bien lobé mais notre ascocarpe ne porte qu‘une seule
boursoufflure (Tableau 13).
La taille de l‘ascocarpe et leur poids donnent une indication sur la maturité de
l‘ascocarpe (Planche I, Fig. B).
Quant à la taille des ascocarpes, ils mesurent environ 3cm ; la dimension de ces ascocarpes
est inférieure à celle rapportée dans d‘autres travaux (TRAPPE et al. (1979), CHEVALIER et
GRENTE (1980), RIOUSSET et al. (2001) (Tableau 13). Le poids est d‘environ 6g ; ces
mesures ont été effectuées sur des ascocarpes conservés congelés. Cette dimension de
l‘ascocarpe se rapproche de celle rapportée par PARGNEY (1994) qui a montré que la
dimension de la truffe noire de Périgord varie de quelques millimètres à quelques centimètres
selon leur âge, il peut atteindre 114,7g (TRAPPE et al., 1979).
Le tableau 13 présente les caractéristiques des ascocarpes de T. melanosporum examinés
par rapport aux résultats d‘autres travaux sur cette même espèce de truffe.

Tableau 13 : Comparaison de nos résultats sur les paramètres de l‘ascocarpe de T.
melanosporum avec ceux de la littérature.
1-2 Examen du péridium de l‘ascocarpe sous la loupe stéréoscopique:
Le péridium a une couleur marron noir et il est verruqueux, il représente l‘écorce externe
ou l‘enveloppe protectrice de l‘ascocarpe (Planche I, Fig. B).
Nos observations se rapprochent de celles décrites par divers auteurs. En effet, le péridum de
Tuber melanosporum est de couleur noire d‘où le nom de truffe noire, (TRAPPE et al.
(1979), GRENTE et CHEVALIER, 1980). Cette couleur noire indique la maturité de
l‘ascocarpe (RIOUSSET et al., 2001) et sa concentration en mélanine joue un rôle dans le
murissement de la truffe (HARKI et al., 2006).
Le péridium a un aspect rugueux car il possède des écailles pyramidales étroites chez Tuber
melanosporum (Planche I, Figs. B et C).
Cette caractéristique du péridium différencie T. melanosporum de T. indicum dont
le péridium porte des écailles pyramidales larges et rigides (DESSOLAS et al., 2008 ;
BONITO et al., 2011) et de Tuber brumale à péridium portant des écailles lâches qui
tombent par un simple frottement au brossage (Tableau 14) (CHEVALIER et GRENTE,
1973 ; CALLOT et al., 1999 ; HALL et al., 2007).

Le péridium porte aussi des veines aérifères ou ports aérifères (3 ports) qui sont disposés à sa
surface (Planche I, Fig. D). Selon CALLOT et al. (1999), ces veines aérifères jouent un rôle
important dans la respiration du champignon.
Tableau 14 : Comparaison de nos résultats sur l‘aspect du péridium de T. melanosporum
avec ceux de la littérature.


Planche I
Morphologie et aspect du péridium
de l‘ascocarpe de la truffe noire
Fig. A : Morphologie générale d‘un ascocarpe de Tuber melanosporum.
Fig. B : Aspect macroscopique d‘un ascocarpe de Tuber melanosporum.
Fig. C : Aspect du péridium de Tuber melanosporum, observation sous la loupe
stéréoscopique.
Fig. D : Port aérifère de Tuber melanosporum observé sous la loupe
stéréoscopique (flèche).

1-3 Examens microscopiques de la glèbe des ascocarpes:
La glèbe ou chair est subdivisée en deux tissus dont l‘un est stérile et l‘autre est fertile. Nos
examens ont été effectués au microscope photonique sur un fragment de la glèbe (tissu
fertile) prélevé à partir d‘un ascocarpe de Tuber melanosporum et monté entre lame et lamelle
dans une goutte d‘eau distillée.
Les asques de Tuber melanosporum ont une forme sub-globuleuse (Planche II, Figs. A, B et
C), de couleur marron foncé mesurant environ (38– 70×55–120) µm. Ces dimensions des
asques se rapprochent de celle de PARGUEY-LEDUC et al. (1987) mais sont inférieures à
celles rapportées par TRAPPE et al. (1979) et par RIOUSSET et al. (2001) qui ont mentionné
respectivement des dimensions de l‘asque de (21-54×32-88) µm et (80-120×90-140) µm chez
Tuber melanosporum.
L‘asque renferme généralement 2 à 4 ascospores comme l‘a décrit RAVOLANIRINA (1986).
Il porte un pétiole allongé qui retient l‘asque au tissu fertile (CHEVALIER et GRENTE,
1980 ; PARGUEY-LEDUC, 1994 ; PARGNEY et MANCHABLON, 2003). L‘ensemble des
asques remplis d‘ascospores forme le tissu fertile, ce dernier accroché au tissu stérile forme la
glèbe recouverte par le péridium ce qui différencie les Ascomycètes des Basidiomycètes
(DELMAS, 1979 ; DANELL, 1997 ; HANSEN, 2006 ; HANSEN et LAESSOE, 2007).
Les ascospores sont de forme ovoïde (Planche II, Fig. A) et ellipsoïde (Planche II, Figs. B et
C).
Cette morphologique des ascospores de Tuber melanosporum a aussi été décrite par
BONITO et al. (2011). Selon CHEVALIER et GRENTE (1973), elle ressemble au ballon de
rugby.
Les ascospores Tuber melanosporum sont ornementées par des épines dressées, elles sont
droites à extrémités aigues (Planche II, Figs. B et C).
Cette ornementation des ascospores par des épines dressées est un critère important
d‘identification de Tuber melanosporum (TRAPPE et al. ,1979 ; CHEVALIER et
GRENTE,1980 ; CHEVALIER et FROCHOT, 1997). Elle différencie les ascospores de
Tuber melanosporum de celles d‘autres espèces de Tuber en particulier celles de Tuber
indicum dont les ascospores portent des épines recourbées (BONITO et al. , 2011).

Les ascospores que nous avons examinées au microscope photonique sont de couleur marron
foncé (Planche II, Figs. B et C).
La couleur des ascospores est également une caractéristique qui différencie T. melanosporum
dont les ascospores matures sont de couleur marron foncé et immatures marron clair tandis
que celles de Tuber indicum matures sont marron clair (BONITO et al., 2011). De
nombreux travaux sur la composition chimique des Tuber rapportent que la mélanine est le
composant principal chez Tuber melanosporum, elle représente 15% dans un ascocarpe
mature (HARKI et al. , 2006) ce qui explique la couleur des ascospores de T. melanosporum
par rapport à celle des autres espèces de truffe (GRENTE et DELMAS, 1974 ; PARGUEY-
LEDUC, 1994 ; WANG et MARCONE, 2011) .
Les ascospores de nos échantillons d‘ascocarpes mesurent (18-25×30-36) µm. Ces dimensions
se rapprochent de celles rapportées par CHEVALIER et GRENTE (1980), MORENO et al.
(1986), PETHITORG et al. (1995) et BONITO et al. (2011) qui ont mentionné des
dimensions des ascospores de T. melanosporum comprises entre (15-28×17-32) µm et (20-
25×30-35) µm.


Planche II
Morphologie de l‘asque et de l‘ascospore
de la truffe noire
Fig. A : Fragment du tissu fertile de la gléba montrant les asques renfermant 3 à 4
ascospores (observation microscopique à Gr x 100).
Fig. B : Morphologie d‘un asque contenant quatre ascospores ellipsoïdes
(observation microscopique à Gr x 400).
Fig. C : Morphologie d‘un asque contenant deux ascospores ornementées par des
épines dressées (observation microscopique à Gr x 400).

2- Etude de la mycorhization des plants de chêne vert :
2-1- Effet de la mycorhization sur la croissance des plants de chêne vert
Pour évaluer la croissance des plants de chêne vert inoculés avec Tuber melanosporum
Vitt. (Truffe noire du Périgord) sur substrat hors sol et âgés d‘une année, nous avons mesuré les
paramètres suivants : la hauteur de la tige, le nombre des feuilles, la surface des feuilles ainsi
que les poids frais et sec de la partie aérienne.
L‘analyse statistique des résultats (Tableau 15) montre que l‘inoculation des plants de chêne
vert par Tuber melanosporum améliore significativement la croissance des parties aériennes
des plants par rapport aux témoins (Planche III, Figs. A et C ; Planche VI, Fig. A).
Ces résultats se rapprochent de ceux de CHEVALIER (1985) qui a montré que le
champignon truffier améliore la croissance des arbres. Cette augmentation de la hauteur de la
partie aérienne des plants de chêne vert inoculés est provoquée par la présence du
champignon mycorhizien associé à la plante hôte (STRULLU, 1991).
La croissance des arbres permet aussi la production de bois pour les sylvicultures
(TAGLIAFERRO, 1996 ; GUINBERTEAU et COURTECUISSE, 1996 ; MORENO-
ARROYO et al., 2001 ; THERVILLE, 2009).
Tableau 15 : L‘évaluation statistique des paramètres de croissance des plants de chêne vert..
N.S : non significatif
S : significatif

Il ya peu de différence significative entre le nombre de feuilles et la surface foliaire des plants
de chêne vert inoculés et ceux des témoins (Tableau 19 ; Planche I V, Fig. B et C). Nous
avons constaté que certaines feuilles des plants inoculés ont une surface foliaire plus
importante que chez les témoins (Planche III, Fig. B).
De nombreux travaux ont pourtant montré que le champignon mycorhizien améliore
des processus physiologiques chez la plante hôte. Il fournit à l‘arbre des substances de
croissance et des vitamines (LE TACON et GARBAYE, 1986), des éléments minéraux qu‘il est
seul capable de prélever dans les particules de substrat en quantité suffisante, ces éléments
minéraux sont transférés à la plante sans modification ou peuvent avoir une transformation au
cours du métabolisme fongique (CHEVALIER et GRENTE, 1980 ; GARBAYE et LE
TACON ,1986 ; FROCHOT et CHEVALIER, 1997 ; PLASSARD et al., 1997). Selon
MOUSSAIN et al. (1994), il y a une relation entre la mycorhization et la croissance ou le
développement de la plante ; les échanges gazeux de la plante mycorhizée sont plus
développés par rapport aux plants non mycorhizés car il existe une forte assimilation de CO2
provoquée par la mycorhization et la plante à son tour fournit au champignon des substances
carbonées simples comme les sucres qui sont nécessaires à son métabolisme de base (LE
TACON et GARBAYE, 1986), et des substances plus complexes pour sa fructification, pour le
développement de l‘ascocarpe et la structure de l‘ascospore (LE TACON, 1996 ; HARKI et
al., 2006 ; WANG et MARCONE, 2011).
Quant à la biomasse fraiche de la partie aérienne des plants de chêne vert inoculés par
T. melanosporum, elle est significativement améliorée par contre, il n‘y a aucune différence
significative de la biomasse sèche par rapport aux témoins (Tableau 19; Planche IV, Figs. D et
E).
Nos résultats montrent que les plants inoculés sont plus riches en eau à l‘état frais que les
témoins. En effet, le champignon mycorhizien joue un rôle important dans l‘absorption
d‘eau comme l‘ont montré LAMINOU MANZO et al. (2009). D‘autres travaux rapportent
que la mycorhization intervient dans la croissance de la plante ainsi que la biomasse fraîche de
la plante car elle limite les pertes d‘eau par transpiration et intervient dans la régulation
stomatique (LE TACON et GARBAYE, 1986 ; SELOSSE, 2000 ; BA et al., 2011).


Planche III
Croissance des plants de chêne vert inoculés par Tuber melanosporum,
après 12 mois de culture hors sol en serre.
Fig. A : Effet de la mycorhization sur la croissance aérienne d‘un plant de chêne
vert inoculé avec Tuber melanosporum par rapport au plant témoin.
Fig. B : Effet de la mycorhization sur l‘importance de la surface foliaire d‘un
plant de chêne vert inoculé avec Tuber melanosporum par rapport à celle du
plant témoin.
Fig. C : Effet de la mycorhization sur le développement du système racinaire d‘un
plant de chêne vert inoculé avec Tuber melanosporum. Remarquer la formation
du chevelu racinaire par rapport au plant témoin.
Fig. D : Morphologie du chevelu racinaire du plant de chêne vert inoculé avec
Tuber melanosporum par rapport au système racinaire du plant témoin.

Planche IV
Effet de l‘inoculation de Tuber melanosporum sur la croissance
aérienne des plants de chêne vert.

2-2 Effet de la mycorhization sur la croissance racinaire des plants de chêne
Nous avons estimé le développement du système racinaire par la mesure de la longueur
de la racine principale et secondaire, le nombre des racines latérales ainsi que le poids frais de
tout le système racinaire.
Après 12 mois de culture, les systèmes racinaires des plants inoculés par T. melanosporum
présentent un système très développé et dense par la présence de nombreuses racines courtes
formant un chevelu racinaire (Planche III, Figs. C et D).
L‘analyse statistique des résultats montrent des différences significatives entre les systèmes
racinaires des plants inoculés et ceux des témoins (Tableau 16 ; Planche IV, Figs. A à D ;
Planche V).
Le poids frais du système racinaire des plants inoculés est significativement plus important
que celui des témoins (Planche V, Fig. B). Cette différence de poids est provoquée par
l‘association mycorhizienne qui colonise la racine latérale dans le but de former le manteau
ainsi que le réseau mycélien (LE TACON, 1996). Le nombre des racines courtes bien
développées par rapport aux témoins indique qu‗il y a une relation entre la biomasse, le
nombre des racines latérales et l‘infection mycélienne.
Nos résultats rejoignent ceux de certains travaux qui ont montré que le système
racinaire des plants de noisetier mycorhizés par Tuber melanosporum forme un chevelu
racinaire ou bouquet racinaire (CHEVALIER et GRENTE ,1973) et dans l‘association entre
Tuber melanosporum et le chêne, il forme un « chignon » (GRENTE et CHEVALIER
1980).
Selon certains auteurs, ce développement racinaire chez les plants mycorhizés est stimulé par
des auxines spéciales qui s‘accumulent autour de la racine produisant ainsi de nombreuses
racines latérales (ZAFFRAN, 1998 ; DUPRE, 2005 ; FELTEN et al., 2009 ; BONFANTE et
GENRE, 2010 ; MARTIN et al., 2010). La croissance mycélienne de la truffe noire induit chez
la plante la sécrétion d‘éthylène qui provoque aussi la formation des racines latérales
(FELTEN et al., 2009 ; KUES et MARTIN, 2011).
Selon DRENOU (2006), les racines longues et courtes du système racinaire des feuilleux se
développent, se multiplient et changent morphologiquement dans le cas de la mycorhize.

Le développement important du système racinaire indique comme l‘ont signalé BONFANTE
et GENRE (2010), que la mycorhization a un effet sur l‘alimentation minérale de l‘arbre qui est
améliorée par la présence du mycélium, ce dernier intervient dans l‘absorption de certains
minéraux.
Tableau 16 : L‘évaluation statistique des paramètres de développement de la partie racinaire
des plants de chêne vert.
N.S : non significatif
S : significatif

Planche V
Effet de l‘inoculation de Tuber melanosporum sur le développement
racinaire des plants de chêne vert.

2-3 L‘étude morphologique des mycorhizes :
Le début de l‘infection mycélienne des racines de chêne vert inoculées avec le Tuber
melanosporum s‘observe par la présence de filaments mycéliens enveloppant la racine
(Planche VI, Fig. A). Les racines courtes mycorhizées se présentent en massue de couleur
marron foncé (Planche VI, Fig. C) par rapport aux non mycorhizées (Planche VI, Fig. B).
Ces résultats se rapprochent de ceux de CHEVALIER et GRENTE (1973) qui ont
montré que les mycorhizes de Tuber melanosporum forment généralement des
ectomycorhizes simples en forme de petites massues de 1 mm de long de couleur marron
situées à l‘extrémité des racines latérales de l‘arbre. L‘association de Tuber melanosporum au
chêne vert forme aussi une mycorhize simple en forme de petite massue, arrondie au
sommet, de couleur brune avec une surface lisse (DOMINGUEZ et al. ,2005 ; PEREZ et al.
,2007). Cette mycorhize est la zone des échanges entre les deux partenaires de la symbiose
(DRENOU et al., 2006 ; PEREZ et al., 2007).
Nous avons observé des glomérules sur les racines inoculées de chêne vert avec le Tuber
melanosporum (Planche VI, Fig. D). Ces structures ont aussi été décrites dans les mycorhizes
du chêne vert formées par Tuber melanosporum où les radicelles porteuses de mycorhizes se
ramifient à sa base et les ramifications se transforment en mycorhizes et donnent en même
temps l‘aspect d‘un petit bouquet nommé les glomérules (GRENTE et CHEVALIER, 1973 ;
GRENTE et CHEVALIER, 1980).
La présence de ces structures a été signalée dans le chignon racinaire des plants truffiers, elles
apparaissent lors d‘un développement intense de la racine, c‘est le résultat d‘un signal
chimique émis par le champignon aux racines latérales entrainant la croissance des racines
courtes (KUES et MARTIN, 2011).


Planche VI
Morphologie des racines de plants de chêne vert inoculés par Tuber
melanosporum observées sous la loupe binoculaire.
Fig. A : Fragment d‘une racine de chêne vert recouvert par des hyphes
du champignon inoculé à la plante.
Fig. B : Racines courtes des plants de chêne vert non mycorhizées
(flèche : les poils absorbants sont recouverts de débris de perlite).
Fig. C : Racines courtes des plants de chêne vert mycorhizés en forme
de massue.
Fig. D : Glomérule formé sur le système racinaire d‘un plant de chêne
vert inoculé avec le Tuber melanosporum.

2-4 L‘étude microscopique de l‘association mycorhizienne :
Après l‘infection mycélienne des racines de chêne vert inoculées avec le Tuber
melanosporum et la formation de filaments mycéliens enveloppant la racine (Planche VI,
Figs. A et F) pour former le manteau fongique. La progression de l‘infection s‘observe par
la présence des hyphes septés parcourant les espaces intercellulaires des cellules corticales
pour le développement du réseau d‘Hartig (Planche VII, Figs. C et D). Il n‘y a aucune
infection dans les racines des plants témoins (Planche VII, Figs. E et G).Toutes ces
observations indiquent qu‘il s‘agit d‘une ectomycorhize.
Ces résultats se rapprochent de ceux de certains auteurs qui ont montré que les espèces
ectomycorhiziennes forment au début le manteau en parallèle avec le développement du
réseau d‘Hartig (MELIN et NILSON, 1957). Le manteau est formé par le même type de
filament mycélien « haploïde » (RUBINI et al., 2011) et ces hyphes s‘insinuent entre les
cellules épidermique (BONFANTE et GENRE, 2010). La formation du manteau repose sur
les composés produits par la photosynthèse de la plante hôte comme produit préliminaire
« sucre » car le champignon est un espèce hétérotrophe (PERIN et al., 1991 ; KUES et
MARTIN, 2011).
La formation des ectomycorhizes débute par une prolifération des cellules fongiques entre les
cellules corticales dans le but de former le réseau d‘Hartig comme premier contact
(STRULLU, 1991), ce site de pénétration est réalisé par des enzymes fongiques qui dégradent
la paroi cellulaire (LE TACON et al. ,2011). Le séquençage du génome de Tuber
melanosporum a révélé des séquences codant pour les enzymes pariétales (cellulase, hémi-
cellulase et autre) (MARTIN et al. ,2010).
Le taux de mycorhization de nos plants inoculés par Tuber melanosporum, est de 10
%. Ce pourcentage est inférieure à celui d‘un plant truffier commercialisable qui doit avoir un
taux de mycorhization de 15% donc les plants que nous avons obtenus ne peuvent pas être
plantés.



Planche VII
Association mycorhizienne entre Chêne vert et Tuber melanosporum,
après 12 mois de culture hors sol en serre.
Fig. A : Fragment de racine de chêne vert mycorhizé par Tuber
melanosporum recouvert de mycélium (Gr x 400).
Fig. B : Surface externe d‘un fragment racinaire de plant chêne vert
mycorhizé envahie par des filaments fongiques. (Gr x 400).
Fig. C : Cellules racinaires de plant chêne vert mycorhizé avec des
hyphes qui s‘insinuent entre les cellules corticales. (Gr x 400).
Fig. D : Cellules corticales de plant chêne vert mycorhizé montrant des
hyphes cloisonnés intercellulaires. (Gr x 400).
Fig. E : Racine témoin de chêne vert. (Gr x 400).
Fig. F : Coupe transversale à main levée d‘une racine d‘un plant de
chêne vert inoculés par Tuber melanosporum (Gr x 100).
Fig. G : Coupe transversales à main levée d‘une racine d‘un plant de
chêne témoin (Gr x 100).

Conclusion
Notre travail porte sur les caractéristiques morphologiques de la truffe noire Tuber
melanosporum Vitt. et son association mycorhizienne avec une essence forestière Quercus
ilex en culture hors sol, en conditions contrôlées.
L‘étude des caractéristiques morphologiques a permis de décrire l‘ascocarpe de Tuber
melanosporum Vitt. qui est de couleur noir marron, sa forme est subglobuleuse, il est
recouvert par le péridium, couche externe rugueuse recouverte par des écailles pyramidales
larges et portant des port aérifères ; La glèbe ou chair contient un tissu fertile regroupant des
asques globuleux munis d‘un pétiole qui les relient au tissu stérile, ils renferment 2 à 4
ascospores de forme ellipsoïde marron ornementées par des épines dressées, elles sont droites
à extrémités aigues. Toutes ces caractéristiques sont des critères importants d‘identification de
Tuber melanosporum Vitt.
L‘inoculation des plants de chêne vert avec Tuber melanosporum Vitt., en culture hors
sol a permis d‘évaluer après 12 mois de culture en serre la croissance des plants de chêne
inoculés estimée par la hauteur de la tige, le nombre des feuilles, la surface des feuilles ainsi
que les poids frais et sec de la partie aérienne.
L‘analyse statistique des résultats montre que l‘inoculation des plants de chêne vert par
Tuber melanosporum améliore significativement la croissance des parties aériennes des
plants par rapport aux témoins. Il ya peu de différence significative entre le nombre de
feuilles et la surface foliaire des plants de chêne vert inoculés et ceux des témoins. Nous avons
constaté que certaines feuilles des plants inoculés ont une surface foliaire plus importante
que chez les témoins.
Quant à la biomasse fraiche de la partie aérienne des plants de chêne vert inoculés par T.
melanosporum, elle est significativement améliorée par contre, il n‘y a aucune différence
significative de la biomasse sèche par rapport aux témoins.
L‘effet de la mycorhization sur le développement racinaire des plants de chêne a été
estimé par la mesure de la longueur de la racine principale et secondaire, le nombre des racines
latérales ainsi que le poids frais de tout le système racinaire.
Après 12 mois de culture, les systèmes racinaires des plants inoculés par T. melanosporum
présentent un système très développé et dense par la présence de nombreuses racines courtes
formant un chevelu racinaire. L‘analyse statistique des résultats montrent des différences
significatives entre les systèmes racinaires des plants inoculés et ceux des témoins.

Le poids frais du système racinaire des plants inoculés est significativement plus important
que celui des témoins
L‘étude morphologique des mycorhizes révèle que l‘infection mycélienne des racines
des plants inoculés débute par le développement des filaments mycéliens qui enveloppent les
racines latérales. Les racines courtes mycorhizées se présentent en massue de couleur marron
foncé. Des glomérules se forment sur les racines inoculées de chêne vert avec le Tuber
melanosporum ; ces structures ont aussi été décrites dans les mycorhizes du chêne vert
formées par Tuber melanosporum.
Les examens microscopiques de l‘association mycorhizienne ont permis d‘observer la
présence de filaments mycéliens enveloppant la racine pour former le manteau fongique. La
progression de l‘infection s‘observe par la présence des hyphes septés parcourant les espaces
intercellulaires des cellules corticales pour le développement du réseau d‘Hartig Il n‘y a
aucune infection dans les racines des plants témoins.
Toutes ces observations indiquent qu‘il s‘agit d‘une ectomycorhize.
Le taux de mycorhization de nos plants inoculés par Tuber melanosporum, est de 10
%. Ce pourcentage est inférieure à celui d‘un plant truffier commercialisable qui doit avoir un
taux de mycorhization de 15% donc les plants que nous avons obtenus ne peuvent pas être
plantés.
En perspectives, ce travail mérite d‘être poursuivi afin de mieux maitriser la technique
de production des plants de chêne ou d‘autres essences forestières mycorhizés par Tuber
melanosporum en culture hors sol et d‘obtenir un taux de mycorhization de 15% pour
envisager des essais de leur transplantation sur un terrain favorable à leur développement.

Références bibliographiques
AGERER R., 2001. Exploration types of ectomycorhizas, proposal classify ectomycorrhizal
mycelia systems according to their patterns of differentiation and putative ecological importance.
Mycorrhiza, 11: 107-114.
AGERER R., 2006. Fungal relationships and structural identity of their ectomycorrhizae. Mycol.
Progress, 5: 67-107.
BA A., DUPONNOIS R., DIABATE M., DREYFUS B., 2011. Champignon mycorhizien des
arbres forestiers en Afrique de l‘ouest. Ed. IRD, Maroc, 268p.
BARBERO M., LOISEL R., 1980. La recherche verte en région méditerranéenne. R.F.F.,
XXXIII (6) : 531-543.
BARCHFIELD J., 2008. Europe‘s truffle shortage blamed on global warming. USA Today, p.2.
BENAC D., 2006. Détail fruit et légumes, CTIFL, 236 : 4p.
BELLAKHDAR J., 2003. Le Maghreb à travers ses plantes. Ed. LE FENAC, pp. 47-57.
BERTAULT G., RAYMOND M., BERTHOMIEU A., CALOT C., FERNANDEZ D., 1998.
Trilling variation in truffles. Nature, 394: 7-34.
BONFANTE P., 1973. Nuclear division in the vegetative hyphae of Tuber species plurimae.
Mycopathologia et Mycologia Applicata, 49 (3): 161-167.
BONFANTE P., GENRE A., 2010. Mechanisms underlying beneficial plant-fungus interactions
in mycorrhizal symbiosis. Nature Communication, 48 (1): 1-11.
BONITO G., TRAPPE J.M., RAWLINSON P., VILGALYS R., 2010. Improved
resolution of major clades within Tuber and taxonomy of species within the Tuber
gibbosum complex. Mycologia, 125 (5): 1042-1057.
BONUSO E., 2008. Metodi moleculari nello studio della variabillita. Genetica di Tuber brochii.
These doctorat. Alma Mater Studiorum Université Bolognia, pp. 4 – 8.
BOULANGER E., 1899. In : DESSOLAS H., PARGNEY J-CL., CHEVALIER G., 2008.
Nouveau manuel de trufficulture. Réédition du manuel du Docteur Pradel (1914).Ed. Dessolas,
Paris, (France), 308p.
BRUNEL A., FASOLO BONFANTE P., 1973. Particularité de mycélium de Tuber
melanosporum. Ann. Phytopath., 5(3) : 317-318.
CAIRNEY J.W.G., 2012. Extramatricial mycelia of ectomycorhizal fungi as moderators of carbon
dynaics in forest soil. Soil biology and biochemistry, 47: 198-208.
CALLOT G., BYE P., RAYMOND M., FERNANDEZ D., PARGNEY J-CL., PARGUEY-
LEDUC A., JANEX-FAVRE M-C., MOUSSA R., PAGES L., 1999. La truffe, la terre, la vie. Ed.
INRA., Paris, (France), 209p.

CASERMEIRO M.A., GARCIA-MONTERO L.G., MANJON J.L., HERNANO I., 2007.
Impact of active soil carbonate and burn size on the capacity of the rockrose Cistus laurifolius to
produce Tuber melanosporum carpophores in truffle culture. Mycological Reasearch, 111: 734-
739.
CHATIN A., 1892. La truffe. Lib. J.B. Ballière et fils, Paris, France, 372p.
CHETIN S.C., MORAR G., 2011. Economically and alimentary important truffle species,
establishment and developpement of the truffle culture in Xiromero region. Agricultura, 3: 79-80.
CHEVALIER G., 1972. Obtention de culture de mycélium de truffe a partir du carpophore et des
mycorhizes. Académie d‘agriculture de France, 53(185) : 981-989.
CHEVALIER G., POLLACSEK A., 1973. Synthèse axénique des mycorhizés de Tuber brumale
Vitt. a partir de culture pures du champignon. Ann. Phytopat., 5 (2) : 163-182.
CHEVALIER G., GRENTE J., 1978. Application pratiques de la symbiose ectomycorhizienne :
production a grand échelle de plants mycorhizé par la truffe Tuber melanosporum Vitt.
Mushrooms Science, 10 : 483-505.
CHEVALIER G. GRENTE J., 1980. La trufficulture un or noir pour la région méditerranéenne.
Forêt Méditerranéenne, 1 (2) : 151–162.
CHEVALIER G., 1985. La mycorhization contrôlée en pépinières forestières possibilité
d‘application aux conteneurs. R.F.F. XXXVII (2): 93-106.
CHEVALIER G., 1996. Le chêne vert et le chêne blanc, essence truffière par excellence. Forêt
méditerranéenne, 17 (3): 235-242.
CHEVALIER G., FROCHOT H., 1997. La maitrise de la culture de la truffe. Rev. For. Fr.,
XLIX n°sp : 201-213.
CHEVALIER G., DESSOLAS H., PARGNEY J-CL., 2008. Nouveau manuel de trufficulture.
Réédition du manuel du Docteur Pradel (1914). Ed. Dessolas, Paris, (France), 308p.
CORTES G., BARROETAVENA C., RAJCHENBERG M., 2011. Argentina the new frontiers
for truffles. In le VI th international workshop on edible mycorhizal mushrooms (IWEMM6).
Rabat, Maroc, 6-10 avril 2011, p. 31.
DANELL E., 1997. Les progrès dans la maitrise de la culture de chanterelles,
Chantharellus cibarius. Rev. For. Fr., XLIX (n° sp.) : 214-21.
DEHMAI-MEGREROUCH M., 2002. Typologie et dynamique des chênaies vertes en Algérie.
Foret Méditerranéenne, XXIII (2) : 117-133.
DEXHEIMER J., 1997. Étude structurale et fonctionnelle des interfaces entre le champignon et la
plante hôte. Rev. For. Fr. XLIX-n°sp. : 43-56.
DIAZ G., HONRUBIA M., GARCIA G., GUTIERREZ A., 1996. Caractérisation des
mycorhizes dans des bois de pin d‘Alep sur le Sistema Ibérico (Espagne), résultats préliminaires.
CIHEAM-Option Méditerranéennes, pp. 43-50.

DIAZ P., IBANEZ E., SENORANS F.J., REGLO G., 2003. Truffle aroma characterization by
headspace solid-phase micro extraction. Journal of Chromatography, 1017: 207-214.
DELMAS J., POITOU N., 1978. La mycorhization de Quercus pubescent par Tuber
melanosporum en condition contrôlées. Mushroom Science, 1 : 995-1005.
DELMAS J., 1979. Les truffes. Revues pour la science, 26 : 69-80.
DESSOLAS H., PARGNEY J-CL., CHEVALIER G., 2008. Nouveau manuel de
trufficulture. Réédition du manuel du Docteur Pradel (1914).Ed. Dessolas, Paris, 308p.
DOMINGUEZ J.A., RODRIGUEZ J.A., SAIZ D‘OMENACA J.A., 2005.
Ectomycorrizas en dos plantaciones trufers de encina Quercus ilex L. ballota Desf. Samp.
En Castellano. Bol. San. Veg. Plagas, 31 : 147-157.
DRENOU CH., 2006. Les racines, face cachée des arbres. Institut pour le développement
forestier service d‘utilité forestière CNPPF, Paris, 335p.
DUMORT E., 1822. In: HANSEN K., LAESSOE TH., 2007. Truffe trouble: what happened
to the Tubérales? Mycological Research, 111: 1075 – 1099.
DUHOUX E., NICOLE M., 2004. Biologie végétale. Association et interaction chez les plantes.
Ed. Dunod, Paris, 166p.
FELTEN J., KOHLER A., MORIN E., BHALERAO RP., PALMEK., MARTIN F.,
DITENGON F.A, LEGUE V., 2009. The ectomycorrhizal fungius Laccaria bicolor
stimulateslaterales root formation in poplar and Arabidopsis through auxin transport and
signaling. Plant Physiol., 151: 1991-2005.
FERKA ZAZOU N., 2006. Impact de l‘occupation spacio- temporelle des espèces sur la
conservation de l‘écosystème forestier. Cas de la commune de Tessala, wilaya de Sidi Bel abbes,
Algérie. Mém. Magister Univ. De Tlemcen, 164p.
FONTANA A., 1968. In. PALENZONA M., CHEVALIER G., FONTANA A., 1972. Sintesi
micorrizica tra i mcel in coltura di Tuber brumale, Tuber melanosporum, Tuber rufum e
semenzali di conifere e latifoglie. Bolletino dell istituto ed orto botanico dell universita di Torino,
18: 41-52.
FONTANA A., PALENZONA M., CHEVALIER G., 1972. Sintesi micorrizica trai mcel in
coltura di Tuber brumale, Tuber melanosporum, Tuber rufum e semenzali di conifere e latifoglie.
Bolletino dell istituto ed orto botanico dell universita di Torino, 18: 41-52.
FONTANA A., GIOVANNETTI G., 1979. Simbiosis micorrizica tra cistus incanus L. ssp ;
incanus e Tuber melanosporum Vitt. Allionia, 23 : 5-11.
FORTAS Z., CHEVALIER G., 1992. Effet des conditions de culture sur la mycorhization de
l‘Helianthemum guttatum par trois espèces de terfez des genres Terfezia et Tirmania d‘Algérie.
Can. J. Bot., 70 : 2453-2460.

GARCIA-MONTERO L.G., MANJON J.L., PASQUAL C., GARCIA-ABRIL A., 2007.
Ecological patterns of Tuber melanosporum and different Quercus Mediterranean forests:
Quantitative production of truffles, burn sizes and soil studies. Forest Ecology and Management,
242: 288-296.
GARCIA-MONTERRO L.G., VELVERDE-ASENJO I., DIAZ P., PASCUAL C., 2008.
Statistical patterns of carbonates and total organic carbon on soils of Tuber melanosporum and
Tuber rufum brulis. Soil Research, 47 (2): 206-212.
GARCIA-MONTERO L.G., QUINTANA A., VALVERDE-ASENJO I., DIAZ P., 2009.
Calcareous amendments in truffle culture: Asoil nutrition hypothesis. Soil Biology et Biochmistry,
41: 1227-1232.
GARCIA-BAREDA S., REYNA S., 2013. Response of Tuber melanosporum fruiting to
canopy opening in a Pinus-Quercus forest. Ecological Engineering, 53: 54-60.
GRENTE J., CHEVALIER G., 1973. Recherche en vue d‘une trufficulture rationnelle, Aspects
mycologiques. B.T.I., 283 : 733-746.
GRENTE J., DELMAS J., 1974. Perspectives pour une trufficulture moderne. Clermont-
Ferrand. INRA. France, 65p.
GUERIN-LAGUETTE A., CUMINGS N., YUN W., 2011. Overview of research on edible
mycorrhizal fungi in New Zealand. In le VI th international workshop on edible mycorhizal
mushrooms (IWEMM6). Rabat, Maroc, 6-10 avril 2011, p. 55.
GUINBERTEAU J., COURTECUISSE R., 1997. Diversité des champignons (surtout
mycorhiziens) dans les écosystèmes forestiers actuels. Rev. For. Fr., XLIX-n°sp : 25-39.
GUIRAUD J.P., 1998. Microbiologie alimentaire. Ed. DUNOD, Paris, pp. 171-193.
HAICHOUR R., 2009. Stress thermique et limite écologique du chêne vert en Algérie. Univ. de
Mentouri. Constantine, 139p.
HALL I.R., BROWN G.T., ZAMBONELLI A., 2007. Taming the truffle. Portland,
Oregon, USA, 304p.
HANSEN K., 2006. Ascomycota truffles: cup fungi go underground. Friends of the
Farlow, 47: 1-12.
HANSEN K., LAESSOE TH., 2007. Truffle trouble: what happened to the Tubérales?
Mycological Research, 111: 1075 – 1099.
HARKI F., KLAEBE A., TALOU T., DARGENT R., 1996. Identification and
quantification of Tuber melanosporum Vitt. Sterols. Steroids, 61: 609-612
HARKI F., BOUYA d., DARGENT R., 2006. Maturation-associated alternations of the
biochemical characteristics of the black truffle Tuber melanosporum Vitt. Food Chemistry, 99:
394-400.

JEANDROZ S., MURATC., WANG Y., BONFANTE P., LE TACON F., Molecular phylogeny
and historical biogeography of the genus Tuber, the truetruffles. Journal of Biogeography, p.1365.
KORF R.P., 1973. Discomycetes and Tuberales. In: RIOUSSET G., RIOUSSET L.,
CHEVALIER G., BRADET M.C., 2001 : Truffe d‘Europe et de chine. Ed. INRA. France, 179p.
KUES U., MARTIN F., 2011. On the road to understanding truffles in the underground. Fungal
genetic and biology, 48: 555-560.
KREMER F., 2005. les arbres. Ed. Lavoisier. France, 230p.
LAMINOU MANZO O., IBRAHIM D., CAMPANELLA B., PAUL R., 2009. Effets de
l'inoculation mycorhizienne du substrat sur la croissance et la résistance au stress hydrique
de cinq espèces fixatrices de dunes : Acacia raddiana Savi ; Acacia nilotica (L.) Willd. Ex
Del. var. adansonii ; Acacia senegal (L.) Willd ; Prosopis chilensis Stunz. et Bauhinia
rufescens Lam. Geo-Eco-Trop., 2009, 33: 115 – 124.
LAURENTE P., 2003. Regard sur les champignons, 52p.
LA LIBERTIE M., 2006. Mauvaise surprise protégé vous. pp. 33-35.
LAQBAQBI A., CHEVALIER G., BERLUREAU D., 2011. Modélisation de la trufficulture au
Royaume du Maroc. In le VI th international workshop on edible mycorhizal mushrooms
(IWEMM6). Rabat, Maroc, 6-10 avril 2011, p. 63.
LE TACON F., GARBAYE J., 1986. La maitrise des associations mycorhiziennes en pépinières
forestière. R.F.F., XXXVIII (3) : 249-259.
LE TACON F., 1996. Conclusions vers une meilleure prise en compte des champignons
mycorhiziens dans la gestion forestière. Rev. For. Fr., XLIX- n°sp : 245-255
LE TACON F., WANG Y., TAN Z.M., MURAT C., JEANDROZS., 2006. Molecular
taxonomie of chinese belongoing to the Tuber rufum and Tuber puberlum groups. Fungal
diversity, VIX: 301-328.
MACONNELL J.E., ESSELEN W.B., 1947. Carbohydrate in cultivated mushrooms. Journal of
Food Research, 12: 118-121.
MABRU D., DOUET J.P., MOUTON A., DUPRE C., RICARD J.M., MEDINA B.,
CASTROVIEJO M., CHEVALIER G., 2004. PCR-RFLP using a SNP on the mitochondrial Lsu-
rDNA as an easy method to differentiate Tuber melanosporum (Périgord truffle) and other truffle
species in cans. International Journal of Food Microbiology, 94: 33-42.
MARTIN F., KOHLER A., MURAT C., BALESTRINI R., COUTINHO P.M., MONTANINI
B., MORIN E., NOEL B., PERCUDANI R., PORCEL B., RUBINI A., AMICUCCI A.,
AMSELEM J., ANTHOUARD V., ARCIONI S., ARTIGUENAVE F., AURY J.M.,
BALLARIO P., BOLCHI A., BRENNA A., BRUN A., BUEE M., CANTAREL B.,
CHEVALIER G., COULOUX A., DA SILVA C., DENOEUD F., DUPLESSIS S.,

GHIGNONE S., HILSELBERGER B., IOTTI M., MARÇAIS B., MELLO A., MIRANDA M.,
PACIONI G., QUESNEVILLE H., RICCIONI C., RUOTOLO R., SPLIVALLO R., STOCHI
V., TISSERANT E., VISCOMI A. R., ZAMBONELLI A., ZAMPIERI E., HENRISSAT B.,
LEBRUN M-H., PAOLOCCI F., BONFANTE P., OTTONELLO S., WINCKER P., 2010.
Périgord black truffle genome uncovers evolutionary origins and mechanisms of symbiosis.
Nature, 464: 1033-1038.
MELLO A., MURAT C., BONFANTE P., 2006. Truffles: much more than a prized and local
fungal delicacy. FEMS Microbiology Letters, 260: 1-8.
MONCHABLON E., PARGNEY J-CL., 2003. Étude du statut mycorhizien d‘arbres producteurs
de truffes, 25 ans après plantation en Meuse. Bulletin de l'Académie Lorraine des Sciences, 42 (1-
4) : 1-9.
MEOTTO F., 1996. Caractérisation des mycorhizes sur le terrain. CIHEM-option
méditerranéenne, pp. 33-41.
MORENO G., GALAN R., ORTEGA A., 1986. Hypogeous fungi from continental
Spain. Cryptogamie Mycol., 7 (3): 201-229.
MOUSAIN D., PLASSARD CL., ARGILLIER C., SARDIN T., LEPRINCE F., EL
KARKOURI KH., ARVIEU J-CL., CLEYET-MAREL J-CL., 1994. Stratégie d‘amélioration de la
qualité des plants forestières et des reboisements méditerranéennes par utilisation de la
mycorhization contrôlée en pépinière. Acta. Bot. Gallica, 141 (4) : 571-580.
MOUSSU CH., LAURIAC ., 2008. La sylviculture truffière. Forêt de France, 518 : 15-26.
MULLER CL., SUSZKA B., BONNET-MASIMBERT M., 1994. Les grains des feuillus
forestiers de la récolte au semis. INRA. Paris,p.24.
MURAT C., DIEZ J., LUIS P., DELARUELLE C., DUPRE C., CHEVALIER G.,
BONFANTE P., MARTIN E., 2004. Polymorphism at the ribosomal DNA ITS and its relation
to postglacial re-colonization routes of the Périgord truffle Tuber melanosporum. New Phytologist,
164: 401-411.
MURAT CL., ZAMPIERI E., VIZZINI A., BONFANTE P., 2008. Is the périgord black truffle
threatene by an invasive species? We dreaded it and it has happened! New Phytologist, 178: 699-
702.
O‘DONNEL K., CIGELNIK E., WEBER N. S., TRAPPE J.M., 1997. In: BONUSO E., 2008.
Metodi moleculari nello studio della variabillita. Genetica di Tuber brochii. These doctorat. Alma
Mater Studiorum , Université Bolognia, p. 20.
OLIACH D., FISHER CH., OLIVERA A., COLINAS C., MARTINEZ de ARAGON J.,
BONET A., 2011. Black truffle production in Catalonia, Spain: the role of the growers association
in their developpement. In le VI th international workshop on edible mycorhizal mushrooms
(IWEMM6). Rabat, Maroc, 6-10 avril 2011, p. 71.

PACIONI G., BELLINA A., D‘ANTONIO M., 1990. Odeur composition of the Tuber
melanosporum complex. Mycol. Res., 94 (2): 201-204.
PARGUEY-LEDUC A., MONTANT C., KULIFAJ M., 1987. Résumé de la
Morphologie et structure de l‘ascocarpe adulte du Tuber melanosporum Vitt. (Black
Périgord Truffle). Microbiological Research, 84 (2) :277-278.
PARGUEY-LEDUC A., DARGENT R., SAIDALI-SARY C., JANEX-FARRE M.C., KULIFAJ
M., 1992. Ultrastructure localization chitine in the asci, ascospore and hyphal wall in Tuber
melanosporum. Mycological Research, 96: 59-64.
PARGNEY J-CL., 1994. Connaissance actuelles sur la structure, la biologie et l‘écologie de la
truffe. Bulletin des académies et société Lorraines des sciences, 3: 159-173.
PARGNEY J-CL., JALADE M., 1995. Etude cytologique des formations fongique en stromas se
développant a la surface des racines des plants truffiers. Bulletin de l'Académie Lorraine des
Sciences, 34 (1) : 27-44.
PARGNEY J-CL., MEUNIER G., 2004. Proposition d‘un nouveau type de truffière academie
Lorraine des Sciences, Villers-les-Nancy , France, 43 (1-4) : 2-11.
PERCUDANI R., TREVISI A., ZAMBONELLI A., OTTONELLO S., 1999. Molecular
Phylogeny of Truffles (Pezizales: Terfeziaceae, Tuberaceae) Derived from Nuclear rDNA
Sequence Analysis. Molecular Phylogenetic and Evolution, 13 (1):169–180.
PEREZ F., PALFNER G., BRUNEL N., 2007. Synthesis and establishement of Tuber
melanosporum Vitt. Ectomycorrhizae on two Nothofagus species in Chile. Mycorrhiza, 17: 627-
632.
PETHITORG J.C., ARDOIN-GUIDON F., CLIQUE M.T., CORMOND S.,
JOUSSERAND P., LE PONNER S., BOURLANDY M., 1995. Vrais et faux parasite
microscopique. Cahier de formation Parasitologique, p. 47.
PINKAS Y., MAIMON M., SHABI E., ELISHA S., SHMULEWICH Y., FREEMAN S., 2000.
Inoculation, isolation and identification of Tuber melanosporum from old new oak hosts. Mycol.
Res. 104 (4): 472-477.
PLASSARD CL., CHALOT M., BOTTON B., MARTIN F., 1997. Le rôle des éctomycorhizes
dans la nutrition azotée des arbres forestiers. Rev. For. Fr. XLIX-n°sp.: 82-98.
PRADEL E., 1914. In : CHEVALIER G., DESSOLAS H., PARGNEY J-CL., 2008. Nouveau
manuel de trufficulture. Réédition du manuel du Docteur Pradel (1914).Ed. Dessolas, Paris,
France, 308p.
PRUETT G.E., BRUN J.N., MIHAIL D.J., 2008. Colonization of pedunculate oak by the
Burgundy truffle fungus is greater with natural than with pelletized lime. Agroforest Syst., 72: 41-
50.

RAVOLANIRINA F., 1986. Etude de l‘influence de quelques facteurs sur la croissance
mycélienne des terfez in vitro et synthèse des mycorhizes. Mém. D.E.A., Univ. Clermont Ferrand,
France, 72p.
RIOUSSET G., RIOUSSET L., CHEVALIER G., BRADET M.C., 2001 : Truffe d‘Europe et de
Chine. Ed. INRA. France, 179p.
ROBIN B., ROBIN C., ROBIN-MASSANES CH., 2008. La mycorhization contrôlée.
Pépinières ROBIN, France, 32p.
ROUQUEROL T., PAYRE H., 1975a. In: ROUQUEROL T., PAYRE H., 1975. Observation
sur le comportement de Tuber melanosporum dans un site naturel. Revue de mycologie, XXXIX
(2) : 107-117.
ROUQUEROL T., PAYRE H., 1975b. Observation sur le comportement de Tuber
melanosporum dans un site naturel. Revue de mycologie, XXXIX (2) : 107-117.
ROUX C., SEJALON-DELMAS N., MARTIN M., PARGUEY-LEDUC A., DARGENT R.,
BECARD G., 1999. Phylogenetc relationships between European and Chinese truffles based on
parsimony and distance analysis of ITS sequences. FEMS. Microbiol. Lett., 180: 147-155.
RUBINI A., BELFIORI B., RICCIONI CL., TISSERANT E., ARCIONI S., MARTIN F.,
PAOLOCCI F., 2011. Isolation and characterization of MAT genes in the symbiotic ascomycete
Tuber melanosporum. New Phytologist, 189: 710-722.
SANTELICES R., PEREZ F., REYNA Y.S, RAMIREZ R., 2004. Introduction cultive de
Tuber melanosporum Vitt. en Chile. Jornadas Técnicas Forestales y Ambientales, 9 :
135-140.
SELOSSE M-A., 2000. La symbiose, structures et fonctions, role écologique et révolutif.
Ed. Veibert, Paris, 160p.
SELOSSE M-A., 2006. Animal ou végétal ? Pour la science, 350 : 66-72.
SHANNON M., 2011. Progress report on the Developpement of Mediterranean truffle orchards
in British Columbia, Canada. In le VI th international workshop on edible mycorhizal mushrooms
(IWEMM6). Rabat, Maroc, 6-10 avril 2011, p. 32.
SHAW P., 1995. Mycological societies of the world: association trufficulture du canton de
LAMBESC et environs. Mycologist August, 9(3): 115-116.
SMITH M.A., READ D.J., 2008. In: BONFANTE P., GENRE A., 2010. Mechanisms
underlying beneficial plant-fungus interactions in mycorrhizal symbiosis. Nature Communication,
48 (1): 1-11.
SOURZAT P., 2011. Black truffle cultivation and competing fungi. In Proceeding of the 7 th
international conference on mushroom biology and mushroom products (ICMBMP7). Section
Mycorhizal mushroom, pp. 516-528.

SPLIVALLO R., BOSSI S., MAFFEI M., BONFANTE P., 2007. Discrimination of truffle
fruiting body versus mycelial aromas by stir bar sorptive extraction. Phytochemistry, 68: 2584-
2598.
SPLIVALLO R., FISHER U., GOBEL C., FEUSSNER I., KARLOVSKY P., 2009. Truffles
regulate plant root morphogenesis via the production of auxine and ethylene. Plant Physiol., 150
(4): 2018-2029.
STRULLU D.G., 1991. Les mycorhizés des arbres et des plantes cultivés. Ed. Lavoisier, Paris,
250p.
TAGLIAFERRO F., 1996. Evaluation de la mycorhization en pot et en pleine terre : principe de
réglementation. CIHEA. Option méditerranéenne, pp. 63-73.
TANG Y-J., ZHOUG J-J., ZHU LI-W., 2008. Significance of fungal elicitor on the production of
ganoderic acid and Ganoderma polysaccharides by the submerged culture of medicinal mushroom
Ganoderma lucidum. Process. Biochemistry, 43 (12): 1359-1370.
THERVILLE CL., 2009. Cractérisation de la place de la trufficulture dans le paysage naturel et
culturel d‘Uzès. Rapport de Master Univ. Des sciences et Technique, Montpellier, 66p.
TORRES P., HUNROBIA M., 1994. Inoculation of containerized Pinus halepensis (Miller)
seedlings ans Suillus collinitus (Fr) OKuntze. Ann. Sci. for., 51: 521-528.
TRAPPE J.M., 1971. A synopsis of the Carbomyceteaceae and Terefeziaceae (Tuberales). Trans.
Br. Mycol. Soc., 57: 85-92.
TRAPPE J-M., 1979. The orders, famillies, and genera of hypogeous Ascomycotina (truffles and
relatives). Mycotaxon, 9: 297-340.
URBAIN A., NEUNER-PLATTNER I., KRISAI-GREILHUBER I., HASEL WANDTER K.,
2004. Molecular studies on trricolous microfungi reveal novel anamorphs of two Tuber species.
Mycol. Res., 108 (7): 749-758.
WANG Y., TAN ZM., ZHANG DC., MURAT C., JEANDROZ S., LE TACON F., 2006.
Phylogenetic and populational study of the Tuber indicum complex. Mycol. Res., 110: 1034-1045.
WANG S., MARCONE F. M., 2011. The biochemistry and biological properties of the World‘s
most expensive underground edible mushroom: Truffles. Food Research International, 44: 2567-
2581.
ZAMPIERI E., RIZZELLO R., BONFANTE P., MELLO A., 2012. The detection of
mating type gene of Tuber melanosporum in productive and non productive soils.
Applied Soil Ecology, 57: 9-15.

Annexe :
La table de Student (DELMAS, 2010)

Colorant : bleu de coton au lactophénol 0.1 % (LANGERON, 1952)
Acide lactique ……………………………………………………………….20g
Glycérol…………………………………………………………………………40g
Phénol……………………………………………………………………………20g
Eau distillé………………………………………………………………………20g
Bleu de méthyle………………………………………………………… ……0.1g

Résumé
Notre travail porte sur l’étude morphologique de la truffe noire Tuber
melanosporum Vitt. et son association mycorhizienne avec une essence forestière
Quercus ilex, en culture hors sol, en conditions contrôlées.
Les examens macroscopiques et microscopiques ont permis de connaître la
couleur des ascocarpes (noir marron), leur forme subglobuleuse recouverte par le
péridium rugueux portant des écailles pyramidales larges et des ports aérifères. La
gléba ou chair contient un tissu fertile, contient des asques globuleux renfermant 2 à
4 ascospores ellipsoïdes, ornementées par des épines dressées à extrémités
aigues. Toutes ces caractéristiques sont des critères importants pour l’identification
de Tuber melanosporum Vitt.
L’inoculation des plants de chêne vert avec Tuber melanosporum Vitt., en
culture hors sol a permis d’estimer la croissance aérienne des plants de chêne
inoculés. Après 12 mois de culture en serre, les résultats montrent que
l’inoculation améliore significativement la croissance des parties aériennes des
plants par rapport aux témoins ainsi que le développement de leur système
racinaire qui est dense par la présence de nombreuses racines courtes formant un
chevelu racinaire. Les racines courtes mycorhizées se forment en massue et des
glomérules se forment sur les racines inoculées. Le taux de mycorhization des
plants est de 10 %.
Les examens microscopiques de l’association mycorhizienne révèlent la
présence de filaments mycéliens enveloppant la racine pour former le manteau
fongique et des hyphes septés parcourant les espaces intercellulaires des cellules
corticales pour former le réseau du Hartig. Toutes ces observations indiquent que
ce champignon forme chez le chêne vert une ectomycorhize.
Mots clés :
Truffe Noire; Tuber Melanosporumvitt.; Quercus Ilex L.; Asque; Ascospore; Chêne
Vert; Inoculation; Culture Hors Sol; Mycorhization; Ectomycorhize.