Embed Size (px)
Citation preview

CLASSIFICATION DES ESPECES
La théorie cellulaire a été émise au 18ème siècle :
« LA CELLULE EST L’UNITE STRUCTURALE ET FONCTIONNELLE
DE TOUS LES ETRES-VIVANTS »
PROCARYOTES
EUCARYOTES
Métazoaire – animal « cellules animales pluricellaires »
Métaphyte – végtal « cellules végétales pluricellaires »
Protozoaire – prophyte – protiste « cellelules animales unicellaires »
ARCHEES BACTERIES
ANIMAUX PLANTES CHAMPIGNONS PROTISTES

GENERALITES SUR LA CELLULE
La cellule est la plus petite unité capable de réaliser toutes les réactions nécessaires à la vie
« respiration, nutrition … » Elle représente donc la plus petite unité de la matière vivante.
Elle s’alimente, respire, communique avec d’autres cellules et se reproduit.
Il existe des êtres-vivants unicellaires « une cellule : exemple les bactéries » et des êtres-
vivants pluricellaires « plusieurs cellules : exemple l’Homme »
Les êtres-vivants présentent une grande diversité. Pourtant tous sont constitués de cellule.
Chez un même organisme, les cellules présentent des différences de forme, de taille et aussi
au niveau de leurs organites. Elles sont spécialisées dans une fonction. Selon celle-ci une
cellule peut être plus ou moins riche en organites cellulaires. Malgré ces différences, les
cellules ont une organisation commune.
On distingue deux types de cellules : les cellules procaryotes et les cellules eucaryotes.
Les cellules eucaryotes possèdent un noyau. Une membrane plasmique délimite le
cytoplasme, dans lequel baignent les organites. Chaque organite exerce une fonction.
Parmi les cellules eucaryotes, les cellules végétales se distinguent des cellules animales par la
présence d’une paroi cellulaire et d’organites spécifiques comme une grande vacuole et des
chloroplastes.
Les cellules procaryotes, par exemple les bactéries, sont constituées d’un cytoplasme
dépourvu d’organites. L’information génétique est libre dans le cytoplasme.
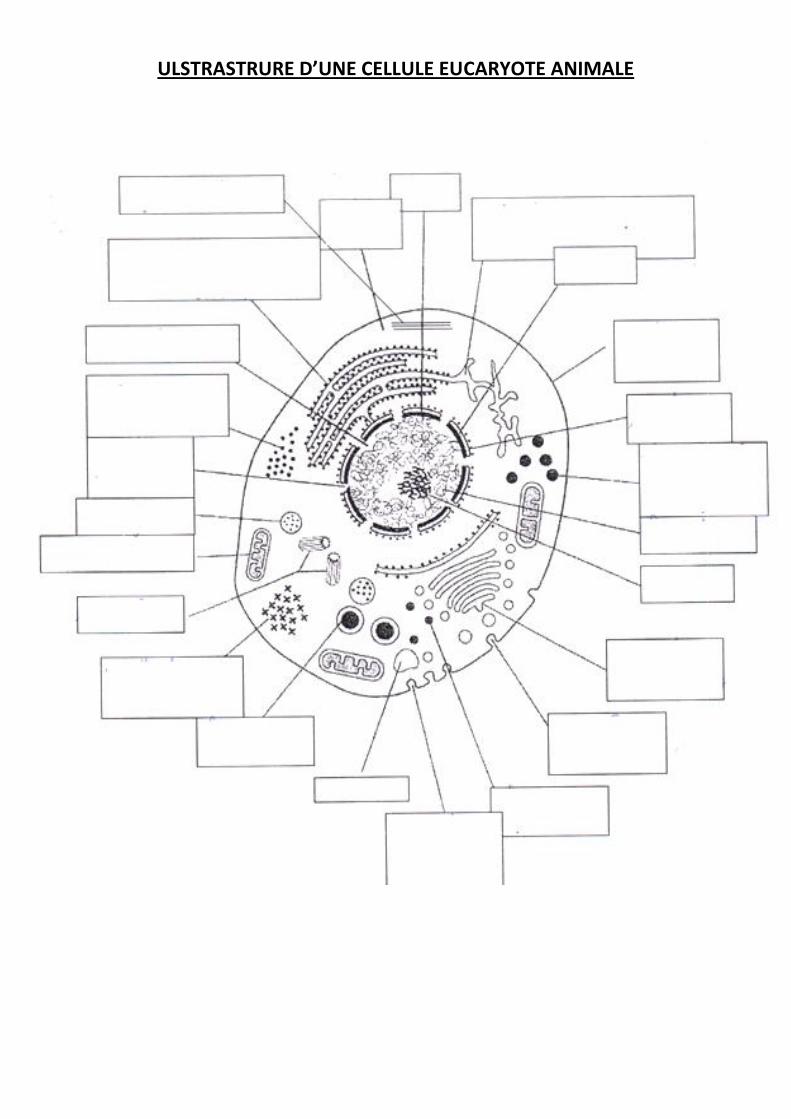
ULSTRASTRURE D’UNE CELLULE EUCARYOTE ANIMALE
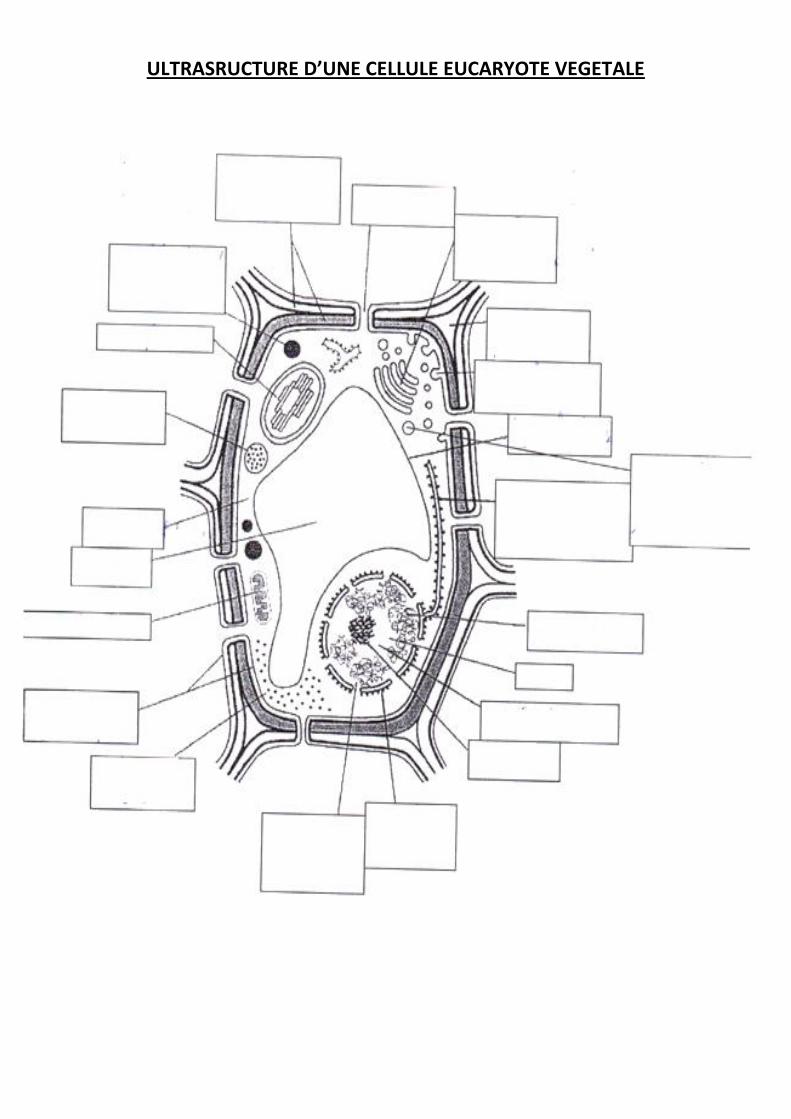
ULTRASRUCTURE D’UNE CELLULE EUCARYOTE VEGETALE
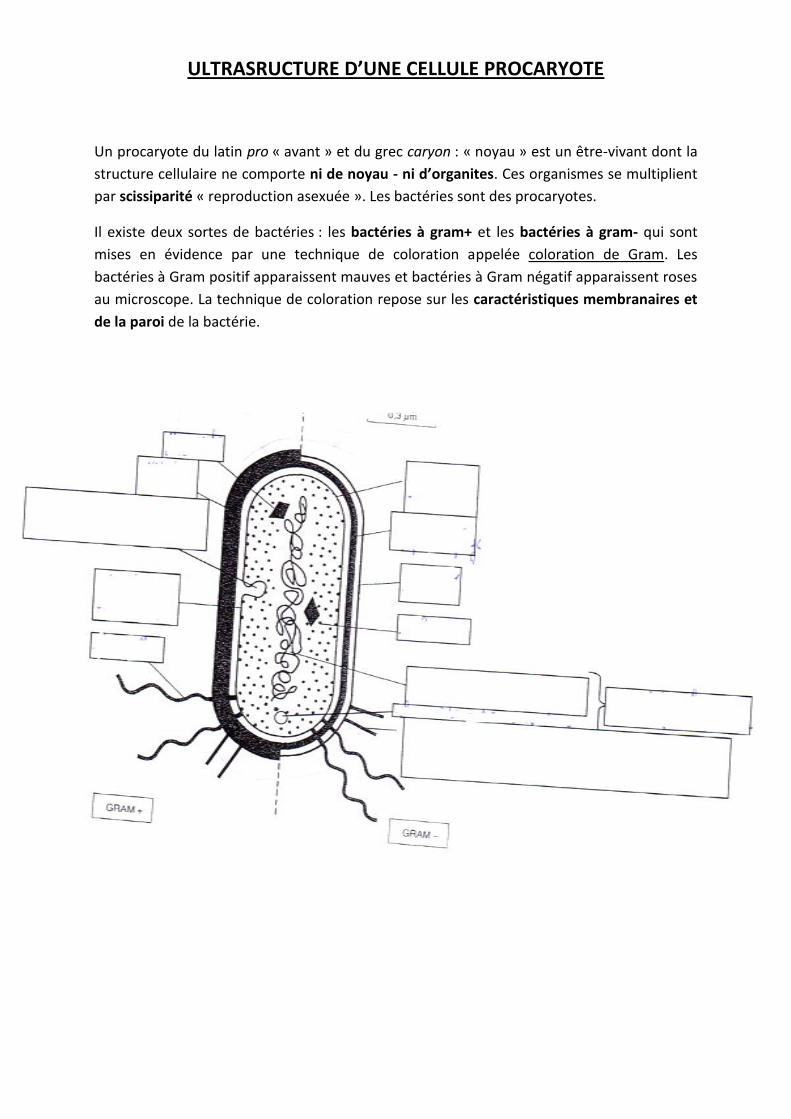
ULTRASRUCTURE D’UNE CELLULE PROCARYOTE
Un procaryote du latin pro « avant » et du grec caryon : « noyau » est un être-vivant dont la
structure cellulaire ne comporte ni de noyau - ni d’organites. Ces organismes se multiplient
par scissiparité « reproduction asexuée ». Les bactéries sont des procaryotes.
Il existe deux sortes de bactéries : les bactéries à gram+ et les bactéries à gram- qui sont
mises en évidence par une technique de coloration appelée coloration de Gram. Les
bactéries à Gram positif apparaissent mauves et bactéries à Gram négatif apparaissent roses
au microscope. La technique de coloration repose sur les caractéristiques membranaires et
de la paroi de la bactérie.

La membrane plasmique
La membrane plasmique sépare le milieu intracellulaire du milieu extracellulaire et délimite
la cellule. Elle est à distinguer des endomembranes qui délimitent les organites.
Elle est constituée de deux couches de phospholipides qui forment une bicouche
phospholipidiques.
Au niveau de la membrane plasmique des cellules eucaryotes animales, on trouve un
manteau appelé le glycocalyx.
En effet, les phospholipides présentent une partie hydrophile et une partie hydrophobe, on
dit qu’ils sont amphiphiles.
Entre les phospholipides, on peut trouver des molécules de cholestérol « liquide de nature
stéroïdiennes » et des protéines transmembranaires qui traverse la bicouche
phospholipidique.
On trouve aussi des protéines extrinsèques, c’est-à-dire accolées à la face externe de la
membrane plasmique et des protéines intrinsèques, c’est-à-dire accolées à la face interne
de la membrane plasmique. Elles assurent différentes fonctions :
- servent de récepteurs membranaires
- servent de reconnaissance de la cellule
- servent à adhérer à d’autres cellules
- permettent la formation de pores ou canaux pour le passage d’eau, des ions et des
substances hydrophiles.

La membrane a un rôle de protection et de zone d’échange entre le milieu intracellulaire, du
milieu extracellulaire.
Plusieurs paramètres vont jouer dans la fluidité de la membrane :
- la composition lipidique : plus il y a de cholestérol, plus la fluidité membranaire
diminue et la membrane devient de plus en plus rigide.
- la nature des acides gras : plus il y a des acides gras insaturés, plus la fluidité
membranaire augmente – de même plus la chaine carbonée des acides gras est
longue plus la fluidité diminue et la membrane devient rigide.
- la composition protéique : plus il y a de protéines, plus la fluidité diminue
- la température : le froid diminue la fluidité membranaire.
Le transport membranaire est l’un des principaux rôles de la membrane plasmique. La
membrane n’est pas perméable à toutes les cellules, on dit qu’elle a une barrière semi-
perméabilité très sélective.
Les mécanismes passifs : les cellules baignent dans une substance riche en substances
dissoutes « glucose, oses, acides gras, vitamines … ». La membrane est perméable à
certaines substances et imperméables à d’autres. Les mécanismes passifs sont ceux qui se
font naturellement c’est-à-dire qu’il n’y a pas besoin d’énergie. Il en existe trois :
- la diffusion simple : elle tend à réaliser un équilibre de part et d’autre de la
membrane plasmique, c’est-à-dire que les particules diffusent selon le gradient de
concentration « les particules vont du milieu le plus concentré, vers le milieu le
moins concentré »
Milieu hypertonique Milieu hypotonique
- l’osmose « concerne l’eau » : la membrane plasmique est très perméable à l’eau. Elle
passe du milieu le moins concentré vers le milieu le plus concentré.
Milieu hypotonique Milieu hypertonique

- la diffusion facilitée : le mécanisme est le même que celui de la diffusion simple :
c’est-à-dire que les particules diffusent selon le gradient de concentration « les
particules vont du milieu le plus concentré, vers le milieu le moins concentré ». Ce
transport est assuré par des transporteurs membranaires de nature protéique :
pompes.
Milieu hypertonique Milieu hypotonique
Les mécanismes actifs ou transports actifs : ce transport ne se fait pas naturellement. Il
nécessite une énergie fournie par l’ATP. Ceci, car les molécules sont transportées dans le
sens contraire du gradient de concentration, c'est-à-dire qu’elles vont passer du milieu le
moins concentré, vers le milieu le plus concentré. Ce transport est assuré par des
transporteurs membranaires de nature protéique : pompes.
Milieu hypotonique Milieu hypertonique
Le transport vésiculaire : ce transport nécessite également la production d’énergie et donc
d’ATP. Il existe différents types de transport vésiculaire :
- l’exocytose : sécrétion ou élimination de substances présentes dans la cellule ; la
substance est enfermée dans une vésicule d’exocytose « sac membraneux qui
fusionne avec la membrane plasmique et s’ouvre vers l’extérieur en relâchant la
substance en question ».
1
2
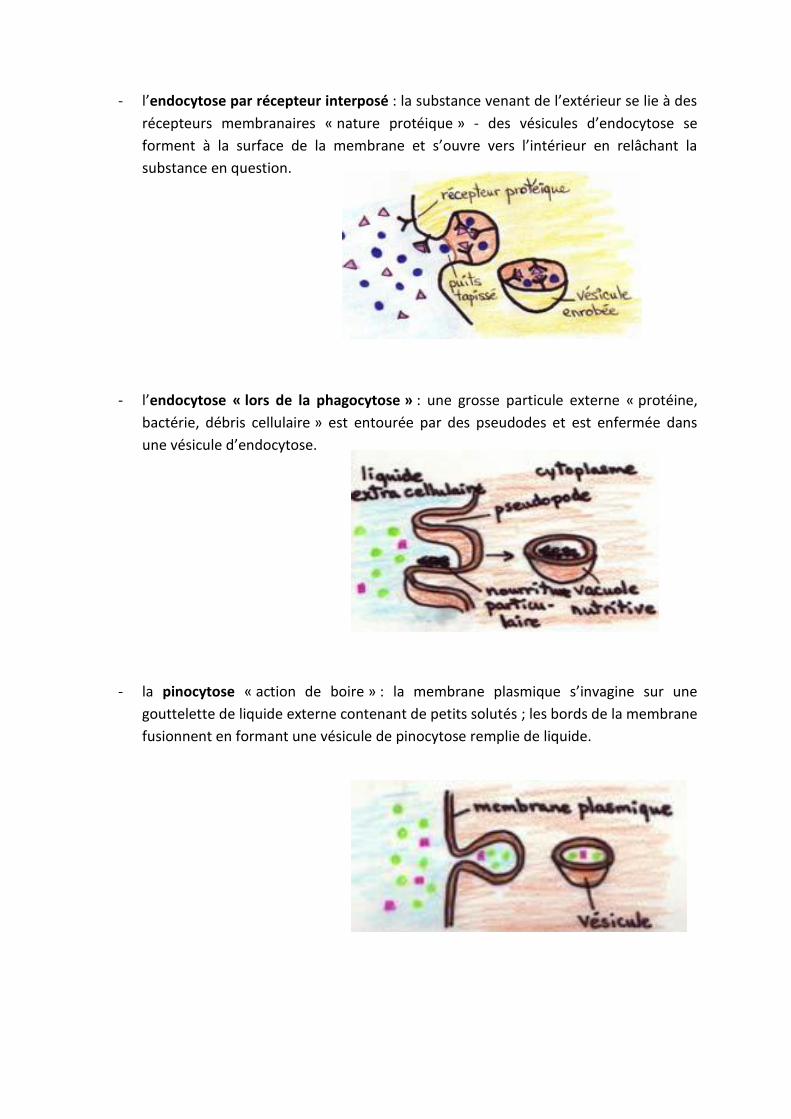
- l’endocytose par récepteur interposé : la substance venant de l’extérieur se lie à des
récepteurs membranaires « nature protéique » - des vésicules d’endocytose se
forment à la surface de la membrane et s’ouvre vers l’intérieur en relâchant la
substance en question.
- l’endocytose « lors de la phagocytose » : une grosse particule externe « protéine,
bactérie, débris cellulaire » est entourée par des pseudodes et est enfermée dans
une vésicule d’endocytose.
- la pinocytose « action de boire » : la membrane plasmique s’invagine sur une
gouttelette de liquide externe contenant de petits solutés ; les bords de la membrane
fusionnent en formant une vésicule de pinocytose remplie de liquide.

Le noyau
Le noyau est l’organite cellulaire contenant le matériel génétique. Il est absent chez les
cellules procaryotes et présent chez les cellules eucaryotes.
Toutes les cellules humaines sont nucléés « possèdent un noyau » à l’exception des globules
rouges (hématies ou érythrocytes). On dit qu’elles sont anucléées.
Le noyau est constitué d’une membrane nucléaire qui peut porter des ribosomes et être en
continuité avec le réticulum endoplasmique rugueux ou granuleux. A certains endroits,
l’enveloppe nucléaire forme des pores nucléaires.
A l’intérieur du noyau se trouve un ou plusieurs nucléoles. Le nucléole est le lieu de synthèse
des ribosomes.
De plus, le nucléoplasme (intérieur du noyau) est composé de chromatine. Celle-ci est
constituée de l’association d’une molécule d’ADN avec des protéines d’histones. L’ADN,
porteur du programme génétique, est localisé au niveau du noyau des eucaryotes. L’ADN se
réplique dans le noyau, c’est-à-dire qu’il produit des répliques de lui-même. Le noyau est
indispensable à la vie cellulaire, car il joue un rôle fondamental dans la synthèse de la
matière vivante, dans la division cellulaire et dans les mécanismes de l’hérédité.
On trouve également un centrosome près du noyau qui contient une paire de centrioles.
L’enveloppe nucléaire ou membrane nucléaire
Elle délimite le noyau. Elle est formée de 2 membranes qui sont séparées par l’espace
périnucléaire. La membrane externe est en continuité avec la membrane du réticulum
endoplasmique et porte des ribosomes.
Elle est percée de larges pores qui permettent le passage de molécules entre le noyau et le
cytoplasme et inversement. Les 2 membranes « interne et externe »se rejoignent au niveau
des pores nucléaires. Ces pores représentent 30% de la surface de l’enveloppe.
Pendant l’interphase, l’enveloppe emprisonne les chromosomes dans le noyau. Au cours de
la mitose « division cellulaire » l’enveloppe nucléaire est rompue momentanément, ses
fragments sont dispersés dans le cytoplasme.
Le nucléole
C’est un amas de chromatine plus ou moins spiralée. Le nucléole est un sous-compartiment cellulaire du noyau et est un lieu où se produit la transcription des ARN ribosomiques, qui constituent avec des protéines, les deux sous-unités des ribosomes. C’est aussi le lieu de la transcription des ARNt.

Le nucléoplasme
Le nucléoplasme est un liquide qui apparaît grisâtre ponctuée de noir en microscopie
électronique contenu dans le noyau. Il contient en moyenne entre 70% et 90% d’eau. Il est
riche en enzymes intervenant entre autres dans la synthèse de l’ADN et de l’ARN, ou
permettant le stockage intracellulaire de produits de réserve, comme des graisses et des
glucides, en nucléotides et en ATP.
Le cytoplasme
Le cytoplasme est le matériel cellulaire situé à l’intérieur de la cellule mais à l’extérieur du
noyau. Il est composé de deux éléments :
Hyaloplasme « appelé aussi le cytosol » : c’est un liquide qui contient des molécules
organiques diverses et des ions et où baignent le cytosquelette et des organites.
Organites : exécutent des fonctions répondant aux besoins de toutes les cellules.
C’est le lieu où se déroulent les principales réactions biochimiques de la cellule.
Les ribosomes
Toutes les cellules y compris les procaryotes contiennent des ribosomes. Le ribosome est un
complexe protéique de deux sous unités : une grande sous-unité de 65S « S : Svedberg,
coefficient de sédimentation » et une petite sous-unité de 40S. L’ensemble des deux sous-
unités fait 80S.
Ils peuvent être libres dans le cytosol - associés à la membrane du réticulum endoplasmique
– en chainette libre « polysomes : plusieurs ribosomes ». Ils participent à la synthèse des
protéines « traduction ».
Le réticulum endoplasmique
Le réticulum endoplasmique forme des cavités tubulaires qui communiquent entre elles. On
distingue trois types de réticulum endoplasmique :
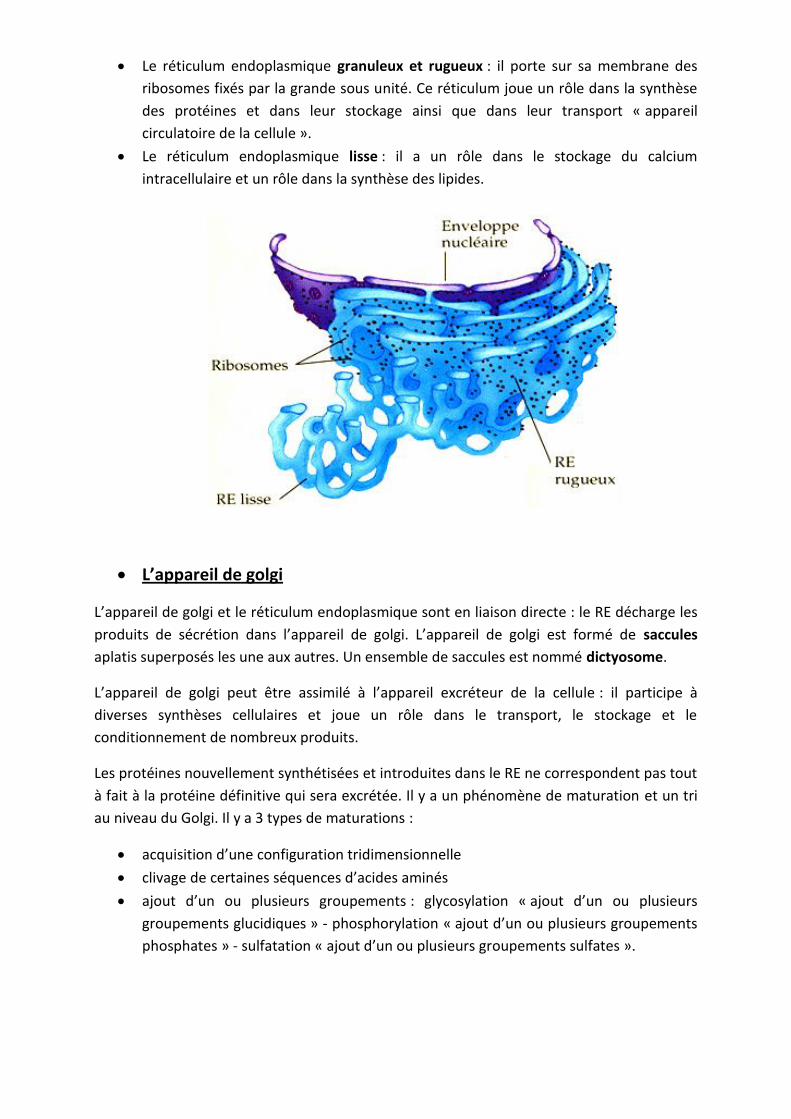
Le réticulum endoplasmique granuleux et rugueux : il porte sur sa membrane des
ribosomes fixés par la grande sous unité. Ce réticulum joue un rôle dans la synthèse
des protéines et dans leur stockage ainsi que dans leur transport « appareil
circulatoire de la cellule ».
Le réticulum endoplasmique lisse : il a un rôle dans le stockage du calcium
intracellulaire et un rôle dans la synthèse des lipides.
L’appareil de golgi
L’appareil de golgi et le réticulum endoplasmique sont en liaison directe : le RE décharge les
produits de sécrétion dans l’appareil de golgi. L’appareil de golgi est formé de saccules
aplatis superposés les une aux autres. Un ensemble de saccules est nommé dictyosome.
L’appareil de golgi peut être assimilé à l’appareil excréteur de la cellule : il participe à
diverses synthèses cellulaires et joue un rôle dans le transport, le stockage et le
conditionnement de nombreux produits.
Les protéines nouvellement synthétisées et introduites dans le RE ne correspondent pas tout
à fait à la protéine définitive qui sera excrétée. Il y a un phénomène de maturation et un tri
au niveau du Golgi. Il y a 3 types de maturations :
acquisition d’une configuration tridimensionnelle
clivage de certaines séquences d’acides aminés
ajout d’un ou plusieurs groupements : glycosylation « ajout d’un ou plusieurs
groupements glucidiques » - phosphorylation « ajout d’un ou plusieurs groupements
phosphates » - sulfatation « ajout d’un ou plusieurs groupements sulfates ».

Le RE et l’appareil de golgi forment un réseau endomembranaire :
Les lysosomes
Les lysosomes se sont des petites vésicules délimitées par une membrane. Ils contiennent
des enzymes « protéines permettant d’accélérer les réactions chimiques » digestives et
interviennent dans la digestion des organites inutilisables dans la cellule ainsi que dans la
digestion de certains matériaux exogènes.

Les peroxysomes
Ce sont des petites vésicules remplies d’enzymes. Ils sont chargés de la détoxification de la
cellule.
Les centrosomes – centrioles
Un centriole est un cylindre comprenant 9 groupes de 3 tubules. Une cellule en interphase
comprend 2 centrioles perpendiculaires l’un à l’autre et en général proches du noyau, ils
sont entourés d’un nuage protéique. Ils forment le centrosome. Une cellule en mitose
comprend 2 centrosomes soit 4 centrioles.
Leur rôle est de coordonner et de réguler les mouvements intracellulaires. Ils jouent un rôle
important lors de la division cellulaire notamment dans la formation du fuseau mitotique.

Le cytosquelette
Le cytosquelette est constitué de microfibres :
les microtubules : se sont des bâtonnets creux ayant un rôle déterminant dans la
cellule. Ils sont formés à partir d’une protéine appelée tubuline. En interphase, ils
sont disposés radialement à partir du centrosome. En mitose, lors de la métaphase,
ils constituent le fuseau mitotique.
les microfilaments : se sont de longues fibres composées de protéines appelées
actine et myosine. Selon le type et le rôle de la cellule, les microfilaments se
présentent sous différentes formes « contraction – mobilité – prolongements
cytoplasmiques ».
les filaments intermédiaires : se sont de longs bâtonnets droits ou courbés,
composés de différentes protéines qui donnent une certaine rigidité au
cytosquelette. On distingue plusieurs types de filaments intermédiaires selon les
différents types cellulaires : « les filaments de kératine – neurofilaments … »

Les mitochondries
Les mitochondries sont des organites filiformes. Elles se composent de 2 membranes
mitochondriales, une interne et une externe, qui délimitent trois milieux : le milieu extra-
mitochondrial – l’espace inter-membranaire – et la matrice. La membrane interne forme des
invaginations qui apparaissent sous forme de crête au microscope électronique. La
mitochondrie contient son propre ADN – l’ADN mitochondrial.
Les mitochondries sont impliquées dans la respiration cellulaire et donc la production
d’énergie. L’énergie est stockée sous forme d’ATP.
C’est dans la mitochondrie que se déroulent les dernières étapes du cycle respiratoire qui
convertit l’énergie des molécules organiques issues de la digestion « glucose » en énergie
directement utilisable par la cellule « ATP » : en présence de d’oxygène c’est-à-dire en
condition d’aérobiose.
En absence d’oxygène, c'est-à-dire en condition d’anaérobiose, la cellule utilise la
fermentation dans le cytoplasme pour produire l’énergie nécessaire à son fonctionnement,
mais c’est un système beaucoup moins efficace, qui dégrade de façon incomplète le substrat
« production d’acide lactique donnant lieu à des crampes par exemple ».
Toutes les cellules de l’organisme ont besoin d’énergie pour assurer leur activité, mais
certaines sont beaucoup plus exigeantes que d’autres : les cellules à fort activité synthétique
« cellule glandulaire – cellule nerveuse – cellule musculaire ».
Il n’existe pas de stock d’ATP dans les cellules – ceci implique que dans les cellules qui
consomment beaucoup d’énergie, il y a nécessité d’une régénération efficace de l’ATP.

La régénération de l’ATP se fait par 3 voies différentes en fonction de l’équipement
enzymatique de la cellule – de l’intensité de l’activité – des conditions d’oxygénation :
la première voie est la voie anaérobie alactique ou voie de la phosphocréatine : c’est
la voie de régénération immédiate, elle est entièrement cytoplasmique. Elle utilise la
phosphocréatine comme donneur de phosphate :
𝐴𝐷𝑃 + 𝑝ℎ𝑜𝑠𝑝ℎ𝑜𝑐𝑟é𝑎𝑡𝑖𝑛𝑒 → 𝐴𝑇𝑃 + 𝑐𝑟é𝑎𝑡𝑖𝑛𝑒
la deuxième voie est la voie anaérobie lactique ou voie de la fermentation lactique :
elle est entièrement cytoplasmique. Elle utilise la dégradation des glucides « en
particulier le glucose » et comprend deux étapes : la glycose « qui à partir d’une
molécule de glucose donne deux molécules d’acide pyruvique puis formation d’acide
lactique ». Lors de la glycolyse, il y a formation de deux moles d’ATP par mole de
glucose oxydée : c’est une dégradation incomplète puisqu’elle aboutit à la formation
de composé organique encore riche en énergie.
la troisième voie est la voie aérobie ou voie de la respiration cellulaire.
La respiration cellulaire est une réaction chimique d’oxydo-réduction « transfert d’électrons
d’une chimique à une autre. On appelle oxydant : l’espèce qui capte les électrons –
réducteur : l’espèce qui cède les électrons » qui fournit l’énergie nécessaire à une cellule
pour fonctionner.
La respiration cellulaire nécessite :
un carburant : il s’agit du glucose – d’acides gras – et autres molécules organiques.
Dans le cas des humains, il provient de la digestion et est amené par la circulation
sanguine. Dans le cas des pantes, il provient de la dégradation du saccharose ou de la
dégradation de l’amidon.
de l’oxygène : dans le cas des humains : il est extrait de l’air par la ventilation
pulmonaire et est amené à la cellule par la circulation sanguine, fixé sur
l’hémoglobine des hématies.
La respiration cellulaire se fait en 4 étapes principales :
glycolyse
l’oxydation du pyruvate
le cycle de Krebs
la chaine respiratoire
La dégradation des molécules organiques glucidiques débute dans le hyaloplasme : c’est la
glycolyse au cours de laquelle la molécule de glucose est oxydée en 2 molécules de pyruvate
et 2 molécules de transporteur oxydé sont réduites – la réaction est :
𝑪𝟔𝑯𝟏𝟐𝑶𝟔 + 𝟐𝑻 → 𝟐 𝑪𝑯𝟑 − 𝑪𝑶 − 𝑪𝑶𝑶𝑯 + 𝟐 𝑻𝑯, 𝑯+
Glucose Transporteur oxydé Pyruvate Transporteur réduit

Cette réaction d’oxydo-réduction est couplée à la synthèse d’ATP à partir d’ADP et de
phosphate inorganique.
Le pyruvate oxydation est ensuite dégradé dans les mitochondries en CO2 et en H2O – c’est
ce processus qui assure la production de la majeure partie de l’ATP nécessaire aux activités
cellulaires : la respiration.
La dégradation du pyruvate débute dans la matrice des mitochondries au cours d’un cycle de
réactions « cycle de Krebs » - il s’agit de réactions de décarboxylation et d’oxydoréduction
aboutissant à la production de CO2 et de transporteur réduit TH, H+.
Ces molécules de transporteur réduit ainsi que celle provenant de la glycolyse sont
réoxydées au niveau de la membrane interne des mitochondries grâce à la chaîne
respiratoire. Le bilan est la réduction du dioxygène en eau et la réoxydation des transports
réduits. Ces réactions sont couplées à la synthèse d’ATP. De là, résulte la production de la
quasi-totalité de l’ATP cellulaire.
Dans ces conditions, la dégradation du glucose est totale. Elle aboutit uniquement à la
formation de composés minéraux non énergétiques « CO2 – H2O ».
Le rendement énergétique est plus important que pour la fermentation : à partir d’un mole
de glucose on forme 36 moles d’ATP.
Equation-bilan :
Schéma-bilan :

Le chloroplaste
Ces organites sont spécifiques des cellules végétales. Ils sont présents dans le cytoplasme
des cellules végétales chlorophylliennes. Ils jouent un rôle essentiel dans le fonctionnement
d’une cellule végétale car ils permettent de capter la lumière à l’origine de la
photosynthèse.
Par l’intermédiaire de la chlorophylle « pigment » qu’ils possèdent et de leurs
ultrastructures, ces organites sont capables de transférer l’énergie véhiculée par les photons
à des molécules chimiques.
Les chloroplastes sont des plastes, c'est-à-dire des organites cellulaires végétaux qui peuvent
se charger de pigments ou de diverses substances nutritives « par exemple : les
amyoloplastes chargés d’amidon ». Les plastes contiennent également leur propres ADN.
Les chloroplastes prennent la forme de disques aplatis. Il s’agit d’un organite composé de
deux membranes séparées par un espace intermédiaire. Il contient un réseau membraneux
constitué de sacs aplatis nommés thylakoides qui baignent dans le stroma « liquide
intrachloroplastique ». Les thylakoides sont composés d’un lumen « lumière » entouré d’une
membrane, et contiennent de la chlorophylle « pigments vert » et des caroténoides
« pigments jaune ». Un empilement de thylakoides se nomme granum « au pluriel des
grana ». Ils possèdent leur propre ADN.
Le chloroplaste est un élément indispensable à la photosynthèse. Il absorbe l’énergie
lumineuse pour la transformer en énergie chimique sous forme d’ATP.
Le chloroplaste absorbe l’ensemble du spectre de la lumière visible émis à part le vert raison
pour laquelle les feuilles des plantes ont un aspect vert. La chlorophylle se trouve dans la
membrane des thylakiodes.

Les différentes étapes de la photosynthèse qui convertissent la lumière en énergie chimique
se déroulent dans les thylakoides tandis que les étapes de la conversion de l’énergie en
glucide se déroulent dans le stroma du chloroplaste.
Le chloroplaste joue aussi un rôle dans la biosynthèse des lipides.
La photosynthèse est le processus bioénergétique qui permet aux plantes de synthétiser
leur matière organique en exploitant l’énergie lumineuse. C’est la fabrication de matière
carbonée organique à partir d’eau et de carbone minéral « CO2 » en présence de lumière.
La photosynthèse utilise l’énergie lumineuse « d’origine solaire ou artificielle » pour
fabriquer du sucre « glucose ». Il s’agit d’une réaction d’oxydo-réduction au cours de
laquelle l’eau est oxydée et le dioxyde de carbone est réduit :
𝟔𝑪𝑶𝟐 + 𝟔𝑯𝟐𝑶 + 𝜺 𝒍𝒖𝒎𝒊𝒏𝒆𝒖𝒔𝒆 → 𝑪𝟔𝑯𝟏𝟐𝑶𝟔 + 𝟔𝑶𝟐
Cette réaction résulte de multiples réactions que l’on peut regrouper en 2 phases :
La 1ère, dite photochimique « face claire » se déroule au niveau des membranes des
thylakoides des chloroplastes où sont localisés les pigments chlorophylliens et les
systèmes enzymatiques nécessaires à sa réalisation « cytochromes ou
photosyntèmes : transporteurs d’électrons et sphères pédonculées – synthèse d’ATP
avec ATP synthétase ». L’énergie des photons captés par les pigments
chlorophylliens au cours de cette phase entretiennent un ensemble de réactions au
cours desquelles l’eau est oxydée « protolyse de l’eau » et des transporteurs T sont
réduits :
𝟐𝑯𝟐𝑶 + 𝟐𝑻 → 𝟐(𝑻𝑯, 𝑯+) + 𝑶𝟐
Cette phase ne nécessite pas de CO2 mais ne peut se dérouler qu’à la lumière et
uniquement si les molécules de transporteur oxydé sont continuellement présentes.
Les réactions d’oxydoréduction qui ont lieu durant cette phase sont couplées à la
synthèse d’ATP.
La 2ème phase dite non photochimique « cycle de calvin – face de fixation du carbone
- face sombre » se déroule dans l’obscurité comme la lumière et a lieu dans le
stroma des chloroplastes. Le CO2 est incorporé dans des molécules organiques « le
glucose, l’amidon stocké dans les plastes » au cours d’un ensemble de réactions
mobilisant les molécules de transporteurs réduits (𝑇𝐻, 𝐻+) et ATP formées au cours
de la 1ère phase.
Les deux phases de la photosynthèse sont couplées. A la suite de cette phase, les molécules
de transporteur se retrouvent à l’état oxydé et l’ATP sous forme ADP + Pi , ce qui permet le
déroulement permanent à la lumière de la phase photochimique.

Equation-bilan :
Schéma-bilan :
La vacuole
La vacuole est une cavité remplie de fluide dans les cellules végétales et dans les cellules des
champignons dont la fonction est emmagasinée les éléments nutritifs et l’eau.
La vacuole centrale des cellules végétales et des champignons a une fonction
homéostatique, assurant la turgescence cellulaire, et sa membrane « le tonoplaste » possède des pompes ioniques. Elle contient de l’eau – des glucides – des ions organiques et
inorganiques – et des pigments. Certaines vacuoles centrales sont également des sites
d’accumulation de réserves ou de substances particulières, parfois toxiques « latex … » voire
de déchets, ce qui leur confère un rôle de détoxification de la cellule.
La vacuole représente entre 80 et 90% du volume cellulaire de la cellule végétale adulte. La
vacuole joue un rôle primordial dans la croissance de la cellule végétale.

La paroi pecto-cellulosique
La paroi végétale est un élément de structure cellulaire qui protège chaque cellule végétale.
C’est est une structure qui évolue en fonction de l’âge des tissus végétaux.
La paroi d’une cellule végétale adulte est constituée de plusieurs couches :
La paroi d’une cellule adulte est constituée :
- d’une membrane plasmique parsemée de plasmodesme.
- d’une paroi secondaire composée de beaucoup de celluloses – d’hémicelluloses – et
de lignines formant une structure résistante et indéformable qui assure à la fois la
protection mécanique et le soutien de la cellule « on parle de paroi squelettique ».
Seules les cellules adultes possèdent cette paroi.
- d’une paroi primaire composée d’un peu de celluloses - d’hémicelluloses – et de
pectines – et de protéines.
- d’une lamelle moyenne composée de pectines : assure l’adhésion entre les cellules.
De par sa composition, on qualifie cette paroi de paroi pecto-cellulosique.
La paroi primaire est capable de se déformer et c’est cette déformation qui est à l’origine de
la croissance cellulaire. Il existe deux types de déformation de la paroi :
- 1 réversible et rapide caractérisant son élasticité.
- 1 irréversible et lente caractérisant sa plasticité.
Les plasmodesmes
Le plasmodesme est le canal traversent la paroi de la cellulaire végétale. Il relie les
membranes plasmiques et les cytoplasmes des cellules adjacentes, qui forment alors un
compartiment continu, le symplasme. L’ensemble de plasmodesmes représente en moyenne
1% de la surface de la paroi, mais certaines zones en présentent une concentration élevée,
alors que d’autres en sont quasiment dépourvues.

LE METABOLISME CELLULAIRE PRIMAIRE
Le métabolisme est l’ensemble des transformations moléculaires et énergétiques qui se
déroulent de manière ininterrompue dans la cellule ou l’organisme vivant. C’est un
processus ordonné, qui fait intervenir des processus de dégradation « catabolisme » et de
synthèse organique « anabolisme ».
Le métabolisme hétérotrophe
Le métabolisme hétérotrophe c’est le métabolisme des cellules eucaryotes animales.
On appelle hétérotrophie la nécessité pour un organisme de disposer d’une source de
matière organique pour synthétiser sa propre matière organique. L’organisme fabrique sa
propre matière organique à partir de la matière organique prélevée dans le milieu extérieur
« extracellulaire ».
L’organisme prélève :
- soit exclusivement des végétaux « phytophage »
- soit exclusivement des animaux « zoophage »
- soit des végétaux et des animaux « omnivore »
- soit de la matière organique en décomposition « saprophage »
Le métabolisme autotrophe
On appelle autotrophie la capacité d’un organisme à synthétiser sa propre matière
organique à partir de matière minérale : eau et ions minéraux prélevées dans le sol en
dioxyde de carbone atmosphérique en présence de lumière.
Cette synthèse de matière organique s’accompagne d’un prélèvement de dioxyde de
carbone et d’un rejet d’O2. Elle nécessite de l’énergie lumineuse et la présence des
chloroplastes : c’est le métabolisme de la photosynthèse.
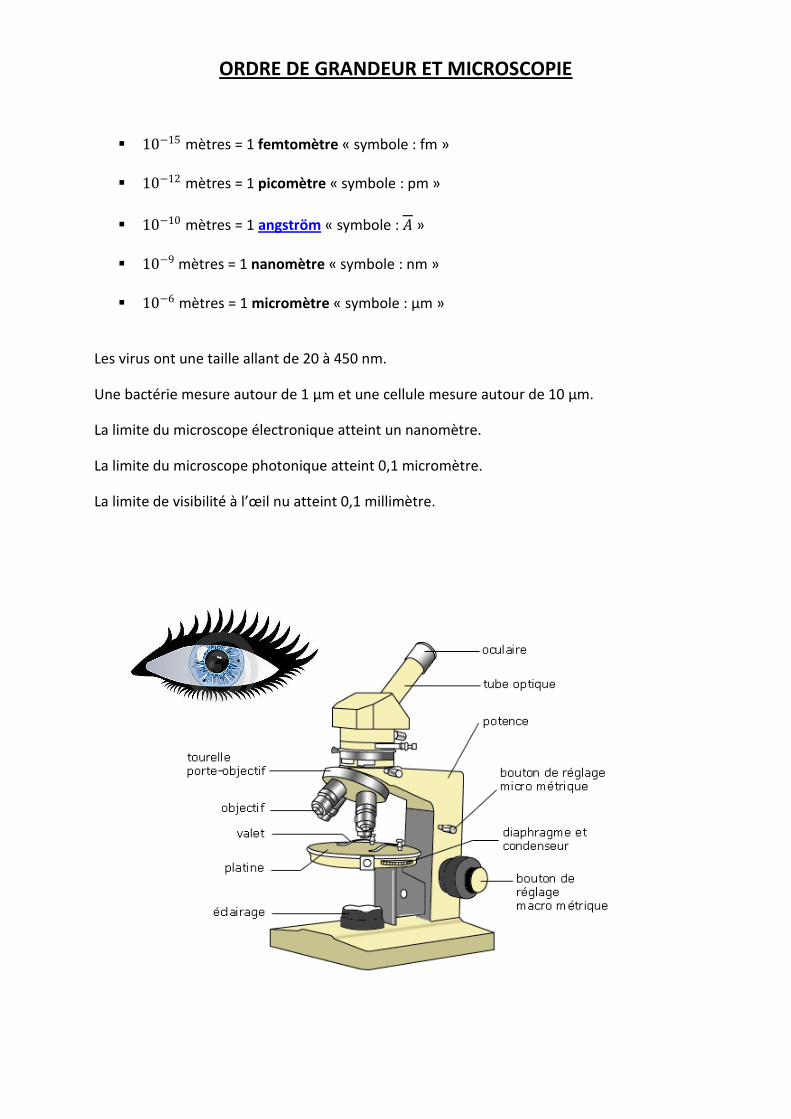
ORDRE DE GRANDEUR ET MICROSCOPIE
10−15 mètres = 1 femtomètre « symbole : fm » 10−12 mètres = 1 picomètre « symbole : pm »
10−10 mètres = 1 angström « symbole : 𝐴 » 10−9 mètres = 1 nanomètre « symbole : nm »
10−6 mètres = 1 micromètre « symbole : µm »
Les virus ont une taille allant de 20 à 450 nm.
Une bactérie mesure autour de 1 µm et une cellule mesure autour de 10 µm.
La limite du microscope électronique atteint un nanomètre.
La limite du microscope photonique atteint 0,1 micromètre.
La limite de visibilité à l’œil nu atteint 0,1 millimètre.

QUELQUES TESTS CYTOCHIMIQUES A CONNAITRE
Le test de Brachet : ce test utilise du vert de méthyl « colorant histologique ou
cytologique basique vert » et de la pyronine « colorant histologique ou cytologique
basique rouge ». Il permet de mettre en évidence l’ADN et/ou l’ARN dans une
cellule. Le plus souvent, il est utilisé pour marquer la chromatine « en vert » dans une
cellule. vert – ADN / rouge – ARN.
La réaction de Feulgen : ce test utilise le réactif de Schiff « composé de fuchsine,
colorant histologique ou cytologique rouge ». Il permet de mettre en évidence la
présence d’ADN dans une cellule. La chromatine devient rouge violacé.
Le test à l’eau iodée « ou lugol » : coloration bleu marine – marron : il permet de
mettre en évidence les polysaccharides « amidon – glycogène ». amidon – bleu
marine / glycogène – marron.
Le test à la liqueur de Fehling « bleu » : coloration rouge brique : il permet de mettre
en évidence les oses.
La réaction de Biuret « avec du sulfate de cuivre » : coloration mauve violacée : il
permet de mettre en évidence les liaisons peptides et donc les protéines.





![African History – Histoire Africaine [L’Unite Culturelle de l’Afrique Noire] Les Fondements Moraux Du Pouvoir Africain](https://img.pdfslide.fr/doc/110x75/577c7eb61a28abe054a229ec/african-history-histoire-africaine-lunite-culturelle-de-lafrique.jpg)