Embed Size (px)
Citation preview

L’un des enjeux de la biologiecellulaire et moléculaire estde comprendre les méca-nismes d’un événement quise produit au sein d’un orga-
nisme ou d’une cellule vivante.Cependant, les techniques dispo-nibles permettant d’étudier la cellulevivante sont encore rares. Le déve-loppement d’une méthode simple
qui permet la visualisation d’événe-ments cellulaires dans leur environ-nement naturel constitue donc uneavancée importante en biologie cellu-laire : cette méthode repose sur lagreen fluorescent protein (GFP), pro-téine fluorescente extraite de laméduse Aequorea victoria. Le clonagedu gène de la GFP dans un premiertemps [1] et l’étude de son expres-
45
La green fluorescentprotein : applicationà la dynamiqueintracellulairedes récepteurs stéroïdiens
La green fluorescent protein (GFP) est une protéine extraitede la méduse Aequorea victoria. Cette protéine a la particu-larité d’émettre de la fluorescence sous UV sans l’ajout desubstrat ou de co-facteurs. Le clonage de son gène etl’étude de son expression dans des bactéries ou dans desorganismes plus complexes tel que C. elegans ont enrichi lesperspectives de son application dans la recherche biomédi-cale. Le gène de la GFP est aujourd’hui utilisé comme gènerapporteur, comme marqueur cellulaire ou encore commemarqueur moléculaire. Le gène codant pour la GFP a parexemple été fusionné aux gènes codant pour des récep-teurs stéroïdiens afin de connaître la dynamique intracellu-laire de ces récepteurs. A ce jour, ont été rapportées lesprotéines de fusion avec le récepteur des glucocorticoïdes(GFP-GR), le récepteur des minéralocorticoïdes (GFP-MR)et le récepteur des androgènes (GFP-AR). La fluorescenceémise par la GFP permet de localiser les protéines dans lescompartiments cellulaires et d’en définir ainsi les déplace-ments lors de stimulations diverses, rendant accessible lavisualisation d’événements cellulaires dynamiques.
ADRESSESV. Georget : étudiante en thèse. J.C. Nicolas :directeur de recherche à l’Inserm. Inserm U. 439,Pathologie moléculaire des récepteursnucléaires, 70, rue de Navacelles, 34090Montpellier, France. C. Sultan : professeur demédecine à l’université de Montpellier I. InsermU. 439, Pathologie moléculaire des récep-teurs nucléaires, 70, rue de Navacelles,34090 Montpellier, France. Unité BEDR,hôpital Lapeyronie, 34295 MontpellierCedex 5, France. Unité d’endocrinologiepédiatrique, Hôpital A.-de-Villeneuve,34295 Montpellier Cedex 5, France.
SYNTHÈSEmédecine/sciences 1999 ; 15 : 45-55
m/s n° 1, vol. 15, janvier 99
Virginie GeorgetJean-Claude NicolasCharles Sultan
34243$1 045-055 2/05/02 17:34 Page 45

sion dans des organismes tels que labactérie Escherischia coli ou le néma-tode Caenorhabditis elegans, dans unsecond temps, [2] ont ouvert denombreuses perspectives dans l’utili-sation de cette protéine en biologiecellulaire et en génétique molécu-laire. Le système GFP est aujourd’huiutilisé comme marqueur cellulairepour identifier puis trier une popula-tion cellulaire transformée [3],comme gène rapporteur pour mesu-rer l’expression d’un gène dans descellules vivantes [2], ou encorecomme marqueur moléculaire pourvisualiser, dans une cellule vivante, ladynamique d’une protéine fusionnéeà la GFP [4]. Dans cet article, nousdévelopperons plus particulièrementce dernier aspect dans l’étude de ladynamique intracellulaire des récep-teurs stéroïdiens.
Green fluorescentprotein : structure,fonctionnement
Parmi les nombreux organismes bio-luminescents, la méduse Aequorea vic-toria peut émettre une lumière vertede ses photocytes jusqu’aux bords deson ombrelle [5]. Cette fluorescenceest engendrée par l’activationséquentielle de deux protéines :l’aequorine, une protéine dépen-dante du calcium et la green fluores-cent protein (GFP) (figure 1). En pré-sence d’oxygène, l’aequorine estcouplée à la coelentérazine. La fixa-tion de calcium provoque un change-ment conformationnel de l’aequo-rine qui induit une oxygénation de lacoelentérazine. Cette réaction d’oxy-génation s’accompagne d’une émis-sion de lumière bleue. Dans laméduse, un transfert d’énergie entrel’aequorine et la GFP conduit àl’émission d’une lumière verte [6].In vitro, la fluorescence de la GFPpeut être obtenue par excitation enlumière bleue ou en UV.La GFP est une protéine de 238 acidesaminés avec une masse molaire de 27à 30 kDa en gel polyacrylamide déna-turant (SDS-PAGE) [7]. Cette pro-téine est très stable et résistante à denombreuses conditions physico-chi-miques dénaturantes. In vitro, la GFPconserve sa fluorescence à des tem-pératures aussi élevées que 65 °C etdans une gamme de pH de 5,5 à 12avec, toutefois, une diminution
d’intensité pour des pH inférieurs à7. La fluorescence de la GFP résistedans des solutions qui contiennentjusqu’à 6M d’hydrochlorure de gua-nidium, 8M d’urée, 1 % de SDS oudans des solvants organiques dilués.La GFP reste aussi fluorescente aprèsfixation des cellules par du glutaral-déhyde ou du paraformaldéhyde.Même si la majeure partie de la pro-téine (acides aminés 7 à 229) estnécessaire à l’acquisition de la fluo-rescence de la GFP [8], son chromo-phore n’est constitué que de troisacides aminés en position Ser65-Tyr66-Gly67 [9]. In vitro, la GFP estdonc fluorescente sans ajout de sub-strat ou de co-facteurs [2]. Sa cristal-lisation révèle que la protéine estconstituée de 11 feuillets β, d’unehélice α qui contient le chromo-phore et de boucles entre les feuillets[10, 11]. Les 11 feuillets β formentune cage dans laquelle est empri-sonné le chromophore, les boucles
formant le plancher de la cage (figure2). Cette forme surprenante de laGFP est appelée forme « β-can ». Laformation du chromophore au seinde la cage se produit par une modifi-cation post-traductionnelle autocata-lytique des acides aminés 65-67. Cestrois résidus sont dans un premiertemps cyclisés puis oxydés pour for-mer le chromophore p-hydroxyben-zylidene-imidazolidinone (figure 3)[7, 12, 13]. C’est seulement aprèsl’oxydation que la GFP est fluores-cente. Cette oxydation est égalementsuivie d’un cycle de protonation-déprotonation. La formation de cechromophore actif se produit en 2 à4 heures environ. La GFP native pré-sente deux pics d’absorption, un picmajeur à 395 nm et un pic d’unemoindre intensité autour de 477 nm,et un pic d’émission à 509 nm. Cesdeux pics d’absorption semblent êtreassociés à deux formes isomères duchromophore qui peuvent être sous
46 m/s n° 1, vol. 15, janvier 99
Lumière bleue
Lumière verte
Coelentérazine
Coelentéramide
Transfertd'énergie
Ca2+
GFP
Apoaequorine Aequorine
Figure 1. Mécanisme d’activation dela GFP dans la méduse Aequorea vic-toria. En présence d’oxygène, l’aequo-rine est une protéine dépendante ducalcium couplée à la coelentérazine.La fixation de calcium provoque unchangement conformationnel del’aequorine qui induit une oxygéna-tion de la coelentérazine. Cette réac-tion d’oxydation s’accompagne d’uneémission de lumière bleue. Il s’établitalors un transfert d’énergie entrel’aequorine et la GFP. Ce transfertd’énergie conduit à l’émission d’unelumière verte de la GFP.
Figure 2. Structure cristallogra-phique de la GFP. La GFP est consti-tuée de 11 feuillets β et d’une héliceα. Les feuillets β sont disposés enparallèle et forment une cage danslaquelle est enfermée l’hélice α quicontient le chromophore. Lesboucles entre les feuillets β formentle plancher de la cage. Cette formesurprenante de la GFP est appelée« β can ». (Cette image a été repro-duite avec l’aimable autorisation deM. T.D. Romo, Université de Rice.)
34243$1 045-055 2/05/02 17:34 Page 46

forme stable et sous forme excitée.Après irradiation à 395 nm, la formestable absorbe à 477 nm. Cette pro-priété est d’ailleurs à la base d’uneapplication de la GFP. Elle consiste àirradier par un laser UV (395 nm) laGFP, ce qui augmente la forme quiabsorbe à 477 nm et la fluorescencede la GFP lorsqu’elle est excitée àcette longueur d’onde d’où son nomde sur-fluorescence [14]. Ces deuxisomères correspondent à des formesprotonées et déprotonées [7] quisont maintenant mieux comprisesdepuis la cristallisation de la GFP sau-vage [11] et qui sont dues à des inter-actions spécifiques entre les acidesaminés du chromophore et les acidesaminés des feuillets β positionnés surla face interne de la cage [15]. A cejour, de nombreuses variantes de la
GFP existent, chacune apportant denouvelles propriétés de fluorescence.La GFP-S65T est mutée dans le chro-mophore en position 65 et substitueune thréonine à la sérine [16]. CetteGFP présente des caractéristiques defluorescence modifiées (Tableau I).En effet, la mutation simplifie lespectre à un seul pic d’absorption à488nm [16], ce qui permet de détec-ter cette GFP avec des filtres clas-siques de fluorescéine isothiocyanate(FITC). De plus, cette substitutionqui induit une fluorescence six foisplus forte que celle de la GFP sau-vage est plus résistante au photoblan-chiment. Certaines mutations tellesque la S65T sont connues pour aug-menter la vitesse de formation duchromophore et notamment la cycli-sation, ce qui réduit le temps delatence entre la synthèse et la fluores-cence de la GFP [16]. Pour la plupartdes GFP, la formation optimale dufluorophore dépend du pH et de latempérature. Par exemple, la GFPnative est plus fluorescente à 28 °Cqu’à 37 °C [17]. Trois GFP mutées,l’α -GFP mutée en positionF100S/M154T/V164A [18], l’EGFP(enhanced-GFP) mutée en F64L/S65T[19, 20] et la GFP mutéeS65T/S147P [21] ont la capacitéd’acquérir la conformation correcteà toute température physiologique[21, 22] (Tableau I). Ces dernières,beaucoup plus fluorescentes que laGFP sauvage, sont les candidatesidéales pour une étude en cellulevivante. Néanmoins, l’intensité defluorescence des GFP S65T et del’EGFP diminue très rapidement àdes pH inférieurs à 7, contrairementà la GFP sauvage et l’α-GFP qui sontrésistantes à une large gamme de pH[22]. Cette propriété est maintenantutilisée pour mesurer des variationsde pH au sein de différents comparti-ments d’une seule cellule [23]. Enrevanche, ces GFP ne sont pas adap-tées pour l’étude de phénomènes quise déroulent dans des compartimentsacides tels que les lysosomes ou lesendosomes. La stabilité de la GFPreprésente un avantage dans desapplications de marquage molécu-laire mais elle peut aussi être uninconvénient quand le gène de laGFP est utilisé comme gène rappor-teur. Cette stabilité ne permet pasd’étudier des phénomènes de syn-thèse rapide et transitoire. C’est
pourquoi, de nouvelles constructionsde la GFP ont été récemment déve-loppées : il s’agit de la fusion à la par-tie carboxy-terminale de la GFP depetits peptides qui seraient suscep-tibles de rendre la GFP plus sensibleaux protéases et donc de réduire sontemps de demi-vie [24]. De plus, unenouvelle GFP déstabilisée estaujourd’hui commercialisée. Elle aun temps de demi-vie réduit à2 heures. Il n’y a donc pas une seuleGFP idéale pour toutes lesrecherches en biologie cellulaire oumoléculaire : chaque GFP présentedes avantages et des inconvénientsdont il faut tenir compte pour uneutilisation efficace [22].Le mutant Y66H et le double mutantY66H/Y145F cristallisé par Watcher[25] émettent une lumière bleue à445 nm lorsqu’ils sont excités à381nm [26] (Tableau I). Ces mutantsont été les premières GFP décrites quiémettent une fluorescence de couleurdifférente et sont donc appelés BFP(blue fluorescent protein). Il existe égale-ment maintenant l’EBFP [27]. Demême que le mutant EYFP(S65G/S72A/T203Y/V68L) émet unefluorescence jaune à 527 nm aprèsune excitation à 513 nm [10]. Toutecette nouvelle gamme de GFP avecdes spectres d’excitation et d’émissionaltérés est importante pour au moinsdeux raisons. Tout d’abord, ces GFPsont un outil supplémentaire pourl’étude de plusieurs molécules expri-mées simultanément dans la mêmecellule vivante. En effet, en utilisantdes filtres très spécifiques, il est pos-sible de détecter jusqu’à trois GFP dif-férentes dans la même cellule [26].Ces mutants peuvent également servirde donneur ou d’accepteur d’énergiedans un système de transfert d’éner-gie (fluorescence resonance energy transfer,FRET) [26].
Green fluorescentprotein : applications
Grâce à toutes ses propriétés, la GFPest une protéine de choix dans la bio-logie du vivant et de nombreusesapplications ont déjà été rapportées àce jour.
GFP gène rapporteur
Le gène de la GFP peut être utiliséen tant que gène rapporteur. Le
47m/s n° 1, vol. 15, janvier 99
OHN
N
NH
OH
N
H
O
O
O2 Oxydation-O
NN
O
O
OH
N
OH
H
O
O
ON
N
O-
O
+H+
-H+
OHN
N
NH
OH
N
H
O
O
CyclisationH2O
NH
N
H
OH
N
H
OH
N
H
Tyr66 Gly67
Ser65
Figure 3. Formation du chromophorede la GFP. Le chromophore de laGFP est constitué des trois acidesaminés Ser-Tyr-Gly en position 65 à67. Sa formation se produit aprèsune cyclisation entre les acides ami-nés 65 et 67 puis une oxydation. Lechromophore p-hydroxybenzylidène-imidazolidinone est alors constitué. Ilsera sous forme protoné ou dépro-toné. C’est seulement après l’oxygé-nation que la GFP peut être fluores-cente.
34243$1 045-055 2/05/02 17:34 Page 47
gène de la GFP, sous la dépendanced’un promoteur d’intérêt, témoignede l’activité de ce promoteur par lasimple fluorescence émise de la GFP.La GFP présente certains avantagespar rapport à d’autres protéinestelles que la chloramphénicol acétyltransférase (CAT) ou la β-galactosi-dase, dont les gènes sont souvent uti-lisés comme gènes rapporteurs. Toutd’abord, la mesure de la fluorescenceest une quantification du nombre deprotéines synthétisées et non pas ledosage d’une activité enzymatique.La GFP est détectable dans des cel-lules vivantes, ce qui permet de mesu-rer une fluorescence émise par descellules ou par un tissu, à plusieurs
48 m/s n° 1, vol. 15, janvier 99
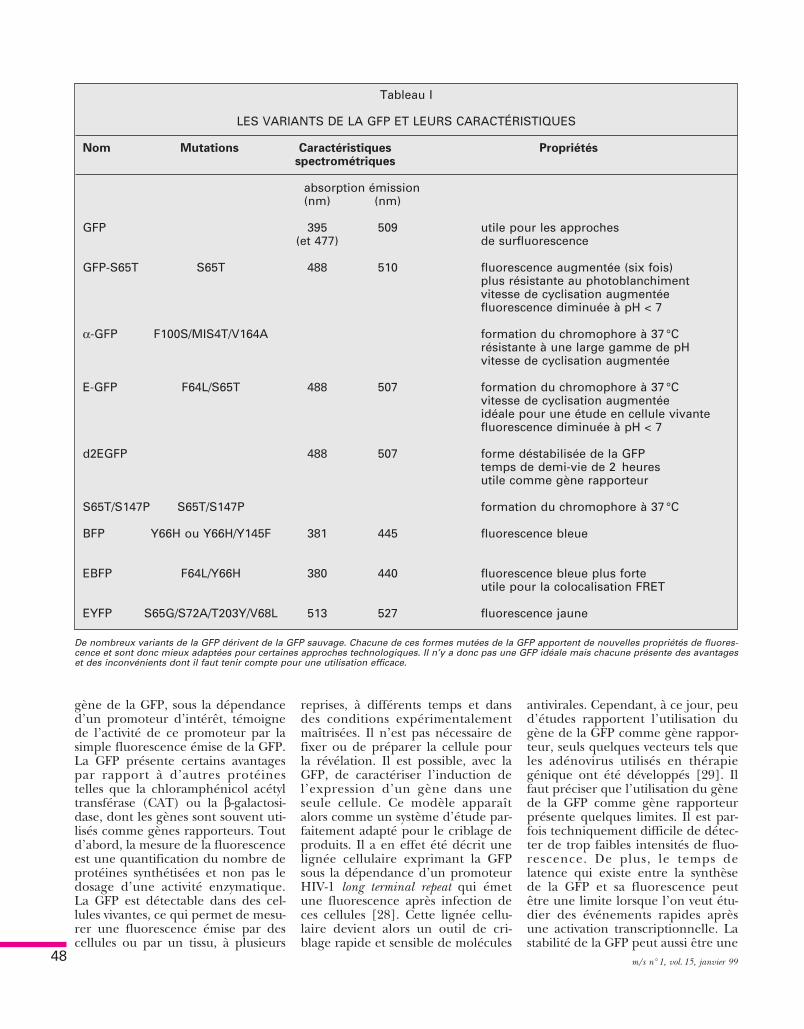
Tableau I
LES VARIANTS DE LA GFP ET LEURS CARACTÉRISTIQUES
Nom Mutations Caractéristiques Propriétésspectrométriques
absorption émission(nm) (nm)
GFP 395 509 utile pour les approches(et 477) de surfluorescence
GFP-S65T S65T 488 510 fluorescence augmentée (six fois)plus résistante au photoblanchimentvitesse de cyclisation augmentéefluorescence diminuée à pH < 7
α-GFP F100S/MIS4T/V164A formation du chromophore à 37 °Crésistante à une large gamme de pHvitesse de cyclisation augmentée
E-GFP F64L/S65T 488 507 formation du chromophore à 37 °Cvitesse de cyclisation augmentéeidéale pour une étude en cellule vivantefluorescence diminuée à pH < 7
d2EGFP 488 507 forme déstabilisée de la GFPtemps de demi-vie de 2 heuresutile comme gène rapporteur
S65T/S147P S65T/S147P formation du chromophore à 37 °C
BFP Y66H ou Y66H/Y145F 381 445 fluorescence bleue
EBFP F64L/Y66H 380 440 fluorescence bleue plus forteutile pour la colocalisation FRET
EYFP S65G/S72A/T203Y/V68L 513 527 fluorescence jaune
De nombreux variants de la GFP dérivent de la GFP sauvage. Chacune de ces formes mutées de la GFP apportent de nouvelles propriétés de fluores-cence et sont donc mieux adaptées pour certaines approches technologiques. Il n’y a donc pas une GFP idéale mais chacune présente des avantageset des inconvénients dont il faut tenir compte pour une utilisation efficace.
reprises, à différents temps et dansdes conditions expérimentalementmaîtrisées. Il n’est pas nécessaire defixer ou de préparer la cellule pourla révélation. Il est possible, avec laGFP, de caractériser l’induction del’expression d’un gène dans uneseule cellule. Ce modèle apparaîtalors comme un système d’étude par-faitement adapté pour le criblage deproduits. Il a en effet été décrit unelignée cellulaire exprimant la GFPsous la dépendance d’un promoteurHIV-1 long terminal repeat qui émetune fluorescence après infection deces cellules [28]. Cette lignée cellu-laire devient alors un outil de cri-blage rapide et sensible de molécules
antivirales. Cependant, à ce jour, peud’études rapportent l’utilisation dugène de la GFP comme gène rappor-teur, seuls quelques vecteurs tels queles adénovirus utilisés en thérapiegénique ont été développés [29]. Ilfaut préciser que l’utilisation du gènede la GFP comme gène rapporteurprésente quelques limites. Il est par-fois techniquement difficile de détec-ter de trop faibles intensités de fluo-rescence. De plus, le temps delatence qui existe entre la synthèsede la GFP et sa fluorescence peutêtre une limite lorsque l’on veut étu-dier des événements rapides aprèsune activation transcriptionnelle. Lastabilité de la GFP peut aussi être une
34243$1 045-055 2/05/02 17:34 Page 48

limite dans des études d’expressiontrès transitoire. Enfin, la restrictionmajeure à l’utilisation du gène de laGFP en tant que gène rapporteur estliée aux limites d’intensité de fluores-cence quantifiables. La limite infé-rieure : le signal doit être distinguédu bruit de fond de fluorescence quiest variable suivant les cellules, et lalimite supérieure : toutes les GFP syn-thétisées ne sont pas fluorescenteslors d’une synthèse importante deprotéines, car l’étape d’oxydation dela tyrosine devient limitante.Apparemment, le gène rapporteurGFP ne constitue pas un progrèsmajeur par rapport au gène rappor-teur luciférase, largement utilisé.Cependant, la synthèse de la GFPsous la dépendance d’un promoteurspécifique permet d’étudier l’expres-sion d’un gène dans une seule celluleet de déterminer le lieu et lemoment de son expression dans unorganisme.
GFP marqueur cellulaire
Inversement, par sa facilité et sa rapi-dité de détection, la GFP est une pro-téine candidate idéale comme mar-queur cellulaire. Cette application estd’ores et déjà très répandue : c’estcertainement celle qui connaîtra leplus grand essor. Lors d’une co-trans-fection de deux vecteurs codant pourdeux protéines, dont la GFP, la fluo-rescence permet de sélectionner,dans une population cellulaire, lescellules qui co-expriment les deuxprotéines par un simple tri des cel-lules fluorescentes. Ce tri se fait com-munément par un FACS (fluorescenceactivated cell sorter) et peut être réalisédans le cadre d’une transfection tran-sitoire dans le but d’établir unelignée stable [3]. Par FACS, il est éga-lement possible de trier des popula-tions cellulaires exprimant des GFPdifférentes : des cellules positivespour la GFP, d’autres positives pourla BFP et des cellules qui co-expri-ment simultanément les deux pro-téines fluorescentes [30, 31]. Dansl’établissement d’animaux transgé-niques, la sélection des cellulessouches exprimant la protéine trans-gène est une étape importante. Ellepeut maintenant se réaliser par unsimple tri de cellules fluorescentesco-exprimant la GFP et la protéinetransgène. De plus, la GFP n’est pas
toxique chez les animaux transgé-niques. Ainsi, par une sélection decellules souches positives pourl’EGFP, Okabe et al. ont obtenu dessouris transgéniques vertes [32] donttous les organes sont fluorescents.La GFP peut également servircomme marqueur pour suivre ledevenir d’un type cellulaire dans unorganisme, par exemple la progres-sion de cellules tumorales GFP-posi-tives dans une souris nude [33].
GFP marqueur moléculaire
La GFP peut aussi être fusionnée àdes protéines afin d’étudier leur loca-lisation cellulaire et d’analyser leurdevenir dans des cellules vivantes. Laprotéine de fusion peut être obtenuepar des techniques classiques de sous-clonage à partir de toute une gammede vecteurs d’expression commercia-lisés possédant des multisites de clo-nage adjacents aux séquences codantpour l’extrémité amino-terminale oucarboxy-terminale du gène de laGFP. Cette construction est intro-duite par transfection de manièretransitoire ou stable dans le systèmed’expression choisi. La fluorescencepeut ensuite être détectée par diffé-rentes technologies microscopiquesmunies d’un filtre FITC.La fluorescence dans la cellule peutêtre analysée par un microscopeconfocal qui permet une reconstitu-tion tri-dimensionnelle de la cellule àpartir des couches successives réali-sées dans la cellule. Le microscopeconfocal apporte une très bonnedéfinition de l’image de la cellule etdonc une localisation précise de lafluorescence. Toutefois, il n’est pastrès adapté dans des études de ciné-tique et est utilisé le plus souventpour observer des cellules fixées : ladurée et la répétition des mesures dela cellule conduisent à un photoblan-chiment de la fluorescence et à lamort de la cellule.Le cytomètre à balayage est une autretechnique d’analyse de fluorescence.Le temps d’acquisition de l’imaged’une cellule est plus long que celuid’un microscope confocal, ce qui per-met d’utiliser une puissance de laserbeaucoup plus faible et ainsi de conser-ver l’intégrité de la cellule. Il est, enrevanche, beaucoup moins résolutif.Ce microscope muni de son laser àpuissance maximale est parfaitement
adapté pour les études de photoblan-chiment décrites ultérieurement.Le microscope à épifluorescenceoffre un bon compromis entre ladéfinition de l’image, la rapiditéd’acquisition et la viabilité de la cel-lule. C’est également le plus adapté àdes études de cinétique sur des cel-lules vivantes : ainsi, contrairementaux techniques d’immunomarquage,la même cellule peut être observéedans différentes conditions. La GFPpermet donc de visualiser en tempsréel et dans l’espace la protéined’intérêt.La GFP seule est distribuée uniformé-ment dans la cellule alors que fusion-née à la protéine analysée, la fluores-cence a pu être détectée dans descompartiments très divers de la cellule[34, 35]. La GFP a notamment étéfusionnée avec des protéines impli-quées dans le cytosquelette [36], desprotéines du réticulum endoplas-mique [37], de l’appareil de Golgi[38], des mitochondries [39], maiségalement des protéines membra-naires pour suivre la localisation desrécepteurs membranaires, leur inter-nalisation et leur recyclage [40]. Préci-sons qu’en général la GFP ne modifiepas la fontion de ces protéines malgréson poids moléculaire élevé (27 kDa).Ont aussi été décrites des protéineschimériques de la GFP avec des pro-téines cytoplasmiques ou nucléaires,telles que les récepteurs stéroïdiens.
Application à ladynamique intracellulairedes récepteursstéroïdiens
Les récepteurs stéroïdiens appartien-nent à la superfamille des récepteursnucléaires. Il s’agit de facteurs trans-criptionnels qui règlent – après acti-vation par un ligand spécifique –l’expression de gènes cibles [41, 42].Cette famille comprend le récepteurdes œstrogènes (ER), le récepteur dela progestérone (PR), le récepteurdes minéralocorticoïdes (MR), lerécepteur des glucocorticoïdes (GR)[43] et enfin le récepteur des andro-gènes (AR).Ces récepteurs présentent une grandehomologie de leur structure primaire,fondée sur la présence de quatregrands domaines : le domaine amino-terminal, le domaine de liaison àl’ADN (DBD), la région charnière et
49m/s n° 1, vol. 15, janvier 99
34243$1 045-055 2/05/02 17:34 Page 49

le domaine de liaison de l’hormone(LBD). Alors que le mécanismed’action moléculaire des différentsmembres de la famille des récepteursnucléaires apparaît très similaire, leurlocalisation cellulaire en l’absenced’hormone est différente [44].L’analyse de la distribution cellulairede ces récepteurs a été réalisée parimmunomarquage de cellules ou detissus. Il est généralement reconnuque ER et PR sont des récepteursnucléaires en l’absence d’hormone[45, 46] avec toutefois une navette(shuttle) entre le cytoplasme et lenoyau [47]. La localisation cellulairede MR, GR et AR a été très long-temps controversée. En fonction del’anticorps utilisé, MR apparaît uni-quement cytoplasmique ou à la foiscytoplasmique et nucléaire [48]. Desdonnées contradictoires ont été éga-lement rapportées pour GR en fonc-tion des conditions de fixation et deperméabilisation des cellules [43].AR a été trouvé dans le noyau des cel-lules dans des tissus cibles [49, 50],alors que dans des cellules transfec-tées, il a été visualisé dans le noyauou le cytoplasme uniquement oudans les deux compartiments cellu-laires simultanément [51, 52]. Cesdiscordances de localisation ont sou-vent été attribuées aux techniquesd’immunomarquage qui nécessitentla fixation et la perméabilisation descellules, ainsi que l’utilisation d’anti-corps avec des spécificités relatives,chacune de ces étapes pouvantconduire à des artéfacts.C’est pourquoi différents groupesont entrepris d’étudier la dynamiquedes récepteurs stéroïdiens. Il a étédécrit une protéine de fusion entrela GFP et le GR (GFP-GR) [53, 54],entre la GFP et le AR (GFP-AR ) [55]et, plus récemment, une protéine defusion entre la GFP et le MR (GFP-MR) [56]. Chacune de ces construc-tions a été transfectée dans plusieurstypes cellulaires. Dans un premiertemps, la localisation cellulaire enl’absence d’hormone a été détermi-née puis la dynamique du récepteur– sous forme de cinétique en pré-sence d’agonistes, mais égalementd’antagonistes – a été analysée.
GFP-GR
C’est le groupe d’Ogawa qui le pre-mier a décrit une GFP-GR en fusion-
nant la GFP à un GR tronqué de ses131 premiers acides aminés [54].GFP-GR est transcriptionnellementactive. Cette protéine chimérique estcytoplasmique en l’absence d’hor-mone dans la lignée cellulaire COS-1,dans une lignée dérivée de tumeurépithéliale de carpe ou encore dansune culture primaire de cellules de larétine d’un embryon de poulet. GFP-GR rentre très rapidement dans lenoyau en présence de dexamétha-sone. La fluorescence est uniformé-ment distribuée dans le noyau maisest exclue des nucléoles. Enrevanche, Htun et al. observent queGFP-GR-C656G, en présence de dexa-méthasone, forme rapidement desregroupements dans le noyau [53].Cette GFP-GR-C656G est une pro-téine de fusion entre la GFP-S65T etune protéine GR mutée. Le rempla-cement de la cystéine 656 par une gly-cine dans le domaine de liaison del’hormone a pour effet d’augmenterl’affinité du récepteur pour la dexa-méthasone d’un facteur 9. A faibleconcentration de dexaméthasone, il ya activation de GFP-GR-C656G sansactivation du GR endogène. GFP-GR-C656G est exprimée dans une lignéecellulaire qui a intégré dans songénome plusieurs copies du gène rap-porteur MMTV-CAT. Ces regroupe-ments sont observés par microscopieconfocale et apparaissent comme despoints très fluorescents dans le noyau.De plus, l’antagoniste RU486 induitle transfert de GFP-GR-C656G dansle noyau mais ne conduit pas à la for-mation de ces points fluorescents.D’après les auteurs, ces regroupe-ments correspondraient donc à dessites potentiels transcriptionnelle-ment actifs [53]. Afin de vérifier laspécificité de ces points fluorescents,les auteurs ont construit une pro-téine de fusion entre la GFP et unGR mutant incapable de lier l’ADN.Cette GFP-GR-mutant conserve lacapacité de rentrer dans le noyaumais ne forme pas de regroupe-ments ; cela confirme l’hypothèse dela formation de ces regoupementssur des sites spécifiques d’ADN. Plusrécemment, ces auteurs ont analyséla distribution de GFP-GR-C656Gdans une autre lignée cellulaire(lignée 3134). Ces cellules contien-nent un large fragment d’ADN inté-gré dans le génome comportant plu-sieurs MMTV en tandem [57]. Dans
cette lignée cellulaire, un large sitefluorescent est observé par microsco-pie confocale, formant une structureen forme de ruban après reconstruc-tion tridimensionnelle. Ce rubanfluorescent caractériserait la fixationde nombreux GFP-GR sur le tandemde MMTV. Cependant, la résolutionde la microscopie confocale ne per-met pas la visualisation de l’ADNsous forme chromatinienne ; la visua-lisation de la fixation de protéinesfluorescentes sur l’ADN n’est doncpas envisageable par cette technique.On ne peut assurer le caractère signi-ficatif de ce ruban fluorescent quepar des méthodes indirectes. Cesauteurs ont également construit uneprotéine de fusion entre la GFP et unGR muté qui, cette fois, modifie laspécificité de reconnaissance de l’élé-ment de réponse au GR [57]. Cemutant reconnaît alors l’élément deréponse au ER. Transfectée dans lalignée 3134, cette nouvelle protéinechimère rentre dans le noyau aprèsincubation avec la dexaméthasonemais ne forme pas de regroupementsen forme de ruban. Cela confirmeune fois encore la spécificité de cespoints fluorescents. La formation decette structure révélerait la liaison deGFP-GR sur le tandem de MMTV.Ainsi, par l’intermédiaire de la GFP,il serait possible de visualiser, dansdes cellules vivantes, la liaison desrécepteurs stéroïdiens sur l’ADN. Ilreste toutefois à faire la preuvedirecte de l’interaction de GFP-GR-C656G sur ces MMTV.
GFP-MR
Une protéine de fusion entre l’EGFPet le MR de rat a été récemment rap-portée [56]. Contrairement à GFP-GR, GFP-MR est à la fois cytoplas-mique et nucléaire en l’absenced’hormone. Cette localisation cellu-laire a été observée dans trois lignéescellulaires différentes : cellules CHO,MDCK et CV-1. En présence d’aldo-stérone, le récepteur est nucléairedans toutes les cellules transfectées.Des études cinétiques ont alors étéréalisées et ont révélé qu’en dixminutes, la totalité du récepteur étaitnucléaire [56]. Après cinq minutesd’incubation apparaissent des pointsconcentrés fluorescents dans lenoyau. En présence d’antagonistes,tels que la spironolactone ou le
50 m/s n° 1, vol. 15, janvier 99
34243$1 045-055 2/05/02 17:34 Page 50

ZK91587, le transfert au noyau estfaible même à de fortes concentra-tions. Ces deux antihormones nonseulement n’induisent pas la forma-tion de points fluorescents dans lenoyau mais inversent leur formationinduite par l’aldostérone. Lesauteurs évaluent à 1 500-4 500 lenombre de points fluorescents parnoyau en présence d’agonistes.Même si ces structures sont retenuessur une préparation de matricenucléaire, il paraît difficile de penserque chaque cluster, soit plusieursGFP-MR, sont fixés sur son élémentde réponse [56]. En effet, contraire-ment à ce qui était réalisé dans le tra-vail de Htun et al. [53], les cellulesutilisées dans cette étude ne contien-nent dans leur génome que les sitesnaturels des gènes réglés par lesminéralocorticoïdes. Le nombre depoints fluorescents ne correspon-drait donc pas au nombre de gènescibles. Les auteurs retiennent l’hypo-thèse d’un regroupement des GFP-MR sur des régions de la chromatinequi dirigeraient ces récepteurs acti-vés vers leurs gènes cibles [56].
GFP-AR
Nous avons développé une étude dela protéine de fusion GFP-AR [55].Dans un premier temps, nous avonsvérifié que la protéine de fusionconservait les caractéristiques fonc-tionnelles de l’AR. Nous avons,notamment, mesuré sa constanted’affinité pour l’androgène non méta-bolisable (le R1881), ainsi que sacapacité d’induire une activité trans-criptionnelle sur un gène rapporteurluciférase sous la dépendance d’unpromoteur spécifique des andro-gènes. Nos résultats montrent que lafusion de la GFP sur le domaineamino-terminal de l’AR ne modifiepas ses caractéristiques fonctionnelles.Ces propriétés étant conservées, nousavons analysé par fluorescence laGFP-AR exprimée dans des cellulesCOS-7 transfectées transitoirement.Alors que la GFP est distribuée uni-formément dans la cellule entre lecytoplasme et le noyau, GFP-ARréside de manière prédominantedans le cytoplasme en absence d’hor-mone et devient nucléaire en pré-sence d’androgène (figure 4). Nousavons alors étudié la cinétique de ladynamique de l’AR en présence
d’androgènes. Les cellules sont incu-bées en présence de R1881 sous lemicroscope à épifluorescence. Cemicroscope est relié à une caméraCCD qui permet une capture et unstockage de l’image de la cellule.Nous observons et nous enregistronsle mouvement de la GFP-AR dans lamême cellule toutes les 15 minutesenviron. Nous avons observé que desenregistrements plus fréquents de lacellule ne diminuent pas la fluores-cence mais sont létales pour la cel-lule. Pour chaque image acquise,nous pouvons quantifier le pourcen-tage de la fluorescence nucléaire parrapport à la fluorescence totale. Nousavons démontré que l’AR, qui estcytoplamique en l’absence d’hor-mone, est rapidement transportédans le noyau. Ce phénomène estvisible après seulement quelquesminutes d’incubation ; il est completaprès une heure d’incubation en pré-sence de forte concentrationd’androgènes (R1881 10– 6 M). Cettedynamique intracellulaire de l’ARdépend de la concentration. Ce n’estpas la vitesse d’entrée du récepteurdans le noyau qui varie en fonctionde la concentration d’hormone maisla quantité de récepteurs importésdans le noyau. La quantité de récep-
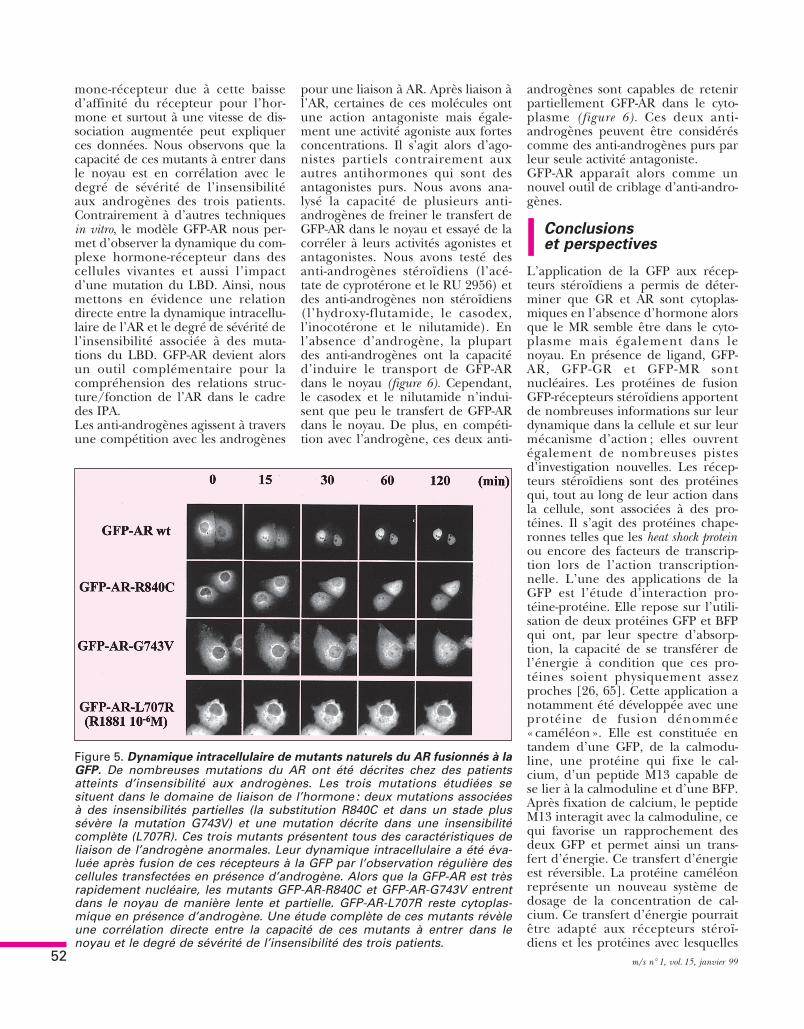
teurs importés dépend également del’affinité du récepteur pour le ligand.Ainsi, pour les trois principauxandrogènes testés, l’importance dutransfert nucléaire est formuléeainsi : R1881 > dihydrotestostérone >testostérone. Le transport nucléairede l’AR est donc également hor-mono-dépendant.Les androgènes, via l’AR, sont impli-qués dans la différenciation, le déve-loppement et le maintien des organesgénitaux internes et externes. Denombreuses mutations de l’AR ontété rapportées au cours d’insensibili-tés partielles aux androgènes (IPA) etd’insensibilités complètes aux andro-gènes (ICA) [58, 59]. Parmi lesdiverses mutations décrites dansnotre groupe [60, 61], nous avonsappliqué le modèle de la GFP-AR àtrois mutations. L’une d’entre elles,la substitution L707R est associée àune ICA [62] et deux autres muta-tions, R840C et dans une forme plussévère G743V sont associées à des IPA[63]. Ces trois mutations sont situéesdans le domaine de liaison de l’hor-mone du récepteur. Elles présententtoutes une altération de l’affinitépour R1881, allant jusqu’à l’absencetotale de liaison de l’hormone pourla mutation L707R. Cependant, unedifférence importante apparaît entreles mutants R840C et G743V : bienqu’ils présentent une diminutionsimilaire de l’affinité pour le R1881,l’augmentation de la vitesse de disso-ciation qu’ils induisent est nettementplus élevée pour G743V que pourR840C. Nous avons réalisé des ciné-tiques sur des cellules COS-7 transfec-tées par des vecteurs d’expressioncodant pour chacun de ces récep-teurs mutés. Les cellules sont alorsincubées en présence de 10– 6, 10– 7 ou10– 9M de dihydrotestostérone. LaGFP-AR-L707R est cytoplasmique enl’absence d’hormone et reste dans lecytoplasme en présence de DHT10– 6M. GFP-AR-R840C et GFP-AR-G743V entrent dans le noyau en pré-sence de 10– 6M de DHT, de manièrequasi identique au récepteur sauvage.En revanche, à une concentrationphysiologique (10– 9M), GFP-AR-R840C entre très lentement et partiel-lement dans le noyau alors que GFP-AR-G743V n’y est que faiblementtransporté même après 24 heuresd’incubation (figure 5) [63]. Une cer-taine instabilité du complexe hor-
51m/s n° 1, vol. 15, janvier 99
Figure 4. Localisation cellulaire de laGFP et de la protéine de fusion GFP-AR. La GFP et la protéine de fusionentre GFP et le récepteur des andro-gènes (GFP-AR) sont exprimées dansdes cellules COS-7 puis observées enmicroscopie à fluorescence. La GFPest distribuée uniformément danstoute la cellule et cette localisationn’est pas modifiée en présenced’androgène non métabolisable(R1881). GFP-AR, en l’absence d’hor-mone, est cytoplasmique et elle estnucléaire après addition de R1881.
34243$1 045-055 2/05/02 17:34 Page 51
mone-récepteur due à cette baissed’affinité du récepteur pour l’hor-mone et surtout à une vitesse de dis-sociation augmentée peut expliquerces données. Nous observons que lacapacité de ces mutants à entrer dansle noyau est en corrélation avec ledegré de sévérité de l’insensibilitéaux androgènes des trois patients.Contrairement à d’autres techniquesin vitro, le modèle GFP-AR nous per-met d’observer la dynamique du com-plexe hormone-récepteur dans descellules vivantes et aussi l’impactd’une mutation du LBD. Ainsi, nousmettons en évidence une relationdirecte entre la dynamique intracellu-laire de l’AR et le degré de sévérité del’insensibilité associée à des muta-tions du LBD. GFP-AR devient alorsun outil complémentaire pour lacompréhension des relations struc-ture/fonction de l’AR dans le cadredes IPA.Les anti-androgènes agissent à traversune compétition avec les androgènes
pour une liaison à AR. Après liaison àl’AR, certaines de ces molécules ontune action antagoniste mais égale-ment une activité agoniste aux fortesconcentrations. Il s’agit alors d’ago-nistes partiels contrairement auxautres antihormones qui sont desantagonistes purs. Nous avons ana-lysé la capacité de plusieurs anti-androgènes de freiner le transfert deGFP-AR dans le noyau et essayé de lacorréler à leurs activités agonistes etantagonistes. Nous avons testé desanti-androgènes stéroïdiens (l’acé-tate de cyprotérone et le RU 2956) etdes anti-androgènes non stéroïdiens(l’hydroxy-flutamide, le casodex,l’inocotérone et le nilutamide). Enl’absence d’androgène, la plupartdes anti-androgènes ont la capacitéd’induire le transport de GFP-ARdans le noyau (figure 6). Cependant,le casodex et le nilutamide n’indui-sent que peu le transfert de GFP-ARdans le noyau. De plus, en compéti-tion avec l’androgène, ces deux anti-
androgènes sont capables de retenirpartiellement GFP-AR dans le cyto-plasme (figure 6). Ces deux anti-androgènes peuvent être considéréscomme des anti-androgènes purs parleur seule activité antagoniste.GFP-AR apparaît alors comme unnouvel outil de criblage d’anti-andro-gènes.
Conclusionset perspectives
L’application de la GFP aux récep-teurs stéroïdiens a permis de déter-miner que GR et AR sont cytoplas-miques en l’absence d’hormone alorsque le MR semble être dans le cyto-plasme mais également dans lenoyau. En présence de ligand, GFP-AR, GFP-GR et GFP-MR sontnucléaires. Les protéines de fusionGFP-récepteurs stéroïdiens apportentde nombreuses informations sur leurdynamique dans la cellule et sur leurmécanisme d’action ; elles ouvrentégalement de nombreuses pistesd’investigation nouvelles. Les récep-teurs stéroïdiens sont des protéinesqui, tout au long de leur action dansla cellule, sont associées à des pro-téines. Il s’agit des protéines chape-ronnes telles que les heat shock proteinou encore des facteurs de transcrip-tion lors de l’action transcription-nelle. L’une des applications de laGFP est l’étude d’interaction pro-téine-protéine. Elle repose sur l’utili-sation de deux protéines GFP et BFPqui ont, par leur spectre d’absorp-tion, la capacité de se transférer del’énergie à condition que ces pro-téines soient physiquement assezproches [26, 65]. Cette application anotamment été développée avec uneprotéine de fusion dénommée« caméléon ». Elle est constituée entandem d’une GFP, de la calmodu-line, une protéine qui fixe le cal-cium, d’un peptide M13 capable dese lier à la calmoduline et d’une BFP.Après fixation de calcium, le peptideM13 interagit avec la calmoduline, cequi favorise un rapprochement desdeux GFP et permet ainsi un trans-fert d’énergie. Ce transfert d’énergieest réversible. La protéine caméléonreprésente un nouveau système dedosage de la concentration de cal-cium. Ce transfert d’énergie pourraitêtre adapté aux récepteurs stéroï-diens et les protéines avec lesquelles
52 m/s n° 1, vol. 15, janvier 99
Figure 5. Dynamique intracellulaire de mutants naturels du AR fusionnés à laGFP. De nombreuses mutations du AR ont été décrites chez des patientsatteints d’insensibilité aux androgènes. Les trois mutations étudiées sesituent dans le domaine de liaison de l’hormone : deux mutations associéesà des insensibilités partielles (la substitution R840C et dans un stade plussévère la mutation G743V) et une mutation décrite dans une insensibilitécomplète (L707R). Ces trois mutants présentent tous des caractéristiques deliaison de l’androgène anormales. Leur dynamique intracellulaire a été éva-luée après fusion de ces récepteurs à la GFP par l’observation régulière descellules transfectées en présence d’androgène. Alors que la GFP-AR est trèsrapidement nucléaire, les mutants GFP-AR-R840C et GFP-AR-G743V entrentdans le noyau de manière lente et partielle. GFP-AR-L707R reste cytoplas-mique en présence d’androgène. Une étude complète de ces mutants révèleune corrélation directe entre la capacité de ces mutants à entrer dans lenoyau et le degré de sévérité de l’insensibilité des trois patients.
34243$1 045-055 2/05/02 17:34 Page 52

ils interagissent en construisant parexemple des protéines chimèresGFP-récepteurs stéroïdiens et BFP-heat shock protein afin de visualiser lesinteractions entre ces protéines insitu dans la cellule. Day et al. ont toutrécemment décrit la visualisation dela formation d’un homodimère dufacteur de transcription Pit-1 parFRET entre GFP- et BFP-Pit1 [66].De plus, l’un des moyens d’étude parla GFP est le photoblanchiment ouinversement la surfluorescence d’unerégion de la cellule. Cette méthodeest aujourd’hui utilisée pour analyserle mouvement de protéines fluores-centes au sein de la cellule. Le pho-toblanchiment est abordé selon deuxapproches : sous le nom de FLIP(fluorescence loss in photobleach) et sousle nom de FRAP (fluorescence recoveryafter photobleach). L’une (FLIP)consiste à éblouir par un laser unpoint de la cellule et de suivre la dis-parition progressive de fluorescencedans toute la cellule, qui révèle lepassage des molécules fluorescentesdans la zone d’irradiation du laser.Le FLIP est déjà utilisé pour des pro-téines du réticulum endoplasmiqueet de l’appareil de Golgi [67].L’autre (FRAP) consiste à blanchirune zone de la cellule et à observer laréapparition de la fluorescence danscette zone qui est due à la migrationde protéines fluorescentes. La sur-fluorescence consiste à illuminerdans un premier temps en UV, ce quimodifie les propriétés de la GFP et
entraîne une augmentation de lafluorescence lors de l’excitation à470 nm. Cette surfluorescence per-met d’étudier la diffusion de la pro-téine marquée en mesurant, soit ladisparition au point d’irradiation,soit l’apparition de la fluorescencedans des zones non irradiées par lelaser UV [14].La vitesse de diffusion (ou coefficientde diffusion) d’une protéine peutêtre également évaluée par la mesurede corrélation de fluorescence. Ildevrait être possible de déterminer lataille du complexe formé contenantla protéine fluorescente et ainsid’étudier les interactions avecd’autres constituants de la cellule.Ces techniques apparaîssent donccomme des nouveaux moyensd’étude de la dynamique des récep-teurs stéroïdiens dans le cytoplasmeet dans le noyau et permettent devisualiser dans une cellule vivante desmodifications de l’environnementcellulaire et des interactions entremacromolécules ■
RÉFÉRENCES1. Prasher DC, Eckenrode VK, Ward WW,Prendergast FG, Cormier MJ. Primary struc-ture of the Aequorea victoria green-fluores-cent protein. Gene 1992 ; 111 : 229-33.
2. Chalfie M, Tu Y, Euskirchen G, WardWW, Prasher DC. Green fluorescent proteinas a marker for gene expression. Science1994 ; 263 : 802-5.
3. Gubin AN, Reddy B, Njoroge JM, MillerJL. Long-term, stable expression of greenfluorescent protein in mammalian cells. Bio-chem Biophys Res Commun 1997 ; 236 : 347-50.
4. Wang S, Hazelrigg T. Implications forbcd mRNA localization from spatial distri-bution of exu protein in Drosophila ooge-nesis. Nature 1994 ; 369 : 400-3.
5. Prasher DC. Using GFP to see the light.Trends Genet 1995 ; 11 : 320-3.
6. Kojima S, Hirano T, Niwa H, Ohashi M,Inouye S, Tsuji FL. Mechanism of the redoxreaction of the Aequorea green fluorescentprotein (GFP). Tetrahedron Lett 1997 ; 38 :2875-8.
7. Heim R, Prasher DC, Tsien RY. Wave-length mutations and posttranslationalautoxidation of green fluorescent protein.Proc Natl Acad Sci USA 1994 ; 91 : 12501-4.
8. Li XQ, Zhang GH, Ngo N, Zhao XN,Kain SR, Huang CC. Deletions of the Aequo-rea victoria green fluorescent protein definethe minimal domain required for fluores-cence. J Biol Chem 1997 ; 272 : 28545-9.
9. Cody CW, Prasher DC, Westler WM,Prendergast FG, Ward WW. Chemical struc-ture of the hexapeptide chromophore ofthe Aequorea green-fluorescent protein.Biochemistry 1993 ; 32 : 1212-8.
10. Örmo M, Cubitt AB, Kallio K, Gross LA,Tsien RY, Remington SJ. Crystal structure ofthe Aequorea victoria green fluorescent pro-tein. Science 1996 ; 273 : 1392-5.
11. Yang FY, Moss LG, N PG. The molecularstructure of green fluorescent protein. NatBiotechnol 1996 ; 14 : 1246-51.
12. Chattoraj M, King BA, Bublitz GU,Boxer SG. Ultra-fast excited state dynamicsin green fluorescent protein : multiple statesand proton transfer. Proc Natl Acad Sci USA1996 ; 93 : 8362-7.
13. Niwa H, Inouye S, Hirano T, et al. Che-mical nature of the light emitter of theAequorea green fluorescent protein. ProcNatl Acad Sci USA 1996 ; 93 : 13617-22.
14. Yokoe H, Meyer T. Spatial dynamics ofGFP-tagged proteins investigated by localfluorescence enhancement. Nat Biotechnol1996 ; 14 : 1252-6.
15. Brejc K, Sixma TK, Kitts PA, et al. Struc-tural basis for dual excitation and photoiso-merization of the Aequorea victoria greenfluorescent protein. Proc Natl Acad Sci USA1997 ; 94 : 2306-11.
16. Heim R, Cubitt AB, Tsien RY. Improvedgreen fluorescence. Nature 1995 ; 373 : 663-4.
53m/s n° 1, vol. 15, janvier 99
Figure 6. Effet des anti-androgènes sur la localisation cellulaire du AR. Lesanti-androgènes, largement utilisés en clinique, sont classés comme ago-nistes partiels ou purs antagonistes. Ont été testés des anti-androgènes sté-roïdiens (acétate de cyprotérone et RU2956) et des anti-androgènes non sté-roïdiens (inocotérone, hydroxy-flutamide, casodex et nilutamide). Leur effetsur la localisation cellulaire d’AR est évalué par l’incubation des cellules enprésence de chaque anti-androgène seul ou en compétition avec le R1881.En l’absence d’androgène, la plupart des anti-androgènes testés induisent untransfert d’AR dans le noyau. Le casodex et le nilutamide ont la capacité deretenir partiellement GFP-AR dans le cytoplasme.
Remerciements
Nous remercions les Drs Béatrice Térouanneet Paul Mangeat pour leur aide scientifique ettechnique. Une partie de ce travail a été soute-nue par l’ARC (V.G. : boursière de l’ARC) etpar un contrat CEE Biomed.
34243$1 045-055 2/05/02 17:34 Page 53

RÉFÉRENCES17. Pines J. GFP in mammalian cells. TrendsGenet 1995 ; 11 : 326-7.
18. Crameri A, Whitehorn EA, Tate E, Stem-mer PC. Improved GFP by molecular evolu-tion using DNA shuffling. Nat Biotechnol1996 ; 14 : 315-9.
19. Zhang G, Gurtu V, Kain SR. An enhan-ced green fluorescent protein allows sensi-tive detection of gene transfer in mamma-lian cells. Biochem Biophys Res Commun 1996 ;227 : 707-11.
20. Cormack B, Valdivia R, Falkow S. FACS-optimized mutants of the green fluorescentprotein (GFP). Gene 1996 ; 173 : 33-8.
21. Kimata Y, Iwaki M, Lim CR, Kohno K. Anovel mutation which enhances the fluores-cence of green fluorescent protein at hightemperatures. Biochem Biophys Res Commun1997 ; 232 : 69-73.
22. Patterson GH, Knobel SM, Sharif WD,Kain SR, Piston DW. Use of the green fluo-rescent protein and its mutants in quantita-tive fluorescence microscopy. Biophys J1997 ; 73 : 2782-90.
23. Llopis J, McCaffery JM, Miyawaki A, Far-quhar MG, Tsien RY. Measurement of cyto-solic, mitochondrial, and Golgi pH in singleliving cells with green fluorescent proteins.Proc Natl Acad Sci USA 1998 ; 95 : 6803-8.
24. Andersen JB, Sternberg C, Poulsen LK,Bjorn SP, Givskov M, Molin S. New unstablevariants of green fluorescent protein for stu-dies of transient gene expression in bacte-ria. Appl Environ Microbiol 1998 ; 64 : 2240-6.
25. Wachter RM, King BA, Heim R, et al.Crystal structure and photodynamic beha-vior of the blue emission variantY66H/Y145F of green fluorescent protein.Biochemistry 1997 ; 36 : 9759-65.
26. Heim R, Tsien R. Engineering greenfluorescent protein for improved bright-ness, longer wavelengths and fluorescenceresonnance energy transfer. Curr Biol 1996 ;6 : 178-82.
27. Yang TT, Sinai P, Green G, et al. Impro-ved fluorescence and dual color detectionwith enhanced blue and green variants ofthe green fluorescent protein. J Biol Chem1998 ; 273 : 8212-6.
28. Gervaix A, West D, Leoni LM, RichmanDD, Wong-Staal, Corbeil J. A new reportercell line to monitor HIV infection and drugsusceptibility in vitro. Proc Natl Acad Sci USA1997 ; 94 : 4653-8.
29. Peel AL, Zolotukhin S, Schrimsher GW,Muzyczka N, Reier PJ. Efficient transduc-tion of green fluorescent protein in spinalcord neurons using adeno-associated virusvectors containing cell type-specific promo-ters. Gene Ther 1997 ; 4 : 16-24.
30. Ropp JD, Donahue CJ, Wolfgang-Kim-ball D, et al. Aequorea green fluorescent pro-tein : simultaneous analysis of wild-type andblue-fluorescing mutant by flow cytometry.Cytometry 1996 ; 24 : 284-8.
31. Anderson MT, Tjioe IM, Lorincz MC, etal. Simultaneous fluorescence-activated cellsorter analysis of two distinct transcriptionalelemnts within a single cell using enginee-red green fluorescent proteins. Proc NatlAcad Sci USA 1996 ; 93 : 8508-11.
32. Okabe M, Ikawa M, Kominami K, Naka-nishi T, Nishimune Y. « Green mice » as asource of ubiquitous green cells. FEBS Lett1997 ; 407 : 313-9.
33. Chishima T, Miyagi Y, Wang XE, et al.Cancer invasion and micrometastasis visuali-zed in live tissue by green fluorescent pro-tein expression. Cancer Res 1997 ; 57 : 2042-7.
34. Rizzuto R, Brini M, Pizzo P, Murgia M,Pozzan T. Chimeric green fluorescent pro-tein as a tool for visualizing subcellularorganelles in living cells. Curr Biol 1995 ; 5 :635-42.
35. Rizzuto R, Brini M, De Giorgi F, et al.Double labelling of subcellular structureswith organelle-targeted GFP mutants invivo. Curr Biol 1996 ; 6 : 183-8.
36. Olson KR, Mcintosh JR, Olmsted JB.Analysis of MAP 4 function in living cellsusing green fluorescent protein (GFP) chi-meras. J Cell Biol 1995 ; 130 : 639-50.
37. Naray-Fejes-Toth A, Fejes-Toth G. Sub-cellular localization of the type 2 11 β-hydroxysteroid dehydrogenase. J Biol Chem1996 ; 271 : 15436-42.
38. Cole N, Smith C, Sciaky N, Teresaki M,Edidin M, Lippincott-Schwartz J. Diffusionalmobility of Golgi proteins in membranes ofliving cells. Science 1996 ; 273 : 797-801.
39. Yano M, Kanazawa M, Terada K, et al.Visualization of mitochondrial proteinimport in cultured mammalian cells withgreen fluorescent protein and effects of ove-rexpression of the human import receptorTom20. J Biol Chem 1997 ; 272 : 8459-65.
40. Tarasova NI, Stauber RH, Choi JK, et al.Visualization of G protein-coupled recetortrafficking with the aid of the green fluores-cent protein. J Biol Chem 1997 ; 272 : 14817-24.
41. Evans RM. The steroid and thyroid hor-mone receptor superfamily. Science 1988 ;240 : 889-95.
42. Mangelsdorf D, Thummel C, Beato M, etal. The nuclear receptor superfamily : thesecond decade. Cell 1995 ; 83 : 835-9.
43. Brink M, Humbel BM, De Kloet ER, VanDriel R. The unliganded glucocorticoidreceptor is localized in the nucleus, not inthe cytoplasm. Endocrinology 1992 ; 130 :3575-81.
44. Guiochon-Mantel A, Delabre K, LescopP, Milgrom E. Intracellular traffic of steroidhormone receptors. J Steroid Biochem MolBiol 1996 ; 56 : 1-6.
45. King WJ, Greene GL. Monolonal antibo-dies localize oestrogene receptor in thenuclei of target cells. Nature 1984 ; 307 : 745-7.
46. Perrot-Applanat M, Logeat F, Groyer-Picard M, Milgrom E. Immunocytochemicalstudy of mammalian progesterone receptorusing monoclonal antibodies. Endocrinology1985 ; 116 : 1473-84.
47. Guiochon-Mantel A, Lescop P, Christin-Maitre S, Loosfelt H, Perrot-Applanat M,Milgrom E. Nucleocytoplasmic shuttling ofthe progesterone receptor. EMBO J 1991 ;10 : 3851-9.
48. Lombes M, Binart N, Delahaye F, Bau-lieu EE, Rafestinoblin ME. Differentialintracellular localization of human minera-locorticosteroid receptor on binding of ago-nists and antagonists. Biochem J 1994 ; 302 :191-7.
49. Husmann DA, McPhaul MJ. Localizationof the androgen receptor in the developingrat gubernaculum. Endocrinology 1991 ; 128 :383-7.
50. Sar M, Lubahn DB, French FS, WilsonEM. Immunohistochemical localization ofthe androgen receptor in rat and humantissues. Endocrinology 1990 ; 127 : 3180-6.
51. Jenster G, Trapman J, Brinkmann AO.Nuclear import of the human androgenreceptor. Biochem J 1993 ; 293 : 761-8.
52. Simental JA, Sar M, Lane MV, FrenchFS, Wilson EM. Transcriptional activationand nuclear targeting signals of the humanandrogen receptor. J Biol Chem 1991 ; 266 :510-8.
53. Htun H, Barsony J, Renyi I, Gould DL,Hager GL. Visualization of glucocorticoidreceptor translocation and intranuclearorganization in living cells with a greenfluorescent chimera. Proc Natl Acad Sci USA1996 ; 93 : 4845-50.
54. Ogawa H, Inouye S, Tsuji FI, Yasuda K,Umesono K. Localization, trafficking, andtemperature-dependence of the aequoreagreen fluorescent protein in cultured verte-brate cells. Proc Natl Acad Sci USA 1995 ; 92 :11899-903.
55. Georget V, Lobaccaro J, Térouanne B,Mangeat P, Nicolas J, Sultan C. Traffickingof the androgen receptor in living cells withfused green fluorescent protein-androgenreceptor. Mol Cell Endocrinol 1997 ; 129 : 17-26.
56. FejesToth G, Pearce D, NarayFejesTothA. Subcellular localization of mineralocorti-coid receptors in living cells : effects ofreceptor agonists and antagonists. Proc NatlAcad Sci USA 1998 ; 95 : 2973-8.
57. Htun H, Walker D, Hager GL. Opticalimaging approach to study protein-DNAinteraction and nuclear organization in cul-tured living cells. Proc 10th Convers BiomolStereodyn 1999 (sous presse).
58. Quigley CA, Debellis A, Marschke KB,Elawady MK, Wilson EM, French FS. Andro-gen receptor defects : historical, clinical,and molecular perspectives. Endocrine Rev1995 ; 16 : 271-321.
54 m/s n° 1, vol. 15, janvier 99
34243$1 045-055 2/05/02 17:34 Page 54

RÉFÉRENCES59. Gottlieb B, Lehvaslaiho H, Beitel LK,Lumbroso R, Pinsky L, Trifiro M. Theandrogen receptor gene mutations data-base. Nucleic Acids Res 1998 ; 26 : 234-8.
60. Sultan C, Lumbroso S, Poujol N, BelonC, Boudon C, Lobaccaro JM. Mutations ofthe androgen receptor gene in androgeninsensitivity syndrome. J Steroid Biochem MolBiol 1993 ; 46 : 519-30.
61. Sultan C, Savage MO. Intersex states. In :Grossman A, ed. Clinical endocrinology.Londres : Blackwell Science, 1998 : 795-809.
62. Lumbroso S, Lobaccaro JM, Georget V,et al. A novel substitution (Leu707Arg) inexon 4 of the androgen receptor genecauses complete androgen resistance. J ClinEndocrinol Metab 1996 ; 81 : 1984-8.
63. Georget V, Terouanne B, Lumbroso S,Nicolas J, Sultan C. Trafficking of theandrogen receptor mutants fused to thegreen fluorescent protein : a new investiga-
tion of partial androgen insensitivity syn-drome. J Clin Endocrinol Metab 1998 ; inpress.
64. Gerdes MJ, Dang TD, Larsen M, RowleyDR. Transforming growth factor-beta 1induces nuclear to cytoplasmic distributionof androgen receptor and inhibits andro-gen response in prostate smooth musclecells. Endocrinology 1998 ; 139 : 3569-77.
65. Miyawaki A, Llopis J, Heim R, et al. Fluo-rescent indicators for Ca2+ based on greenfluorescent proteins and calmodulin. Nature1997 ; 388 : 882-7.
66. Day RN. Visualization of Pit-1 transcrip-tion factor interactions in the living cellnucleus by fluorescence resonance energytransfer microscopy. Mol Endocrinol 1998 ;12 : 1410-9.
67. Lippincott-Schwartz J, Smith CL. Insightsinto secretory and endocytic membrane traf-fic using green fluorescent protein chime-ras. Curr Opin Neurobiol 1997 ; 7 : 631-9.
55m/s n° 1, vol. 15, janvier 99
SummaryGreen fluorescent protein :its application to the studyof intracellular dynamicsof steroid receptors
The green fluorescent protein(GFP) comes from the Aequoreavictoria jellyfish. In vivo, it acquiresthe capacity of fluorescence byactivation of aequorin. In vitro, it isactivated by simple exposure tostandard long-wave ultraviolet orblue light. GFP does not requirethe addition of substrate or cofac-tors for generation of its greenlight and it represents a new toolfor monitoring the dynamic pro-cesses in living cells and organisms.It can produce fluorescence invarious organisms, such as bacte-ria, plants and mammals. GFP isnow applied as a reporter for geneexpression, as a marker to studycell lineage and as a tag to localizeproteins in living cells. GFP doesnot require cell fixation or otherchemical steps for analysis,contrary to immunocytochemistry.Immunodetection needs fixationand permeabilization steps, whichmay lead to artefacts in the patternof localization. This would explainparticularly the contradictoryresults about the localization of thesteroid receptors. Several studieshave thus been developed usingGFP tagged to different steroidreceptors : glucocorticoid receptor(GR), mineralocorticoid receptor(MR) and androgen receptor(AR). In absence of hormone,GFP-GR and GFP-AR were descri-bed to be cytoplasmic, while theGFP-MR was both cytoplasmic andnuclear. In presence of each speci-fic ligand, GFP-steroid receptorswere all nuclear. Formation ofnuclear clusters were observed inthe presence of agonists for GFP-GR and GFP-MR. GFP permits theanalysis of dynamic cellular eventsand has already been etablished asan important tool for manybranches of biological research.
TIRÉS À PARTC. Sultan.
B. Malissen : Molecular interaction in first signal.T. Strom : Manipulation of allo-recognition through T-cell growth factor.J. Thomas : Tolerance induction using anti-CD3 immunotoxine antibodies.L. Chatenoud : Immuno-intervention through lymphocyte receptor targeting.R. Zhong : Prevention rejection and induction of tolerance by monoclonal
antibodies against CD45RB.J. Bluestone : Second signals in allo and xeno-recognition.D. Latinne : Immuno-intervention through CD2/LFA3 inhibition.T.C. Pearson : Manipulation of B7/CD28-CTL44 in primates.S. Knechtle : Inhibition of CD40 L pathway.A. Wörn : Engineering of scFv fragments for extracellular and intracellular
applications.B. Vanhove : ScFv and intra-cellular bioreagents.R. Dunbar : HLA tetramers.G. Grassy : Computer-assisted rational design of immunosuppressive
peptides.N. Suciu-Foca : Altered peptides in vivo.F. Sanfilippo : Inhibition of complement in vivo.J.S. Pober : Targeting second signals provided by vascular endothelial cells.
Renseignements et formulaires d’Abstracts :NAT Secrétariat ITERT, CHU Hôtel-Dieu, 30, boulevard Jean-Monnet,44093 Nantes (France).Fax : (33) 2 40 08 74 11.
Inscriptions :1 900FF (déjeuners et dîners compris).Date limite de remise des abstracts : 1er avril 1999.
Sixième NAT (Nantes-Actualités-Transplantation)10-11 juin 1999 – Nantes (France)
Targeting Recipient ImmuneResponse through Bioreagents
34243$1 045-055 2/05/02 17:34 Page 55








![LINE CONSTRUCTION FOREST EVENT COLORS PRO TECH...MARQUAGE HAUTEMENT FLUORESCENT [12 MOIS] FLASH MARKER,est un traceur forestier hautement fluorescent. Sa formulation à base de cires,](https://img.pdfslide.fr/doc/110x75/614a37b612c9616cbc6945fd/line-construction-forest-event-colors-pro-tech-marquage-hautement-fluorescent.jpg)













![[ZX] [méca 1.9D] Le joint de culasse de KaraPitchou, saison 2](https://img.pdfslide.fr/doc/110x75/629e562936790f2f1200a707/zx-mca-19d-le-joint-de-culasse-de-karapitchou-saison-2.jpg)






