Embed Size (px)
Citation preview

This article was downloaded by: [University of New Hampshire]On: 05 October 2014, At: 09:24Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH,UK
Acta Botanica Gallica: BotanyLettersPublication details, including instructions forauthors and subscription information:http://www.tandfonline.com/loi/tabg20
Les composés phénoliques etla caractérisation des niveauxtaxonomiques inférieurs dumonde végétalMaurice Jay a , Joël Reynaud a , Olivier Raymond a &Cédric Grossi aa Laboratoire de Biologie micromoléculaire etPhytochimie , Université Claude Bernard Lyon-I , 43Bd du 11 novembre 1918, F-69622 , VilleurbannePublished online: 27 Apr 2013.
To cite this article: Maurice Jay , Joël Reynaud , Olivier Raymond & Cédric Grossi(1996) Les composés phénoliques et la caractérisation des niveaux taxonomiquesinférieurs du monde végétal, Acta Botanica Gallica: Botany Letters, 143:6, 481-491,DOI: 10.1080/12538078.1996.10515345
To link to this article: http://dx.doi.org/10.1080/12538078.1996.10515345
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all theinformation (the “Content”) contained in the publications on our platform.However, Taylor & Francis, our agents, and our licensors make norepresentations or warranties whatsoever as to the accuracy, completeness,or suitability for any purpose of the Content. Any opinions and viewsexpressed in this publication are the opinions and views of the authors, andare not the views of or endorsed by Taylor & Francis. The accuracy of theContent should not be relied upon and should be independently verified withprimary sources of information. Taylor and Francis shall not be liable for anylosses, actions, claims, proceedings, demands, costs, expenses, damages,and other liabilities whatsoever or howsoever caused arising directly orindirectly in connection with, in relation to or arising out of the use of theContent.

This article may be used for research, teaching, and private study purposes.Any substantial or systematic reproduction, redistribution, reselling, loan,sub-licensing, systematic supply, or distribution in any form to anyone isexpressly forbidden. Terms & Conditions of access and use can be found athttp://www.tandfonline.com/page/terms-and-conditions
Dow
nloa
ded
by [
Uni
vers
ity o
f N
ew H
amps
hire
] at
09:
24 0
5 O
ctob
er 2
014

Acta bot. Gallica, 1996, 143 (6), 481-491.
Les composes phenoliques et Ia caracterisation des niveaux taxonomiques inferieurs du monde vegetal
par Maurice Jay, Joel Reynaud, Olivier Raymond et Cedric Grossi
Laboratoire de Biologie micromoteculaire et Phytochimie, Universite Claude Bernard Lyon-1, 43 Bd du 11 novembre 1918, F-69622 Villeurbanne
INTRODUCTION
Resume.- Les marqueurs chimiques auxquels appartiennent les composes phenoliques, ant ete largement utilises dans les etudes a caractere systematique ; les niveaux taxonomiques concernes ant ete les niveaux superieurs d'organisation : ordre, famille, genre. Par suite du developpement de nouvelles methodes analytiques alliant sensibilite et automatisme, l'etude d'un patron metabolique (phenolique) a l'echelle de l'individu est devenue possible: ainsi est nee Ia Biologie micromoleculaire qui s'interesse a Ia structure et au fonctionnement des populations vegetales. Trois examples illustreront ce propos : Dactylis glomerafa, Rosa x hybrida, Lotus comiculatus.
Summary.- The chemical markers such as phenolic compounds, have been extensively used in botanical chemosystematic studies ; these last mainly concerned the high taxonomic levels : order, family, genus. In consequence of new more sensitive and automatic analytical procedures, it is possible to approach the metabolic (phenolic) profile of individual genotypes : so the Micromolecular Biology is born, concerned with the organisation and functioning of plant populations. Three examples will give support to this proposal : Dactylis glomerata, Rosa x hybrida, Lotus corniculatus.
Key words : phenolic metabolism- plant population biology.
II est d'une pratique classique en Biologie de Populations de mesurer Ia diversite genetique a I' aide des proteines enzymatiques et de Ia technique d' electrophorese (Ducousso, 1985 ; HaywardetMcAdam, 1977 ;Lumaret 1981); cette pratique releve de I' etude du polymorphisme isozymique et conduit a interpreter Ia
structure genetique d'un groupe (population, metapopulation, espece) vegetal sur Ia base des frequences des divers electromorphes pour chacune des activites enzymatiques eprouvees. C' est une voie tres actuelle de prospection des ressources genetiques, particulierement exploitee pour Ies especes dont certaines formes ont connu un developpement agronomique.
©Societe botanique de France 1996. ISSN 1253-8078.
Dow
nloa
ded
by [
Uni
vers
ity o
f N
ew H
amps
hire
] at
09:
24 0
5 O
ctob
er 2
014

482 ACTA BOTANICA GALLICA
A cote de cette demarche, et de maniere plus recente, certains biologistes ont realise que dans le monde vegetal, d'autres marqueurs metaboliques pouvaient etre utilises dans le but de mieux cerner Ia structuration taxonomique et de lui donner un sens fonctionnel (Ardouin, 1985 ; Boyet 1985). Ces marqueurs nouveaux qui ne relevent plus du metabolisme primaire comme c'est le cas pour Ies proteines, vont concerner un metabolisme qui constitue une originalite des vegetaux : le metabolisme secondaire.
II faut entendre par Metabolisme Secondaire uncertain nombre de voies s'articulant sur des molecules charnieres banales telles que acetate, shikimate, mevalonate, etc. De ces molecules naitront trois grandes families biogenetiques : phenols, terpenes, alcaloldes, plus un certain nombre d'autres families plus petites: glucosinolates, glycosides cyanogenetiques, betalaines, etc. Chacune de ces voies ouvre, grace a des reactions sequentielles de types conjugaison ou substitution, sur des centaines de molecules differentes (Ia plupart monomeres), c'est-a-dire sur un registre de diversite extraordinaire. Correlativement, chaque molecule est tres faiblement representee en terme de teneurs, ce qui leur a valu Ie label de micromolecules, et celui de biologie (ou polymorphisme) micromoleculaire pour leur mise en oeuvre comme marqueurs en Biologie des Populations (Jay, 1988).
A vrai dire le terme de marqueur au sens strict est impropre dans notre acceptation de Ia biologie micromoleculaire, et il est bien preferable de parler de marquage metabolique dans Ia mesure ou les molecules d'une voie etudiee sont biogenetiquement reliees ; dans ces conditions nous accordons plus d'importance a l'equilibre entre ces molecules qui est regi par des regulations metaboliques genetiquement fixees, qu'a !'existence d'une molecule particuliere. Ceci se traduit par I' obtention de cartes metaboliques propres a chaque individu et par
Ia comparaison des individus deux a deux au niveau de leur profil moleculaire norme encore appele empreinte individuelle pour telle ou telle voie metabolique (Boyet et Jay, 1989 ; Lauranson et a/., 1995). Les developpements d'un tel concept sont maintenant nombreux en particulier avec Ia voie metabolique des phenols, sachant que les applications peuvent etre classees en deux periodes:
- les premieres applications s'appuyant essentiellement sur Ies methodes de chromatographie papier et couches minces, remontent aux travaux de Bate-Smith ( 1956, 1962, 1965) et a ceux des auteurs qui de 1956 a 1980 ont oeuvre en chimiotaxonomie (Lebreton, 1962 ; Jay, 1970 ; Voirin et Jay, 1978),
- les applications ulterieures reposant sur une techno Iogie automatisee sur base chromatographie liquide haute pression, ont concerne aussi bien Ia caracterisation varietale (Biolley eta/., 1994 ; Jay et Ferrero, 1989; Ledeme et Jay, 1987) que !'etude des populations vegetales (Gonnet, 1993) et celle de Ia structure fonctionnelle ou evolutive des especes (Jay et al., 1994)
Dans Ie cadre de ces dernieres applications, les regulations metaboliques responsables de I' empreinte observee ont ete exploitees pour leur signification fonctionnelle ; en effet, Ia diversite mesuree a !'aide du metabolisme secondaire, phenolique en particulier, est I' expression de Ia reponse du vegetal aux pressions de selection du milieu qui le porte: c'est ainsi que Ia synthese de pigments floraux est en relation avec les pollinisateurs, celle de tannins en relation avec les predateurs, celles de phytoalexines en relation avec les pathogenes. Et dans ce sens, I' empreinte metabolique que livre l'individu peut d'une certaine fa\=on etre consideree comme une image hautement integree de sa « microniche ».
C' est so us cet angle que no us presenterons quelques resultats de nos recents travaux.
Dow
nloa
ded
by [
Uni
vers
ity o
f N
ew H
amps
hire
] at
09:
24 0
5 O
ctob
er 2
014

M. JAY ET AL. 483
RESULTATS ET DISCUSSION
1 - Les marqueurs phenoliques et Ia structure taxonomique do Dactylis glomera/a (Poaceae)
Dactylis glome rata est une espece botaniquement interessante puisque cette graminee perenne allogame existe sous Ia forme d'un complexe polyploide, avec un cytotype diplo"ide represente par 14 sous-especes et un cytotype tetraploide illustre par 3 sous-especes (Tableau I). En outre ces diverses sous-especes se reconnaissent sous trois types climatiques : tempere, mediterraneen et subtropical.
La collection d'individus a conceme une centaine de populations echantillonnees sur toute I' aire de I' espece, et representees chacune par 5 a 50 individus selon les cas. L'analyse du metabolisme phenolique a porte sur le materiel foliaire d'une repousse d'automne dans Ies conditions du Jardin Experimental du Centre CNRS de Montpellier. Les resultats presentes sont ex traits de Ardouin ( 1985), Ardouin et al. ( 1985, 1988), Fiasson et al. ( 1986, 1987).
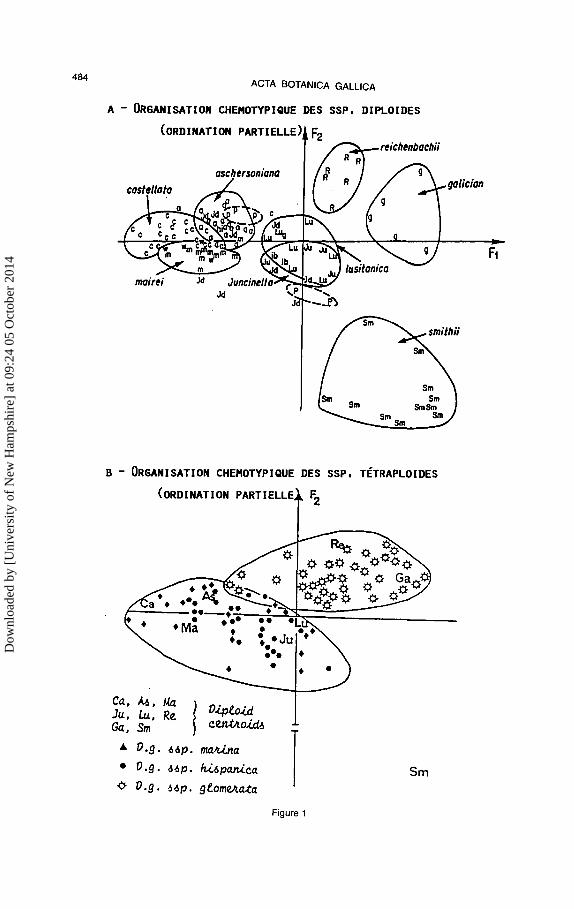
La matrice des donnees (genotypes/variables phenoliques) a ete soumise a une analyse factorielle des correspondances; les resultats sont rapportes dans Ia figure I qui represente le plan 1/2 de !'ordination avec 58% d'inertie objectivee; pour plus de clarte, les ensembles diploldes et tetraploldes ont ete extraits separement d'une ordination initialement unique.
Les sous-especes diploides se resolvent en unites assez bien delimitees, done differenciees et specialisees au plan metabolique; ces unites s'organisent apparemment par rapport a trois poles.
Les sous-especes tetraploldes occupent des volumes plus importants qui traduisent une amplitude de regulations metaboliques plus importante que pour Ies diplo"ides. II existe une nette demarcation du type tempere (ssp. glome rata) et du type mediterraneen (ssp. hispanica) incluant Ie type subtropical (ssp. marina). Ces deux unites se
superposent a deux des trois poles presentes par les diploi"des ; seule Ia sous-espece diplolde smithii se singularise comme representant du troisieme pole.
Cette organisation nee de I' emploi du descripteur micromoleculaire apporte une bonne illustration du caractere specialiste des diploides qui etant le resultat d'une forte radiation adaptative, sont aujourd'hui infeodes a des milieux tres precis quant a leurs caracteristiques physico-chimiques et biotiques. A !'inverse, Ies tetraploi"des dont les performances physiologiques ont ete accrues par le doublement du nombre des alleles, se comportent comme des generalistes capables d'une grande souplesse d'adaptation metabolique pour coloniser de nouveaux milieux.
D'une maniere plus subtile, dans Ia composante tetraploi"de, Ia strategie metabolique generaliste de Ia ssp. hispanica est
Tableau 1.- Composition subspecifique du compexe Dactylis g/omerata L. (Poaceae).
Table 1.- Subspecific composition of the Dactylis glomerata L. (Poaceae) complex.
EURASIEN
Tl!llpf!re
IEDITERRANEEN
Mfcli terraneen
Subtropical
Diploides
aschersoniana
himalayensis
parthiana
"galicien•
Rei chenbachi i
juncinella
castellata
judaica
woronowii
lusitanica
SIIUhi i
santai
mairei
ibizensis
Tl!traploides
gl011erata
(slovenica)
hispanica
urina
(hylodes)
Dow
nloa
ded
by [
Uni
vers
ity o
f N
ew H
amps
hire
] at
09:
24 0
5 O
ctob
er 2
014
484 ACTA BOTANICA GALLICA
A - ORGANISATION CHEMOTYPIQUE DES SSP, DIPLOIDES
(ORDINATION PARTIELLE)
galician
B - ORGANISATION CHEMOTYPIQUE DES SSP, TETRAPLOIDES
(ORDINATION PARTIELLE F2
Ca, M, Ma
t V.i.pto.ui Ju., Lu., Re
Ga, Sm centlto.td6
... V.g • 6 6 p • mai!.Uta I • V.g . 66p. h.i.6pan-i.ca Sm ¢- V.g. 66p. glomvr.a.ta
Figure 1
Dow
nloa
ded
by [
Uni
vers
ity o
f N
ew H
amps
hire
] at
09:
24 0
5 O
ctob
er 2
014

M. JAY ET AL. 485
differente de celle de Ia ssp. glome rata. En effet, Ia ssp. hispanica base sa diversification adaptative sur 10 molecules environ alors que pour Ia ssp. glomerata plus de vingt molecules sont concemees ; cela semble vouloir dire que les tetraploldes mediterraneens fondent leur innovation principalement sur des mecanismes de regulation metabolique, alors que les temperes s'appuient davantage sur le travail des genes de structure.
2 - Les composes flavoniques, marqueurs de l'ascendance dans les premieres etapes de Ia domestication du rosier Rosa x hybrida (Rosaceae)
Les premieres etapes de Ia domestication du Rosier ont repose sur les rencontres de genotypes sauvages aujourd'hui classes dans trois des dix sections du sous-genre Eurosa : Gallicanae, Synstylae et Chinenses. Ces rencontres ont donne naissance a des families de roses anciennes (Tableau 2) dont les plus connues et reputees sont :
- les rosiers de Bengale issus de Rosa chinensis sempeiflorens
- les rosiers Noisette hybrides de Rosa moschata et de Rosa indica vulgaris
- les rosiers de Bourbon dont l'origine est plus complexe et s' appuie probablement sur les trois sections precitees.
Une trentaine de varietes de roses anciennes, choisies pour illustrer les trois families Bengale, Noisette et Bourbon, ont ete recoltees a Ia Roseraie departementale de l'Hay-les-Roses. Les petales de fleurs prelevees au Stade jeune ont ete Soumis a un melange alcoolique et les composes flavoniques ex traits ont ete separes par chromatographie liquide haute pression qui permet de detecter une vingtaine d'heterosides de quercetine et de kaempferol.
Les resultats analytiques ainsi que Ia proportion de quercetine par rapport au kaem-
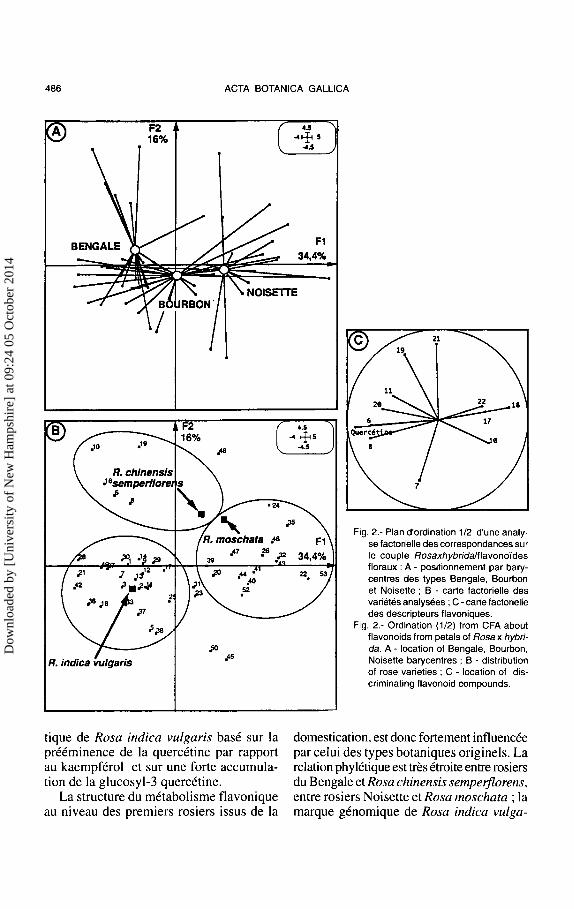
pferol ont ete soumis a une analyse multivariee de type ACP normee dont un plan d'ordination emportant 59% de l'inertie totale est rapporte dans Ia figure 2 (Raymond et al., 1995).
Trois groupes de varietes ressortent de cette analyse neutre :
- le premier groupe reunit les varietes dont le profil metabolique flavonique est semblable a celui de Rosa chinensis sempeiflorens : Ia marque discriminante de ce profil est le glycoside de kaempferol n° 19. Toutes ces varietes se rattachent au type rosier du Bengale ;
- le second groupe rassemble des varietes dont le metabolisme flavonique reproduit assez etroitement celui de Rosa moschata, base sur Ia preeminence du kaempferol par rapport a Ia quercetine et sur Ia forte representation du rhamnosyl-3 kaempferol. Ces varietes sont pour l'essentiel des representants du type Noisette, associes a quelques representants du type Bourbon ;
- le troisieme groupe est plus composite puisqu'on y trouve des representants des trois types : Noisette, Bengale et Bourbon, avec neanmoins une predominance pour les deux derniers. Tous ces exemples developpent un metabolisme flavonique caracteris-
Tableau 2.- Origines botaniques des rosiers Bengale, Bourbon et Noisette.
Table 2.- Botanical origins of Bengale, Bourbon and Noisette roses.
section CHINENSI!S
R. Indica vulgaris
NOISETTE
section SYNSTYLAI!
R.mo5chltl
5acdon GALL/CANAl! R.datnUCIIIIII
Fig. 1.- Plan d'ordination 1/2 (58% d'inertie) d'une analyse factorielle des correspondances sur le couple genotype de dactyles/flavono'ides foliaires.
Fig. 1.- Ordination (1/2 =58% inertia) from Cfill. about flavonoids from the leaves of Oactylis glomerata.
Dow
nloa
ded
by [
Uni
vers
ity o
f N
ew H
amps
hire
] at
09:
24 0
5 O
ctob
er 2
014
486 ACTA BOTANICA GALLICA
A
B
R. indica vulgaris
F2 16%
tique de Rosa indica vulgaris base sur Ia preeminence de Ia quercetine par rapport au kaempferol et sur une forte accumulation de Ia glucosyl-3 quercetine.
La structure du metabolisme flavonique au niveau des premiers rosiers issus de Ia
F1 34,4%
Fig. 2.- Plan d'ordination 1/2 d'une analyse factorielle des correspondances sur le couple Rosaxhybridalllavono'ldes floraux : A - positionnement par barycentres des types Bengale, Bourbon et Noisette ; B - carte factorielle des variates analysees ; C -carte factorielle des descripteurs flavoniques.
Fig. 2.- Ordination (1/2) from CFA about flavonoids from petals of Rosa x hybrida. A - location of Bengale, Bourbon, Noisette barycentres ; B - distribution of rose varieties ; C - location of discriminating flavonoid compounds.
domestication, est done fortement influencee par celui des types botaniques originels. La relation phyletique est tres etroite entre rosiers du Bengale et Rosa chinensis semperjlorens, entre rosiers Noisette et Rosa moschata ; Ia marque genomique de Rosa indica vulga-
Dow
nloa
ded
by [
Uni
vers
ity o
f N
ew H
amps
hire
] at
09:
24 0
5 O
ctob
er 2
014

M. JAY ET AL. 487
ris est plus large puisque son empreinte metabolique se retrouve dans des varietes relevant des trois families de roses anciennes.
Ces observations sont par ailleurs en bon accord avec Ia connaissance generalement admise:
- d'un role precis d'especes telles que Rosa chinensis et Rosa moschata,
- d'une influence plus large de l'espece Rosa indica,
dans les etapes initiales de Ia selection du rosier cultive (Wylie, 1954 ; Maia et Venard, 1976).
Cet exemple est particulierement remarquable par le role qu'il reconnait aux marqueurs flavoniques en tant que temoins et jalons d'un processus de domestication.
3 - Les composes flavoniques, marqueurs de differenciation infraspecifique et de strategies au sein de l'espece Lotus corniculatus (Papilionaceae)
Lars des demieres glaciations, le sud des Alpes a servi de refuge aux Lotiers et plus particulierement aux populations diploldes (Lotus alpinus) qui formaient alors une aire continue. II s'en est suivi une fragmentation de cette aire et un retrait des lotiers diploldes, soit a l'interieur de Ia chaine alpine (Nunataks), soit a sa peripherie (Mt Ventoux, Montagne de Lure, par exemple ). Lars du rechauffement, ces Lotiers dip Joides relictuels auraient foumi les pionniers de Ia recolonisation ; ils auraient en outre donne naissance a des formes tetraploides (Lotus corniculatus) qui plus competitives que leurs and~tres, auraient reussi Ia reconquete des milieux de basse altitude (Favarger, 1975 ; Urbanska et Schwank, 1980).
Partant sur cette hypothese, notre etude a porte sur une collection de genotypes de Lotiers recoltes dans Ies Massifs du Mercantour, du Mont Ventoux, de Ia Montagne de Lure, du Haut Verdon et des Trois Eveches. Apres a voir determine Ies niveaux de ploldie, le materiel vegetal a ete analyse chromatographiquement pour son metabolisme flavonique foliaire.
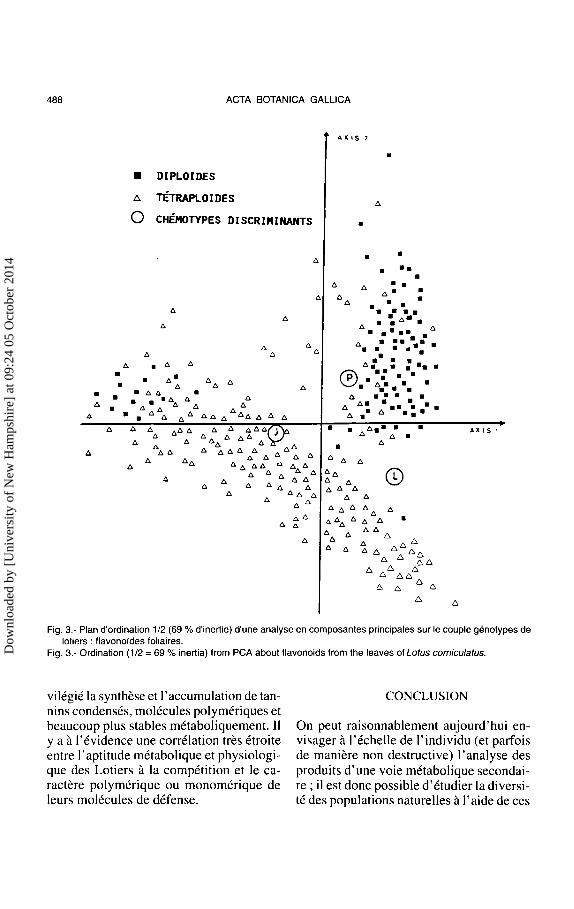
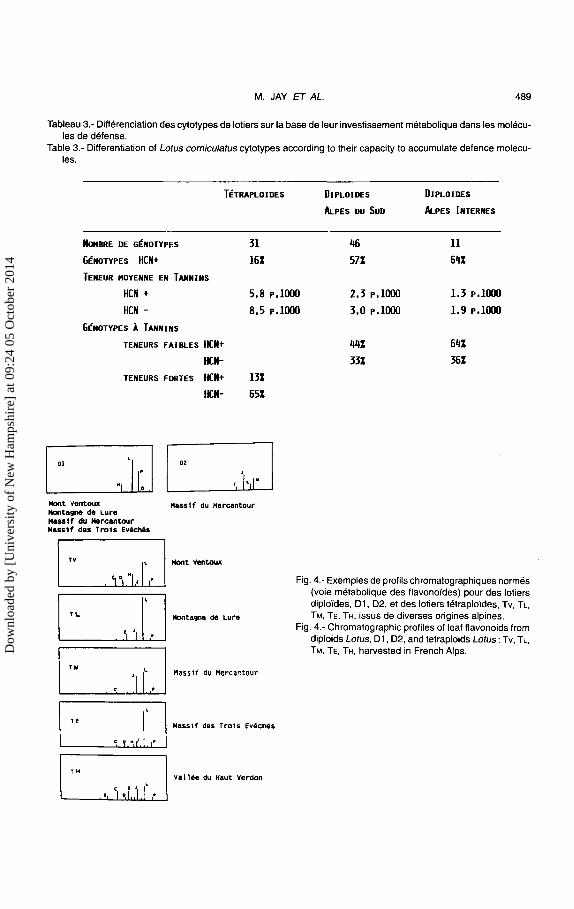
Les resultats traites par analyse en composantes principales donnent lieu a I' ordination presentee dans Ia figure 3 qui rassemble 69% de l'inertie totale (Reynaud et Jay, 1991 ). Globalement, les dip Joides se separent des tetraploldes en deux nuages assez bien discrimines les premiers par des profils flavoniques simples ou Ies composes Let P sont majoritaires, les seconds par des profils beaucoup plus complexes ou seulle compose Lest dominant (Fig. 4).
En ce qui conceme les diploi"des, Ie Massif du Mercantour s'est revele tres interessant : en effet, au Col des Champs, Ies genotypes diplo"ides se repartissent en deux chemotypes tres distincts, l'un identique a celui des dip Joides duMont Ventoux, I' autre entierement nouveau apportant une dominance du flavonolde J. Ces deux types de diplo"ides, morphologiquement non discernables, cohabitent dans Ia meme station, et apparemment ne s'hybrident pas. C'est Ia premiere fois qu'une telle differenciation est montree, et )'hypothese est avancee selon laquelle le chemotype original pourrait provenir des Alpes centrales.
Uncertain nombre de tetraplo"ides originaires en particulier du Mont Ventoux mantrent des profils flavoniques restes proche de ceux des diplo!des, situation temoignant probablement d'un phenomene de tetraploldisation recente. Pour les autres, nous relevons une marque subtile de differenciation pour chaque massif et une marque forte d'hybridation ou d'introgression entre genotypes issus des deux types diplo"ides.
D'un point de vue differenciation fonctionnelle, nous avons essaye de caracteriser les strategies d'adaptation chimique de chacun des deux cytotypes et des trois chemotypes precedemment decrits (Jay et al., 1991 ; Jay et al., 1994 ). Les resultats rappartes dans le tableau 3, montrent que les diplo"ides des deux origines ont adopte une strategie commune de defense qui privilegie Ia cyanogenese et done des composes de faible poids moleculaire et a tum-over rapide. Par contre, Ies tetraploldes ont pri-
Dow
nloa
ded
by [
Uni
vers
ity o
f N
ew H
amps
hire
] at
09:
24 0
5 O
ctob
er 2
014
488 ACTA BOTANICA GALLICA
A X IS 2
• • DIPLOIDES
e. TETRAPLOIDES
() CHEMOTYPES DISCRIMINANTS •
6 6
6
I> 6 6
6 • 6 6
• 6• • • 66 6 6
6
• •
• • • •
6 a • • 6. •
•
66 ••••
••••
•
• .. 6 ••
.6. • • • • • 6
A a •• • •• a . · ..... • • •
.0. •••• • •••
® ••. .• Aa• • •. a
6 • • • •
6 a• • •• •. • • 6 a• A a a a
• .o..• • 6 6 •
6
•
6 6 6
AXIS 1
"6 6 "
Fig. 3.- Plan d'ordination 1/2 (69% d'inertie) d'une analyse en composantes principales sur le couple genotypes de lotiers : flavono"ides foliaires.
Fig. 3.- Ordination (1/2 = 69% inertia) from PCA about flavonoids from the leaves of Lotus corniculatus.
vilegie Ia synthese et I' accumulation de tannins condenses, molecules polymeriques et beaucoup plus stables metaboliquement. II y a a I' evidence une correlation tres etroite entre I' aptitude metabolique et physiologique des Lotiers a Ia competition et le caractere polymerique ou monomerique de leurs molecules de defense.
CONCLUSION
On peut raisonnablement aujourd'hui envisager a l'echelle de l'individu (et parfois de maniere non destructive) l'analyse des produits d'une voie metabolique secondaire; il est done possible d' etudier Ia diversite des populations naturelles a I' aide de ces
Dow
nloa
ded
by [
Uni
vers
ity o
f N
ew H
amps
hire
] at
09:
24 0
5 O
ctob
er 2
014
M. JAY ET AL. 489
Tableau 3.- Differenciation des cytotypes de lotiers sur Ia base de leur investissement metabolique dans les molecules de defense.
Table 3.- Differentiation of Lotus cornicu/atus cytotypes according to their capacity to accumulate defence molecules.
TfTRAPLOIDES DIPLOIDES DIPLOIDES
ALPES DU Suo ALPES INTERNES
NDMBRE DE GENOTYPES 31 46 11
GENOTYPES HCN+ 16Z 57Z 64Z TENEUR HOYENNE EN TANNINS
HCN + 5.8 p,lOOO 2.3 p,lOOO 1.3 p,l()()()
HCN - 8.5 P.lOOO 3.0 p,lOOO 1.9 p,}()()()
GENOTYPES A TANNINS
TENEURS FAIBLES HCN+
HCN-
TENEURS FORTES HCN+ 13Z
01
Mont Ventoux Montagne de· Lure Massif du Mercantour Massif des Trois Ev6clwls
I' c 0 " I I 1,, !'
Jl
, .. J .. I
HCN- 65Z
Mass t f du Mercantour
Mont Ventoux
Montagne de Lure
Massif du Mercar.tour
Massif des T ro 1 s Ev8ches
Vallee du Haut verdon
44Z 64Z 33Z 36Z
Fig. 4.- Exemples de profils chromatographiques normes (voie metabolique des flavono"ides) pour des lotiers diplo"ides, 01, 02, et des lotiers tetraplo"ides, Tv, TL, TM, TE, TH, issus de diverses origines alpines.
Fig. 4.- Chromatographic profiles of leaf flavonoids from diploids Lotus, 01, 02, and tetraploids Lotus: Tv, TL, TM, TE, TH, harvested in French Alps. D
ownl
oade
d by
[U
nive
rsity
of
New
Ham
pshi
re]
at 0
9:24
05
Oct
ober
201
4

490 ACTA BOTANICA GALLICA
descripteurs et d'acceder a une nouvelle perception de leur organisation.
Si nous pouvons joindre a !'etude de Ia diversite chimique, celle de Ia biologie de reproduction pour I' espece concemee, no us aurons I' opportunite de com prendre le fonctionnement de Ia diversification chimique au sein de cette espece.
Finalement, si nous pouvons completer cette approche synthetique du couple genotypes-milieu par quelques mesures des pressions de selection qui s 'y exercent, il deviendra possible de comprendre comment fonctionne le filtre du milieu, et de faire hypothese sur Ia valeur adaptative ou fonctionnelle de certaines voies ou de certaines molecules.
BIBLIOGRAPHIE
Ardouin P., 1985.- Analyse, par Ia biochimie flavonique, de Ia structure genetique et de !'organisation d'un Complexe polyploide Dactylis glomerata L. These Doc. Univ. Lyon Fr., 143p.
Ardouin P., J.L. Fiasson & M. Jay, 1988.- La composante diploide du complexe specifique Dactylis glomerata L. : organisation sur Ia base de son metabolisme phenolique. Taxon, 37, 272-281.
Aardouin P., J.L. Fiasson, M. Jay, R. Lumaret & J.M. Hubac, 1985.- Chemical diversification within the Dactylis glomerata L. polyploid Complex (Gramineae). In : Genetic Differentiation and Dispersal in Plants. Jacquard P., G. Heim & J. Antonovics (Edit.), Springer Verlag, Berlin, 3-15.
Bate-Smith E.G., 1956.- Commoner phenolic constituents of plants and their systematic distribution. Sc. Proc. Roy Dublin Soc., 27, 165-176.
Bate-Smith E.G., 1962.- The phenolic constituents of plants and their taxonomic significance I. Dicotyledons. J. Linn. Soc. (Bot.) London, 58, 95-173.
Bate-Smith E.G., 1965.- Recent progress in the chemical taxonomy of some phenolic constituents of plants. Bull. Soc. Bot. Fr., Memoires, 16-28.
Biolley J.P. , M. JAY & M.A. Viricel, 1994.- Flavonoid diversity and metabolism in 1 00 Rosaxhybrida cultivars. Phytochemistry, 35, 413-419.
Boyet C., 1985.- Organisation du genre Oryza (Gramineae) il.l'aide de marqueurs micromoleculaires. These Doc. Univ. Lyon Fr., 144p.
Boyet C. & M. Jay, 1989. -Taxonomical and biological organisation in the genus Oryza : 0. latifolia complex. Biochem. Syst. & Ecol., 17, 443-447.
Ducousso A., 1985.- Comparaison de populations adjacentes d'Arrhenaterum elatius sur des sols normaux et toxiques : analyse de Ia diversite genetique et du regime de Ia reproduction. These Doc. Univ. Lille Flandres Artois Fr., 98p.
Favarger C., 1975.- Cytotaxonomie et histoire de Ia flare orophile des Alpes et de quelques massifs montagneux d'Europe. Lejeunia, 77, 1-45.
Fiasson J.L., P. Aardouin & M. Jay, 1986.- La camposante tetraploide du complexe specifique Dactylis glomerata L. Organisation et signification fonctionnelle sur Ia base de son metabolisme flavonique. An.
Sc. Nat. Bot. Paris, 8, 35-43. Fiasson J.L., P. Ardouin & M. Jay, 1987.- About a phylo
genetic groundplan of the specific complex Dactylis glomerata (Poaceae). Biochem. Syst. & Ecol., 15, 225-229.
Gannet J.F., 1993.- Flavonoid glycoside variation in wild specimen of Centaurea triumpheti and comments on its relashionships with Centaurea montana based on flavonoid fingerprintings. Biochem. Syst. & Ecol., 21, 389-396.
Hayward M.D. & N.J. Me Adam, 1977.- Isozyme polymorphism as a measure of distinctiveness and stability in cultivars of Lolium perenne. Z. Pflanzenzuchtg, 79, 59-68.
Jay M., 1970.- Quelques problemas taxinomiques et phylogenetiques des Saxifragacees vus a Ia lumiere de Ia biochimie flavonique. Bull. Mus. nat/. Hist. nat. Paris, 42, 754-775.
Jay M., 1988.- La biologie micromoleculaire: une nouvelle approche phytochimique de !'organisation du Monde vegetal. Proc. XIVth Intern. Conference of Polyphenols, Brock University, St Catharines, Canada, 20-32.
Jay M. & F. Ferrero, 1989.- An integrated varietal identification approach using modern technology : application to pink Carnation varieties. Plant Varieties and Seeds, 2, 63-71.
Jay M., J. Reynaud, S. Blaise & D. Cartier, 1991.- Evolution and differentiation of populations of Lotus corniculatusl. in French Alps: conclusions. Evolut. Trends in Plants, 5, 157-160.
Jay M., J. Reynaud, D. Cartier & S. Blaise, 1994.- Diversification strategies of Lotus corniculatus in light of chemical markers. First Intern. Lotus Symposium, Missouri Bot. Gard. StLouis (USA), 20-23.
Lauranson J., X. Vekemans, C. Lefebvre & M. Jay, 1995.Fiavonoid profiles variation in Armeria maritima (Mill.) Willd. Biochem. Syst. & Ecol., 23,319-329.
Lebreton P., 1962.- Contribution il.l'etude flavonique du Houblon (Humulus lupulus) et d'autres Urticales. These Doc. Univ. Lyon Fr., 94p.
Ledeme P. & M. Jay, 1987.- Application of colour analysis and chemical fingerprint in identification of Carnation cultivars. Acta Hort., 216,219-226.
Dow
nloa
ded
by [
Uni
vers
ity o
f N
ew H
amps
hire
] at
09:
24 0
5 O
ctob
er 2
014

M. JAY ET AL. 491
Lumaret R., 1981.- Structure genetique d'un complexe polyploide : Dactylis glomerata L. These Doc. Univ. Montpellier Fr., 168p.
Maia N. & P. Venard, 1976.- Cy1otaxonomie du genre Rosa et origine des rosiers cultives. In: Travaux sur le Rosier de Serre, INRA, station d'Antibes Fr.
Raymond 0., J.P. Biolley & M. Jay, 1995.- Fingerprinting the selection process of Ancient Roses by means of floral phenolic metabolism. Biochem. Syst. Eco/., 23, 555-565.
Reynaud J. & M. Jay, 1991.- Evolution and differentiation of populations of Lotus corniculatus L. from
French Alps : Contribution of phenolic markers. Evolut. Trends in Plants, 5, 149-156.
Urbanska K. & 0. Schwank, 1980.-Variation within Lotus corniculatus L. from Switzerland. Ill. Microdifferentiation in Lotus a/pinus (DC) Schleicher above the timberline. Ber. Geobot. lnst. Eidg. Techn. Hochschule, Stiftung Rubel, 47, 29-47.
Voirin B. & M. Jay, 1978.- Etude chimiotaxonomique des Lycopodiales, lsoetales, Selaginellales et Psilotales. Biochem. Syst. & Eco/., 6, 99-102.
Wylie A.P., 1954.- The history of garden roses. J. Roy. Hort. Soc., 12, 555-571.
Dow
nloa
ded
by [
Uni
vers
ity o
f N
ew H
amps
hire
] at
09:
24 0
5 O
ctob
er 2
014