Embed Size (px)
Citation preview

UNIVERSITE CHEIKH ANTA DIOP -DAKAR
FACULTE DE MEDECINE ET DE PHARIVlACIEi'"
ANNEE 1991
LES HEPATITES NON-A NON-B :DIAGNOSTIC BIOLOGIQUE ET ETUDE EPIDEMIOLOGIGUE'
A DAKAR ET DANS SA REGION
soutenue le 26 janvier 1991
par
Madame Aminata 5ALL/ r ----, ---=, !. ONSEll AFRICAïN~ '- "" '-, -,
i POUR l'ENSEIGNEM MAtGACH~,: C. A. MES ENT SUPERIEUR!
1... - OUAGAD 1
Arr;v~e 2.J .ADU.r .1995 OUGOU!Enregistré ~OUSn. - ' f
MEMBRES DU JURY~~~" '," 'H. D'1' ~: '1, 3' . ,:
Président ~ M. René NDOYE, Professeur
Membres: M. Amadou Moustapha SOW, Professeur
M. Doudou BA. Professeur
M. Lamine DIAKHATE, Professeur
M. Abibou SAMB, Professeur
Directeur de Thèse: M. Jacques PILLOT, Professeur
Co-directeur: Docteur Bernard LEGUENNO
Invité: Docteur Jean-Pierre DIGOUTTE

Ce travail a été réalisé avec l'aide d'un contrat de recherche
DREP n° 90/071 en collaboration avec
- L'Institut Pasteur de Dakar (service virologie médicale)
- L'Institut Pasteur de Paris (service immunologie microbienne)
- L'hôpital Antoine Béclère Clamart (service immunologie)

JE DEDIE CE TRAVAIL
- A mon MARI
Ta patience et ta compréhension sont inestimables. Sans cesdeux qualités qui te glorifient, ce travail n'aurait jamaispu être réalisé avec autant de sérénité et de lucidité.
Ce travail est donc le tien.
- A mes PARENTS
Que de sacrifices consentis pour la réussite de vosenfants !Puisse ce travail vous donner satisfaction et vous procurerde la joie.
- A ma soeur Aïssatou SALL, et à mon frère Cheikh T. SALL
In memoriam.
- A mes enfants
- A toute ma famille
- A toute ma belle-famille
- A mes amies Tout particulièrement Mme DIENG Khady SECK etMme DIOP Awa GUEYE

A NOS MAITRES ET JUGES
Toutes VOS ,critiques et vos suggestions seront bienacceptées.
Votre ardeur et votre passion du travail font de vous desEtres appréciés.Vos réalisation scientifiques font de vous des Hommesrespectés.Vous êtes donc des juges dignes de ce nom.
Au Professeur Jacques PILLOT
- Directeur de l'Unité d'Immunologie MicrobienneInstitut Pasteur Paris
- Chef du Service de Bactériologie-Immunologiede l'Hôpital Antoine BECLERE
- Professeur de Microbiologie à la Faculté deMédecine de l'Université Paris VI
Vous nous avez accueilli dans vos Laboratoires et permis laréalisation de ce travail que vous avez dirigé avecbienveillance et enthousiasme.Votre support financier et technique ont été indispensablesà son accomplissement.Soyez-en remercié et soyez assuré de mon admiration.
Au Docteur Bernard LEGUENNO
- Chef du Service de virologie Médicale del'Institut Pasteur de Dakar
Vous nous avez accueilli dans votre service plusieurs annéesdurant.Vos conseils et suggestions tout au long de ce travail ontété d1un grand apport.En témoignage de mes remerciements.

JE REMERCIE
- Tout le personnel du laboratoire d'analyses biologiques del'Institut d'Hygiène Sociale tout particulièrement Mme DIOP SataDIAGNE pour sa participation à l'étude expérimentale.
Tout le personnel du laboratoire de Pharmacologie/Physiologie de l'UCAD plus particulièrement le Professeur BabacarFAYE, et Mme DIENG Cathy FAYE.
- Toute l'équipe de Physiologie Médicale de l'UCAD.
Le personnel du laboratoire d'immunologie de l' HôpitalAntoine BECLERE et tout particulièrement :
Mme LAZIZI Yamina auprès de qui j'ai trouvé un soutien moral etscientifique sans faille durant mes multiples séjours en France.
Mme Liliane KEROS
Mme Solange D' AZAMBUJA, Mme Marie José BRIANTAIS, Serge, Edi the"Marie Françoise.
- Le personnel du laboratoire d'Immunologie Microbienne del'Institut Pasteur de Paris, en particulier Hakima, Bernadette,Agathe.
L'équipe du Département Informatique de l'ENSUT toutparticulièrement Mr CORENTHIN et Mr BASSENE.
- Toute la famille BLOT pour leur sincère amitié

-p L A li
INTRODUCTION
SITUATION DU PROBLEME
PREMIERE PARTIE = DIAGNOSTIC BIOLOGIQUE DE L'HEPATITE E :MISE AU POINT D'UN TEST DE DEPISTAGE DES SUJETS INFECTES PAR LEVHE
ETUDE EXPERIMENTALE SUR L'ANIMALDU VHE AU SINGE.
1 - Matériel et méthode2 - Résultats3 - Tests d'inoculations croisées
TRANSMISSION
PURIFICATION DES IMMUNOGLOBULINES ET MARQUAGE
1 - PREPARATION DES IgG ANTI VHE A PARTIR DE SERUMS DESUJETS INFECTES PAR LE VHE.
1) Matériel
2) Méthode
II - SEPARATION DES ANTICORPS ANTI VHE DE SINGESPAR METHODES CHROMATOGRAPHIQUES
1) - Préparation des IgM anti VHE
2) - Préparation des IgG anti VHE
3) Controle de pureté des IgM et IgG préparées
4) - Concentration desÉ.·,:immunoglobulines préparéeset déterminat~~r~e-l~:t~neur en proteines.
III - MARQUAGE DES IgG FRACTIONNEES' { (~'1;, \\ "::.\
1) - pour immunoe\z}mol~e / '1:C', """ ,~'/.,,<:".~"
a) marquage à la,p$r.oxy~ase, ,t>~r:;lJpe.' . •
b) marquage a la oe-'t?a,""'galactosldase
2) - pour immunofluorescence.
3) - Controle du marquage.

METHODES ANALYTIQUES
RECHERCHE DES ANTICORPS ANTI VHE DANS LES SERUMS DE SINGES
- Etude de la réaction immunoglobuline fluorescentesur coupe de foie de singes infectés
RECHERCHE DE L'ANTIGENE VHE DANS LES SELLES DESUJETS INFECTES :
l - par microscopie. électronique
II - par culture cellulaire
III - par techniques immunoenzymatiques
DEUXIEME PARTIE : DIAGNOSTIC BIOLOGIQUE DE L'HEPATITE C = ETUDECRITIQUE DES METHODES DE DETECTION DES ANTICORPS ANTI HCV.
A - METHODES DE DETECTION DES ANTICORPS ANTI HCV
1) Points communs
2) Points différentiels
B - ETUDE DE LA SPECIFICITE DES TESTS DE DETECTION
1) Méthode
2) Résultats.
TROISIEME PARTIE : ETUDE EPIDEMIOLOGIQUE DES HEPATITES NA NB ADAKAR ET DANS SA REGION
A - RECHERCHE DE L'HEPATITE E DANS UNE POPULATION DESUJETS ATTEINTS D'HEPATITE AlGUE
1) Méthode
2) Résultats
B - ETUDE EPIDEMIOLOGIQUE DE L'HEPATITE C
1) Détermination de la prévalence du portage desanticorps antiHCV dans la population générale
2) Détermination de la prévalence du portage desanticorps antiHCV chez les donneurs de sang.
3) Recherche des facteurs de risque de portage desanticorps antiHCV

4) Evaluation du risque de transmission del'hépatite C pour 100 unités de sangtransfusées par rapport à d'autres maladiestransmissibles par voie parentérale
QUATRIEME PARTIE : DISCUSSION GENERALE
CINQUIEME PARTIE CONCLUSION
SIXIEME PARTIE : BIBLIOGRAPHIE.

AgHBs
AgVHE
BSA
B W
CMV
DEAE
DO
EBV
EDTA
ELISA
E l A
Fc
HIV
IgM
IgG
ITCF
K D
Kb
NANB
NANBNC
NaBH4
ONPG
OPD
PAGE
PBS
PEG
- LISTE DES ABREVIATIONS -
Antigène Australia
Antigène du virus de l'Hépatite E
Bovin serum Albumin
Bordet Wassermann
cytomegalovirus
Diethylaminoéthyl
Densité optique
virus d'Epstein Barr
Sel de l'acide éthylène diamine tétracétique
Enzyme Linked Immuno-Sorbent Assay
Electro Immuno Assay
Partie constante de l'IgG
Human Immunodeficiency Virus
Immunoglobuline type M
Immunoglobuline type G
Isothiocyamate de fluorescéine
Kilodalton
Kilobase
non-A, non-B
non A, non B, non C
Borohydrure de Sodium
Ortho nitrophényl Bêta galactopyrannoside
Ortho phénylène diamine
Electrophorèse en gel de polyacrylamide
Phosphate buffer salin
Polyethylène glycol

PCR
SGOT
SGPT
SOD
VCA
VHA HAV
VHB
VHC
VHE
YF
Polymérisation Chain Reaction.
Sel de l'acide glutamo-oxaloacétique
Sel de l'acide glutamo-pyruvique
Superoxyde dismutase
Antigène Viral Capsidiaire
Virus de l'Hépatite A
Virus de l'Hépatite B
Virus de l'Hépatite C
Virus de l'Hépatite E
Yellow Fever (fièvre jaune)

- LISTE DES TABLEAUX ET FIGURES-
Tableau 1 Le virus de l'Hépatite Ccaractéristiques
Principales
Tableau 2 situation taxonomique probable du virusde l'Hépatite C
Tableau 3 : Principales caractéristiques du VHE
Tableau 4 Diagnostic par soustraction des Hépatitesnon A, non B
Tableau 5 Résultat des selles de malades de Constantinepar méthode d'immunocapture des IgM
Tableau 6 Résultat des selles de malades de Constantinedans le système THjSpH avec des IgG marquées àla péroxydase
Tableau 7 Résultat des selles de malades de Constantinetestées dans les systèmes THjSpH avec des IgGmarquées à la Bêta galactosidase
Tableau 8 Résultat des selles de malades de Constantinedans le système TSjSpS avec des IgG marquées àla péroxydase
Tableau 9 Résultat des selles de malades de Constantinetestées dans le système TsjSpS avec des IgG marquéesà la Bêta galactosidase
Tableau 10
Tableau Il
Tableau 12
Tableau 13
Tableau 14
Tableau 15
Résultat des selles de Constantine testées dans lesystème TsjSpS avec comme liquide de saturation lelait écrémé à 4 %
Recherche de la protéine Fv dans les sellespositives de malades de Constantine
Résultat des selles de Constantine après passagesur immunoadsorbant
Réexamen des selles de malades de Constantineaprès passage sur immunoadsorbant d'IgM monoclonale
Résultat des selles de Bangkok après passage surimmunoadsorbant
Résultat du contrôle de spécif~cité de la réactionimmuno-enzymatique par recherche de l'antigène VHE

Tableau 16
Tableau 17
Tableau 18
Tableau 19
Tableau 20
Tableau 21
Tableau 22
Tableau 23
Tableau 24
Tableau 25
Tableau 26
Résultat du contrôle de la spécificité de laréaction immuno-enzymatique par réactionsd'inhibition
Résultat du contrôle de la spécificité de laréaction immunoenzymatique par réactionsd'inhibition avec des IgM et avec des IgG
Résultat du contrôle de la spécificité de laréaction immunoenzymatique par des réactions témoins
Résultat du contrôle de la spécificité des méthodesde détection du VHC
Résultat de la recherche des marqueurs viraux dansles sérums de sujets DAKAROIS atteints d'Hépatiteaiguë
Résultat de la recherche des anticorps anti HCVdans les serums de sujets atteints d'Hépatite aiguëà DAKAR
Résultat de la recherche de l'AgVHE dans lesselles de sujets atteints d'Hépatite aiguë à DAKAR
Résultat de la recherche de l'AgVHE dans les sellesde sujets atteints d'Hépatite aiguë à DAKAR aprèspassage sur immunoadsorbant
Résultat de la recherche des facteurs de risquede portage des anticorps antiHCV à DAKAR
Résultat de la recherche des anticorps antiHCVen relation avec des marqueurs indirects desHépatites
Propriétés physico-chimiques et biologiques desagents responsables des Hépatites NANB

- FIG URE S -
Figure 1
Figure 2
Figure 3
Evolution des transaminases chez le singe témoinSI
Evolution des transaminases chez le singe témoinS2
Evolution des transaminases chez le singe témoinS3
Figure 4 Evolution des transaminases chez le singe témoinS4
Figure 5 Evolution des transaminases chez le singe PATAS S5inoculé avec l'extrait de selles chloroformé
Figure 6 Evolution des transaminases chez le singe PATAS S6inoculé avec l'extrait de selles non chloroformé
Figure 7 Evolution des transaminases chez le singecercopithèque S7 inoculé avec l'extrait de selleschloroformé
Figure 8 Evolution des transaminases chez le singecercopithèque S8 inoculé avec l'extrait de sellesnon chloroformé
Figure 9 Evolution des transaminases chez le singe S9 aprèsinoculation croisée
Figure 10 Evolution des transaminases chez le singe S10 aprèsinoculation croisée
Figure Il Résultat du fractionnement des anticorps antiVHE surS300
Figure 12 Evolution de la "proteine Fv" chez le singe (parrapport au SGPT)
Figure 13 Evolution de la protéine Fv chez le singe (parrapport au SGPT)
Figure 14 Evolution de l'AgVHE chez le singe S8 (par rapportau SGOT)
Figure 15 Evolution de l'AgVHE chez le singe S8 (par rapportau SGPT)
Figure 16 Histogramme de répartition de la prévalence desanticorps antiVHE en fonction de la tranche d'âge
Figure 17 Cinétique de l'évolution des anticorps antiVHC enfonction du temps (séroconversion précoce)

Figure 18 Cinétique de l'évolution des anticorps antiVHC enfonction du temps (séroconversion tardive).

INTRODUCTION

si de nombreuses infections virales sont susceptibles de
provoquer des hépatites, le vocable "hépatite virale" a surtout
été réservé aux hépatites A et B qui se différencient par des
critères cliniques, épidémiologiques et étiologiques.
En effet depuis la découverte de l'antigène Australia (AgHBs)
par BLUMBERG et collaborateurs en 1967 (13), et la visualisation
du virus de l'hépatite A au microscope électronique en 1973 (27),
les agents viraux de ces deux types d'hépatite sont actuellement
bien connus; et des tests très performants pour la caractérisation
des virions ou de leurs antigènes, ou la mise en évidence
d'anticorps spécifiques furent mis au point.
c'est dans ce contexte que l'on s'est rendu compte, que des
agents viraux, autres que le virus de l'hépatite A (VHA) ou le
virus de l'hépatite B (VHB) , étaient à l'origine d'hépatites
transmissibles donc infectieuses, qui jusque là, échappaient au
diagnostic biologique.
De telles hépatites furent groupées sous le nom d'''hépatite
non-A non-B".
Mais il convient de préciser que ce groupe bien que très
hétérogène exclut néanmoins :
- les hépatites bactériennes,
- les hépatites parasitaires,
- les hépatites médicamenteuses
les hépatites auto-immunes et même
les hépatites virales dues au cytomégalovirus, ou au virus
d'Epstein Barr, ou encore au virus Delta.
Très rapidement, des données épidémiologiques, virologiques
et cliniques ont permis d'individualiser deux formes d'hépatites
non-A non-B (39-48-70-77-89).
La première ressemble à l'hépatite A ; elle est transmise par
voie entérale, est dénuée de tout risque de chronicité ; elle est
surtout fréquente dans les pays sous développés où elle est
responsable de morbidité et mortalité accrues chez la femme
enceinte au troisième trimestre de la grossesse (3-23-26-49-57-64-
1

80) .
L'agent causal récemment découvert est dénommé VHE (17) et
l'on parle alors d'hépatite E.
La seconde forme ressemble à l'hépatite B elle est
essentiellement transmise par voie parentérale (5-14-32-40-41) et
évoluerait vers la chronicité dans 50 à 80 % des cas (50-66-68)
aboutissant à la cirrhose et au cancer primitif du foie (12-81).
Plusieurs agents en seraient probablement responsables i mais le
principal, identifié récemment, a été dénommé VHC par la majorité
des auteurs, et l'on parle alors d'hépatite C~
si l'identification du VHC et du VHE constitue à l' heure
actuelle un progrès décisif pour la santé publique, le diagnostic
biologique des infections correspondantes pose encore problème.
En effet, les tests de dépistage des sujets infectés par le
VHC ou le VHE, sont à l'état de recherche pour l'hépatite Ei alors
que pour l'hépatite C, nous disposons à l'heure actuelle de tests
immuno-enzymatiques, qui détectent les anticorps dirigés contre un
antigène exprimé lors de l'affection par le virus C (anti HCV) dans
le sérum ou le plasma. Mais ces tests posent des problèmes de
spécificité et les sérologies positives doivent être interprétées
avec prudence lorsque le sérum contient des immuncomplexes, dans
les cas de dysglobulinémie, et même dans certaines affections avec
dysfonctionnement hépatique.
Ce sont ces deux problèmes majeurs qui nous ont conduit :
1- à la mise au point d'un test de dépistage des sujets
infectés par le VHE.
2- à une étude critique des tests de détection des anticorps
antiHcV, avant d'entreprendre l'étude épidémiologique des hépatites
NA NB à Dakar et dans sa région.
2

SITUATION DU PROBlEME

Depuis de nombreuses années déjà, et grâce à la microscopie
électronique, les agents responsables des hépatites A et B sont
bien connus.
Le virus de l'hépatite A ou VHA est un virus de très petite
taille, non enveloppé et dont l'information génétique est portée
par un génome à ARN. Il détermine une virémie de courte durée, de
deux à trois semaines. Pendant cette période le virus est excrété
en abondance et uniquement dans les matières fécales, ce qui
détermine un mode de contamination féco-orale autour du suj et
atteint, du fait des conditions d'hygiène précaires. L'immunité
conférée par ce virus est très stable.
Le virus de l'hépatite B ou VHB est très différent. Sa partie
centrale comprend un déterminant antigénique appelé HBc et un
·génome à ADN qui porte l'information génétique. Ce virus est
enveloppé et son enveloppe porte le déterminant antigénique HBs.
Si l'on exclut les formes fulminantes, la virémie déterminée par
ce virus est toujours prolongée et deux cas de figure peuvent se
présenter: - soit le sujet guérit dans 90 % des cas après une
virémie de six à huit semaines ;
- soit le sujet devient porteur chronique dans 10 % des cas.
La transmission se fait principalement par voie sanguine ou
sexuelle. Le virus n'est pas éliminé dans les matières fécales mais
on le retrouve dans les sécrétions telles que la salive, les
larmes, la sueur ou encore les sécrétions génitales.
Cette parfaite connaissance des VHA et VHB a permis à la
recherche dans le domaine des hépatites de faire un grand bond et
la terminologie "hépatites non-A, non-B" fut introduite pour
désigner des hépatites dont les agents étiologiques n'étaient pas
identifiés, mais qui apparaissent sérologiquement distinctes des
hépatites A et B (4-8-28-29-33-78).
Quels sont donc ces virus des hépatites non-A non-B ?
A l'heure actuelle, deux virus peuvent être sûrement
incriminés (2-14-48-62)
3

- l'un ubiquitaire, déterminerait une virémie prolongée. Il n'est
pas apparemment éliminé par les matières fécales et son mode de
contamination est très voisin de celui du virus B. Ce virus est
dit virus non-A non-B, liB like", mais c'est surtout la dénomination
de VHC (virus de l'hépatite C) qui est reconnue par la majorité des
auteurs depuis 1989.
Il s'agit d'un virus à ARN dont la classification taxonomique
est encore mal définie.
En effet, les comparaisons de séquences en nucléotides
indiquent des homologies avec le groupe des Flavivirus (en
particulier virus de la dengue et de la fièvre jaune), des
Pestivirus et même de virus de plantes (20-51-55-62).
Les principales caractéristiques du VHC et la position
taxonomique des virus voisins sont mentionnées dans les tableaux
suivants :
4
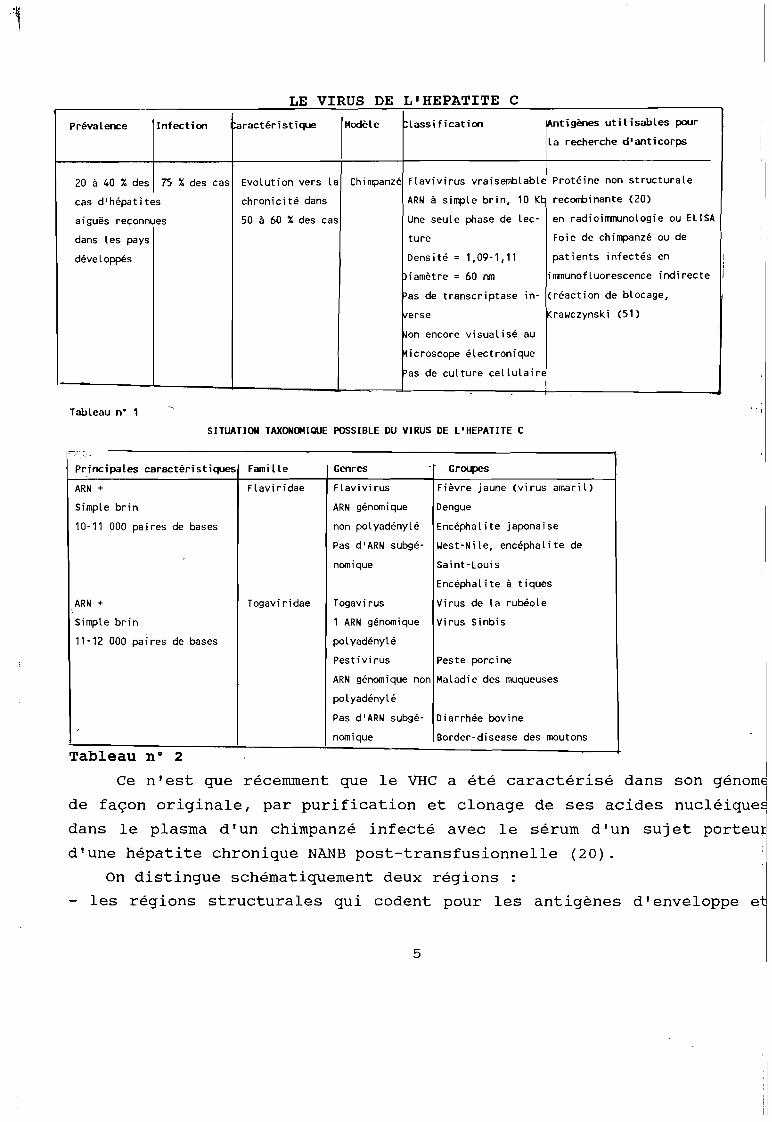
LE VIRUS DE L'HEPATITE C
Prévalence Infection aractéristique Modèle lassification ~tigènes utilisables pour
la recherche d'anticorps
20 à 40 % des 75 %des cas Evolution vers la Chimpanzé Flavivirus vraisemblable Protéine non structurale
cas d'hépatites chronicité dans ARN à simple brin, 10 KI: recombinante (20)
aiguës reconnues 50 à 60 %des cas Une seule phase de lec- en radioimmunologie ou ELISA
dans les pays ture Foie de chimpanzé ou de
développés Densité = 1,09-1,11 patients infectés en ii
Diamètre = 60 nm immunofluorescence indirecte 1
Pas de transcriptase in- (réaction de blocage,
verse Krawczynski (51)
~on encore visualisé au
Microscope électronique
Pas de culture cellulaire1
Tableau n" 1
SITUATION TAXONOMIQUE POSSIBLE DU VIRUS DE L'HEPATITE C
ARN génomique Dengue
non polyadénylé Encéphalite japonaise
Pas d'ARN subgé- Yest-Nile, encéphalite de
Principales caractéristiques Famille
ARN + Flaviridae
Simple brin
10-11 000 paires de bases
Genres
Flavivirus
Groupes
Fièvre jaune (virus amaril)
1 ARN génomique Virus Sinbis
polyadénylé
ARN +
Simple brin
11-12 000 paires de bases
Togaviridae
nomique
Togavi rus
Pestivirus
Saint-Louis
Encéphalite à tiques
Virus de la rubéole
Peste porcine
ARN génomique non Maladie des muqueuses
polyadénylé
Pas d'ARN subgé- Diarrhée bovine
nomique Border-disease des moutons
Tableau n° 2
Ce n'est que récemment que le VHC a été caractérisé dans son génomJ
de façon originale, par purification et clonage de ses acides nUcléiqueJ
dans le plasma d'un chimpanzé infecté avec le sérum d'un sujet porteuJ
d'une hépatite chronique NANB post-transfusionnelle (20). ;1
On distingue schématiquement deux régions : jles régions structurales qui codent pour les antigènes d'enveloppe e
5

......:
de capside.
- la région non structurale qui code en particulier pour des protéineE
impliquées dans la réplication virale. :1
Les figures 1 et 2 proposent un schéma de son organisatio~ généralE
et de son génome que l'on subdivise en gènes st.ructuraux (C, PrM, E) e,~\gènes non structuraux (NSl, NS2, NS3, NS4, NSS).
Les déterminants codés par les régions E et NSI semblent
particulièrement importants pour l' immunogénici té et l' antigénicité d~~particules virales.
$" 9! 1Protéine)\", .1b::~r. ~ d'enveloppe E·
{ J:' y., . .&J...F- Protéine~. . ..':'."~ de membrane M
~: ~~ .~ Prot{oineif .... .~~"B de nucléocapsic1e C
~'. ",:~
~J!J[~~Jt~Figure 1 : Représentati.on schém~~iquedu virus C par analogie aux f1avlvlrus.
Figure 2 : Représentation schématique de l'organisation génétique des flavivirus(réalisée par Vincent Deubel, Laboratoire des Arbovirus, Institut Pasteur Fondation)
AUG 10 723 nucléotides stop
3'
NS5(900)
·IAllr~NS4A NS48(150) (248)
C5-1C 100-3
NS3(618)
NON STRUCTURAL5' STRUCTURAL
IA_A_~_AII·IC prM E NS1 NS2A NS2B(113) (166) (495) (352) (218) (130)
II Â Ipr M
(75)La région 5' du génome virai code pour des protéines de structure intervenant dans la composition de la capside et de I"enveloppe virale.La région 3' code pour des protéines de régulation dont le rôle reste mal connu Les régions E et NS1 semblent très importantes pour l'anligénicilé desparticules.La région du génome viral soulignée en trait plein est celle qui a été initialement clonée par Chiron. Le test anti-HCV (Chiron-Ortho-Abbott) repose donc surl'expression de l'antigène correspondant.
6

L'autre virus incriminé dans ces hépatites NANB est proche du virus
A. La contamination se fait par les aliments souillés par les matières
fécales de sujets infectés par ce virus NANB (9). Ce virus est dit "NANB",
liA like" ou également "VRE".
Dans certains pays d'Afrique ou d'Asie, ce virus existe à l'état
endémique, mais il sévit également soit de manière sporadique soit sous
la forme d'épidémies, dans de nombreux pays sous développés. Déjà en 1955
en Inde, 29 mille cas d'hépatites à transmission orale avaient été
identifiés (60-83). En 1975, une épidémie semblable survenait en Inde dans
la ville de Ahmedabad (71). Des études sérologiques ont révélé qu'il ne
s'agissait ni d'hépatite A, ni d'hépatite B.
Plus récemment encore, en 1980, dans la ville de Kashmir, une épidémie
fut rapportée (79-86). Des études épidémiologiques et cliniques menées dans
d'autres pays sur les hépatites ont révélé des épidémies en Asie du Sud
Est (58), au Népal dans la vallée du Katmandu (38-46), au Pakistan (21).
De telles épidémies sont également survenues en Afrique du Nord, en ALGERIE
(10-59), en Afrique de l'Ouest en COTE d'IVOIRE (69), en Afrique de
l'Est au Soudan (1) et en Somalie (57), et plus récemment encore au Mexique
(80), en Amérique du Nord et au Japon(88). La plus importante est apparue
en Chine où plus de 100 mille cas ont été recensés entre septembre 1986 et
avril 1988.
Dans toutes ces épidémies, le mode de transmission retenu est la
transmission hydrique(1-8-19-36-37-45) .
7
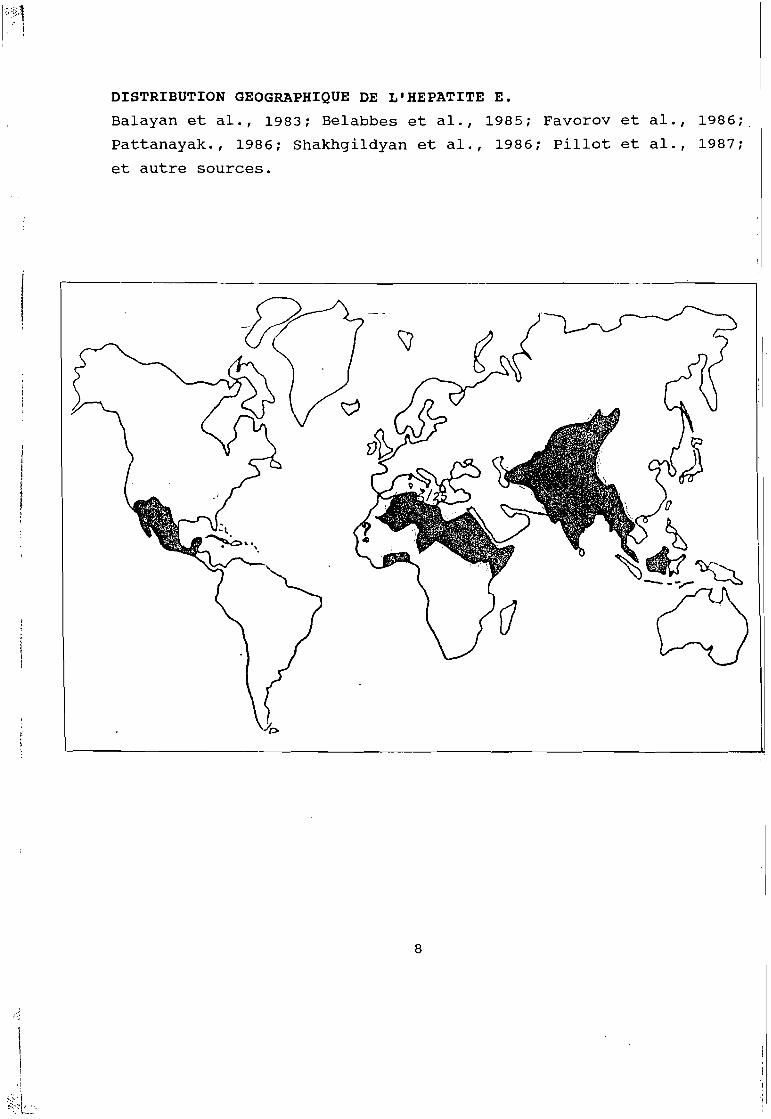
1~4. ,
DISTRIBUTION GEOGRAPHIQUE DE L'HEPATITE E.
Balayan et al., 1983; Belabbes et al., 1985; Favorov et al., 1986;.
Pattanayak., 1986; Shakhgildyan et al., 1986; pillot et al., 1987;
et autre sources.
8

Pour caractériser le virus, de nombreuses recherches au microscope
électronique ont été réalisées, soit à partir de selles de patients ou de
singes infectés (15), soit dans des coupes de foie de singes infectés (7
15), soit dans la bile où le virus semble présent en grande quantité et où
même un clonage du génome viral a pu être obtenu. Dans 4 % des cas, des
particules virales d'un diamètre de 27-38 nm ont été mises en évidence
(15); et on admet aujourd'hui qu'il s'agit d'un virus non enveloppé de 32
nm de diamètre en moyenne.
Son génome a été cloné et séquencé (16-67).
Il est constitué d'un ARN simple brin de 7600 nucléotides. Il fait
partie de la famille des Calicivirus (16).
Deux particularités caractérisent ce virus
- celle de donner des hépatites fulminantes chez la femme enceinte avec
10 à 40 % de mortalité
- une variabilité très significative de souches provenant de différentes
régions du globe (16).
Les principales caractéristiques de ce virus sont mentionnées dans
le tableau suivant :
Prévalence
IMe
Cas sporadiques
50 % des hépatites
aiguës chez l'enfant:
80 % chez les femmes
enceintes
Népal
Epidémies dans La
saison humide
Cas sporadiques entre
les épidémies
80 % des hépatites
Tableau n D 3
Caractéristique clinique Modèle animal
Infection asymptomatique Sensibilité de presque
dans 30 à 50 %des cas toutes les espèces de
Taux de mortalité 1-3 % singes (chimpanzé,
sauf chez la femme encein- Rhésus, cynomolgus,
te cercopithèque)
Pas d'évoLution vers La
chronicité
9
Classification
CaLicivirus
ARN à simple brun
8,5 Kb
Densité = 1,29
Diamètre =
30 - 33 nm
Visualisé dans les
seLLes, La bile
Pas de culture

1
1
l'
1
Si des progrès importants ont été réalisés ces dernières années pOUl
la caractérisation du VHC et du VHE, le diagnostic des infectio~~correspondantes, jusqu'à très récemment ne se faisait que par exclusidl
l '
des autres formes d'hépatites à partir de leur marqueurs viraux.1
Le tableau suivant récapitule les principaux marqueurs qui peuveBi
être recherchés, mais il convient de préciser que l'on se contenB(
généralement d'un seul marqueur considéré comme optimal, plus fiable, Ji
inscrit en caractère gras dans notre tableau. il
DIAGNOSTIC PAR SOUSTRACTION DES HEPATITES NON-A NON-B :1
Hépatites
A B Virus delta CMV EBV YF,
antigène viral -(selles)
1
Hybridation DNA -ou RNA (selles)
IgM anti-virusA -(sérum)
DNA polyrnérase -(sérum)
1
Hybridation DNA - 1
1
(sérum) ,
Antigène pré-S2 -,
(sérum) 1
anti-HBc,
IgM - 1
(sérum)
Antigène viral -(sérum)
1
...,~-;"{J('IvE;:-.
~\IgM anti-Delta ~ _ ft.. ,
,'- "'" ;
(sujets!~ ,
AgHBs+) <' ? :,
~~(~. "
IgM anti-CMV
~IgM anti-VCA \O~~ -\. ,..
trgM anti-YF -;, ::..•.•' e,v< -",..~<":. fl~.m.•{\
Les tests à la fois les plus simples et les plus significatifs pou~l'exclusion du virus considéré sont en caractère gras. Tableau n° 4
10

1
1 1
:1
Cette méthode très laborieuse de diagnostic des hépatites NANB, d~même que la gravité du problème de santé publique qu'elles représenten~1devaient très vite orienter vers la recherche de marqueurs plus SPécifiqu~!
1
en vue d'un diagnostic plus fiable.
11
, ,i t

PREMIERE PARTIE:
MISE AU POINT D'UN TEST DE
DEPISTAGE
DES, SUJETS INFECTES PARlEVHE

Dans la perspective d'un dépistage des sujets infectés par
le VHE, nous avons élaboré différents protocoles de recherche,
les uns visant à rechercher les déterminants antigéniques du
virus dans les selles et sa visualisation au microscope les
autres visant à mettre en évidence des anticorps antiVHE.
Pour la recherche du virus ou de ses déterminants
antigéniques, nous avons mis en jeu :
. l' immunomicroscopie électronique pour la recherche directe
du virus dans les selles,
. la culture cellulaire à partir des selles,
les tests immunoenzymatiques de capture d'antigène sur
les selles.
Pour la recherche des anticorps anti VHE, c'est la
microscopie à immunofluorescence qui a été mise en jeu.
Toutes ces techniques nécessitent soit une source d'antigène
VHE, soit une source d'anticorps anti VHE. c'est pourquoi, nous
avons eu recours :
- A) à la transmission du VHE au singe
B) à des méthodes préparatives à savoir
1. la préparation d' IgG anti VHE à partir de sérums de
sujets infectés par le VHE,
2. la séparation par méthodes chromatographiques des
anticorps anti VHE (IgM et IgG) à partir de sérums de singes
infectés.
3. le marquage des IgG fractionnées pour leur utilisation
comme outil analytique.
12

ETUDE EXPERIMENTALE SUR L'ANIMAL:
TRANSMISSION DU VHE AU
SINGE

..l'existencei Depuis que d'un virus NANB à transmission oro-
fécale fut reconnue, plusieurs publications se succèdèrent
confirmant la présence de ce virus dans de nombreux pays,
notamment dans les pays en voie de développement.
Des protocoles expérimentaux d'étude furent proposés et le
modèle retenu fut la transmission du virus à la race simienne.
Différentes espèces de singes ont été testées cynomolgus,
Rhésus, Saïmiri ... (6-15-69-73-74-76-77).
Dans le même sens, nous avons donc tenté de transmettre
l'hépatite NANB à deux espèces de singe retrouvées au Sénégal:
Erythrocebus Patas et Cercopithecus Eathiops.
1. Matériel et méthode
a) - matériel
Lors d'une épidémie d' hépatite virale à transmission hydrique
survenue en Constantine, des selles et des sérums de suj ets
atteints avaient été collectés pendant la phase aiguë de la
maladie. A partir des sérums, différents marqueurs viraux des
hépatites infectieuses ont été recherchés. L'absence d'IgM anti
HAV, d'IgM anti-HBc, d'IgM anti-Delta, d'IgM anti-CMV, d'IgM
anti-VCA a permis de retenir le diagnostic d'hépatite non-A non
B. Le mode de transmission a permis de retenir le diagnostic
d'hépatite non-A non-B type E. A partir des selles, des
extractions du virus ont été réalisées suivant des méthodes
classiques d'extraction d'entérovirus.
1ère technique
- Faire une suspension de selles à 10 % en PBS (cf préparation
des tampons)
- centrifuger à 5 000 tours/minute pendant dix minutes
Agiter le surnageant avec 0,5 ml de chloroforme
centrifuger à 5 000 tours/minute pendant dix minutes
Recueillir la phase aqueuse
- Passer sur filtre 0,22 f et récupérer le filtrat qui constitue
l'inoculum.
13

2éme technique
- Faire une suspension de selles à la % en PB8
- Centrifuger à 5 000 tours/minute pendant trente minutes
- Filtrer sur filtre 0,22 p et récupérer le filtrat.
b) Méthode
- Phase d'observation ou de pré-inoculation
Pendant cette période, les singes après avoir été
déparasités (lvermec lM ou Mintézol suspension), sont suivis sur
le plan biologique. Un bilan hépatique a été réalisé et les
transaminases sériques ont été dosées chaque semaine pendant un
mois. Les selles ont été également recueillies pendant cette
période.
- Phase d'inoculation
Un lot de 8 singes ne possédant ni anticorps antiHAV, ni
anticorps anti-HBc, répartis en 3 groupes selon l'espèce a été
utilisé dans notre expérimentation.
- Le groupe 1 est constitué de 4 singes témoins dont 2 de type
Patas (81, 8_2) et 2 de type Cercopithèque (83,84). Ces singes
témoins ont été inoculés avec des selles humaines ne comportant
aucun agent infectieux reconnu.
- Le groupe 2 comprend 1 singe Patas (85) ayant reçu 0,5 ml
d'extrait chloroformé de selles d'un sujet atteint d'hépatite E
(El), 1 singe Patas (86) ayant reçu 0,5 ml d'extrait de selles
non chloroformé du même sujet (E2).
est la voie intraveineuse.
1 singe Cercopithèque
1 singe Cercopithèque
- Le groupe 3 comprend également
inoculé avec 0,5 ml dl extrai t El,
inoculé avec 0,5 ml d'extrait E2.
La voie d'administration utilisée
(87)
(88)
Les selles de ces différents singes furent collectées
quotidiennement, les sérums hebdomadairement, et conservés
à -80·C.
2. Résultats
14

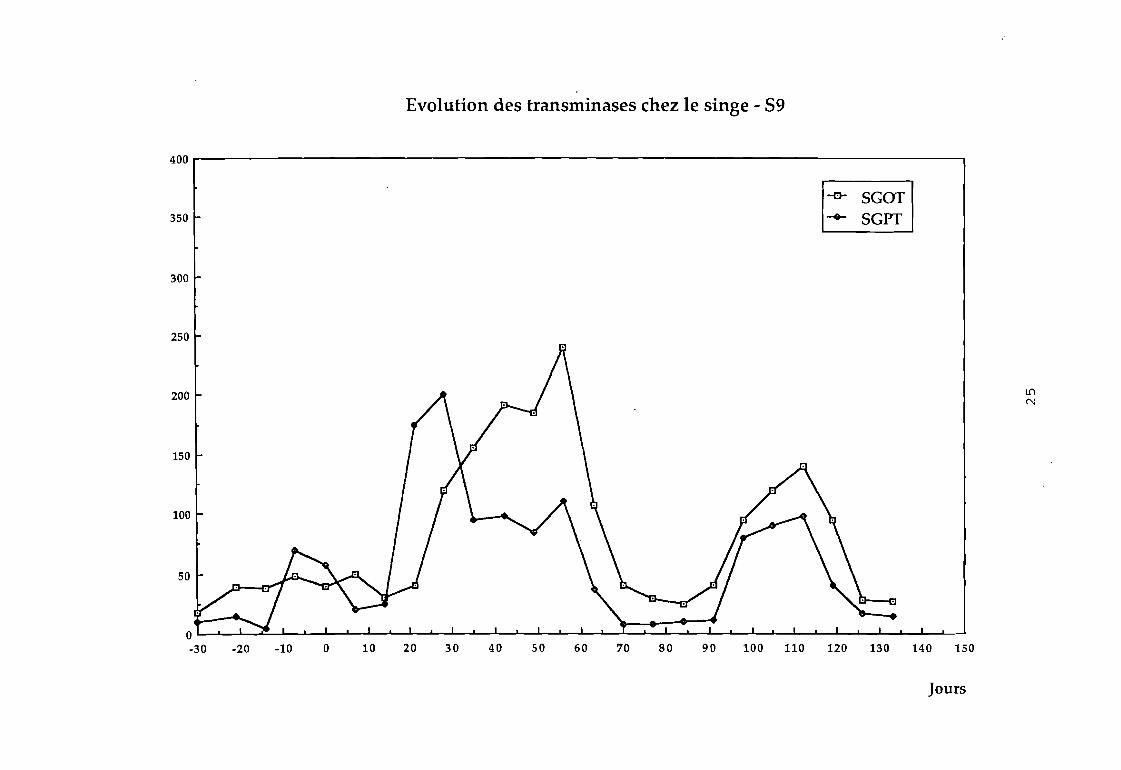
Dans le groupe 1 : aucune variation sensible des transaminases
n'a été observée (cf courbes 81, 82, 83, 84).
Dans le groupe 2 :
85 pas d'élévation des transaminases (cf courbe 85),
86 élévation des transaminases au bout de 10 jours (cf
courbe 86).
Dans le groupe 3 :
87 pas d'élévation des transaminases (cf courbe 87),
88 on observe une augmentation considérable des transaminases
au bout de 4 semaines (cf courbe 88).
15
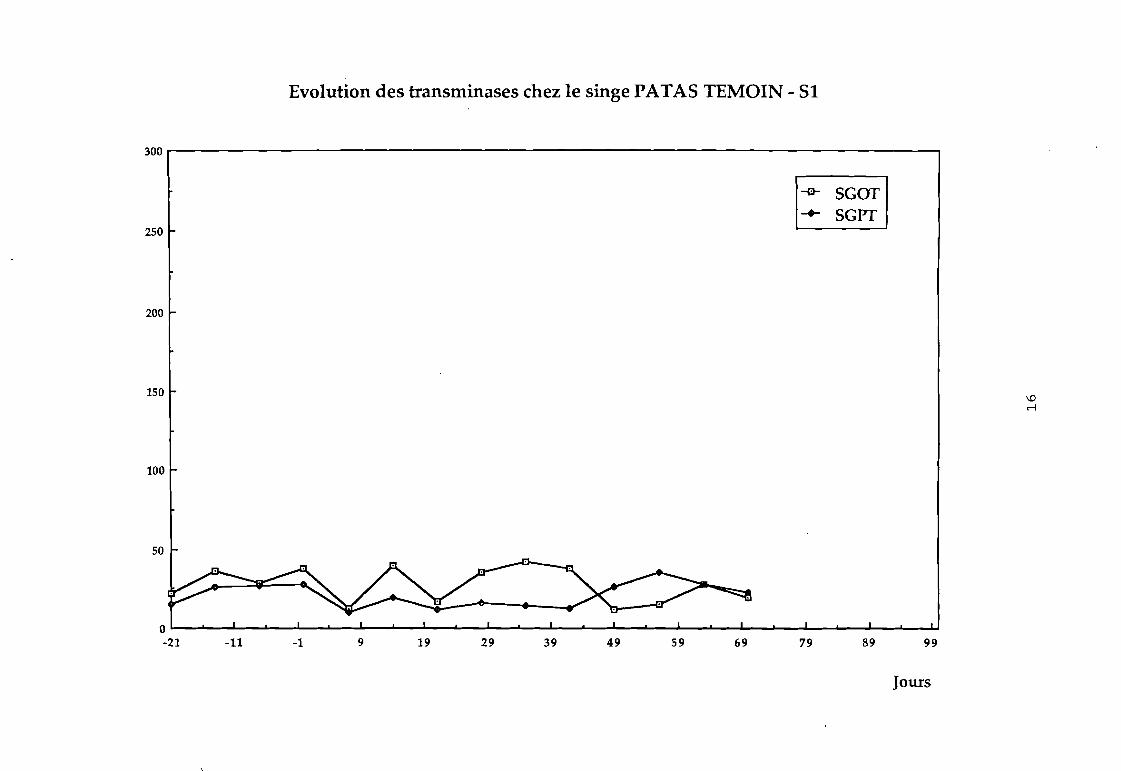
Evolution des transminases chez le singe PATA5 TEMOIN - 51
300 i 1
250
I~ SGor 1...- SGPT
200
150\0.--i
100
50
9989796959493929199-1-11
0' , l , , , , , 1 , 1 , , • , , l , , ft , , , , Il
-21
Jours
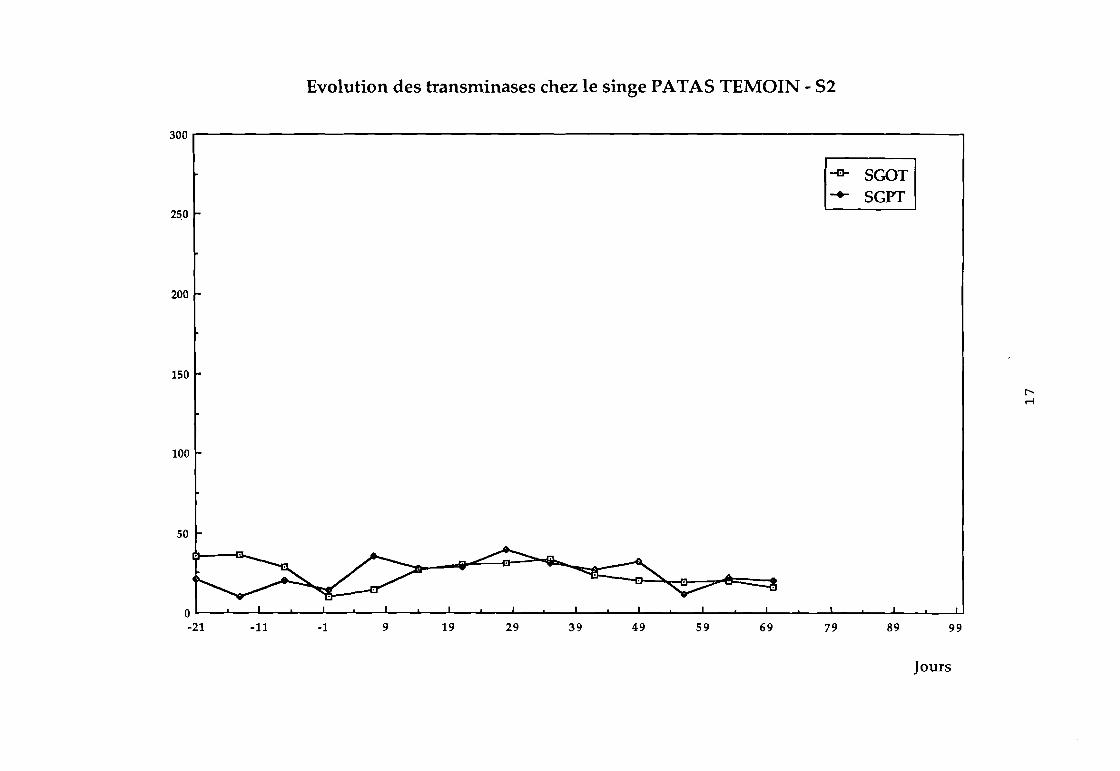
Evolution des transminases chez le singe PATA5 TEMOIN - 52
300 i ,
250
200
150
100
50
I~ SGOT 1-+- SGPT
['r-i
-11 -1 9 19 29 39 49 59 69 79 89 99
Jours
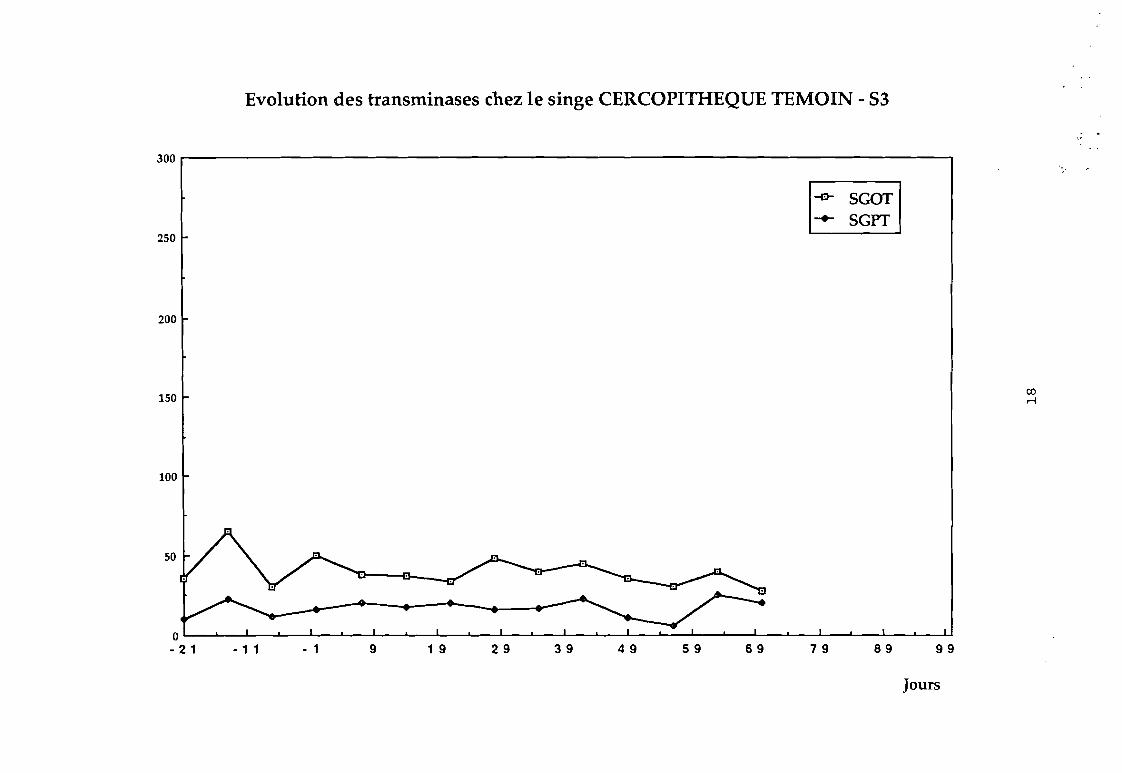
Evolution des transminases chez le singe CERCOPITHEQUE TEMOIN - 53
300 i i
250
1- SGOf1-+- SGPT
200
150 cori
100
99897969594939291 99- 1- 1 1
o l , l , l , l , l , l , l , l , 1 1 l , l , l , 1 1
- 2 1
50
Jours
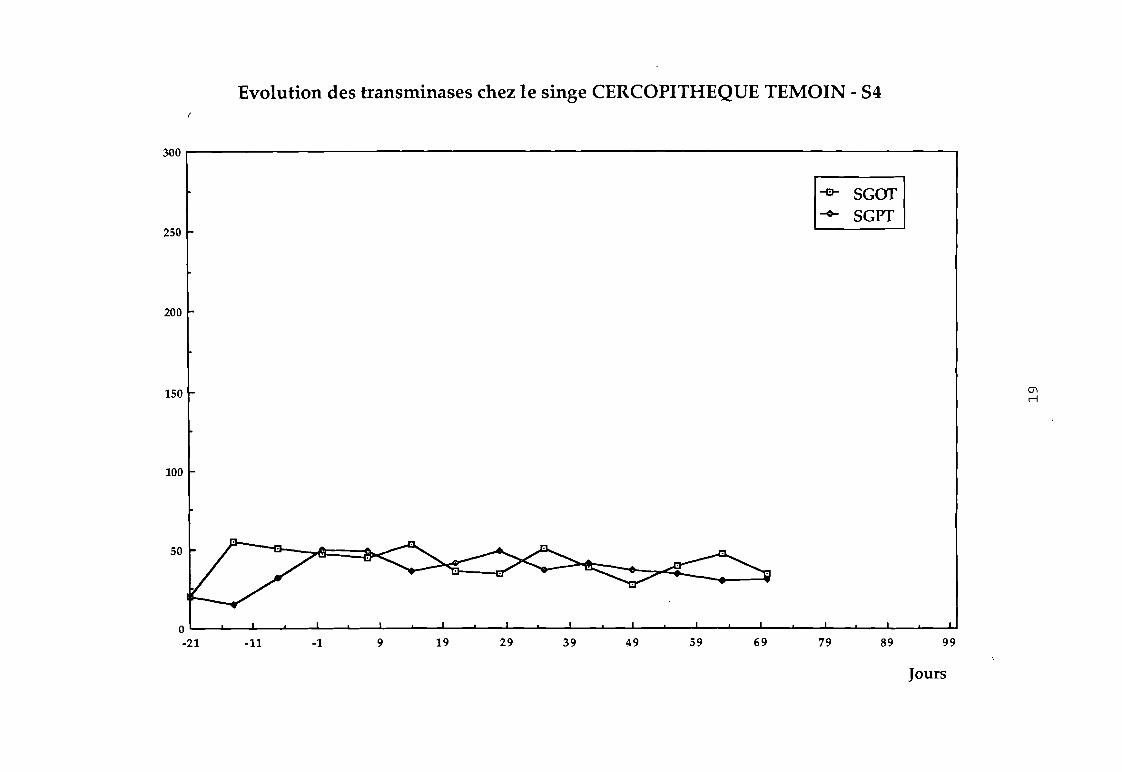
Evolution des transminases chez le singe CERCOPITHEQUE TEMOIN - 54
300 1 1
250
I~ SGor1-+ SGPT
200
150 0'\ri
100
50
9989796959493929199-1-11
o l , , , l , , , l , l , l , l , l , l , l , l , Il
-21
Jours
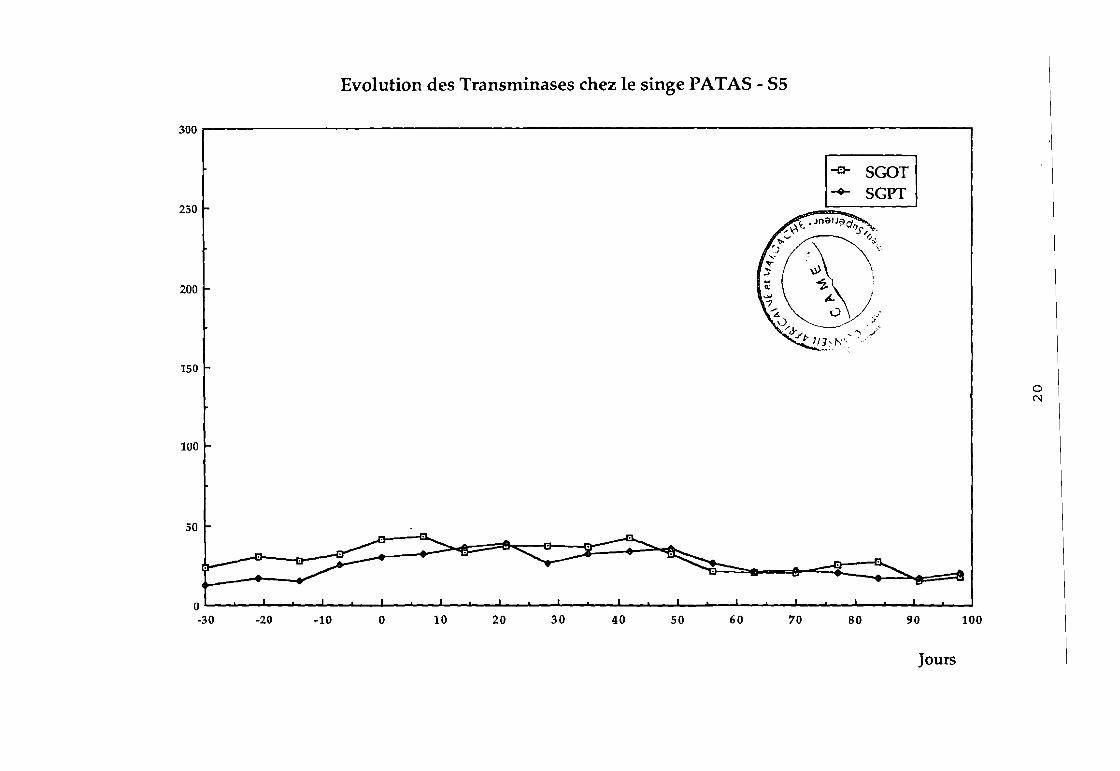
Evolution des Transminases chez le singe PATA5 - 55
300 i i
250
200
I~ san 1-+- SGPT
150
oN
100
50
100908070605040302010o-10-20
o 1 ft 1 ft , J , ,1ft l , l , l , 1 t 1 ! ' , 1 1 1 , J
-30
Jours
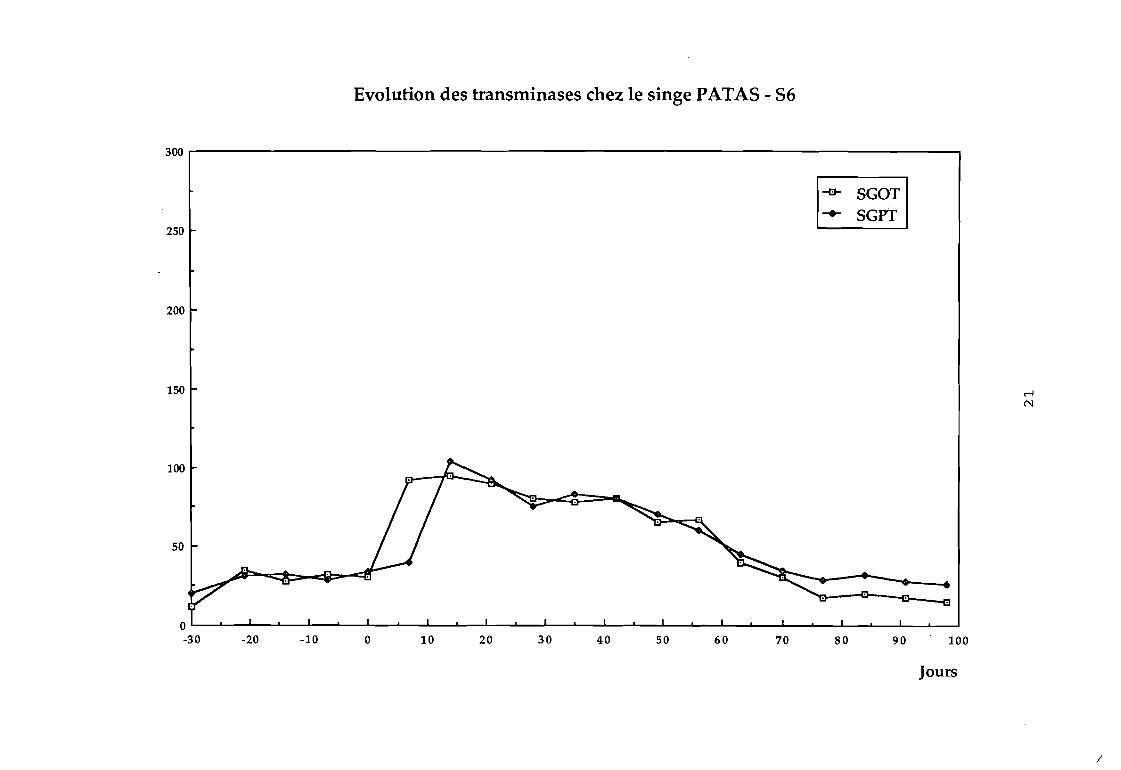
Evolution des transminases chez le singe PATA5 - 56
300. i
-a- SGOT...... SGIT
250
200
150 r-iN
100
50
100908070605040302010o-10-20
o ' , l , l , , ,1 ft l , l , , ft , , , ft 1 1 l , 1 1 1
-30
Jours
/
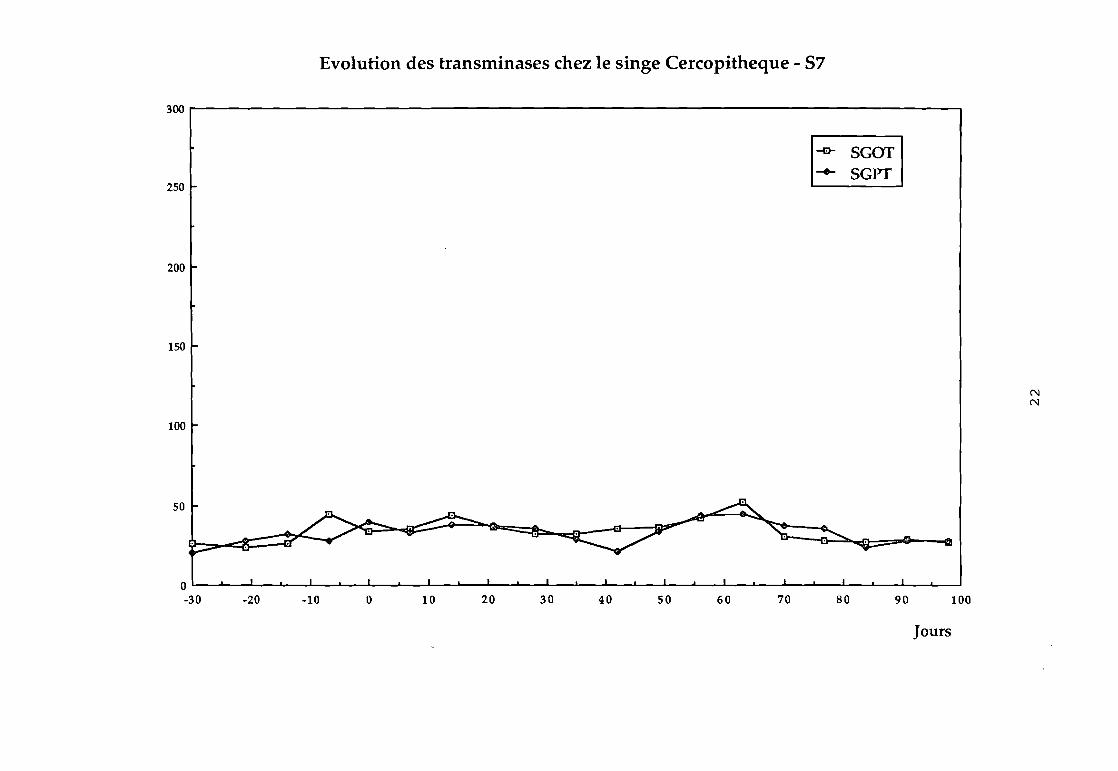
Evolution des transminases chez le singe Cercopitheque - 57
300' ,
250
I~ SGOT1...... SGPT
200
150
NN
100
50
100908070605040302010o-10-20
o ' , l , , 1 l , , ft l , 1 1 l , , • l , l , 1 1 l , 1
-30
Jours
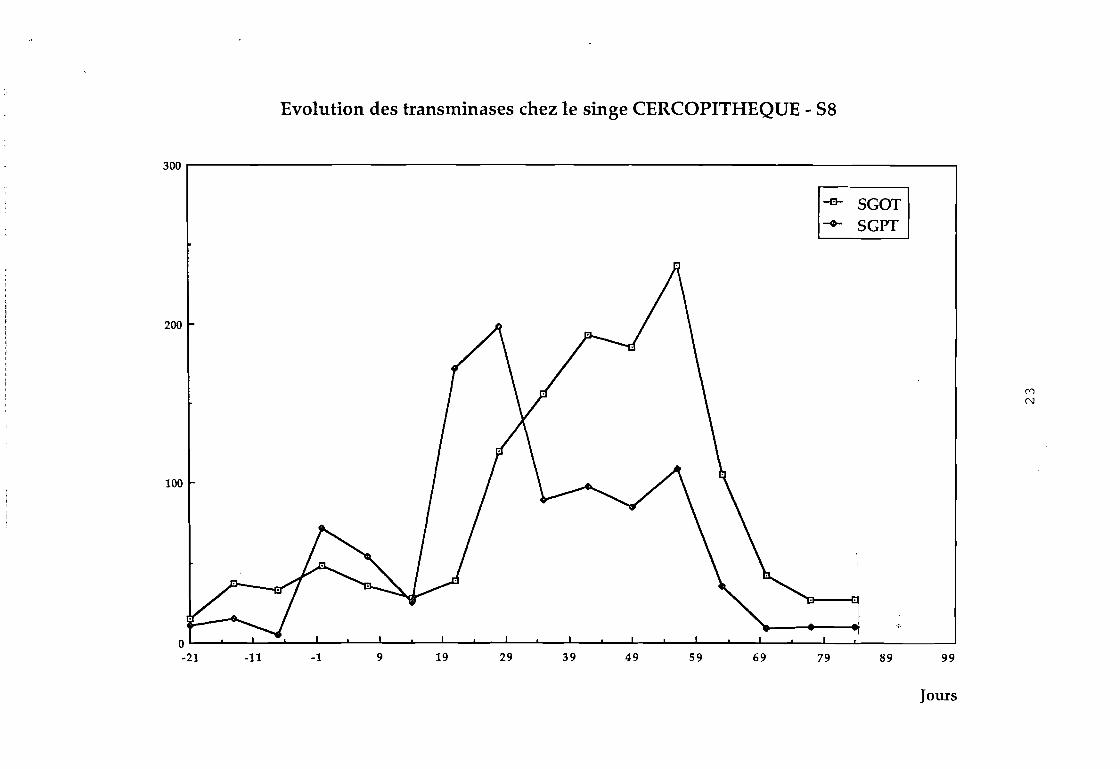
Evolution des transminases chez le singe CERCOPITHEQUE - S8
300. 1
1- SGar 1...... SGPT
MN
99
.,~,.
89796959493929199-1-11
o l , 1 l , , , , , , , 1 l , , , l , l , 1 1 1
-21
200
100
JOUIS

3. Tests d'inoculations croisées
Ne disposant d'aucun test biologique de dépistage des
marqu~urs du VHE, et pour vérifier que le virus inoculé était
différent du VHC et ne possédait aucune communauté antigénique
avec ce dernier, des tests d'inoculations croisées ont été
pratiqués.
a) Technique
Ce test porte sur deux singes type Cercopithèque: l'un S9
fut infecté expérimentalement par des selles de sujets atteints
d'hépatite NANB épidémique l'autre, SlO, par le sérum d'un
malade atteint d'une hépâtite NANB post-transfusionnelle.
S9 4 semaines après l'inoculation, une élévation importante
des transaminases fut observée suivie d'un retour à la
normale 4 semaines plus tard (cf courbe) .
S10 : Une élévation plus modérée des transaminases fut obtenue
au bout de 5 semaines avec retour à la normale 4 semaines plus
tard. C'est à ce moment là que les inoculations croisées furent
pratiquées, c'est-à-dire que le singe S9 était inoculé avec le
sérum d'un malade atteint d'hépatite C à la phase aiguë et le
singe S10 avec les selles de sujets atteints d'hépatite E pendant
la montée des transaminases.
b) Résultats
si l'on exclut les formes d'hépatite à rechute, on peut
conclure que le virus inoculé est différent du VHC et qu'il ne
possède aucune communauté antigénique avec celui-ci; puisque une
élévation des transaminases sériques, témoin d' une hépatite
clinique, fut observée 20 jours après ces réactions croisées (cf
courbes S9, 810).
24
Evolution des transminases chez le singe - 59
400 1 1
350l;r SGar 1-+- SGPr
300
250
l!)
N
100 110 120 130 140 150908070605040302010oo l , 1 ?" l , l , l , l , l , 1 l , 1 1 1 1 l , l , 1 l , l , , , l , l , 1
-30 -20 -10
50
100
150
200
Jours
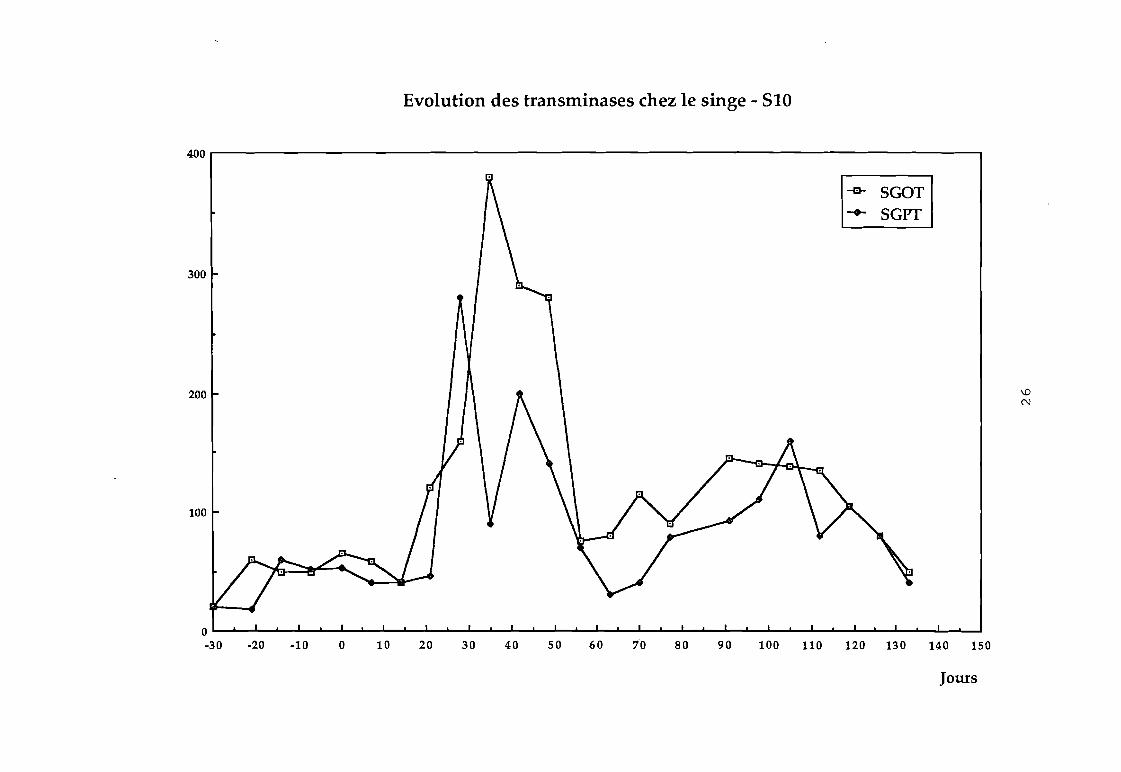
Evolution des transminases chez le singe - 510
400 1 1
300
-Go SGOT-+- SGPT
200
100
\.0N
100 110 120 130 140 150908070605040302010oo 1 ft , , , , , , , , l , l , l , , ft , , , l , , l , l , , e , , , , , l ,
-30 -20 -10
Jours

PURIFICATION DES IMMUNOGLOBULINES
ET MARQUAGE

A N T l - V H E
DES U JET S
LEV H E
l - PRE PAR A T ION DES l g G
A PAR TIR DES E R U M S
l N FEe TES PAR
1) Matériel
il s'agit de sérum de malades atteints d'hépatite virale à
transmission hydrique, survenue à Bangkok lors d'une épidémie.
Les sérums ont été prélevés chez ces sujets pendant la phase
convalescence de la maladie, c'est-à-dire après le retour à la
normale des transaminases.
2) Méthode
La préparation se fait à partir des sérums des malades
convalescents par relargage à l'aide de sels neutres.
a. Principe
Le sulfate d'ammonium à demi-saturation précipite les
fractions riches en IgG.
b. Technique
- Préparation de la solution saturée de sulfate d'ammonium
Dissoudre 1 g de sulfate d'ammonium par ml d'eau distillée à
50°C. Ajouter du sulfate jusqu'à saturation, ajuster le pH
à 7.
Saturation du sérum
Les IgG précipitent en grande partie à 40 % de saturation.
Il faut calculer la quantité de solution saturée à utiliser
par la formule
saturation 2 - saturation 1
1 - saturation 2
Saturation 1 degré de saturation de départ du sérum
Saturation 2 degré de saturation que l'on veut obtenir.
Exemple : à partir de sérum frais, combien de ml de sulfate
d'ammonium saturé faut-il ajouter pour obtenir une solution à
40 % ?
27

0,40 - ° 0,4 = 0,66 soit 66 %
1 - 0,4 0,6
Donc pour 100 ml de sérum, il faudra ajouter 66 ml de solution
saturée de sulfate d'ammonium.
- Mettre le mélange une nuit à +4°C.
- Centrifuger à faible vitesse (2 000 t/mn) pendant la minutes
et recueillir le culot de centrifugation qui contient les IgG.
Le culot est alors repris dans du tampon PBS sous un volume
de 1 ml.
Dessaler cette solution en dialysant dans du tampon PBS
pendant 48 heures.
- Vérifier au troisième jour que l'eau de dialyse ne contient
plus de traces de sulfate d'ammonium en effectuant la réaction
de Nessler.
- Réaction de Nessler
A 2 gouttes d'eau de dialyse, ajouter 0,7 ml de réactif de
Nessler, puis 3 ml de lessive de soude l' apparition d'une
coloration rouille indique la présence de sulfate d'ammonium.
Dialyser à nouveau jusqu'à disparition de cette coloration de
l'eau de dialyse. Les IgG ainsi préparées peuvent être conservées
à +4°C.
28

II SEP A RAT ION
A N TIC ° R P SAN T l
DE SINGES
DES
- V H E (IgM - IgG)
1. préparation des IgM antiVHE
Elle se fait à partir d'un pool de sérums du singe S8
recueillis pendant la montée des transaminases par un
fractionnement en gel filtration sur colonne.
a) - principe
C'est une méthode de filtration sur tamis moléculaire qui
sépare des protéines de même solubilité mais de poids
moléculaires différents.
b) - Technique
Préparation de la colonne
Elle nécessite un choix judicieux de la colonne, du tampon
et du gel.
Choix de la colonne : Il dépend de la quantité de produit
à injecter et de son volume.
5 à 10 ml de sérum : colonne Pharmacia K50-100 (soit 5 cm de
diamètre, 100 cm de longueur). Volume de la colonne = 1,8 1.
1 à 2 ml de sérum : colonne Pharmacia K25-100 ou SR25-100 ou
colonne Wright GA-22 x 90. Volume de la colonne = 530 ml.
0,5 ml : colonne Pharmacia K15 x 100 ou K16 x 100 ou colonne
Wright GA-16 x 90. Volume de la colonne = 200 ml.
0,25 à 0,5 ml de sérum: colonne Pharmacia K9-60 ou colonne
Wright GA-10 x 60. Volume de la colonne = 38 ml.
Débit de la colonne : il dépend de sa taille, de la nature
du gel et de la qualité de la séparation. Les valeurs moyennes
sont" les suivantes
- pour une K50-100 30 à
- pour une K25-100 7 à
- pour une K15-100 2,5 à
choix du gel en fonction
Poids moléculaire > 1 million
CLB) .
50 ml/heure
15 ml/heure
8 ml/heure.
du poids moléculaire à séparer
: Sepharoses (notamment Sepharose
29

Poids moléculaire compris entre 80 mille et 1 million Sephacryl
Sephacryl
magnétique, puis
(sinon risque de
le tampon est
moitié dans du
S300.
Poids moléculaire compris entre 20 mille et 150 mille
S200.
Poids moléculaire < 20 mille: Sephadex G75 ou G50.
Choix du tampon : Habituellement, on utilise du tampon
PBS-azide à 1/1000 0,15 M PH 7,2.
Remarque : Le tampon utilisé, de même que le gel doivent être
dégazés.
Comment dégazer le tampon ?- Dans une fiole à vide, mettre environ deux fois le volume de
la colonne de tampon.
- Mettre sous agitation magnétique faible.
Recouvrir avec un bouchon trop large, puis relier à une trompe
à eau puissante pendant un quart d'heure.
- Pour casser le vide, arrêter l'agitateur
retirer le tuyau avant d'arrêter la pompe
reflux) .
Comment dégazer le gel ?
La même technique que celle pour dégazer
utilisée, mais diluer préalablement le gel de
tampon dégazé.
- Mettre sous agitation lente afin de ne pas briser les billes
de gel.
Installation de la colonne
La colonne est installée verticalement (vérifier dans toutes
les directions avec un niveau à eau).
- installer un réservoir en haut de la colonne
en bas de la colonne, mettre un tuyau de longueur supérieure
à celle de la colonne
- remplir entièrement la colonne avec du tampon dégazé et purger
le tuyau de sortie
- verser le gel doucement dans la colonne au-dessus du tampon
dans lequel il tombe en formant des volutes
installer la pompe qui doit être reliée à l ' extrémité
inférieure de la colonne par un tuyau le plus court possible,
30

et dont le débit est supérieur au débit d'utilisation pour
permettre au gel de se tasser.
- laisser le gel se tasser. Le gel est correctement tassé
lorsque son niveau supérieur dans le haut de la colonne ne
varie plus
- enlever l'excès de gel
- enlever le réservoir
- installer l'adaptateur en l'enfonçant légèrement dans le gel
le visser solidement
Pour purger l'air du tuyau du bas, laisser couler l'eau de
la colonne. Pour purger la pompe, la faire marcher à vide avant
de la relier à la colonne. Le tuyau d'arrivée à la pompe plonge
dans le tampon par l'intermédiaire d'un microtube en verre qui
l'empêche de flotter. Le tuyau de sortie de la colonne est relié
directement au collecteur de fractions où la collection se fait
en gouttes (100 gouttes = environ 5 ml) ou en temps. Ce
collecteur est en relation avec un enregistreur qui inscrit la
forme globale de la courbe.
La lecture
Les fractions sont analysées par absorption dans l'UV à 280
nm. Les densités optiques obtenues au spectrophotomètre sont
reportées sur une coufbe en fonction du volume d'élution : la
courbe présente alors trois pics s'il s'agit d'un sérum:
1er pic correspondant aux IgM (PM = 1000 Kd)
2éme pic est celui des IgG (PM = 150 Kd)
3éme pic représente l'albumine (PM = 68 Kd).
Dans notre étude, pour la séparation des immunoglobulines,
nous avons choisi une colonne Pharmacia K2S-100, le gel Sephacryl
S300 et du tampon PBS azide à 1/1000 comme éluant.
31
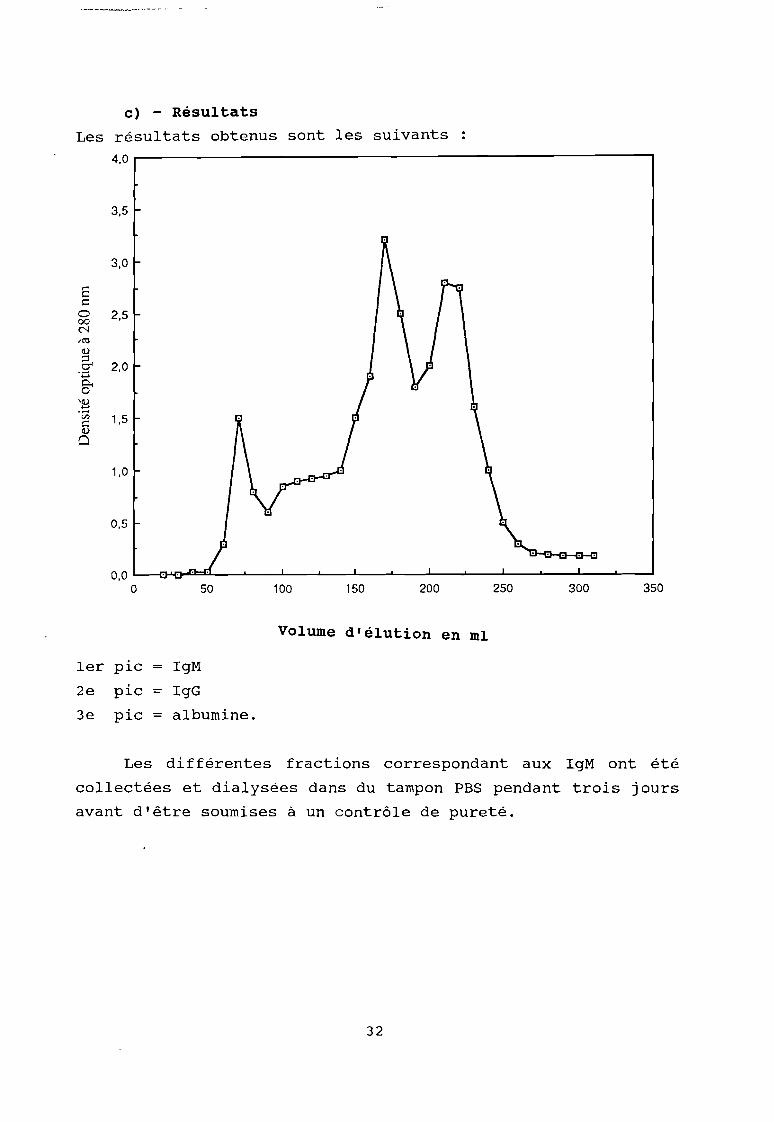
c) - Résultats
Les résultats obtenus sont les suivants
4,0
3,5
3,0
8t::
0 2,500N,C\i0:::lcr' 2,0
'P§<'0~.....en 1,5t::00.
1,0
0,5
0,00 50 100 150 200 250 300 350
Volume d1élution en ml
1er pic IgM
2e pic = IgG
3e pic = albumine.
Les différentes fractions correspondant aux IgM ont été
collectées et dialysées dans du tampon PBS pendant trois jours
avant d'être soumises à un contrôle de pureté.
32

2. préparation des IgG antiVHE
On utilise un pool de sérums du singe S8 recueillis pendant
la phase de convalescence de la maladie, que l'on fractionne par
chromatographie pur résines échangeuses d'ions. Nous avons
utilisé comme support: DEAE Trisacryl M, IBF.
a) - principe
Il est basé sur la propriété des protéines à s'associer aux
groupes chargés fixés sur les résines. Plus la protéine est
chargée, c'est-à-dire éloignée de son point isoélectrique, plus
fortement elle se fixe. On obtient l' élution en augmentant
progressivement la force ionique.
b) - Technique
Après avoir séparé les fractions d' IgM et d' IgG par
chromatographie sur S300, dialyser les IgG dans du tampon Tris
HCl 0,025 M, Nacl 0,035 M, pH 8,8 pendant 48 heures,
- Déposer l'échantillon sur la colonne à raison de 5 ml de
sérum pour 15 ml de gel équilibré dans le même tampon (volume
gel/volume d'échantillon = 3).
Le débit est de l'ordre de 50 ml/heure. Sous ces conditions,
seules les IgG ne sont pas retenues par le support et sont éluées
dans un pic étroit. Le degré de pureté des IgG éluées est de
99,5% .
Remarque : La capacité maximale de la DEAE Trisacryl M est de
50 mg de protéines par ml de gel. Cette technique est applicable
à différentes espèces animales mais avec un degré de pureté
variable.
Espèce Pureté IgG
Rat ++
Souris
Lapin ++
Mouton ++
Chèvre +
Porc ++
Chien ++
Homme +
33

3. Contrôle de pureté des IqM et IqG préparées
Il se fait par immunodiffusion sur gel suivant la technique
classique d'outcherlony.
a) - principe
L'antigène en solution diffuse dans un gel contenant
l'anticorps. Les deux réactifs (antigène et anticorps) diffusent
l'un vers l'autre dans une couche de gel vierge et lorsqu'ils se
rencontrent, il y a formation d'un arc de précipitation.
b) - Technique
Préparation du gel
Le gel est composé d'agarose = 0,6 g
PEG 6000 3 g
tampon 100 ml
Tris 0,02 M 2,42 g
glycine 0,01 M 0,75 g
EDTA 0,01 M 2,9 g
eau QSP = 1 000 ml
agitateur chauffant et il est
Le gel est alors coulé sur une
Le tampon est ainsi préparé
Le mélange se fait sur
conservé en chambre humide.
surface de verre (plaque de verre ou boîte de Pétri) et après
solidification, des sources de diffusion sous forme de puits
circulaires sont creusées avec un emporte-pièce dans la gélose.
On dépose dans deux puits différents l'antigène et l'anticorps,
et la diffusion se fait pendant 48 h en chambre humide.
c) - Résultats
Par cette technique nous avons pu contrôler que les
immunoglobulines que nous avons préparées étaient pures.
34

4. Concentration des immunoglobulines préparées et
détermination de la teneur en protéines
Cette concentration se fait par ultrafiltration sous
différence de pression.
a) - Technique
Dans une fiole à vide, on place un boyau à dialyse
parfaitement étanche dans lequel on introduit la suspension
d'immunoglobulines à concentrer. La fiole est reliée à une trompe
à eau et bouchée hermétiquement. Il règne ainsi dans la fiole,
une pression inférieure à celle existant dans le boyau, de telle
sorte que l'eau puisse s'échapper du boyau alors que les
protéines sont retenues à l'intérieur du sac à dialyse qui
constitue en fait une membrane semi-perméable. La teneur finale
en protéines est évaluée au spectrophotomètre UV à 280 nm.
35

acétique carbonate
- sodium métapériodate
III - MAR QUA G EDE SIg G FRA C T ION NEE S
Nos IgG fractionnées ont été marquées soit pour usage
ultérieur en immunoenzymologie (peroxydase-bêta-galactosidase),
soit pour immunofluorescence (fluorescéine).
1) - pour immunoenzymoloqie
a) Marquage à la péroxydase
Réactifs
- eau parfaitement déionisée, bidistillée
- peroxydase
acétate de sodium anhydre Acide
disodique anhydre - carbonate monosodique
- sodium borohydrure.
Technique
- Premier jour A 8 mg de peroxydase dans 1 ml H20, ajouter 0,8
ml de NaI04 0,1 M (21 mg/ml) en agitant. Laisser 20 à 30 min
maximum à température ambiante. Dialyser cette solution une nuit
à +4°C contre un tampon acide acétique-acétate de sodium 10-3 M
pH4, 4 (1 1) (cf préparation des tampons).
Dialyser simultanément 5 mg d'IgG (volume compris entre 0,2 et
0,5 ml) contre du tampon carbonate 10-2 M pH 9,5 une nuit à +4°C
(2 XII) (cf préparation des tampons).
- Deuxième jour: Mélanger les solutions d'IgG et de péroxydase.
Volume final compris entre 2 et 2,5 ml. La solution se trouble
parfois : elle redevient limpide après alcalinisation.
Alcalinisation ajouter 40 à 60 pl de tampon carbonate 1 M.
Contrôler le pH. si pH < 9,0 ajouter du tampon carbonate 1 M par
aliquotes de 10 pl.
Laisser 3 heures à température ambiante à l'obscurité.
Ajouter 100 pl de NaBH4 à 4 mg/ml en eau distillée et laisser 2
heures à +4°C.
Dialyser contre du PBS (2 xlI) une nuit à +4°C.
- Troisième jour: Diluer à l'aide d'un tube gradué à 50 % en
glycérol. Bien homogénéiser.
Le conjugué est stable à -20°C pendant au moins un an.
36

b) Marquage à la bêta-galactosidase
Principe
La préparation du conjugué anticorpsjbêta-galactosidase se
fait par la glutaraldéhyde qui est fixée aux anticorps, selon la
technique de GUESDON-AVRAMEAS et TERNYNCK (34).
Cette méthode permet l'association de l'enzyme à la protéine
par l'intermédiaire des groupements aldéhydes de la
glutaraldéhyde qui réagissent avec les groupements aminés de
l'anticorps et de l'enzyme.
Technique
La bêta-galactosidase, à raison de la mg est additionnée
de 5 mg d'anticorps sous un volume total de 2 ml ; le mélange
est dialysé toute la nuit contre du tampon phosphate 0,1 M pH
6,8 (cf préparation des tampons). On ajoute le lendemain à ce
mélange, maintenu sous agitation, 100pl d'une solution de
glutaraldéhyde à 1 % dans du tampon phosphate 0,1 M pH 6,8. Après
3 heures de contact à la température du laboratoire, on ajoute
une solution de lysine (ou de tout autre réactif contenant des
groupements NH2 libres) en tampon phosphate 0,1 M pH 6,8 pour
obtenir une concentration finale en lysine de 0,1 M. Celle-ci,
laissée en contact pendant 2 heures à la température du
laboratoire, permet le blocage de l'excès de glutaraldéhyde.
Conservation
Les conjugués à la péroxydase ou à la bêta-galactosidase,
sont enfin dialysés contre du PBS pendant 3 jours à +4°C, puis
centrifugés 20 minutes à 3.000 g, et un volume égal de glycérol
est ajouté comme conservateur. Le mélange peut alors être
conservé sans antiseptique à -20°C.
37

2. pour immunofluorescence
a) Marquage des IgG par la fluoresceine
Technique
On utilise l'isothiocyanate de fluorescéine Mérieux (ITCF).
La solution de protéines à conjuguer est ajustée à 20 mg/l
(mesure de la DO à 280 nm avec précision). Avant de mettre
l'ITCF, il faut ajuster le pH à 9 en ajoutant l/lOème du volume
de tampon carbonate O,S M aux protéines.
L'isothiocyanate de fluorescéine ne se dissout qu'à PH 9.
Ajouter l'ITCF à la solution de protéines à conjuguer en raison
de 1 mg pour 40 mg de protéines
- Laisser en agitation douce et continue une nuit à +4°C
- Séparer alors les protéines marquées sur Sephadex GSO.
Séparation des protéines marquées sur Sephadex GSO
- Laver le gel avec du PBS 2 à 3 fois puisque le gel est en
tampon phosphate. Pour celà centrifuger à 3.000 tours/minute
pendant S minutes et éliminer le surnageant
- Verser alors le gel dans une colonne de manière à ce que la
hauteur de la colonne soit dix fois supérieure à son diamètre.
Il faut S ml de gel pour 1 ml de solution à chromatographier ;
-Laver la colonne en continu pendant 40 minutes environ avec
l'eau physiologique tamponnée:
- Déposer délicatement la solution conjuguée sur le papier filtre
en haut du gel et laisser couler.
Résultats
Les protéines marquées sont éluées les premières et les
fractions correspondantes sont colorées, suivies des protéines
non marquées dont les fractions correspondantes sont incolores.
L'ITCF libre est éliminée dans les dernières fractions colorées.
On fait un pool des premières fractions colorées qui ont
été recueillies et on évalue la teneur en protéines à 280 nm
puis à 496 nm.
La concentration finale en protéines exprimée en ng/ml est
donnée par la formule
38

DO 280 nm - 0,46 (DO 496 nm)
p =-~---------------------------
J~ :Rj 0,46
1,4
est
1,4
= concentration en protéines
= facteur de correction dû à l'ITCF
= absorption dans l'UV de 1 mg de protéines.
Après quoi, on calcule le poids de l'ITCF en gamma/ml. Il
donné par la formule :
DO (496)
F = -------0,21
1 gamma d'ITCF a une absorption de 0,21 à 496 nm
Calculer le rapport F/P. Ce rapport doit être compris entre
3 et 6. Les immunoglobulines marquées à la fluorescéine ainsi
préparées sont épuisées sur de la poudre de foie de boeuf.
Technique d'épuisement
- On utilise 100 mg de poudre de foie de boeuf pour 20 mg de
protéines marquées ;
- A cette poudre réhydratée au préalable, ajouter la solution
de protéines marquées ;
- Laisser le mélange une heure à la température du laboratoire,
puis une nuit à +4°C.
Cette méthode permet de capter tous les anticorps anti-foie,
il ne reste que les anticorps anti-virus donc toute fluorescence
observée au microscope sera spécifique du virus.
De plus les immunoglobulines non marquées sont faiblement
chargées tandis que celles qui sont marquées sont plus chargées
négativement. Dans ce cas, les anticorps anti-tissus ou anti
cellules qui portent des charges positives vont se lier par des
liaisons électrostatiques avec les protéines marquées à la
fluorescéine. Il s'agit de liaisons non spécifiques.
L'épuisement par la poudre débarrasse de ces forces
électronégatives donc les seules liaisons présentes seront
ensuite des liaisons immunologiques entre le virus et les
anticorps marqués.
Remarque : A la place de la poudre de foie, on peut utiliser un
sérum polymérisé avec du glutaraldéhyde qui donne un gel. La
technique d'épuisement reste inchangée.
39

3. Contrôle du marquage
a) - Matériel
- Plaques à microtitration
SI : substrat spécifique pour béta-galactosidase (ONPG 8 mg dans
2 ml d'H20+ PM2 8 ml + mercaptoéthanol 56 pl).
S2 : substrat spécifique pour péroxydase (OPD 2 mg/ml + tampon
citrate +1 ul/ml d'H202).
- Tampon de saturation (BSA 0,5/1000 + Tween 20 à 1/1000 + PBS)
- Tampon de lavage (PBS + Tween 20 à 1/1000)
- R3 : IgG de lapin anti-Fc(gamma) humain dont la méthode de
préparation est décrite en annexe).
b) - Technique
Sur une plaque Nunc, incuber à 37°C l'anti-Fc (gamma) à une
concentration de 5 gamma/ml de PBS, puis une nuit à +4°C ;
- A l'aide du tampon de saturation (200 Pl/puits), saturer la
plaque 2 à 3 heures à 37°C;
- Laver avec la solution de lavage
Ajouter l'IgG marquée diluée dans le tampon de saturation à
40 gamma/ml et incuber 1 heure à 37°C;
- Laver plusieurs fois.
si l'IgG est marquée à la bêta-galactosidase, ajouter 100
pl de substrat 1, puis 50 pl de carbonate de Na et lire à 405
nm. si l'IgG est marquée à la péroxydase, ajouter 100 pl de
substrat 2, puis 50 pl d'acide sulfurique 4N et lire à 496 nm.
40

METHODES ANALYTIQUES

RECHERCHE DES ANTICORPS
ANTIVHE DANS LE SERUM

ETUDE DE LA REACTION IMMUNOGLOBULINE FLUORESCENTE
SUR COUPE DE FOIE DE SINGE INFECTE
1. Principe
Des coupes de foie de singe infecté expérimentalement sont
mis en contact avec les IgG marquées à la fluorescéine des
malades à tester.
si le virus est présent dans les coupes de foie, il se forme
une réaction antigène/anticorps que l'on visualise au microscope
à irnrnunofluorescence.
2. Technique
La technique utilisée est l'irnrnunofluorescence directe
- A partir des biopsies de foie de singe infecté par le VHE et
montées sur cryobed, on réalise à l'aide d'un cryostat des coupes
de 5 u que l'on dépose sur lames
- Sécher à l'air libre et fixer à l'éther pendant 5 minutes.
Ajouter 50 à 100 fI d'IgG de malades à tester, marquées à la
fluorescéine ;
Incuber 30 minutes à 37°C en chambre humide ~
- Laver 3 ou 4 fois en PBS puis en eau distillée
- Monter en glycérine tamponnée avec une lamelle
- Observer à l'aide d'un microscope à irnrnunofluorescence.
3. Résultats
Nous avons utilisé comme source d'antigène VHE le foie du
singe Patas infecté expérimentalement prélevé chaque semaine par
biopsie à l'aiguille, après inoculation.
41

Les résultats ont été constamment négatifs.
sérum de singe
Avant inoculation
J7
(7 jours après inoculation)
J14
J21
J30
J37
J44
42
IFD

RECHERCHE DE L'ANTIGENE VHE DANS
LES SELLES DE SUJETS INFECTES

l - RECHERCHE DE LIANTIGENE VHE PAR IMMUNOMICROSCOPIE
ELECTRONIQUE
1. Technique
- Des suspensions à 10 % en PBS ont été réalisées avec les selles
du singe S8 recueillies quotidiennement
- centrifuger à 5.000 tours/mn pendant 30 minutes
- Passer sur filtre 0,22 p- A partir d'l ml de filtrat, faire une ultracentrifugation à
30.000 tours/mn pendant 4 heures soit 100.000 g (BECKMANN R50)
- Reprendre le culot dans un volume minimum de surnageant (50
ul environ)
- Faire une coloration négative en mettant en contact une goutte
du culot viral avec une goutte de réactif phospho-tungstique (cf
préparation en annexe)
- Déposer une goutte du mélange sur une grille et laisser sécher
- Observer au microscope électronique.
Par cette technique, nous n'avons pas pu détecter le VHE
dans les selles que nous avons testées, ce qui nous a conduit à
améliorer cette technique de la manière suivante :
- A deux gouttes de la suspension du culot viral, ajouter une
quantité équivalente d'IgM de malades atteints d'hépatite E
- Mettre à l'étuve à 37°C pendant une heure
- Aj outer alors l' anti-p commercial (monoclonal anti-IgM humaine)
dilué au l/lOOème
- Faire une ultracentrifugation à 95.000 tours/mn (400.000 g)
pendant 5 minutes sur BECKMANN TL 100.
- Remettre le culot en suspension dans un volume minimum de
surnageant
- Observer alors au microscope électronique.
2. Résultats
Ils sont restés tout aussi décevants : la recherche du VHE
dans les mêmes selles que précédemment, s'est avérée négative.
43

fait par
de l'effet
II - RECHERCHE DU VHE PAR CULTURE CELLULAIRE
1. Culture des lignées cellulaires utilisées
Les lignées cellulaires utilisées sont :
- La lignée PLC PRF5 (Alexander) = lignée d'hépatocytes dérivée
d'un hépatocarcinome d'un sujet porteur d'antigène HBs.
- La lignée HepG2 = lignée dérivée d'un hépatoblastome humain.
Ces cellules se cultivent dans des boîtes de culture
stériles (Falcon) en plastique de 25 cm2, dans une étuve à 37°C
en atmosphère à 5 % de C02.
Le milieu de culture est
Le milieu Dulbecco MEM (GIBCO-BRL) additionné de 10 %
de sérum de veau foetal.
2. Inoculation des cultures cellulaires
- Les extraits de selles supposés contenir le VHE sont passés
sur filtre 0,22 p puis dilués dans le milieu de culture sans
sérum de veau foetal.
- Déposer 0,5 ml de cet extrait sur la culture.
- Incuber 1 heure à 37°C.
- Ajouter alors 4,5 ml de milieu complet (l'inoculum est alors
au 1/10ème).
Le premier renouvellement du milieu se fait 24 heures après,
puis toutes les 48 heures.
3. Lecture
La recherche de l'antigène VHE se
immunofluorescence indirecte, et la recherche
cytopathogène par microscope électronique.
Recherche de l'antigène VHE par immunofluorescence indirecte.
Mettre sur les cellules inoculées 50 à 100 pl de sérum
contenant des immunoglobulines antiVHE, et dilué en PBS (dilution
minimale au 1/20ème)
- Laisser incuber à 37 OC en chambre humide une à deux heures.
44

- Laver en tampon PBS trois fois.
Ajouter 50 à 100pl d'antiglobulines humaines marquées à la
fluorescéine (antilgG, M humaines Pasteur ou Mérieux) diluées
en PBS.
- Faire une contre coloration au Bleu Evans (5mn).
- Laver en eau distillée et monter en glycérine tamponnée avec
une lamelle.
Recherche de l'effet cytopathogène
Elle se fait directement au microscope électronique au
grossissement 40.000.
4. Résultats
En microscopie électronique aucun effet cytopathogène n'a
été observé de même la culture sIest avérée négative. Mais il
faut souligner à ce propos que jusqu'à ce jour aucune technique
de culture du VHE n'a pu être mise au point malgré des recherches
laborieuses menées par plusieurs équipes.
45
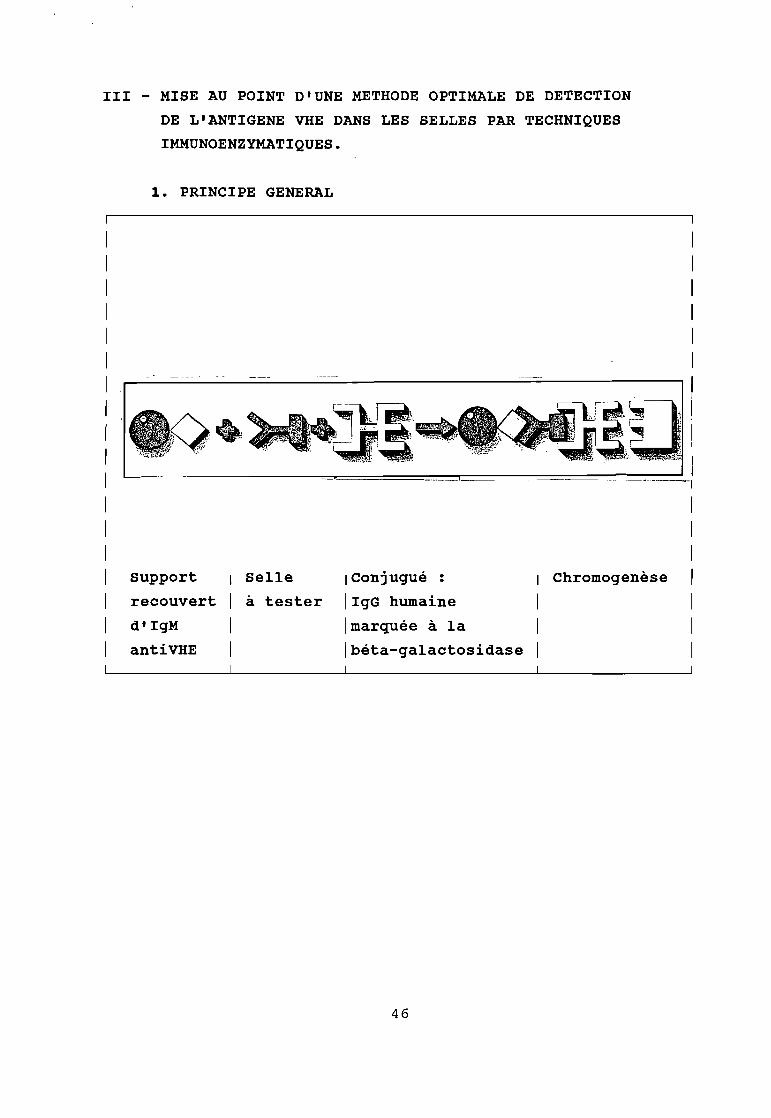
III - MISE AU POINT D'UNE METHODE OPTIMALE DE DETECTION
DE L'ANTIGENE VHE DANS LES SELLES PAR TECHNIQUES
IMMUNOENZYMATIQUES.
1. PRINCIPE GENERAL
Selle IConjugué :
à tester IIgG humaine
Imarquée à la
Ibéta-galactosidase
d'IgM
antiVHE
Support
recouvert
1
1
1
1
1
1
,..........:---=::...:...:..:..--,-:....::..---=:..=.:......---------=---------,1
~~U.!i. ~.- .~ 1
-------11
1
1
Chromogenèse 1
1
1
1
46

extraits de selles à tester et des
sains ne comportant aucun agent
2. TECHNIQUES
1) Technique par immunocapture des IgM
a) Matériel et réactifs :
- Plaques de microtitration,
- Tampon PBS
- Solution d'anti-p commercial,
- Sérum total de malades en phase aiguë d'hépatite E,
- R1 = solution de lavage (PBS, Tween 20 : 1 %0),
- R2 = IgG humaine de sujets présumés convalescents d'hépatite
E,marquée à la péroxydase,
R3 = milieu réactionnel de l'enzyme comprimé d'OPD tampon
citrate + H202,
- R4 = solution d'arrêt (H2S044N).
b) Méthode
Premier jour
- Sensibiliser les plaques avec 100 pl/puits de solution d'anti
u à 5 gamma/ml de PBS. Placer les plaques 1 heure à 37°C puis une
nuit à +4°C.
- Laver alors 3 fois les plaques avec la solution R1. Ajouter
100 pl de sérums de sujets infectés dilués au 1/100e et incuber
1 heure à 37°C.
Deuxième jour
saturer les plaques avec une solution de bovalbumine à 1/1000
pendant 2 heures à 37°C;
- Préparer parallèlement les
selles témoins de sujets
infectieux (au moins 5)
- Pour celà,
centrifuger à
surnageant sur
- Laver 3 fois
faire une suspension de selles à 10 % en PBS,
5.000 tours/mn pendant 30 minutes. Passer le
filtre 0,22 p et recueillir le filtrat;
les plaques
47

infectés par VHE
la phase aiguë de
- Ajouter les extraits de selles à tester et les selles témoins
en raison de 100 pl/puits
- Mettre les plaques 1 heure à 37°C puis une nuit à +4°C.
Troisième jour
- Laver les plaques 3 fois ;
Mettre 100 pl de R2 à une concentration optimale qui est de 2
gamma/ml et incuber 1 heure à 37°C
- Laver plusieurs fois les plaques (au moins 3 fois) et ajouter
100 pl de réactif R3 à préparer extemporanèment
- Laisser agir 5 minutes à l'obscurité;
Arrêter la réaction avec 50 pl de R4/puits. Lire à 496 nm sur
lecteur plaque ELISA.
L' intensité de la coloration lue en densité optique, est
proportionnelle à la quantité de conjugué lié, et donc
éventuellement à la concentration d'antigène VHE présent dans
les échantillons de selles.
Les résultats sont exprimés en PIN, c'est-à-dire
DO de l'échantillon
moyenne des DO des témoins négatifs
Le résultat est considéré comme positif lorsque PIN > 2,1 soit
une DO supérieure à 2 a des DO témoins.
c) - Résultats
Nous avons testé des selles de sujets
lors d'une épidémie de Constantine, pendant
la maladie dans deux systèmes :
- Un système témoin constitué de sérum de sujet sain n'ayant
jamais été en contact avec le VHE et les IgG marquées à la
péroxydase du même sujet.
- Un système spécifique comprenant un pool de sérums de sujets
infectés par le VHE pendant la phase aiguë de la maladie, et les
IgG marquées à la péroxydase de ces mêmes sujets pendant la phase
de convalescence.
48
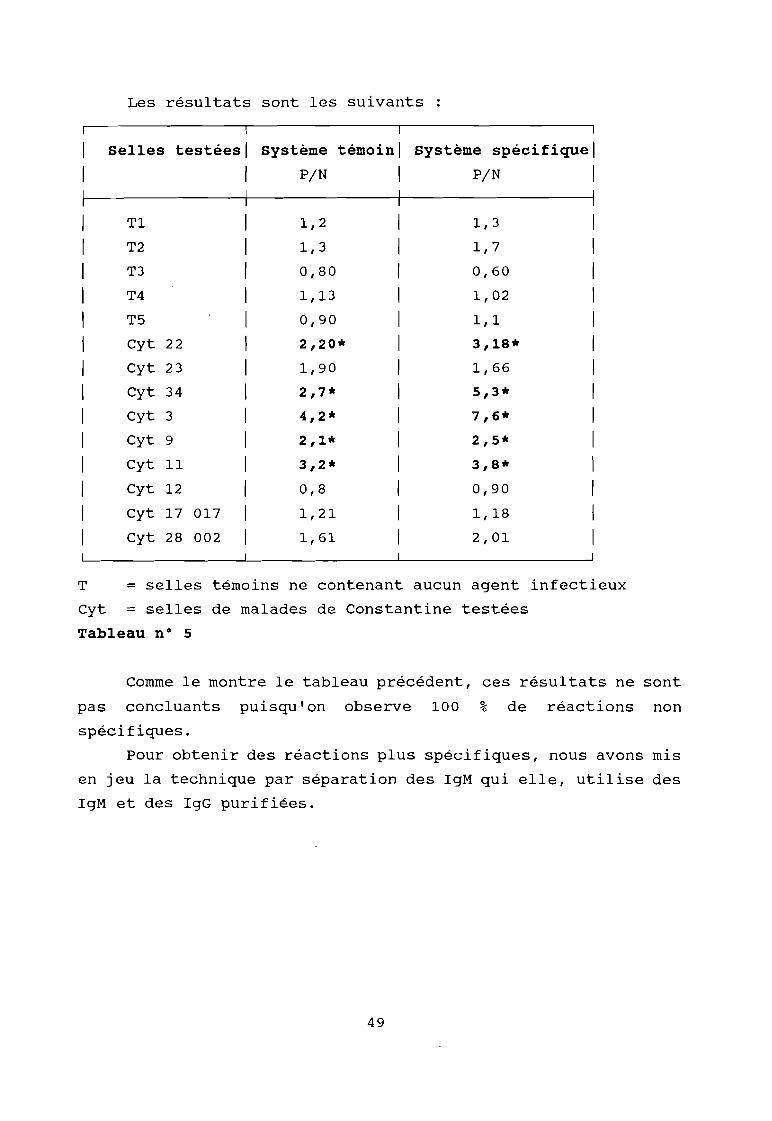
Les résultats sont les suivants
Selles testées
Tl
T2
T3
T4
T5
Cyt 22
Cyt 23
Cyt 34
Cyt 3
Cyt 9
Cyt 11
Cyt 12
Cyt 17 017
Cyt 28 002
Système témoin
PIN
1,2
1,3
0,80
1,13
0,90
2,20*
1,90
2,7*
4,2*
2,1*
3,2*
0,8
1,21
1,61
système spécifique 1
PIN 1
1
1,3
1,7
0,60
1,02
1,1
3,18*
1,66
5,3*
7,6*
2,5*
3,8*
0,90
1,18
2,01
T selles témoins ne contenant aucun agent infectieux
Cyt = selles de malades de Constantine testées
Tableau n° 5
Comme le montre le tableau précédent, ces résultats ne sont
pas concluants puisqu'on observe 100 % de réactions non
spécifiques.
Pour obtenir des réactions plus spécifiques, nous avons mis
en jeu la technique par séparation des IgM qui elle, utilise des
IgM et des IgG purifiées.
49

2. Technique par fixation directe des IgM sur support
a) Matériel et réactifs
Quatre systèmes ont été réalisés
- Un système témoin humain TH qui consiste à sensibiliser les
plaques à microtitration avec de l'IgM fractionnée de sujet sain
et révéler avec l'IgG marquée du même sujet.
- Un système témoin singe TS qui consiste à sensibiliser les
plaques avec l'IgM du singe S8 avant inoculation et révéler avec
l'IgG marquée du même singe avant inoculation.
- Un système spécifique humain SpH qui consiste à sensibiliser
les plaques avec l'IgM de sujet atteint d'hépatite E pendant la
phase aiguë et révéler avec l'IgG marquée du même sujet pendant
la phase de convalescence.
- Un système spécifique singe Sp qui consiste à sensibiliser les
plaques avec l'IgM du singe S8 pendant la montée des
transaminases et révéler avec l'IgG marquée du même singe après
retour à la normale des transaminases.
Les IgG utilisées ont été marquées avec la bêta
galactosidase et la péroxydase, et les substrats révélateurs de
ces enzymes sont respectivement
- le mélange: ONPG 4 mg/ml; PM2 ; bêta-mercapto~thanol 0,1 M
7 pl/ml PM2 ;
- l'OPD + tampon citrate + H202.
50

b) Méthode
Les plaques sensibilisées sont placées 1 heure à 37°C, puis
une nuit à +4°C.
- Laver les plaques avec le liquide de lavage PBS + Tween 20
à 1/1000.
- Saturer alors les plaques avec 200 ul de liquide de saturation
(BSA 0,5/1000, Tween 20 0,5/1000, PBS 100 ml)
- Placer les plaques à l'étuve à 37°C pendant 2 heures
Laver les plaques 3 fois et déposer les extraits de selles à
tester à raison de 100 pl/puits, laisser une heure à 37°C puis
une nuit à +4°C ;
- Laver les plaques 3 fois ;
Ajouter l'IgG marquée diluée dans le liquide de saturation à
la concentration de 1 gamma/ml
- Laver les plaques au moins 3 fois
- Ajouter les substrats spécifiques (100 pl/puib)
- Bloquer les réactions soit avec du carbonate 1,43 M si l'IgG
est marquée à la bêta-galactosidase soit avec l'acide sulfurique
4N si l'IgG est marquée avec la péroxydase.
c) - Résultats
51
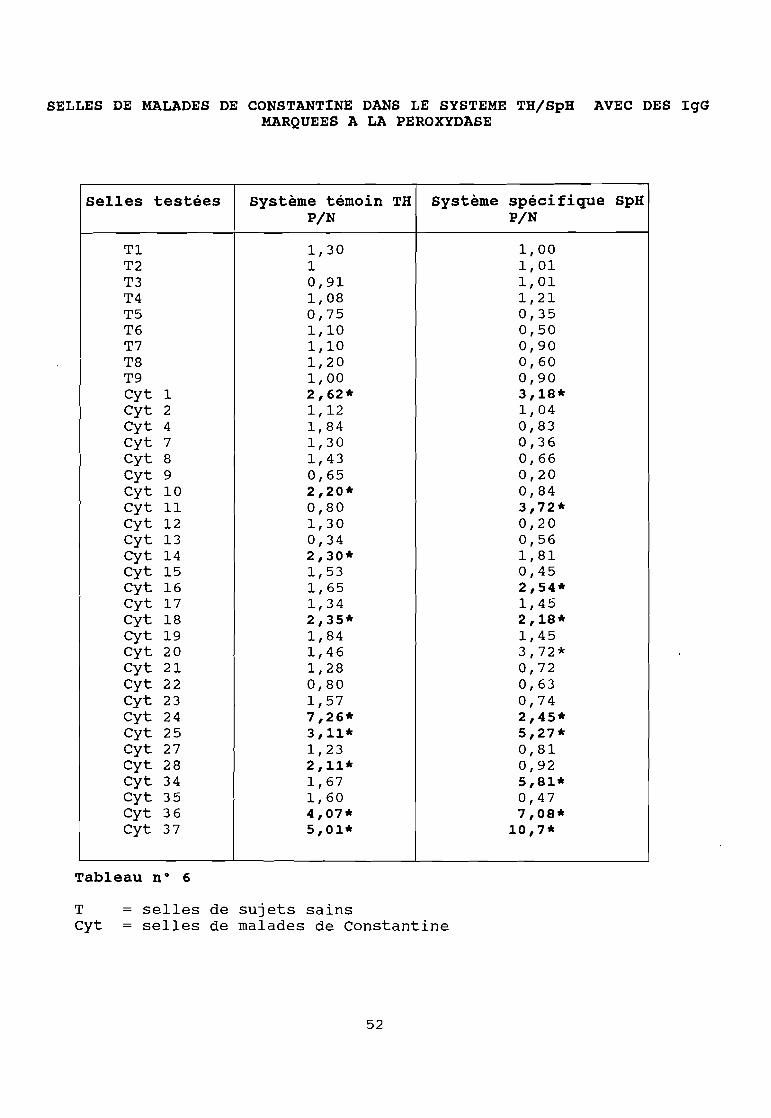
SELLES DE MALADES DE CONSTANTINE DANS LE SYSTEME TH/SpH AVEC DES IgGMARQUEES A LA PEROXYDASE
Selles testées système témoin TH système spécifique SpHPIN PIN
Tl 1,30 1,00T2 1 1,01T3 0,91 1,01T4 1,08 1,21T5 0,75 0,35T6 1,10 0,50T7 1,10 0,90T8 1,20 0,60T9 1,00 0,90Cyt 1 2,62* 3,18*Cyt 2 1,12 1,04Cyt 4 1,84 0,83Cyt 7 1,30 0,36Cyt 8 1,43 0,66Cyt 9 0,65 0,20Cyt 10 2,20* 0,84Cyt Il 0,80 3,72*Cyt 12 1,30 0,20Cyt 13 0,34 0,56Cyt 14 2,30* 1,81Cyt 15 1,53 0,45Cyt 16 1,65 2,54*Cyt 17 1,34 1,45Cyt 18 2,35* 2,18*Cyt 19 1,84 1,45Cyt 20 1,46 3,72*Cyt 21 1,28 0,72Cyt 22 0,80 0,63Cyt 23 1,57 0,74Cyt 24 7,26* 2,45*Cyt 25 3,11* 5,27*Cyt 27 1,23 0,81Cyt 28 2,11* 0,92Cyt 34 1,67 5,81*Cyt 35 1,60 0,47Cyt 36 4,07* 7,08*Cyt 37 5,01* 10,7*
Tableau n° 6
T = selles de sujets sainsCyt = selles de malades de Constantine
52
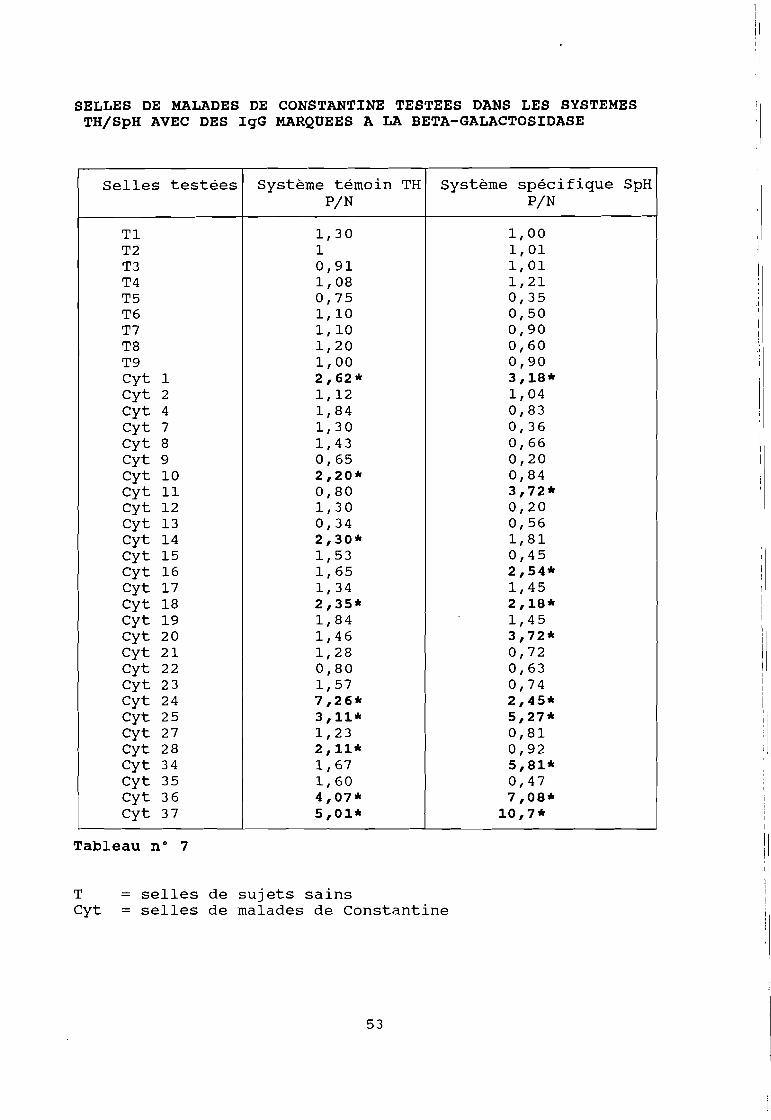
SELLES DE MALADES DE CONSTANTINE TESTEES DANS LES SYSTEMESTH/SpH AVEC DES IgG MARQUEES A LA BETA-GALACTOSIDASE
Selles testées Système témoin TH Système spécifique SpHPIN PIN
Tl 1,30 1,00T2 1 1,01T3 0,91 1,01T4 1,08 1,21T5 0,75 0,35T6 1,10 0,50T7 1,10 0,90T8 1,20 0,60T9 1,00 0,90Cyt 1 2,62 * 3,18*Cyt 2 1,12 1,04Cyt 4 1,84 0,83Cyt 7 1,30 0,36Cyt 8 1,43 0,66Cyt 9 0,65 0,20Cyt 10 2,20* 0,84Cyt Il 0,80 3,72*cyt 12 1,30 0,20Cyt 13 0,34 0,56Cyt 14 2,30* 1,81Cyt 15 l,53 0,45Cyt 16 1,65 2,54*Cyt 17 1,34 1,45Cyt 18 2,35* 2,18*Cyt 19 1,84 1,45Cyt 20 1,46 3,72*Cyt 21 1,28 0,72Cyt 22 0,80 0,63Cyt 23 l,57 0,74Cyt 24 7,26* 2,45*Cyt 25 3,11* 5,27*Cyt 27 1,23 0,81Cyt 28 2,11* 0,92Cyt 34 1,67 5,81*Cyt 35 1,60 0,47Cyt 36 4,07* 7,08*Cyt 37 5,01* 10,7*
Tableau n° 7
T = sellés de sujets sainsCyt selles de malades de Constantine
53
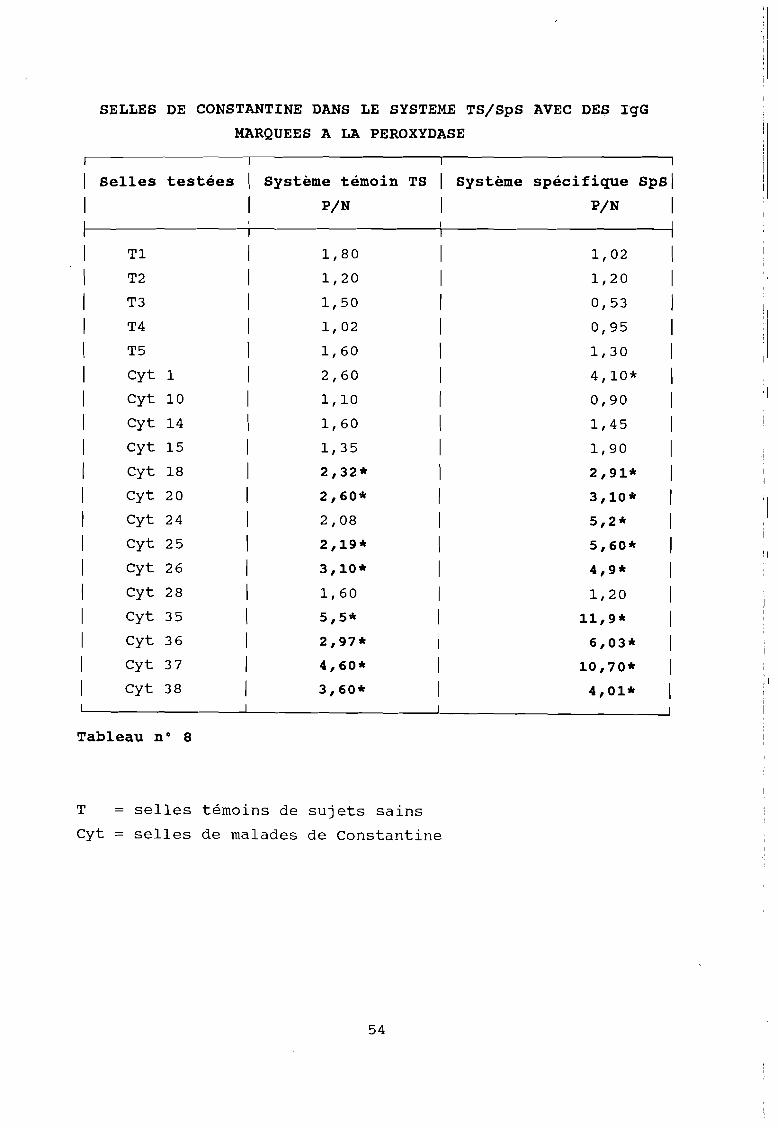
SELLES DE CONSTANTINE DANS LE SYSTEME TS/SpS AVEC DES IgG
MARQUEES A LA PEROXYDASE
1 Selles testées Système témoin TS Système spécifique spsi
1 PIN PIN 1
1 1
1 Tl 1,80 1,02 1
1 T2 1,20 1,20 1
1 T3 l,50 0,53 )
1 T4 1,02 0,95 1
1 T5 1,60 1,30 1
1 Cyt 1 2,60 4,10* 1
1 Cyt 10 1,10 0,90 1
1 Cyt 14 1,60 1,45 1
1 Cyt 15 1,35 1,90 1
1 Cyt 18 2,32* 2,91* 1
1 Cyt 20 2,60* 3,10* 1
1 Cyt 24 2,08 5,2* 1
1 Cyt 25 2,19* 5,60* 1
1 Cyt 26 3,10* 4,9* 1
1 Cyt 28 1,60 1,20 1
1 Cyt 35 5,5* 11,9* 1
1 Cyt 36 2,97* 6,03* 1
1 Cyt 37 4,60* 10,70* 1
1 Cyt 38 3,60* 4,01* 1
Tableau n° 8
T selles témoins de sujets sains
Cyt = selles de malades de Constantine
54
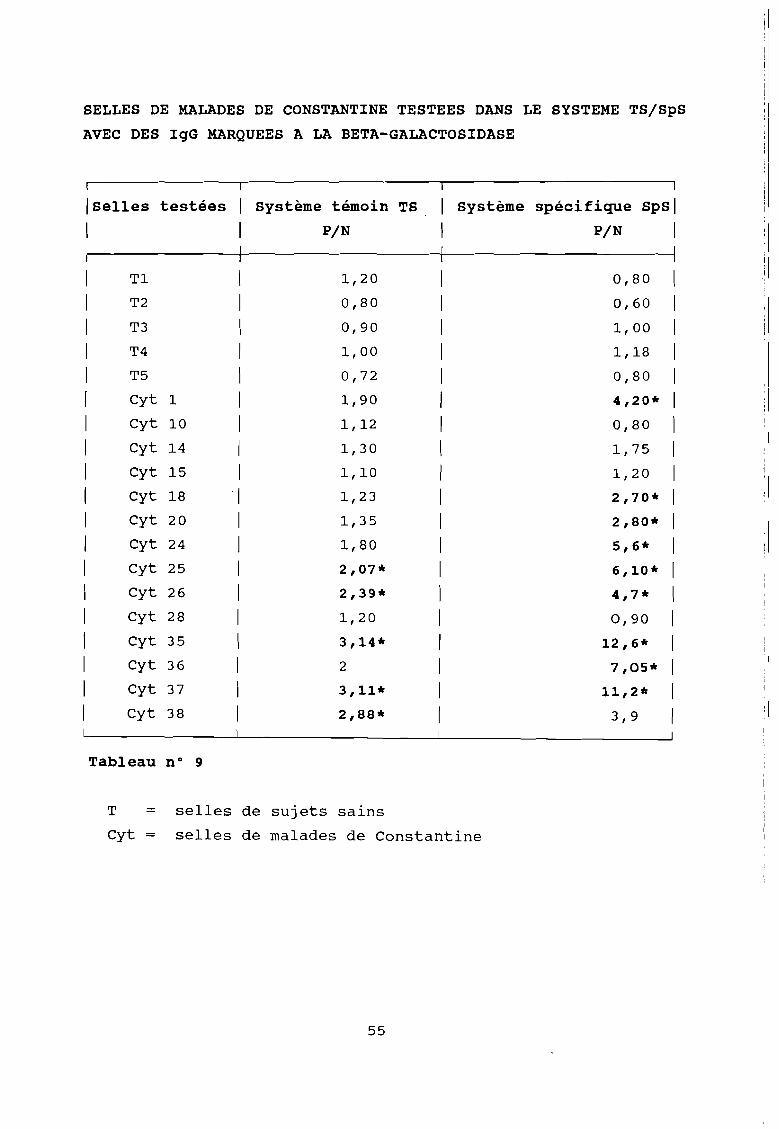
SELLES DE MALADES DE CONSTANTINE TESTEES DANS LE SYSTEME TSISpS
AVEC DES IgG MARQUEES A LA BETA-GALACTOSIDASE
Iselles testées 1 système témoin TS système spécifique SpS
1 1 PIN PIN
1 1
1 Tl 1 1,20 0,80
1 T2 1 0,80 0,60
1 T3 1 0,90 1,00
1 T4 1 1,00 1,18
1 T5 1 0,72 0,80
1 Cyt 1 1 1,90 4,20*
Cyt 10 1 1,12 0,80
Cyt 14 1 1,30 1,75
Cyt 15 1 1,10 1,20
Cyt 18 -1 1,23 2,70*
Cyt 20 1 1,35 2,80*
Cyt 24 1 1,80 5,6*
Cyt 25 1 2,07* 6,10*
Cyt 26 1 2,39* 4,7*
Cyt 281 1,20 0,90
Cyt 35 1 3,14* 12,6*
Cyt 36 1 2 7,05*
Cyt 37 1 3,11* 11,2*
Cyt 38 1 2,88* 3,9
Tableau n° 9
T = selles de sujets sains
Cyt = selles de malades de Constantine
55

Ces résultats montrent :
que le marquage à la péroxydase donne des réactions non
spécifiques dans 95 % des cas contre 64 % pour le marquage à la
bêta-galactosidase pour lequel nous avons opté.
- de même 60 % des réactions positives sont non spécifiques dans
le système humain contre 40 % dans le système singe. Nous avons
donc abandonné le système humain au profit du système singe que
nous avons cherché à améliorer.
Pour celà le lait écrémé à 4 % connu comme agent de blocage
non spécifique en Westernblot et qui en Elisa permet
l'utilisation de certains systèmes où les réactions non
spécifiques sont fréquentes (au risque de diminuer la
sensibilité) a été mis en jeu comme agent de saturation et de
dilution à la.place de la BSA.
Comme en atteste le tableau suivant aucune amélioration
sensible n'a été obtenue, seul le bruit de fond est diminué sans
que la positivité des réactions ne soit affectée.
56
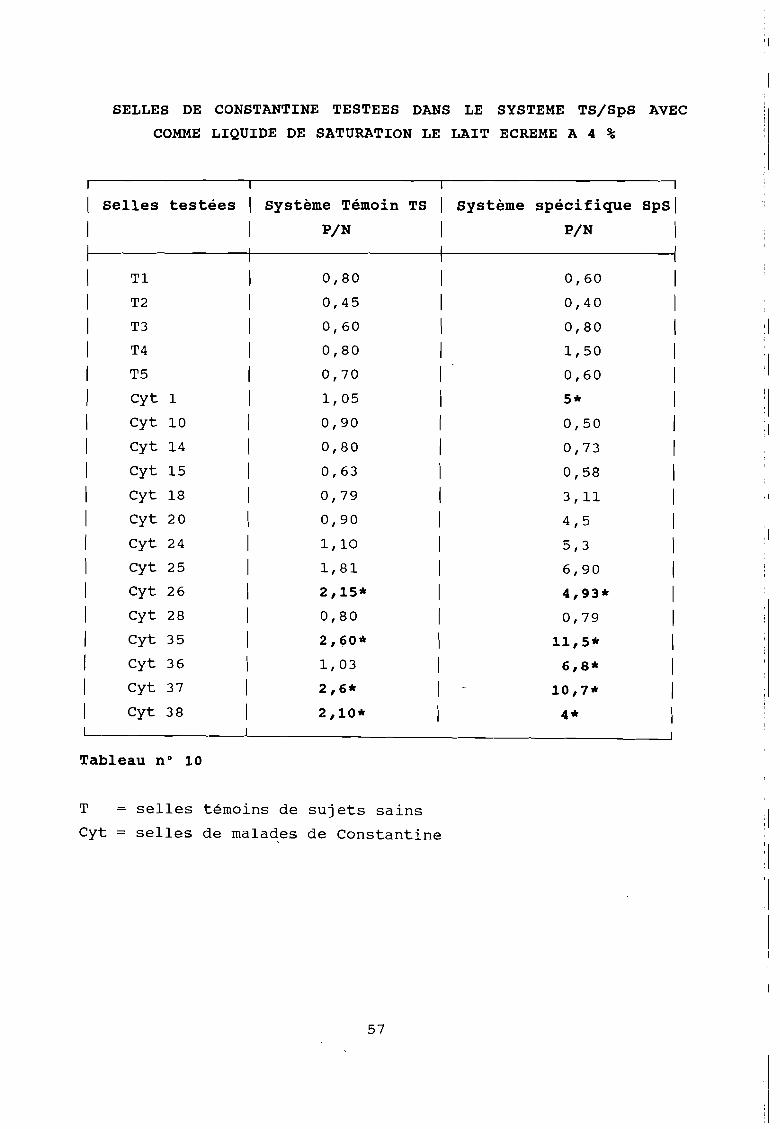
SELLES DE CONSTANTINE TESTEES DANS LE SYSTEME TSISpS AVEC
COMME LIQUIDE DE SATURATION LE LAIT ECREME A 4 %
Selles testées système Témoin TS système spécifique spsi
PIN PIN 1
1
Tl 0,80 0,60
T2 0,45 0,40
T3 0,60 0,80
T4 0,80 1,50
T5 0,70 0,60
Cyt 1 1,05 5*
Cyt 10 0,90 0,50
Cyt 14 0,80 0,73
Cyt 15 0,63 0,58
Cyt 18 0,79 3,11
Cyt 20 0,90 4,5
Cyt 24 1,10 5,3
Cyt 25 1,81 6,90
Cyt 26 2,15* 4,93*
Cyt 28 0,80 0,79
Cyt 35 2,60* 11,5*
Cyt 36 1,03 6,8*
Cyt 37 2,6* 10,7*
Cyt 38 2,10* 4*
Tableau n° 10
T selles témoins de sujets sains
Cyt = selles de malaqes de Constantine
57

c'est dans ce contexte qu'une étude menée par l'équipe du
Pr J. PILLOT (61) devait nous orienter vers la recherche d'une
protéine non spécifique appelée "Protéine Fv". En effet, cette
étude met en évidence l'existence de cette protéine dans les
selles d'hépatite non-A non-B. Cette protéine est capable de se
combiner de manière non immune avec la maj orité des
immunoglobulines humaines quelque soit leur isotype ou leur
allotype. Ce facteur est également présent chez 35 % des
hépatites B ainsi que chez 6,7 % des témoins (fréquence attendue
des porteurs sains).
La protéine est spécifique du fragment Fab et plus
précisément du fragment Fv et du domaine VH.
La molécule est une sialoglycoprotéine acide (pHi = 4)
hydrophobe, trypsine et chymotrypsine résistante. Al' état natif,
son poids moléculaire apparent est de 175 Kda en SDS-PAGE
préparative, il est de 85 Kda, ce qui suggère une structure
dimérique. L'origine hépatique a été démontrée par sa détection
dans la bile et le foie de patients ayant subi une
transplantation hépatique pour hépatite non-A non-B fulminante.
Les meilleures conditions permettant de caractériser cette
protéine Fv mettent en jeu une IgM monoclonale préparée à partir
du sérum d'un sujet atteint de la maladie de Waldenstrom et une
IgG d'enfant de 6 mois à 1 an marquée à la péroxydase.
Par cette technique, nous avons recherché la protéine Fv
dans les selles du singe infecté expérimentalement par le VHE;
et dans les différentes selles de Constantine positives.
C'est ainsi que nous avons pu détecter chez le singe ce
facteur qui apparaît avant la montée des transaminases comme le
montrent les f igures suivantes; et qui serait un marqueur précoce
des hépatites.
58
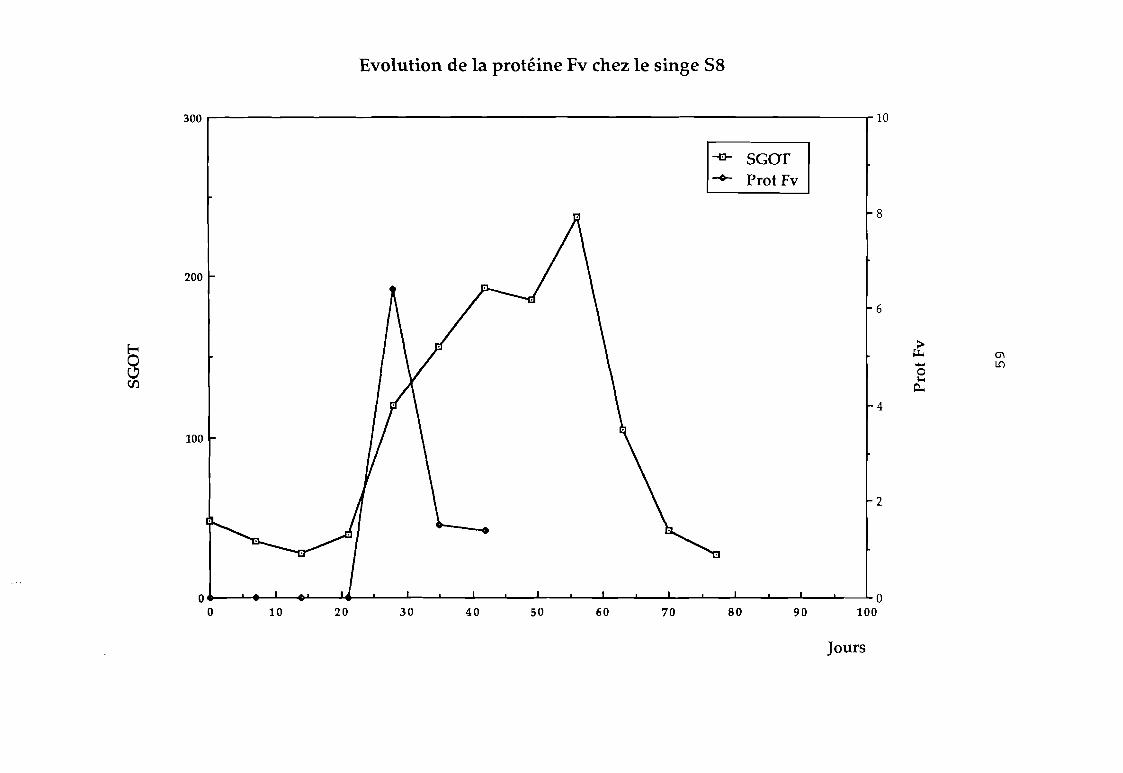
Evolution de la protéine Fv chez le singe 58
300 1 1 10
-e- SGar-+- Prot Fv
2
8
200
1 X ?---i \ 16
§ t IV \ ~>~ en~ l!)01-<Cf'.J ~
4
1 1/ \ ~ 1100
908070605040302010
• '. 1 .' ., ' 1 1 1 1 1 1 1 1 l , l , I,l 0100
Jours
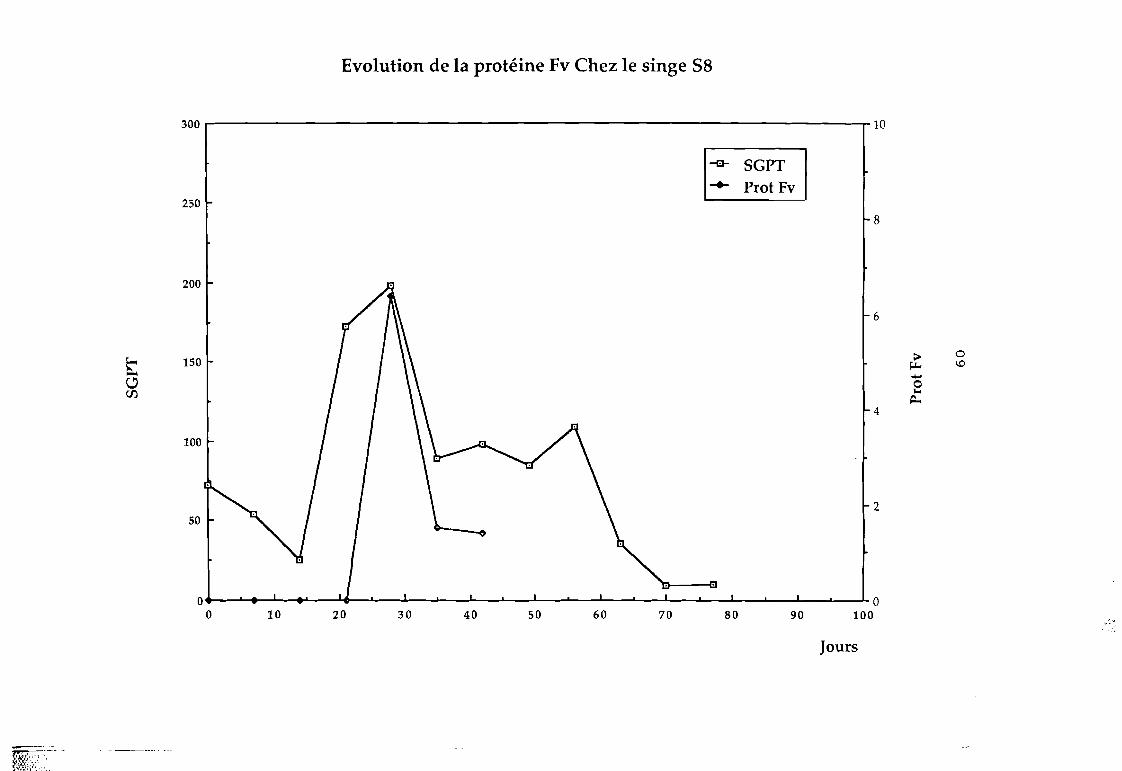
Evolution de la protéine Fv Chez le singe S8
300 1 1 la
-e- SGPT~ Prot Fv
2508
250
200
1 /~ 16
t 1S0r 1 1 \\ t;;. 0~ \0....
0 0Cf) '"'~
4
1 1 1 \\ A 1100
908070605040302010
• ',' l' " ' l , l , , , l , , , l , , 1 1 a100
Jours
f~t7-,--'- -
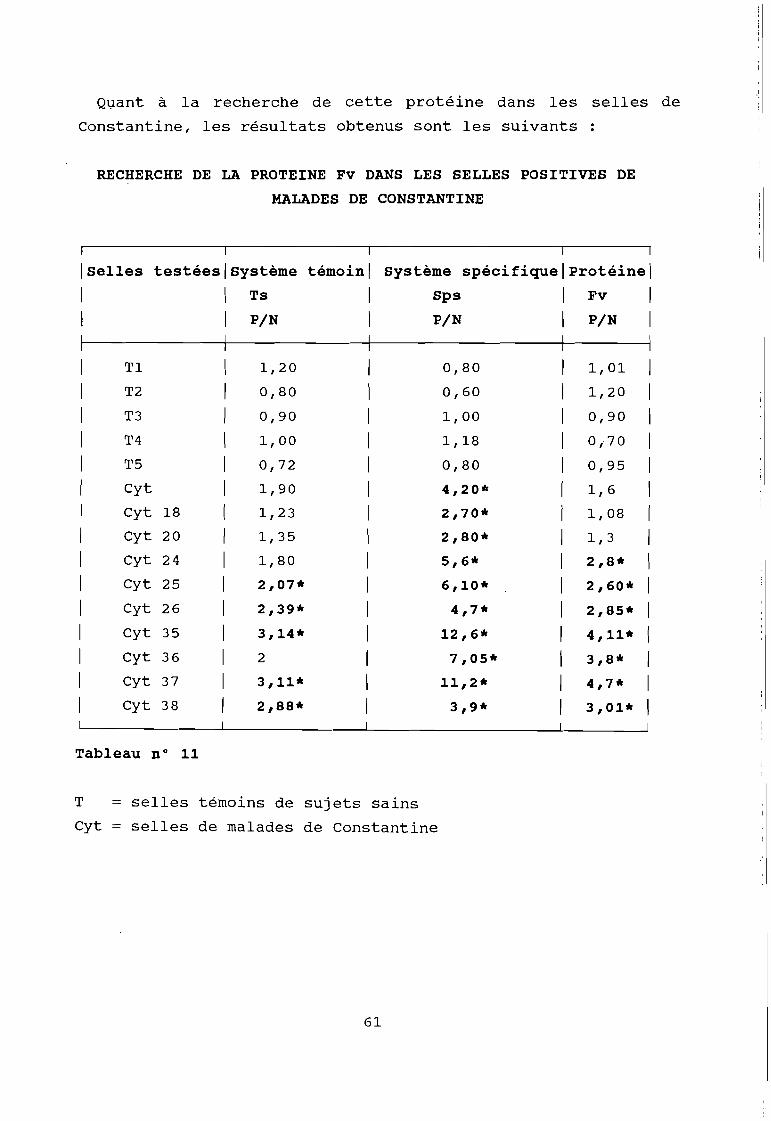
Ql,lant à la recherche de cette protéine dans les selles de
Constantine, les résultats obtenus sont les suivants
RECHERCHE DE LA PROTEINE Fv DANS LES SELLES POSITIVES DE
MALADES DE CONSTANTINE
1
Selles testées 1 système témoin système spécifique Protéine
Ts sps Fv
PIN PIN PIN
Tl 1,20 0,80 1,01
T2 0,80 0,60 1,20
T3 0,90 1,00 0,90
T4 1,00 1,18 0,70
T5 0,72 0,80 0,95
Cyt 1,90 4,20* 1,6
Cyt 18 1,23 2,70* 1,08
Cyt 20 1,35 2,80* 1,3
Cyt 24 1,80 5,6* 2,8*
Cyt 25 2,07* 6,10* 2,60*
Cyt 26 2,39* 4,7* 2,85*
Cyt 35 3,14* 12,6* 4,11*
Cyt 36 2 7,05* 3,8*
Cyt 37 3,11* 11,2* 4,7*
Cyt 38 2,88* 3,9* 3,01*
Tableau n° 11
T = selles témoins de sujets sains
Cyt = selles de malades de Constantine
61

D'après ces résultats, ces réactions non spécifiques
semblent liées à l'existence de cette protéine Fv qui est
présente dans 70 % des selles positives. Nous avons alors émis
l'hypothèse que le passage des selles sur immunoadsorbant d'IgM
monoclonale devrait donc éliminer ces réactions non spécifiques.
Préparation de L'immunoadsorbant
L'immunoadsorption sur Sépharose activé a été effectuée selon
la méthode de March et Cuatrecasas modifiée par l'introduction
de Bromure de cyanogène (BrCn) en solution dans l'acétronitrile.
Pour 50 mg de protéines à fixer, on utilise 10 ml de Sépharose
4B (Pharmacia) .A un volume de Sépharose 4B tassé par décantation,
on ajoute un volume d'eau distillée et 2 volumes de carbonate de
sodium 2M. Dans cette suspension, on introduit 1/20e de volume
de BrCn dissout dans l'acétonitrile, à raison de 2 g de BrCn pour
1 ml d'acétonitrile. Cette opération est effectuée rapidement,
sous hotte aspirante et avec agitation pendant 2 minutes.
Ensuite, le Sépharose est rapidement lavé sur un verre fritté n°
3 raccordé à une fiole à vide, avec un grand excès d'eau
distillée froide (4°C). On lave ensuite en tampon carbonate 0,1
M pH9 à +4°C. Après ce dernier lavage, le Sépharose est dispersé
toujours à +4°C, dans le tampon carbonate (10 ml pour 10 ml de
Sépharose). Les protéines dialysées en tampon carbonate 0,1 M pH
9 sont ajoutées, goutte à goutte en agitant, puis laissées sous
agitation douce à +4°C pendant 18 heures. Le Sépharose est
ensuite refiltré sur verre fritté, le filtrat est recueilli en
vue de calculer le rendement de fixation de la protéine au
Sépharose activé, par absorption à 280 nm. On traite ensuite
l'immunoadsorbant par l'éthanolaminé 1 M pH 9 pendant 2 h à la
température du laboratoire, en agitation douce afin de saturer
les sites actifs lib~es. Puis, l'immunoadsorbant est traité par
l'agent d'élution qui sera utilisé ultérieurement pour extraire
la substance fixée chlorure de magnésium 5 M ou un tampon
glycine pH 2,8 ou encore un mélange d'acide acétique dioxane pH
2,9 (acide acétique 0,2 M + 10 % de dioxane). Après lavage en eau
physiologique, l'immunoadsorbant est prêt à l'emploi.
62
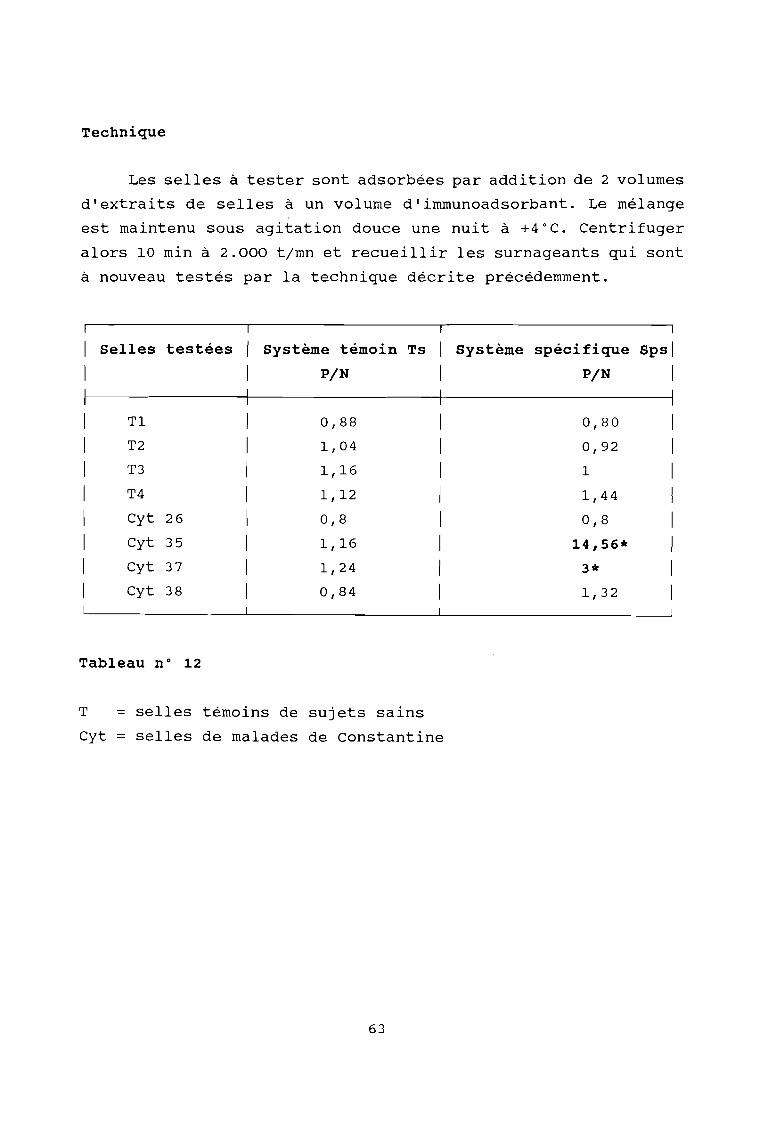
Technique
Les selles à tester sont adsorbées par addition de 2 volumes
d'extraits de selles à un volume d'immunoadsorbant. Le mélange
est maintenu sous agitation douce une nuit à +4°C. Centrifuger
alors 10 min à 2.000 tlmn et recueillir les surnageants qui sont
à nouveau testés par la technique décrite précédemment.
selles testées système témoin Ts système spécifique spsl
PIN PIN 1
1
Tl 0,88 0,80 1
T2 1,04 0,92 1
T3 1,16 1 1
T4 1,12 1,44 1
Cyt 26 0,8 0,8 1
Cyt 35 1,16 14,56* 1
Cyt 37 1,24 3* 1
Cyt 38 0,84 1,32 1
Tableau n° 12
T selles témoins de sujets sains
Cyt selles de malades de Constantine
63

Comme le montre le tableau précédent, pour les quatre selles
étudiées, deux conservent leur activité en système spécifique
tandis que les deux autres la perdent. Ce qui est en faveur pour
ces dernières d'une interférence de la protéine Fv.
Ces résultats semblent donc confirmer notre hypothèse et
nous ont conduit à la recherche de l'antigène VHE et de la
protéine Fv dans les selles du singe infecté, dans différentes
selles de sujets atteints d'hépatite épidémique à Constantine
et à Bangkok après passage systématique sur immunoadsorbant,
avant d'entreprendre cette même étude sur les selles de sujets
atteints d'hépatite aiguë à Dakar et dans sa région.
Résultats :
1) chez le singe infecté l'antigène VHE est détectable dès
la troisième semaine après l'inoculation c'est-à-dire pendant la
période d' incubation, avant la montée des transaminases (cf
courbes suivantes).
64
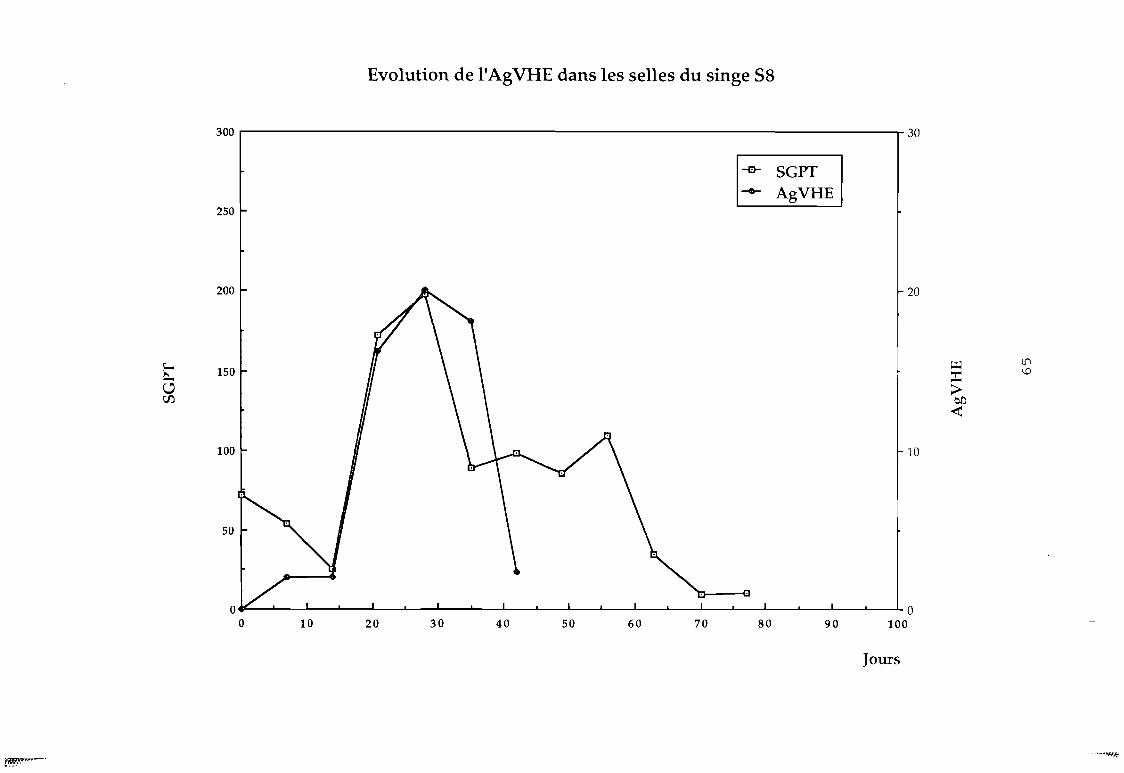
Evolution de l'AgVHE dans les selles du singe S8
300 i 1 30
250
-0-
-+-SGPTAgVHE
toU')
200
150
100
50
20
10
~:r:;>00~
lîI\.0
908070605040302010......' l , l , Iii , 1 1 l , l , 1 1 1 1 1 0
100
Jours
t~_·
....•.•••""'y
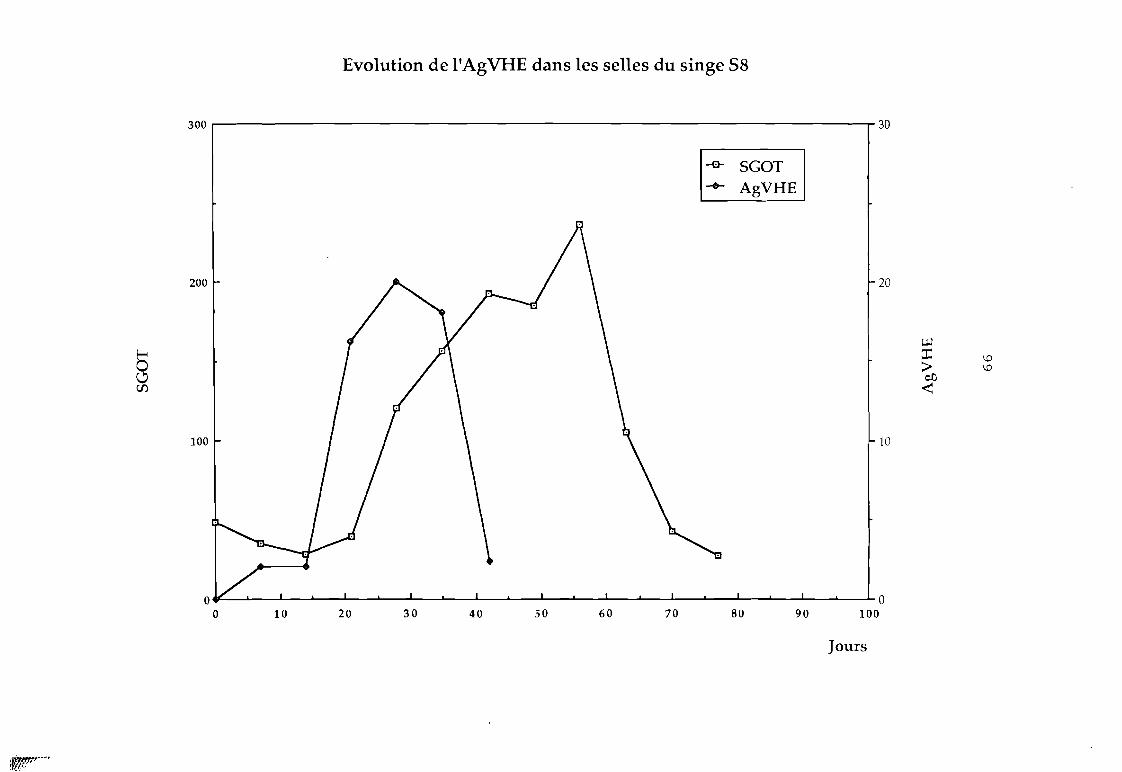
Evolution de l'AgVHE dans les selles du singe 58
300 1 1 30
-0-
......SGOTAgVHE
boCf)
200
100
20
10
~
::c:>0.0~
\0\0
908070605040302010
.".. , l , , l , 11ft , 1 ) 1 l , l , , 1 1 a100
Jours
<rWmr'-"~fl(.('
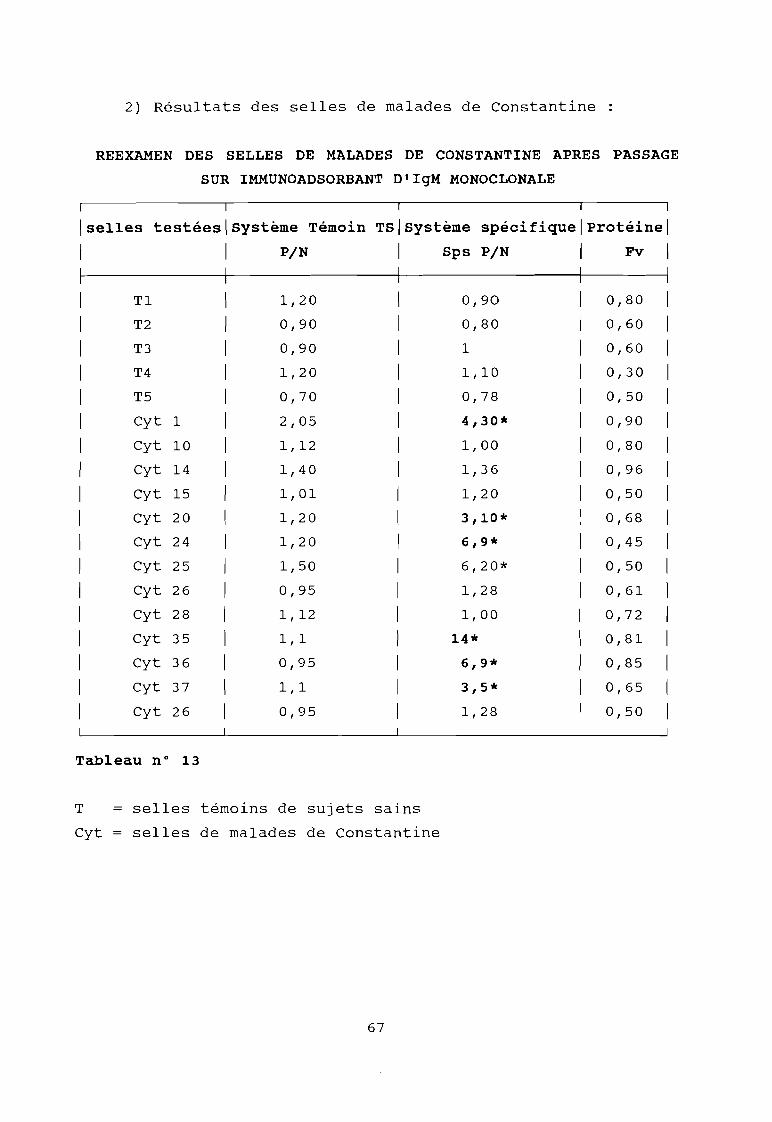
2) Résultats des selles de malades de Constantine
REEXAMEN DES SELLES DE MALADES DE CONSTANTINE APRES PASSAGE
SUR IMMUNOADSORBANT D'IgM MONOCLONALE
1 1
selles testéeslsystème Témoin Tslsystème spécifique Protéine 1
PIN Sps PIN Fv
Tl 1,20 0,90 0,80
T2 0,90 0,80 0,60
T3 0,90 1 0,60
T4 1,20 1,10 0,30
T5 0,70 0,78 0,50
Cyt 1 2,05 4,30* 0,90
Cyt 10 1,12 1,00 0,80
Cyt 14 1,40 1,36 0,96
Cyt 15 1,01 1,20 0,50
Cyt 20 1,20 3,10* 0,68
Cyt 24 1,20 6,9* 0,45
Cyt 25 l,50 6,20* 0,50
Cyt 26 0,95 1,28 0,61
Cyt 28 1,12 1,00 0,72
Cyt 35 1,1 14* 0,81
Cyt 36 0,95 6,9* 0,85
Cyt 37 1,1 3,5* 0,65
Cyt 26 0,95 1,28 0,50
Tableau n° 13
T = selles témoins de sujets sains
Cyt selles de malades de Constantine
67
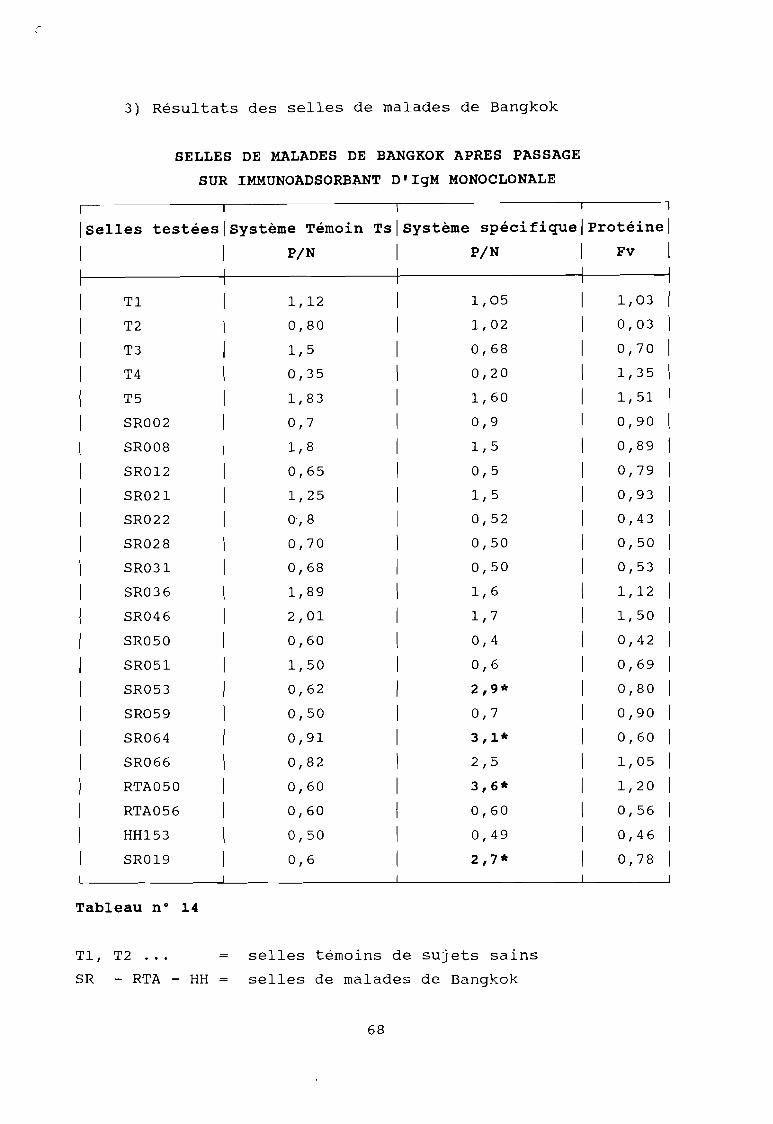
3) Résultats des selles de malades de Bangkok
SELLES DE MALADES DE BANGKOK APRES PASSAGE
SUR IMMUNOADSORBANT D'IgM MONOCLONALE
1 1 1
Selles testéeslsystème Témoin Tslsystème spécifique Protéine 1
Tl
T2
T3
T4
T5
SR002
SR008
SR012
SR021
SR022
SR028
SR031
SR036
SR046
SR050
SR051
SR053
SR059
SR064
SR066
RTA050
RTA056
HH153
SR019
Tableau n° 14
Tl, T2 . . .SR - RTA - HH
PIN PIN Fv
1,12 l,OS 1,03
0,80 1,02 0,03
1,5 0,68 0,70
0,35 0,20 1,35
1,83 l,6O l,51
0,7 0,9 0,90
1,8 1,5 0,89
0,65 0,5 0,79
1,25 1,5 0,93
0,8 0,52 0,43
0,70 0,50 0,50
0,68 0,50 0,53
1,89 1,6 1,12
2,01 1,7 l,50
0,60 0,4 0,42
l,50 0,6 0,69
0,62 2,9* 0,80
0,50 0,7 0,90
0,91 3,1* 0,60
0,82 2,5 l,OS
0,60 3,6* l,2O
0,60 0,60 0,56
0,50 0,49 0,46
0,6 2,7* 0,78
selles témoins de sujets sains
selles de malades de Bangkok
68

Dès lors, nous avons alors entrepris un contrôle de la
spécificité du système de détection que nous avons mis au point.
Pour ce faire :
- 18 selles de malades atteints d'hépatite aiguë à Dakar ont été
testées dans notre système de même que 18 selles de sujets
témoins ne souffrant d'aucune atteinte hépatique évidente.
- des réactions d'inhibition et des réactions témoins utilisant
différents anticorps d'une part, et les selles positives d'autre
part, ont été réalisées.
69
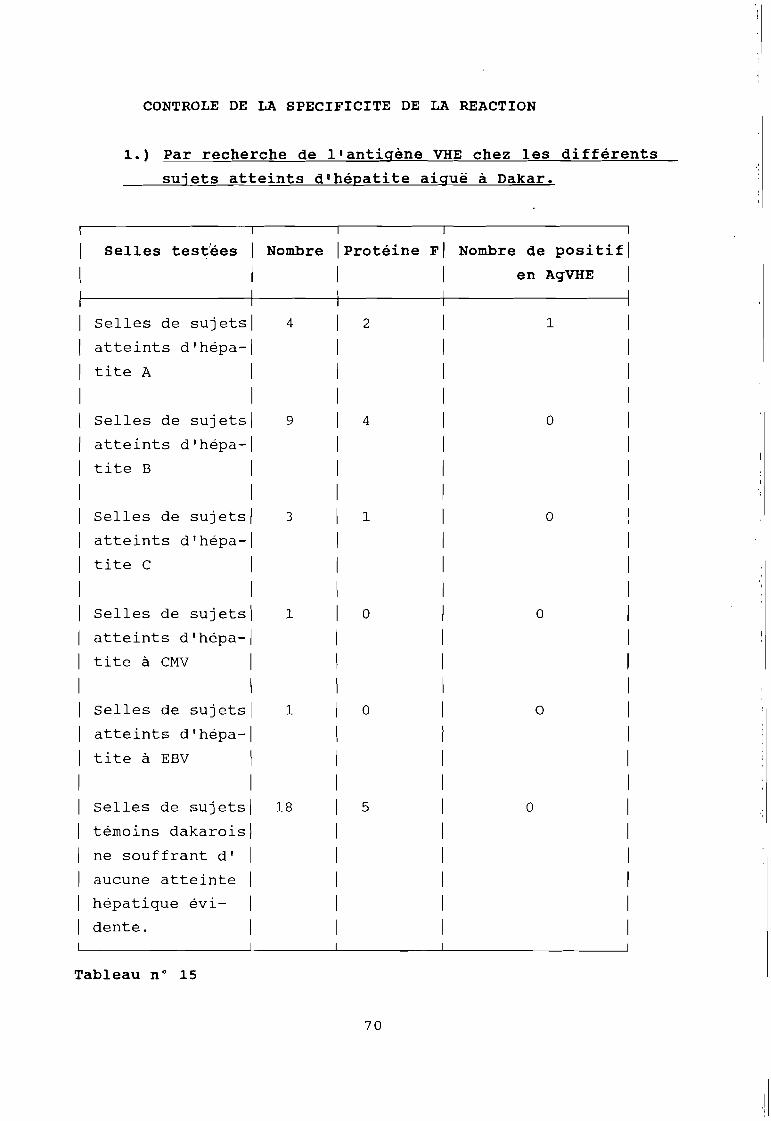
CONTROLE DE LA SPECIFICITE DE LA REACTION
1.) Par recherche de l'antigène VHE chez les différents
sujets atteints d'hépatite aiguë à Dakar.
Selles test'ées Nombre 1 Protéine F Nombre de positifl
en AgVHE 1
1
Selles de sujets 4 2 1 1
atteints d'hépa-
tite A
Selles de sujets 9 4 0
atteints d'hépa-
tite B
Selles de sujets 3 1 0
atteints d'hépa-
tite C
Selles de sujets 1 0 0
atteints d'hépa-
tite à CMV
selles de sujetsl 1 0 0
atteints d'hépa-I
tite à EBV
Selles de sujetsl 18 5 0
témoins dakaroisl
ne souffrant d'
aucune atteinte
hépatique évi-
dente.
Tableau n° 15
70

2.) Réactions d'inhibition
a) principe
Les selles adsorbées après passage sur immunoadsorbant d'une
IgM monoclonale sont incubées avec des sérums contenant des
anticorps antiVHE.
Les antigènes VHE sont alors supposés se complexer et ne
devraient plus réagir ultérieurement, lorsqu'ils sont mis en
réaction dans le système de détection précédemment défini.
b) Technique
On réalise différentes dilutions (1/10, 1/20, 1/40, 1/80,
1/100) à partir d'un mélange d'IgM et d'IgG provenant du singe
S8 infecté par le VHE, à 2,5 mg/ml pour chaque immunoglobuline.
A 100 pl de cette solution d'anticorps spécifiques, on
ajoute 100 fI de selles antérieurement reconnues comme contenant
l'antigène associé à l'infection par le VHE.
Laisser en contact 2 heures à 37°C puis une nuit à +4°C.
Passer à nouveau les selles dans le système spécifique.
c) résultats
71
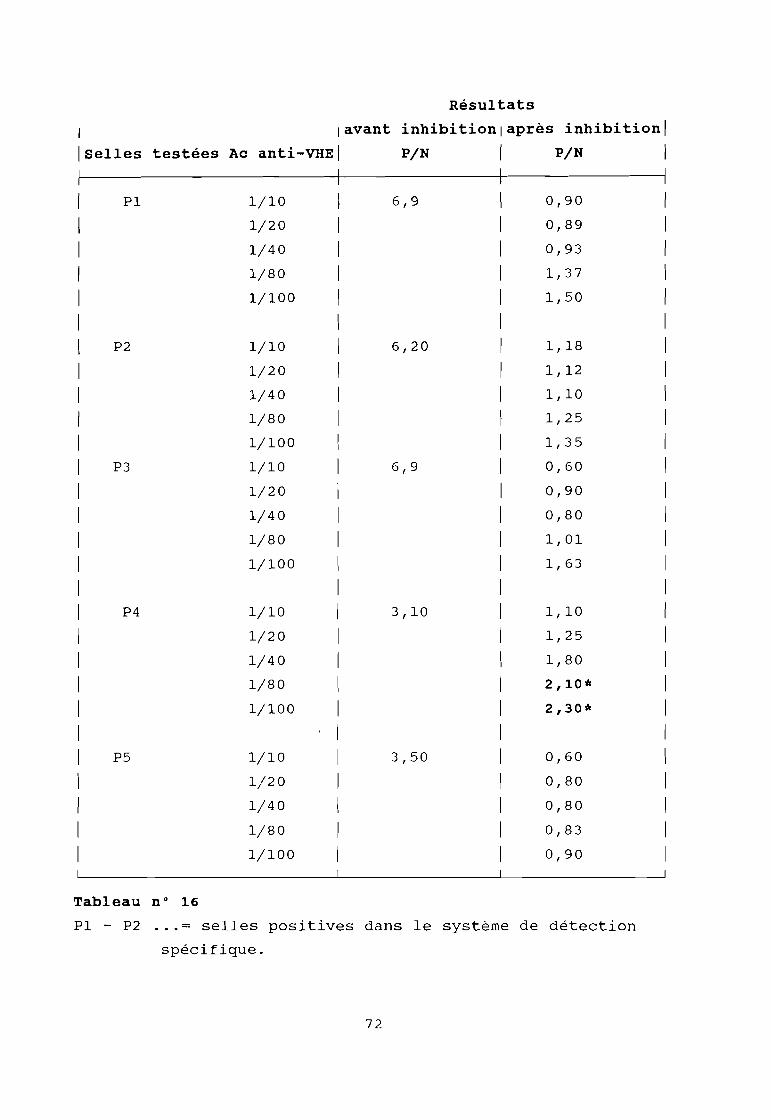
Résultats
1 lavant inhibition 1après inhibition
Iselles testées Ac anti-VHEI PIN PIN
1
1 Pl 1/10 6,9 0,90
1 1/20 0,89
1 1/40 0,93
1 1/80 1,37
1 1/100 l,50
1
1 P2 1/10 6,20 1,18
1 1/20 1,12
1 1/40 1,10
1 1/80 1,25
1 1/100 1,35
1 P3 1/10 6,9 0,60
1 1/20 0,90
1 1/40 0,80
1 1/80 1,01
1/100 1,63
P4 1/10 3,10 1,10
1/20 1,25
1/40 1,80
1/80 2,10*
1/100 2,30*
P5 1/10 3,50 0,60
1/20 0,80
1/40 0,80
1/80 0,83
1/100 0,90
Tableau n° 16
Pl - P2 .•• = selles positives dans le système de détection
spécifique.
72
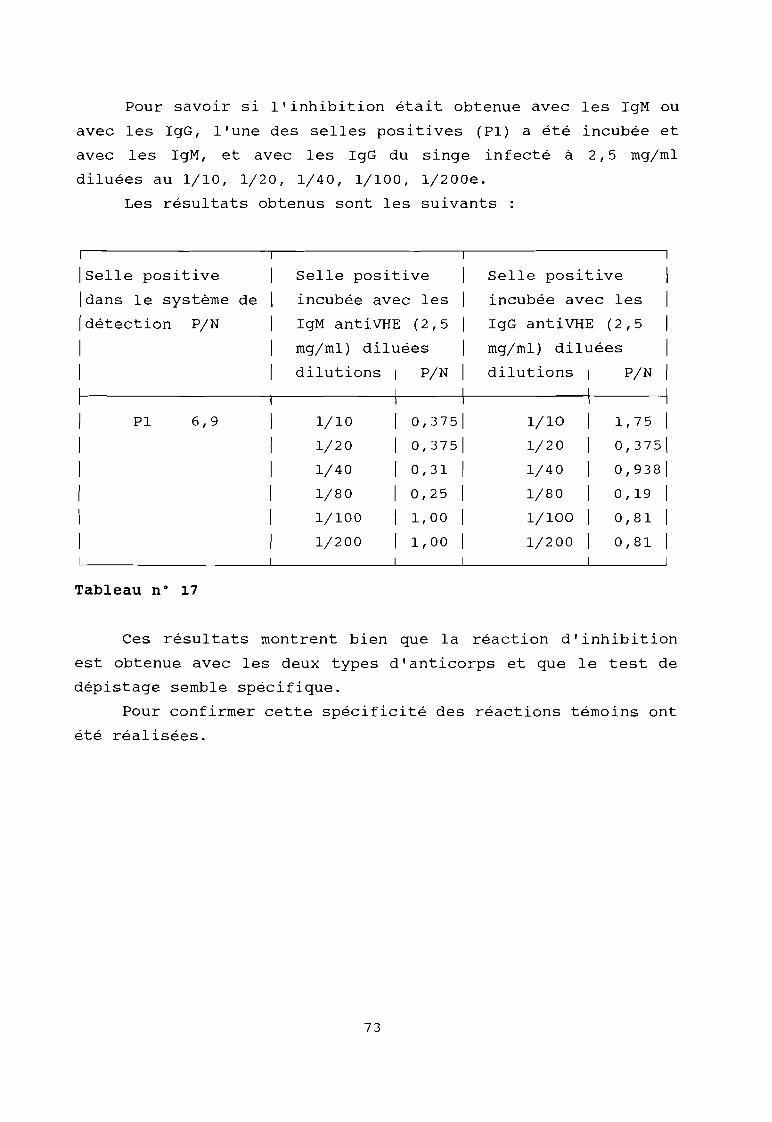
Pour savoir si l'inhibition était obtenue avec les IgM ou
avec les IgG, l'une des selles positives (Pl) a été incubée et
avec les IgM, et avec les IgG du singe infecté à 2,5 mg/ml
diluées au 1/10, 1/20, 1/40, 1/100, 1/200e.
Les résultats obtenus sont les suivants
Iselle positive Selle positive Selle positive 1
1dans le système de incubée avec les incubée avec les 11détection PIN IgM antiVHE (2,5 IgG antiVHE (2,5 1
mg/ml) diluées mg/ml) diluées 1dilutions PIN dilutions PIN 1
1
Pl 6,9 1/10 0,375 1/10 1,75 11/20 0,375 1/20 0,3751
1/40 0,31 1/40 0,9381
1/80 0,25 1/80 0,19 11/100 1,00 1/100 0,81 1
1/200 1,00 1/200 0,81 1
Tableau n° 17
Ces résultats montrent bien que la réaction d'inhibition
est obtenue avec les deux types d'anticorps et que le test de
dépistage semble spécifique.
Pour confirmer cette spécificité des réactions témoins ont
été réalisées.
73
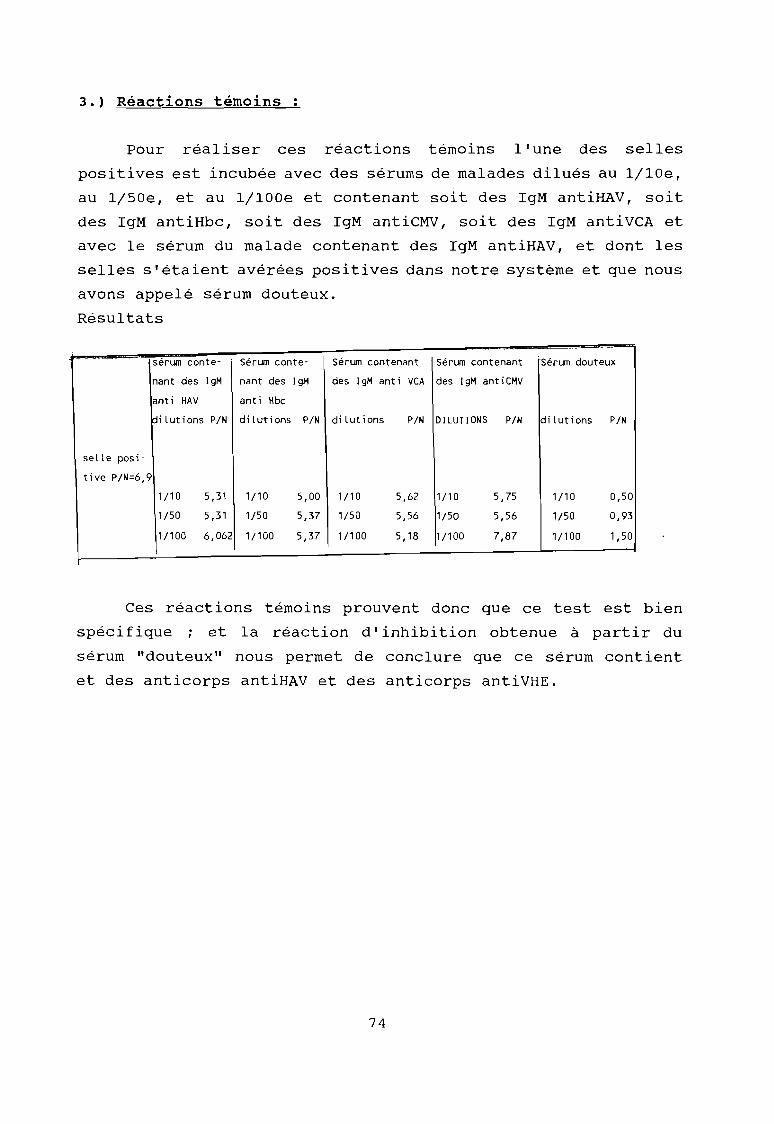
3.) Réactions témoins
Pour réaliser ces réactions témoins l'une des selles
positives est incubée avec des sérums de malades dilués au 1110e,
au 1/50e, et au 11100e et contenant soit des IgM antiHAV, soit
des IgM antiHbc, soit des IgM antiCMV, soit des IgM antiVCA et
avec le sérum du malade contenant des IgM antiHAV, et dont les
selles s'étaient avérées positives dans notre système et que nous
avons appelé sérum douteux.
Résultats
serum conte- Sérum conte- Sérum contenant Sérum contenant Sérum douteux
nant des IgM nant des IgM des IgM anti VCA des 1gM ant i CMV
anti HAV ant i Hbc
dilutions PIN di luti ons PIN di lutions PIN DILUTIONS PIN di lutions PIN
selle posi-
tive P/N=6,9
1/10 5,31 1/10 5,00 1/10 5,62 1/10 5,75 1/10 D,50
1/50 5,31 1/50 5,37 1/50 5,56 1/50 5,56 1/50 0,93
1/100 6,062 1/100 5,37 1/100 5,18 1/100 7,87 1/100 1,50
Ces réactions témoins prouvent donc que ce test est bien
spécifique et la réaction d'inhibition obtenue à partir du
sérum "douteux" nous permet de conclure que ce sérum contient
et des anticorps antiHAV et des anticorps antiVHE.
74

DEUXIEME PARTIE:
DIAGNOSTIC BIOLOGIQUE
DE l'HEPATITE C :
ETUDE CRITIQUE DES TESTS
DE DETECTION
DES ANTICORPS ANTIHCVg

Depuis que la recherche systématique de l'antigène HBs chez
les donneurs de sang a été rendue obligatoire dans de nombreux
pays, l'incidence des hépatites post-transfusionnelles a diminué
de manière très sensible mais beaucoup moins qu'on ne pouvait
l'espérer.
En effet, il a été montré que l'hépatite C représente à
l'heure actuelle 50 à 90 % des hépatites post-transfusionnelles
(81) et constitue pour certains pays un véritable fléau en santé
publique.
Des tests de détection des anticorps anti VHC dans le sérum
ou dans le plasma ont été mis au point permettant certes de
réduire cette redoutable complication des transfusions sanguines.
Ces anticorps reconnaissent une protéine de la région non
structurale du VHC probablement impliquée dans la réplication
virale. L'avantage de ce point est qu'il existerait une relation
entre le taux de l'anticorps anti-HCV et la réplication virale
et donc l'infectiosité du sérum.
Des études expérimentales sur le chimpanzé menées par Weiner
et al. (85) ont permis de le démontrer.
Mais ces tests posent des problèmes de spécificité comme
le prouvent les études suivantes.
75

AI METHODES DE DETECTION DES ANTICORPS ANTIHCV
A l'heure actuelle, pour la recherche des anticorps antiHCV
nous disposons de techniques immunoenzymatiques :
- tests de screening en Elisa (Ortho Elisa et Abbott ElA).
- tests de confirmation par immunoblot (Chiron Riba, Abbott
Inhibition) (53).
Tous ces tests présentent des points communs et quelques
points différentiels.
1) - POINTS COMMUNS
a) - principe
Tous ces tests ont le même principe à savoir
un support réactionnel revêtu d'un antigène recombinant
spécifique de HCV est mis en contact avec le sérum ou le plasma
à tester.
si l'anticorps antiHCV est présent dans le prélèvement, des
complexes antigène-anticorps vont se former à la surface du
support réactionnel après incubation.
Dans le cas contraire, aucun complexe ne se fixera, et les
protéines plasmatiques ou sériques libres seront évacuées lors
du lavage.
Une immunoglobuline anti humaine marquée à la peroxydase
est alors ajoutée et peut se fixer aux anticorps spécifiques
déjà couplés aux antigènes ,viraux.
si ces complexes immuns sont absents, le conjugué libre est
évacué au cours du lavage.
Enfin, un système de détection enzymatique spécifique est
ajouté entraînant l'apparition d'une coloration révélatrice de
la présence d'anticorps antiHCV.
La réaction est stoppée soit grâce à un réactif bloquant
(H2S04) , soit par lavage et l' intensité de la coloration est
proportionnelle à la quantité de conjugué lié et par delà à la
concentration d'anticorps anti-HCV présents dans le prélèvement.
b) - l'antigène
76
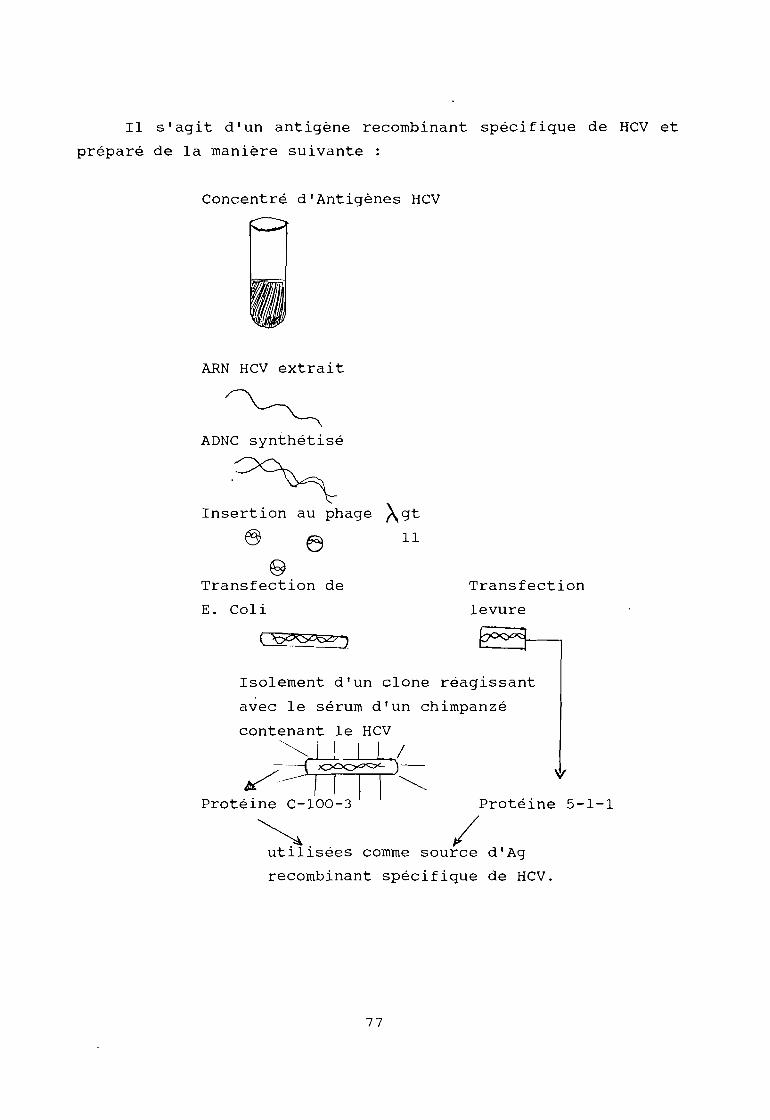
Il s'agit d'un antigène recombinant spécifique de HCV et
préparé de la manière suivante :
Concentré d'Antigènes HCV
ARN HCV extrait
ADNC synthétisé
Insertion au phage
9Transfection de
E. Coli
( 5QZ><SZ>e;z;>J
~gt
11
Transfection
levure
Protéine 5-1-1
Isolement d'un clone réagissant
avec le sérum d'un chimpanzé
contenant le HCV...~ 1 1 bJ-{
0===lTFr~Protéine C-100-3
~ /utilisées comme source d'Ag
recombinant spécifique de HCV.
77

2) - POINTS DIFFERENTIELS
a) - le support
Il est constitué
- soit de micropuits revêtus de l'antigène pour le kit Ortho,
- soit de billes revêtues du même antigène pour le kit Abbott
ou encore
- de membrane de nitrocellulose pour les tests de confirmation.
b) - les immunoglobulines marquées
Pour les tests screening, l'immunoglobuline marquée est une
immunoglobuline monoclonale de souris antihumaine dans le système
ortho, alors qu'il s'agit d'une immunoglobuline polyclonale de
lapin antihumaine dans le test Abbott.
Quant aux tests de confirmation, les immunoglobulines
marquées utilisées sont des immunoglobulines de chèvre anti IgG
humaine.
78

BI - ETUDE DE LA SPECIFICITE DE CES TESTS
1) Méthode
Pour étudier la spécificité de ces tests une étude
rétrospective a été réalisée sur différents sérums présentant
une réaction positive :
en IgM anti HAV = 10
en latex waaler rose = 10
en facteur antinucléaire, antiDNA = 20
et sur des sérums de sujets atteints d'hépatites auto
immunes = 13
- ou de cirrhoses alcooliques = 142
Chacun de ces sérums a été étudié dans les 4 tests
précédemment cités.
2) Résultats
Les résultats obtenus ont été consignés dans le tableau
suivant :
79
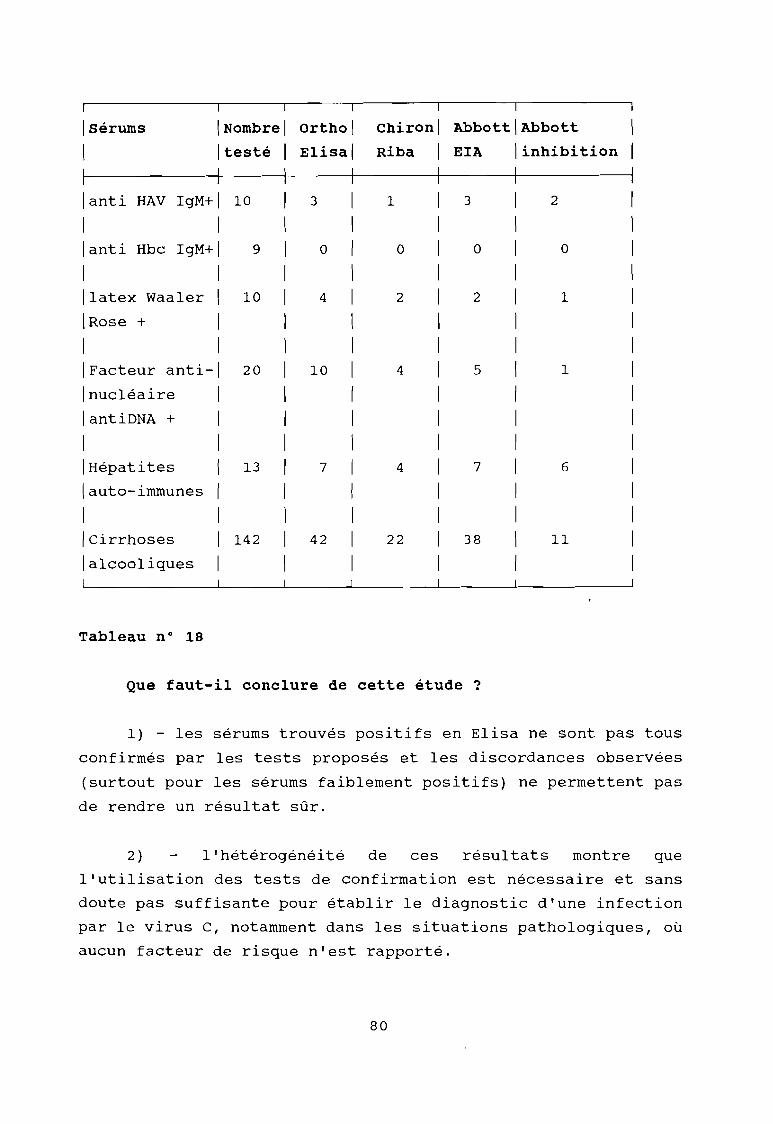
1Sérums Nombre Ortho 1 Chiron Abbott Abbott
1 testé Elisal Riba ElA inhibition
1 1
lanti HAV IgM+ la 3 1 1 3 2
1 1
lanti Hbc IgM+ 9 a / a a a
1 1
1 latex Waaler la 4 1 2 2 1
IRose + 1
1
1 Facteur anti- 20 la 1 4 5 1
1 nucléaire 1
lantiDNA + 1
1
1 Hépatites 13 7 1 4 7 6
1auto-immunes 1
1
/cirrhoses 142 42 1 22 38 11
1 alcooliques 1
1
Tableau n° 18
Que faut-il conclure de cette étude ?
1) - les sérums trouvés positifs en Elisa ne sont pas tous
confirmés par les tests proposés et les discordances observées
(surtout pour les sérums faiblement positifs) ne permettent pas
de rendre un résultat sûr.
2) l' hétérogénéité de ces résultats montre que
llutilisation des tests de confirmation est nécessaire et sans
doute pas suffisante pour établir le diagnostic d'une infection
par le virus c, notamment dans les situations pathologiques, où
aucun facteur de risque n1est rapporté.
80

TROISIEME PARTIE:
ETUDE EPIDEMIOLOGIQUE
DES HEPATITES
NON-A NON-B A DAKAR ET
DANS SA- REGION

~S~..1l
A - RECHERCHE DE L'HEPATITE E DANS UNE POPULATION
DE SUJETS ATTEINTS D'HEPATITE AlGUE A DAKAR
1) Méthode
Cette étude porte sur 30 cas de sujets adultes atteints
d'hépatite aiguë parfaitement définie sur le plan clinique. Sur
le plan biologique, le dosage des transaminases sériques (SGOT
SGPT) , de la bilirubine (directe, indirecte et totale), des
phosphatases alcalines et des gammaGT ont permis de confirmer le
diagnostic d'hépatite.
Le retour à la normale des transaminases 3 mois après, et
un contrôle clinique, ont permis de confirmer le diagnostic
d'hépatite aiguë.
A partir des sérums de ces malades, différents marqueurs
viraux ont été recherchés, à savoir :
- les IgM anti-HAV,
- l'antigène HBs et les IgM anti-HBc,
- les IgM anti-CMV,
- les IgM anti-vCA,
- les marqueurs de la fièvre jaune.
2) Résultats
Les résultats obtenus sont consignés dans le tableau suivant:
82

RECHERCHE DES MARQUEURS VIRAUX DANS LES SERUMS DE SUJETS
ATTEINTS D'HEPATITE AlGUE
Marqueurs d'hépatite
IgM anti-HAV
Antigène HBs
dont IgMHbc+
dont IgMHbc
IgM anti-VCA
IgM anti-CMV
IgM anti-virus amaril 1
Tableau n° 19
Nombre de cas positifs
4
9
5
4
1
1
o
D'après ces résultats, nous pouvons déduire que 50 % de ces
hépatites aiguës de l'adulte sont NANB.
Nous avons alors entrepris la recherche des anticorps anti
HCV dans les sérums correspondants. Pour ce faire, les sérums
d'hépatite NANB ont été testés dans les kits Abbott et dans les
kits Ortho.
Les résultats suivants ont été obtenus
Noobre Noobre X Noobre X
SérUllS NANB Positifs Ortho Positivité Ortho Positifs Abbott Positivité Abbott
15 3 20 3 20
Tableau n° 20
83

Ces résultats ont été confirmés par des tests de
confirmations disponibles également, et commercialisés par les
mêmes laboratoires que précédemment cités. Ces tests montrent
que les anticorps anti-HCV sont effectivement présents dans 20%
des cas.
A partir des selles de ces sujets et des selles témoins,
nous avons recherché l'antigène VHE, et les résultats obtenus
sont consignés dans le tableau suivant :
Nombre de positifsen AgVHE
Nombre1
\selles testées
1
1,--------------+----------------
Selles de sujetsatteints d'hépatite A
4 1
Selles de sujetsatteints d'hépatite B
9 o
Selles de sujetsatteints d'hépatite C
3 o
Selles de sujetsatteints d'hépatiteà CMV
1 o
Selles de sujetsatteints d'hépatiteà EBV
1 o
Selles de sujetsatteints d'hépatiteà sérologie négative
12 5
Selles de sujets témoinsDakarois ne souffrantd'aucune atteinte héptique évidente
30 1
Tableau n° 21
84
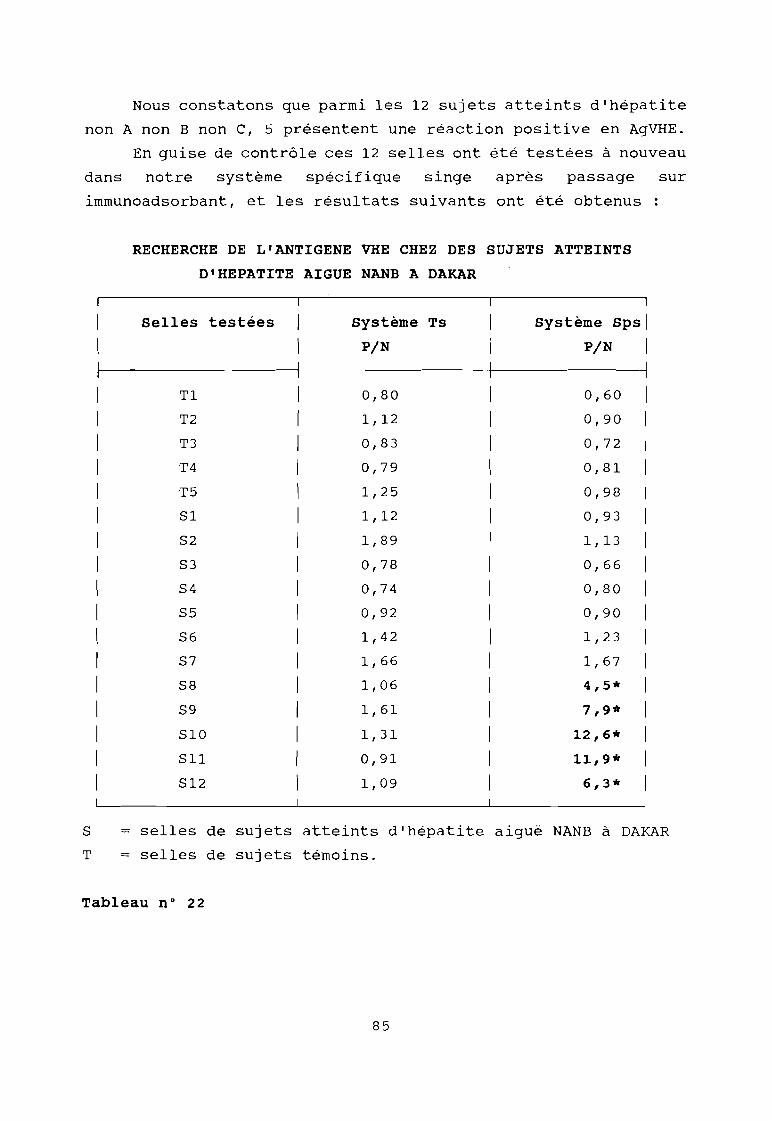
Nous constatons que parmi les 12 sujets atteints d'hépatite
non A non B non C, 5 présentent une réaction positive en AgVHE.
En guise de contrôle ces 12 selles ont été testées à nouveau
dans notre système spécifique singe après passage sur
immunoadsorbant, et les résultats suivants ont été obtenus
RECHERCHE DE L'ANTIGENE VHE CHEZ DES SUJETS ATTEINTS
D'HEPATITE AlGUE NANB A DAKAR
Selles testées système Ts Système Sps
PIN PIN
Tl 0,80 0,60
T2 1,12 0,90
T3 0,83 0,72
T4 0,79 0,81
T5 1,25 0,98
81 1,12 0,93
82 1,89 1,13
83 0,78 0,66
84 0,74 0,80
85 0,92 0,90
86 1,42 1,23
87 1,66 1,67
88 1,06 4,5*
89 1,61 7,9*
810 1,31 12,6*
811 0,91 11,9*
812 1,09 6,3*
8 = selles de sujets atteints d'hépatite aiguë NANB à DAKAR
T selles de sujets témoins.
Tableau n° 22
85

Cinq de ces selles sont effectivement positives en AgVHE
soit un pourcentage de 41,66 % des hépatites non A non B
non C.
D'après cette étude, si aucun cas d'épidémie d'hépatite n'a
été signalé à ce jour au Sénégal, nous pouvons cependant affirmer
qu'il existe des cas sporadiques d ' hépatite non A non B qui
représentent 50 % des hépatites aiguës dans notre étude dont 17
% sont confirmés en VHE et 10% en VHC.
86

B - ETUDE EPIDEMIOLOGIQUE DE L'HEPATITE C
Cette étude a pour but :
- de déterminer la prévalence du portage des anticorps anti HCV
dans la population générale et chez les donneurs de sang ;
- de rechercher les facteurs de risque de portage des anticorps
antiHCV ;
- de rechercher une corrélation entre la présence des anticorps
anti HCV et des marqueurs indirects tels que les transaminases
sériques (SGOT - SGPT) dont la recherche systématique est rendue
obligatoire dans certains pays, mais non encore pratiquée au
Sénégal.
- d'évaluer le risque de transmission de l'hépatite C pour 100
unités de sang transfusées par rapport à d' autres affections
transmissibles par voie parentérale telles que la syphilis, le
SIDA ou encore l'hépatite B.
1 - Détermination de la prévalence du portage des
anticorps anti HCV dans la population générale.
Cette étude porte sur 950 suj ets apparemment sains, répartis
en deux groupes, representatifs de la population sénégalaise.
Le premier groupe comprend 500 sujets de 15 à 55 ans et le
deuxieme groupe 450 sujets de 55 à 90 ans.
Ces suj ets sont groupés par tranche d'âge et les sérums
sont étudiés unitairement à l'aide du test Elisa screening Ortho.
Chaque échantillon positif ou douteux est réétudié et
confirmé en immunoblot (Riba anti HCV Ortho) puis reclassé parmi
les sérums anti HCV positifs ou négatifs.
Les résultats montrent :
a) une prévalence globale de portage de 0,73 % avec une
certaine variabilité suivant la tranche d'âge considérée (cf.
histogramme de répartition des âges).
87
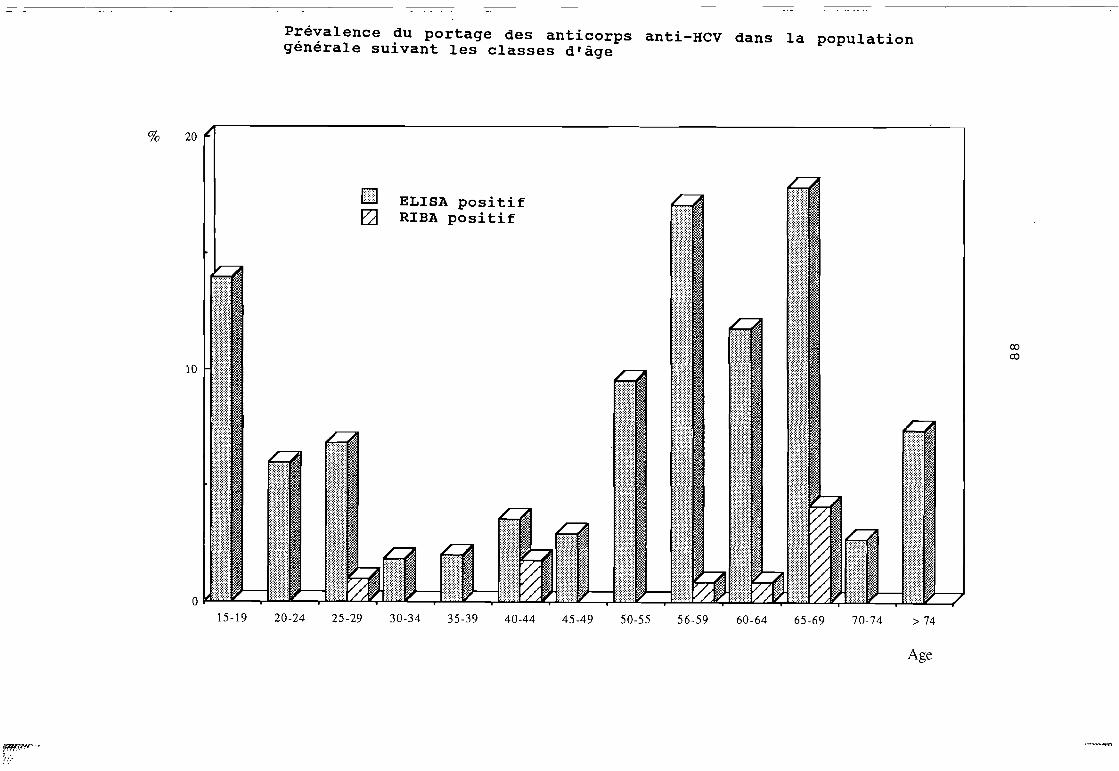
Lill ELISA positifEJ RIBA positif
% 20
10
Prévalence du portage des anticorps anti-HCV dans la populationgénérale suivant les classes d'âge
coco
ifll'f,~""'····, .~:.:.~
15-19 20-24 25-29 30-34 35-39 40-44 45-49 50-55 56-59 60-64 65-69 70-74 > 74
Age
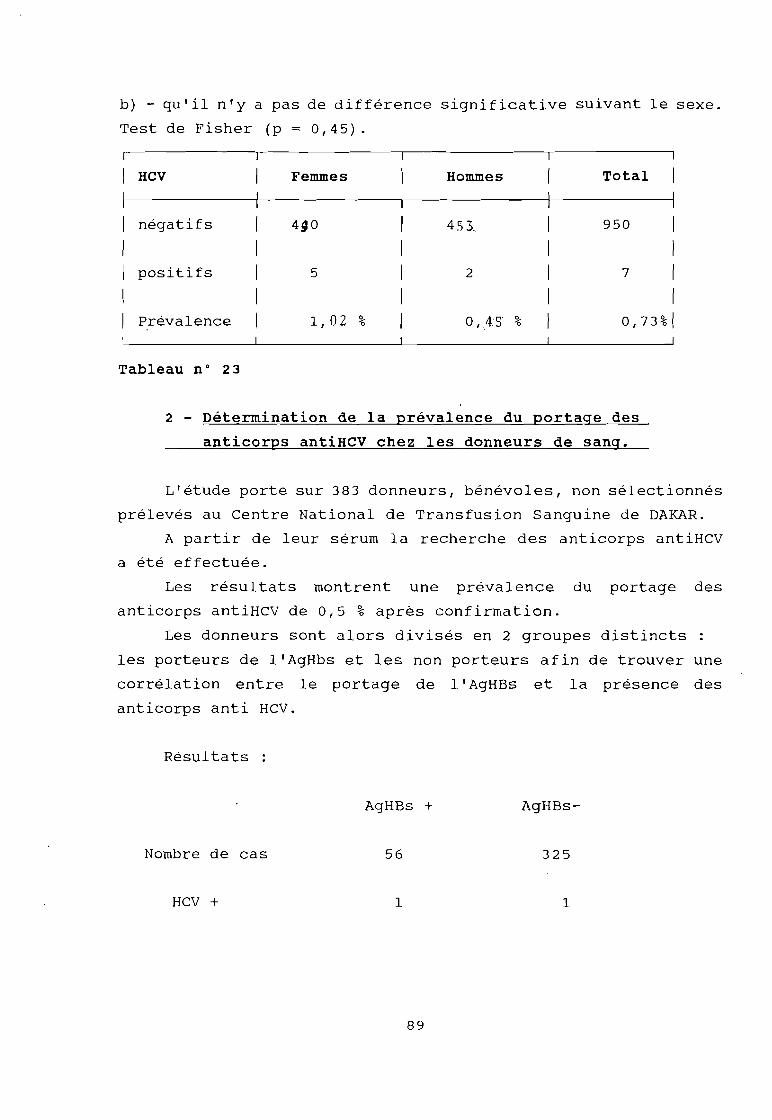
b) - qu'il n'y a pas de différence significative suivant le sexe.
Test de Fisher (p = 0,45).
HCV Femmes Hommes Total
négatifs 490 4S3. 950
positifs 5 2 7
Prévalence 1,02 ~ O,4S ~ 0,73%10 0
1
Tableau n° 23
2 - Détermination de la prévalence du portage des
anticorps antiHCV chez les donneurs de sang.
L'étude porte sur 383 donneurs, bénévoles, non sélectionnés
prélevés au Centre National de Transfusion Sanguine de DAKAR.
A partir de leur sérum la recherche des anticorps antiHCV
a été effectuée.
Les résultats montrent une prévalence du portage des
anticorps antiHcV de 0,5 % après confirmation.
Les donneurs sont alors divisés en 2 groupes distincts :
les porteurs de l'AgHbs et les non porteurs afin de trouver une
corrélation entre le portage de l' AgHBs et la présence des
anticorps anti HCV.
Résultats
Nombre de cas
HCV +
AgHBs +
56
1
89
AgHBs-
325
1

Soit une prévalence du portage des anticorps anti HCV de
1,78% chez les donneurs AgHBs+ versus 0,3 % chez les donneurs
AgHBs-. P = 0,27 non significatif.
3 - Recherche des facteurs de risque de portage des
anticorps antiHCV.
Compte tenu des données épidémiologiques et cliniques en
ce qui concerne l' hépati te C, 3 facteurs de risque ont été
recherchés :
- l'hospitalisation,
- la transfusion,
- antécédent d'hépatite.
Les résultats obtenus sont les suivants
Facteurs de 1 Nombre total HCV HCV
risque 1 positifs négatifsl
1
Hospitalisation 1 83 1 82
1
Transfusion 1 5 0 5
1
Antécédent 1 17 2 15
d'hépatite 1
1
Sans antécédent 1 769 4 765
Tableau n° 24
90

D'après ces résultats il n' y aurait pas de différence
statistiquement significative entre
le groupe témoin et le groupe de suj ets ayant déj à été
hospitalisés (p = 0,21),
- le groupe témoin et les sujets ayant déjà été transfusés.
Par contre le risque est net chez des sujets ayant des
antécédents d'hépatite (p = 0,000016).
Nous avons également cherché à établir une relation entre
la présence des anticorps antiHCV et des marqueurs indirects des
hépatites. Pour ce faire les sujets sont divisés en 2 groupes
- ceux ayant des transaminases normales (SGOT-SGPT < 25 UI/I)
- ceux ayant des transaminases élevées (SGOT-SGPT > 25 UI/I).
(technique Biomérieux)
Les résultats montrent
6
96
102
sujets ayant des
transaminases sé-I
riques élevées
1
847
848
sujets ayant des
transaminases sériques
normales
1
1
1
1---------------------------.-,
IHCV
Ipositifs
1
IHCV
1 négatifs
1
1Total
Tableau na 25
faut souligner que les deux donneurs
ont des transaminases élevées.
les 2 groupes est statistiquement
et on peut dire qu'il existe une
des anticorps antiHCV et le taux
La différence entre
significative (p = 8.10-6
corrélation entre le portage
de transaminases sériques.
Dans le même sens, il
antiHCV+ dans notre étude,
91

4 - Evaluation du risque de transmission de l'hépatite C
pour 100 unités de sang transfusées par rapport a
d'autres maladies transmissibles par voie parentérale.
Chez les donneurs de sang, si le dépistage n'était pas fait,
,le risque de transmission de :
- l'hépatite B serait de 14,62 % (73 donneurs/500 sont AgHbs+)
- celui de transmettre le SIDA serait de 1,04 % (6 donneurs/500
sont HIV+)
celui de transmettre la syphilis serait de 0,78 % (4
donneurs/500 sont BW +)
- et enfin celui de transmettre l'hépatite C est de 0,50 % (2
donneurs/500 sont antiVHC +) .
92

DISCUSSION GENERALE

l - DIAGNOSTIC BIOLOGIQUE DE L'HEPATITE E
A - ETUDE EXPERIMENTALE SUR L'ANIMAL
Cette étude que nous avons menée, permet de conclure
1. Les espèces de singe retrouvées au Sénégal sont
sensibles au VHE et pourront ainsi être utilisés comme source
d'antigène pour nos travaux ultérieurs.
2. La sensibilité des singes au VHE est variable d'une
espèce à l'autre.
3. Le temps d'incubation de la maladie varie de 10 à 30
jours. Ce délai a largement été confirmé par de nombreux auteurs
(23-26-65) et lion retient aujourd'hui que la période
d'incubation varie en moyenne de 20 à 32 jours.
4. L'élévation des transaminases constitue un bon
marqueur biochimique de l'hépatite E.
5. Le virus de l' hépatite E est probablement sensible
au chloroforme voir même inactivé (30).
Cette sensibilité du virus à différents agents chimiques a
également été rapportée par d'autres auteurs (31-71) et
récapitulée dans le tableau suivant
PROPRIETES PHYSICO-CHIMIQUES ET BIOLOGIQUE DES AGENTSRESPONSABLES DES HEPATITES NANB
Filtration 220 nm
chaleur (60 0, 10h)
Formol (1/1000)
Chloroforme 10%
Ethanol
Tableau n° 26
93
passe
sensible
sensible
sensible
sensible ou non

Quant aux méthodes que nous avons utilisées pour le
dépistage des sujets infectés par le VHE, une étude critique
nous permettra de mieux opter pour une technique.
B - ETUDE CRITIQUE DES TECHNIQUES UTILISEES
POUR LE DEPISTAGE DES SUJETS INFECTES PAR LE VHE
1 - L1immunofluorescence pour la recherche des
anticorps antiVHE
Par la méthode de recherche que nous avons utilisée, nous
n'avons pu détecter aucun anticorps antiVHE dans les sérums
testés parce que très probablement le foie que nous avons utilisé
comme source d'antigène ne contenait pas le VHE.
En effet, une étude américaine ultérieure menée au Center
of Diseases Control par BRADLEY et collaborateurs (16), et
utilisant la même technique, s'est soldée par des résultats
positifs. Ils ont pu retrouvé le VHE dans les hépatocytes de
foie de singe infecté expérimentalement le premier jour de la
montée des transaminases, et ont mis au point un système de
détection des anticorps antiVHE à partir de ce foie (17).
Nous avons donc déduit que la réplication du VHE dans le
foie était très brève et que les biopsies que nous avons
utilisées avaient été réalisées trop tardivement.
Par ailleurs, il nous plaît de souligner que nos différents
sérums ont été contrôlés par la même équipe et que le sérum de
notre singe S8 a été retrouvé positif en anticorps antiVHE après
inoculation et négati f avant inoculation. ·Mais il faut le
reconnaître il s'agit là d'une méthode qu'on ne peut utiliser en
routine et qui reste réservée à la recherche; parce que :
- les réactifs ne sont pas renouvelables,
- il y a difficulté de standardisation.
Les appl ications possibles seraient l 'util isation dl un sérum
confirmé positif en anticorps, marqué, qui servirait à rechercher
l'antigène VHE dans les coupes de foie; ou bien l'utilisation
d'un système foie positif/sérum marqué positif utilisable pour
94

la sérologie en série dans un test de blocage (les sérums étudiés
seront alors utilisés non marqués) .
2 - Recherche du virion dans les selles
a) - par microscopie électronique
Bien qu'ayant multiplié nos recherches et essayé de
nombreuses techniques de microscopie électronique, les résultats
sont restés négatifs, cependant qu'il était montré dans certaines
études la présence de particules de 27 à 32 nm dans les selles
de suj ets atteints d 1 hépatite E (15). Mais il convient de
préciser que ces particules n'étaient retrouvées que dans 4 % des
cas.
De ces expériences nous avons déduit :
- que la quantité de VHE éliminée dans les selles était faible
nécessitant donc un enrichissement. Cette déduction était
d'autant plus logique que des méthodes de purification virale
par gradient de sucrose ou de chlorure de Césium que nous avons
entreprises ne nous ont jamais permis d'isoler le virus à partir
des selles. Toutes ces difficultés, de même que le faible
pourcentage de positivité constaté limitent donc l'utilisation
de cette méthode.
b) - par culture cellulaire
Tous nos essais de culture cellulaire sur lignée
hépatocytaire HepG2 et PLcPRF5 ne nous ont pas permis d'observer
un effet cytopathogène caractéristique du VHE.
Le VHE ne provoque-t-il pas d'effet cytopathogène ? In
vitro, il est difficile de répondre à cette question puisque
jusqu'à ce jour aucune équipe de chercheurs n'a pu cultiver ce
virus.
si cet échec constaté face aux techniques usuelles de
culture virale pourrait s'expliquer par une fragilité du virus,
ou encore par des conditions de conservation difficiles à
respecter il semble plutôt lié à la nature du produit
95

pathologique en l'occurrence les selles qui peuvent contenir de
nombreuses substances cytotoxiques.
Il faudrait dans ce cas peut-être avoir recours à des
techniques de purification virale à partir des selles, ou de la
bile, où le virus semble également présent avant d'entreprendre
la culture cellulaire.
In vivo, les biopsies hépatiques que nous avons effectuées
chez le singe infecté expérimentalement n'ont révélé aucune
lésion caractéristique au niveau des hépatocytes.
Par contre chez l'homme des études bien documentées portant
sur des biopsies de foie réalisées chez des sujets décédés
d'hépatite E fulminante ont permis de décrire des lésions
caractéristiques (22-35-64-65). Mais les différents résultats
présentés ne sont pas tous concordants et des investigations plus
poussées seraient certes nécessaires.
3 - Recherche de l'antigène VHE par techniques
immuno-enzymatiques.
Les différentes techniques mises au point permettent de
tirer les conclusions suivantes :
La recherche de l'antigène VHE dans les selles à partir
d'un système humain donne de moins bons résultats que le système
singe. Nous avons émis l'hypothèse que dans les selles on peut
retrouver des substances telles que les lipopolysaccharides
bactériens, et certains germes saprophytes de la flore
intestinale, qui peuvent se lier aux sites anticorps des IgM
fixées sur le support réactionnel, et aux fragments Fab des IgG
marquées donnant des réactions non spécifiques (95 % dans notre
étude). C'est la raison pour laquelle nous avons abandonné le
système humain au profit du système singe que nous avons
optimisé.
En fait, à côté des substances d'origine bactérienne de la
flore intestinale saprophyte il existe d'autres substances non
spécifiques pouvant se lier soit aux IgG, soit aux IgM de manière
non immune et pouvant interférer avec le système spécifique.
En effet, d'après nos résultats, la protéine Fv que nous
96

avons décrite précédemment semble être la principale cause de
ces réactions non spécifiques puisque le passage des selles sur
immunoadsorbant d' IgM monoclonale qui élimine la protéine Fv fait
disparaître ces réactions dans 100 % des cas.
Mais il faut cependant remarquer que lorsque le seuil de
positivité est faible (PIN < 3) l'utilisation de
l'immunoadsorbant peut perturber les réactions dans la mesure
où une partie de l'antigène spécifique peut se fixer sur
l'immunoadsorbant donnant ainsi de faux résultats négatifs. Dans
de tels cas, le contrôle devra Si effectuer par des réactions
d'inhibition sur des selles non adsorbées.
Mais il faut cependant dire que si la protéine Fv est source
de réactions non spécifiques, sa recherche semble fort
intéressante dans la mesure où elle est le témoin précoce d'une
inflammation hépatique. Chez le singe, elle apparait 20 jours
après 11 inoculation alors que les transaminases ne s'élèvent
qu'au bout de 25 jours.
Chez l'homme, à défaut d'étude menée avant la montée des
transaminases, ont peut dire que son apparition dans les selles
est contemporaine de l'élévation des transaminases sériques.
Le passage en revue de toutes les techniques que nous avons
util isées pour le dépistage des suj ets infectés par le VHE,
révèle que les meilleurs résultats ont été obtenus avec les
techniques immunoenzymatiques; qui pour être valables, rappelons
le doivent utiliser :
- des anticorps antiVHE d'origine non humaine
- des IgG marquées à la béta-galactosidase qui révèlent moins
la protéine F que les IgG marquées à la péroxydase. Des études
menées par l'équipe du Professeur Jacques PILLOT (61) l'ont
effectivement prouvé.
et enfin des selles passées sur immunoadsorbant d'IgM
monoclonale pour éliminer les réactions non spécifiques.
97

· '..~"!§,
;.
C- ACQUISITIONS ET PERSPECTIVES
Grâce à ces techniques immuno-enzymatiques, nous avons pu
mettre au point une méthode spécifique reproductible de
détection de l'antigène VHE dans les selles des malades infectés.
Malgré une variabilité des souches de VHE provenant de
différentes régions du globe, signalée par certains auteurs,
nous pouvons dire qu'il existe au moins une communauté
antigénique entre ces différentes souches.
En effet par cette technique, nous avons pu retrouver
l'antigène VHE dans les selles de malades atteints d'hépatite E
de différentes provenances
- de la Thaïlande (dans 26 % des selles de malades de Bangkok)
- de l'Afrique du Nord (dans 38 % des selles de malades de
Constantine) .
- de l'Afrique de l'Ouest, au Sénégal où nous avons retrouvé des
cas sporadiques d'hépatite E à Dakar (33 % des hépatites NANB
dans notre étude).
Mais il faut cependant remarquer que cet antigène apparaît
précocement dans les selles, dès la période d'incubation chez
l'animal, pendant la montée des transaminases chez l'homme et
que par cette technique le diagnostic de l'hépatite E ne peut se
faire que pendant la phase aiguë de la maladie.
Mais si l'on sait aujourd'hui qu'aucun cas d'hépatite E
chronique n'a été signalé chez l'homme ou chez l'animal, on peut
dire que la majeure partie des hépatites E pourront être
diagnostiquées par cette méthode. Il s'agit là d'une contribution
importante dans le domaine de recherche des hépatites, surtout
si l'on sait qu'à l' heure actuelle nous ne disposons d'aucun test
utilisable en routine pour le diagnostic de l'hépatite E.
Cependant une question subsiste : Quand disparaît l'antigène
VHE chez l'homme?
98

Si nous pouvons donner une réponse à cette question en ce
qui concerne le singe où l'antigène n'est plus détectable au
delà du 42ème jour après inoculation, chez l'homme un suivi dans
le temps de l'évolution de cet antigène s'avère nécessaire.
Des études prochaines nous permettront certainement de
répondre à cette question.
Mais l'apparition précoce de l'antigène détecté qui apparaît
avant la montée des transaminases chez l'animal, sa présence de
courte durée dans les selles, de même que les problèmes inhérents
au traitement et à la conservation des selles, rendent bien
compte de l'intérêt de développer un test sérologique de
détection des anticorps antiVHE, à partir d'antigène produit chez
le singe.
C'est pourquoi nous incluons dans nos perspectives
immédiates de recherche l'inoculation de plusieurs singes genre
cercopithèque, afin d'obtenir à partir de leur foie, ou de leur
selle, ou de leur bile, une quantité suffisante en antigène VHE.
Ainsi, nous pourrons réaliser:
- soit des monoclonaux à partir du VHE pour la recherche des
anticorps antiVHE dans le sérum ou le plasma;
- soit mettre au point une technique de dépistage par
"Dot-Blot" par fixation directe de l'antigène VHE sur membrane.
Ainsi nous pourrons étudier l'évolution des anticorps
antiVHE chez le singe et par transposition chez l' homme; et
disposer de tests d'utilisation plus malléables en routine.
Mais dores et déjà nous pouvons dire que nous sommes armés
si une épidémie d'hépatite E arrivait à se déclarer au Sénégal.
99

Par ailleurs, dans le cadre de la prévention de l'hépatite,
nous incluons dans nos perspectives de recherche, la préparation
d'immunoglobulines à partir de sujets ayant été infectés par le
VHE, et ayant acquis une immunité. L'injection de telles
immunoglobulines pourrait assurer une immunisation passive chez
des sujets à risque, et plus particulièrement chez la femme
enceinte, chez qui une telle prophylaxie peut être
particulièrement intéressante en attendant de disposer d'un
vaccin éventuel.
100
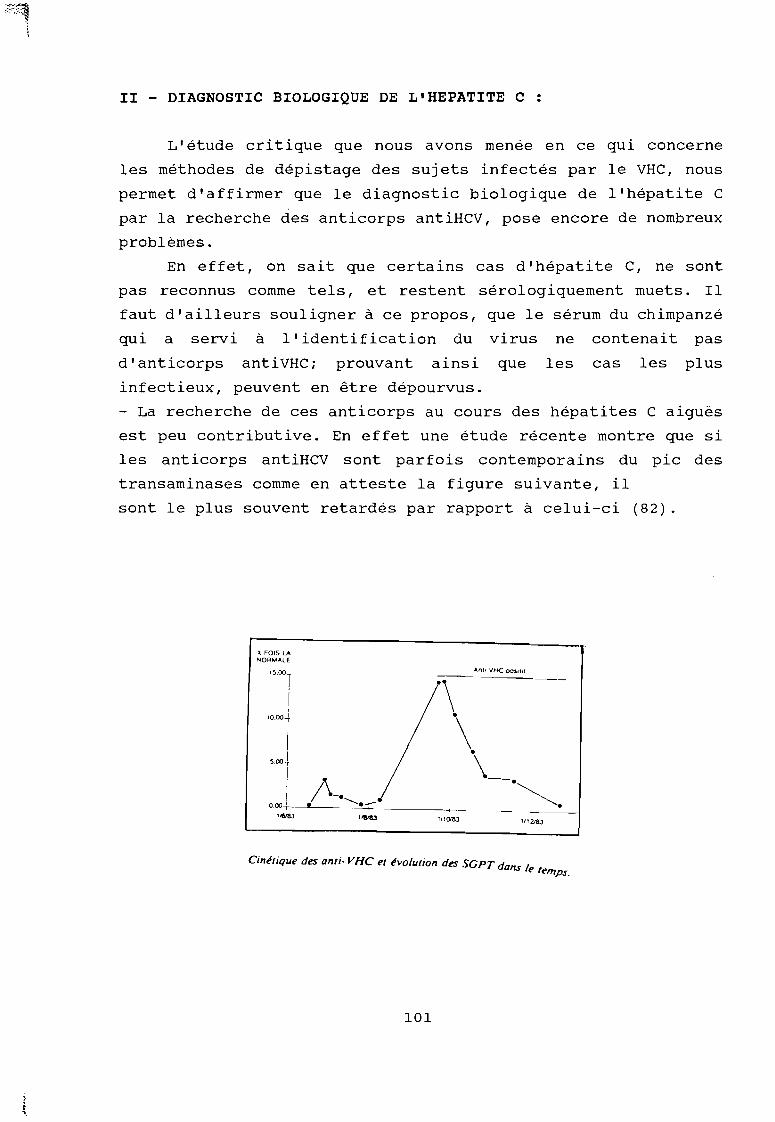
II - DIAGNOSTIC BIOLOGIQUE DE L'HEPATITE C :
L'étude critique que nous avons menée en ce qui concerne
les méthodes de dépistage des sujets infectés par le VHC, nous
permet d'affirmer que le diagnostic biologique de l'hépatite C
par la recherche des anticorps antiHCV, pose encore de nombreux
problèmes.
En effet, on sait que certains cas d'hépatite C, ne sont
pas reconnus comme tels, et restent sérologiquement muets. Il
faut d'ailleurs souligner à ce propos, que le sérum du chimpanzé
qUl a servl à l'identification du virus ne contenait pas
d'anticorps antiVHC; prouvant ainsi que les cas les plus
infectieux, peuvent en être dépourvus.
- La recherche de ces anticorps au cours des hépatites C aiguës
est peu contributive. En effet une étude récente montre que si
les anticorps antiHCV sont parfois contemporains du pic des
transaminases comme en atteste la figure suivante, il
sont le plus souvent retardés par rapport à celui-ci (82).
1/12r8J1/1QI8J
X FOIS LANORM~lE
l~.ooT .\i
"Wr \5.00~ e\
1 _
o.oo~1----I../_\_-._.........=.---=--_--+-__.~~~11618J , l8I8J
Cinétique des anti- VHC et évolution des SGPT d 1
aIlS le temps.
101
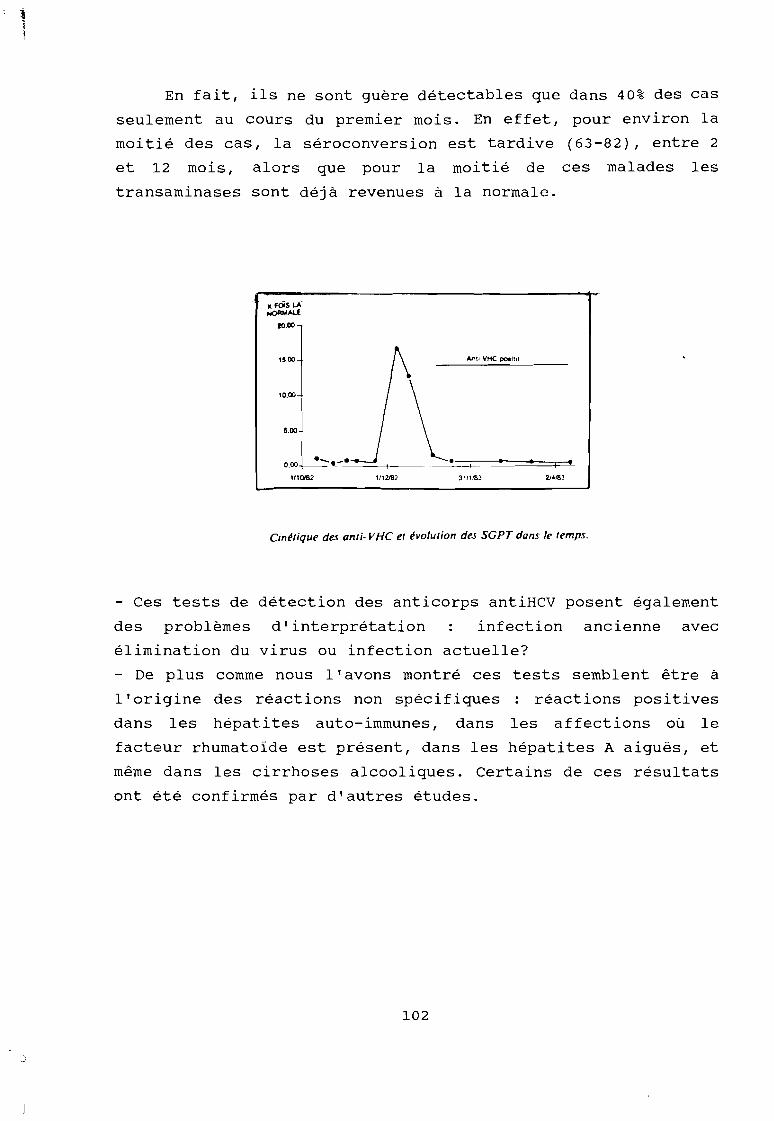
l1,
En fait, ils ne sont guère détectables que dans 40% des cas
seulement au cours du premier mois. En effet, pour environ la
moitié des cas, la séroconversion est tardive (63-82), entre 2
et 12 mois, alors que pour la moitié de ces malades les
transaminases sont déjà revenues à la normale.
1D~1
u.oo1
,0.ooJ \
~'ooL0 00 .~.-. •• 1
11 10m2 1/12182
•3111/83
•1 •
V4/8.:l
j
Cin~tique des anti· VHC et ~volution des SGPT dans le temps.
- Ces tests de détection des anticorps antiHCV posent également
des problèmes d'interprétation infection ancienne avec
élimination du virus ou infection actuelle?
- De plus comme nous l'avons montré ces tests semblent être à
l'origine des réactions non spécifiques réactions positives
dans les hépatites auto-immunes, dans les affections où le
facteur rhumatoïde est présent, dans les hépatites A aiguës, et
même dans les cirrhoses alcooliques. Certains de ces résultats
ont été confirmés par d'autres études.
102

Plusieurs hypothèses sont avancées pour expliquer ces
probables faux positifs.
- La présence au cours des hépatites auto-immunes d'un (auto)
anticorps oligoclonal de spécificité indéterminée, réagissant
également avec l'antigène C 100-3 du kit. (54).
- La présence d'un anticorps anti-idiotype se fixant à la phase
solide du kit et liant également les IgG. (54).
- Ou bien la présence d'anticorps circulants dirigés contre la
superoxyde-dismutase (SaD). En effet on sait que l'antigène cible
des tests HCV ELISA est un polypeptide de fusion C 100-3/S00; et
le contrôle des sérums de 19 sujets antiHCV positifs, dont 10
sont atteints d'hépatite NANB chronique post-transfusionnelle,
et 9 d'hépatite chronique active auto-immune, par une recherche
d'anticorps anti-sOO, montre que les antigènes SaD sont toujours
absents dans le premier groupe et touj ours présents dans l'autre.
(43) .
- Ou la présence d'anticorps de faible affinité, plus sensibles
au lavage à l'urée 8 moles/l, seraient également responsables de
certains faux positifs des tests HCV ELISA, observés au cours
d'hépatopathies variées (87).
- Ou encore la présence d'anticorps non spécifiques dirigés
contre des Togavirus ou de Flavivirus autres que le virus de
l'hépatite C,circulant en milieu tropical ,qui réagiraient
également avec la proteine C 100-3 du kit (étude en cours par
notre équipe) .
De plus des discordances très nettes apparaissent suivant
les techniques utilisées A titre d'exemple 4 % de
séropositivité en anticorps antiHCV ont été observés chez les
donneurs de sang sénégalais en "screening Elisa" versus 0,4 %
en "Immunoblot".
Cette situation semble préoccupante surtout si l'on sait
que la maj eure partie des études épidémiologiques dont nous
disposons à l' heure actuelle ont été effectuées avant
l'utilisation de ces tests de confirmation (24-25-44-52). Ces
études ne doivent être interprétées qu'après contrôle par ces
tests.
103

Tous ces problèmes mettent l'accent sur la nécessité de
disposer de tests plus performants , plus spécifiques donc plus
fiables surtout si l'on sait que le VHC est caractérisé par un
vaste réservoir de porteurs chroniques asymptomatiques dont
certains évoluent de manière insidieuse vers la cirrhose et le
cancer primitif du foie.
104

III - ETUDE EPIDEMIOLOGIQUE DES HEPATITES NANB
L'étude épidémiologique que nous avons entreprise permet
de tirer ies conclusions suivantes :
- l'hépatite E existe bien au Sénégal. Dans notre échantillon,
elle représente 42 % des hépatites NANBNC et 17 % des hépatites
aiguës chez l'adulte.
Ce pourcentage semble nettement inférieur à celui observé
dans certains pays en voie de développement où l'on rapporte que
l'hépatite E représente entre 50 et 90% des hépatites virales
aiguës (65). Une enquête à plus large échelle, serait peut-être
nécessaire pour confirmer le pourcentage que nous avons obtenu.
Cependant ce pourcentage relativement faible, ne doit
nullement détourner notre attention du risque d'épidémie qui
existe; épidémies, rappelons le, sont survenues dans de nombreux
pays d'Afrique et d'Asie.
- Toujours dans notre étude, cette hépatite ne concerne que les
jeunes adultes de 21 à 40 ans. Ces résultats semblent donc
confirmer les conclusions de l'étude menée par RAMALINGASWANI
V. et collaborateurs (59) qui disaient je cite : "l' hépatite
NANB à transmission entérale touche avec une égale fréquence
l'homme et la femme et s'observe principalement chez le jeune
adulte entre 15 et 40 ans".
Quant à l'étude épidémiologique de l'hépatite C, elle nous
permet de constater :
- le faible pourcentage du portage des anticorps antiHCV dans
la population générale (0, 73 %) et chez les donneurs de sang
(0,5%); pourcentages certes sous estimés, si l'on sait qu'une
étude récente a montré la possibilité de détecter par PCR l'ARN
HCV dans le sérum et le foie de sujets apparemment séronégatifs,
incluant des sujets avec hépatite chronique, mais également des
donneurs de sang. Elle nous permet également de constater que la
prévalence des anticorps anti-HCV augmente avec l'âge jusqu'à la
quatrième décennie, puis décroit ensuite, reflétant probablement
la disparition des anticorps détectables avec le temps. Il faut
remarquer que de telles conclusions sont également rapportées
105

par une étude américaine récente. (72).
Cependant, on constate également dans notre étude une
prévalence relativement élevée de 4,12% (4 antiHCV+/97 étudiés)
dans la tranche d'âge G5~G9 ans. Des études ultérieures pourront
certainement nous éclairer sur cette forte prévalence.
de formuler les recommandations suivantes les tests
actuellement commercialisés pour la détection des anticorps
antiHCV par screening Elisa ne doivent pas être utilisés de
manière systématique dans nos centres de transfusion sanguine.
En effet ils donnent 90% de réactions positives non spécifiques.
Et si l'on sait que près de 20% des donneurs de sang sont
éliminés puisque porteurs de l'AgHBs, plus de 2% éliminés puisque
porteurs du HIV ou du Tréponéma pallidum; ces fausses réactions
positives, ne feraient que limiter à tort, le nombre de donneurs
déjà nettement insuffisant; sans vouloir parler du coût élevé de
ces recherches, qui serait de 12.500 F CFA pour un seul
dépistage.
A défaut, l'utilisation de mélanges de solvants et de
détergents destinés à l'inactivation virale serait peut être à
envisager au moins pour la préparation des fractions
plasmatiques. Ces procédés sont aujourd'hui couramment employés
et ont fait la preuve de leur efficacité dans certains pays
d'Europe. (42).
- De proposer un schéma simple, qui pourrait être retenu devant
tout don de sang en attendant de disposer de tests plus fiables
pour le diagnostic de l'hépatite C, et plus accessibles sur le
plan économique.
lOG
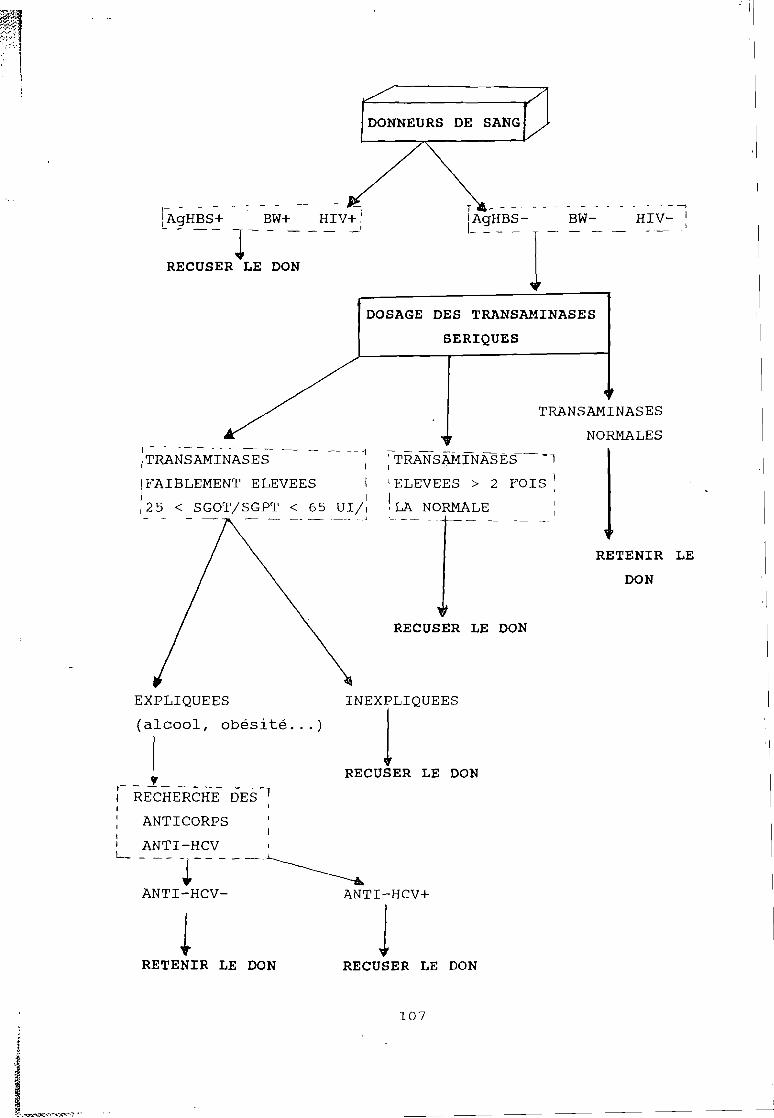
DONNEURS DE SANG
- - - - -
lA9"HBS~ BW+ HIV+ :1- - - --RECUSER LE DON
.. 1
DOSAGE DES TRANSAMINASES
SERIQUES
TRANSAMINASES
NORMALES1 -- -. - - - - - - - - -- 1,TRANSAMINASES 1
IFAIBLEMENT ELEVEES 11 1
,25 < SGOT/SGPT < 65 UII,
~TRANSAMfuASES-- ~l,'ELEVEES> 2 FOIS J
!LA NORMALE '
RETENIR LE
DON
RECUSER LE DON
INEXPLIQUEES
lRECUSER LE DON
RECUSER LE DONRETENIR LE DON
EXPLIQUEES
(alcool, obésité ... )
,__ L_ _1 RECHERCHE DES l1 1
: ANTICORPS11 ANTI-HCV 1
L---l-----~
ANTI-HCV- ANTI-HCV+
1107

En effet ce schéma s'inspire des résultats d'une étude
hollandaise (56) qui montre une prévalence du portage des
anticorps antiHCVde 0,6% chez des donneurs de sang à
transaminasémie normale; de 10% à transaminasémie faiblement
élevée (une à deux fois la normale), mais de manière répétée;
de 24% chez les donneurs dont le taux de transaminases sériques
est supérieur à deux fois la normale.
Il s'inspire également de notre étude dakaroise où nous
avons constaté que 80% des sujets porteurs des anticorps anti
HCV, et 100% des donneurs de sang anti-HCV+, ont un taux de
transaminases sériques élevé avec une moyenne de 70 UI/l pour
les SGOT et les SGPT.
Donc en attendant de disposer de tests plus fiables pour
le diagnostic de l'hépatite C, le dosage systématique des
transaminases sériques chez les donneurs de sang constituerait
une prévention au moins partiellement efficace de l' hépatite
NANB post-transfusionnelle.
108

CONCLUSION

Au terme de notre étude, nous pouvons dire que les tests
commercialisés actuellement pour le dépistage des sujets infectés
par le VHC permettent de détecter des anticorps dirigés contre
un antigène exprimé lors d'une hépatite NANB appelée hépatite
« C ».Ces tests permettent certes de détecter 50 à 80 % des
hépatites transmises par voie parentérale autres que l'hépatite
B permettant ainsi de réduire considérablement les hépatites
post-transfusionnelles.
cependant ces tests posent encore de sérieux problèmes :
- les anticorps antiHCV ne sont détectables en moyenne que dans
30 à 40 % des hépatites aiguës, ils n'apparaissent qu'après 3 à
4 mois d'évolution. Il paraît donc essentiel de pouvoir disposer
d'un marqueur plus précoce de l'infection, tant en ce qui
concerne les exigences de la sécurité transfusionnelle, que
celles du diagnostic clinique des infections aiguës.
De plus comme nous l'avons montré, ces tests semblent être
à l'origine de nombreuses réactions non spécifiques; 50 à 60 %
dans certaines situations pathologiques, 90 % en milieu tropical
plus précisément à DAKAR où notre étude a été menée.
Ces réactions non spécifiques semblent liées au fait que
l'antigène utilisé pour la détection des anticorps antiVHC est
une protéine fonctionnelle et non pas structurale du VHC.
Dès lors, il parait imperleux de se diriger vers le
recherche d'autres marqueurs plus spécifiques, pour un diagnostic
biologique fiable de l'hépatite C.
L'espoir est permis, si l'on sait aujourd'hui,qu'une équipe
de chercheurs japonais a pu isoler une protéine de structure du
VHC de 22 KD, reconnue par les anticorps présents dans le sérum
d'un patient atteint d'hépatite NANB chronique.
Cette protéine a déjà été partiellement purifiée en vue de
la préparation d'un test ELISA pour la détection d'un anticorps
anti 22KD dans les sérums de patients atteints d'hépatite NANB.
Les essais préliminaires utilisant ce test encore à l'état
de recherche montrent que non seulement la majorité des hépatites
C chroniques sont diagnostiquées, mais également les hépatites
C aiguës.
109

diagnostic
en pleine
De tels tests sembleraient donc plus performants pour le
diagnostic biologique de l'hépatite C.
Sinon, un appel à l'amplification genlque comme l'a déjà
suggéré HOUGHTON serait peut-être nécessaire.
En effet, la PCR permet la détection de l'ARN HCV, et ce
de façon précoce, puisque dans les expériences de transmission
à des chimpanzés, l'ARN a pu être détecté dans les premiers jours
suivant l'inoculation. La PCR devrait donc pouvoir permettre une
estimation sensible et très directe de la virémie. La PCR peut
cependant poser plusieurs problèmes
- risque de contamination inhérent à ces techniques.
- nécessité d'utiliser des séquences adaptées à l'isolat du fait
de la variabilité génétique éventuelle. (47).
- nécessité d'une bonne conservation des sérums car l'ARN HCV
est manifestement facilement dégradé.
Seules des comparaisons permettront de dire si l'estimation
de la virémie peut être réalisée au mieux par des tests
sérologiques, ou par des techniques de biologie moléculaire comme
la PCR.
C'est dire donc que les méthodes actuelles de
de l' hépatite C loin d'être satisfaisantes, sont
évolution et ouvrent ainsi une voie de recherche.
Quant au diagnostic biologique de l'hépatite E, comme nous
l'avons exposé précédemment, nous avons pu mettre au point une
méthode spécifique de détection de l'antigène VHE, qui a permis
de nous rendre compte de l'existence du VHE au Sénégal.
Même si le nombre de cas étudié semble limité, les résultats
sont statistiquement interprétables et nous pouvons dire que
l'hépatite E représente 42 % des hépatites NANBNC et 17 % des
hépatites aiguës de l'adulte dans notre échantillon.
110

Cependant en combinant les tests actuels de détection des
anticorps antiHCV et notre méthode de détection de l'antigène
VHE, on se rend compte que 47 % des hépatites NANB aiguës à DAKAR
restent encore non identifiées.
Nous savons déj à qu'il existe des formes d' hépatite C
séronégatives, et des études en immunomarquage et amplification
moléculaire dans le foie ont déjà permis de le démontrer dans au
moins 15 % des cas (18). Cette situation est finalement
comparable à celle toujours d'actualité des hépatites B
séronégatives au cours desquelles le VHB est uniquement
détectable par PCR, ces cas étant toujours classés
sérologiquement dans le groupe des hépatites NANB résiduelles
(18). Cette situation démontre bien la nécessité de disposer de
tests spécifiques, fiables, très performants pour pouvoir parler
d'hépatite C ou d'hépatite E et pose par là même le problème de
la terminologie d' hépatite NANB qui devient impropre, et la
suggestion de certains auteurs (18-81) de parler "d'hépatites
séronégatives" devient justifiée.
Ce terme refléterait ainsi le besoin de rechercher de
nouveaux marqueurs, tant pour le VHB que pour le VHC ou le VHE
ou tout autre virus. Et ce n'est que grâce à de tels progrès
qu'il sera possible de préciser l'existence de virus
supplémentaires, et de pouvoir répondre à la question que l'on
se pose actuellement, à savoir
"Existe-t-il plus de cinq virus des hépatites 7".
III

BIBLIOGRAPHIE

1
2
3
4
5
6
7
Al Arabi MA, Hyams KC, Mahgoub M, Al Haq AA, El
Ghoreb N. (1987). Non-A non-B hepatitis in Omdurman,
Sudan. J Med Virol, 21, 217-222.
Alter HJ, Purcell RH, Holland PV et al. (1978).
Transmissible agent in non-A non-B hepatitis. Lancet
1, 459-463.
Alter MJ, Gerety RJ et al. (1982). Sporadic non-A
non-B hepatitis. Frequency and epidemiology in an
urban US population. J. Infect Dis, 145, 886-893.
Alter, Purcell HJ et al. (1984). Non-A non-B
hepatitis. A review and interim report of an
ongoing prospective study. In : Viral hepatitis.
Edited by GN Vyas, Franklin Institute Press.
Philadelphia, 359-369.
Alter MJ, purcell RH, shih JW, Melpolder JC,
Houghton M, Choo QL, Kuo G. (1989). Detection
of antibody to hepatitis C virus in prospectively
followed transfusion recipients with acute and
chronic non-A non-B hepatitis. N Engl J Med, 321,
1494-1500.
Andzhaparidze AG, Balayan MS, Savinov AP, BRAGINSKY
DM et al. (1986). Non-A non-B hepatitis transmitted
by the fecal oral mode experimentally produced in
monkeys. Vopr. Virusol., 31, 73-81.
Arankalle VA, Ticehurst J, Sreenivasan MA et al.
(1988). Aetiological association of a virus-like
particle with enterically transmitted non-A non-B
hepatitis. Lancet, 1, 1550-553.
112

8
9
10
11
12
13
14
Balayan MS, Agafonov VI, Andzhaparidze AG et al.
(1982). Viral hepatitis, similar in clinical picture
and epidemiology to hepatitis A but differing from
it, etiologically. Microbiol Epidemiol Irnrnunobiol,
8, 79-84.
Balayan MS, Andjaparidze AG, Savinskaya SS et al.
(1983). Evidence for a virus in non-A non-B
hepatits transmitted via the fecal oral route.
Intervirology, 20, 23-31.
Bellabes H, Bougerrnouh A, Benatallah A, Illoul G
(1982). Study of an outbreak of non-A non-B
hepatitis in Algeria. Proceeding of the VIII IFIPD
Congress Session Non-A non-B hepatitis, Stockholm,
Sweden.
Bellabes EH, Bourguerrnouh A, Benatallah A, Illoul G
(1985). Epidemie non-A non-B hepatitis in Algeria
Strong evidence for its spreading by water. J. Med
Virology, 16, 257-263.
Betman M, Alter HJ, Ishak KG, purcell RH et al.
(1979). The chronic sequelae of non~A non-B
hepatitis. Ann Inter Med., 91, 1-6.
Blurnberg BS, Gerstley BJS, Hugerford DA, London WT,
Sutnick AI. (1967). A serum antigen (Australia
antigen) in down's syndrome, leukaemia and
hepatitis. Ann Inter Med., 66, 924-930.
Bradley DW, Maynard JE, Popper H et al. (1983).
Post-transfusion non-A non-B hepatitis
physico-chemical properties and two distinct agents.
J Inf Dis, 148, 254-265.
113

15
16
17
Bradley DW, Krawczynski K., Cook EH Jr et al.
(1987). Enterically transmitted non-A non-B
hepatitis : seriaI passage of disease in cynomolgus
macaques and tamarins and recovery of disease
associated 27 to 34 nm virus-like particles. Proc
Natl Acad Sei, USA, 84, 6277-6281.
Bradley DW, Andjaparidze, Cook EH et al. (1988).
Aetiological agent of enterically transmitted non-A
non-B hepatitis. J Gen Virol, 69, 731-738.
Bradley DW (1989). The virus of enterically
transmitted non-A non-B hepatitis. Symposium of
non-A non-B hepatitis and blood borne infectious
diseases, Tokyo, Elsevier (à paraître) .
18 Brechot C. (1990) Le virus de l' hépatite C
découverte de la biologie moléculaire.
Gastroentérol. Clin Biol, 14, 54-56.
une
19
20
21
Charkraborty S, Datta M, Pasha ST, Kumar S. (1982).
Non-A non-B hepatitis. A common source outbreak
traced to savage contamination of drinking water.
J. Commun Dis, 14, 41-46.
Choo QL, Kuo G, Weiner AJ, Overby LR, Bradley DW,
Houghton M. (1989). Isolation of a cDNA clone
derived from a blood borne non-A non-B hepatitis
genome. Science, 244, 359-361.
Decodk KM, Bradley DW, Sandford NG et al. (1987).
Epidemie non-A non-B hepatitis in patients from
Pakistan. Ann Internat. Med., 106, 227-230.
114

22
23
24
25
26
27
28
29
Desmet VJ, Devos R. (1985). Ultrastructural findings
in non-A non-B hepatitis. In Brunner H, Thaler H
(eds). "Hepatology : A fetschrift for Hans Popper",
New-York, Raven Press, 159-175.
Dienstag JL. (1983). Non-A non-B hepatitis.
1. Recognition, epidemiology and clinical features.
II. Experimental transmission putative virus agents
and markers and prevention. Gastroenterology, 85,
439-462.
Esteban M, Viladomin l, Gonzales A, Roget M et al.
(1989). Hepatitis C virus antibodies among risk
group in Spain. Lancet, 2, 294-297.
Farrow LJ, Stewart JS, Stern H, Clifford RE et al.
(1981). Non-A non-B hepatitis in West London. Lancet
1, 982-984.
Favorov MO, Khukhlovich PA, Zavrov GK et al. (1986).
Clinical and epidemiological features and diagnosis
of viral non-A non-B hepatitis with fecal oral
transmission mechanism. Vopr Virusol, 1, 65-69.
Y;>--,\, \~j9.oJFeinstone SM, Zapilian AZ, Purcell RH. (1973).
Detection by immune electronmicroscopy of a virus
like antigen associated with acute illness.
Science, 182, 1026-1028.
Feinstone SN, Kaprikran AZ, Purcell RH. (1975).
Transfusion associated hepatitis not due to viral
hepatitis A or B. The New England Journal of
Medicine, 292, 767-770.
Feinstone SN, Hoofnagle JM. (1984). Non-A non-B
may be B hepatitis. The New England Journal of
Medicine, 311, 185-188.
115

30
31
32
33
34
35
36
37
Feinstone SN, Mihalih et al. (1987). Inactivation
of hepatitis B virus and non-A non-B hepatitis by
chloroform. Infect Immun, 41, 816-821.
Fields HA, Mc Caustland KA, Bradley DW et al. (1982)
Enterically Non-A, Non-B virus : physical, chemical
and immunological properties. J Immunol Methods, 51,
149-157.
Fletcher ML, Trowell JM, Craske J, Pawen K,
Rizza CR. (1983). Non-A non-B hepatitis after
transfusion of factor VIII in unfrequently treated
patients. Br Med J, 287, 1754-1757.
Francis DP, Hadler SC, Prendergast TJ. (1984).
Occurence of hepatitis A, Band non-A non-B in the
united states.CDC sentinal country hepatitis study.
Amer J Med, 76, 69-74.
Guesdon JL, Ternynck T, Avrameas. (1979). In
Histochem Cytochem. Technics Handbook.
Gupta DN, Smetana HF. (1957). The histopathology
of viral hepatitis as seen in Dehli epidemic
(1955-56). Indian Journal of Medical Research, 1,
145-151.
Gust ID, Purcell RH. (1987). Report of a workshop
waterborne non-A non-B hepatitis. J. infect Dis,
156, 630-635.
Gust ID, Purcell RH. (1987). Report of a workshop :
Waterborne non-A non-B hepatitis from the virus
laboratory, Fairfield Hospital, Fairfield Victoria,
Australia and the National Institute of Allergy and
Infectious Diseases, National institute of Health,
Bethesda, Maryland.
116

38
39
40
41
42
43
44
Hillis A, Shrestha SM, Saha NK. (1973). An epidemic
of infectious hepatitis in the Kathmandu Valley.
Journal of Nepal Medical Association, 11, 145-151.
Hollinger FB, Mosley JW, Szmuness W et al. (1980).
Transfusion transmitted viruses study : experimental
evidence of two non-A non-B hepatitis agents. J
Infect Dis, 142, 400-407.
Hollinger FB, Khan et al. (1983). NANB hepatitis
following blood transfusion risk factors associated
with donor characterization. Viral Hepatitis,
International Symposium, Edited by Szumess, Alter et
al., Franklin Institute Press, Philadelphia, 361- ~
Hopf U, Moller B, Kuther 0 et al. (1990). Long-term
follow-up of post transfusion and sporadic chronic
hepatitis non-A, non-B and frequency of circulating
antibodies to hepatitis C virus (HCV)
J Hepatol, 10, 69-76.
Horowitz MS, Horowitz B, Rooks C, Hilgartner MW.
(1988). Virus safety solvent/detergent~treated
antihaemophilic factor concentrate. Lancet, 2,
186-189.
Ikeda Y. et al. (1990) Correlation anticorps anti
SOD/anticorps anti-HCV, d'après une étude sur 19 sujets
anti-HCV positifs. Lancet, 335, 1345-1346.
Janot C, Couroucé AM, Maniez M. (1989). Antibodies
to hepatitis C virus in French blood donors. Lancet,
2, 796-797.
117

45
46
. 47
48
49
50
51
52
Joshi YK, Babu S, Sarin S, Tandon BN et al. (1985).
Immunoprophylaxis of epidemic non-A non-B hepatitis.
Indian J Med Res, 81, 18-19.
Kane MA, Bradley DW, Shrestha et al. (1984).
Epidemie non-A non-B hepatitis in Nepal : recovery
of a possible etiologic agent and transmission
studies. J. Amer Med Ass, 252, 3140-3145.
Kaneko S. et al. (1990). Variabilité génétique du
virus de l'hépatite C. Lancet, 1, 976.
Khuroo MS (1980). Study of an epidemic of non-A non-B
hepatitis : possibility of another human hepatitis
virus different from post-transfusion non-A non-B
type. Am J Med, 68, 818-824.
Khuroo MS, Duermeyer W, Zargar et al. (1983). Acute
sporadic non-A non-B hepatitis in India. Am J
Epidemiology, 118, 360-364.
Koff RS, Galambos JJ. (1987). Viral hepatitis In :
Schiff L, Schiff ER JB ed. Diseases of the Liver.
Philadelphia Lippincott Company: 457-581.
Krawczynski C. (1989). Immunohistochemical
identification of hepatitis C associated antigen
in liver. International Symposium on non-A non-B
hepatitis and blood borne infectious diseases,
Tokyo, Elsevier (à paraître).
Kuhn LP, Seidl S, Stangel W et al. (1989). Antibody
to hepatitis C virus in German blood donors. Lancet,
2, 324.
118

53
54
55
56
t 57
58
59
Kuo G, Choo QL, Alter HJ, Gitnick AG, Redeker AG,
Purcell RH, Miyamura T, Dienstag Alter MJ, stevens
CE, Tegtmever GE, Bonino F, Colombo M, Lee WS, Kuo
C, Berger K, Shus~er JR, Overby LR, Bradley DW,
Houghton M. (1989). An assay for circulating
antibodies to a major etiologic virus of human
non-A non-B hepatitis. Science, 244, 362-363.
Mc Farlane IG. et al. (1990). Hépatites chroniques
actives auto-immunes faux positifs des tests HCV
Elisa? . Lancet, 335, 754-757.
Miller RH, Purcell RH. (1990). HCV and virus like.
Proc. Natl. Acad. Sciences, 87, 2057-2061.
Morgan C. et al. (1990). Attitude don-donneur
arbre décisionnel. Lancet, l, 921.
z.:--------- ------Morbidity and mortality weekly report. Centers of
Disease Control. (1987). Enterically transmitted
non-A non-B hepatitis East Africa. US Department~Health and Human Services/Public Health Service,
36/n D 10, 241-244.
Myint H, Soe MM, Khin T, Myint Tin KM. (1985).
A clinical and epidemiological study of an epidemic
non-A non-B hepatitis in Rangoon. Am J Tropical Med
Hyg, 345, 1183-1188.
Nouasria B, Trepo C, Larouze B et al.(1987). Non-A
non-B acute hepatitis in Eastern Aigerian adults.
Transactions of the Royal Society of Tropical
Medicine and Hygiene, 78, 137-138.
119

60
61
62
63
64
65
66
67
Pavri KM, BNiphadkar et al. (1972). Retrospective
studies on Australia antigen in sera collected
during the epidemic of viral hepatitis at Delhi
in 1956. india J Med Res, 60, 1575-1578.
Pillot J, Sharma MD, Lazizi Y, Budkowska A, Dauguet
C, Galimand M, Sarthou JL (1987). Immunological
characterization of a viral agent involved epidemic
and sporadic non-A non-B hepatitis. Ann Inst
Pasteur/Virol, 138, 145-158.
pillot J (1990). Données récentes sur les virus des
hépatites non-A non-B. Qui sont-ils ? Combien
sont-ils ? Ann Inst Pasteur/Actualités, 83-89.
Prince, Brotman AM, Grady et al. (1989). Long
incubation post-transfusion hepatitis without
serological evidence of exposure to hepatitis B
virus. Lancet, 2, 241-246.
Purcell RH, Ticehurst JR. (1988). Enterically
transmitted non-A non-B hepatitis. Epidemiology
and clinical charateristics. Zuckerman AJ (ed) :
"Viral Hepatitis and liver Disease". New-York:
Alan R, liss, inc, 131-137.
Ramalingaswani V, Purcell RH. (1988). Waterborne
Non-A, Non-B, hepatitis. Lancet, 1, 571-573.
Reesink HW, Van Der Poel CL. (1989). Blood
transfusion and hepatitis : still a threat?
Blut, 58, 1-6.
Reyes GR, Purdy MA, Kim JP et al. (1989).
Isolation of a cDNA from the virus responsible for
enterically transmitted non-A, non-B hepatitis.
Science, 247, 1335-1399.
120

68 Rosenbaum J, Carneiro B, Dhumeaux D, Trepo C.
(1984). Hépatites virales Non-A, Non-B.
Gastroenterol Clin Biol, 8, 273-287.
69 Sarthou JL, Budkowska A, Sharma MD, Lhuillier M,
pillot J. (1986). Characterization of an antigen
antibody system associated with epidemic non-A non-B
hepatitis in West Africa and experimental
transmission of an infectious agent to primates.
Ann Inst Pasteur/Virol., 137E, 225-232.
70 Smuness W, Alter HJ, Maynard JE. (1981). Viral
hepatitis. International Symposium, Philadelphia,
Franklin Institute Press, 305-317.
71 Sreenvasan MA, Banerjee et al. (1978).
Epidemiological investigations of an oubreak of
infectious hepatitis in Ahmedabad City during 1975
1976. Indian J Med Res, 67, 197-206.
72 Stevens CE. et al. (1990). A preliminary study in
volunteer blood donors. In New York Blood Center,
JAMA, 263, 49-53.
73 Tabor E., Gerety RJ, Drucker JA, Sef et al. (1978).
Transmission of non-A non-B hepatitis from man to
chimpanzee. Lancet, 2, 463-466.
74 Tabor E, April M, Seef LB, Gerety RJ. (1979).
Acquired immunity to human non-A non-B hepatitis.
Cross-challenge of chimpanzees with three infectious
human sera. J Inf Dis, 140, 789-793.
75 Tabor E, Gerety ·RJ. (1980). Inactivation of an agent
of non-A non-B hepatitis by formaI in. J inf Dis,
142, 767-770.
121

76
77
78
79
80
81
82
83
Tabor E (1981). Development and application to the
chimpanzee animal model for human NANB hepatitis.
Edited by Gerety, Academie Press, New-York,
189-206.
Tabor E, Snoy P, Jackson DR et al. (1984).
Additional evidence for more than one afflut
of human non-A non-B hepatitis. Transmission and
passage studies in chimpanzees. Transfusion, 24,
224-230.
Tabor E. (1985). The three viruses of non-A non-B
hepatitis. Lancet, l, 743-745.
Tandon BN, Joshi SK, Gandhi BM et al. (1982).
An epidemic of non-A non-B hepatitis in North
India. indian J Med Res, 75, 739-744.
Tavera C, Velazquez 0, Avila C et al. (1987).
Enterically transmitted non-A non-B hepatitis
Mexico Morbidity and Mortality Weekly Report.
Atlanta. United states Department of Health,
Education and Welfare, Public Health Service,
Center for Disease Control, 36, 597-602.
Trepo C. (1990). Identification du virus de
l'hépatite C (VHC) : un progrès décisif pour la
santé publique. Med Sciences, 6, 98-107.
Van Der Poel et al. (1989). Anti-hepatitis C
antibodies and non-A non-B post-transfusion
hepatitis in the Netherlands. Lancet, 2, 297
298.
Viswanathan R. (1957). Infectious hepatitis in Delhi
(1955-1956). A critical epidemiological study.
Indian Journal of Medical Research, 34, 1-29.
122

84
85
86
87
88
89
Wands JR, Lieberman HM, Muchmore E, Isselbacher K,
Shafritz DA. (1982). Detection and transmission in
Chimpanzees of hepatitis B virus-related agents
formerly designated Non-A, Non-B hepatitis.
Proc Natl Acad science, 79, 7552-7556.
Weiner AJ, Kuo G, Bradley DW et al. (1990).
Detection of hepatitis C viral sequences in non-A,
non-B hepatitis. Lancet, 1, 355.
Wong OC, Purcell RH, Streentvasan MA, Prasad SR,
Pavri M. (1980). Epidemie and endemic hepatitis in
India Evidence for a non-A non-B hepatitis virus
etiology. Lancet, 1980, 2, 876-879.
Wreghitt TG, et al. (1990). False positive reactions'
with ortho ELISA : immunological and Chemical
considerations. Lancet, 1, 789.
Yamauchi M, Nakayima H, Kinura K et al. (1983).
An epidemic of non-A non-B hepatitis in Japan. Am
J Gastroenterol, 78, 652-655.
Yoshizawa H, Ithoh, Iwahin. (1983). Demonstration of
two different types of non-A non-B hepatitis by
reinfection and cross-challenge studies in
chimpanzees. Gastroenterology, 81, 197-213.

AIlTIlTEXE
l - PREPARATION DES IgG DE LAPIN ANTI Fc (t) HUMAIN
Elle se fait en deux étapes :- Préparation du fragment Fc (gamma) humain par la méthode dePORTER qui est appliquée à des IgG humaines purifiées sur OEAEcellulose.
A 1 g d' IgG dissoute dans 100 ml de tampon phosphatemonopotassique disodique 0,1 M pH 7,2, on ajoute 44,6 mg d'acideéthylène diamine tétracétique (EOTA), puis 96,6 mg de L-cystéineet 10 mg de papaïne cristallisée (Merck). Après incubation de 3heures· à 37 oC, on dialyse contre du tampon phosphatemonopotassique disodique 0,1 M pH 7,2. On filtre sur SéphadexG100 dans ce même tampon pour éliminer les IgG non digérées quisortent dans le premier pic exclu et les produits de dégradationqui sortent dans un troisième pic. Le deuxième pic qui contientles fragments d'IgG est concentré et dialysé en tampon phosphatemonopotassique disodique 0,005 M pH 7 , 2. A ce stade, des cristauxde Fc (gamma) apparaissent dans le sac de dialyse. on lesrecueille par centrifugation et les purifie par sixcristallisations successives réalisées par dialyse en eaudistillée froide, avec redissolution en tampon acétate 0,1 M pH5,4. Le contenu du deuxième pic dialysé est ensuitefractionné sur OEAE 52 équilibrée en tampon phosphate 0,005 MpH 7,2. Les fragments Fab ne sont pas retenus dans cesconditions. Les fragments Fc (gamma) sont élués en tamponphosphate 0,3 M pH 7,2. A ce stade, les fractions ne sont pasgénéralement rigoureusement pures, ce qui rend nécessaire unedernière purification. Pour ce faire, les fragments Fc (gamma)sont passés sur un immunoadsorbant d' IgG de mouton anti-Fab,pour éliminer les impuretés résiduelles.
La caractérisation immunologique des produits est réaliséepar immunodiffusion double en gel vis-à-vis d'anti-sérum humaintotal, d'anti-IgG, d'anti-Fab et d'anti-Fc (gamma). La puretébiochimique est établie et le contrôle des masses moléculairesest réalisé par électrophorèse en gel de polyacrylamide sos.
- Immunisation du lapin
Elle se fait par injection intramusculaire, toutes les 3semaines, de 100 mg de protéines dans 0,2 ml d'eau physiogiqueen présence d'un volurne d'adj uvant complet de Freund. L' évolutiondu taux des anticorps et leur spécificité est suivie parimmunodiffusion en gel et hémagglutination passive, et lessaignées sont réalisées au moment jugé opportun, en général après3 ou 4 injections de l'antigène.
//

II - PREPARATION DES REACTIFS ET TAMPONS UTILISES
A - T A M P 0 N S
1 - Tampon Tris (hydroxymethyl) aminométhane :
A : 0,2 M solution de Tris hydroxyméthyl aminométhane(24.2 g dans 1000 ml)
B : 0.2 M HeL50 ml de A + x ml de B, compléter à 200 ml.
x
5.08.1
12.216.521.926.832.538.441.444.2
2 - Tampon bicarbonate-carbonate
pH
9.08.88.68.48.28.07.87.67.47.2
A 0.2 M solution de carbonate anhydre de sodium(21.2 g dans 1000 ml).
B 0.2 M solution bicarbonate de sodium (16.8 g dans1000 ml).x ml de A + y ml de B, compléter à 200 ml.
x
4.07.59.5
13.016.019.522.025.027.530.033.035.538.540.542.545.0
y
46.042.540.537.034.030.528.025.022.520.017.014.511. 59.57.55.0
pH
9.29.39.49.59.69.79.89.9
10.010.110.210.310.410.510.610.7
G.E. Delory et E. J. King, J. Biochem., 1945, 39, 245
11
1
1
/

3 - Tampon phosphate:
A 0.2 M solution phosphate monobasique de sodium(27.8 g dans 1000 ml).
B 0.2 M solution de phosphate dibasique de sodium(53.65 g de Na2HP047 H20 ou 71.7 g de NaHP0412 H20 dans 1000 ml).
x ml de A + y ml de B, compléter à 200 ml.
x
93.592.090.087.785.081.577.573.568.562.556.551.0
y
6.58.0
10.012.315.018.522.526.531. 537.543.549.0
pH
5.75.85.96.06.16.26.36.46.56.66.76.8
x
45.039.033.028.023.019.016.013.010.58.57.05.3
y
55.061.067.072.077.081. 084.087.090.591.593.094.7
pH
6.97.07.17.27.37.47.57.67.77.87.98.0
S.P.L. Sorensen, Biochem. Z., 1909, 21, 1311909, 22, 352
4 - Tampon acétate:
A 0.2 M solution diacide (11.55 ml dans 100 ml)B 0.2 M solution dlacétate de sodium (16.4 de C2H302Na
ou 27.2 g de C2H302Na 3H20 dans 1000 ml).x ml de A + y ml de B, compléter à 100 ml.
x y pHacide acétique acétate de Na
46.3 3.7 3.644.0 6.0 3.841.0 9.0 4.036.8 13.2 4.230.5 19.5 4.425.5 24.5 4.620.0 30.0 4.814.8 35.2 5.010.5 39.5 5.28.8 41.2 5.44.8 45.2 5.6
G.S. walpole, J. Chem. Soc. , 1914, 105, 2501

3 - Tampon phosphate ..A 0.2 M solution phosphate monobasique de sodium
(27.8 g dans 1000 ml).B 0.2 M solution de phosphate dibasique de sodium
(53.65 g de Na2HP047 H20 ou 71. 7 g de NaHP0412 H20 dans 1000 ml).
x ml de A + y ml de B, compléter à 200 ml.
x y pH x y pH
93.5 6.5 5.7 45.0 55.0 6.992.0 8.0 5.8 39.0 61. 0 7.090.0 10.0 5.9 33.0 67.0 7.187.7 12.3 6.0 28.0 72.0 7.285.0 15.0 6.1 23.0 77.0 7.381. 5 18.5 6.2 19.0 81. 0 7.477.5 22.5 6.3 16.0 84.0 7.573.5 26.5 6.4 13.0 87.0 7.668.5 31. 5 6.5 10.5 90.5 7.762.5 37.5 6.6 8.5 91. 5 7.856.5 43.5 6.7 7.0 93.0 7.951.0 49.0 6.8 5.3 94.7 8.0
S.P.L. Sorensen, Biochem. Z. , 1909, 21, 1311909, 22, 352
4 - Tampon acétate ..A 0.2 M solution d'acide (11.55 ml dans 100 ml)B 0.2 M solution d'acétate de sodium (16.4 de C2H302Na
ou 27.2 g de C2H302Na 3H20 dans 1000 ml).x ml de A + y ml de B, compléter à 100 ml.
x y pHacide acétique acétate de Na
46.3 3.7 3.644.0 6.0 3.841.0 9.0 4.036.8 13.2 4.230.5 19.5 4.425.5 24.5 4.620.0 30.0 4.814.8 35.2 5.010.5 39.5 5.28.8 41. 2 5.44.8 45.2 5.6
G.S. Walpole, J. Chem. Soc. , 1914, 105, 2501

,. '- .
B - REACTIFS
1 - PM2
Tampon phosphate 0,1 M PH 7,4Titriplex : 2 mMMgS04 1 mMMnS04 : 0,2 mM
2 - Réactif phospho-tunqstique
Acide ortho-phosphoriqueTungstate de sodium
0,01 N0,5 mM