Embed Size (px)
Citation preview

21/02/2018
1
Partie 2 : Physiologie neuronale
Plan :
1. Potentiels membranaires
2. Synapse
3. Neurotransmetteurs et récepteurs
4. Intégration nerveuse
1. Potentiels membranaires1. Principes fondamentaux d’électricité
2. Potentiel de repos de la membrane
3. Fonction de signalisation
1. Potentiels gradués
2. Potentiels d�action

21/02/2018
2
1. Potentiels membranaires1. Principes fondamentaux d’électricité
2. Potentiel de repos de la membrane
3. Fonction de signalisation
1. Potentiels gradués
2. Potentiels d�action
Principes fondamentaux
Membrane perméable
+ -
+ -
+ -
+ -
=0 V =0 V
- Equilibre électrique
- Déséquilibre chimique = gradientde concentration
+ - =NaCl
Situation initiale :

21/02/2018
3
Principes fondamentaux
Membrane perméable
+
-
+-
+
-
+ -
=0 V =0 V
+ - =NaCl
- Equilibre électrique
- Equilibre chimique
Situation finale :
Principes fondamentaux
Membrane imperméable
+ -
+ -
+ -
+ -
=0 V =0 V
+ - =NaCl
- Equilibre électrique
- Déséquilibre chimique = gradientde concentration
Situation initiale :
- Equilibre électrique
- Déséquilibre chimique
Situation finale :

21/02/2018
4
Principes fondamentaux
Membrane semi-perméable
+ -
+ -
+ -
+ -
Charge=0
Charge=0
Déséquilibre chimique = gradientqui pousse les ions - du compartimentde gauche vers celui de droite
Principes fondamentaux
Membrane semi-perméable
+-
+
-
+ -
+-
Charge=+3
Charge=-3
Déséquilibre électrique = gradientqui pousse les ions - du compartimentde droite vers celui de gauche
21/02/2018
5
Principes fondamentaux
Membrane semi-perméable
+
-
+-
+-
+
-
Charge=+2
Charge=-2
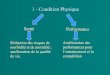
Nouvel état d’équilibre résultant du gradient chimique et du gradient électrique =
Équilibre électro-chimique (avec mouvement d’eau possible)
Notion de potentiel d’équilibre (les deux forces se compensent)
Principes fondamentaux
Gradient électrochimique
Gradient chimique
A A A A
ABB B B
B
A Gradient électrique
+ + +
- -
-
-
+
+

21/02/2018
6
Principes fondamentaux
Ainsi, lorsque la concentration de cet ion sera dix fois plus importante à l'intérieurde la cellule qu'à l'extérieur, son potentiel d'équilibre (différence de potentielengendrée par l'ion en question) sera de – 58 mV à 20°C et de – 61 mV à 37°Cpuisque le logarithme de 10 est égal à 1. Si maintenant, on applique cette formuleaux principaux ions en présence au niveau d'un neurone de mammifère, ons'aperçoit qu'à 20 °C le potentiel d'équilibre est :
y pour les ions sodium de + 58 mV ,
y pour les ions potassium de – 84 mV,
y pour les ions calcium de + 116 mV,
y pour les ions chlore de – 58 mV.
Comme la membrane est perméable à tous ces ions (les anions, à l'exception duchlore, ne traversent pas ou peu la membrane), le potentiel de membrane necorrespond à aucune de ces valeurs mais résulte de la combinaison des différentsflux ioniques générés par leurs gradients de concentration et leurs gradientsélectriques.
On obtient ainsi :
y un flux net entrant de sodium du à la somme des gradients ;
y un flux net sortant de potassium du à la différence des gradients ;
y un flux net entrant de calcium du à la somme des gradients ;
y un flux nul de chlore puisque les deux gradients s'annulent.
Or, il se trouve qu'au repos, très peu de canaux au sodium et au calcium sontouverts. Il en résulte que le potentiel de repos est principalement du à unesortie de potassium de la cellule, ce qui a pour effet de négativer le milieuintracellulaire aux abords de la membrane.
Il reste maintenant à expliquer pourquoi le potentiel est stable au cours du temps.En effet, si le potassium sort en permanence de la cellule, il finira par annuler songradient de concentration. Il faut donc concevoir un mécanisme faisant entrer dupotassium à l'intérieur de la cellule contre son gradient de manière à maintenir sonéquilibre ionique.
Un tel mécanisme existe : il s'agit de la pompe membranaire à activité ATPasiquedécouverte en 1955, la pompe Na-K qui transporte activement, contre leursgradients, trois ions Na+ du milieu intracellulaire vers le milieu extracellulaire enéchange de deux ions K+ du milieu extracellulaire vers le milieu intracellulaire.
Propriétés électriques du neurone
CUEEP-Université Lille 1 Sciences et Technologies25
3. Potentiels de membrane1. Principes fondamentaux d�électricité
2. Potentiel de repos de la membrane
3. Fonction de signalisation
1. Potentiels gradués
2. Potentiels d�action
21/02/2018
7
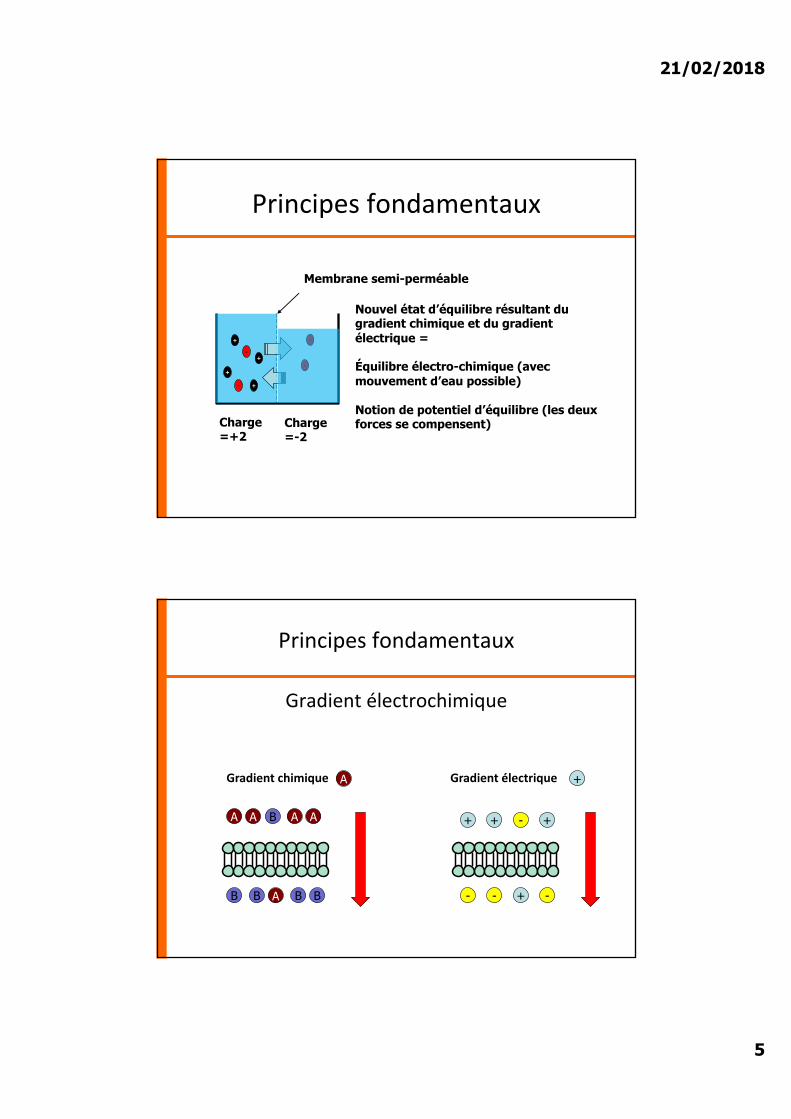
3.2. Potentiel de repos de la membraneIons Intracellulaire
(mmol/l)Extracellulaire
(mmol/l)
K+ 150 5
Na+ 2 140
Cl- 10 105
Anions organiques (Pr-)
65 0
Déséquilibre chimique et électrique
Production d�un potentiel de repos (mesuré en mV)
Membrane 100 % imperméable
Cellule artificielle
3.2. Potentiel de repos de la membrane
Neurone
Micro-électrodeen verre
• Différence de charge électrique entre liquide intra et extra cellulaire
• Valeur représente la différence de charge du milieu intracellulaire vs extracellulaire (fixé à 0 arbitrairement)
• En moyenne –70 mV dans les neurones
21/02/2018
8
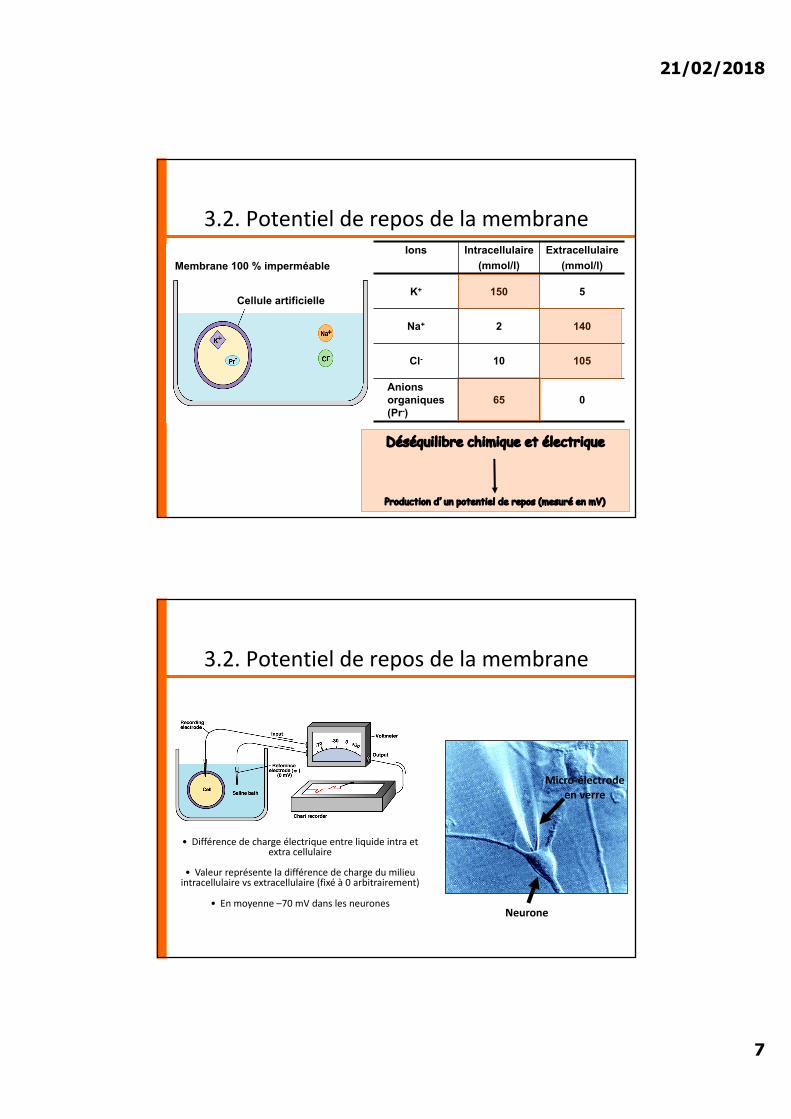
3.2. Potentiel de repos de la membrane
Quelle est l’origine de la différence de potentiel ?
3.2. Potentiel de repos de la membrane
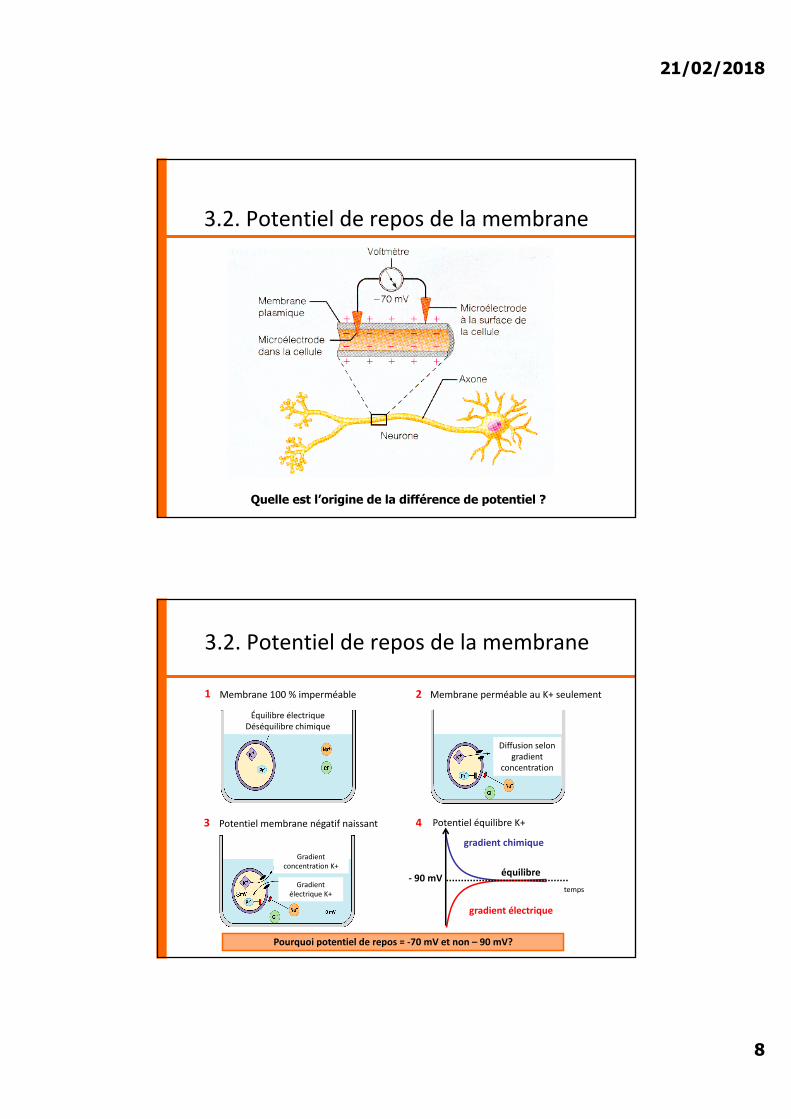
Équilibre électriqueDéséquilibre chimique
1 2
3
Membrane 100 % imperméable Membrane perméable au K+ seulement
Diffusion selon gradient
concentration
Potentiel membrane négatif naissant
gradient chimique
gradient électrique
équilibre- 90 mV
Gradient concentration K+
Gradient électrique K+
4
Pourquoi potentiel de repos = -70 mV et non – 90 mV?
Potentiel équilibre K+
temps

21/02/2018
9
3.2. Potentiel de repos de la membrane
• Potentiel d’équilibre Na+ = +60 mV
• Perméabilité membrane au Na+
est très inférieure au K+
Attention : le mouvement de Na+ sous l’influence du gradient électrique est ici sortant parce que l’intérieur de la cellule artificielle est positif. Dans une « vraie » cellule, l’intérieur étant plus électronégatif que l’extérieur, les ions Na+ ont un déplacement de l’extérieur vers l’intérieur de la cellule (voir diapo suivante)
3.2. Potentiel de repos de la membrane
Ainsi, lorsque la concentration de cet ion sera dix fois plus importante à l'intérieurde la cellule qu'à l'extérieur, son potentiel d'équilibre (différence de potentielengendrée par l'ion en question) sera de – 58 mV à 20°C et de – 61 mV à 37°Cpuisque le logarithme de 10 est égal à 1. Si maintenant, on applique cette formuleaux principaux ions en présence au niveau d'un neurone de mammifère, ons'aperçoit qu'à 20 °C le potentiel d'équilibre est :
y pour les ions sodium de + 58 mV ,
y pour les ions potassium de – 84 mV,
y pour les ions calcium de + 116 mV,
y pour les ions chlore de – 58 mV.
Comme la membrane est perméable à tous ces ions (les anions, à l'exception duchlore, ne traversent pas ou peu la membrane), le potentiel de membrane necorrespond à aucune de ces valeurs mais résulte de la combinaison des différentsflux ioniques générés par leurs gradients de concentration et leurs gradientsélectriques.
On obtient ainsi :
y un flux net entrant de sodium du à la somme des gradients ;
y un flux net sortant de potassium du à la différence des gradients ;
y un flux net entrant de calcium du à la somme des gradients ;
y un flux nul de chlore puisque les deux gradients s'annulent.
Or, il se trouve qu'au repos, très peu de canaux au sodium et au calcium sontouverts. Il en résulte que le potentiel de repos est principalement du à unesortie de potassium de la cellule, ce qui a pour effet de négativer le milieuintracellulaire aux abords de la membrane.
Il reste maintenant à expliquer pourquoi le potentiel est stable au cours du temps.En effet, si le potassium sort en permanence de la cellule, il finira par annuler songradient de concentration. Il faut donc concevoir un mécanisme faisant entrer dupotassium à l'intérieur de la cellule contre son gradient de manière à maintenir sonéquilibre ionique.
Un tel mécanisme existe : il s'agit de la pompe membranaire à activité ATPasiquedécouverte en 1955, la pompe Na-K qui transporte activement, contre leursgradients, trois ions Na+ du milieu intracellulaire vers le milieu extracellulaire enéchange de deux ions K+ du milieu extracellulaire vers le milieu intracellulaire.
Propriétés électriques du neurone
CUEEP-Université Lille 1 Sciences et Technologies25
Au repos, très peu de canaux Ca2+ et Na+ ouverts. Le potentiel de membrane se rapproche donc du potentiel d’équilibre du potassium
21/02/2018
10
3.2. Potentiel de repos de la membrane
Ainsi, lorsque la concentration de cet ion sera dix fois plus importante à l'intérieurde la cellule qu'à l'extérieur, son potentiel d'équilibre (différence de potentielengendrée par l'ion en question) sera de – 58 mV à 20°C et de – 61 mV à 37°Cpuisque le logarithme de 10 est égal à 1. Si maintenant, on applique cette formuleaux principaux ions en présence au niveau d'un neurone de mammifère, ons'aperçoit qu'à 20 °C le potentiel d'équilibre est :
y pour les ions sodium de + 58 mV ,
y pour les ions potassium de – 84 mV,
y pour les ions calcium de + 116 mV,
y pour les ions chlore de – 58 mV.
Comme la membrane est perméable à tous ces ions (les anions, à l'exception duchlore, ne traversent pas ou peu la membrane), le potentiel de membrane necorrespond à aucune de ces valeurs mais résulte de la combinaison des différentsflux ioniques générés par leurs gradients de concentration et leurs gradientsélectriques.
On obtient ainsi :
y un flux net entrant de sodium du à la somme des gradients ;
y un flux net sortant de potassium du à la différence des gradients ;
y un flux net entrant de calcium du à la somme des gradients ;
y un flux nul de chlore puisque les deux gradients s'annulent.
Or, il se trouve qu'au repos, très peu de canaux au sodium et au calcium sontouverts. Il en résulte que le potentiel de repos est principalement du à unesortie de potassium de la cellule, ce qui a pour effet de négativer le milieuintracellulaire aux abords de la membrane.
Il reste maintenant à expliquer pourquoi le potentiel est stable au cours du temps.En effet, si le potassium sort en permanence de la cellule, il finira par annuler songradient de concentration. Il faut donc concevoir un mécanisme faisant entrer dupotassium à l'intérieur de la cellule contre son gradient de manière à maintenir sonéquilibre ionique.
Un tel mécanisme existe : il s'agit de la pompe membranaire à activité ATPasiquedécouverte en 1955, la pompe Na-K qui transporte activement, contre leursgradients, trois ions Na+ du milieu intracellulaire vers le milieu extracellulaire enéchange de deux ions K+ du milieu extracellulaire vers le milieu intracellulaire.
Propriétés électriques du neurone
CUEEP-Université Lille 1 Sciences et Technologies25
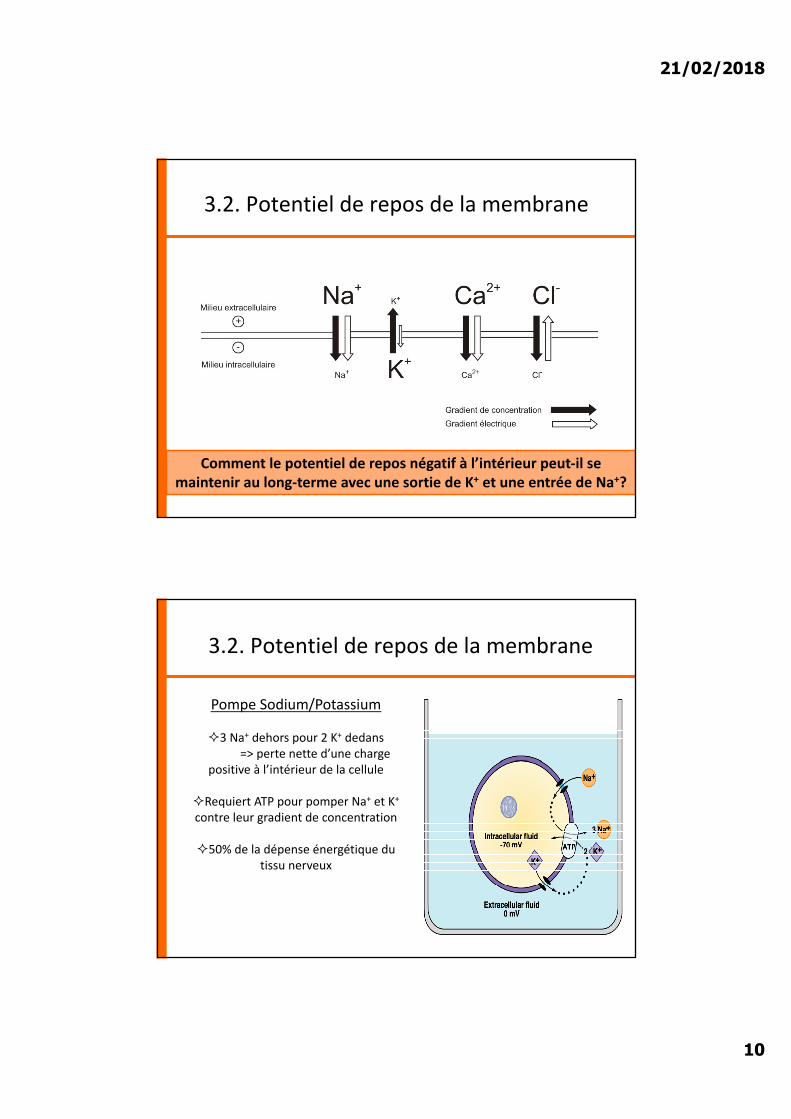
Comment le potentiel de repos négatif à l’intérieur peut-il se maintenir au long-terme avec une sortie de K+ et une entrée de Na+?
3.2. Potentiel de repos de la membrane
Pompe Sodium/Potassium
²3 Na+ dehors pour 2 K+ dedans => perte nette d’une charge
positive à l’intérieur de la cellule
²Requiert ATP pour pomper Na+ et K+
contre leur gradient de concentration
²50% de la dépense énergétique du tissu nerveux

21/02/2018
11
3.2. Potentiel de repos de la membrane
Pompe Sodium/Potassium
²Preuve expérimentale apportée par utilisation de Na+ radioactif injecté à
l’intérieur d’un neurone et qui se retrouve à l’extérieur (transport contre
son gradient de concentration)
²Blocage de la pompe par cyanure qui bloque la production d’ATP ou la
ouabaïne extraite d’un arbre qui pousse en Ethiopie qui bloque spécifiquement
la pompe
3. Potentiels de membrane1. Principes fondamentaux d�électricité
2. Potentiel de repos de la membrane
3. Fonction de signalisation
1. Potentiels gradués
2. Potentiels d�action
21/02/2018
12
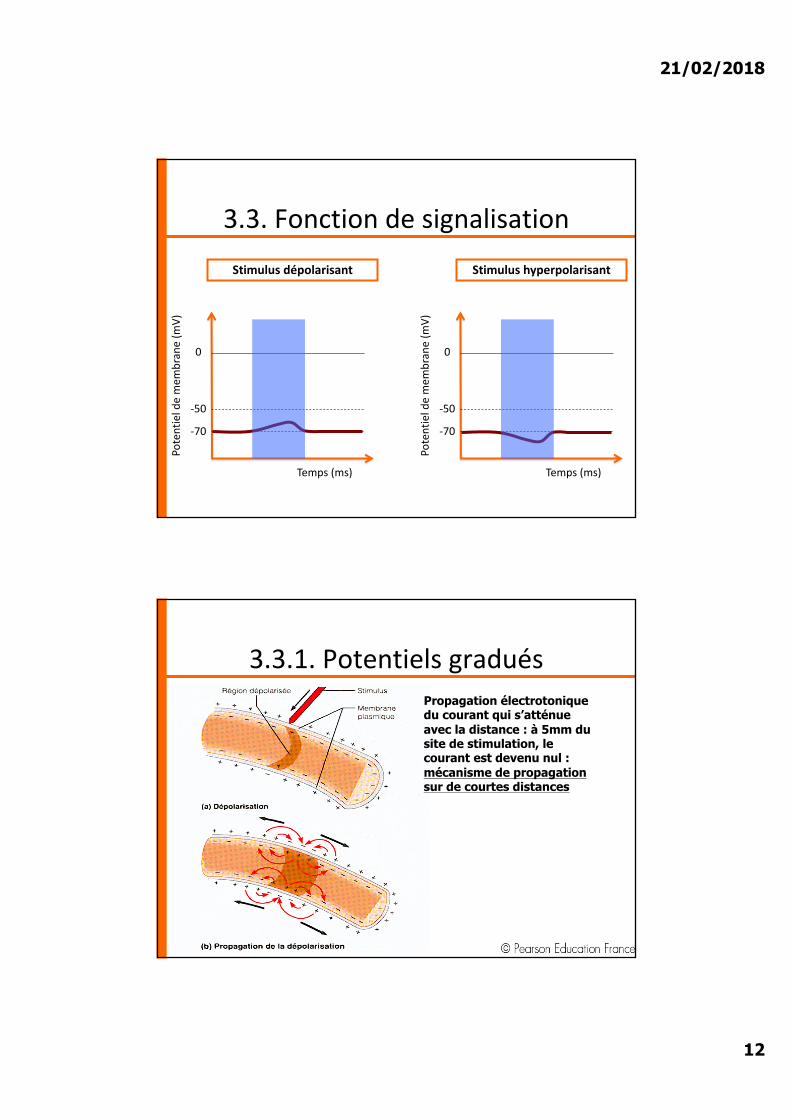
3.3. Fonction de signalisation
-70
-50
0
Temps (ms)
Pote
ntie
l de
mem
bran
e (m
V)
Stimulus dépolarisant
-70
-50
0
Temps (ms)
Pote
ntie
l de
mem
bran
e (m
V)
Stimulus hyperpolarisant
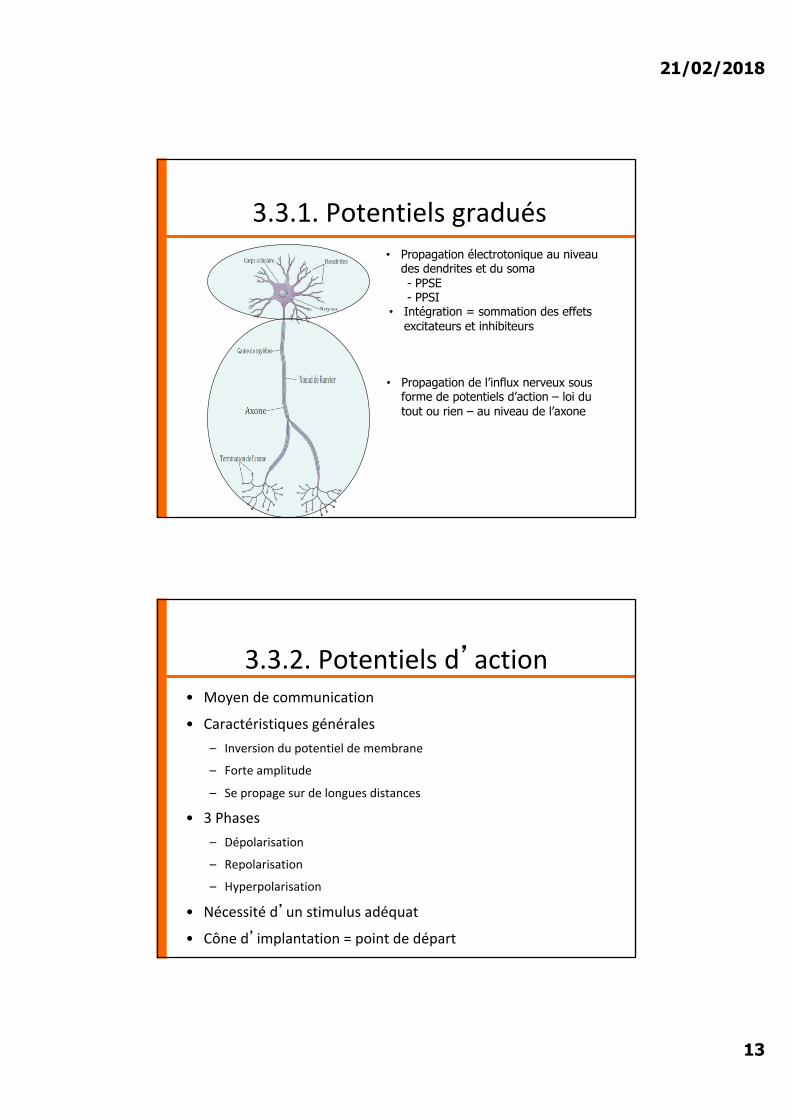
3.3.1. Potentiels graduésPropagation électrotoniquedu courant qui s’atténue avec la distance : à 5mm du site de stimulation, le courant est devenu nul : mécanisme de propagation sur de courtes distances
21/02/2018
13
• Propagation électrotonique au niveau des dendrites et du soma- PPSE- PPSI
• Intégration = sommation des effets excitateurs et inhibiteurs
3.3.1. Potentiels gradués
• Propagation de l’influx nerveux sous forme de potentiels d’action – loi du tout ou rien – au niveau de l’axone
3.3.2. Potentiels d�action• Moyen de communication• Caractéristiques générales
– Inversion du potentiel de membrane– Forte amplitude
– Se propage sur de longues distances
• 3 Phases – Dépolarisation– Repolarisation– Hyperpolarisation
• Nécessité d�un stimulus adéquat• Cône d�implantation = point de départ
21/02/2018
14
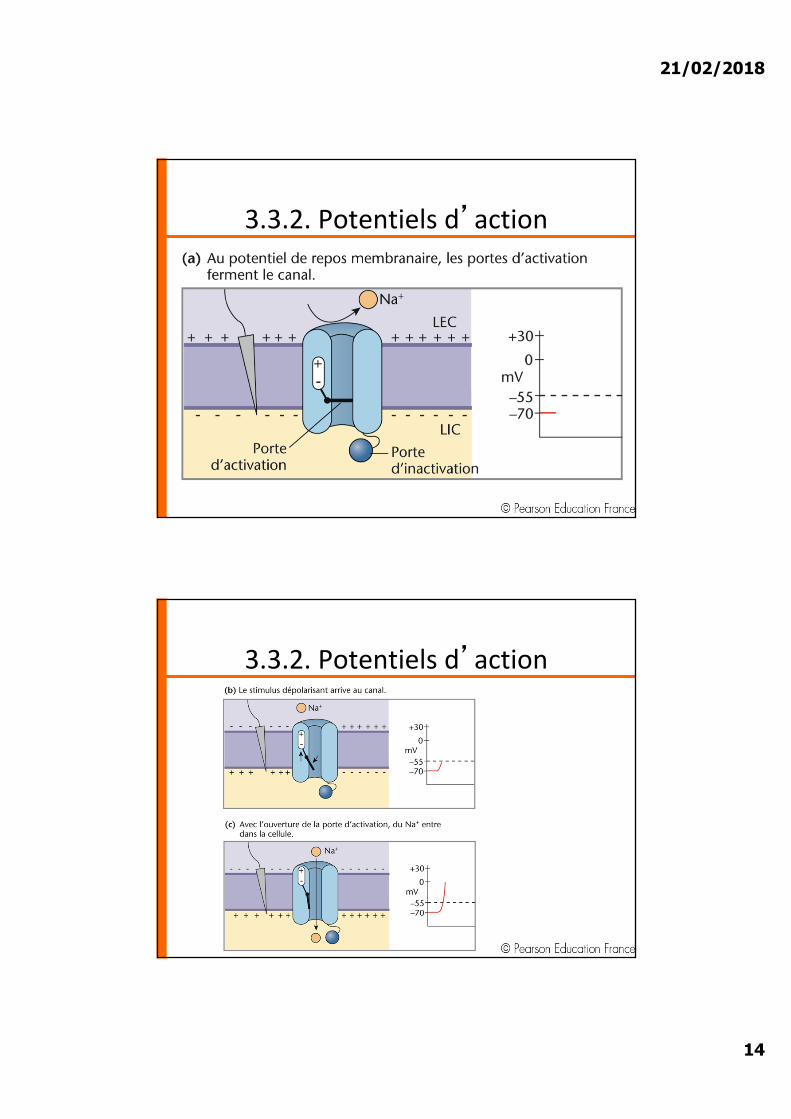
3.3.2. Potentiels d�action
3.3.2. Potentiels d�action
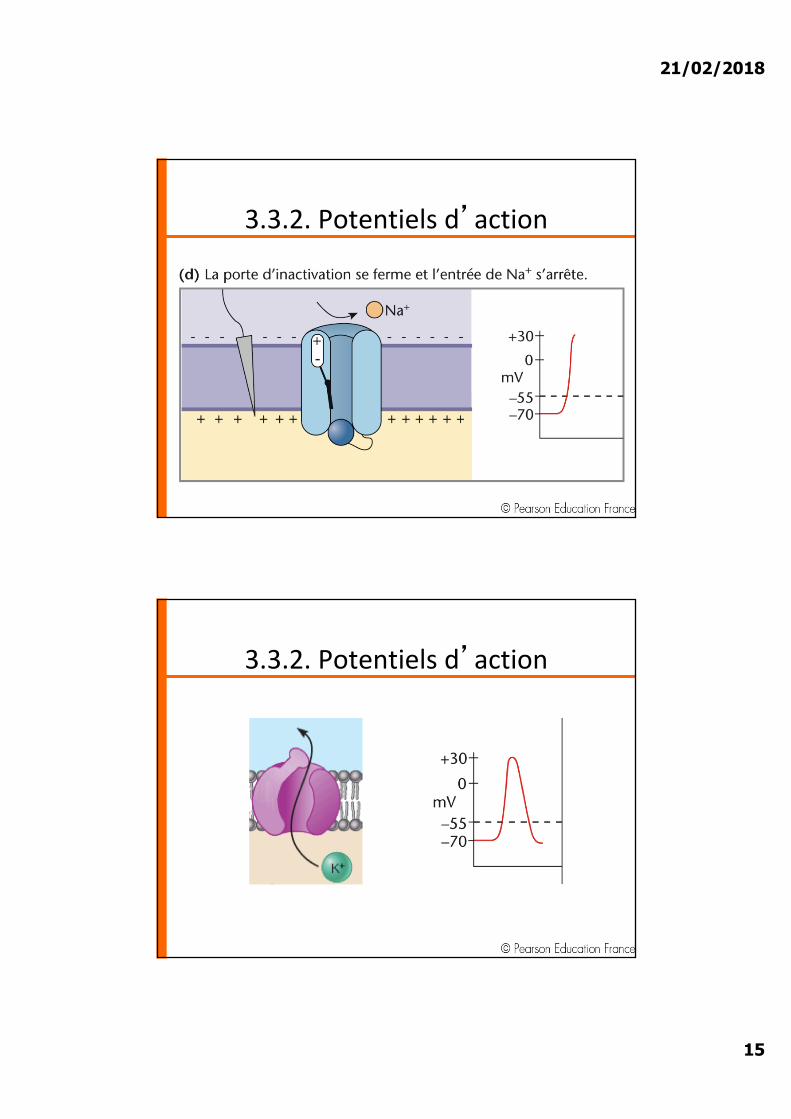
21/02/2018
15
3.3.2. Potentiels d�action
3.3.2. Potentiels d�action

21/02/2018
16
3.3.2. Potentiels d�action
3.3.2. Potentiels d�action
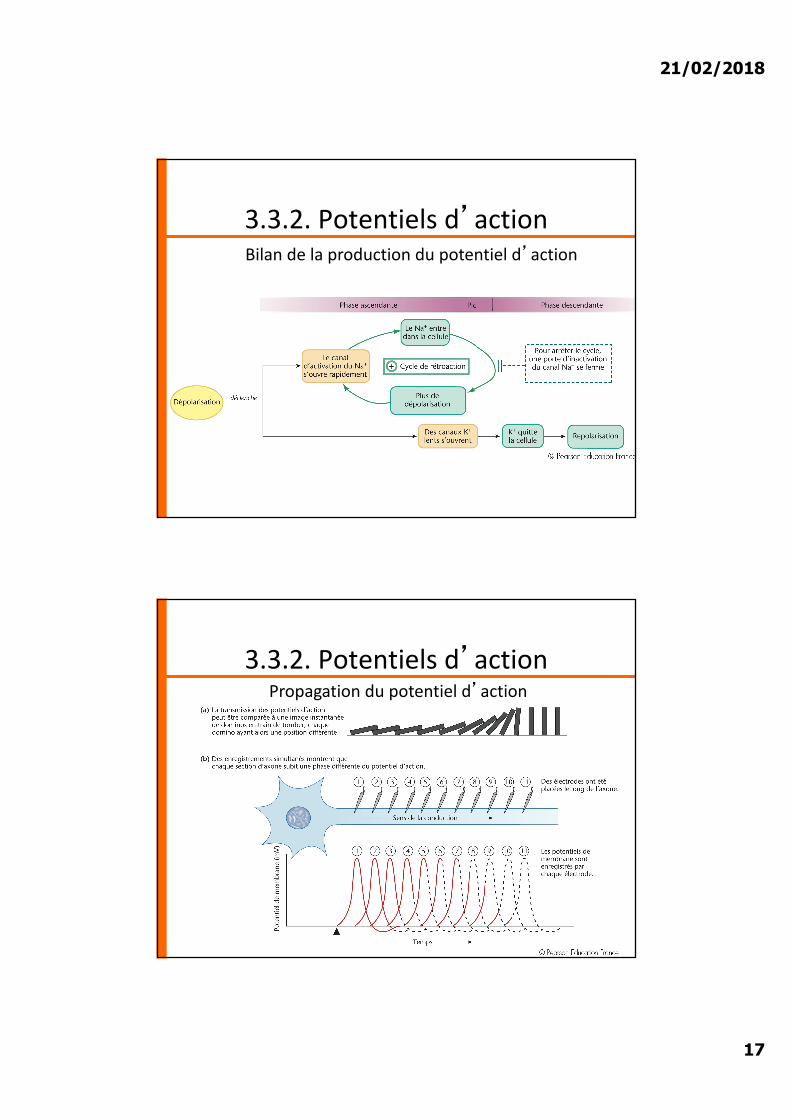
21/02/2018
17
3.3.2. Potentiels d�actionBilan de la production du potentiel d�action
3.3.2. Potentiels d�actionPropagation du potentiel d�action
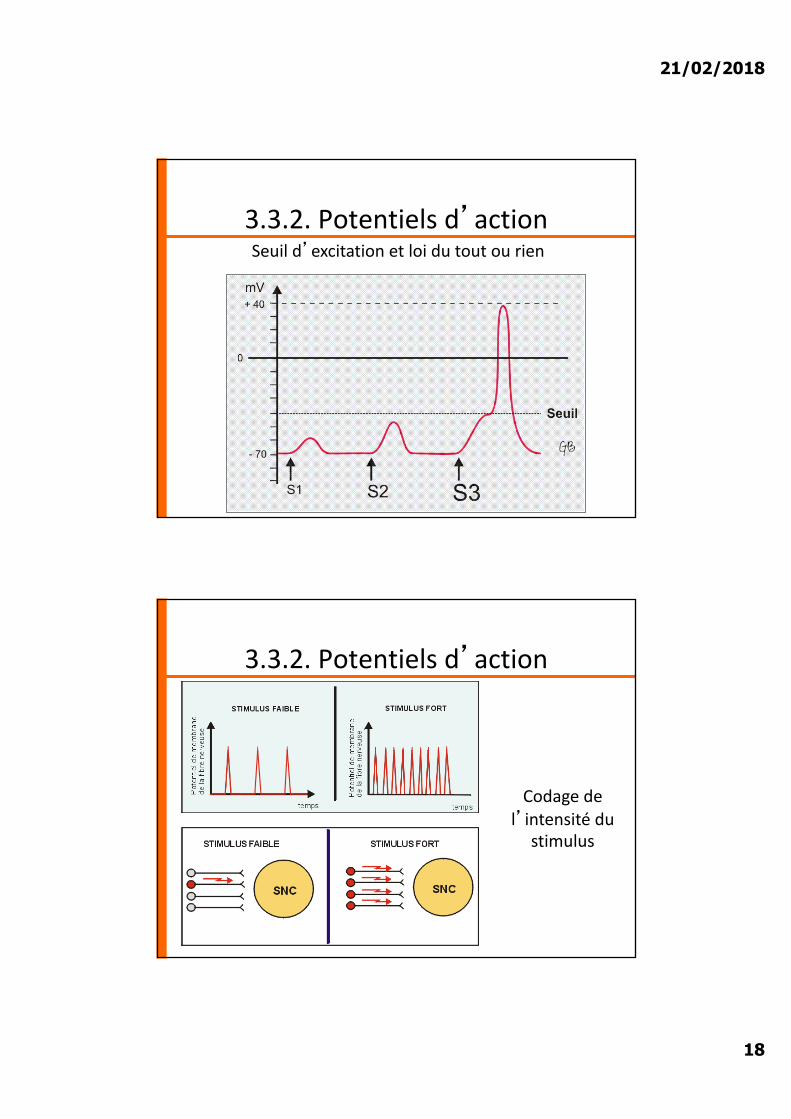
21/02/2018
18
3.3.2. Potentiels d�actionSeuil d�excitation et loi du tout ou rien
3.3.2. Potentiels d�action
Codage de l�intensité du
stimulus
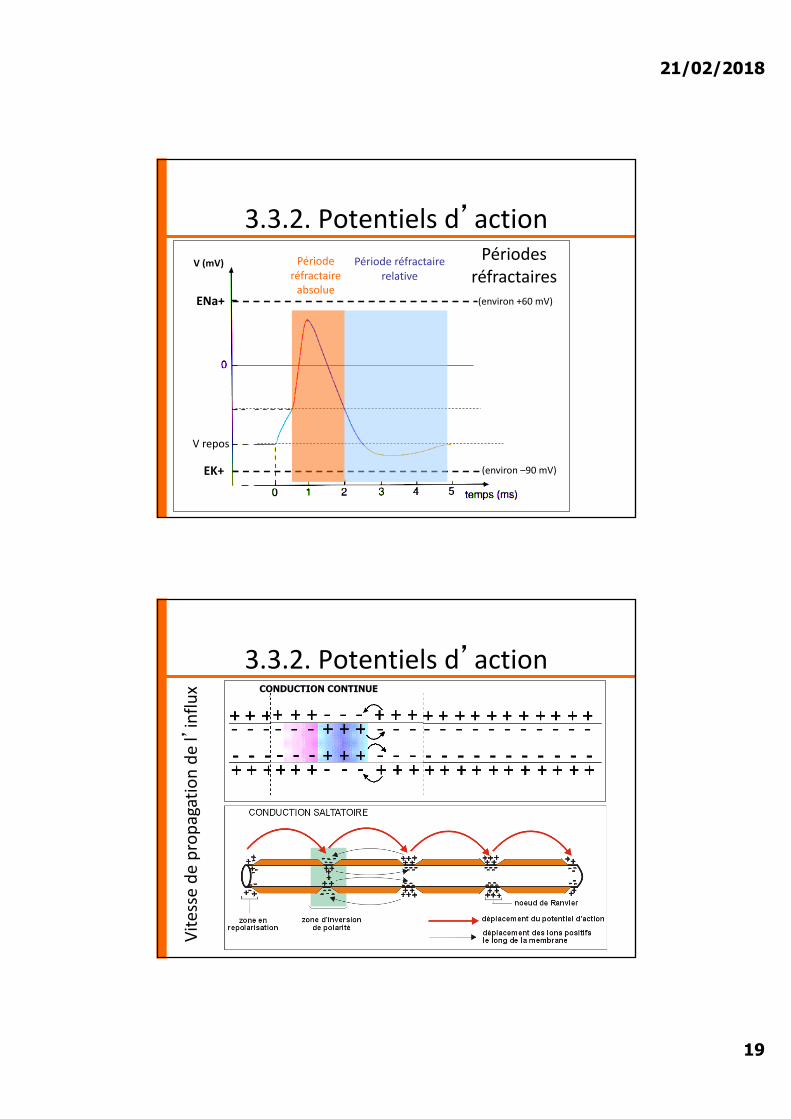
21/02/2018
19
3.3.2. Potentiels d�actionV (mV)
EK+ (environ –90 mV)
ENa+ (environ +60 mV)
V repos
Période réfractaire
absolue
Période réfractaire relative
Périodes réfractaires
3.3.2. Potentiels d�action
Vite
sse
de p
ropa
gatio
n de
l�in
flux CONDUCTION CONTINUE
21/02/2018
20
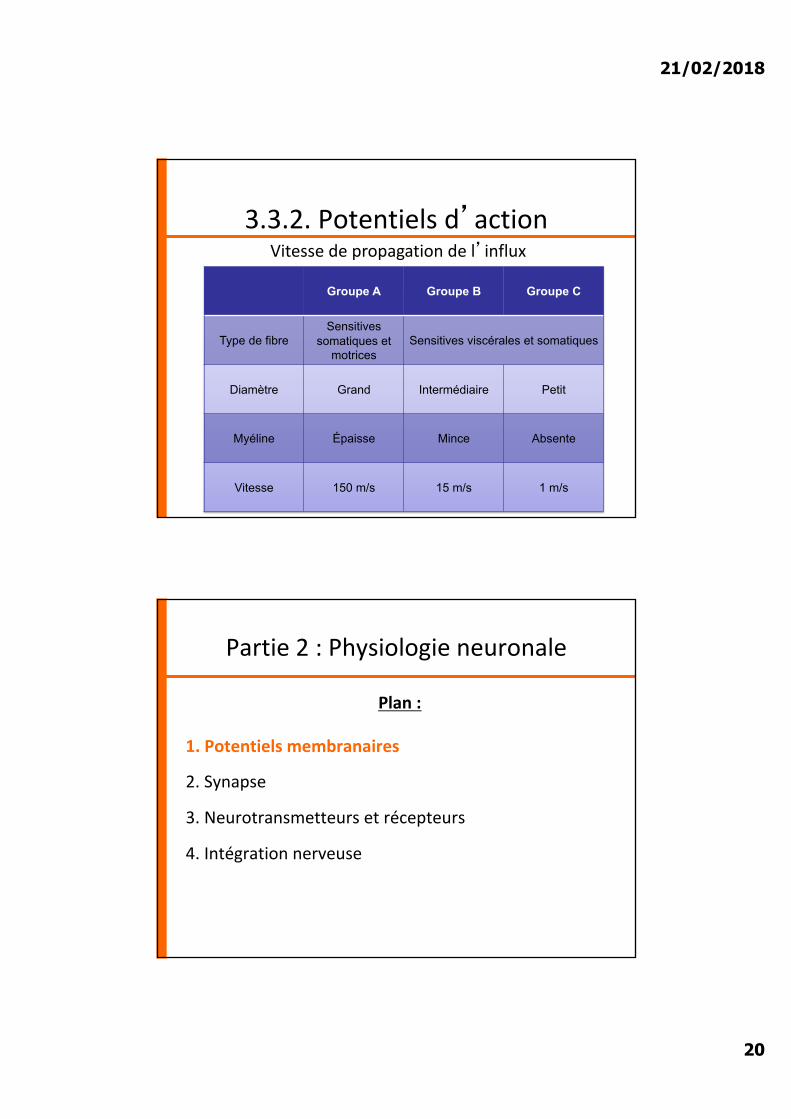
3.3.2. Potentiels d�actionVitesse de propagation de l�influx
Groupe A Groupe B Groupe C
Type de fibreSensitives
somatiques et motrices
Sensitives viscérales et somatiques
Diamètre Grand Intermédiaire Petit
Myéline Épaisse Mince Absente
Vitesse 150 m/s 15 m/s 1 m/s
Partie 2 : Physiologie neuronale
Plan :
1. Potentiels membranaires
2. Synapse
3. Neurotransmetteurs et récepteurs
4. Intégration nerveuse
21/02/2018
21
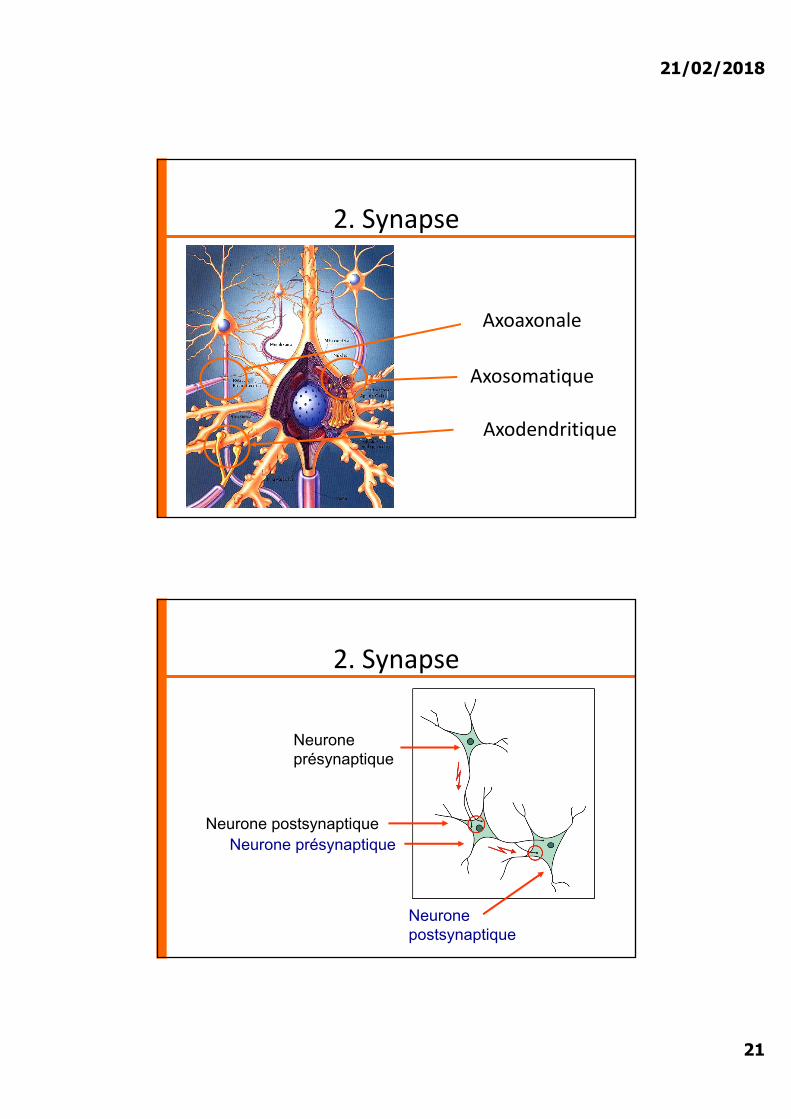
2. Synapse
Axoaxonale
Axosomatique
Axodendritique
2. Synapse
Neurone présynaptique
Neurone postsynaptiqueNeurone présynaptique
Neurone postsynaptique

21/02/2018
22
2. Synapse
1. Synapses électriques
2. Synapses chimiques
3. Potentiels postsynaptiques et intégration
synaptique
2. Synapse
1. Synapses électriques
2. Synapses chimiques
3. Potentiels postsynaptiques et intégration
synaptique
21/02/2018
23
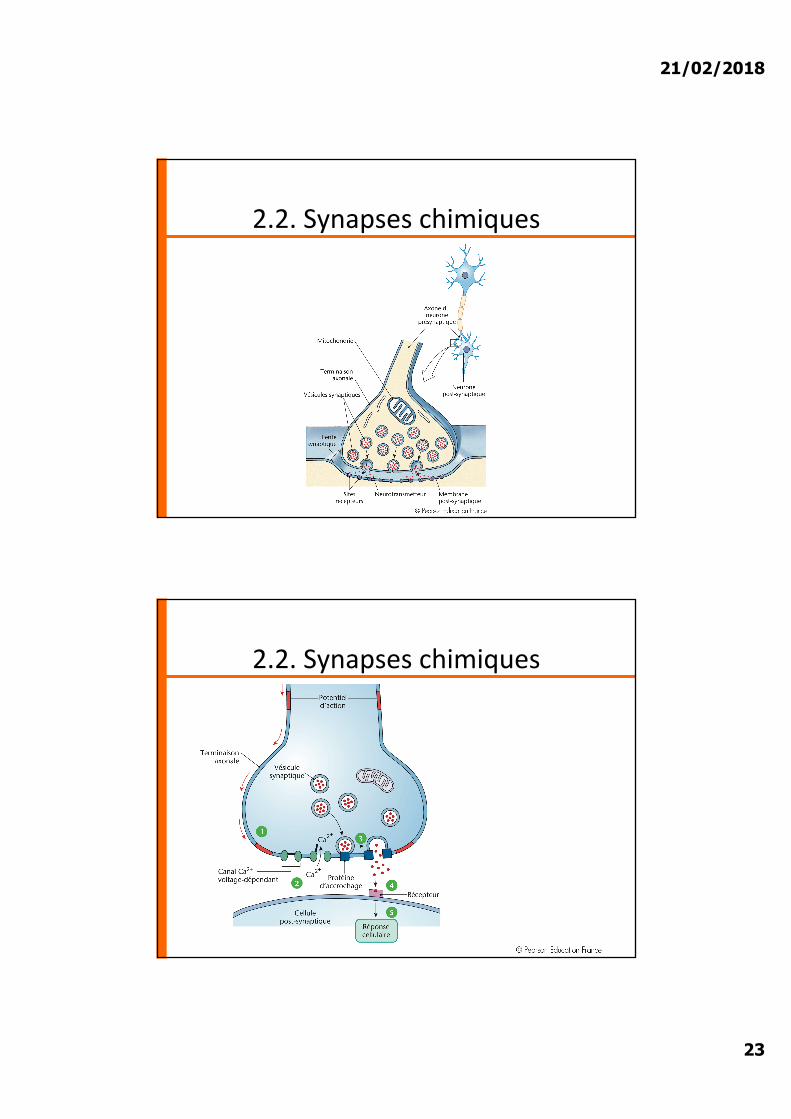
2.2. Synapses chimiques
2.2. Synapses chimiques
21/02/2018
24
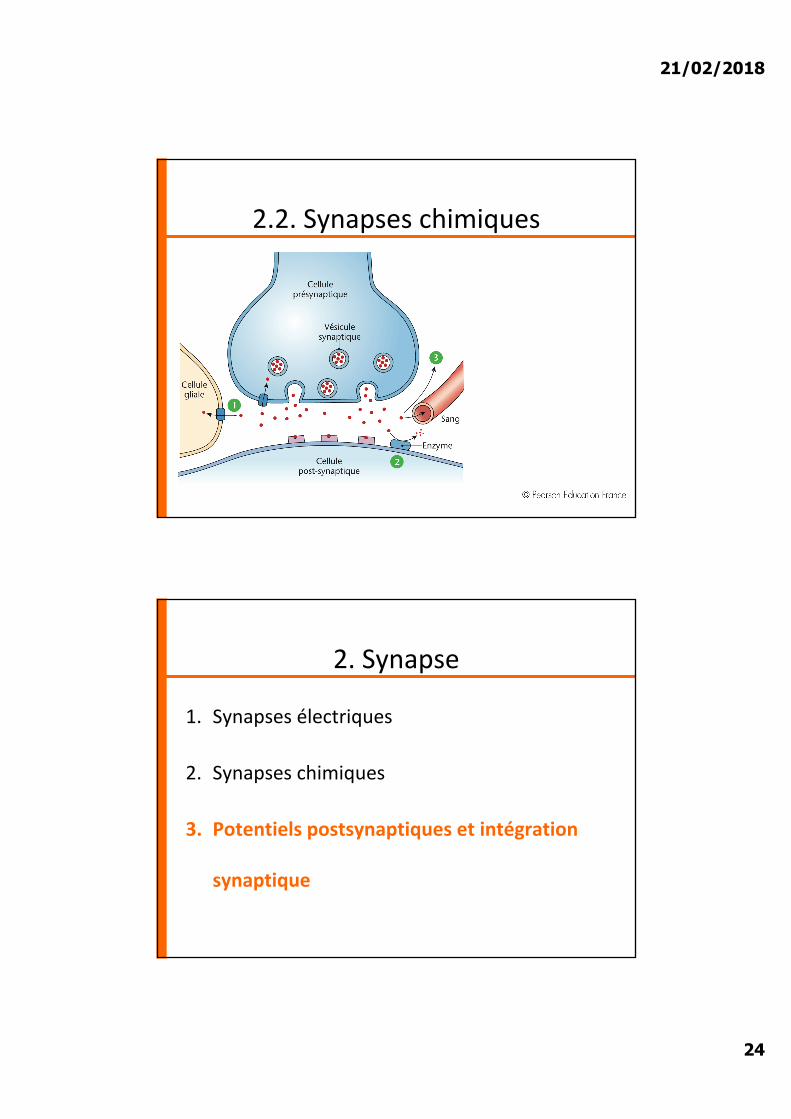
2.2. Synapses chimiques
2. Synapse
1. Synapses électriques
2. Synapses chimiques
3. Potentiels postsynaptiques et intégration
synaptique
21/02/2018
25
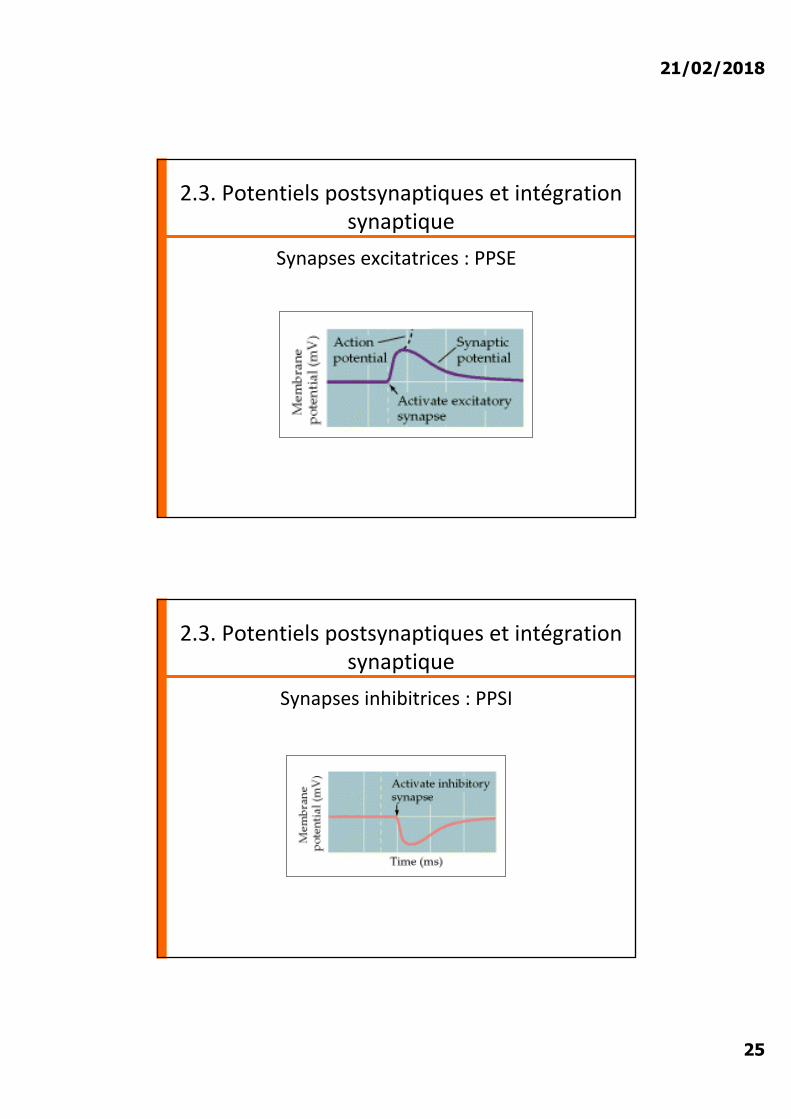
2.3. Potentiels postsynaptiques et intégration synaptique
Synapses excitatrices : PPSE
2.3. Potentiels postsynaptiques et intégration synaptique
Synapses inhibitrices : PPSI
21/02/2018
26
2.3. Potentiels postsynaptiques et intégration synaptique
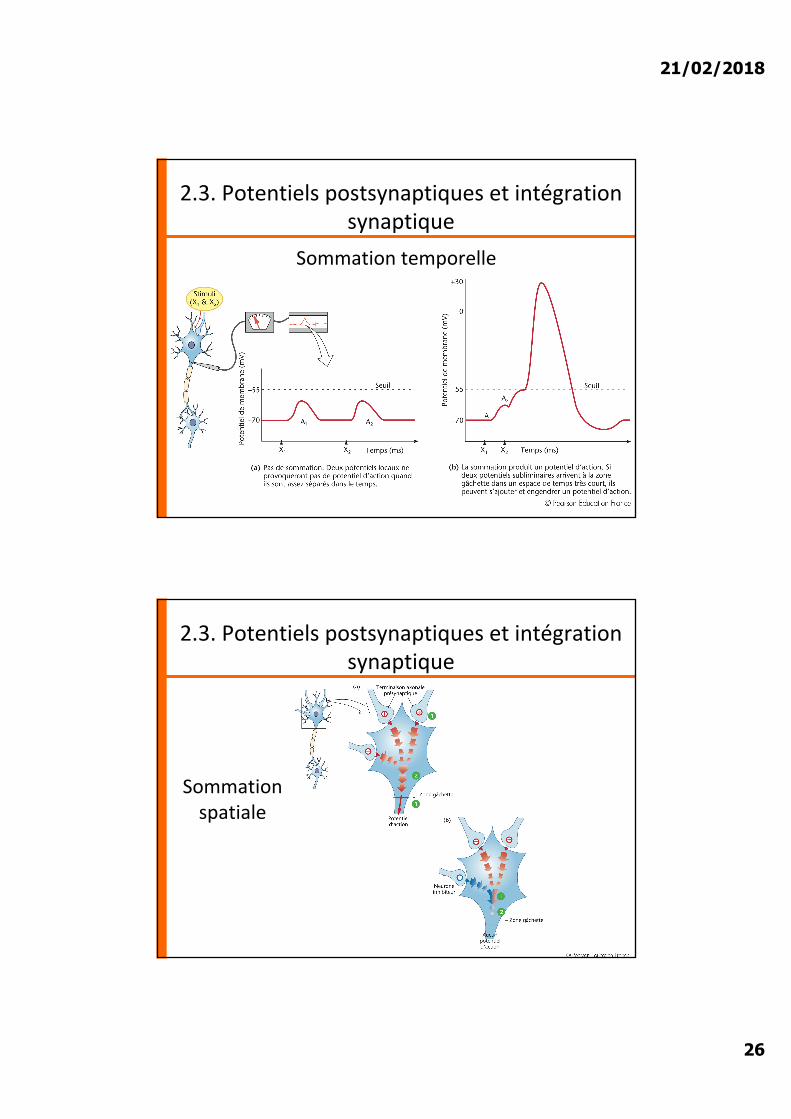
Sommation temporelle
2.3. Potentiels postsynaptiques et intégration synaptique
Sommation spatiale
21/02/2018
27
2.3. Potentiels postsynaptiques et intégration synaptique
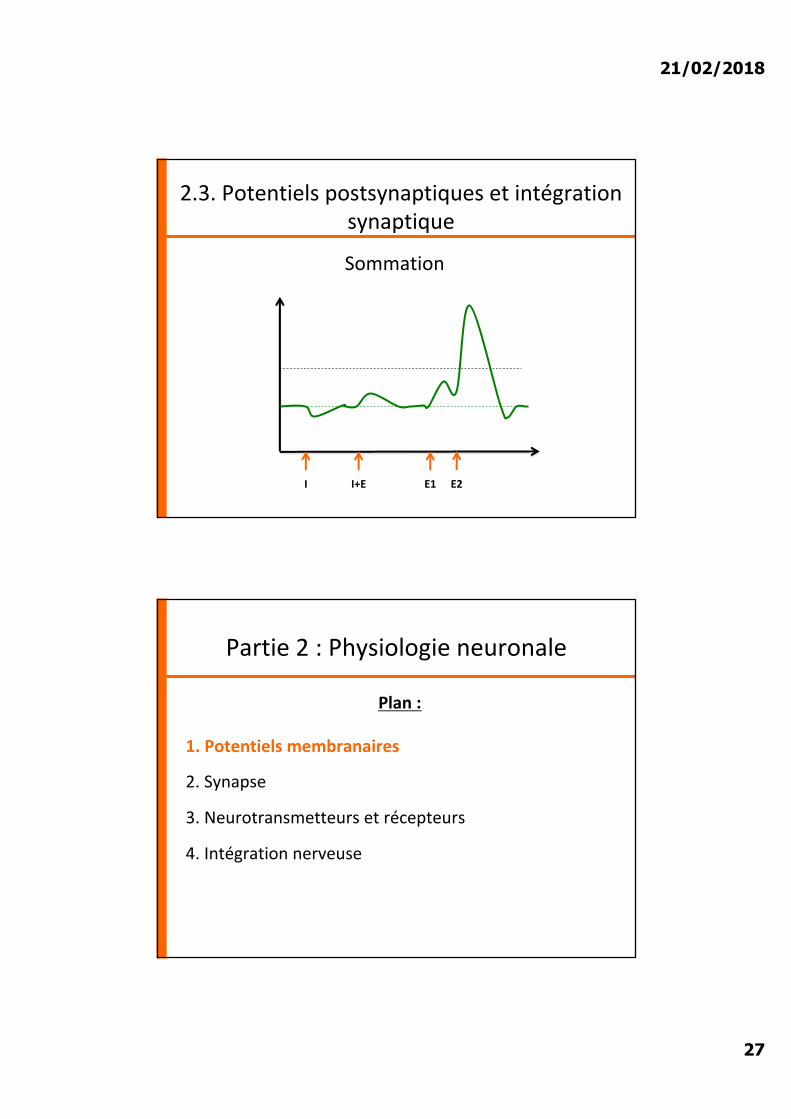
Sommation
I I+E E1 E2
Partie 2 : Physiologie neuronale
Plan :
1. Potentiels membranaires
2. Synapse
3. Neurotransmetteurs et récepteurs
4. Intégration nerveuse
21/02/2018
28
3. Neurotransmetteurs et récepteurs
Caractéristiques :
1. Présent dans le corpuscule terminal
2. Production de PPSE et PPSI
3. Élimination naturelle
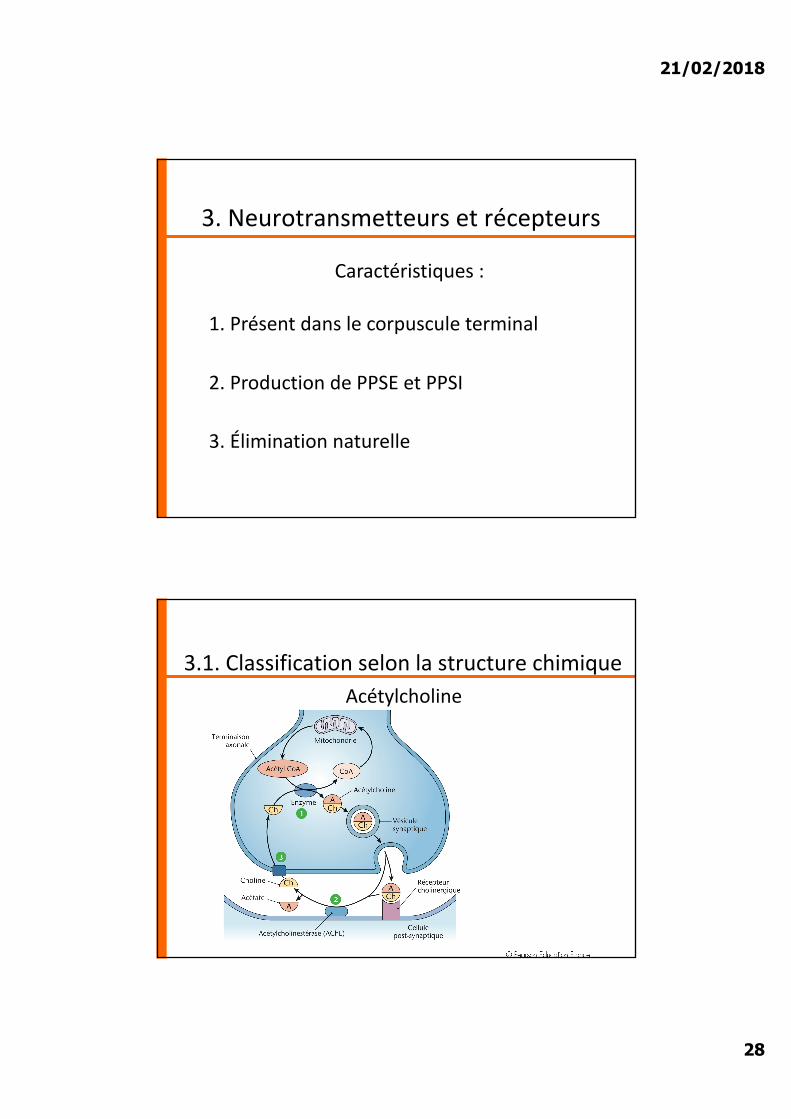
3.1. Classification selon la structure chimiqueAcétylcholine

21/02/2018
29
Neurotransmetteurs Classes fonctionnelles Sites de sécrétion
Noradrénaline (NA) + ou – selon récepteurAction indirecte SNC et SNP
Dopamine + ou – selon récepteurAction indirecte SNC et SNP
Sérotonie (5-HT)– en généralAction indirecte ou directe SNC
Histamine+ ou – selon récepteurAction indirecte SNC
Amines biogènes
3.1. Classification selon la structure chimique