Embed Size (px)
Citation preview

1
Programme Intégré de Recherches en Agroforesterie à ResTinclières
( PIRAT )
RAPPORT D'ETUDE 2008 Document remis au Conseil Général du Département de l’Hérault
SOMMAIRE
Equipes de recherche
INRA UMR SYSTEM – Dynamique du couple pucerons du blé - auxiliaires en parcelles agroforestières
p 2
Equipes de développement
Chambre d’Agriculture de l’Hérault – Interactions arbre/vigne Agroof Développement – FARM SAFE, un outil d’aide à la décision en Agroforesterie. Calibrage et premières simulations Centre Régional de la Propriété Forestière du Languedoc-Roussillon – Suivi phytosanitaire des arbres
p 13
p 23
p 42

2
Approche de la répartition spatiale des insectes ra vageurs des cultures intercalaires en parcelles agroforestières : cas de s pucerons du blé dur
INRA Montpellier, CBGP – UMR SYSTEM
Responsable du projet : SMITS Nathalie INRA – SYSTEM, 2 place Viala 34060 Montpellier cedex tél. 04.99.61.30.55, fax. 04.99.61.25.52 [email protected] Collaboration scientifique : Chrstian Dupraz (agroforesterie) Collaboration technique : Equipe METAFOR de l'UMR SYSTEM
I. INTRODUCTION
Dans les parcelles agroforestières, arbres et cultures intercalaires sont exposés aux
risques d'attaques de ravageurs ou de maladies fongiques d'une manière particulière, liée à
la structuration particulière de ces systèmes de production.
Ainsi, plusieurs phénomènes peuvent entrer en jeu concernant l'influence du système
agroforestier sur les risques phytosanitaires des espèces cultivées. On peut considérer que
la diversité végétale introduite par les linéaires d'arbres leur permet de servir de réservoir
aux ravageurs, ou au contraire à leurs ennemis naturels, comme le feraient des lisières de
champs, mais au niveau infra-parcellaire ; le microclimat auquel sont soumis les ravageurs
(et leurs ennemis naturels) et les agents de maladies cryptogamiques peut aussi être
sensiblement modifié par la présence d'arbres dans la culture, et ainsi entraîner une
modification substantielle de la vitesse de développement des insectes considérés ou du
risque d'incidence des maladies ; la structure en bandes de la culture intercalaire peut elle-
même contribuer à modifier l'accessibilité et la disponibilité de la ressource pour les
ravageurs, ou la dissémination des agents pathogènes.
C'est l'influence de la structure agroforestière sur le risque ravageur pour la culture
associée aux arbres qui est étudiée ici, appliquée au cas des pucerons des céréales sur du
blé dur en parcelles agroforestières à noyers, dans la partie sud du Domaine de
Restinclières, à Prades-le-Lez (34).
Dans ces parcelles agroforestières, les populations de pucerons et de certains de
leurs ennemis naturels ont été suivies depuis deux ans pour estimer ces phénomènes.
Cependant, ce suivi a été fait grâce à des échantillonnages aléatoires, qui ne tenaient pas
compte de la structuration particulière des populations de pucerons d'une part (formation de
colonies par des individus peu mobiles) ni de la disposition particulière du blé dur dans ces
parcelles d'autre part (en bandes séparées par des linéaires d'arbres). En effet, le
3
comportement peu mobile de ravageurs comme les pucerons des céréales, qui colonisent
entre autres la culture de proche en proche pendant leur phase aptère, peut être nettement
modifié par la rupture structurelle dans la ressource que constituent les linéaires d'arbres de
la parcelle agroforestière. Cette discontinuité de la ressource peut limiter la dissémination
des pucerons dans une période où les conditions abiotiques leurs sont par ailleurs
favorables.
Pour essayer de mettre en évidence une structuration spatiale de la population de
pucerons liée aux linéaires d'arbres dans la parcelle agroforestière, la répartition spatiale des
pucerons aux abords d'un linéaire et en s'éloignant de celui-ci a été comparée à la répartition
observée dans une parcelle de culture de blé dur pure au printemps 2008.
II. OBJETS D'ETUDE ET METHODES
1 . Dispositif expérimental
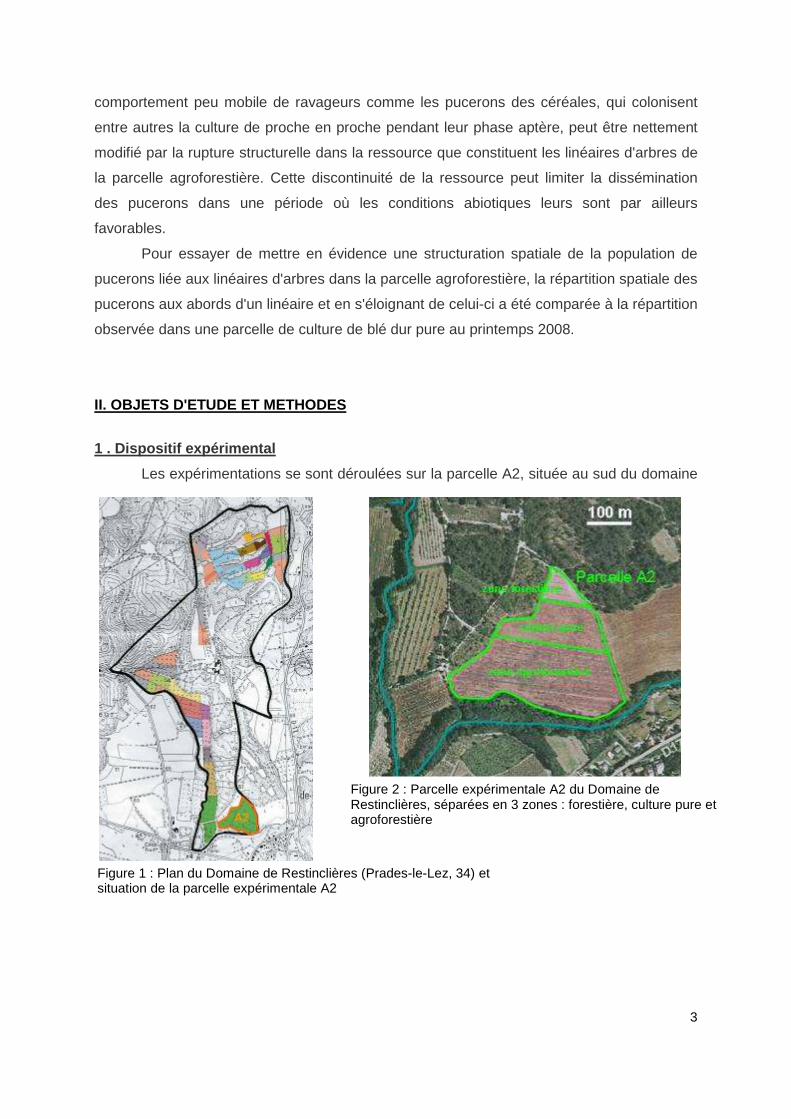
Les expérimentations se sont déroulées sur la parcelle A2, située au sud du domaine
Figure 1 : Plan du Domaine de Restinclières (Prades-le-Lez, 34) et situation de la parcelle expérimentale A2
Figure 2 : Parcelle expérimentale A2 du Domaine de Restinclières, séparées en 3 zones : forestière, culture pure et agroforestière

4
de Restinclières (Figure 1), constituée d'une partie agroforestière, d'une partie en culture
pure et d'une partie forestière accolées (Figure 2).
La culture annuelle suivie est du blé dur, au sein d'une rotation colza – blé dur ; dans
la zone agroforestière il est installé en bandes de 13 m de large séparées par des linéaires
de noyers hybrides âgés de 12 ans, initialement plantés tous les 4 mètres et éclaircis depuis.
2 . Protocoles expérimentaux
2.1 Insectes étudiés : Pucerons des céréales
Plusieurs milliers d'espèces de pucerons sont recensés dans le monde, et plusieurs
centaines d'entre elles sont considérées comme des ravageurs en agriculture. Les cycles
biologiques des pucerons sont variés, il leur faut le plus souvent deux hôtes végétaux, avec
un œuf d'hiver pondu sur l'hôte primaire. Ces caractéristiques se sont parfois perdues quand
l'hôte primaire se raréfie. Sur l'hôte secondaire et si les conditions sont favorables, les
femelles pondent directement de jeunes larves issues de reproduction asexuée
(parthénogénèse). Ce type de reproduction permet une accélération du cycle de
développement, et favorise la création de colonies de pucerons dès que les conditions
environnementales le permettent. Elle rend cependant la population très sensible au moindre
changement des conditions environnementales, notamment à la disponibilité de la ressource
végétale.
Les pucerons se nourrissent en pompant la sève élaborée dans les vaisseaux de
leurs plantes hôtes. Ces sont des phytophages assez spécifiques, chaque espèce ayant une
gamme d'hôte plutôt restreinte. Ainsi, sur céréales et sous nos climats, trois espèces de
pucerons dominent :
- Sitobion avenae Fabricius (figure 3a) a le plus souvent une seule plante hôte et est
inféodé aux graminées (Poacées). Les individus colonisent les céréales en
s’installant sur le limbe des feuilles puis migrent dans les épis dès leur sortie. Cette
Figure 3 : Pucerons des céréales. a : Sitobion avenae (adulte ailé et larves) ; b : Ropalosiphum padi (adulte aptère) ; c : Metopolophium dirhodum (larves de différents âges, dont une avec ébauches alaires).

5
espèce survit en hiver sur des résidus de culture ou des graminées spontanées. C'est
souvent l'espèce la plus abondante dans notre région, surtout après l'émergence des
épis. Les pucerons de cette espèce sont de couleurs très variées, mais avec toujours
des tarses et des cornicules noirs.
- Rhopalosiphum padi L (figure 3b) a conservé le plus souvent un cycle sur deux
hôtes, son hôte primaire est Prunus padus, le merisier à grappes. On le reconnaît à
sa forme très arrondie et à ses couleurs caractéristiques olive et bronze.
- Metopolophium dirhodum Walker (figure 3c) pond son œuf d’hiver sur des plantes du
genre Rosa. Les individus de cette espèce sont de couleur verte très claire, portant
souvent une ligne médiane plus foncée sur le dos.
Les dégâts provoqués par les pucerons des céréales peuvent être directs : les
prélèvements de sève entraînent une croissance et un développement réduits de la plante ;
ils peuvent être liés au dépôt de miellat : sur ces excréments se développent souvent un
ensemble de champignons saprophytes, regroupés sous le terme de fumagine, qui
diminuent le rayonnement reçu par les feuilles, et donc la photosynthèse ; enfin, les
pucerons des céréales sont des vecteurs potentiels de viroses des plantes, transmettant
notamment le virus de la jaunisse nanisante de l'orge, à l'origine de pertes importantes de
rendement certaines années, surtout si elle arrive précocement sur de jeunes plants. Il est à
noter cependant que la culture de blé dur en Languedoc Roussillon est très rarement victime
de fortes pullulations de pucerons induisant le recours à des interventions chimiques.
2.2 Suivi de la répartition spatiale des insectes et analyses
La répartition spatiale des pucerons a été estimée à deux reprises au cours du
printemps 2008 : les 21-22 mai et les 4-5 juin. La première date correspond à un blé
comportant encore des feuilles vertes et des épis jeunes, encore au stade laiteux, alors qu'à
la deuxième date les feuilles étaient sèches et le grain pâteux.
L'échantillonnage, le décompte des insectes et l'analyse de ces données ont été
réalisés suivant une méthode dite "SADIE" (Spatial Analysis by Distance IndicEs), décrite
notamment par Perry et al (1999)1.
Les notations ont été faites de manière non destructive sur des plants de blé en place
: chaque plant est examiné soigneusement pour compter tous les pucerons de chaque
espèce qu'il héberge de la manière la plus délicate possible. A chaque lieu de comptage, 5
1 Perry, J.N., Winder, L., Holland, J.M., Alston, R.D., 1999. Red-blue plots for detecting clusters in count data. Ecology Letters 2 (2), 106-113.

6
plants sont ainsi examinés et repérés pour être examinés à nouveau lors du comptage
suivant. Le nombre total de pucerons de chaque espèce sur l'ensemble des 5 plants est
collecté et utilisé pour l'analyse.
Ces comptages sont réalisés à tous les noeuds d'une grille dont la taille de la maille
dépend de la finesse avec laquelle on désire suivre la répartition spatiale. Trois grilles ont
ainsi été suivies, comportant chacune 6*6 points (Figure 4). Dans la partie agroforestière,
une "petite" grille, avec une maille de 1 m de côté a été étudiée, jouxtant un linéaire d'arbre,
pour rendre compte de l'organisation de la population de pucerons depuis le linéaire jusqu'à
environ le milieu de la bande de blé ; dans la zone en culture pure, une "petite" grille similaire
a été étudiée, proche du milieu de la parcelle, et une "grande" grille a également été utilisée,
incluant la petite grille comme une de ses mailles, et comportant également 6*6 points, mais
AF CP
30 m
5 m
LA
BLE
1 m
Figure 4 : Représentation schématique des points de prélèvement pour le suivi de la répartition
spatiale des pucerons du blé. AF : zone agroforestière (LA : linéaire d'arbres et bande enherbée
associée) ; CP : culture pure ; chaque point représente l'examen de 5 talles de blé.

7
avec une maile de 5 m de côté. Le côté de la grille est parallèle à la dernière ligne d'arbre,
lisière de la partie en culture pure de la parcelle A2 (Figure 5).
En tout, 180 talles ont été examinées à chaque fois dans la partie agroforestière et
340 talles dans la culture de blé pure.
III. RESULTATS - DISCUSSION
III.1 Abondance générale des pucerons
Sur l'ensemble des plants observés, on a dénombré un total de 698 pucerons les 21-
22 mai et 211 pucerons les 4 et 5 juin. De plus, on a pu constater lors de ce dernier
comptage que la très grande majorité des pucerons trouvés étaient localisés dans les épis.
Ceci est cohérent avec la sénéscence avancée des feuilles de blé lors du comptage de juin,
et place le deuxième décompte dans la phase de décroissance de la population des
pucerons pour la saison.
On distingue cependant de nettes différences entre les trois espèces de pucerons.
L'espèce S. avenae est largement dominante sur l'ensemble des talles observées,
puisqu'elle représente 72% des individus observés les 21 et 22 mai (M. dirhodum : 27% et
<1% pour R. padi) et environ 95% pour le décompte des 4 et 5 juin (M. dirhodum : 5% et
aucun R. padi). A nouveau, on peut mettre ces chiffres en relation avec la phénologie du blé.
En effet, seuls les épis étaient encore consommables par les pucerons les 4 et 5 juin ; or
l'espèce M. dirhodum est en général essentiellement présente sur feuilles : il est donc
Figure 5 : Implantation des zones d'échantillonnage fin (quadrillage bleu clair) et large (grille noir, points de relevés verts) de pucerons dans la parcelle A2
8
compréhensible que la proportion de cette espèce dans le total des pucerons soit plus faible
lors du deuxième comptage, où très peu de feuilles persistaient.
Si on considère séparément les prélèvements de la zone agroforestière et ceux de la
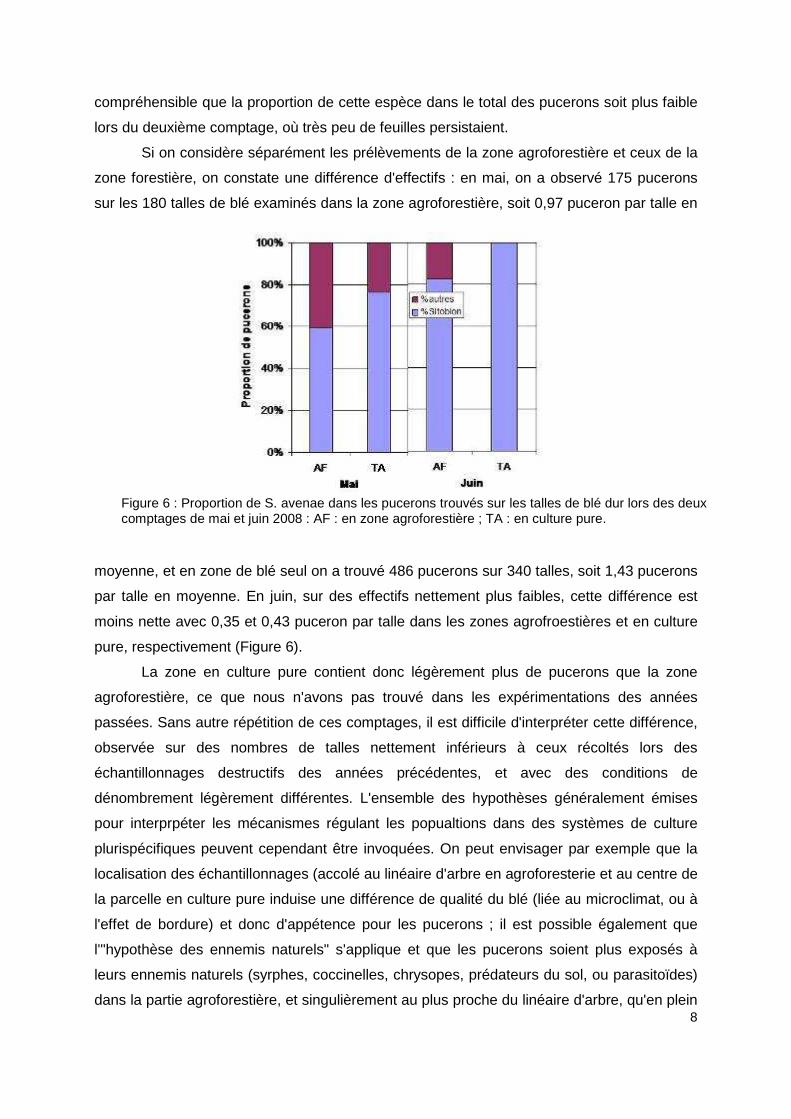
zone forestière, on constate une différence d'effectifs : en mai, on a observé 175 pucerons
sur les 180 talles de blé examinés dans la zone agroforestière, soit 0,97 puceron par talle en
moyenne, et en zone de blé seul on a trouvé 486 pucerons sur 340 talles, soit 1,43 pucerons
par talle en moyenne. En juin, sur des effectifs nettement plus faibles, cette différence est
moins nette avec 0,35 et 0,43 puceron par talle dans les zones agrofroestières et en culture
pure, respectivement (Figure 6).
La zone en culture pure contient donc légèrement plus de pucerons que la zone
agroforestière, ce que nous n'avons pas trouvé dans les expérimentations des années
passées. Sans autre répétition de ces comptages, il est difficile d'interpréter cette différence,
observée sur des nombres de talles nettement inférieurs à ceux récoltés lors des
échantillonnages destructifs des années précédentes, et avec des conditions de
dénombrement légèrement différentes. L'ensemble des hypothèses généralement émises
pour interprpéter les mécanismes régulant les popualtions dans des systèmes de culture
plurispécifiques peuvent cependant être invoquées. On peut envisager par exemple que la
localisation des échantillonnages (accolé au linéaire d'arbre en agroforesterie et au centre de
la parcelle en culture pure induise une différence de qualité du blé (liée au microclimat, ou à
l'effet de bordure) et donc d'appétence pour les pucerons ; il est possible également que
l'"hypothèse des ennemis naturels" s'applique et que les pucerons soient plus exposés à
leurs ennemis naturels (syrphes, coccinelles, chrysopes, prédateurs du sol, ou parasitoïdes)
dans la partie agroforestière, et singulièrement au plus proche du linéaire d'arbre, qu'en plein
Figure 6 : Proportion de S. avenae dans les pucerons trouvés sur les talles de blé dur lors des deux comptages de mai et juin 2008 : AF : en zone agroforestière ; TA : en culture pure.

9
centre de la zone en culture pure. Nous n'avons cependant pas les éléments pour tester ces
hypothèses dans les données collectées en 2008.
Les différences entre les deux types de système de culture (agroforestier / culture
pure) apparaissent aussi quant à la proportion de S. avenae présents sur les talles
observées. En effet, en mai, 59% des pucerons observés sur les talles de la zone
agroforestière étaient de cete espèce, alors qu'ils représentaient près de 71% des pucerons
échantillonnés dans la culture pure. En juin, les comptages en zone agroforestière
contenaient 82% de S. avenae, alors que c'était la seule espèce encore présente sur
l'ensemble des talles observés en culture de blé dur pure. Ceci peut être rapproché du fait
que les observations rapportées par l'équipe Agroforesterie de l'INRA indique un léger retard
dans la phénologie du blé dans les zones agroforestières par rapport à la culture pure ; or la
progression de l'âge physiologique correspond à la sénéscence des feuilles et est donc
défavorable aux pucerons des feuilles (M. dirhodum), tandis que les pucerons des épis (S.
avenae) peuvent y survivre plus longtemps.
Quoi qu'il en soit, ces observations ont été réalisées en un seul lieu de la zone
agroforestière (carré de 5 m par 5 m) et un seul lieu de la zone en culture pure (30 m par 30
m), et il serait hasardeux d'en tirer des conclusions sur l'ensemble de la parcelle, même s'il
est vraisemblable que les tendances observées sur ces échantillons sont représentatives de
ce qu'on aurait pu mesurer en multipliant les emplacement de décompte, d'après nos
expériences antérieures.
III.2 Répartition spatiale des pucerons
10
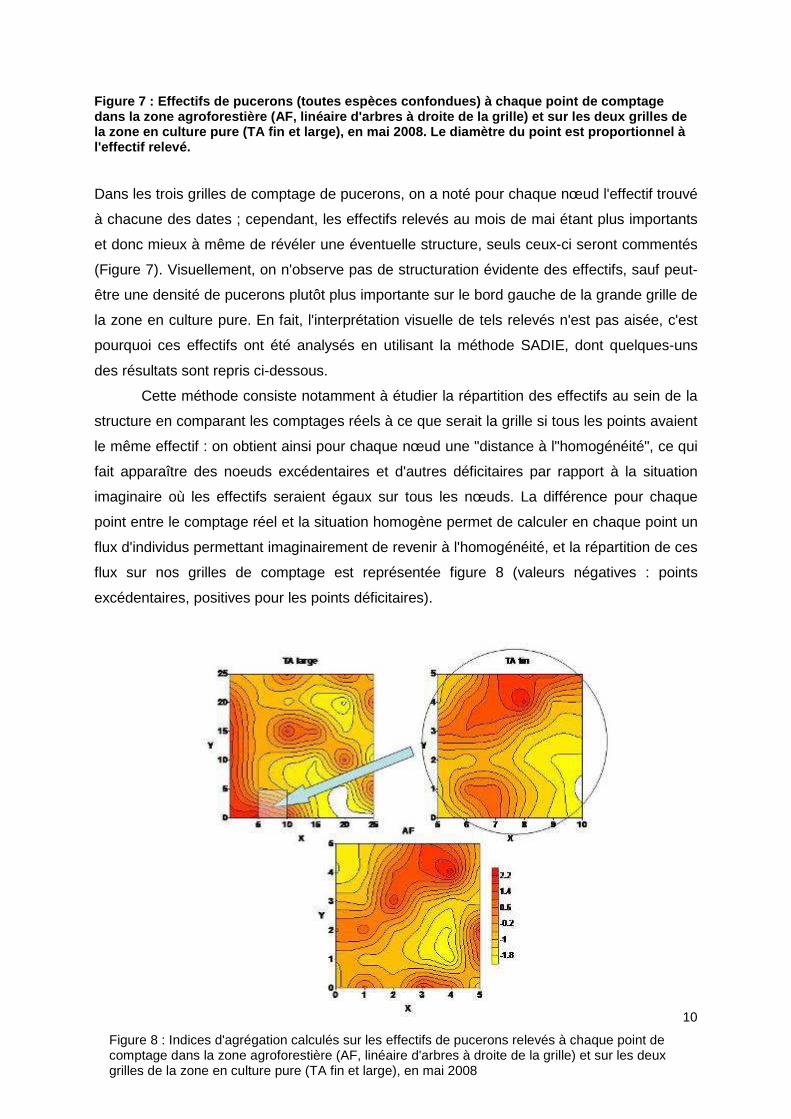
Figure 7 : Effectifs de pucerons (toutes espèces co nfondues) à chaque point de comptage dans la zone agroforestière (AF, linéaire d'arbres à droite de la grille) et sur les deux grilles de la zone en culture pure (TA fin et large), en mai 2 008. Le diamètre du point est proportionnel à l'effectif relevé. Dans les trois grilles de comptage de pucerons, on a noté pour chaque nœud l'effectif trouvé
à chacune des dates ; cependant, les effectifs relevés au mois de mai étant plus importants
et donc mieux à même de révéler une éventuelle structure, seuls ceux-ci seront commentés
(Figure 7). Visuellement, on n'observe pas de structuration évidente des effectifs, sauf peut-
être une densité de pucerons plutôt plus importante sur le bord gauche de la grande grille de
la zone en culture pure. En fait, l'interprétation visuelle de tels relevés n'est pas aisée, c'est
pourquoi ces effectifs ont été analysés en utilisant la méthode SADIE, dont quelques-uns
des résultats sont repris ci-dessous.
Cette méthode consiste notamment à étudier la répartition des effectifs au sein de la
structure en comparant les comptages réels à ce que serait la grille si tous les points avaient
le même effectif : on obtient ainsi pour chaque nœud une "distance à l"homogénéité", ce qui
fait apparaître des noeuds excédentaires et d'autres déficitaires par rapport à la situation
imaginaire où les effectifs seraient égaux sur tous les nœuds. La différence pour chaque
point entre le comptage réel et la situation homogène permet de calculer en chaque point un
flux d'individus permettant imaginairement de revenir à l'homogénéité, et la répartition de ces
flux sur nos grilles de comptage est représentée figure 8 (valeurs négatives : points
excédentaires, positives pour les points déficitaires).
Figure 8 : Indices d'agrégation calculés sur les effectifs de pucerons relevés à chaque point de comptage dans la zone agroforestière (AF, linéaire d'arbres à droite de la grille) et sur les deux grilles de la zone en culture pure (TA fin et large), en mai 2008

11
Les paramètres ainsi calculés et un nombre important de répartitions simulées
aléatoirement permettent de réaliser un test statistique sur nos données, en vue de
déterminer si la répartition observée des effectifs peut ou non être le résultat d'une répartition
aléatoire, ou si les effectifs aux points de prélèvement ont au contraire un arrangement
structuré.
On peut constater visuellement qu'il est difficile de mettre en évidence une
structuration nette des flux ainsi calculés sur notre dispositif. Les gammes de valeurs de ces
flux sont peu différentes entre les zones étudiées, variant de -2.12 à 2.42 en zone
agroforestière, de -2.5 à 2.29 dans la grille fine de la zone en culture pure et de -3.52 à 2.42
dans la grille large. Cette similarité dans les gammes de variation des valeurs de flux indique
peu de différence de structuration entre les trois grilles, ce qui montre que la grille en zone
agroforestière, bien que située le long d'un linéaire d'arbres, ne présente de structures
agrégatives plus fortes que la grille de même taille située en milieu de parcelle de blé. Il ne
semble donc pas que les arbres influencent significativement la répartition spatiale des
pucerons dans ce cas.
Ceci est confirmé par les tests statistiques réalisés sur ces données, qui donnent la
gamme de valeurs de flux correspondant à une répartition aléatoire de ces comptages, pour
chacune des trois grilles. Puis on calcule sur chaque un indice d'agrégation Ia, qui
représente les degré avec lequel les comptages observés sont agrégés, ce qui indique une
répartition éloignée de l'aléatoire pour des valeurs de Ia > 1, et une situation aléatoire pour
Ia=1. Dans la zone agroforestière, on trouve Ia = 1.055, et la probabilité que les données
observées ne soient pas plus ni moins agrégées que si les comptages étaient répartis
aléatoirement sur les nœuds est estimées à 32%, ce qui est faible par rapport aux 95%
généralement retenus pour ce genre de test statistique ; dans la grille fine de la zone en
culture pure, l'indice d'agrégation vaut 1.121, et la probabilité passe à 21%, et dans la grille
large Ia vaut 1.512 et la probabilité 9,6%. Cela conduit à accepter l'hypothèse de répartition
aléatoire de ces comptages dans les trois cas, et donc à considérer que les effectifs de
pucerons observés sur la parcelle sont répartis de manière aléatoire au sein des zones
étudiées.
IV - CONCLUSION
Les parcelles agroforestières de Restinclières ont fait l'objet, deux années de suite,
de suivis de populations de pucerons des céréales et de quelques ennemis naturels dans la
culture intercalaire de blé dur, dans trois lieux différents au sein du domaine. Lors de ces
divers suivis, aucune différence n'a pu être mise en évidence, en terme de dynamique des
populations de ces insectes, entre les parcelles agroforestières et les zones de culture pure

12
attenantes étudiées simultanément. Or les différentes hypothèses écologiques concernant
les équilibres entre pucerons ravageurs et auxiliaires prédateurs ou parasites de ceux-ci, et
leur lien avec l'habitat (diversité végétale, structure de la ressource en blé, influence sur le
microclimat…) auraient pu laisser prévoir des dynamiques différentes dans ces deux
systèmes de culture.
L'environnement particulièrement riche, en terme de biodiversité, que constitue le
Domaine de Restinclières et la relativement petite taille des parcelle en culture pure étudiées
ont notamment pu être invoqués pour expliquer que, si de telles différences sont
susceptibles d'apparaître, elles ne puissent pas être mises en évidence dans nos conditions
expérimentales. Cependant, il n'était pas à exclure que notre mode d'échantillonnage lors de
ces mesures (prélèvement aléatoire de lots de 25 talles dans des zones définies) soit peu
adapté à l'estimation de populations pucerons dont on connaît par ailleurs le mode de
développement en colonies, et qui en outre s'installent, pour les parcelles agroforestières,
dans un environnement par définition morcelé en bandes. Il était donc utile de vérifier qu'une
très forte structuration spatiale de la répartition des pucerons au sein des bandes de blé
dans la parcelle agroforestière ne rendait pas inopérant le suivi de la dynamique des
populations tel que réalisé lors des campagnes précédentes. Les mesures de répartition
spatiale réalisées cette année montrent une variabilité de la population de pucerons entre les
points de prélèvement, mais pas ou très peu d'agrégation des effectifs, ce qui valide un
échantillonnage aléatoire comme mode de suivi de la population, mais incite à maintenir un
nombre de talles par échantillon assez important.
Outre la validation du mode d'échantillonnage adopté précédemment, ces comptages
nous ont indiqué que la présence des linéaires d'arbres dans la parcelle agroforestière
semblait peu influencer la répartition spatiale des colonies de pucerons des céréales sur le
blé dur en tant que culture intercalaire. Ceci a cependant été démontré à seulement deux
dates, dont l'une correspond à des effectifs assez faibles et en toute fin de développement
des pucerons sur cette culture. Un suivi spatio-temporel des effectifs de pucerons sur
l'ensemble d'une saison pourrait permettre de tester l'influence ou non des linéaires d'arbres
à différents stades de la dynamique du puceron sur cette culture. La question de la
répartition spatiale des bioagresseurs (ravageurs ou agents pathogènes cryptogamiques)
dans une culture intercalaire en parcelle agroforestière et de l'influence des arbres sur celle-
ci représente en effet un des phénomènes à bien connaître pour pouvoir estimer la
modification du risque phytosanitaire sur une culture donnée, liée à la présence des arbres
au sein de la parcelle cultivée.

13
Étude des interactions entre l'arbre et la vigne en agroforesterie
Chambre d’Agriculture de l’Hérault
Adresse Correspondant du projet
Chambre d'agriculture de l'Hérault ; 15 rue V. Hugo 34120 Pézenas
Nathalie Goma-Fortin
Chercheurs impliqués Nathalie Goma-Fortin, William Trambouze
Objectifs de l'étude Dans le cadre de parcelles agroforestières, l'objectif est d'évaluer l'impact des arbres sur la culture intercalaire, la vigne dans cette partie de l'étude. L'évolution inter-annuelle de la qualité et de la quantité de raisins produits seront les premiers indicateurs d'une concurrence éventuelle entre les arbres et les ceps de vigne et de son mode d'installation. Compte tenu du caractère pérenne de la vigne, l'étude sera menée à court et à moyen terme sur la récolte de raisins et à plus long terme sur l’expression végétative, la puissance et le système racinaire des ceps.
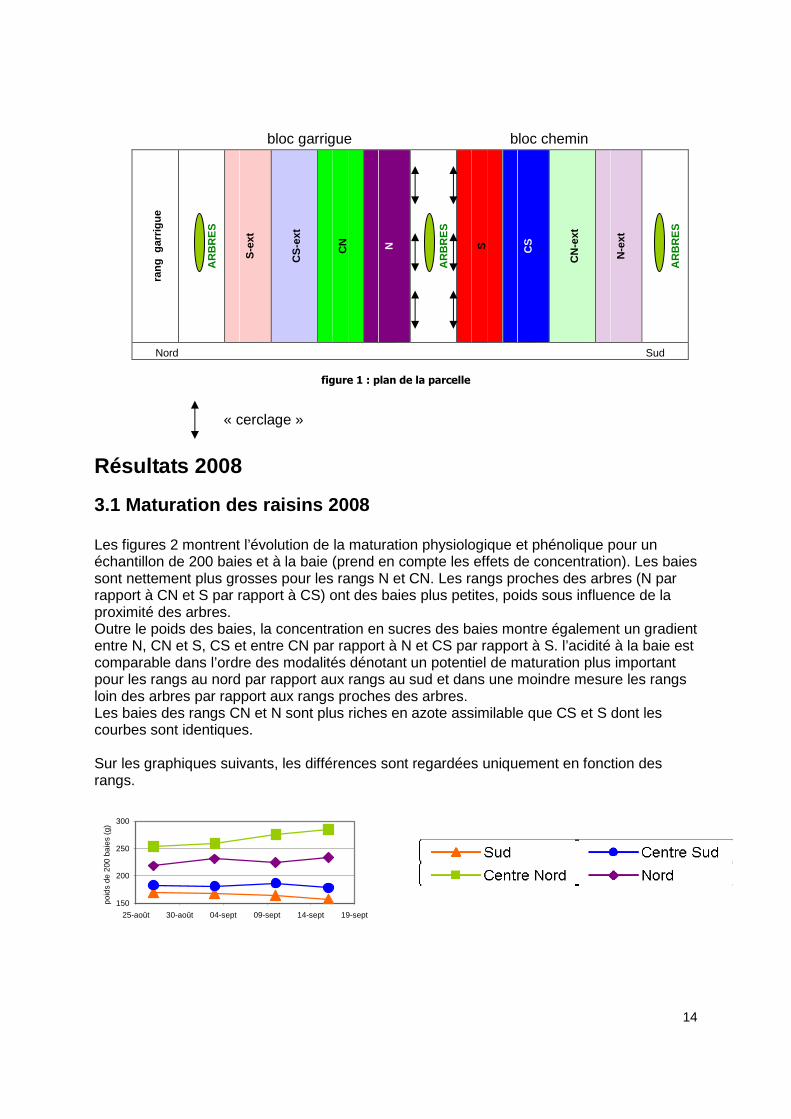
Protocole Les parcelles choisies sur le domaine de Restinclières sont la B4 et, depuis 2004, la B10. La parcelle B4, suivie depuis 2001, est plantée en syrah-cormier (feuillus). Elle regroupe des plantations extensives (la rangée d’arbres remplace deux rangs de vigne) et intensives (la rangée d’arbres remplace un rang de vigne). Cette parcelle ne donnant toujours pas de différences significatives entre les modalités. Il a été décidé d’arrêter temporairement son suivi pour se consacrer pleinement au suivi de la parcelle B10 plantée en syrah-pins brutia (résineux). Ces arbres se sont développés nettement plus rapidement que les cormiers. Cette parcelle est plantée en extensif uniquement. En 2007, les modalités ont été doublées par « cerclage » des arbres centraux. L’idée est de limiter la concurrence en coupant les racines des arbres. Ce cerclage a été obtenu par le passage d’une dent de sous-soleuse à 1m75 des arbres et des rangs de vigne immédiats (N et S). 3 répétitions « cerclé » et « non cerclé » ont été réalisées. La figure 1 représente le plan de la parcelle. En 2008, le suivi a porté sur les paramètres classiques de la production de raisins. Les caractéristiques portant sur la taille (poids de bois de taille, vigueur…) et la production totale de matière sèche pour le millésime (puissance) seront abordées dans le compte-rendu 2009 (la taille se faisant en décembre, janvier). Seuls les rangs autour du rang d’arbre central (CN, N, S, CS) ont été suivis sur les paramètres de contrôle de maturité, d’azote assimilable et du δ13C donnant une idée de la contrainte hydrique subie par les plantes.
14
rang
gar
rigue
AR
BR
ES
S-e
xt
CS
-ext
CN
N
AR
BR
ES
S
CS
CN
-ext
N-e
xt
AR
BR
ES
Nord Sud
figure 1 : plan de la parcelle
Résultats 2008
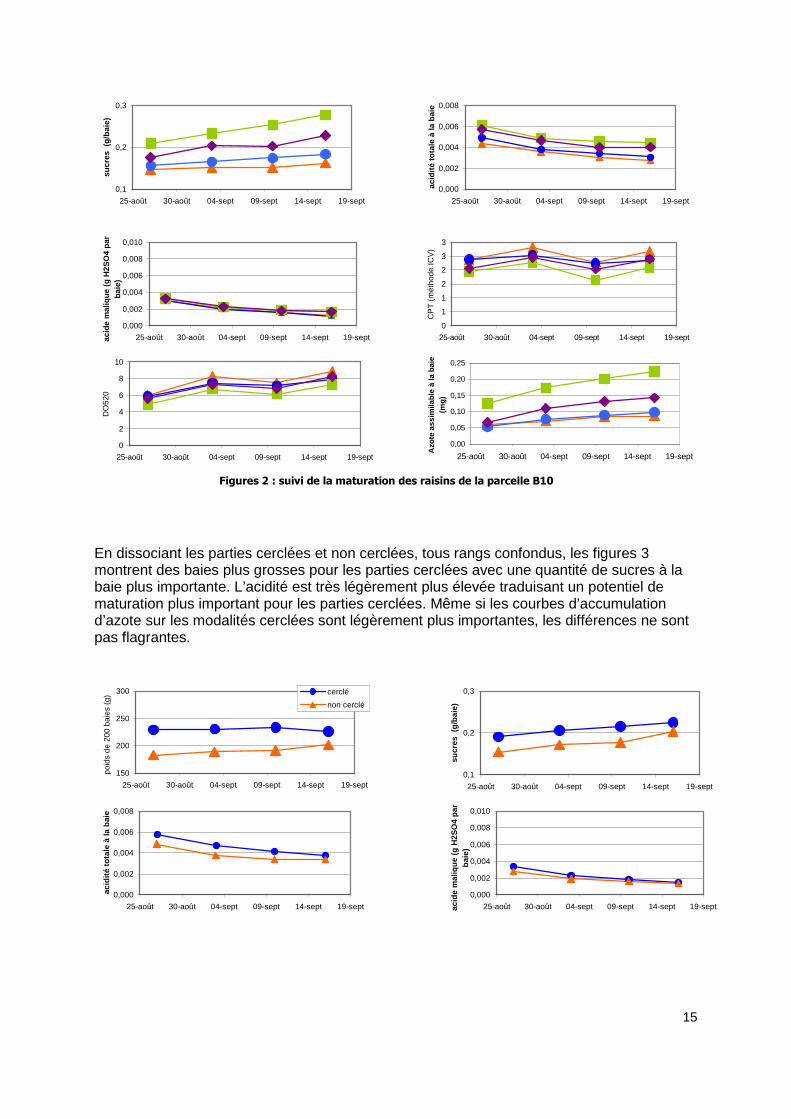
3.1 Maturation des raisins 2008 Les figures 2 montrent l’évolution de la maturation physiologique et phénolique pour un échantillon de 200 baies et à la baie (prend en compte les effets de concentration). Les baies sont nettement plus grosses pour les rangs N et CN. Les rangs proches des arbres (N par rapport à CN et S par rapport à CS) ont des baies plus petites, poids sous influence de la proximité des arbres. Outre le poids des baies, la concentration en sucres des baies montre également un gradient entre N, CN et S, CS et entre CN par rapport à N et CS par rapport à S. l’acidité à la baie est comparable dans l’ordre des modalités dénotant un potentiel de maturation plus important pour les rangs au nord par rapport aux rangs au sud et dans une moindre mesure les rangs loin des arbres par rapport aux rangs proches des arbres. Les baies des rangs CN et N sont plus riches en azote assimilable que CS et S dont les courbes sont identiques. Sur les graphiques suivants, les différences sont regardées uniquement en fonction des rangs.
150
200
250
300
25-août 30-août 04-sept 09-sept 14-sept 19-sept
poid
s de
200
bai
es (
g)
bloc chemin bloc garrigue
« cerclage »
15
0,1
0,2
0,3
25-août 30-août 04-sept 09-sept 14-sept 19-sept
sucr
es (
g/ba
ie)
0,000
0,002
0,004
0,006
0,008
25-août 30-août 04-sept 09-sept 14-sept 19-sept
acid
ité to
tale
à la
bai
e
0,000
0,002
0,004
0,006
0,008
0,010
25-août 30-août 04-sept 09-sept 14-sept 19-septacid
e m
aliq
ue (
g H
2SO
4 pa
r ba
ie)
0
1
1
2
2
3
3
25-août 30-août 04-sept 09-sept 14-sept 19-sept
CP
T (
mét
hode
ICV
)
0
2
4
6
8
10
25-août 30-août 04-sept 09-sept 14-sept 19-sept
DO
520
0,00
0,05
0,10
0,15
0,20
0,25
25-août 30-août 04-sept 09-sept 14-sept 19-sept
Azo
te a
ssim
ilabl
e à
la b
aie
(mg)
Figures 2 : suivi de la maturation des raisins de la parcelle B10
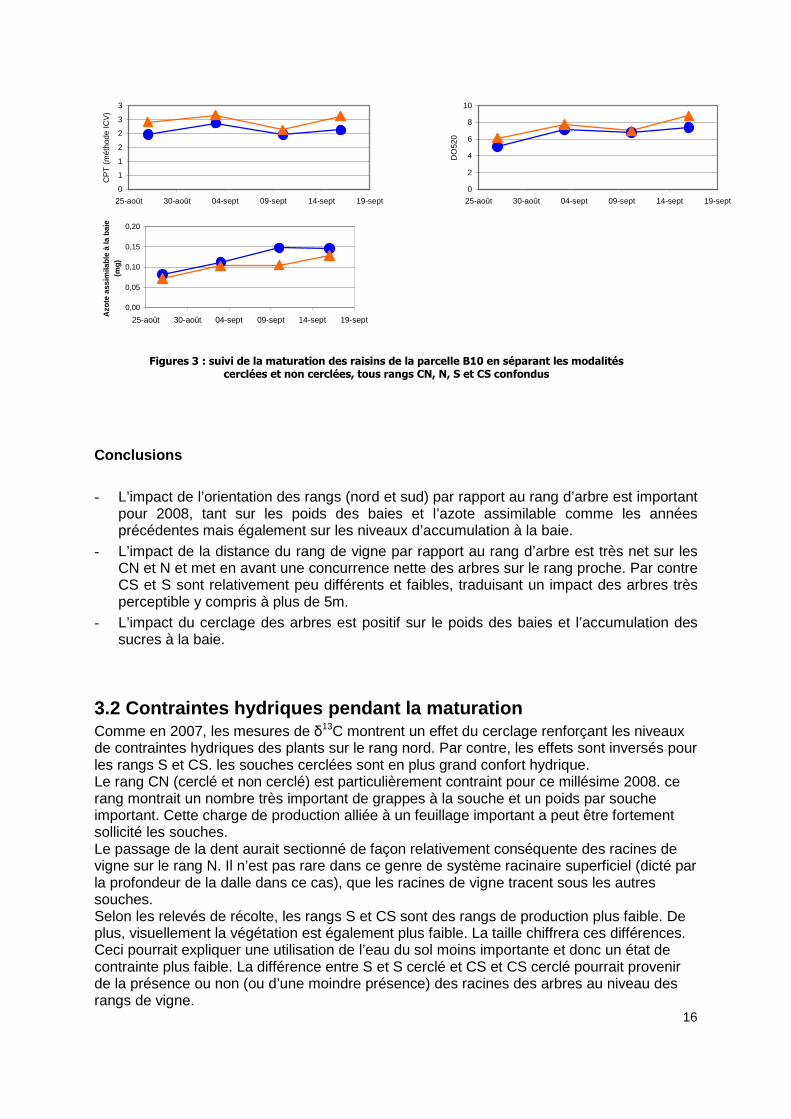
En dissociant les parties cerclées et non cerclées, tous rangs confondus, les figures 3 montrent des baies plus grosses pour les parties cerclées avec une quantité de sucres à la baie plus importante. L’acidité est très légèrement plus élevée traduisant un potentiel de maturation plus important pour les parties cerclées. Même si les courbes d’accumulation d’azote sur les modalités cerclées sont légèrement plus importantes, les différences ne sont pas flagrantes.
150
200
250
300
25-août 30-août 04-sept 09-sept 14-sept 19-sept
poid
s de
200
bai
es (
g)
cerclé
non cerclé
0,1
0,2
0,3
25-août 30-août 04-sept 09-sept 14-sept 19-sept
sucr
es (
g/ba
ie)
0,000
0,002
0,004
0,006
0,008
25-août 30-août 04-sept 09-sept 14-sept 19-sept
acid
ité to
tale
à la
bai
e
0,000
0,002
0,004
0,006
0,008
0,010
25-août 30-août 04-sept 09-sept 14-sept 19-septacid
e m
aliq
ue (
g H
2SO
4 pa
r ba
ie)
16
0
1
1
2
2
3
3
25-août 30-août 04-sept 09-sept 14-sept 19-sept
CP
T (
mét
hode
ICV
)
0
2
4
6
8
10
25-août 30-août 04-sept 09-sept 14-sept 19-sept
DO
520
0,00
0,05
0,10
0,15
0,20
25-août 30-août 04-sept 09-sept 14-sept 19-sept
Azo
te a
ssim
ilabl
e à
la b
aie
(mg)
Figures 3 : suivi de la maturation des raisins de la parcelle B10 en séparant les modalités cerclées et non cerclées, tous rangs CN, N, S et CS confondus
Conclusions
- L’impact de l’orientation des rangs (nord et sud) par rapport au rang d’arbre est important pour 2008, tant sur les poids des baies et l’azote assimilable comme les années précédentes mais également sur les niveaux d’accumulation à la baie.
- L’impact de la distance du rang de vigne par rapport au rang d’arbre est très net sur les CN et N et met en avant une concurrence nette des arbres sur le rang proche. Par contre CS et S sont relativement peu différents et faibles, traduisant un impact des arbres très perceptible y compris à plus de 5m.
- L’impact du cerclage des arbres est positif sur le poids des baies et l’accumulation des sucres à la baie.
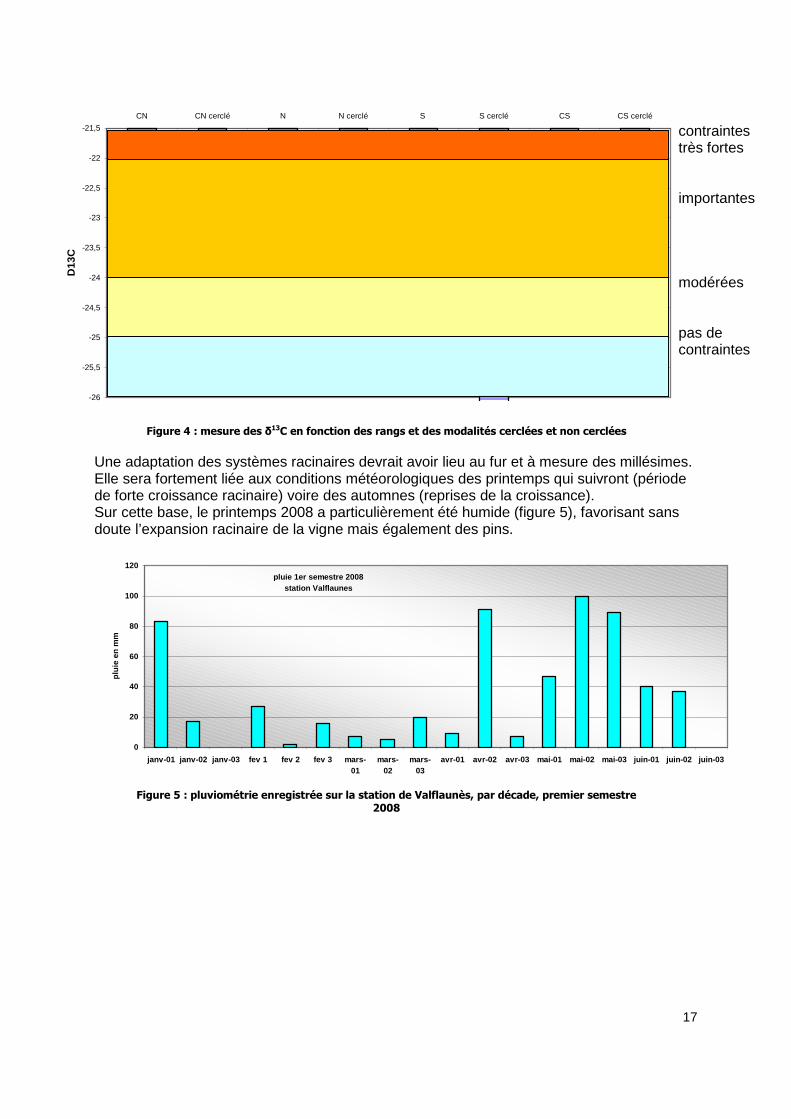
3.2 Contraintes hydriques pendant la maturation Comme en 2007, les mesures de δ13C montrent un effet du cerclage renforçant les niveaux de contraintes hydriques des plants sur le rang nord. Par contre, les effets sont inversés pour les rangs S et CS. les souches cerclées sont en plus grand confort hydrique. Le rang CN (cerclé et non cerclé) est particulièrement contraint pour ce millésime 2008. ce rang montrait un nombre très important de grappes à la souche et un poids par souche important. Cette charge de production alliée à un feuillage important a peut être fortement sollicité les souches. Le passage de la dent aurait sectionné de façon relativement conséquente des racines de vigne sur le rang N. Il n’est pas rare dans ce genre de système racinaire superficiel (dicté par la profondeur de la dalle dans ce cas), que les racines de vigne tracent sous les autres souches. Selon les relevés de récolte, les rangs S et CS sont des rangs de production plus faible. De plus, visuellement la végétation est également plus faible. La taille chiffrera ces différences. Ceci pourrait expliquer une utilisation de l’eau du sol moins importante et donc un état de contrainte plus faible. La différence entre S et S cerclé et CS et CS cerclé pourrait provenir de la présence ou non (ou d’une moindre présence) des racines des arbres au niveau des rangs de vigne.
17
-26
-25,5
-25
-24,5
-24
-23,5
-23
-22,5
-22
-21,5
CN CN cerclé N N cerclé S S cerclé CS CS cerclé
D13
C
Figure 4 : mesure des δ13C en fonction des rangs et des modalités cerclées et non cerclées
Une adaptation des systèmes racinaires devrait avoir lieu au fur et à mesure des millésimes. Elle sera fortement liée aux conditions météorologiques des printemps qui suivront (période de forte croissance racinaire) voire des automnes (reprises de la croissance). Sur cette base, le printemps 2008 a particulièrement été humide (figure 5), favorisant sans doute l’expansion racinaire de la vigne mais également des pins.
pluie 1er semestre 2008 station Valflaunes
0
20
40
60
80
100
120
janv-01 janv-02 janv-03 fev 1 fev 2 fev 3 mars-01
mars-02
mars-03
avr-01 avr-02 avr-03 mai-01 mai-02 mai-03 juin-01 juin-02 j uin-03
plui
e en
mm
Figure 5 : pluviométrie enregistrée sur la station de Valflaunès, par décade, premier semestre
2008
contraintes très fortes importantes modérées pas de contraintes
18
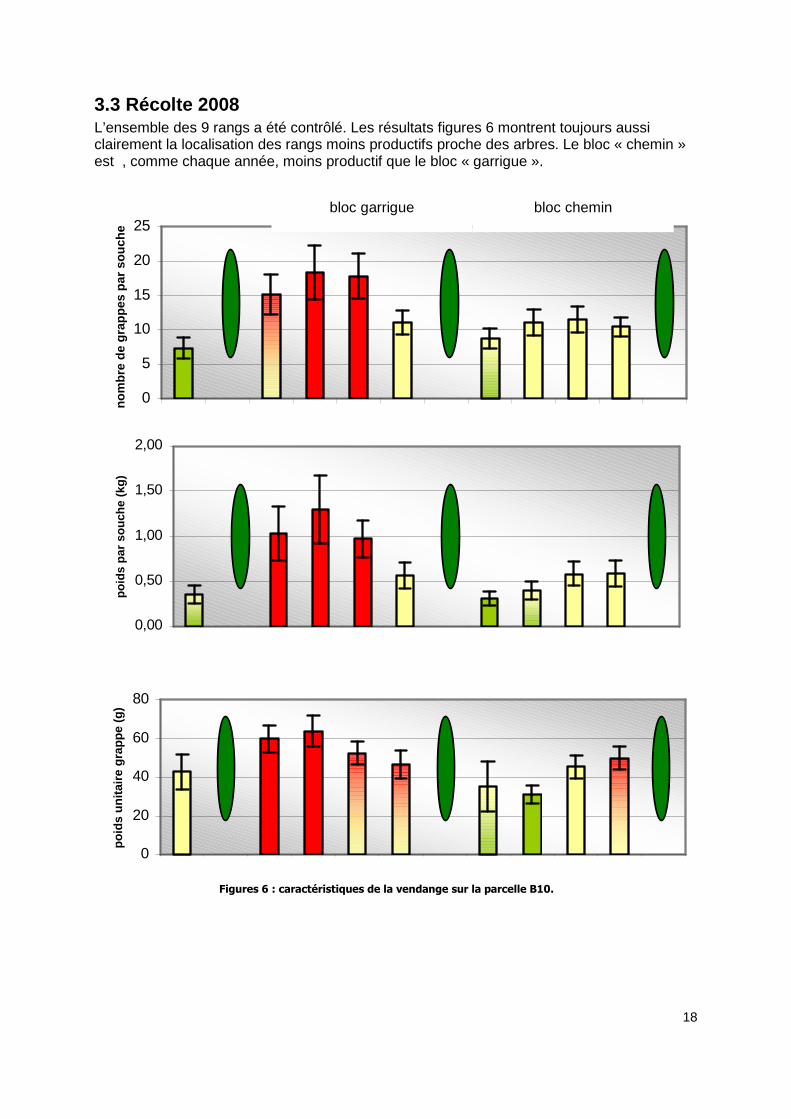
3.3 Récolte 2008 L’ensemble des 9 rangs a été contrôlé. Les résultats figures 6 montrent toujours aussi clairement la localisation des rangs moins productifs proche des arbres. Le bloc « chemin » est , comme chaque année, moins productif que le bloc « garrigue ».
0
5
10
15
20
25
nom
bre
de g
rapp
es p
ar s
ouch
e
0,00
0,50
1,00
1,50
2,00
poid
s pa
r so
uche
(kg
)
0
20
40
60
80
poid
s un
itaire
gra
ppe
(g)
Figures 6 : caractéristiques de la vendange sur la parcelle B10.
bloc chemin bloc garrigue
19
3.4 Taille 2008 La taille 2008 n’a pas encore été réalisée à la date du rapport. Elle le sera en décembre 08 / janvier 09 et fera l’objet d’un chapitre au prochain compte rendu. .
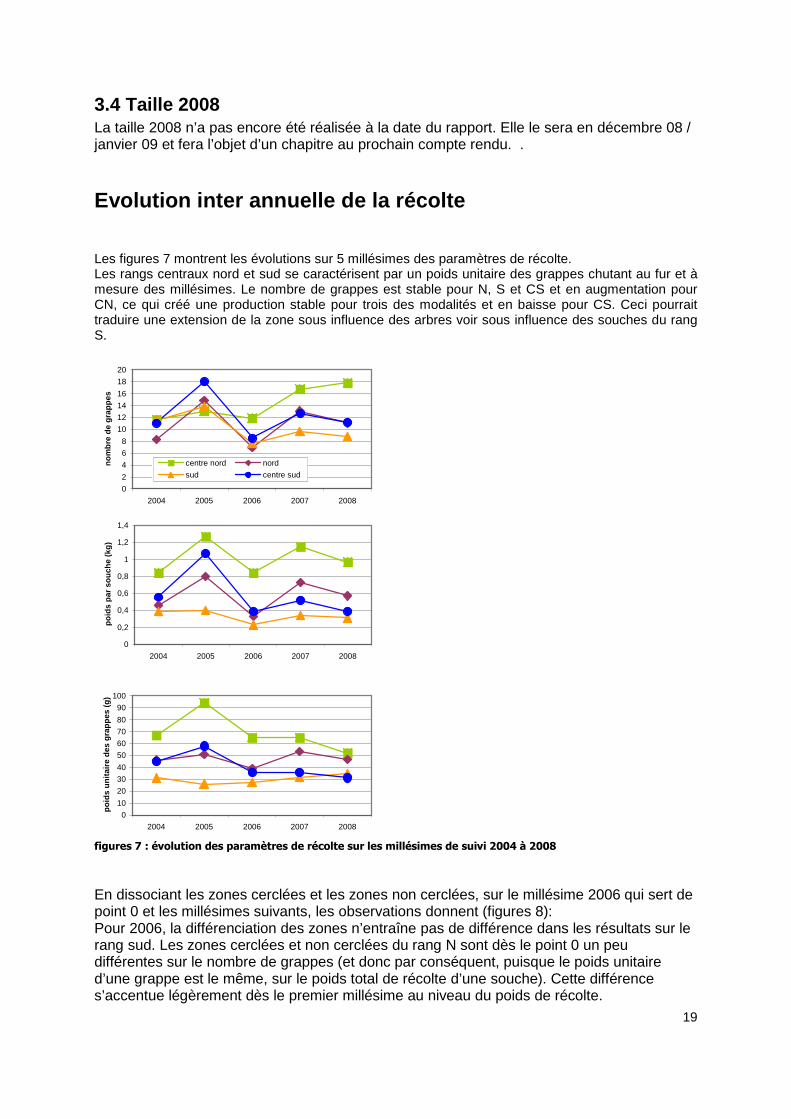
Evolution inter annuelle de la récolte Les figures 7 montrent les évolutions sur 5 millésimes des paramètres de récolte. Les rangs centraux nord et sud se caractérisent par un poids unitaire des grappes chutant au fur et à mesure des millésimes. Le nombre de grappes est stable pour N, S et CS et en augmentation pour CN, ce qui créé une production stable pour trois des modalités et en baisse pour CS. Ceci pourrait traduire une extension de la zone sous influence des arbres voir sous influence des souches du rang S.
0
2
4
6
8
10
12
14
16
18
20
2004 2005 2006 2007 2008
nom
bre
de g
rapp
es
centre nord nord
sud centre sud
0
0,2
0,4
0,6
0,8
1
1,2
1,4
2004 2005 2006 2007 2008
poid
s pa
r so
uche
(kg
)
0
10
20
30
40
50
60
70
80
90
100
2004 2005 2006 2007 2008
poid
s un
itaire
des
gra
ppes
(g)
figures 7 : évolution des paramètres de récolte sur les millésimes de suivi 2004 à 2008
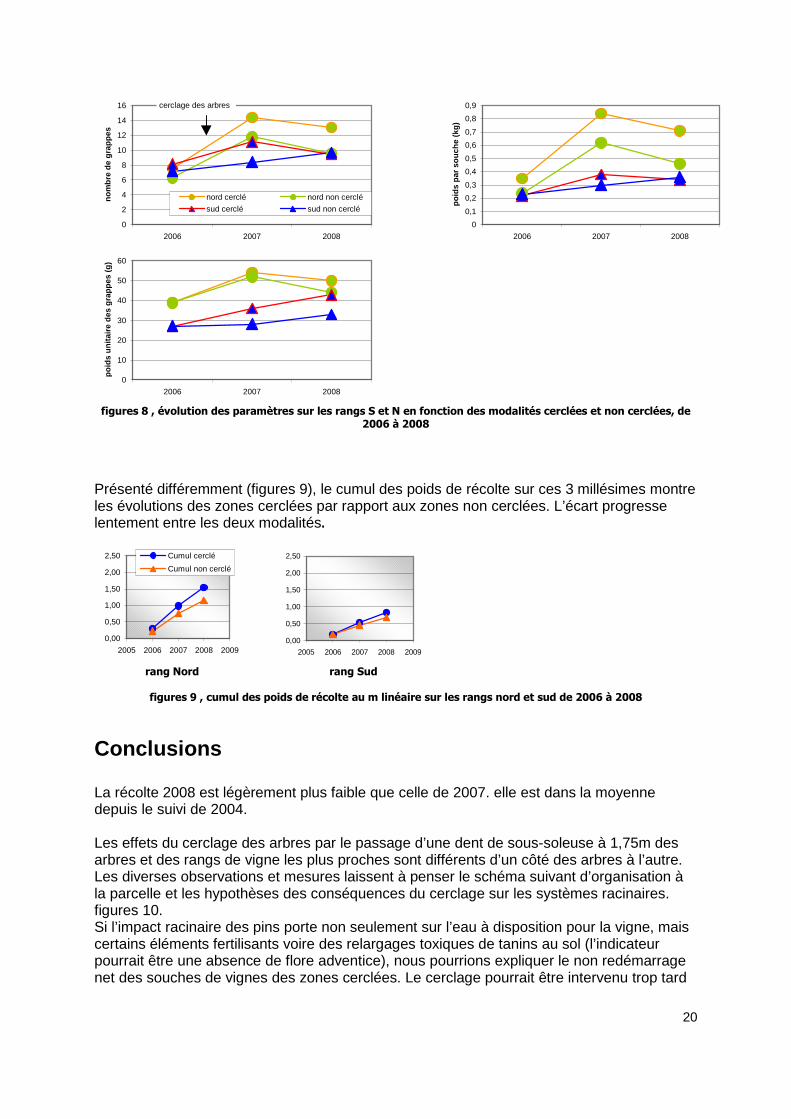
En dissociant les zones cerclées et les zones non cerclées, sur le millésime 2006 qui sert de point 0 et les millésimes suivants, les observations donnent (figures 8): Pour 2006, la différenciation des zones n’entraîne pas de différence dans les résultats sur le rang sud. Les zones cerclées et non cerclées du rang N sont dès le point 0 un peu différentes sur le nombre de grappes (et donc par conséquent, puisque le poids unitaire d’une grappe est le même, sur le poids total de récolte d’une souche). Cette différence s’accentue légèrement dès le premier millésime au niveau du poids de récolte.
20
0
2
4
6
8
10
12
14
16
2006 2007 2008
nom
bre
de g
rapp
es
nord cerclé nord non cerclé
sud cerclé sud non cerclé
cerclage des arbres
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
2006 2007 2008
poid
s pa
r so
uche
(kg
)
0
10
20
30
40
50
60
2006 2007 2008
poid
s un
itaire
des
gra
ppes
(g)
figures 8 , évolution des paramètres sur les rangs S et N en fonction des modalités cerclées et non cerclées, de 2006 à 2008
Présenté différemment (figures 9), le cumul des poids de récolte sur ces 3 millésimes montre les évolutions des zones cerclées par rapport aux zones non cerclées. L’écart progresse lentement entre les deux modalités.
0,00
0,50
1,00
1,50
2,00
2,50
2005 2006 2007 2008 2009
Cumul cerclé
Cumul non cerclé
0,00
0,50
1,00
1,50
2,00
2,50
2005 2006 2007 2008 2009
rang Nord rang Sud
figures 9 , cumul des poids de récolte au m linéaire sur les rangs nord et sud de 2006 à 2008
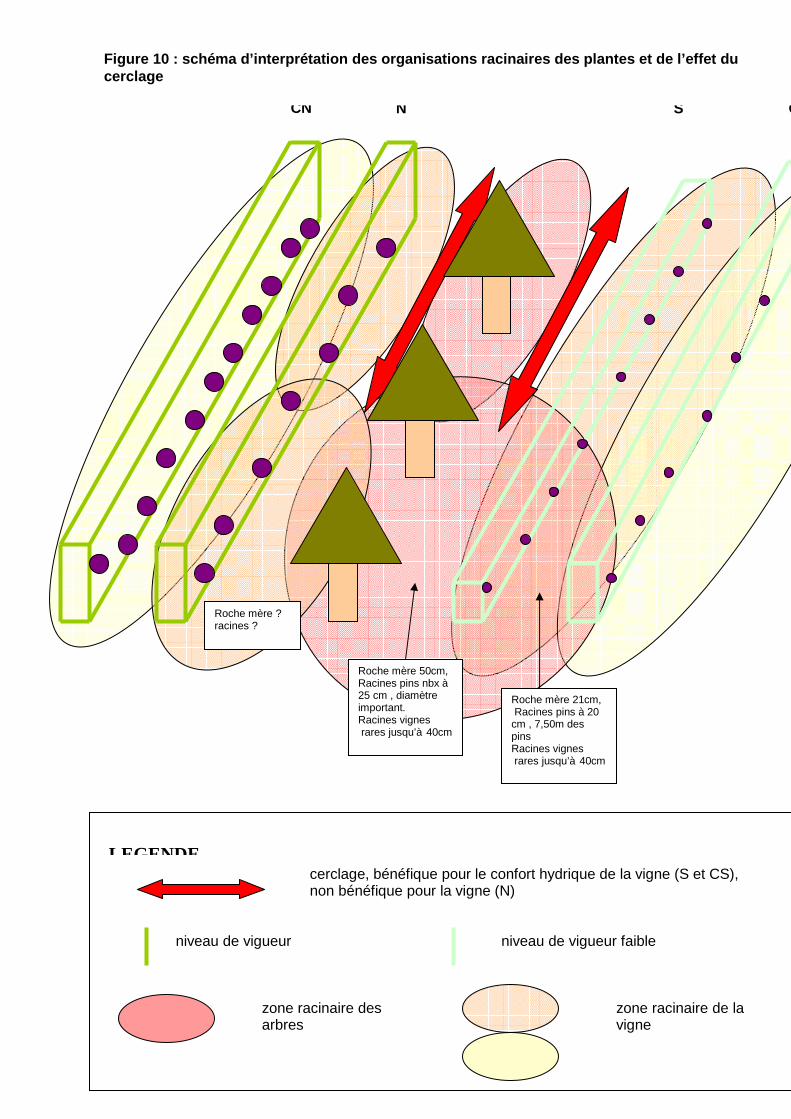
Conclusions La récolte 2008 est légèrement plus faible que celle de 2007. elle est dans la moyenne depuis le suivi de 2004. Les effets du cerclage des arbres par le passage d’une dent de sous-soleuse à 1,75m des arbres et des rangs de vigne les plus proches sont différents d’un côté des arbres à l’autre. Les diverses observations et mesures laissent à penser le schéma suivant d’organisation à la parcelle et les hypothèses des conséquences du cerclage sur les systèmes racinaires. figures 10. Si l’impact racinaire des pins porte non seulement sur l’eau à disposition pour la vigne, mais certains éléments fertilisants voire des relargages toxiques de tanins au sol (l’indicateur pourrait être une absence de flore adventice), nous pourrions expliquer le non redémarrage net des souches de vignes des zones cerclées. Le cerclage pourrait être intervenu trop tard

21
dans la vie de la parcelle et la consommation des éléments fertilisants ou le relargage d’exudats racinaires a peut être été trop important. Une dernière année de suivi des blocs cerclés et non cerclés parait indispensable pour prendre en compte les différentes conditions de millésimes. Des fosses dans le bloc « garrigue » permettraient d’étayer l’hypothèse de la figure 10 et dans des parcelles différentes, la parcelle en pin parasol au dessus par exemple. Cependant, il est possible de donner le schéma de conseils suivants qui seraient affinés après la prochaine campagne : Un système agroforestier sur sol de garrigue ne peut pas être conseillé, même en plantation extensive sans envisager l’ouverture de plusieurs fosses avant plantation. Ces fosses ont pour objectif d’évaluer les contraintes à l’enracinement de la vigne : dalle superficielle, roche mère difficile à pénétrer, horizons compactés … - Si le sol est assez profond sur la majeure partie de la parcelle (exemple B4), alors un
système agroforestier peut être envisagé en faisant attention au choix de l’arbre. L’idée est que l’arbre implanté n’émette pas trop rapidement un système racinaire superficiel important (les pins brutia avec des racines à plus de 7,50m dès les premières années par exemple). L’entretien de la parcelle liée aux arbres pourrait porter sur un cerclage régulier des arbres (si ceux-ci ne sont pas gravement affectés). Ce cerclage imposerait une plantation en extensif.
- Si le sol est superficiel (exemple B10 avec la roche mère entre 21cm et 50 cm de profondeur). Une plantation agroforestière ne peut pas être envisagée.
22
CN N S CS
cerclage, bénéfique pour le confort hydrique de la vigne (S et CS), non bénéfique pour la vigne (N)
niveau de vigueur moyen
niveau de vigueur faible
zone racinaire des arbres
zone racinaire de la vigne
Roche mère ? racines ?
Roche mère 50cm, Racines pins nbx à 25 cm , diamètre important. Racines vignes rares jusqu’à 40cm
Roche mère 21cm, Racines pins à 20 cm , 7,50m des pins Racines vignes rares jusqu’à 40cm
LEGENDE
Figure 10 : schéma d’interprétation des organisations racinai res des plantes et de l’effet du cerclage

23
FARM SAFE, un outil d’aide à la décision en agrofor esterie – Calibrage et premières simulations
Agroof – Développement
Adresse : 120 Impasse des 4 vents, Le poulverel
30140 ANDUZE
Correspondant du projet :
Fabien Liagre
Ingénieurs et techniciens impliqués :
Xavier HAMON (ingénieur)
Tél : 04 66 56 85 47 Email : [email protected]
Introduction L’équipe de développement d’AGROOF a continué sur cette année 2008 de travailler sur le logiciel d’aide à la décision en agroforesterie : Farm sAFe. De plus en plus de projets agroforestiers se montent en France, notamment en Languedoc Roussillon, et le conseil technique demandé par les agriculteurs ne trouvent parfois que peu d’écho au niveau local. Un réel besoin d’appui technique se fait sentir même si la formation de techniciens (administration, instituts techniques, chambre d’agriculture) progresse. Cependant, face à la diversité des aménagements, il est bien difficile de donner des réponses appropriées aux questions des agriculteurs : Comment vont évoluer mes rendements si je plante plus d’arbres ? Combien d’heures de travail supplémentaires la plantation va –t-elle me demander ? Quelle rentabilité a mon projet ? Plusieurs scénarii à tester donc. L’objectif est de mettre à disposition un outil simple et convivial qui permette aux techniciens et in fine aux agriculteurs d’imaginer, tester et prévoir l’aménagement de parcelles agroforestières et leur insertion dans le projet d’exploitation. Après une présentation de l’architecture et des finalités du logiciel nous présenterons les résultats des simulations de différents scénarii basés sur les données technico-économiques du domaine de Restinclières. Nous aborderons finalement l’état de progression du prototype et les voies d’amélioration.
Matériel et méthodes
Présentation du logiciel et de son fonctionnement Le logiciel de prédiction technico-économique est un assemblage de deux modèles créés sur tableurs Excel : FarmSafe et LerSafe. Le principe du logiciel est de permettre à l’utilisateur de prédire les résultats possibles de son projet agroforestier en termes de rendement, temps de travail, trésorerie et rentabilité. Des fonctions avancées permettront d’évaluer la valeur patrimoniale du projet, de tester différents

24
paramètres important par des études de sensibilité et d’affiner le projet par des calculs d’optimisation en fonction de critères technico-économiques. Les résultats seront affichés à trois échelles : l’hectare, la parcelle et l’exploitation. Suite à un manque de temps et de moyen, le module exploitation est encore incomplet. Son analyse a été menée à bien, l’architecture du logiciel est prête mais sa programmation reste encore à finaliser. Comme annoncé en préambule, la principale difficulté du modèle tient à la prédiction des rendements pour la parcelle agroforestière. Pour cela deux sources de données seront possibles :
� Les données générées par Hi-sAFe : dans ce cas, l’utilisateur aura la possibilité d’importer les tables de données des différents scénarii qui seront calculées sur Hi-sAFe, en toute indépendance de fonctionnement avec le logiciel.
� Les données générées par le module LerSafe, intégré au logiciel. Ce sera dans un premier temps la source de données principales pour tout utilisateur non confirmé.
Le logiciel permet de comparer un ou plusieurs scénarios agroforestiers à un scénario agricole sans arbre (scénario d’origine) et à un scénario de boisement pur (type boisement de terre agricole).
25
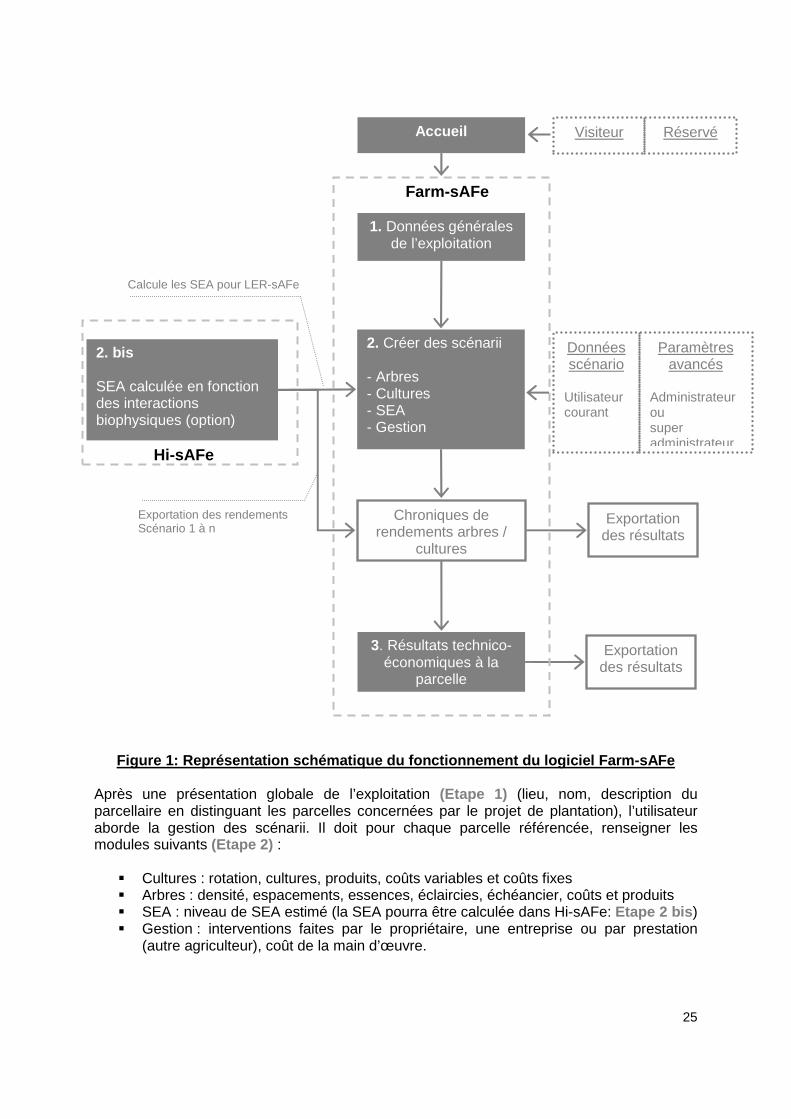
Figure 1: Représentation schématique du fonctionnem ent du logiciel Farm-sAFe
Après une présentation globale de l’exploitation (Etape 1) (lieu, nom, description du parcellaire en distinguant les parcelles concernées par le projet de plantation), l’utilisateur aborde la gestion des scénarii. Il doit pour chaque parcelle référencée, renseigner les modules suivants (Etape 2) :
� Cultures : rotation, cultures, produits, coûts variables et coûts fixes � Arbres : densité, espacements, essences, éclaircies, échéancier, coûts et produits � SEA : niveau de SEA estimé (la SEA pourra être calculée dans Hi-sAFe: Etape 2 bis ) � Gestion : interventions faites par le propriétaire, une entreprise ou par prestation
(autre agriculteur), coût de la main d’œuvre.
Calcule les SEA pour LER-sAFe
2. Créer des scénarii - Arbres - Cultures - SEA - Gestion
1. Données générales de l’exploitation
3. Résultats technico-économiques à la
parcelle
2. bis SEA calculée en fonction des interactions biophysiques (option)
Exportation des rendements Scénario 1 à n
Exportation des résultats
Chroniques de rendements arbres /
cultures
Exportation des résultats
Hi-sAFe
Farm-sAFe
Accueil Visiteur Réservé
Données scénario
Utilisateur courant
Paramètres avancés
Administrateur ou super administrateur
26
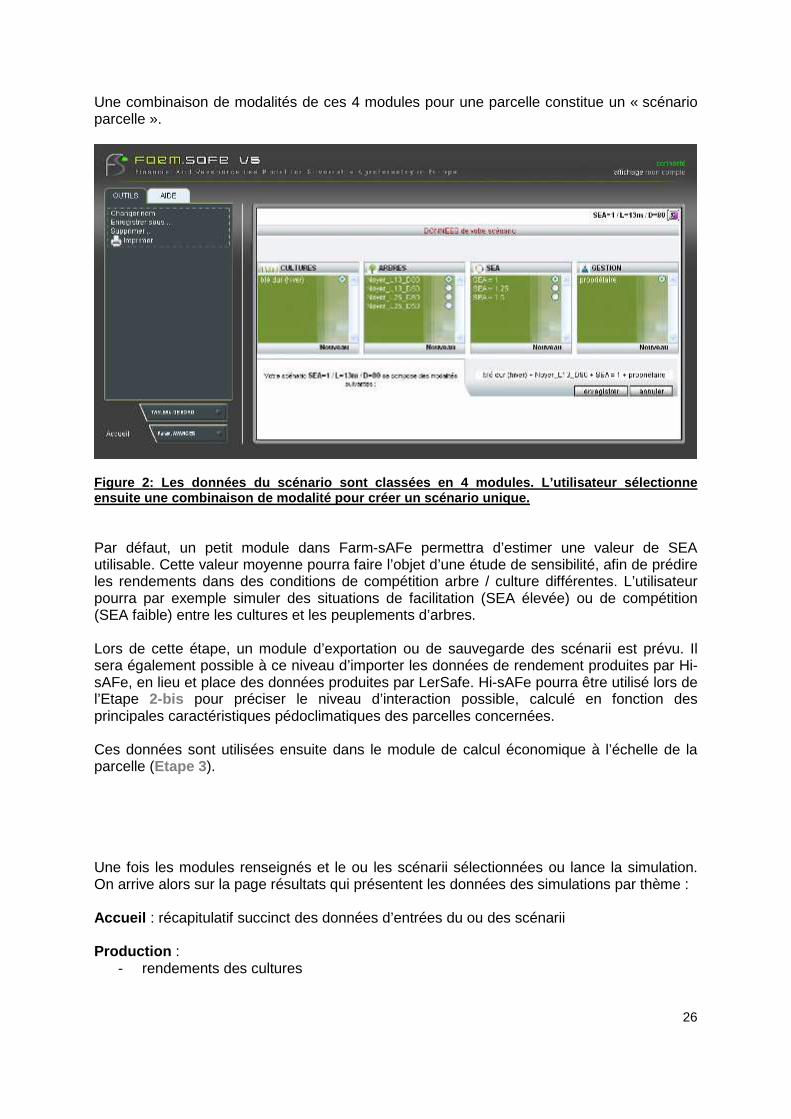
Une combinaison de modalités de ces 4 modules pour une parcelle constitue un « scénario parcelle ».
Figure 2: Les données du scénario sont classées en 4 modules. L’utilisateur sélectionne ensuite une combinaison de modalité pour créer un s cénario unique. Par défaut, un petit module dans Farm-sAFe permettra d’estimer une valeur de SEA utilisable. Cette valeur moyenne pourra faire l’objet d’une étude de sensibilité, afin de prédire les rendements dans des conditions de compétition arbre / culture différentes. L’utilisateur pourra par exemple simuler des situations de facilitation (SEA élevée) ou de compétition (SEA faible) entre les cultures et les peuplements d’arbres. Lors de cette étape, un module d’exportation ou de sauvegarde des scénarii est prévu. Il sera également possible à ce niveau d’importer les données de rendement produites par Hi-sAFe, en lieu et place des données produites par LerSafe. Hi-sAFe pourra être utilisé lors de l’Etape 2-bis pour préciser le niveau d’interaction possible, calculé en fonction des principales caractéristiques pédoclimatiques des parcelles concernées. Ces données sont utilisées ensuite dans le module de calcul économique à l’échelle de la parcelle (Etape 3 ). Une fois les modules renseignés et le ou les scénarii sélectionnées ou lance la simulation. On arrive alors sur la page résultats qui présentent les données des simulations par thème : Accueil : récapitulatif succinct des données d’entrées du ou des scénarii Production :
- rendements des cultures
27
- Production de bois (AF/BTA) par hecatre et par catégorie (bois d’œuvre, industrie,autre)
- Production de bois sur la parcelle - SEA (évolution de la SEA annuelle et de ses composantes)
Temps de travail : AF/BTA
- Plantation - Entretien - Total
Trésorerie :
- Marge détaillée (culture, arbres) - Marge brute absolue et relative - Marge nette absolue et relative - Marge nette cumulée - Valeur Actualisée Nette (VAN) - Valeur d’avenir
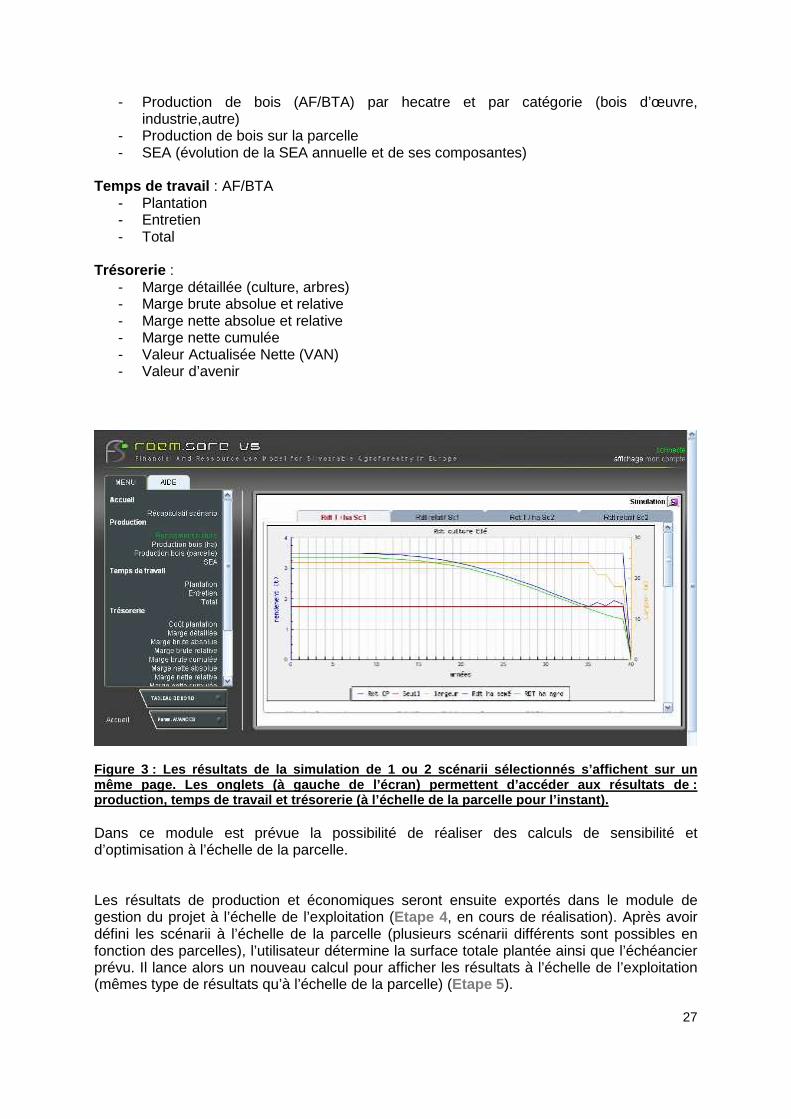
Figure 3 : Les résultats de la simulation de 1 ou 2 scénarii sélectionnés s’affichent sur un même page. Les onglets (à gauche de l’écran) permet tent d’accéder aux résultats de : production, temps de travail et trésorerie (à l’éch elle de la parcelle pour l’instant). Dans ce module est prévue la possibilité de réaliser des calculs de sensibilité et d’optimisation à l’échelle de la parcelle. Les résultats de production et économiques seront ensuite exportés dans le module de gestion du projet à l’échelle de l’exploitation (Etape 4 , en cours de réalisation). Après avoir défini les scénarii à l’échelle de la parcelle (plusieurs scénarii différents sont possibles en fonction des parcelles), l’utilisateur détermine la surface totale plantée ainsi que l’échéancier prévu. Il lance alors un nouveau calcul pour afficher les résultats à l’échelle de l’exploitation (mêmes type de résultats qu’à l’échelle de la parcelle) (Etape 5 ).

28
De même qu’à l’échelle de la parcelle il est prévu de lancer des calculs de sensibilité et d’optimisation technico-économiques.
Exemple – Scénario « Restinclières » Etape 1 : Données générales
� Exploitation
Pays Région Nom SAU France Languedoc-
Roussillon Domaine de
Restinclières 50
� Parcelles : nombre et nom des parcelles. Dans notre cas nous prenons une parcelle de 4 ha de forme rectangulaire (400 x 100 m). Les paramètres pédoclimatiques serviront à renseigner Hi-sAFe pour le calcul de la SEA.
Etape 2 : Paramétrage des modules Les données renseignées dans ces modules sont identiques pour chaque scénario, seuls 3 paramètres sont ici à l’étude : la largeur des allées cultivées, la densité d’arbre s (finale et initiale) ainsi que la SEA (scénario plus ou moins optimiste).
� Cultures La rotation est ici une monoculture de blé d’hiver. Nous avons bien entendu introduit plusieurs type de cultures dans le logiciel mais dont le comportement en système agroforestier est encore mal connu (ex : modélisation d’une culture de printemps type maïs). Dans cette étude nous nous focaliserons sur les cultures d’hiver mais envisagerons le cas
Nom Surface A2 4 ha
Figure 4: Modules qui composent un scénario dans Farm sAFe
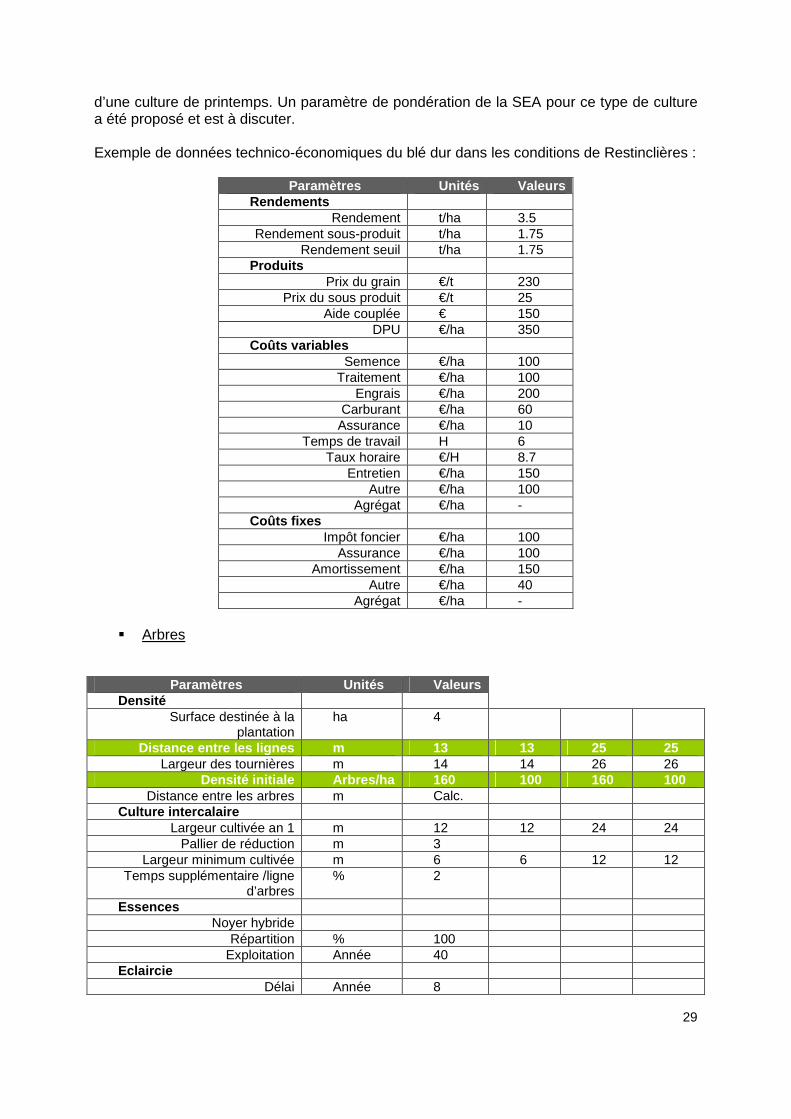
29
d’une culture de printemps. Un paramètre de pondération de la SEA pour ce type de culture a été proposé et est à discuter. Exemple de données technico-économiques du blé dur dans les conditions de Restinclières :
Paramètres Unités Valeurs Rendements
Rendement t/ha 3.5 Rendement sous-produit t/ha 1.75
Rendement seuil t/ha 1.75 Produits
Prix du grain €/t 230 Prix du sous produit €/t 25
Aide couplée € 150 DPU €/ha 350
Coûts variables Semence €/ha 100
Traitement €/ha 100 Engrais €/ha 200
Carburant €/ha 60 Assurance €/ha 10
Temps de travail H 6 Taux horaire €/H 8.7
Entretien €/ha 150 Autre €/ha 100
Agrégat €/ha - Coûts fixes
Impôt foncier €/ha 100 Assurance €/ha 100
Amortissement €/ha 150 Autre €/ha 40
Agrégat €/ha -
� Arbres
Paramètres Unités Valeurs Densité
Surface destinée à la plantation
ha 4
Distance entre les lignes m 13 13 25 25 Largeur des tournières m 14 14 26 26
Densité initiale Arbres/ha 160 100 160 100 Distance entre les arbres m Calc.
Culture intercalaire Largeur cultivée an 1 m 12 12 24 24
Pallier de réduction m 3 Largeur minimum cultivée m 6 6 12 12
Temps supplémentaire /ligne d’arbres
% 2
Essences Noyer hybride
Répartition % 100 Exploitation Année 40
Eclaircie Délai Année 8

30
Nombre d’arbres retirés /ha 80 50 80 50
Echéancier Année plantation An 2008 Surface plantée ha 4
Charges AF Coût unitaire plants (noyer
hybride) €/plant 4.5
Protection €/arbre 1.9 Bourrage €/arbre 0 Paillage €/arbre 0.3
� SEA La SEA est pour l’instant fixée par l’utilisateur. Dans nos calcules nous l’avons fait varier de 1.1 à 1.5 avec des valeurs intermédiaires fines, puisque Farm safe autorise plus de décimales (5) que le tableur Ler qui arrondi à 2 chiffres après la virgule.
� Gestion Une fois les paramètres des modules rentrés, l’utilisateur peut alors combiner différentes modalités pour créer plusieurs scénarii. Ces scénarii sont enregistrés et sont donc mobilisables par l’utilisateur à chaque connexion.
Thèmes d’analyse Etant donné l’état d’avancement du logiciel nous nous concentrerons dans ce rapport sur la crédibilité des résultats du logiciel et l’interprétation de quelques thèmes d’analyse :
� Thème 1 : Comparaison des sorties de Ler et Farm Constat : Farm-safe est plus précis que LER dans les calculs de productions de biomasse. Si les volumes sur pieds sont calculés selon les mêmes formules, le calcul des volumes d’éclaircie est fonction du volume sur pied dans Farm, alors qu’il est fixé par l’utilisateur dans Ler. Objectif : Vérifier la similarité des résultats entre LER-safe et FARM-safe en terme de surfaces relatives agricoles et de chroniques de rendements Méthode : Pour des données d’entrées identiques nous avons calculés à partir des productions de bois la valeur de la surface relative forestière (SRF farm). Nous forçons
Propriétaire Entreprise Prestation Coût horaire main d’œuvre
(€/h) 8.71 - -
Coût horaire machine (€/h) 30 - -

31
ensuite LER-safe à calculer, à partir de cette valeur de SRF, les chroniques de rendements et la surface relative agricole que nous comparons avec celles données par Farm Safe.
� Thème 2 : Recherche de SEA optimum pour gommer l’effet de compensation dû au taux d’occupation initial en parcelle à plus larges espacements
Constat : à densité et SEA égales, on n’obtient des rendements de cultures finaux inférieurs pour des allées larges de 25 m comparées à celles de 13 m. Cet écart vient du fait que le rendement objectif (production agricole sur la durée de la rotation) est le même dans les 2 cas (en effet la SRA ne change pas). Le gain de production en début de rotation (taux d’occupation supérieur en 25m égal à 96%) est alors compensé par une perte plus importante en fin de rotation. Objectif : rechercher par « encadrement » la valeur de SEA optimum pour atteindre dans un scénario à grands espacements (25 m), des rendements finaux plus cohérents avec la réalité. Méthode : Pour un scénario donné (ex : SEA=1.2 / Densité : 80 arbres / Largeur :13m) Nous allons tester pour une largeur de 25m, des valeurs de SEA pour atteindre : - au minimum le rendement final de la culture intercalaire en 13m - de 40 à 50% du rendement initial en 25 m, ce qui correspond à des mesures observées sur parcelles traditionnelles (dire d’expert).
� Thème 3 : Influence de différents coefficients de dominance des arbres sur l’évolution des surfaces relatives
Constat : Les arbres agroforestiers, arbres de pleine lumière en dessous d’une certaine densité (ex : 50 arbres/ha) poussent mieux qu’en ambiance forestière. Dans certains cas on peut observer des arbres 2 fois plus gros que dans le témoin forestier. Objectif : Dans toutes nos simulations, nous avons considéré que les arbres isolés développaient une biomasse 20% plus élevée qu’en situation forestière en dessous du seuil de 50arbres/ha. L’objectif est donc de tester l’impact d’un coefficient de dominance (valeur qui traduit le gain total de biomasse d’un arbre de pleine lumière) plus élevé sur l’évolution des rendements de la culture intercalaire. Méthode : tester un panel de couples Coefficient de dominance / SEA. On prendra pour le coefficient de dominance les valeurs suivantes : 1.2 / 1.4 / 1.6 / 1.8 / 2.0 (densité et largeurs constantes).
Résultats
� Thème 1 : Comparaison des sorties entre Ler et Farm
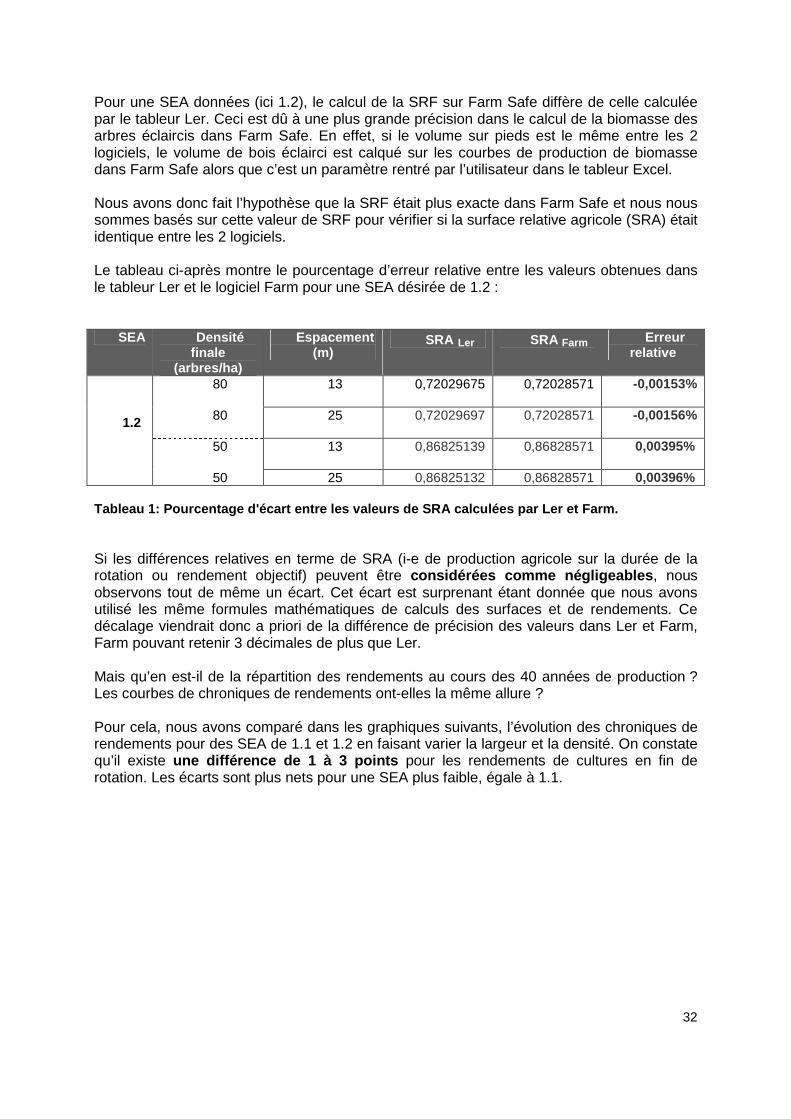
32
Pour une SEA données (ici 1.2), le calcul de la SRF sur Farm Safe diffère de celle calculée par le tableur Ler. Ceci est dû à une plus grande précision dans le calcul de la biomasse des arbres éclaircis dans Farm Safe. En effet, si le volume sur pieds est le même entre les 2 logiciels, le volume de bois éclairci est calqué sur les courbes de production de biomasse dans Farm Safe alors que c’est un paramètre rentré par l’utilisateur dans le tableur Excel. Nous avons donc fait l’hypothèse que la SRF était plus exacte dans Farm Safe et nous nous sommes basés sur cette valeur de SRF pour vérifier si la surface relative agricole (SRA) était identique entre les 2 logiciels. Le tableau ci-après montre le pourcentage d’erreur relative entre les valeurs obtenues dans le tableur Ler et le logiciel Farm pour une SEA désirée de 1.2 :
SEA Densité finale
(arbres/ha)
Espacement (m)
SRA Ler SRA Farm Erreur relative
80 13 0,72029675
0,72028571
-0,00153%
80 25 0,72029697 0,72028571
-0,00156%
50 13 0,86825139 0,86828571 0,00395%
1.2
50 25 0,86825132 0,86828571 0,00396% Tableau 1: Pourcentage d'écart entre les valeurs de SRA calculées par Ler et Farm. Si les différences relatives en terme de SRA (i-e de production agricole sur la durée de la rotation ou rendement objectif) peuvent être considérées comme négligeables , nous observons tout de même un écart. Cet écart est surprenant étant donnée que nous avons utilisé les même formules mathématiques de calculs des surfaces et de rendements. Ce décalage viendrait donc a priori de la différence de précision des valeurs dans Ler et Farm, Farm pouvant retenir 3 décimales de plus que Ler. Mais qu’en est-il de la répartition des rendements au cours des 40 années de production ? Les courbes de chroniques de rendements ont-elles la même allure ? Pour cela, nous avons comparé dans les graphiques suivants, l’évolution des chroniques de rendements pour des SEA de 1.1 et 1.2 en faisant varier la largeur et la densité. On constate qu’il existe une différence de 1 à 3 points pour les rendements de cultures en fin de rotation. Les écarts sont plus nets pour une SEA plus faible, égale à 1.1.
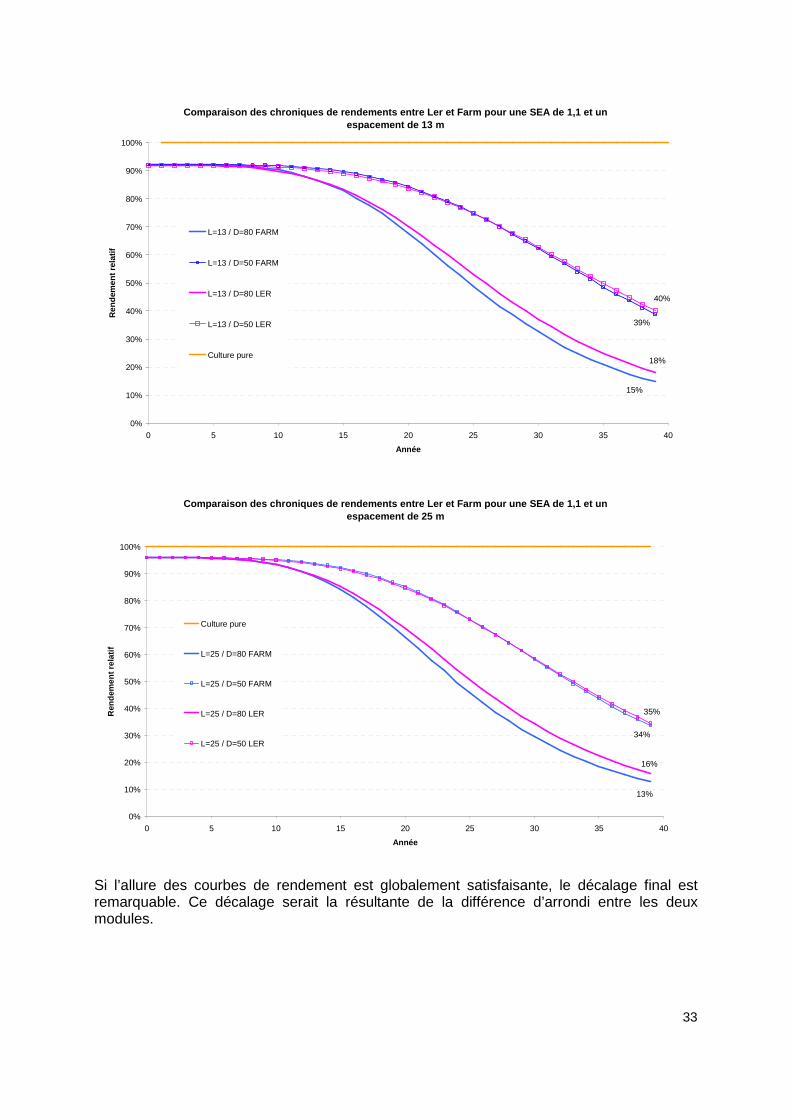
33
Comparaison des chroniques de rendements entre Ler et Farm pour une SEA de 1,1 et un espacement de 13 m
15%
39%
18%
40%
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
0 5 10 15 20 25 30 35 40
Année
Ren
dem
ent r
elat
if
L=13 / D=80 FARM
L=13 / D=50 FARM
L=13 / D=80 LER
L=13 / D=50 LER
Culture pure
Comparaison des chroniques de rendements entre Ler et Farm pour une SEA de 1,1 et un espacement de 25 m
13%
34%
16%
35%
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
0 5 10 15 20 25 30 35 40
Année
Ren
dem
ent r
elat
if
Culture pure
L=25 / D=80 FARM
L=25 / D=50 FARM
L=25 / D=80 LER
L=25 / D=50 LER
Si l’allure des courbes de rendement est globalement satisfaisante, le décalage final est remarquable. Ce décalage serait la résultante de la différence d’arrondi entre les deux modules.
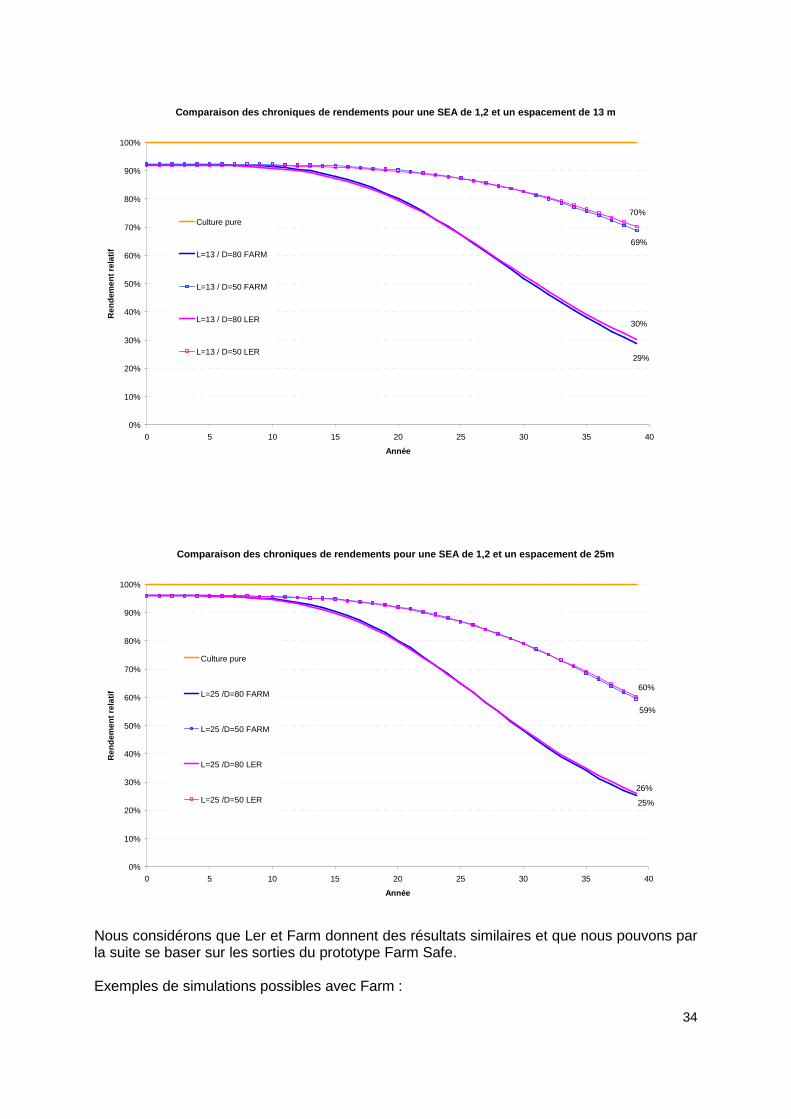
34
Comparaison des chroniques de rendements pour une S EA de 1,2 et un espacement de 13 m
29%
69%
30%
70%
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
0 5 10 15 20 25 30 35 40
Année
Ren
dem
ent r
elat
if
Culture pure
L=13 / D=80 FARM
L=13 / D=50 FARM
L=13 / D=80 LER
L=13 / D=50 LER
Comparaison des chroniques de rendements pour une S EA de 1,2 et un espacement de 25m
25%
59%
26%
60%
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
0 5 10 15 20 25 30 35 40
Année
Ren
dem
ent r
elat
if
Culture pure
L=25 /D=80 FARM
L=25 /D=50 FARM
L=25 /D=80 LER
L=25 /D=50 LER
Nous considérons que Ler et Farm donnent des résultats similaires et que nous pouvons par la suite se baser sur les sorties du prototype Farm Safe. Exemples de simulations possibles avec Farm :
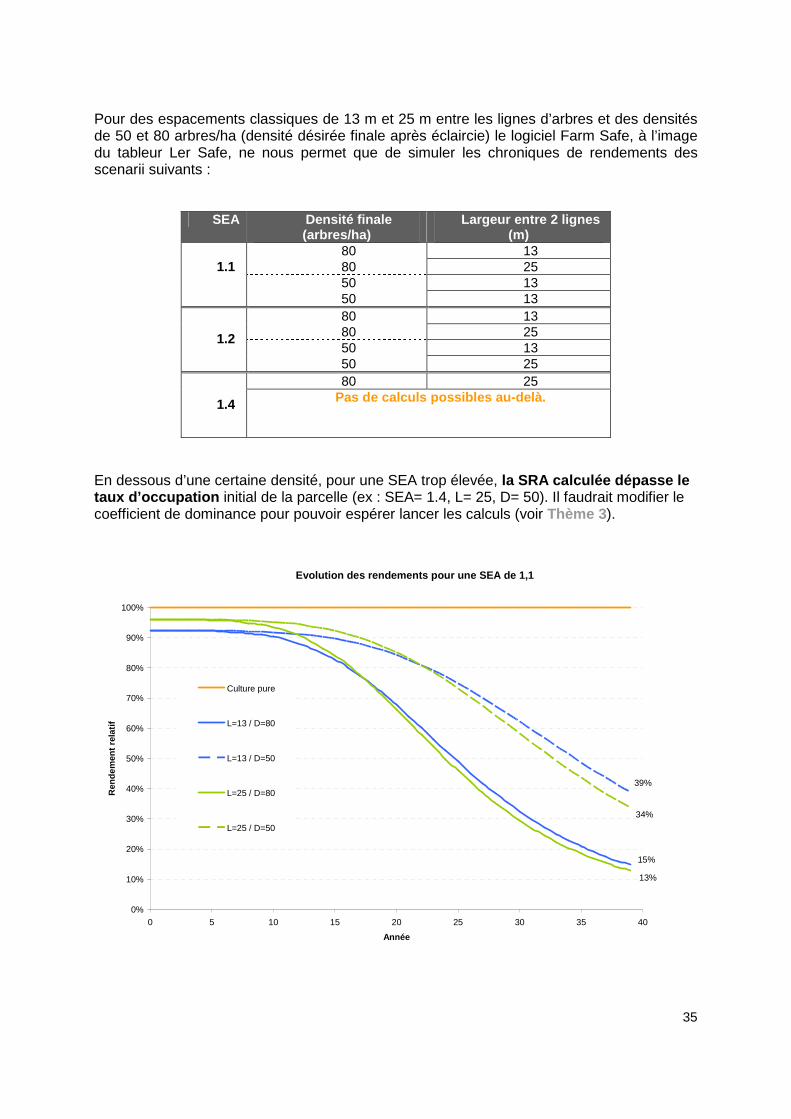
35
Pour des espacements classiques de 13 m et 25 m entre les lignes d’arbres et des densités de 50 et 80 arbres/ha (densité désirée finale après éclaircie) le logiciel Farm Safe, à l’image du tableur Ler Safe, ne nous permet que de simuler les chroniques de rendements des scenarii suivants :
SEA Densité finale (arbres/ha)
Largeur entre 2 lignes (m)
80 13 80 25 50 13
1.1
50 13 80 13 80 25 50 13
1.2
50 25 80 25
1.4 Pas de calculs possibles au-delà.
En dessous d’une certaine densité, pour une SEA trop élevée, la SRA calculée dépasse le taux d’occupation initial de la parcelle (ex : SEA= 1.4, L= 25, D= 50). Il faudrait modifier le coefficient de dominance pour pouvoir espérer lancer les calculs (voir Thème 3).
Evolution des rendements pour une SEA de 1,1
15%
39%
13%
34%
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
0 5 10 15 20 25 30 35 40
Année
Ren
dem
ent r
elat
if
Culture pure
L=13 / D=80
L=13 / D=50
L=25 / D=80
L=25 / D=50
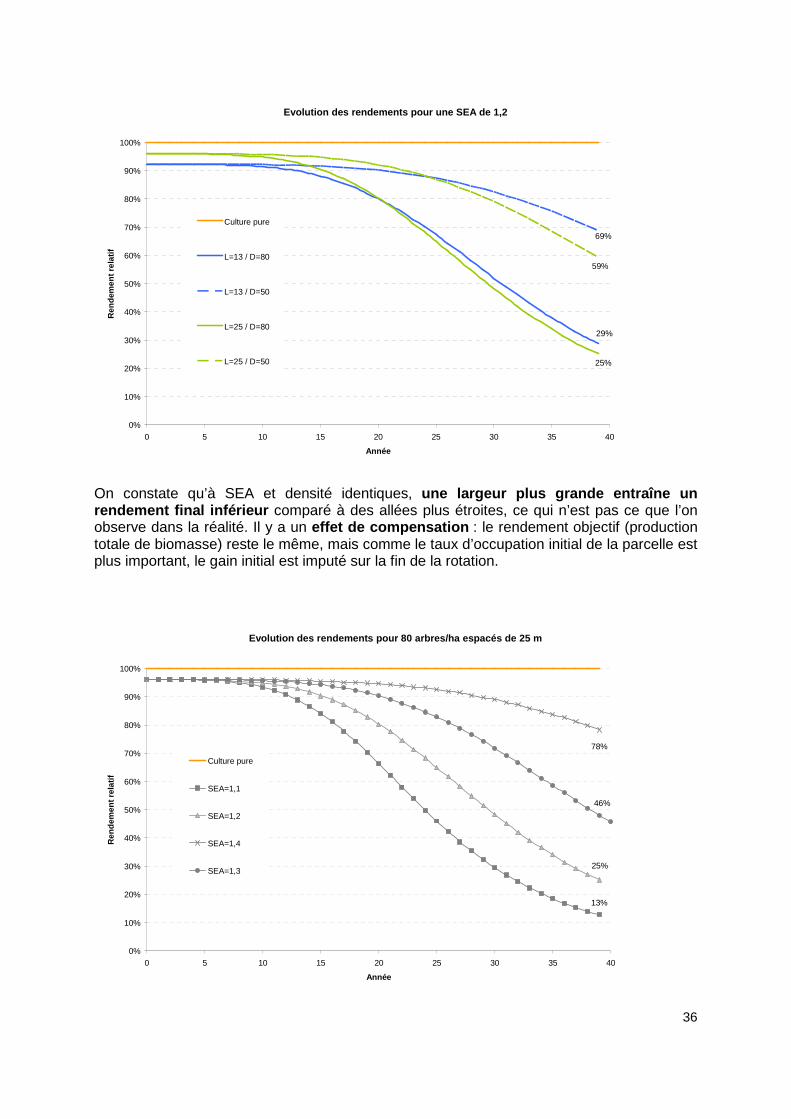
36
Evolution des rendements pour une SEA de 1,2
29%
69%
25%
59%
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
0 5 10 15 20 25 30 35 40
Année
Ren
dem
ent r
elat
if
Culture pure
L=13 / D=80
L=13 / D=50
L=25 / D=80
L=25 / D=50
On constate qu’à SEA et densité identiques, une largeur plus grande entraîne un rendement final inférieur comparé à des allées plus étroites, ce qui n’est pas ce que l’on observe dans la réalité. Il y a un effet de compensation : le rendement objectif (production totale de biomasse) reste le même, mais comme le taux d’occupation initial de la parcelle est plus important, le gain initial est imputé sur la fin de la rotation.
Evolution des rendements pour 80 arbres/ha espacés de 25 m
13%
25%
78%
46%
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
0 5 10 15 20 25 30 35 40
Année
Ren
dem
ent r
elat
if
Culture pure
SEA=1,1
SEA=1,2
SEA=1,4
SEA=1,3
37
A Restinclières la SEA prédite est de 1.6. Avec des largeurs de 13m et une densité de 80 arbres/ha, Farm ne peut pas calculer ce scénario pour un coefficient de dominance ici fixé à 1.2. Il faudrait relever la SRF pour pouvoir que la SRA retombe en dessous du taux d’occupation initial. (ex : SRF=0.8 et SRA=0.8).
� Thème 2 : Recherche de SEA optimum pour gommer l’ef fet de compensation dû au taux d’occupation initial en par celle à plus larges espacements
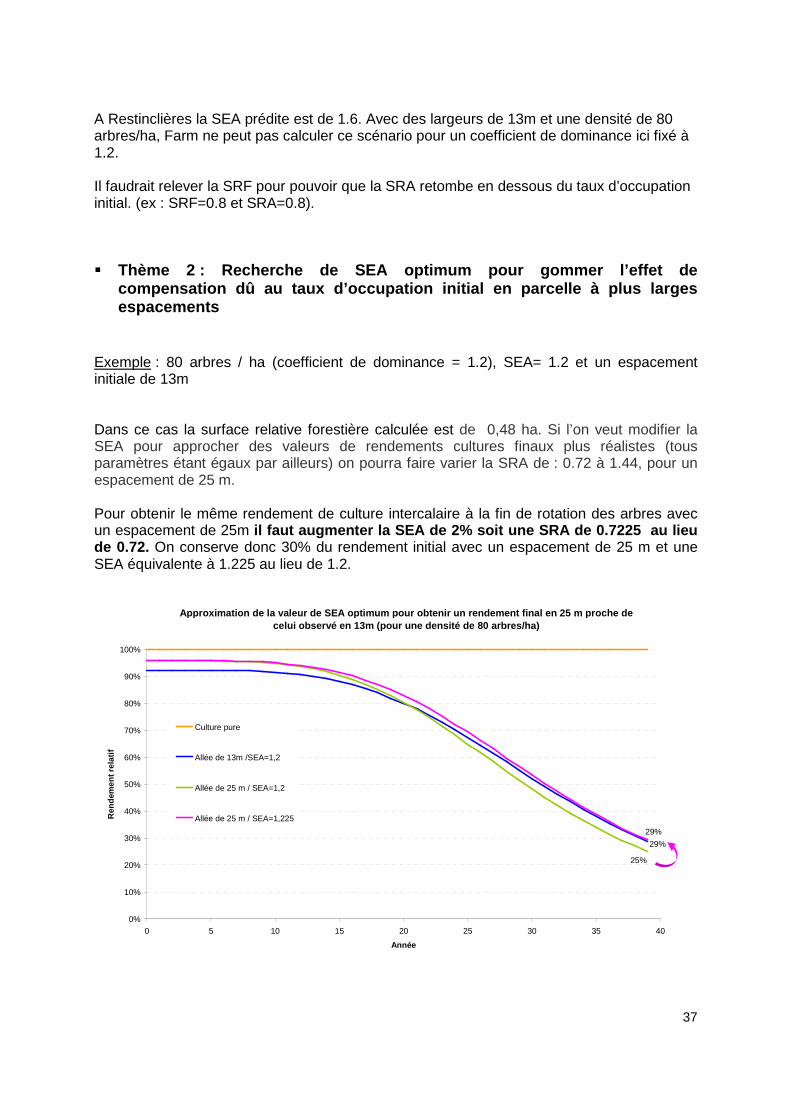
Exemple : 80 arbres / ha (coefficient de dominance = 1.2), SEA= 1.2 et un espacement initiale de 13m Dans ce cas la surface relative forestière calculée est de 0,48 ha. Si l’on veut modifier la SEA pour approcher des valeurs de rendements cultures finaux plus réalistes (tous paramètres étant égaux par ailleurs) on pourra faire varier la SRA de : 0.72 à 1.44, pour un espacement de 25 m. Pour obtenir le même rendement de culture intercalaire à la fin de rotation des arbres avec un espacement de 25m il faut augmenter la SEA de 2% soit une SRA de 0.72 25 au lieu de 0.72. On conserve donc 30% du rendement initial avec un espacement de 25 m et une SEA équivalente à 1.225 au lieu de 1.2.
Approximation de la valeur de SEA optimum pour obte nir un rendement final en 25 m proche de celui observé en 13m (pour une densité de 80 arbres /ha)
29%
25%
29%
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
0 5 10 15 20 25 30 35 40
Année
Ren
dem
ent r
elat
if
Culture pure
Allée de 13m /SEA=1,2
Allée de 25 m / SEA=1,2
Allée de 25 m / SEA=1,225
38
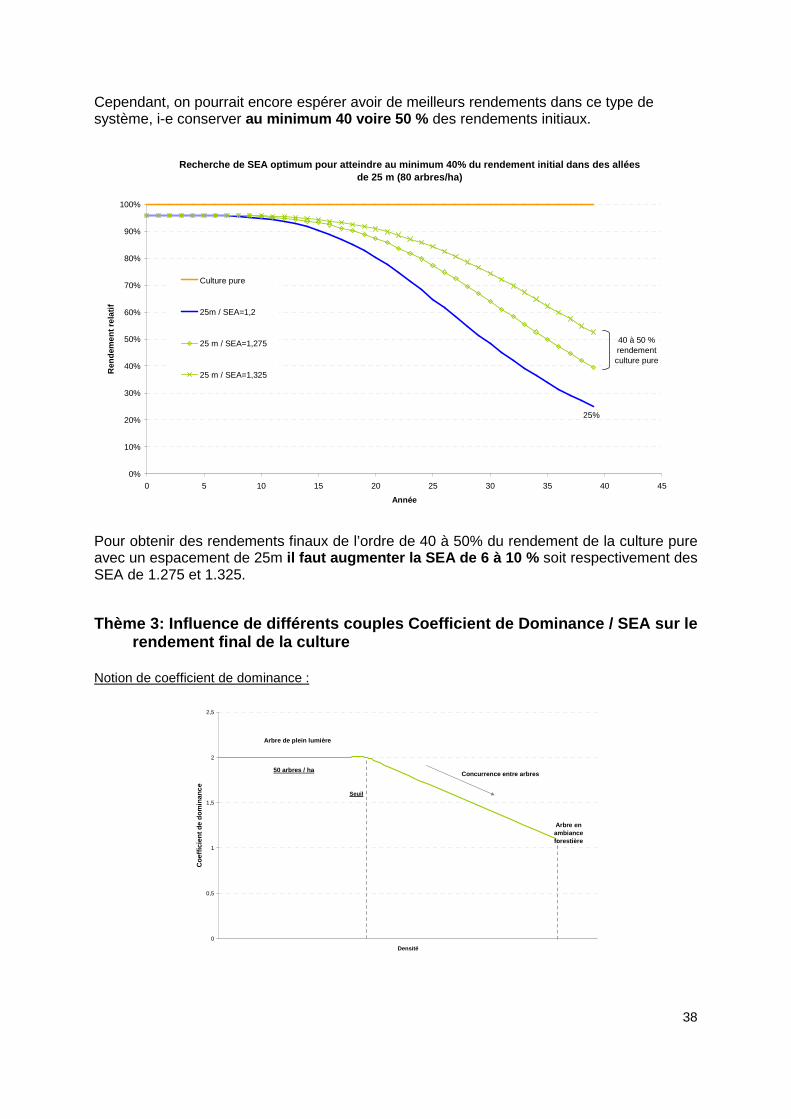
Cependant, on pourrait encore espérer avoir de meilleurs rendements dans ce type de système, i-e conserver au minimum 40 voire 50 % des rendements initiaux.
Recherche de SEA optimum pour atteindre au minimum 40% du rendement initial dans des allées de 25 m (80 arbres/ha)
25%
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
0 5 10 15 20 25 30 35 40 45
Année
Ren
dem
ent r
elat
if
Culture pure
25m / SEA=1,2
25 m / SEA=1,275
25 m / SEA=1,325
40 à 50 %rendement culture pure
Pour obtenir des rendements finaux de l’ordre de 40 à 50% du rendement de la culture pure avec un espacement de 25m il faut augmenter la SEA de 6 à 10 % soit respectivement des SEA de 1.275 et 1.325.
Thème 3: Influence de différents couples Coefficien t de Dominance / SEA sur le rendement final de la culture
Notion de coefficient de dominance :
0
0,5
1
1,5
2
2,5
Densité
Coe
ffici
ent d
e do
min
ance
Arbre de plein lumière
Concurrence entre arbres
Arbre en ambiance forestière
50 arbres / ha
Seuil
39
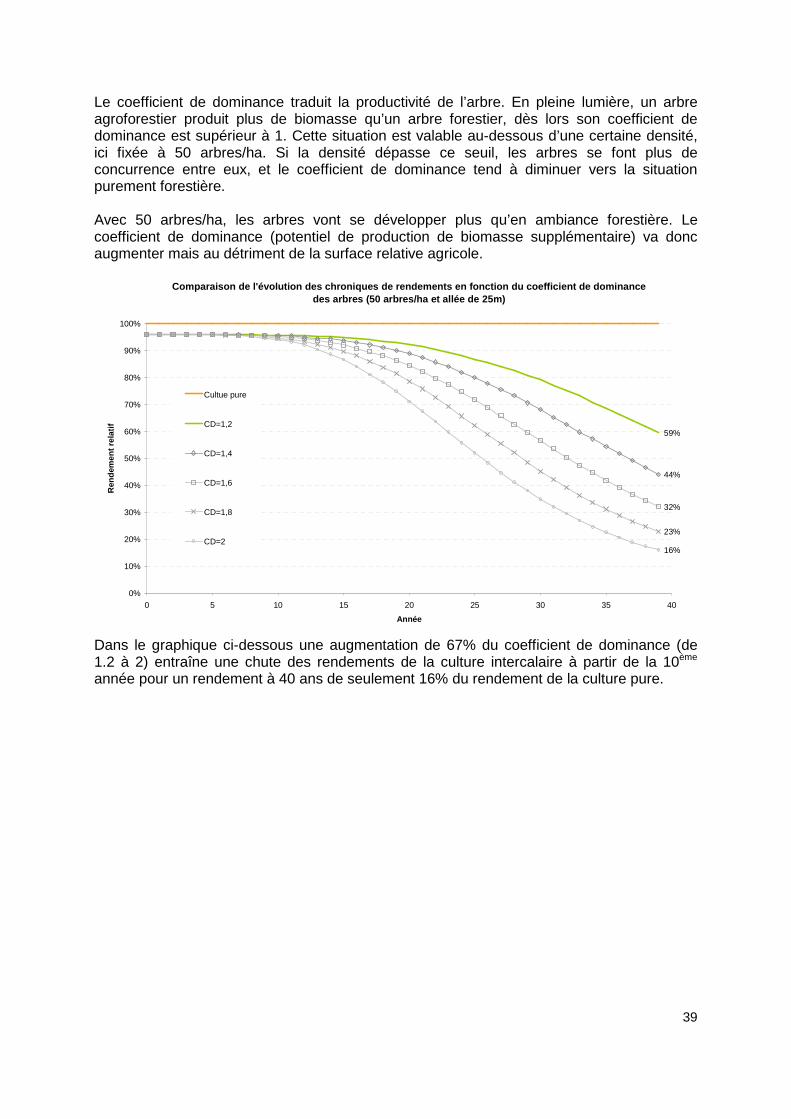
Le coefficient de dominance traduit la productivité de l’arbre. En pleine lumière, un arbre agroforestier produit plus de biomasse qu’un arbre forestier, dès lors son coefficient de dominance est supérieur à 1. Cette situation est valable au-dessous d’une certaine densité, ici fixée à 50 arbres/ha. Si la densité dépasse ce seuil, les arbres se font plus de concurrence entre eux, et le coefficient de dominance tend à diminuer vers la situation purement forestière. Avec 50 arbres/ha, les arbres vont se développer plus qu’en ambiance forestière. Le coefficient de dominance (potentiel de production de biomasse supplémentaire) va donc augmenter mais au détriment de la surface relative agricole.
Comparaison de l'évolution des chroniques de rendem ents en fonction du coefficient de dominance des arbres (50 arbres/ha et allée de 25m)
59%
44%
32%
23%
16%
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
0 5 10 15 20 25 30 35 40
Année
Ren
dem
ent r
elat
if
Cultue pure
CD=1,2
CD=1,4
CD=1,6
CD=1,8
CD=2
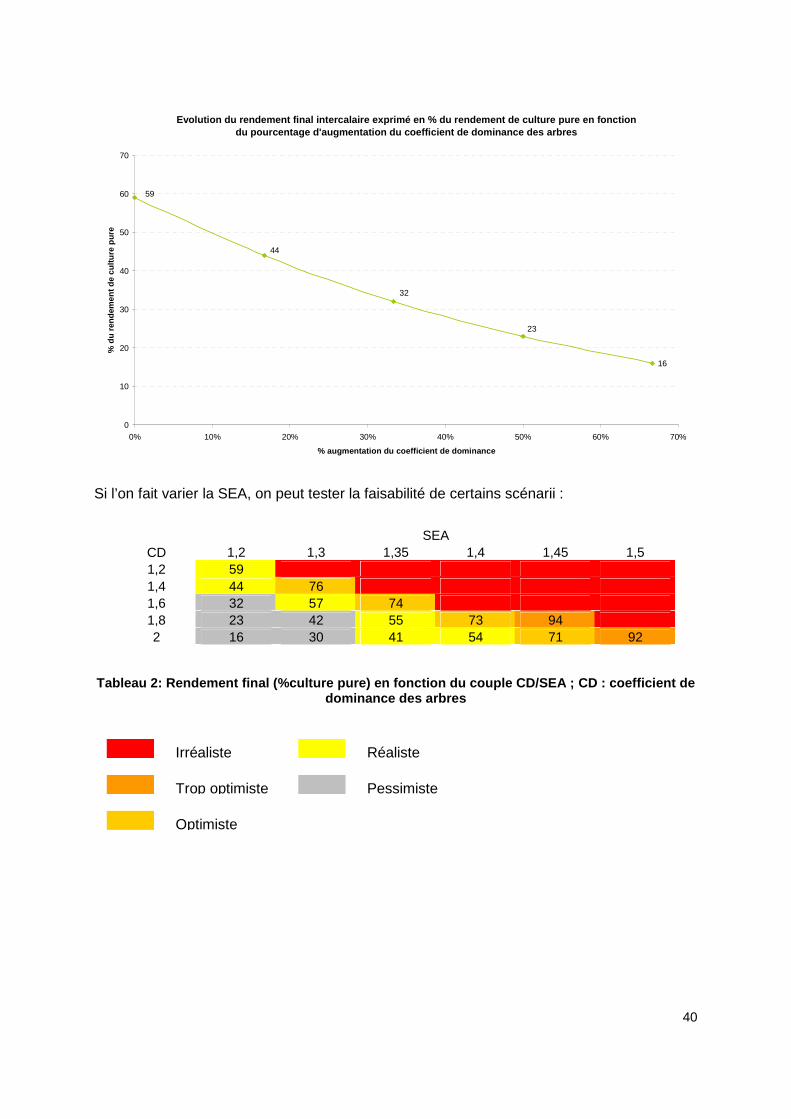
Dans le graphique ci-dessous une augmentation de 67% du coefficient de dominance (de 1.2 à 2) entraîne une chute des rendements de la culture intercalaire à partir de la 10ème année pour un rendement à 40 ans de seulement 16% du rendement de la culture pure.
40
Irréaliste
Trop optimiste
Optimiste
Réaliste
Pessimiste
Evolution du rendement final intercalaire exprimé e n % du rendement de culture pure en fonction du pourcentage d'augmentation du coefficient de dom inance des arbres
16
23
32
44
59
0
10
20
30
40
50
60
70
0% 10% 20% 30% 40% 50% 60% 70%
% augmentation du coefficient de dominance
% d
u re
ndem
ent d
e cu
lture
pur
e
Si l’on fait varier la SEA, on peut tester la faisabilité de certains scénarii :
Tableau 2: Rendement final (%culture pure) en fonct ion du couple CD/SEA ; CD : coefficient de dominance des arbres
SEA CD 1,2 1,3 1,35 1,4 1,45 1,5 1,2 59 1,4 44 76 1,6 32 57 74 1,8 23 42 55 73 94 2 16 30 41 54 71 92

41
Les cases en rouges représentent les scénarii irréalistes. Dans ce cas le logiciel ne peut pas calculer les chroniques de rendement soit parce que la SRF et donc le coefficient de dominance n’est pas assez élevé. Compte tenu de la SRF fixée, la SRA calculée dépasse le taux d’occupation de la parcelle. Les scénarii « trop optimistes » ont pu être calculés par le logiciel mais ne sont pas plausibles dans la réalité. Une fonction de sécurité (contrôle, avertissement) sera à installer dans une prochaine version, pour prévenir l’utilisateur. Les scénarii « optimiste » et « réaliste » représentent des associations où la culture intercalaire est rentable jusqu’en fin de rotation des arbres avec des rendements finaux situés entre 40 et 75 % du rendement de la culture pure. Enfin, les cas « pessimiste » ont généralement une trop forte SRF pour une faible SEA. La surface relative forestière augmente vite et au détriment des cultures. Les rendements de la culture en fin de rotation sont généralement inférieurs à 40%.
Conclusion - Discussion La programmation PHP du logiciel à partir du tableur Excel donne des résultats satisfaisants sur les thèmes d’analyse présentés. Le logiciel présente l’intérêt d’être beaucoup plus rapide en temps de calculs que le tableur et possède la particularité de gérer et enregistrer des scénarii. L’interface est conviviale et il reste encore à concevoir le texte de l’aide pour un utilisateur non-initié (structure de l’aide déjà en place). Cependant, nous n’avons pas pu présenter dans ce rapport l’évaluation du temps de travail et la rentabilité des différents scénarii. Cette partie reste encore à vérifier et/ou à corriger. De plus des modifications restent encore à apporter sur la mise en forme des résultats et leur possible exportation sous fichier excel. Le prototype doit être tester prochainement pour vérifier le fonctionnement de certains modules :
- Tester la pondération affectée à la SEA en présence de cultures de printemps dans la rotation des cultures
- Tester l’impact d’un mélange d’essences sur les rendements de la culture - Evaluer et comparer les temps de travail en fonction de différentes modalités de
gestion de la parcelle (ligne, pieds des arbres).
- Calculer la rentabilité d’un projet agroforestier à travers le calcul de la marge brute et de la VAN de différents scénarii.
Les modules complémentaires à élaborer :
- Module exploitation (Etape 5) - Mise en ligne et gestion des mots de passe - Câbler les pages d’accueil (présentation de l’agroforesterie, diaporama) - Réaliser la programmation de la base de données actuellement sous format Access.

42
Suivi phytosanitaire des arbres dans les parcelles agroforestières expérimentales du Domaine de Restinclières
CRPF Languedoc Roussillon
Adresse : 378, rue de la Galéra, BP4228
34097 MONTPELLIER CEDEX 5
Correspondant du projet :
Michèle LAGACHERIE
Ingénieurs et techniciens impliqués :
Tél : 04 67 41 68 10 Mail : [email protected]
Campagne de mesures 2008 Le but de cette campagne de mesures est de suivre l’évolution des essences installées, au niveau des risques et évènements sanitaires . Cette proposition vient d’un besoin devenu évident vu le comportement de certaines essences qui ont dépéri en nombre ces dernières années (à ce sujet, le cas de l’aulne de Corse est emblématique). S’il y a un intérêt pour ces mesures sur une campagne d’un an, il faut souligner l’intérêt encore plus grand de réaliser ce type de suivi de manière régulière et sur plusieurs années. En effet, il est parfois difficile de juger de l’importance d’un problème sanitaire sans prendre en compte l’évolution de ce problème sur le temps (cas des dépérissements sur plusieurs années). Dans un premier temps, un protocole d’observation spécifique à cette campagne est décrit, fonction des contraintes de cette campagne : mesures effectuées en été, sélection d’un sous-ensemble d’essences parmi toutes celles qui ont été plantées. Ensuite, pour ce sous-ensemble d’essences, les symptômes rencontrés sont listés, et pour chacun, une identification possible est proposée. Un tableau récapitule alors pour chaque essence le ou les symptômes rencontrés, immédiatement suivi d’une synthèse sur cette campagne. En annexe sont proposés d’abord une description synthétique des ravageurs et pathogènes rencontrés ou suspectés, puis une courte bibliographie. Note : Les références entre crochets [] correspondent à la bibliographie donnée en fin de document.
Protocole d’observation sanitaire Pour les raisons évoquées, nous nous sommes limités pour cette campagne de mesures, au diagnostic de terrain : symptômes observables et si possible identifiables sans analyse plus précise (microscope, analyse chimique). Les essences suivantes ont été observées :
� Noyer hybride

43
� Érable plane
� Cormier
� Aulne de Corse
� Merisier
� Poirier commun Comme proposé dans [1], l’observation est limitée à trois catégories de symptômes , méthode permettant l’exhaustivité des relevés, adaptée au diagnostic mis en place cette année :
� Sur feuilles :
- Densité de feuillage (chute prématurée, éclaircissement partiel…)
- Couleur suspecte ou décoloration suspecte, dépérissement (dessèchement)
- Flétrissement
- Taille et forme
- Lésions – trous
- Fructifications de champignons, dépôts divers
� Sur bourgeons, pousses et rameaux : Note : l’observation sur bourgeons et sur pousses n’a pas été possible du fait de l’époque tardive des relevés, et devrait donc être réalisée dans un second temps, à la bonne époque (hiver, printemps).
- Décoloration jaunâtre, brunissement
- Dépérissement, flétrissement
- Anomalies de croissance, déformations
- Lésions
- Fructifications de champignons, dépôts divers
� Sur tronc et branches :
- Chute
- Décoloration jaunâtre
- Flétrissement
- Signes de dépérissement (branches, nécrose de l’écorce, écoulements)
- Anomalies de croissance, déformations
- Lésions (blessures)
- Fructifications de champignons, dépôts divers

44
Observations et causes Plutôt que de donner les observations parcelle par parcelle, les observations sont listées par essence, ce qui permet de comparer le comportement de chacune d’entre elles suivant les conditions dans lesquelles elle a été installée et observée (témoin agricole, forestier, etc...). Il n’est pas toujours possible, suivant le stade de développement des maladies ou attaques, de déceler la ou les causes initiales, ce qui souligne l’intérêt de réaliser un tel suivi de manière plus régulière . Note : chaque ravageur ou pathogène dont il est question dans ce chapitre, et qui est annoté d’un astérisque, est référencé dans le chapitre « Ravageurs et pathogènes ».
Noyer hybride Note : seule la parcelle A2 - Agroforesterie grandes cultures a été parcourue, qui propose un ensemble de conditions diverses pour cette essence, y compris un témoin forestier.
� Symptôme : Coulure de tanins Une tige du témoin agricole avec une cicatrice d’élagage tardif (grosse branche)
présentant une coulure de tanin. => Présence possible d’un xylophage type zeuzère*, à redouter : À surveiller en
2009 (apparition de trous de sortie avec sciure).
Photo 1 - Noyer hybride : coulure de tanins

45
� Symptôme : Branches sèches Seulement 2 arbres sur les 42 du témoin forestier ont une ou deux branches sèches. => Origine inconnue. À surveiller en 2009.
� Symptôme : Feuilles jaunissantes et de taille rédui te Un très petit nombre de tiges sont concernées – 2, uniquement dans le témoin
agricole. Le problème se situe sur les feuilles près du tronc et à l’intérieur du houppier. => Il s’agit d’un problème de type pathologique : possible anthracnose du noyer
(Gnomonia sp.*). À confirmer.
Photo 2 - Noyer hybride : feuilles jaunissantes et de taille réduite
Commentaire sur les noyers hybrides
L’élagage et la taille de formation étaient en cours au moment des mesures (août 2008)
dans le témoin agricole. Une intervention visant à dessoucher les souches des arbres

46
enlevés en éclaircis était programmée, afin d’éviter d’avoir à traiter les rejets (parfois
vigoureux).
Quant au témoin forestier, rappelons qu’aucune éclaircie n’a encore été réalisée jusqu’au
moment des dernières mesures (août 2008). Une éclaircie pourrait toutefois être
envisagée dans les 2 ans, car les houppiers des beaux sujets se touchent déjà. Notons
que le taux de survie reste très élevé (plus de 80%) dans cette partie.
Érable plane � Symptôme : Écorce se craquelant au pied du tronc
Quelques érables ont leur écorce qui se détache, craquelle, à la base du tronc, sur une
hauteur de quelques dizaines de centimètres, toujours avec présence d’une malformation
(blessure / élagage).
=> L’origine est très probablement abiotique : cicatrice suite à une blessure (cf photo).
Photo 3 – Érable plane : écorce se craquelant au pi ed du tronc

47
Aulne de Corse � Symptôme : Écorce se craquelant d’abord au pied pui s sur le tronc
Un très fort pourcentage – plus de 90% – des aulnes vivants présentent ce problème (plus ou moins important) dans les parcelles agro-forestières .
Par contre, le témoin forestier (parcelle B2) affiche plus de 90% d’aulnes qui n’ont pas ce problème d’écorce se craquelant , les sujets restants ayant au moins la cime sèche et l’écorce du tronc craquelée (possiblement pour une tout autre raison).
Ce qui différencie les deux témoins est d’une part leur situation (soleil direct, culture en parcelle agro-forestière), et d’autre part un élagage déjà effectué (pas encore dans le témoin forestier, parcelle B2).
=> L’origine est probablement abiotique : coup de s oleil notamment , station, ambiance…
Photo 4 – Aulne de Corse : écorce se craquelant d’a bord au pied puis
sur le tronc

48
� Symptôme : Écorce se craquelant d’abord au pied pui s sur le tronc, avec présence de carpophores Les craquelures et fissures, par rapport au symptôme précédent, sont accentuées,
l’écorce se détachant quasiment complètement par endroits. Le carpophore est celui du champignon Schizophyllum commune* : c’est un champignon de blessure qui entraîne une pourriture blanche fibreuse : le facteur favorable d’installation est « le coup de soleil » sur hêtre et fruitiers en forêt, c’est un pathogène connu en arboriculture .
=> Comme précédemment, l’origine est probablement a biotique : coup de soleil , mais dans un stade d’évolution plus avancé.
Photo 5 – Aulne de Corse : écorce se craquelant d’a bord au pied puis sur le tronc, avec
apparition de carpophores
� Symptôme : Galles (érinoses) sur feuilles Bombements sur la face supérieure, feuilles déformées. Une seule tige a été
observée avec ce symptôme, dans une parcelle agro-forestière avec céréales (parcelle A5). => Attaque d’acariens qui forment des galles (érinoses), problème commun sur
érable et tilleul : l’espèce est vraisemblablement Acalitus brevitarsus* (la galle des plaques feutrées de l’aulne).

49
Photo 6 – Aulne de Corse : galles (érinoses) sur fe uilles
Cormier � Symptôme : Écorce se craquelant au pied du tronc
Que ce soit en parcelle agro-forestière ou en témoin forestier, la quasi-totalité des cormiers vivants présentent ce symptôme (plus ou moins important), dont la hauteur peut varier de 10cm à 1m.

50
=> Il s’agit d’un caractère naturel de l’essence, l ’écorce restant lisse jusqu’aux environs de 7 années [1], et pas d’un problème abiotique ou qui soit lié à la protection contre le gibier.
Photo 7 – Cormier : écorce se craquelant au pied du tronc
Merisier � Symptôme : Écorce se craquelant au pied du tronc ou plus haut, feuillage
affaibli Un peu moins de la moitié des merisiers vivants présentent ce problème (plus ou
moins important) en parcelles agro-forestières à céréales (A6, A8). => L’origine est probablement abiotique : cicatrisation suite à blessure ou élagage
« fort », ou encore coup de soleils. => Un dépérissement bactérien à Pseudomonas * est peu probable mais reste
tout à fait possible (à confirmer avec une analyse plus précise), du fait des importantes cicatrices suite à élagage [1].

51
Photo 8 – Merisier : écorce se craquelant au pied d u tronc ou plus haut, feuillage
affaibli
� Symptôme : Feuilles se crispant et qui « brûlent » Les feuilles se recroquevillent, crispées en cuiller, en réaction aux piqûres de
pucerons (toxines), et apparaissent comme des « brûlures » des feuilles. Deux tiges ont été observées avec ce symptôme, dans les parcelles agro-forestières avec céréales (parcelles A6, A8).
=> Attaque de pucerons noirs du merisier : Myzus cerasi*.
Photo 9 – Merisier : Feuilles se crispant et qui « brûlent »

52
Poirier commun � Symptôme : Taches grises à marron sur feuilles
Taches circulaires grises à marron clair, bordées de noir, évoluant en plaques. Quelques tiges ont été observées avec ce symptôme, dans les parcelles agro-forestières avec céréales ou avec vignes (parcelles A6, A8, B10).
=> Attaque de septoriose : Septoria piricola*.
Photo 10 – Poirier commun : Taches grises à marron sur feuilles

53
� Symptôme : Écorce se craquelant au pied du tronc Quelques poiriers communs répartis sur les parcelles (agroforesterie avec céréales ou
vignes) ont leur écorce qui se détache, craquelle, à la base du tronc, sur une hauteur de
quelques dizaines de centimètres au plus.
=> L’origine est très probablement abiotique : coup de soleil. À confirmer / ne pas écarter, avec une analyse plus poussée ultérieure : phytotoxicité évoluant en nécrose chancreuse ou en chancre (avec pathogène ou non).
Photo 11 – Poirier commun : Écorce se craquelant au pied du tronc
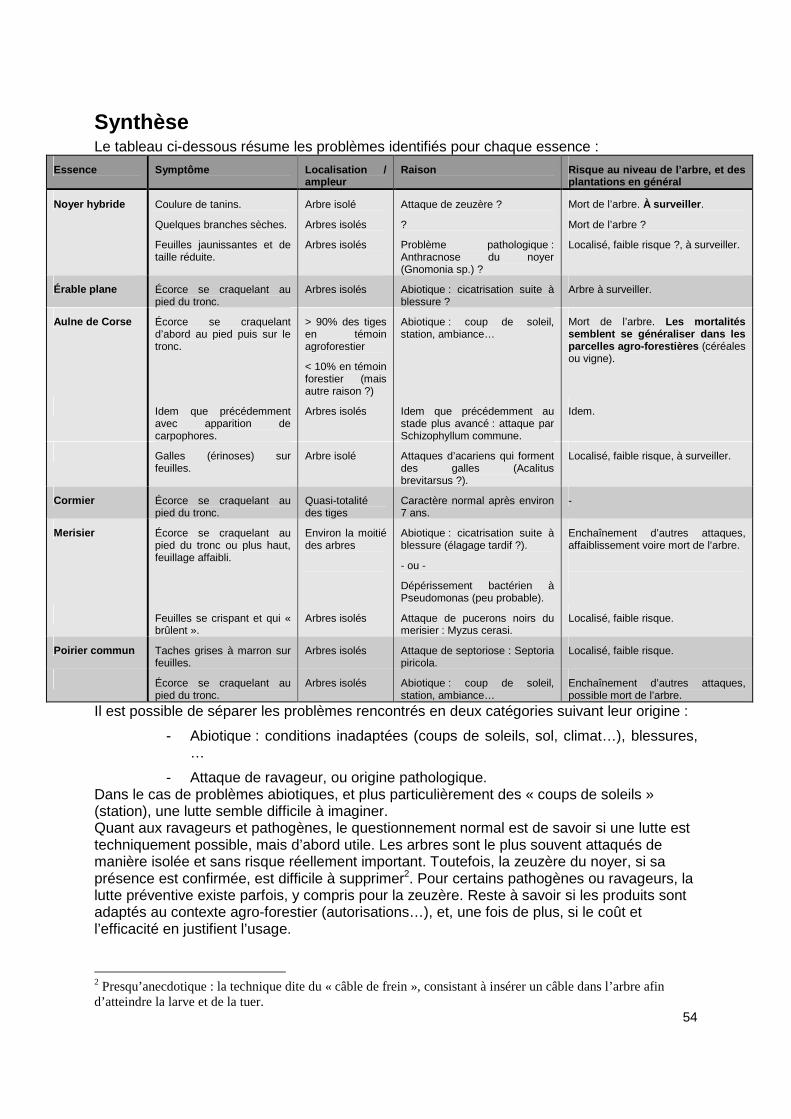
54
Synthèse Le tableau ci-dessous résume les problèmes identifiés pour chaque essence :
Essence Symptôme Localisation / ampleur
Raison Risque au niveau de l’arbre, et des plantations en général
Noyer hybride Coulure de tanins.
Quelques branches sèches.
Feuilles jaunissantes et de taille réduite.
Arbre isolé
Arbres isolés
Arbres isolés
Attaque de zeuzère ?
?
Problème pathologique : Anthracnose du noyer (Gnomonia sp.) ?
Mort de l’arbre. À surveiller .
Mort de l’arbre ?
Localisé, faible risque ?, à surveiller.
Érable plane Écorce se craquelant au pied du tronc.
Arbres isolés Abiotique : cicatrisation suite à blessure ?
Arbre à surveiller.
Aulne de Corse Écorce se craquelant d’abord au pied puis sur le tronc.
> 90% des tiges en témoin agroforestier
< 10% en témoin forestier (mais autre raison ?)
Abiotique : coup de soleil, station, ambiance…
Mort de l’arbre. Les mortalités semblent se généraliser dans les parcelles agro-forestières (céréales ou vigne).
Idem que précédemment avec apparition de carpophores.
Arbres isolés Idem que précédemment au stade plus avancé : attaque par Schizophyllum commune.
Idem.
Galles (érinoses) sur feuilles.
Arbre isolé Attaques d’acariens qui forment des galles (Acalitus brevitarsus ?).
Localisé, faible risque, à surveiller.
Cormier Écorce se craquelant au pied du tronc.
Quasi-totalité des tiges
Caractère normal après environ 7 ans.
-
Merisier Écorce se craquelant au pied du tronc ou plus haut, feuillage affaibli.
Environ la moitié des arbres
Abiotique : cicatrisation suite à blessure (élagage tardif ?).
- ou -
Dépérissement bactérien à Pseudomonas (peu probable).
Enchaînement d’autres attaques, affaiblissement voire mort de l’arbre.
Feuilles se crispant et qui « brûlent ».
Arbres isolés Attaque de pucerons noirs du merisier : Myzus cerasi.
Localisé, faible risque.
Poirier commun Taches grises à marron sur feuilles.
Arbres isolés Attaque de septoriose : Septoria piricola.
Localisé, faible risque.
Écorce se craquelant au pied du tronc.
Arbres isolés Abiotique : coup de soleil, station, ambiance…
Enchaînement d’autres attaques, possible mort de l’arbre.
Il est possible de séparer les problèmes rencontrés en deux catégories suivant leur origine :
- Abiotique : conditions inadaptées (coups de soleils, sol, climat…), blessures, …
- Attaque de ravageur, ou origine pathologique. Dans le cas de problèmes abiotiques, et plus particulièrement des « coups de soleils » (station), une lutte semble difficile à imaginer. Quant aux ravageurs et pathogènes, le questionnement normal est de savoir si une lutte est techniquement possible, mais d’abord utile. Les arbres sont le plus souvent attaqués de manière isolée et sans risque réellement important. Toutefois, la zeuzère du noyer, si sa présence est confirmée, est difficile à supprimer2. Pour certains pathogènes ou ravageurs, la lutte préventive existe parfois, y compris pour la zeuzère. Reste à savoir si les produits sont adaptés au contexte agro-forestier (autorisations…), et, une fois de plus, si le coût et l’efficacité en justifient l’usage.
2 Presqu’anecdotique : la technique dite du « câble de frein », consistant à insérer un câble dans l’arbre afin d’atteindre la larve et de la tuer.

55
Au-delà de l’intérêt de cette première campagne, vue comme un constat de l’état sanitaire des plantations à Restinclières, pour les essences considérées, la nécessité d’un suivi plus régulier se fait sentir , qui devrait permettre, entre autres, de mieux comprendre l’évolution des problèmes rencontrés, et peut-être d’en détecter de nouveaux, de manière précoce.
Ravageurs et pathogènes : risques sur l’arbre Voici un descriptif concis de chaque ravageur ou pathogène identifié (ou supposé) précédemment, et des risques qu’il peut entraîner sur l’arbre.
� Zeuzère (du noyer) : papillon. L’attaque sur le noyer peut provoquer le bris des pousses terminales, ou des galeries importantes sur de grosses sections, pouvant entraîner des ruptures de tige. [3] et [6].
� Pseudomonas mors-prunorum (sur merisier) : bactérie. Entraîne le dépérissement des feuilles et rameaux au-dessus de la partie infectée [1].
� Myzus cerasi (sur le genre Prunus) : puceron de teinte sombre. Dégâts pouvant être important chez de jeunes sujets [4] et [5].
� Schizophyllum commune : champignon, « pourriture blanche ». Ce champignon saprophyte envahit souvent un tronc dont l’écorce a été dégarnie du fait d’un problème abiotique (coup de soleil, …).[1]
� Anthracnose du noyer - Gnomonia leptostyla : champignon. S’attaque aux feuilles et aux fruits, pouvant entraîner une défoliation précoce et un dessèchement des fruits [7].
� Galle des plaques feutrées de l’aulne - Acalitus brevitarsus (aulnes) : galle due à un acarien. Déformation bombée sur la face supérieure de la feuille, et feutrage blanc sur la face intérieure.
� Septoriose - Septoria piricola : champignon. Affecte les feuilles et plus rarement les fruits. L’infection se poursuit jusqu’à la chute des feuilles à l’automne [8].
Bibliographie � [1] Les symptômes de dépérissement des arbres forestiers (HARTMAN, NIENHAUS,
BUTIN), Ed. IDF, 1991.
� [2] Site internet du projet SEBA Favoriser les essences rares par l’Office fédéral de l’environnement des forêts et des paysages de Suisse (OFEFP), fiche Cormier http://www.seba.ethz.ch/lieblinge_f/frame_lieb_f.htm .
� [3] Fiche noyer du CRPF Poitou-Charentes : www.crpf-poitou-charentes.fr/IMG/pdf/noyers .pdf
� [4] Guide des maladies des arbres (parcs et jardins) (NIENHAUS, BUTIN, BÖHMER), Ed. Ulmer, 1996.
� [5] Fiche Myzus cerasi (F.) de l’INRA : http://www.inra.fr/hyppz/RAVAGEUR/3myzcer.htm .
� [6] Fiche Zeuzera pyrina (L.) de l’INRA : http://www.inra.fr/hyppz/RAVAGEUR/3zeupyr.htm .
� [7] Fiche Gnomonia leptostyla (Fr.) de l’INRA : http://www.inra.fr/hyp3/pathogene/3gnolep.htm

56
� [8] Les champignons. Mycologie fondamentale et appliquée, BOUCHET P., GUIGNARD J. L., VILLARD J., Ed. Masson, 1999.
Remerciements L’expertise des correspondants observateurs du Département Santé des Forêts - DSF – de la région Languedoc-Roussillon a été précieuse pour la réalisation de cette campagne, et plus particulièrement celle de M. Bernard BOUTTE, expert DSF à Avignon.





























