Embed Size (px)
Citation preview

BIOCHIMIE, 1978, 60, 203-207.
Synth se et activation de ddrivds N-aminopolyacrylamide comme support dans la ddgradation en phase solide des protdines.
J e a n - C l a u d e CAVADORE.
(22-12-1977).
Unit~ de Recherches INSERM U 1~7, 60, rue de Navacelles, 3-~100 Montpellier.
Summary.
N-amino polyacryi'amide derivatives were syn- thesized and their activation by di[[erent tech- niques were examined. The use o[ these deriva- tives as solid support in sequential degradation of proteins is proposed. An example is reported.
Introduction.
La s t r u c t u r e p r i m a i r e des p r o t 6 i n e s et des pep - t ides est une donn6e de base p o u r des 6 tudes enzy- m o l o g i q u e s , p h a r m a c o l o g i q u e s , i m m u n o l o g i q u e s ; iI en r~sul te que les m ~ t h o d e s qu i c o n v e r g e n t ve rs la d 6 t e r m i n a t i o n d ' u n e s6quence font l ' ob je t d ' u n essai d ' a u t o m a t i s a t i o n de p lus en p lus ponss6. L ' a p p a r i t i o n dans te c o m m e r c e de s 6 q u e n c e u r s a u t o m a t i q u e s en phase l i q u i d e ou en phase so l ide en sont deux exemples . Ces d e u x t e c h n i q u e s 6 ta ient cons id6 r6es i l y a p e u de t e m p s c o m m e c o m p l 6 m e n t a i r e s : la p h a s e l i q u i d e 6ta i t u t i l i s6e p o u r les p r o t 6 i n e s et les gros pep t ides , la p h a s e so l ide ~tait r6se rv6e aux pep,t ides de fa ib le p o i d s mo l6cu l a i r e . De r6cen t s d 6 v e l o p p e m e n t s t ech - n iques tels l ' u t i l i s a t i on du p o l y b r 6 n e [1, 2, 3] ont ~largi le c h a m p d ' a p p t i c a t i o n des deux t e c h n i q u e s . E n p h a s e so l ide te c h o i x du s u p p o r t est p r i m o r - d ia l ; un s u p p o r t id6al d e v a n l pos s6de r les ca r ac - t 6 r i s t i ques su ivan t e s :
a) 6tre i n e r t e v is -h-vis des r6ac t i f s utilis.6s au cou r s de l a d~6gradation ;
Abbrdoialions :
DITC = -phenyl$ne diisothiocyanate. DMF = dimethglformamide. CSe = disulfure de Carbone. DCCD = dicgclo hex!tlcarbodiimide. TFA = acide trifluoroac~tique.
b) 6tre f o r t e m e n t subs t i tu6 en r e g r o u p e m e n t s f o n c t i o n n e l s ap tes h r6ag i r a v e c les p r o t 6 i n e s ou les p e p t i d e s ;
c) pos s6de r de b o n n e s p rop r i6 t6 s r h6o log iques p o u r p e r m e t t r e un 6 c o u l e m e n t des so lvants , lors- que la r6s ine est p lac6e darts une co lonne , sans a u g m e n t a t i o n i m p o r t a n t e de p ress ion .
Le p o l y s l y r b n e amin6 [4] et le v e r r e p o r e u x [5] on t 6t6 u l i l i s6s h ce j o u r ; l eurs avan tages et l eurs fa ib lesses sont b i e n c o n n u s [6] : les d6r iv6s du p o l y s t y r b n e sont t r6s sens ib les aux effets de sol- r an t s , le v e r r e p o r e u x b i en que r i g i d e est pa r t i e l l e - m e n t so lubi l i s6 dans des c o n d i t i o n s a lca l ines .
Nous p r o p o s o n s d ' 6 t u d i e r des d6r iv~s de l ' a c i d e p o l y a c r y l i q u e qu i ne p r 6 s e n t e n t pas ces i nconv6- n ien t s ; l eu r syn th6se et d i f f6 ren t s m o d e s d ' a c t i v a - t i on ' sont d6cr i t s i c i ; un e x e m p l e d ' u t i l i s a t i o n est r e p o r t &
Materiel et M~thodes.
Sgnth~se de ddrivds N-aminopolgacrglamide.
5 g de ehlorure de l 'aeide polyaerylique [3] sont suspendns dans une solution contenant un exe6s de diamine et la quantit~ de base ter t ia ire n~eessaire pour neutral iser le ehlore port~ par la r6sine. Le m~lange est agit~ ~ 40 ° pendant 12' heures, il est e n s u r e filtr~, lav~ par du m~thanol, de l 'ac~tone et s~eh~ sous vide : tes flltrats sont r~unis et la quantit~ de ehlore ]ib~r~ est estim6e par la m~thode de Volhard.
Dans le cas de l '~thyl6ne diamine et de l 'acide 2-5 diamino benz6ne sulfonique le contenu en diamine peut 6tre d6termin~ directement apr6s hydrolyse acide (HG1 6N, 110% 32 heures) sur autoanalyseur d'acides amin6s (Beckman 119; r~sine AA 15).
Activation des d~riv~s N-aminopolyacrylamide :
Les regroupements amines des r~sines obtennes sont transform6s en isothioeyanate par diff6rentes tech- niques.

204 J.-C. Cavadore.
a) Activat ion par le thiophosg~ne : On suspend 1 g de r6sine polyacrylamide duns 10 ml de chloroforme contenant 10 p. cent de thiophosg~ne et on met reflux pendant 4 heures. La r6sine filtr6e, puts lav6e au chloroforme et s6ch6e sous vide, doit dtre utilis~e rapidement .
b) Act ivat ion par le chlorure de th iocarbamoyle [7~ 1 g de r6sine polyacrylamide est mis h reflux duns le benz6ne avee un excbs de chlorure de th iocarbamoyle (4 g) ; apr~s 4 heures, la r6sine est filtr6e, lav6e h l '6thanol et h l '6ther 6thylique, puts s6eh6e sous vide.
e) Activat ion par le p-phenyl~nedi i so th iocyanate [8]. On suspend 1 g de r6sine polyacrylamide duns 0,5 ml de d imethy l fo rmamide contenant 2 p. cent de N-6thyl- morpholine, un tr~s large exc6s (50 fois la quanti t6 d 'amino groupes) de DITC dissout duns le DMF est alors ajout6. La suspension est agit~e h 40 ° pendant 3 heures sous un balayage d'azote ; elle est ensui te ill- tr~e, lav~e au DMF, h l '6thanol et s~ch~e sous vide.
d) Act ivat ion par le mdlange d isul fure de carbone/ carbodi imide [9, 101. 1 g de r6sine polyaerylamide est suspendu duns 5 ml de CSe, le m~lange est refroidi h 10 ° et 5 g de dicyclohexylcarbodiimide sont ajout6s, la suspension est agit6e pendant 12 heures. La r6sine est ensuite filtr6e, iav6e au DMF, h l '6thanol et s6ch6e sous vide.
D~terminat ion sar un peptide module des groupe- men t s isothiocyanate portds par les rdsines.
10t) mg de r~sine activ~e sont suspendus duns 1 ml de solut ion aqueuse de N-6thylmorphol ine (5 p. cent v /v ) contenant 100. ~tmole de leucyl-glycine ; on agite h 40 ° pendant 4 heures puts la r~sine est filtr6e, lav6e h l 'eau, h l '6thanol, h l '~ther 6thylique puts s6ch6e sous vide. La r6sine est reprise par 1 ml d'acide trifluoro- ae6tique et la suspension est agit~e h 44),°C pendant 30 minutes . Le TFA par cyeHsation et coupure provoque la l iberat ion d 'une quanti t6 de glycine ~gale h la quan- tit6 de dipeptide fix6 par les groupelnents isothiocya- nate de la r6sine. La quanti t6 de glycine ainsi lib~r~e est d6termin~e sur autoanalyseur.
At tachemen t des protdines aux rdsines.
La prot~ine (3,00 h 500 nmole) est dissoute dans 1 ml d 'une solution aqueuse de N-6thylmorphol ine (2 p. cent v /v ) h laquelle on ajoute 100 mg de r6sine polyaeryla- mide activ6e, la suspension est main tenue en agi tat ion 4 heures h 40°C. En fin de r6action, pour bloquer les gronpements amines et isothiocyanate dventuel lement libres, on ajoute suceessivement 100 mg de m6thyliso- th ioeyanate et 100 !~1 d '6thanolamine. La r6sine est alors lav6e lean, solut ion aqueuse de TFA (20 p. cent v/v), ~thanol] puis s~eh~e sous vide, la quantit~ de prot6ine aceroch6e est d6termin6e par hydrolyse acide d 'un ~chantillon de 10 mg de r~sine (HC1 6N, 110 °, 30 heures).
Applicat ion dt la d~gradation des protdines.
Le lysozyme de blanc d'ceuf de poule utilis6 comme prot6ine mod61e a 6t~ li~ h une r~sine du type suivant (voir tableau H)
CH~ I SO3H CH-CO-NH~NCS
qui s 'est montr6e la plus apte h l 'accrochage des pro- t6ines. La d~gradation a 6t6 effectu6e 6tape par 6tape h l 'aide du dithioae6tate de m~thyle [11], selon un pro- gramme d6jh d~crit [3], duns un s6queneeur automa- t ique du type Laursen [12] construi t duns notre labo- ratoire.
R~sultats et Discussion.
La r6s ine p o l y a c r y l c h l o r u r e (3-5 m e q de c h l o r e p a r g r a m m e de r6s ine) r6agi t f a c i l e m e n t avec u n n u c l 6 o p h i l e c o m m e 1 ' amino g r o u p e d ' u n p e p t i d e ; duns le cas de l ' a l a n y l - g l y c i n e p a r e x e m p l e le r en - d e m e n t d ' a c c r o c h a g e de 0,5 m e q / g est s a t i s f a i s a n t m a t s t o u s l e s essa i s t e n d a n t h a c c r o c h e r u n e p r o - t6 ine sur ce t y p e de r,6sine son t res t6s i n f r u c t u e u x .
Duns ]e but d ' 6 l i m i n e r des e f fe ts s t 6 r i ques d u s au s u p p o r t n o u s a v o n s i n t r o d u i t u n e s6r ie de dia- m i n e s p e r m e t t a n t de c o n s t r u i r e h l ' u n e des ex t r6- mi t~s un g r o u p e m e n t r 6 ac t i f v is -h-vis des p r o - t~ ines , se lon le s c h 6 m a s u i v a n t :
CH2 + NH2-~CH2~NH 2 -HCI
CH2 I CH--CO-NH~CH ~-NH z
Les r~su l ta t s o b t e n u s se lon la n a t u r e de ces s u b s t i t u a n t s s o n t r e p o r t 6 s duns le t a b l e a u I. La r 6 a c t i o n est comprlbte au bou t de 4 h e u r e s , le r e n - d e m e n t de c o n v e r s i o n du p o l y a c r y l c h l o r u r e en p o l y a c r y l a m i d e d 6 c r o i s s a n t avec la l o n g u e u r du s u b s t i t u a n t ou son e n c o m b r e m e n t s t6r ique .
I1 est t rbs p r o b a b l e que la r6 s ine p r 6 s e n t e u n e p a r t i e des g r o u p e m e n t s c h l o r u r e d ' a c i d e en pos i - t i o n ex p o s 6 e et q u ' u n e p a r t i e est p lu s ou m o i n s m a s q u 6 e ou s l 6 r i q u e m e n t e n c o m b r 6 e p a r des mol6- cu les de d i a m i n e a y a n t d6jh r6agi ; le d 6 p l a c e m e n t du c h l o r e a p p a r a i t c o m m e t rbs l a r g e m e n t g o u v e rn 6 p a r des f a c t e u r s s t 6 r i ques ( t ab leau I).
Les d i f f 6 r e n t e s t e c h n i q u e s u t i l i s6es p o u r t r a n s - f o r m e r t es r~s ines a m i n 6 e s en r~s ines i s o t h i o c y a - nares m o n t r e n t :
- - que le D I T C est le p lus e f f icace ; - - et que le r e n d e m e n t de c o n v e r s i o n d 6 p e n d de
la n a t u r e du s u b s t i t u a n t , les a r o m a t i q u e s d o n n a n t de m e i l l e u r s r6su t t a t s que les a l i p h a t i q u e s .
BIOCHIMIE, 1978, £0, n ° 2.
Ddgradation des protdines en phase solide. 205
C e p e n d a n t , l ' e f f icac i t6 des r 6 s i n e s i s o t h i o c y a - n a t e s h f ixer une p r o t 6 i n e d 6 p e n d n o n s e u l e m e n t du degr6 de s u b s t i t u t i o n en g r o u p e m e n t s i so t h i o - c y a n a t e , m a i s auss i de la n a t u r e du s u b s t i t u a n t et
de son m o d e d ' a c t i v a t i o n . E n pa r t i c t f l i e r , le c a r a c - t~re h y d r o p h i l e du s u b s t i t u a n t s p 6 c i a l e m e n t au v o i s i n a g e du g r o u p e m e n t i s o t h i o e y a n a t e , s e m b l e e s sen t i e l p o u r o r i e n t e r la p , rot6ine d a n s u n e p o s i -
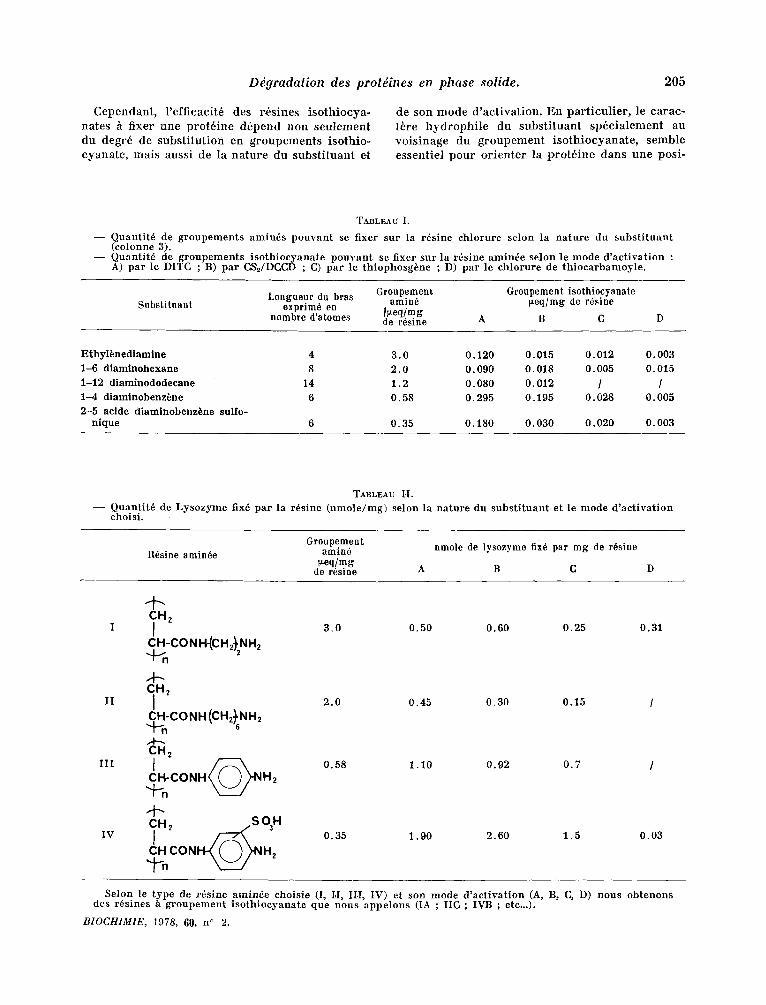
TABLEAU I.
- - Quantit6 de groupements amin6s pouvant se fixer sur la r6sine chlorure selon la nature du subs t i tuant (colonne 3).
- - Quantit6 de groupements isothioeyanate pouvant se fixer sur la r6sine amin6e selon le mode d 'act ivat ion : A) par le D,ITC ; B) par CS'~/DCCD ; C) par le thiophosg6ne ; D) par le chlorure de thiocarbamoyle.
Substituant Groupement Groupement isothiocyanale
Longueur du bras amin6 i~eq/nag de r6sine exprim6 en
hombre d'atonaes ~eq /mg de r6sine A B C
Ethyl~nediamine 4 3.0 0.120 0.015 0. 012 0.003 1-6 diaminohexane 8 2.0 0.090 0.018 0.005 0.015 1-12 diaminododecane 14 1.2 0. 080 0. 012 / / 1-4 diaminobenz~ne 6 0.58 0.295 0.I95 0.028 0.005 2-5 acide diaminobenz~ne sulfo-
nique 6 0.35 0.180 0.030 0.020 0.003
TABLEAU H. - - Quantit6 de Lysozyme fix6 par la r6sine (nmole/mg) selon la nature du subs t i tuant et le mode d 'aet ivat ion
ehoisi.
R6sine aminde Groupement
amia6 nmole de lysozyme fiX6 par nag de r6sine rteq/nag
de r6sine A B C
CHz I I 3 .0 0.50 0.60 0.25 0.31
CH-CO N H-(C H 2~'NH z
CH2 II I 2.0 0.45 0.30 0.15 /
CH-CONH (CH2~'sNH2 M--. CH2
m t ~ 0.58 1.10 0.92 0 .7 / C H - C O N H ( ( ) ) ' N H 2
~H, -S 031"I IV ~ 0.35 1.90 2.60 1.5 0.03
,~n CON H2
Selon le type de r~sine amin6e ehoisie (I, I,I, II.I, IV) et son mode d 'act ivat ion (A, B, G, D) nous obtenons des r6sines a groupement isothioeyanate que nous appelons (IA ; IIC ; IVB ; etc...).
BIOCH1MIE, t978, 60, n ° 2.
206 J . -C. C a v a d o r e .
t ion favor isant la r6act ion de couplage. Ceci est mis en 6vidence dans le tableau II.
A- CH 2
C~ H-CONH-~NCS "+',,. (=B)
Consid6rons deux r6sines amin6es di f f6remment substitu6es I I I ; IV) mats t0utes deux activ6es par la vote B.
Le tableau II nous mont re que c'est la r6sine (IV B) por teuse du r eg roupemen t sulfonique qui fix.e le mieux la prot6ine.
CH2
I / ~ NH-C NH//--~ NcS CH-CONH-(, ~, ,) )- ,- -~t j/,- ~ r ~ __xx--~03 H S L - - J
IW'A~
Gonsid6rons alors la m6me r6sine amin6e (IV) mats activ6e pa r deux votes diff6rentes A et B.
n mole d'acide amine regi~rl6r~
500--
300_
100.
10.
ys Va l Phe G ly A r g C y s Giu L e u A fa A ia A ia M e t L y s Em
' . . . . ' ' ' I b ' ' ' . . . .
5 15 N ° res idu
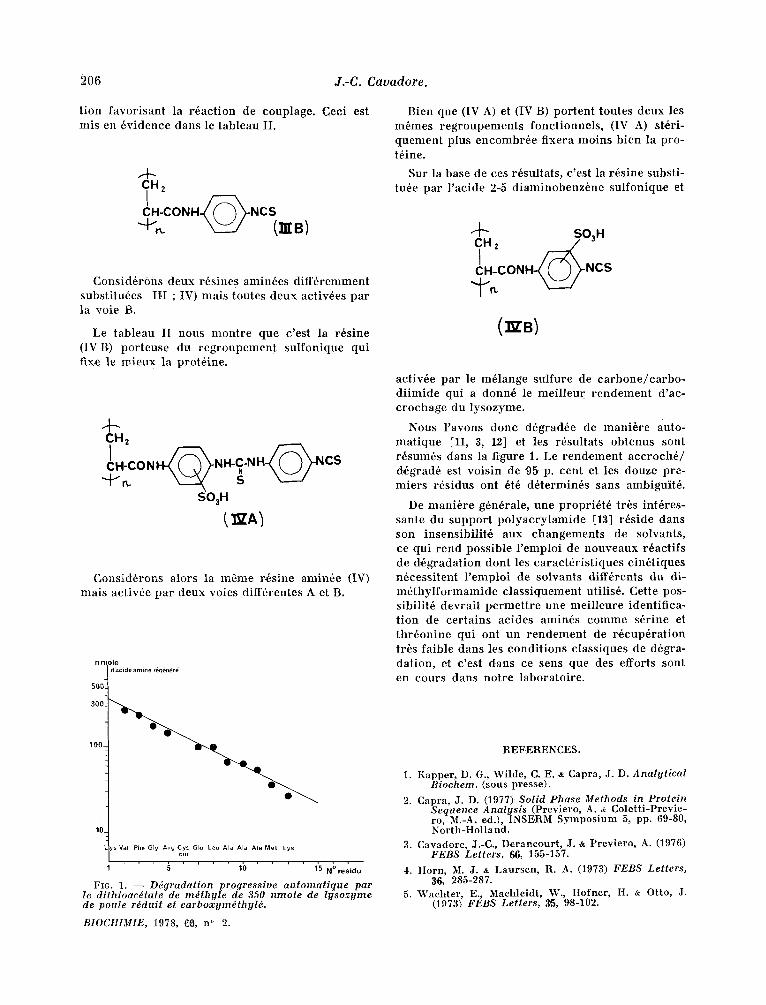
F16. 1. - - Ddgradaiion progressive au tomat ique par le di thioac~tale de m~thy le de 350 nmole de l y sozyme de poule rdduit el carboxym~thyld.
BIOCHIMIE, 1978, £0, n ° 2.
Bien que (IV A) et (IV B) por tent routes deux ]es m6mes regroup.ements fonct ionnels , (IV A) st6ri- quement plus encombr6e fixera moins bien la pro- 16ine.
Sur la base de ces r6sultats, c 'est la r6sine substi- tu6e par l ' ac ide 2-5 d iaminobenz6ne sulfonique et
CC•IC ~ O a H
ONH~NCS
activ6e par le m61ange sulfure de c a r b o n e / c a r b o - d i imide qui a donn6 le mei l leur r endement d 'ac- c rochage du lysozyme.
Nous l 'avons donc d6grad6e de mani6re auto- mat ique Ell, 3, 12] et les r6sultats obtenus sont r6sum6s dans la ~igure 1. Le rendement acc roch6 / d,6grad6 est voisin de 95 p. cent et Ies douze pre- miers r6sidus ont 6t6 d6termin6s sans ambiguit6.
De mani6re g6n6rale, une propri6t6 tr6s int6res- sante du support po lyac ry lamide [13] r6side dans son insensibil i t~ aux changements de solvants, ce qui rend possible t ' emploi de nouveaux r6actifs de d6gradat ion dont les caract6r is t iques cindtiques n6cessitent l ' emplo i de solvanls diff6rents du di- m6thylforxnamide c lass iquement utilis6. Cette pos- sibilit6 devrai t pe rmet t re une mei l leure identif ica- t ion de cer tains acides amin6s comme s6rine et thr6onine qui ont un rendement de r6cup6rat ion tr6s faible dans les condi t ions classiques de d6gra- dalton, et c 'est darts ce sens que des efforts sont en cours dans notre laboratoire.
REFERENCES.
1. Kapper, D. G., Wilde, C. E. a Capra, J. D. Analy t ica l Biochem. (sous presse).
2. Capra, J. D. (1977) Solid Phase Methods in Protein Sequence Analys is (Previero, A. ~ Coletti-Previe- ro, M.-A. ed.), INSERM Symposium 5, pp. 69-80, North-Holland.
3. Cavadore, J.-C., Derancourt, J. ,¢ Previero, A. (1976) FEBS Letters, 66, 155-157.
4. Horn, M. J. ~ Laursen, R. A. (1973) FEBS Letters, 36, 285-287.
5. ~Vachter, E., Maehleidt, W., Hofner, H. ~ Otto, J. (1973) FEBS Letters, 35, 9.8-102.

Ddgradation des prot~ines en phase solide. 207
6. Inman, J. K., Dubois, G. ~, Appella, E. (19.77) Solid Phase Methods in Prote in Sequence Analys i s (Previero, A. ,~ Coletti-Previero, M.-A. ed.) IN~:ERM Symposium 5, pp. 81-94, North-Holland.
7. Sayigh, A. A., Ulrich, It. & Potts, J. S. (1966) J. Org. Chem., 2~65-2466.
8. Laursen, R. A., Horn, M. J. ~, Bonner, A. G. (1972) FEBS Letters, 12, 62-70.
9. Joachims, J. C. ~ Seeligera (1967) Augew. Chem. Int . Ed. Engl., 6, 174-176.
10. Joachims, J. C. (1968) Ber., 101, 1748-1751. 11. Previero, A. & Cavadore, J.-C. (1975) Sol id Phase
Methods in Prote in Sequence Analys i s (Laursen, R. A. ed. Pierce, Rockford), pp. 63-72,.
12. Laursen, R. A. (1971) Eur. J. Biochem., 20, 89-102. 13. Atherton, E., Bridgen, J. ~ Sheppard, R. G. (1976)
FEBS Letters, 64, 178-175.
BIOCHIMIE, 1978, 60, n ° 2.





























