Embed Size (px)
Citation preview

ECOLE NATIONALE VETERINAIRE DE LYON
Année 2006 - Thèse n° 25
INTÉRÊT DES PRIMATES AFRICAINS DANS LA RECHERCHE SUR LE PALUDISME
THESE
Présentée à l’UNIVERSITE CLAUDE-BERNARD - LYON I (Médecine - Pharmacie)
et soutenue publiquement le 3 février 2006 pour obtenir le grade de Docteur Vétérinaire
par
SALLÉ Bettina Née le 4 octobre 1974
à Clermont de l’Oise


ECOLE NATIONALE VETERINAIRE DE LYON
Année 2006 - Thèse n° 25
INTÉRÊT DES PRIMATES AFRICAINS DANS LA RECHERCHE SUR LE PALUDISME
THESE
Présentée à l’UNIVERSITE CLAUDE-BERNARD - LYON I (Médecine - Pharmacie)
et soutenue publiquement le 3 février 2006 pour obtenir le grade de Docteur Vétérinaire
par
SALLÉ Bettina Née le 4 octobre 1974
à Clermont de l’Oise




Au Professeur PICOT,
De la Faculté de Médecine Claude Bernard - Lyon I, Qui nous a fait l’honneur d’accepter la présidence de ce jury de thèse, Hommages respectueux. Au Professeur BOURDOISEAU
De l’Ecole Nationale Vétérinaire de Lyon, Qui a accepté de diriger notre thèse à distance, Qu’il trouve ici l’expression de notre profonde reconnaissance. Au Professeur CADORÉ
De l’Ecole Nationale Vétérinaire de Lyon, Qui a aimablement accepté de participer à notre jury de thèse, Qu’il trouve ici l’expression de nos sincères remerciements.


Au Docteur Pierre ROUQUET, ex responsable du Centre de Primatologie du CIRMF,
Pour son expérience, sa confiance, sa loyauté et son soutien. Longue route parmi la faune sauvage.
Au Professeur Philippe BLOT, Directeur Général du CIRMF,
Pour son accueil eu sein du centre et ses encouragements dans mes travaux. Salutations respectueuses.
Au Docteur Pierre DRUILHE, Directeur de l’Unité de Parasitologie Biomédicale de l’Institut Pasteur,
Pour le partage de ses connaissances et sa disponibilité. Sincères remerciements.
Au Professeur Dominique MAZIER, Directrice de l’Unité Immunologie cellulaire et moléculaire des infections parasitaires de l’INSERM,
Pour sa gentillesse et ses précieux conseils, Amicales salutations.
Au Docteur Benjamin OLLOMO, chercheur en parasitologie au CIRMF,
Pour m’avoir initiée aux méandres du paludisme. Salutations.
A l’ensemble du personnel du Centre de Primatologie du CIRMF,
Pour leur contribution toujours souriante à mon travail quotidien. Lévéléwé.


A Nicole SALLÉ,
Ma mère, ma Lumière. Pour son Amour inconditionnel. Pour ce qu’elle est. Mes plus belles émotions. A Magali, Marinette, Florent, Jean-Marie et leurs tribus respectives,
Mon sang, mes ancres… Pour leur amour, sans limite, sans frontière. Mes plus tendres pensées.
Au Docteur Émilie BAUDRY,
Pour donner, chaque jour, ses lettres de noblesse à l’Amitié. Infinie tendresse.
Aux Docteurs Véronique GONEL, Caroline JACQUES et Karin LEMBERGER,
Pour ne pas avoir laisser les distances et les années ronger notre complicité. Amour, gloire et beauté.
Aux Docteurs Bertrand LEMENERT, Olivier BISSEAUD, Christelle RIBOULET et Nathalie PALHA,
Ô mes anciens, ô ma mère de clinique, Pour m’avoir sortie de la sombre ignorance et m’avoir initiée aux joies de notre métier.
Reconnaissance éternelle. A Romain, Sandrine, Gilles, Olivier, Marie, Agnès, Paul, Trish, Valérie, Olivier, Sosthène, Isaac et les autres, gabonais de souche ou de cœur,
Pour le partage de leur amour pour ce pays et pour ce continent. Tendresses.
A mes étoiles et mes lunes, Pour avoir allumé mes nuits, coloré mes jours et fait mon cœur plus riche.
Douceur caressante. A ces soleils rencontrés en terres rouges dont la lumière continue d’irradier de là-haut :
Annelle, Esteban,
Bettina, Bibiche.
Tendres pensées, sourire en coin et cœur au bord des yeux. A tous ceux qui, pour quelques gènes de différence, nous offrent leur liberté et parfois leur vie pour rendre la nôtre plus facile.
Respect et reconnaissance infinis.


Au Docteur Jean-Louis SALLÉ, 22 février 1939 – 14 janvier 2000
Chirurgien de mon cœur, Pour m’avoir invitée dans cette Vie
et avoir contribué à me la faire belle. Trois notes de blues.
Evolution inéluctable qui, Parallèlement à ce grand courant

Partant du singe pour aboutir à l’homme, Part de l’homme pour aboutir à l’imbécile.
Boris Vian


TABLE DES MATIERES
Introduction ___________________________________________________ 17
PREMIERE PARTIE : BILAN ET ORIENTATION DE LA RECHERCHE SUR LE PALUDISME 17
A- SITUATION ACTUELLE DU PALUDISME _____________________________ 21 A.1- PRESENTATION ZOOLOGIQUE DU GENRE PLASMODIUM____________ 21 A.2- EPIDEMIOLOGIE DU PALUDISME__________________________________ 21
A.2.1- Importance du paludisme dans la santé publique_______________________ 21 A.2.2- Importances relatives des différentes espèces plasmodiales humaines ______ 23 A.2.3- Epidémiologie mondiale__________________________________________ 24
A.3- INFECTION ______________________________________________________ 27 A.3.1- Cycle du plasmodium____________________________________________ 27 A.3.2- Epidémiologie de la transmission___________________________________ 28 A.3.3- Clinique ______________________________________________________ 29 A.3.4- Diagnostic_____________________________________________________ 31 A.3.5- Thérapeutique__________________________________________________ 33 A.3.6- Prophylaxie____________________________________________________ 34
B- THEMATIQUES DE RECHERCHE SUR LE PALUDISME ________________ 35 B.1- PROBLEMATIQUE : LA COMPLEXITE INSOUPÇONNEE DU PALUDISME35
B.1.1- La complexité du plasmodium : polymorphisme et apparitions de résistance_ 35 B.1.2- La complexité de la relation hôte-parasite ____________________________ 37 B.1.3- Le coût des recherches ___________________________________________ 37
B.2- GRANDS AXES DE RECHERCHE SUR LE PALUDISME ________________ 38 B.2.1- Etude du génome du parasite ______________________________________ 38 B.2.2- Développement de la transgénèse __________________________________ 39 B.2.3- Phylogénie des plasmodiums ______________________________________ 40 B.2.4- Développement d’outils diagnostiques_______________________________ 43 B.2.5- Etudes de physiopathologie _______________________________________ 44 B.2.6- Etudes épidémiologiques _________________________________________ 44 B.2.7- Etude de l’immunité liée au paludisme ______________________________ 45 B.2.8- Etude des apparitions de résistance aux anti-paludiques _________________ 46 B.2.9- Développement de nouvelles stratégies thérapeutiques __________________ 48 B.2.10- Développement d’un vaccin antipaludique __________________________ 50

DEUXIEME PARTIE : LES PRIMATES AFRICAINS ET LA RECHERCHE SUR LE PALUDISME_____________________________________________________________ 57
A- UN POINT SUR L’UTILISATION DES PRIMATES NON-HUMAINS DANS LA RECHERCHE __________________________________________________________ 59
A.1- IMPORTANCE QUANTITATIVE DES PRIMATES PAR RAPPORT AUX AUTRES ANIMAUX___________________________________________________ 60
A.1.1- Cas de l’Amérique du Nord _______________________________________ 60 A.1.2- Cas de l’Europe ________________________________________________ 60
A.2- UTILISATION DES PRIMATES PAR SECTEUR DE RECHERCHE ________ 61 A.3- REPARTITION DES DIFFERENTES ESPECES DE PRIMATES ___________ 62
A.3.1- Cas de l’Amérique du Nord _______________________________________ 62 A.3.2- Cas de l’Europe ________________________________________________ 63
A.4- LES PRIMATES UTILISES POUR LA RECHERCHE SUR LE PALUDISME _ 63
B- LES PLASMODIUMS DES PRIMATES NON-HUMAINS _________________ 64 B.1- HISTORIQUE_____________________________________________________ 64 B.2- LES PLASMODIUMS ET LEURS HOTES NATURELS __________________ 66 B.3- CYCLE DU PLASMODIUM CHEZ LES PRIMATES_____________________ 67 B.4- CLINIQUE _______________________________________________________ 67 B.5- DIAGNOSTIC ____________________________________________________ 68 B.6- THERAPEUTIQUE ________________________________________________ 69
C- INTERETS DES PRIMATES AFRICAINS DANS LA RECHERCHE SUR LE PALUDISME___________________________________________________________ 69
C.1- INTERET EN TANT QUE MODELE ANIMAL _________________________ 69 C.1.1- Par rapport aux modèles bio-informatiques ___________________________ 70 C.1.2- Par rapport aux modèles in vitro ___________________________________ 70 C.1.3- Par rapport à une étude directe sur l’humain __________________________ 71
C.2- INTERET PAR RAPPORT AUX MODELES NON PRIMATES ____________ 72 C.2.1- Sensibilité croisée des plasmodiums ________________________________ 72 C.2.2- Proximité des tableaux cliniques ___________________________________ 76 C.2.3- Proximité des systèmes immunitaires _______________________________ 77
C.3- INTERET PAR RAPPORT AUX AUTRES PRIMATES ___________________ 78 C.3.1- Proximité phylogénétique des primates africains avec l’homme___________ 79 C.3.2- Sensibilité aux quatre plasmodiums humains _________________________ 81 C.3.3- Sensibilité aux infections par les sporozoïtes de P. falciparum __ 82 C.3.4- Forte reproductibilité des résultats __________________________________ 82 C.3.5- Sensibilité à d’autres plasmodiums simiens___________________________ 82 C.3.6- Proximité phylogénétique de leurs plasmodiums naturels avec P. falciparum 83 C.3.7- Disponibilité et facilité d’élevage __________________________________ 83
D- LIMITES D’UTILISATION DES PRIMATES AFRICAINS DANS LA RECHERCHE SUR LE PALUDISME______________________________________ 84
D.1- LIMITES BIOLOGIQUES ___________________________________________ 84 D.1.1- Intérêt de l’utilisation des techniques in vitro _________________________ 84 D.1.2- Limites du modèle animal ________________________________________ 84 D.1.3- Avantages des autres modèles animaux par rapport aux primates africains __ 85
D.2- CONTRAINTES FINANCIERES _____________________________________ 89 D.3- CONTRAINTES ECOLOGIQUES ____________________________________ 90
D.3.1- Disponibilité du modèle__________________________________________ 90

D.3.2- Envoi des prélèvements __________________________________________ 90 D.4- CONTRAINTES ETHIQUES ET LEGISLATIVES _______________________ 91
D.4.1- Ethique et utilisation des primates __________________________________ 91 D.4.2- Ethique et législation ____________________________________________ 93
TROISIEME PARTIE : EXEMPLES D’UTILISATION DES PRIMATES AFRICAINS DANS LA RECHERCHE SUR LE PALUDISME _____________________________ 97
A- UTILISATIONS DE PRIMATES AFRICAINS AVEC LEUR PLASMODIUM NATUREL _____________________________________________________________ 99
A.1- ETUDE D’UNE NOUVELLE SOUCHE PLASMODIALE NATURELLE DU CHIMPANZE _________________________________________________________ 99
A.1.1- Caractéristiques morphologiques ___________________________________ 99 A.1.2- Caractéristiques phylogénétiques ___________________________________ 99 A.1.3- Caractéristiques biologiques______________________________________ 102 A.1.4- Conclusion et applications possibles _______________________________ 111
A.2- ESSSAI D’UN VACCIN ERYTHROCYTAIRE ET D’UNE THERAPIE PAR ANTICORPS RECOMBINANTS SUR DES CHIMPANZES INFECTES PAR PLASMODIUM REICHENOWI _________________________________________ 112
A.2.1- Principe de l’étude _____________________________________________ 112 A.2.2- Matériel et méthode ____________________________________________ 113 A.2.3- Résultats _____________________________________________________ 116 A.2.4- Discussion____________________________________________________ 119 A.2.5- Conclusion ___________________________________________________ 122
B- UTILISATIONS DE PRIMATES AFRICAINS EN TANT QU’HOTES ARTIFICIELS DE PLASMODIUMS HUMAINS____________________________ 123
B.1- RECHERCHE D’UN NOUVEAU MODELE POUR LE PALUDISME : ESSAI D’INFECTION DE QUATRE MANDRILLS PAR PLASMODIUM FALCIPARUM123
B.1.1- Principe de l’étude _____________________________________________ 123 B.1.2- Matériel et méthode ____________________________________________ 125 B.1.3- Résultats _____________________________________________________ 125 B.1.4- Discussion et conclusion ________________________________________ 126
B.2- ESSAI D’UN VACCIN PRE-ERYTHROCYTAIRE AVEC DES CHIMPANZES INFECTES PAR PLASMODIUM FALCIPARUM___________________________ 127
B.2.1- Principe de l’étude _____________________________________________ 127 B.2.2- Matériel et méthode ____________________________________________ 127 B.2.3- Résultats _____________________________________________________ 128 B.2.4- Discussion____________________________________________________ 132 B.2.5- Conclusion ___________________________________________________ 134
Conclusion ___________________________________________________ 137
BIBLIOGRAPHIE_____________________________________________ 139
ANNEXES ___________________________________________________ 175


17
Introduction
Le paludisme est l’une des plus importantes des maladies qui frappent l'espèce humaine. Il est responsable de centaines de millions de malades chaque année et de millions de morts à travers le monde.
Le paludisme (du latin pattes ou palus, marais) est provoqué par un parasite des globules rouges du sang. Ce parasite est transmis du sujet malade au sujet sain par l'intermédiaire de moustiques femelles du genre Anopheles ; il subit dans l'organisme du malade comme dans le corps du moustique un certain nombre de transformations qui constituent son cycle évolutif.
Dans les régions chaudes et humides du globe qui représentent son domaine de prédilection, le paludisme, communément appelé aussi malaria (signifiant « mauvais air »), peut se comporter comme un véritable fléau décimant les populations indigènes et les voyageurs. Avec un million de morts par an dont 95% d’enfants, c’est la première cause de décès chez les enfants de moins de 5 ans dans la région Afrique (Organisation Mondiale de la Santé, 2003).
C’est pourquoi la lutte antipaludique est primordiale au regard de la santé publique. Mais la considérable morbidité du paludisme a aussi des conséquences sérieuses sur le plan économique et social. Alors combattre le paludisme, c'est non seulement œuvrer pour la santé des hommes, mais c'est aussi permettre à des provinces ou à des nations entières de prendre leur essor et d'accéder à un niveau de vie supérieur.
En matière de lutte contre le paludisme, la recherche médicale occupe bien sûr le premier plan.
Un siècle de travaux s’est déjà écoulé depuis la découverte des agents infectieux. Si le paludisme était au départ l’affaire de quelques biologistes et médecins, il regroupe aujourd’hui des chercheurs de toutes les spécialités à la pointe de leur domaine. En effet, le paludisme est longtemps apparu comme une maladie qui serait assez simple à éradiquer. Mais les programmes mis en place dans la seconde moitié du XXème siècle n’ont pas tenu leurs promesses et le paludisme s’est avéré être un ennemi redoutable à combattre.
L’une des difficultés auxquelles se confronte le chercheur est l’absence de modèle expérimental fiable. A ce jour, aucune technique ni aucun animal ne peut reproduire fidèlement le profil de la malaria humaine.
Les primates ont toujours été de précieux partenaires pour le chercheur. Proches des humains, ils permettent d’anticiper la réponse obtenue lors d’essais cliniques ultérieurs chez des humains et d’éviter ainsi de risquer des vies humaines ou d’entrer inutilement dans de coûteuses études sur des volontaires humains. Cependant, aucun simien n’est un modèle parfait et l’utilisation des primates non-humains soulève de nombreuses questions éthiques et financières.

18
Parmi les primates, les singes africains ont très vite été délaissés au profit de leurs cousins sud-américains, ces derniers étant considérés comme de meilleurs modèles pour l’infection paludique. Mais aujourd’hui les progrès de l’immunologie et de la biologie moléculaire semblent partiellement remettre en question l’hégémonie des singes américains dans ce domaine. Il convient donc de faire un point sur les avantages et les inconvénients de l’utilisation des primates africains dans le contexte actuel de la recherche sur le paludisme, en le comparant aux autres modèles existants et aux études menées chez l’homme. Enfin, quelques exemples expérimentaux réalisés au Centre International de Recherches Médicales de Franceville (CIRMF) au Gabon illustreront le rôle possible des primates africains dans la lutte contre le paludisme.

PPRREEMMIIEERREE PPAARRTTIIEE :: BBIILLAANN EETT OORRIIEENNTTAATTIIOONN DDEE LLAA
RREECCHHEERRCCHHEE SSUURR LLEE PPAALLUUDDIISSMMEE


Bilan et orientation de la recherche sur le paludisme
21
A- SITUATION ACTUELLE DU PALUDISME
A.1- PRESENTATION ZOOLOGIQUE DU GENRE PLASMODIUM
- Règne animal, sous règne des Protozoaires : ce sont des êtres vivants unicellulaires. - Embranchement des Sporozoa (Apicomplexa) : ce sont des parasites dont le cycle
débute toujours par un germe vermiforme et mobile, appelé sporozoïte. Il est également caractérisé par une phase de croissance et de division des noyaux, la schizogonie, ainsi que par l’existence de gamètes bien différenciés.
- Classe des Haemosporidea - Super-ordre des Sarcosporidies, - Ordre des Haemosporida (Danilevski, 1885) : les micro et macrogamétocytes se
développent indépendamment ; pas de conoïde ; les microgamétocyte produisent 8 microgamètes ; le zygote est mobile (ookinète) ; les sporozoïtes sont nus ; endodyogénie absente ; hétéroxène ; schizogonie (mérogonie) chez le vertébré ; pigment (hémozoïne) visible au microscope optique ; sporogonie chez l'invertébré ; transmision par insectes hématophages.
- sous-ordre des Haemasporina - Famille des Plasmodiidae (Mesnil, 1903) : les caractères de la famille sont les mêmes
que ceux de l'ordre : l'hôte invertébré est un moustique ; il y a une schizogonie dans le sang ; les gamétocytes mûrs sont dans les érythrocytes et au cours de la maturation dans l'érythrocyte, il y a apparition de pigment malarien.
- Genre Plasmodium : les gamétocytes mâle et femelle sont de même taille et situés dans les érythrocytes ; les mâles produisent un nombre limité (8) de microgamètes ; le résultat de la fécondation est un ookinète mobile qui se transforme en oocyste dont la taille augmente au cours de la maturation et qui donne naissance à de très nombreux sporozoïtes ; la schizogonie (mérogonie) a un développement synchrone chez l'hôte vertébré et aboutit à la formation de schizontes présents dans les érythrocytes et dans certains autres tissus.
- Environ 200 espèces qui touchent les reptiles, les batraciens, les oiseaux et les mammifères (Grasse, 1953) responsables ou non de maladies chez leurs hôtes.
L’existence de nombreuses espèces de Plasmodium et de nombreux hôtes sont autant d’arguments pour la recherche d’un bon modèle animal dans la recherche sur le paludisme.
A.2- EPIDEMIOLOGIE DU PALUDISME
A.2.1- Importance du paludisme dans la santé publique
Le paludisme, déjà décrit 2700 ans avant Jésus Christ par les chinois, a souvent modifié profondément le cours de l’histoire. Ainsi, la malaria n’est pas étrangère au déclin des civilisations grecque et romaine, ni à la chute de l’Empire. Sans elle, le destin d’Alexandre ne se serait pas arrêté à l’apogée de sa gloire militaire. La malaria est intimement liée à différents événements tels la conquête de Madagascar, la construction du canal de Panama ou encore la guerre du Pacifique.

Bilan et orientation de la recherche sur le paludisme
22
Dans le monde, le paludisme représente la 5ème cause de mortalité liée à une maladie infectieuse, derrière les maladies respiratoires aiguës (3 millions de décès par an), le SIDA (3 millions de décès en 2001), les maladies diarrhéiques (2,5 millions de décès par an). Le paludisme tue autant que la tuberculose (près de 2 millions de décès par an), le cancer (1,2 millions de décès en 2002) ou les accidents de la route (1,2 millions de décès en 2002) (Organisation Mondiale de la Santé, 2003). Selon les estimations de l'OMS (Organisation Mondiale de la Santé, 2003; Organisation Mondiale de la Santé, 2005):
• entre 2 et 3,2 milliards d'individus, soit près de la moitié de la population mondiale, sont exposés au paludisme.
• 350 à 500 millions nouveaux cas cliniques surviennent chaque année (contre 300 millions en 1940). Certaines études estiment ces chiffres à 660 millions uniquement pour le Plasmodium falciparum (Snow et al., 2005).
• Le paludisme tue chaque année 1,5 à 2,7 millions de personnes (contre 3 millions en 1940) dont 600 000 jeunes enfants.
• 90% des morts résidaient en Afrique. • Le paludisme tue une personne toutes les 12 secondes dans le monde. • Le paludisme tue un enfant toutes les 30 secondes en Afrique.
Le paludisme touche 107 pays dans le monde (contre 140 dans les années 50), dans toutes les régions tropicales et sub-tropicales du globe (sauf dans quelques îles du Pacifique) (Organisation Mondiale de la Santé, 2005). L’endémie est particulièrement élevée dans les zones tropicales défavorisées d'Afrique, d'Asie et d'Amérique Latine. L'Afrique est, de loin, le continent le plus touché avec 90% des cas de paludisme recensés dans ses zones tropicales. Des épidémies peuvent également survenir lors de mouvements de populations peu exposées au paludisme vers des zones hautement endémiques. L'Europe connaît aussi des cas de paludisme, dits d'importation. En France par exemple, on déplore plusieurs milliers de malades et une vingtaine de décès par an. C’est le pays le plus touché d’Europe. La majorité de ces cas survient chez des personnes n'ayant pas suivi de prophylaxie lors de leur séjour à l’étranger. Le nombre de cas de paludisme d’importation en France est assez corrélé au flux de voyageurs vers les zones d’endémie (près de 3,5 millions en 2001). Ce nombre a augmenté régulièrement depuis 1996 (5 000 cas en 1998, 7 000 cas en 1999 selon la Direction Générale de la Santé) pour se stabiliser en 2001 avec 7 300 personnes malades et enfin diminuer de 10% en 2002 (Institut de veille sanitaire, 2003). Les pays de contamination sont majoritairement situés en Afrique subsaharienne (95 % des cas), ce qui explique que plus de 80 % des cas sont dus à Plasmodium falciparum (espèce africaine et mortelle). En ce qui concerne les cas importés en France, l’incidence pour 100 000 voyageurs et par an est en Afrique respectivement 15 à 23 fois plus élevée qu’en Amérique du Sud ou en Asie. Le paludisme est un fléau mondial. Ces données épidémiologiques édifiantes justifient des investissements financiers majeurs dans la recherche.

Bilan et orientation de la recherche sur le paludisme
23
A.2.2- Importances relatives des différentes espèces plasmodiales humaines
Quatre espèces de parasites du genre Plasmodium décrites à la fin du XIXème siècle sont responsables de la maladie chez l'homme : Plasmodium falciparum, Plasmodium vivax, Plasmodium ovale et Plasmodium malariae.
- Plasmodium falciparum est l'espèce la plus pathogène et est responsable de l’immense majorité des cas mortels. Elle est présente dans les zones tropicales d'Afrique, d'Amérique Latine et d'Asie, et elle est dominante en Afrique. D’après le rapport 2003 de l’OMS, des signes inquiétants témoignent de l'extension du paludisme à P. falciparum à de nouvelles régions et de sa résurgence dans des zones où il avait été éliminé (Organisation Mondiale de la Santé, 2003).
- Plasmodium vivax co-existe avec P. falciparum dans le reste du monde et est présent dans certaines régions tempérées. P. vivax prédomine en Amérique centrale, dans certains pays d'Amérique du sud, au Moyen-Orient et en Inde. Ailleurs (Asie du Sud-Est et Indonésie notamment), les prévalences de P. vivax et P. falciparum sont proches.
Les deux dernières espèces plasmodiales, P. malariae et P. ovale, sont beaucoup plus rares.
- Plasmodium ovale est une espèce principalement trouvée en Afrique de l'Ouest ; cette espèce n’est pas mortelle mais peut entraîner des rechutes 4 à 5 ans après la primo infection.
- Plasmodium malariae a une distribution mondiale mais très inégale. Elle n'est pas meurtrière mais peut entraîner des rechutes jusqu'à 20 ans après la primo infection.

Bilan et orientation de la recherche sur le paludisme
24
A.2.3- Epidémiologie mondiale
A.2.3.1- répartition mondiale du paludisme
Figure 1 : incidence estimée des cas cliniques de paludisme (quelle que soit l’espèce) résultant d’une transmission locale – moyenne par pays en 2004 (Organisation Mondiale de la Santé, 2005) En France, les zones mondiales sont classées en 3 groupes par le Centre National de Référence de l'Épidémiologie du Paludisme d'Importation et Autochtone (C.N.R.E.P.I.A.) (voir annexe 3). Les données, publiées dans le bulletin épidémiologique hebdomadaire (B.E.H.) (Institut de veille sanitaire, 2003), suivent trois critères :
• présence ou absence de paludisme, • espèce de Plasmodium incriminée, • degré de résistance de la souche.
pas de paludisme

Bilan et orientation de la recherche sur le paludisme
25
Par ailleurs, l’OMS définit de son côté, selon leur risque, trois groupes de régions en constante évolution selon les nouvelles résistances plasmodiales (les recommandations de prophylaxie françaises et les données de l’OMS ne se recoupant pas toujours (Droy & J., 2003)).
Risque faible. Régions où le paludisme est présent seulement dans de petites zones délimitées. Pas de souche de P. falciparum résistante à la chloroquine. Risque intermédiaire, pas de souche de P. falciparum résistante à la chloroquine. Risque intermédiaire avec présence de souche de P. falciparum résistante à la chloroquine. Risque élevé de souche de P. falciparum résistante à la chloroquine.
Figure 2 : carte de répartition mondiale du paludisme d’après les données OMS (Organisation Mondiale de la Santé, 2003)
Le paludisme concerne tous les pays et son fléau est en constante évolution : il faut surveiller l’apparition de résistances aux traitements et le développement du paludisme dit d’importation lié aux possibilités de voyages accrues (paludisme souvent grave car il touche des individus non immuns). Par ailleurs, chaque région ainsi définie possède un intérêt différent pour la recherche, en fonction du type d’individus à qui l’étude s’adresse (immuns ou non) et de la souche plasmodiale que l’on souhaite étudier (espèce plasmodiale, éventuelle résistance, etc.).

Bilan et orientation de la recherche sur le paludisme
26
A.2.3.2- liens entre le paludisme et le niveau de vie
Le paludisme concerne une centaine de pays mais près de 60 % de tous les décès dus au paludisme concernent les 20 % les plus pauvres de la population mondiale, ce qui en fait la maladie la plus étroitement associée à la pauvreté (Organisation Mondiale de la Santé, 2002).
Le paludisme affecte la santé et la richesse des pays et des personnes. En Afrique aujourd'hui, on reconnaît que le paludisme est à la fois une maladie due à la pauvreté et une cause de pauvreté. Les économistes attribuent au paludisme un déficit de croissance annuel pouvant atteindre 1,3% dans certains pays d'Afrique. Ainsi, au fil des années, l'écart se creuse entre le PIB des pays selon qu'ils sont touchés ou non par le paludisme et c'est la croissance économique de toute la région qui est pénalisée (Sachs & Malaney, 2002). Le problème financier est donc au cœur des préoccupations de la recherche sur le paludisme ; les pays les plus touchés n’ont pas les moyens d’apporter leur contribution à la recherche et les fonds proviennent de pays peu touchés. Le paludisme ne faisant pas partie des priorités de santé publique des pays payeurs, les sommes allouées s’avèrent trop aléatoires et insuffisantes (Gallup et al., 1999).
Figure 3 : estimation de la chargemondiale d’accès palustres en 1998(Gallup et al., 1999)
Figure 4 : estimation de la pauvretédans le monde en 1998 (Gallup &Sachs, 1999)
Charge croissante de paludisme
Charge croissante de pauvreté
Bilan et orientation de la recherche sur le paludisme
27
A.3- INFECTION
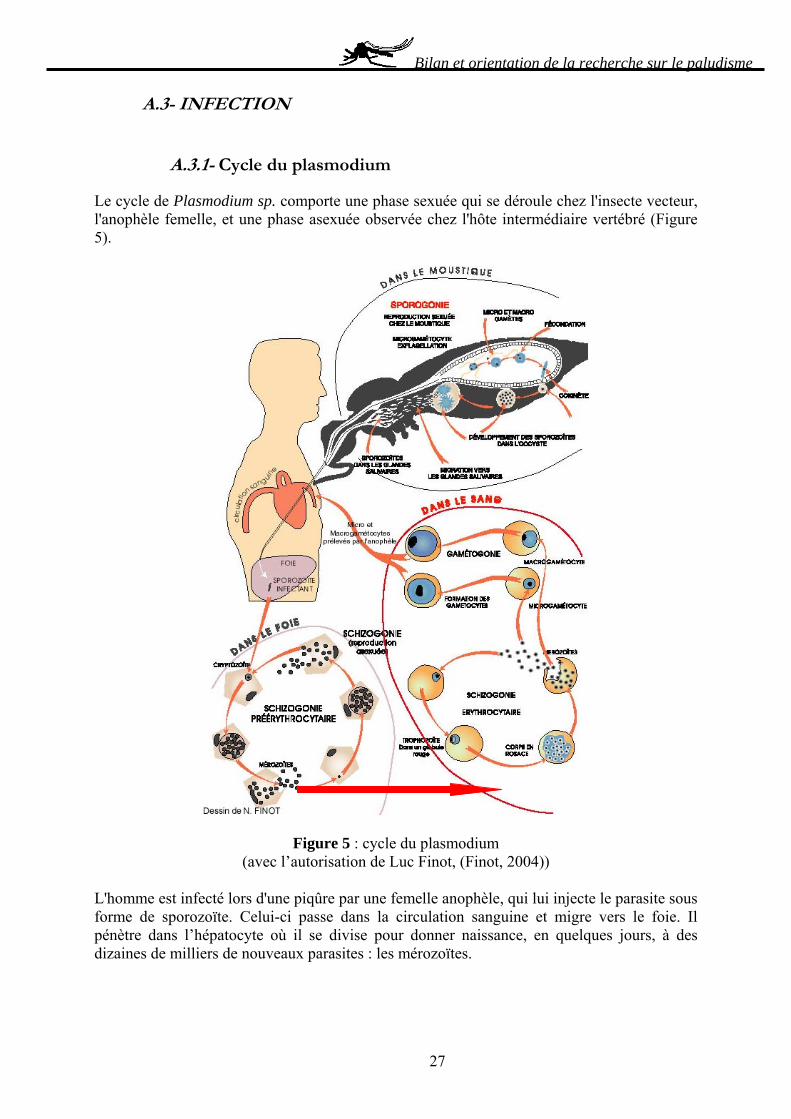
A.3.1- Cycle du plasmodium
Le cycle de Plasmodium sp. comporte une phase sexuée qui se déroule chez l'insecte vecteur, l'anophèle femelle, et une phase asexuée observée chez l'hôte intermédiaire vertébré (Figure 5).
Figure 5 : cycle du plasmodium (avec l’autorisation de Luc Finot, (Finot, 2004))
L'homme est infecté lors d'une piqûre par une femelle anophèle, qui lui injecte le parasite sous forme de sporozoïte. Celui-ci passe dans la circulation sanguine et migre vers le foie. Il pénètre dans l’hépatocyte où il se divise pour donner naissance, en quelques jours, à des dizaines de milliers de nouveaux parasites : les mérozoïtes.

Bilan et orientation de la recherche sur le paludisme
28
La cellule du foie éclate en libérant ces parasites dans le sang. Les mérozoïtes pénètrent dans les érythrocytes et s’y multiplient. Les globules rouges éclatent et les mérozoïtes ainsi libérés en infectent de nouveaux. En parallèle, des cellules sexuées mâles et femelles, les gamétocytes, se forment dans le sang du sujet infecté. Ainsi, lorsqu'un moustique pique un humain infecté, il ingère ces gamétocytes. Ceux-ci se transforment en gamètes et leur fécondation engendre un œuf. L’œuf migre vers les glandes salivaires du moustique et se différencie en sporozoïte. Un nouveau cycle peut alors commencer. L’infection de ce parasite est complexe : parasite dixène, existence de plusieurs stades du protozoaire vivant dans des parties différentes de ses hôtes, existence de nombreuses phases intracellulaires, etc. La recherche sur le paludisme devra donc explorer chaque stade de vie du parasite pour mieux le cerner.
A.3.2- Epidémiologie de la transmission
Le paludisme est une anthropozoonose, avec 3 modes de transmission possible : - piqûre d'une anophèle infectée, - inoculation lors de transfusions (sang de donneur infecté) et de piqûres (matériel ayant précédemment servi à des patients infectés), - transmission de la mère à l’enfant.
En outre, trois types de conditions doivent être remplies pour qu’il y ait paludisme : des conditions concernant la vie de l’anophèle, des conditions de transmission et des conditions concernant l’homme (Duriez, 2002).
1- La condition nécessaire pour la pullulation des anophèles est la présence d’eau (indispensable au développement larvaire et nymphal). Cette condition est remplie toute l'année dans les régions humides (tropiques, équateur).
2- La condition nécessaire à la transmission du paludisme est une température minimale continue. Suivant les cas, soit la transmission sera continue toute l'année et entraînera un état immun, soit elle sera saisonnière, soit encore elle sera intermittente, auquel cas il n'y aura pas d'installation d'état immun.
3- Concernant l’humain, il existe des éléments favorables à la transmission qui entraînent un caractère aggravant du paludisme :
déficience grave en glucose-6-phosphate deshydrogénase (G6PDH) liée à un risque hémolytique et à une contre-indication au traitement par la chloroquine et la primaquine),
déficit immunitaire (SIDA (Grimwade et al., 2004), enfants de 3 mois à 2 ans, splénectomie, etc.)
grossesse (perte d’immunité surtout pour les premières grossesses)
En revanche, le déficit en G6PDH sans hémolyse ou la présence d'hémoglobine S (drépanocytose) entraîne une protection contre l’infection à P. falciparum en diminuant la possibilité de paludisme sévère (Pasvol et al., 1978). Il en est de même pour l’hémoglobine F (alpha-thalassémie), mais de façon plus controversée (Williams et al., 1996; Allen et al., 1997; Mockenhaupt et al.,

Bilan et orientation de la recherche sur le paludisme
29
2004). Les personnes de groupe Duffy négatif seraient quant à elles plus résistantes à P. vivax (Miller et al., 1976). D’autres facteurs génétiques liés aux globules rouges (Pathirana et al., 2005) ou à l’immunité (complexe HLA) exercent une influence sur le plasmodium, mais sans doute dans une moindre mesure. Enfin, des facteurs non génétiques, comme un régime exclusivement lacté déficient en acide para-aminobenzoïque ou une malnutrition protéique, entraîneraient une relative protection vis-à-vis du paludisme.
L’épidémiologie de transmission du paludisme est elle aussi complexe : facteurs génétiques protecteurs ou aggravants, facteurs économiques et sociaux, facteurs climatiques, etc. Tous ces facteurs sont autant de difficultés difficilement maîtrisables.
A.3.3- Clinique
Les symptômes du paludisme peuvent apparaître à partir de 7 jours après une piqûre infectante, mais la maladie peut demeurer latente pendant plusieurs semaines.
Cycle durée symptomatologie
passage dans la circulation sanguine 30 minutes
cycle dans le foie 5 à 12 jours 5 à 7 pour P. falciparum
cycle dans le globule rouge quelques jours
phase asymptomatique
éclatement du globule rouge quelques heures apparition des symptômes Tableau 1 : chronologie des évènements chez l’humain après la piqûre par une femelle anophèle infectée (Site portail de la Guyane) Les signes ne sont pas spécifiques : fièvre d'apparition brutale (39 à 40 °C), grands frissons, fatigue, courbatures, maux de tête, troubles digestifs (diarrhée, douleurs abdominales, surtout chez l'enfant). Contrairement aux notions classiques, la fièvre est plus souvent continue qu'intermittente. Il n'est donc pas possible de faire le diagnostic de paludisme sur ces éléments cliniques, que l'on retrouve dans beaucoup de maladies infectieuses, notamment virales. Seul un examen sanguin, réalisé dans un laboratoire ayant l'expérience du diagnostic de cette parasitose, permet de confirmer le paludisme. Enfin, chaque espèce plasmodiale présente des particularités quant à leurs tableaux cliniques.
A.3.3.1- particularités de Plasmodium vivax
• fièvre tierce bénigne (cycle sanguin en 48h) ; • exceptionnel dans la race noire ; • paludisme bénin avec rechutes pendant 2 années dues à l’existence
d’hypnozoïtes hépatiques ; • paludisme viscéral évolutif si infestations répétées : fébricule permanent avec
poussées irrégulières de fièvre, splénomégalie, amaigrissement et anémie.

Bilan et orientation de la recherche sur le paludisme
30
A.3.3.2- particularités de Plasmodium ovale
• fièvre tierce bénigne (cycle sanguin en 48h) ; • localisation essentiellement africaine, notamment dans les pays de la côte ouest
de l'Afrique Centrale ; • clinique voisine de celle de P. vivax : paludisme bénin avec rechutes ; • rechutes pendant 5 ans au maximum dues aux hypnozoïtes hépatiques.
A.3.3.3- particularités de Plasmodium malariae
• phase d'état : fièvre quarte bénigne (cycle sanguin en 72h) ; • recrudescence jusque 10 à 20 ans ou plus par réactivation de formes
érythrocytaires latentes (pas d'hypnozoïtes) ; • complication : néphropathie quartane (immunopathologie par dépôt de
complexes immuns qui entraîne une insuffisance rénale grave).
A.3.3.4- particularités de Plasmodium falciparum
• prédomine sur le continent africain ; • fièvre tierce maligne (cycle sanguin en 48h) ; • c’est la seule espèce susceptible d'entraîner le décès du patient par paludisme
grave dit aussi "pernicieux" (Gachot et al., 2001). Ce sont les enfants qui en sont le plus souvent atteints. Les femmes enceintes en zone d'endémie et le voyageur sans immunité anti-palustre sont également susceptibles de présenter cette pathologie. Il se présente chez le grand enfant sous forme d’un neuropaludisme, et chez le petit enfant plutôt sous forme d’une anémie grave (Saissy et al., 2003). L’encéphalopathie se manifeste avec une fièvre élevée, des troubles neurologiques, des convulsions, un coma, souvent une hépatomégalie, une insuffisance rénale, une anémie et une thrombopénie. Les hématies parasitées sont séquestrées au sein des micro-vaisseaux cérébraux et d’autres organes. Le mécanisme précis de ce phénomène de cytoadhérence et de séquestration est encore mal connu, mais les plaquettes semblent jouer un rôle important (Wassmer et al., 2003a). Sans traitement, l’évolution est fatale en quelques jours dans 30% des cas. Le patient peut également présenter des séquelles neurologiques définitives, surtout chez l'enfant.
• A l'inverse, un accès simple de paludisme à P. falciparum traité tôt et correctement guérit toujours (Gachot & Pays, 2002), et définitivement dans l'immense majorité des cas (sauf bien sûr si le patient est à nouveau exposé au risque, c'est-à-dire retourne ou séjourne en zone impaludée).
Le tableau clinique diffère selon l’espèce plasmodiale incriminée. La recherche sur le paludisme devra tenir compte de ces différentes pathologies, d’une part pour être plus ciblée et donc plus efficace, d’autre part parce que toutes les espèces ne sont pas aussi menaçantes que P. falciparum. Ainsi, la clinique moins spectaculaire de P. vivax, qui est pourtant aussi répandu que P. falciparum, le relègue bien derrière ce dernier pour les efforts de recherches (Mendis et al., 2001).

Bilan et orientation de la recherche sur le paludisme
31
A.3.4- Diagnostic
A.3.4.1- diagnostic clinique
Les symptômes d’appel sont une fièvre et/ou un syndrome grippal, 8 jours ou plus après l'arrivée en pays d'endémie, souvent atypiques (Duriez, 2002).
A.3.4.2- diagnostic direct : mise en évidence du parasite, de ses composants ou de ses produits
Le diagnostic est le même, quelle que soit l’espèce plasmodiale, et est facile à réaliser.Toutefois, le diagnostic de l'espèce est ensuite indispensable en raison du risque de neuropaludisme lié à P. falciparum. L’identification se fait sur frottis mince et/ou goutte épaisse (les deux pouvant être réalisés sur une même lame) colorés au Giemsa. Le coût de cette méthode est de l’ordre du centime d’euro par test (Lema et al., 1999). Il faut d’abord prélever une goutte de sang à l’aide d’une aiguille ou d’une lancette sur le doigt ou l’oreille préalablement désinfecté. Dans l’idéal le sang devra être pris en phase ascendante d’hyperthermie. On dépose la goutte épaisse et on réalise le frottis mince sur une même lame puis on laisse sécher. Il est préférable de fixer le frottis mince au méthanol absolu dès qu’il est sec. La coloration s’obtient idéalement après 45 minutes dans du Giemsa à pH 7,25. Lorsque le temps est compté (par exemple lors de diagnostic sur un animal anesthésié), on peut doubler la concentration de Giemsa et réduire le temps de trempage à 15 minutes. En deçà de cette durée, la lecture est moins bonne. Quand un grand nombre de gouttes épaisses doivent être colorées, le colorant de Field est indiqué parce que son effet est plus rapide (quelques secondes) (Noland et al., 2003) ; le cytoplasme de l'hématie parasitée est rose (annexe 4). L’observation de la goutte épaisse doit durer au moins 5 minutes, ce qui correspond à environ 100 champs observés. Dans le même principe, on ne peut déclarer un frottis négatif avant 15 minutes de lecture attentive. Un exemple de frottis négatif et un exemple de frottis positif sont présentés en annexe 5. Il est également important de chiffrer la parasitémie (en % d'hématies parasitées ou en nombre de parasites/μL) pour évaluer le risque de neuropaludisme dans le cas de P. falciparum :
- on peut compter le nombre de globules rouges parasités parmi 10 000 globules rouges pour obtenir ainsi une parasitémie exprimée en pourcentage ;
- on peut aussi, plus rapidement, compter le nombre de globules rouges infectés parmi 100 globules blancs. Il faut ensuite multiplier ce chiffre par 80 (car on considère une moyenne de 8000 leucocytes/μL) pour obtenir une approximation de la parasitémie exprimée en nombre de parasites par microlitre. Cette méthode est bien sûr faussée si une autre infection est présente simultanément en raison de l’éventuelle leucocytose associée.
Encadré 1 : Technique d’identification sur frottis mince et goutte épaisse

Bilan et orientation de la recherche sur le paludisme
32
L’examen de la goutte épaisse doit constituer la première étape étant donné qu’elle présente l’avantage de concentrer 20 fois plus de parasites qu’un frottis mince (détection de 20 parasites par microlitre). L’inconvénient est que les parasites peuvent paraître déformés, rendant leur identification difficile. Si la présence de parasites est détectée, l’espèce devra alors être identifiée par l’examen du frottis mince.
Quantitative Buffy Coat malaria test (QBC) : coloration fluorescente des acides nucléiques par l'acridine orange ; ce test semi-quantitatif ne permet pas le diagnostic d’espèce.
Méthode facile, rapide, mais chère (environ 5 euros par test, (Lema et al., 1999)).
Polymerase Chain Reaction (PCR) : possibilité de détection de 1 plasmodium dans 10 ml de sang.
Test très sensible mais compliqué et cher.
Recherche d'antigènes circulants utilisation d’anticorps monoclonaux anti plasmodium (techniques IFI ou ELISA )
• Parasight F et ICT Malaria Pf : recherche de l’antigène HRP-2 (histidine-rich protein-2) spécifique de P. falciparum.
Test facile, sensible, spécifique, mais cher (d’environ 1 euro en Ouganda à 8 euros en Europe (Singh & Shukla, 2001)).
• ICT Malaria Pf/Pv : détecte l’antigène HRP2 de P. falciparum et un antigène commun aux 4 espèces de Plasmodium.
• OptiMAL : recherche de LDH (lactate deshydrogénase) spécifique du parasite ; la nouvelle version d’OptiMal permet de faire le diagnostic des 4 espèces plasmodiales (VanderJagt et al., 2005).
Test simple, sensible (seuil de 50-100 parasites/μL), spécifique, mais cher (environ 1,50 euro (Lab.shoklo-unit, 2002)).
Encadré 2 : autres techniques de diagnostic direct nécessitant plus de matériel et de technicité
A.3.4.3- diagnostic indirect : recherche d'anticorps sériques
Il existe plusieurs techniques qui sont assez compliquées et chères ; du fait du délai d’apparition et de la persistance des anticorps, mais aussi de la possibilité de réactions croisées, le diagnostic indirect n’est pas intéressant dans une démarche purement diagnostique. Cependant il est utile dans les cas suivants :
- en cas de parasitémie nulle et de fièvre d'origine inconnue, - en cas de prise d'antipaludique préalable à la démarche diagnostique, - en cas de surveillance post thérapeutique, - dans le cas de donneurs de sang (risque de paludisme transfusionnel pour le
receveur), - lors d'enquêtes épidémiologiques.

Bilan et orientation de la recherche sur le paludisme
33
Les techniques de diagnostic direct s’utilisent pour toutes les espèces plasmodiales et pour tous les hôtes de plasmodium. Dans le cadre de la recherche sur primates, les kits QBC et les gouttes épaisses sont très couramment utilisées en diagnostic direct. Les sérologies sont également faciles pour les chimpanzés pour lesquels on utilise les mêmes réactifs que les humains.
A.3.5- Thérapeutique
Le choix du traitement dépend avant tout de la gravité et ces critères de choix sont déterminés par l’OMS. La liste des noms déposés disponibles en France et dans quelques autres pays est consultable en annexe 6.
A.3.5.1- molécules disponibles
On peut classer ces médicaments en fonction de leur cible :
• les schizonticides à action rapide Quinine Amino 4 quinoléines : Chloroquine, Amodiaquine
• les schizonticides à action lente
antifoliques : Dapsone, Sulfadiazine antifoliniques : Proguanil, Pyriméthamine association Pyriméthamine-Sulfadoxine antibiotique : Doxycycline
• les schizonticides de découverte plus récente
Méfloquine association Pyriméthamine-Sulfadoxine-Méfloquine Halofantrine Artémisinine et ses dérivés Pyronaridine (extrême-orient uniquement)
• les gamétocytocide ou hypnozoïtocide
Amino 8 quinoléine : Primaquine (pays anglo-saxons uniquement, très efficace mais dotée d'une toxicité élevée, surtout vis-à-vis de certains sujets africains.)
La gamme d’antipaludiques disponibles est assez réduite. Un des objectifs de la recherche sur le paludisme est donc le développement de nouvelles thérapeutiques.
A.3.5.2- schémas thérapeutiques
Les molécules utilisables contre le paludisme sont malheureusement peu nombreuses et la plupart d’entre elles possède une certaine toxicité (Taylor & White, 2004). Ces effets secondaires vont du simple prurit engendré par la chloroquine aux accidents cardiovasculaires possibles avec la quinine et l’halofantrine ou aux troubles neuropsychiatriques sévères liés à la méfloquine.

Bilan et orientation de la recherche sur le paludisme
34
Leur emploi doit donc être raisonné et tenir compte de plusieurs points :
le coût local et la disponibilité des médicaments antipaludiques, la gravité de l’accès palustre, l’espèce plasmodiale incriminée, la région où la maladie a été contractée (par exemple pour la situation de la résistance
de P. falciparum aux antipaludiques), la chimioprophylaxie antécédente, les allergies connues, les maladies concomitantes, l'âge et l'état de grossesse (Stauffer & Fischer, 2003), les chances d'acceptabilité du traitement par le patient, le risque de nouvelle exposition au paludisme après le traitement.
Malgré cela, on estime globalement qu’environ 1% des sujets atteints par P. falciparum meurent de la maladie (Organisation Mondiale de la Santé, 2003). La recherche sur le paludisme comporte un important volet recherche et développement. Du fait de la toxicité de la plupart des antipaludiques, les essais avant la mise sur le marché d’une nouvelle molécule doivent être très rigoureux et la pharmacovigilance assidue.
A.3.6- Prophylaxie
Le paludisme est transmis par la piqûre d’anophèle, la prévention du paludisme passe donc d’abord par la lutte contre les piqûres de moustiques (évitant aussi la transmission d'autres maladies, comme la dengue transmise par des ædes ou l'encéphalite japonaise transmise par les culex) (Droy et al., 2003). Ensuite, la prévention passe par la prise d’une chimioprévention adaptée. Les recommandations sanitaires pour les voyageurs (Institut de veille sanitaire, 2003; Organisation Mondiale de la Santé, 2003) découlent du découpage mondial vu précédemment en quatre groupes.
• Séjour dans un pays du groupe 0 (zones sans paludisme) : pas de chimioprophylaxie. • Séjour dans un pays du groupe 1 (zones sans chloroquinorésistance) : chloroquine • Séjour dans un pays du groupe 2 (zones de chloroquinorésistance) : association de
chloroquine-proguanil ou association atovaquone-proguanil
• Séjour dans un pays du groupe 3 (zones de prévalence élevée de chloroquinorésistance ou multirésistance) : méfloquine ou association atovaquone-proguanil (si intolérance à la méfloquine, monohydrate de doxycycline)
De même que pour le traitement du paludisme, la gamme des produits utilisés en prévention est limitée et les risques toxiques sont élevés. Il est donc important de chercher à développer de nouvelles molécules et/ou un vaccin contre le paludisme, notamment pour les pays situés en zone 3.

Bilan et orientation de la recherche sur le paludisme
35
B- THEMATIQUES DE RECHERCHE SUR LE PALUDISME
Dans le cadre de ce travail la recherche concernant les anophèles ne sera pas abordée.
En un siècle de recherche, quatre prix Nobel ont été attribués à des travaux concernant le paludisme :
- Sir Ronald Ross (1902) : cycle biologique du parasite paludéen chez l’anophèle ;
- Alphonse Laveran (1907) : découverte du parasite dans le sang humain ; - Julius Wagner-Jauregg (1927) : traitement pour une réduction des troubles
psychiatriques à partir de l'inoculation du paludisme ; - Hermann Muller (1948) : utilisation du DDT comme insecticide.
B.1- PROBLEMATIQUE : LA COMPLEXITE INSOUPÇONNEE DU PALUDISME
Une vague d’optimisme a envahi les esprits dans les années 60-70. Le problème semblait simple, il suffisait de développer l’usage de la quinine et des insecticides en attendant l’arrivée proche du vaccin. Malheureusement, au fur et à mesure des recherches, les scientifiques ont découvert l’extrême complexité du paludisme et les dernières décennies nous offrent une réalité alarmante :
- augmentation des épidémies de paludisme (Tyagi, 2004) ; - accroissement de la mortalité depuis trente ans (Sitalakshmi et al., 2003), y
compris en Afrique subsaharienne ; - augmentation de cas de résistance de P. falciparum (May & Meyer, 2003) ; - réémergence de P. vivax dans des zones où il avait été éradiqué, comme le
Caucase et l’Asie Centrale (Sharma, 1996; Povoa et al., 2003 ) ; - augmentation des cas de paludisme d’importation dans les pays développés
(Harling et al., 2004). Aujourd’hui, le vaccin tant espéré n’est toujours pas disponible et les traitements doivent se renouveler pour faire face aux mutations du parasite. Désormais la seule certitude est qu’il n’y a pas de solution unique et miraculeuse pour éradiquer le paludisme. On sait d’une part qu’il s’agira d’une lutte sur plusieurs fronts et d’autre part que cette lutte n’aboutira pas nécessairement à la disparition du paludisme, mais plutôt à une atténuation de ses méfaits.
B.1.1- La complexité du plasmodium : polymorphisme et apparitions de résistance
Avec les anti-pyrétiques, les anti-paludiques sont les molécules les plus utilisées dans les régions tropicales. L’emploi massif de traitements dans les cinquante dernières années a provoqué une importante pression de sélection sur le plasmodium.

Bilan et orientation de la recherche sur le paludisme
36
Les niveaux de chimiorésistance des souches plasmodiales sont évalués après un traitement correctement suivi (Institut de veille sanitaire, 2003). Les souches de Plasmodium isolées des patients sont alors classées en S, R1, R2 et R3 :
- S : disparition complète des formes sanguines sans réapparition (sensible) ; - R1 : disparition des formes sanguines en 1 semaine, réapparition avant 1 mois ; - R2 : diminution du nombre des formes sanguines ; - R3 : aucune diminution du nombre des formes sanguines.
Aujourd’hui on observe de nombreuses souches de P. falciparum résistantes aux antipaludiques, en particulier en Asie du Sud-Est et de plus en plus en Afrique (annexe 3). L’émergence de ces résistances est une des causes majeures de résurgence du paludisme de ces trois dernières décennies (Marsh, 1998) et sans doute la principale cause du doublement de mortalité infantile par paludisme en Afrique de l’Est et du Sud (Korenromp et al., 2003). Ces résistances continuent de progresser, non seulement de façon géographique mais aussi par rapport aux molécules :
- les premières résistances ont concerné la quinine (Nussenzweig et al., 1967) et s’étendent aujourd’hui aux nombreux pays des zones 2 et 3 (Le Bras & Durand, 2003).
- Une autre molécule de première intention, la chloroquine, a entraîné une chimiorésistance dans presque tous les pays impaludés (Ndyomugyenyi et al., 2004; Abdel-Aziz et al., 2005).
- Les résistances aux molécules de deuxième intention sont aussi fréquentes : les résistances à l’association sulfadoxine-pyriméthamine se sont répandues rapidement et largement (Ahmed et al., 2004).
- Les résistances à la méfloquine (Ramharter et al., 2004) sont encore localisées à certaines zones asiatiques où elle a été largement utilisée (Nosten et al., 2000).
- Des résistances à l'halofantrine (Ndong et al., 2003) sont observées depuis une quinzaine d’années.
- On rencontre également régulièrement des cas de résistance à l’association atovaquone-proguanil (Muehlen et al., 2004).
Les résistances de P. vivax sont moins étudiées. Il existe une résistance à la chloroquinine depuis 15 ans dans la région indonésienne et depuis peu sud-américaine (Ruebush et al., 2003) et à l’association sulfadoxine-pyrimethamine de façon plus répandue (Pukrittayakamee et al., 2004). D’autre part, l’effet de la primaquine semble décroître, en particulier en Océanie. L’étude de ces résistances a bénéficié des progrès de la biologie moléculaire de ces vingt dernières années. L’apparition d’une résistance procède de deux évènements : tout d’abord un « accident » génétique qui produit un mutant résistant, puis le processus de sélection par lequel le mutant possèdera un avantage face à la molécule antipaludique. Ce processus favorise alors sa transmission et ainsi l’extension de la résistance (White & Pongtanornpinyo, 2003). A ce jour, les mécanismes exacts et complets d’apparition de ces résistances sont encore débattus ou inconnus. Il faudra décrypter les quelques 5000 gènes et anticiper le haut degré de polymorphisme antigénique du plasmodium pour espérer pouvoir un jour devancer ses mutations (Hayton & Su, 2004).

Bilan et orientation de la recherche sur le paludisme
37
B.1.2- La complexité de la relation hôte-parasite
La complexité du paludisme n’est malheureusement pas une exception, puisqu’il n'existe actuellement aucun vaccin efficace dans le domaine de la parasitologie humaine. En premier lieu il faut atteindre le parasite au sein de son hôte alors que le plasmodium est souvent intracellulaire. Par ailleurs, on ne connaît pas bien les mécanismes de l’immunité liée au paludisme : on connaît l’existence d’un état de « prémunition » qui correspond à une protection contre la maladie (et non contre le parasite), mais on ne sait malheureusement pas reproduire cet état. On s’est d’abord beaucoup intéressé à l’immunité acquise. Les facteurs humoraux ont été très étudiés, dans l’espoir de développer une sorte de sérum antipaludique, mais sans succès. Il semble désormais évident que les cellules immunocompétentes circulantes, bien que fonctionnelles, ne reflètent pas la réalité des mécanismes immunologiques de défense mis en jeu par l'homme contre P. falciparum (ou n’en reflètent qu'une partie infime) ; l’immunité innée joue elle aussi un rôle important (Stevenson & Riley, 2004). Ainsi, actuellement les facteurs humoraux sont surtout utilisés pour tester les antigènes candidats pour un futur vaccin (Garraud et al., 2003) mais beaucoup moins dans l’espoir de développer une protection. La complexité de cette immunité a deux conséquences sur le développement d’un futur vaccin :
- la première est d’ordre général : le choix des antigènes candidats doit se faire de façon parfois empirique et les prédictions in vitro sont souvent éloignées des résultats in vivo.
- La seconde conséquence concerne les régions endémiques, c’est-à-dire les populations immunes : il est difficile mais indispensable d’intégrer les conditions particulières rencontrées sur le terrain pour l’élaboration d’un vaccin.
B.1.3- Le coût des recherches
Le paludisme ne tue presque que dans les pays en voie de développement (Gallup et al., 1999) et l’implication des pays riches est encore insuffisante.
Par ailleurs, on ne peut que déplorer l'hésitation des industries pharmaceutiques à investir dans ces projets. En 1990, alors que les dépenses globales consacrées à la recherche sur le paludisme représentaient 65 dollars par victime, elles s'élevaient à 789 dollars dans le cas de l'asthme et à 3 274 dollars dans celui du sida.
La recherche dans le domaine des maladies tropicales a été la principale victime des fusions observées dans l'industrie pharmaceutique au cours des dix dernières années, les firmes tendant à concentrer leurs ressources sur des domaines restreints mais potentiellement plus profitables (Bouguerra, 1998).

Bilan et orientation de la recherche sur le paludisme
38
Des exceptions voient cependant le jour, comme le prouve le récent accord de collaboration entre la fondation DNDI (Drugs for Neglected Diseases Initiative créée en 2003) et Sanofi-Aventis pour le développement d’un nouveau médicament contre le paludisme à un prix cible inférieur à 0,8 euro. Il s’agit d’une combinaison à dose fixe d’artésunate et d’amodiaquine à un prix moins élevé que les autres combinaisons actuellement disponibles. Ce médicament devrait pouvoir être mis à la disposition des malades dans le courant 2006. De son côté, en 1998, l’OMS présentait le projet Roll Back Malaria (Faire Reculer le Paludisme). Son objectif est de diminuer de moitié la mortalité liée au paludisme d’ici 2010 et encore de moitié d’ici 2015. Au départ, il était estimé qu’il faudrait environ 1,6 milliards d’euros annuels pour réaliser ce plan (Narasimhan & Attaran, 2003). En 2005 ces estimations étaient passées à 2,6 milliards d’euros par an pour lutter de manière efficace contre le paludisme dans les 82 pays où la charge de morbidité est la plus forte. En 2005 donc, malgré un effort de 480 millions fournis pour la lutte contre cette maladie, ces fonds restent très insuffisants. La Banque Mondiale a récemment annoncé un plan visant à fournir 400 à 800 millions d’euros au cours des cinq années à venir, ce qui permettra simplement d’améliorer la couverture préventive et thérapeutique essentielle (Organisation Mondiale de la Santé, 2005).
B.2- GRANDS AXES DE RECHERCHE SUR LE PALUDISME
Pour faire face à un problème aussi complexe que celui du paludisme, la recherche doit être pluridisciplinaire et profiter des derniers progrès dans chaque domaine.
B.2.1- Etude du génome du parasite
L’étude du génome du parasite est à la base de nombreuses applications, si ce n’est de toutes les recherches actuelles sur le paludisme (Hemingway & Craig, 2004). En effet, connaître les gènes et leur expression permet :
- de comparer des parasites à d’autres (de même espèce, de même genre ou de même embranchement) pour mieux comprendre l’histoire et l’évolution du paludisme,
- de détecter le parasite (test diagnostique par recherche d’ADN parasitaire chez un individu, épidémiologie à l’échelle d’une population),
- de comprendre ses relations avec son hôte (échappement à l’immunité d’un individu, apparition de résistances aux traitement à l’échelle individuelle ou planétaire),
- de développer de nouvelles stratégies thérapeutiques et prophylactiques (antipaludiques, vaccin, thérapie génique, etc.).
B.2.1.1- techniques d’étude
a- la PCR
Mise au point en 1985, cette technique a connu le développement le plus rapide et le plus spectaculaire de la biologie. Les progrès en PCR (Polymerase Chain Reaction) ont été déterminants pour l’étude du genre Plasmodium.

Bilan et orientation de la recherche sur le paludisme
39
En général, une des limites d’utilisations de la PCR pour tester la présence d’agent pathogène, est qu’elle n’amplifie pas seulement des fragments d’organismes vivants, mais aussi de simples débris d’ADN. Dans le cadre du paludisme, il semble cependant que seuls des parasites viables puissent être détectés par PCR (Jarra & Snounou, 1998).
b- le séquençage
La très grande majorité des séquences réalisées et publiées aujourd'hui sont réalisées sur des séquenceurs automatiques. Ceux-ci sont capables de réaliser les réactions de séquence (technique proche de la PCR mais les brins synthétisés sont de longueurs différentes), puis de les lire. Le développement des séquenceurs automatiques récent a permis un gain de temps appréciable et une baisse considérable du coût de revient.
B.2.1.2- principaux résultats
La séquence génomique de P. falciparum a récemment été publiée : le génome a été analysé, le polymorphisme de l’ADN, les ARN messagers ainsi que le profil d’expression des protéines sont étudiés (Gardner et al., 2002). Les chercheurs ont surmonté d'importants obstacles techniques avant de parvenir à leurs fins, notamment la difficulté de découpage de l’ADN du parasite. En effet, peut-être à cause de sa composition peu ordinaire (richesse en A+T), le matériel génétique de Plasmodium ne peut être sectionné en morceaux de longueur différente, il se rompt seulement en de très petits morceaux. Dans le même temps, un séquençage a été effectué chez l’anophèle et chez l’humain (Hoffman et al., 2002). Cette banque de données extraordinaire n’est pas encore complètement exploitée mais elle oriente déjà les stratégies vaccinales (Carucci, 2005). On peut espérer que ce travail permettra également bientôt une meilleure compréhension des interactions vecteur-humain et hôte-parasite.
B.2.2- Développement de la transgénèse
B.2.2.1- principe
La transgenèse consiste à ajouter un gène étranger au génome d'un organisme vivant ou au contraire à remplacer très précisément un gène endogène par un autre gène. Cette dernière opération, qui implique la mise en oeuvre d'un processus de recombinaison homologue, peut conduire en pratique à l'inactivation ou à la mutation d'un gène chez l'animal. Elle permet tout aussi bien de remplacer un gène donné par un tout autre gène. Un gène peut être transféré in vivo dans les seules cellules somatiques et on procède alors à une thérapie génique. Le gène peut également être transféré dans les cellules germinales, on obtient alors des lignées d'animaux transgéniques.

Bilan et orientation de la recherche sur le paludisme
40
B.2.2.2- applications
Quatre applications sont possibles, selon l’organisme modifié :
i. production d’un ou plusieurs gènes vaccinaux, par un microorganisme, par une plante ou dans le lait d’un mammifère transgénique (Stowers et al., 2002a) ;
ii. production de parasites vaccinaux déplétés de leurs gènes de pathogénicité (Carvalho & Menard, 2005; Mueller et al., 2005) ;
iii. étude du rôle de certains gènes chez le parasite (Ozwara et al., 2003a; Healer et al., 2004), l’hôte (Tsuji & Zavala, 2003) ou le vecteur du paludisme (Catteruccia et al., 2003; Moreira et al., 2004) ;
iv. vaccination génétique dans un futur plus ou moins proche : transférer aux animaux des gènes qui confèrent de manière définitive une résistance constitutive ou induite à des maladies données. Deux voies sont possibles :
la première, déjà utilisée, consiste à injecter dans les cellules musculaires de l’hôte le gène codant pour l’antigène parasitaire afin que celles-ci produisent elles-mêmes l’antigène vaccinal (vraie vaccination à ADN nu) (Epstein et al., 2004) ;
la seconde voie, plus futuriste, consisterait à injecter à l’hôte directement un gène de résistance. Ces gènes de résistance agiraient indépendamment du système immunitaire et l'effet de protection ne serait donc pas au sens strict le résultat d'une vaccination. Les gènes de résistance pourraient être des gènes naturels identifiés chez les individus d'une espèce animale naturellement résistants à la maladie ou être ceux qui codent pour des anticorps monoclonaux.
B.2.3- Phylogénie des plasmodiums
Cette discipline découle directement des techniques vues précédemment puisqu’il s’agit de comparer différentes séquences d’ADN entre elles.
B.2.3.1- intérêts de la phylogénie
On peut distinguer trois niveaux d’études phylogénétiques : • la place du genre Plasmodium parmi les autres Apicomplexa ou même parmi les
Protozoaires, • études comparatives de différentes espèces du genre Plasmodium, • le polymorphisme au sein de l’espèce P. falciparum.
Les intérêts et les applications de cette discipline sont multiples (Rich & Ayala, 2003) :
répertorier, avec d’autres méthodes que l’observation directe, les différentes espèces plasmodiales (Perkins, 2000) ; retracer l’histoire des plasmodiums humains afin de mieux comprendre ce
qu’ils sont aujourd’hui (par exemple en mettant en parallèle l’augmentation actuelle des cas de paludisme et celle d’il y a 10 000 ans) (Hartl, 2004) ; déterminer le point de divergence des plasmodies humaines dans un but
anthropologique (Conway & Baum, 2002; Carter, 2003 ) ;

Bilan et orientation de la recherche sur le paludisme
41
comparer, à un moment donné, les différentes souches plasmodiales présentes au sein d’une population pour mieux en appréhender l’épidémiologie (Cui et al., 2003) ; chercher le meilleur modèle pour remplacer le plasmodium humain, par
exemple en cherchant une homologie de séquences vaccinales (Janssen et al., 2002) ou au contraire, en contrebalançant les ressemblances biologiques (Kissinger et al., 1998) ; comparer l’arbre obtenu avec les plasmodies à l’arbre de leurs hôtes
naturels pour étudier les mécanismes de coévolution de l’hôte et de son parasite ; évaluer le risque de mutation d’un gène candidat pour un vaccin (Wolfe et
al., 1998) ; choisir un produit de gène ou une voie métabolique très conservée comme
cible de nouveaux médicaments afin de ralentir l’apparition de résistance (Wolfe et al., 1998).
B.2.3.2- principe de la phylogénie
En fonction du niveau de comparaison que l’on souhaite effectuer on choisit différents types de gènes : Pour construire un arbre phylogénétique de l’ensemble des Protozoaires : on peut utiliser les gènes d’enzyme communes, comme la dihydrofolate reductase-
thymidylate transferase (O'Neil et al., 2003) ; on peut également s’intéresser à l’évolution de gènes précis, comme les gènes
extrachromosomaux, au sein de ces groupes pour mieux en appréhender le rôle (Gardner et al., 1994; Vinkenoog et al., 1995).
Pour une étude des 200 espèces du genre Plasmodium ces deux objectifs se retrouvent : comparaison des régions codantes et non-codantes de deux espèces plasmodiales,
assez éloignées pour mieux comprendre les « pressions d’évolution » qui se sont exercées (Castillo-Davis et al., 2004) et aussi l’éventuel rôle des introns (Pizzi & Frontali, 1999) ;
comparaison des gènes communs (ou des parties de gènes), assez bien conservés entre les différentes espèces et peu sujets à la pression de sélection liée à l’immunité (Kedzierski et al., 2002). Les trois gènes classiquement utilisés sont ceux du cytochrome b (Escalante et al., 1998), de l’ARNr codant pour la sous unité 18S (Escalante & Ayala, 1994; Escalante et al., 1997) et de la protéine circumsporozoïte (Escalante et al., 1995). L’idéal est bien sûr de cumuler ces trois gènes dans une même étude (Ayala et al., 1999). On obtient alors une idée de l’apparentement entre deux espèces plasmodiales et l’on peut également discriminer deux espèces morphologiquement identiques (Perkins, 2000).
Pour une étude au sein d’une espèce de Plasmodium : on choisit au contraire des gènes présentant un polymorphisme marqué comme le SNP
(Mu et al., 2002), CSP ou MSP (Rich & Ayala, 2000). Ces études essaient de retracer l’histoire de P. falciparum (Joy et al., 2003) ou de P. vivax (Escalante et al., 2005);
une autre possibilité consiste à sous-grouper une souche de P. falciparum connue en fonction de certains gènes impliqués dans la réponse immunitaire pour mesurer l’étendue de leur polymorphisme (pour la création d’un vaccin) et mieux cerner leur rôle (Lavstsen et al., 2003).
Encadré 3 : présentation de quelques techniques d’études phylogénétiques

Bilan et orientation de la recherche sur le paludisme
42
L’étude phylogénétique, quel que soit son objectif et quelque soit le gène visé, comporte toujours cinq étapes :
1- isolation et purification de l’ADN, 2- amplification du fragment choisi par PCR, 3- clonage et séquençage des fragments, 4- alignement des fragments grâce à un logiciel (clustalW par exemple), 5- construction de l’arbre par deux à trois méthodes : le « neighbor joining », le
« maximum likilihood » et parfois le « unweighted mawimum parsimony ». L’arbre doit être construit en tenant compte des particularités du génome du genre Plasmodium, notamment de sa forte proportion de A+T.
B.2.3.3- principaux résultats
• Les arbres de comparaisons des plasmodies humaines et animales mettent en évidence plusieurs points (Ayala et al., 1999) :
- les trois espèces plasmodiales humaines P. falciparum, P. malariae et P. vivax ne sont pas proches les unes des autres (P. ovale n’apparaît même pas sur certains arbres). Leur divergence précède largement l’origine des hominidés.
- P. falciparum est très proche de P. reichenowi, le plasmodium des chimpanzés. Ils auraient divergé il y a 8 à 12 millions d’années, ce qui correspond approximativement à la divergence d’espèces entre les deux hôtes.
- P. malariae est indistinguable génétiquement de P. brasilianum (parasite de primates sud-américains) et P. vivax de P. simium (idem), ce qui va dans le sens d’une récente transmission de ces plasmodies d’un hôte à l’autre.
• Concernant l’étude ciblée de P. falciparum, on a longtemps cru que sa virulence était liée à sa récente apparition. Or il semble d’après cet arbre que les origines de ce Plasmodium soient anciennes (5 à 50 millions d’années). D’autre part, on constate une absence de mutation silencieuse, comme si le parasite évoluait depuis peu de temps, ce qui serait en contradiction avec le haut niveau de polymorphisme allélique constaté sur les gènes CSP ou MSP. Une hypothèse peut alors être émise (Rich et al., 1998) : l’origine de P. falciparum serait bien ancienne, sans doute en Afrique tropicale, mais ce parasite serait longtemps resté cantonné à la même zone. Puis, il y a peut-être 5 à 10 000 ans, trois facteurs auraient conjointement permis la récente expansion du parasite dans le monde :
- un réchauffement climatique, favorable aux anophèles, - le développement post-néolithique de l’agriculture qui a augmenté la densité
de population, - des modifications génétiques au sein de la triade vecteur-parasite-hôte qui ont
peut-être augmenté leur affinité. Cependant, cette expansion serait trop récente pour permettre l’apparition de mutations silencieuses.
Par ailleurs, une particularité du génome de P. falciparum est de posséder des séquences très répétitives (par exemple les zones NANP et NVDP du gène CSP). Une recombinaison intragénique au sein de ces séquences peut alors engendrer des variations bien plus rapides que les mutations classiques. Ces changements situés sur des gènes participant à l’échappement à l’immunité de l’hôte seraient naturellement sélectionnés et seraient à l’origine du haut degré de polymorphisme de P. falciparum.
Encadré 4 : quelques résultats des études phylogénétiques, historique du plasmodium

Bilan et orientation de la recherche sur le paludisme
43
Les études des phylogénies profitent des avancées dans le séquençage mais les résultats sont parfois encore contradictoires et le paludisme révèle encore ici sa complexité (Hartl, 2004). Il est cependant indispensable de poursuivre les recherches dans ce sens : déterminer l’étendue de la diversité des gènes et identifier les mécanismes de la création et de la persistance de ces variations est une clé importante pour le développement d’un vaccin (Rich et al., 2003; Kwiatkowski, 2005).
B.2.4- Développement d’outils diagnostiques
Si la goutte épaisse et le frottis sont d’excellents examens (faible coût et peu de matériel nécessaire), ils requièrent du temps, de l’expérience et une quantité importante de parasites dans le sang pour leur détection (Zakai, 2003). Cette parasitémie minimale représente une quantité bénigne pour une population immune, mais il n’en est pas forcément de même pour un enfant non-immun. Ces techniques, très utilisées, ont par ailleurs eu le défaut de sous-estimer les cas de co-infections plasmodiales (Mayxay et al., 2004). Par ailleurs, les populations exposées au parasite ont des besoins différents en matière de diagnostic, ce qui permet de définir quatre types de tests nécessaires :
1- les populations dites immunes peuvent utiliser des tests d’une sensibilité moyenne mais dont le prix est modeste (car ces populations appartiennent aux pays en voie de développement) ; frottis et goutte épaisse remplissent bien ces conditions.
2- Les populations non immunes ont toutes besoin d’un test sensible capable de faire un
diagnostic d’espèce (danger lié à P. falciparum). Parmi cette population, certaines personnes peuvent avoir besoin de réaliser elles-mêmes ces tests comme, par exemple, les expatriés des campements forestiers, les techniciens encadrant les dons de sang, etc. Ces tests de diagnostic doivent alors être simples et sensibles même si cela a un coût (Marx et al., 2005).
3- On rencontre aussi au sein de la population non immune et/ou fragile, des gens à faible
revenu (malades du SIDA et femmes enceintes des pays en développement par exemple) pour lesquels le test doit, en plus de sa sensibilité, être abordable.
4- Par ailleurs, l’usage de tests diagnostiques capables de détecter des différences de
souches d’une même espèce plasmodiale permet de faire la différence entre nouvelle infection et recrudescence de paludisme chez un malade (Happi et al., 2004). C’est à partir de cette différence que l’efficacité d’une molécule peut être testée. Ce dernier type de test, alors réalisé en laboratoire, doit surtout être très spécifique et sensible, son coût et sa maniabilité étant moins cruciaux.
La PCR joue donc, ici encore, un rôle extrêmement important (Andrews et al., 2005; Mangold et al., 2005). Dans le cadre du diagnostic, les défauts de cette technique découlent de sa qualité principale : extrêmement sensible, il existe des possibilités de réactions croisées (à Toxoplasma par exemple) et/ou de contamination par un ADN non désiré. C’est pourquoi les laboratoires pharmaceutiques tentent de réduire le nombre de manipulations nécessaires et ainsi le risque de contamination du prélèvement (Perandin et al., 2004).

Bilan et orientation de la recherche sur le paludisme
44
Concrètement, les techniques récentes permettent un diagnostic en temps réel, capable de reconnaître plusieurs espèces plasmodiales dont évidemment P. falciparum avec des sensibilités de l’ordre du parasite par microlitre :
- les tests de détection des antigènes ont beaucoup progressé sur leur sensibilité (Farcas et al., 2003) mais pas encore sur leur coût, ce qui limite leur utilisation aux pays riches ;
- les tests sérologiques quant à eux ne sont quasiment pas utilisés comme outils de diagnostic. Leur défaut principal est de nécessiter un délai de plusieurs jours entre l’infection et la possibilité de détection des anticorps. Toutefois des recherches sont menées sur le recours à des antigènes de synthèse pour des études épidémiologiques (Noya et al., 2003).
L’idéal serait bien sûr de développer un test sensible, capable de différencier les espèces de Plasmodium, de taille minime, utilisable par le malade lui-même et vendu à un prix accessible aux plus démunis… mais ce test idéal n’existe pas encore. De nombreux tests existent, d’assez bonne qualité diagnostique, mais leur coût semble maintenant la barrière la plus difficile à dépasser (Gascoyne et al., 2004).
B.2.5- Etudes de physiopathologie
La clinique de l’accès palustre simple est désormais assez bien décrite et n’est plus beaucoup étudiée. En revanche, de nombreux travaux concernent le paludisme de la femme enceinte et le neuropaludisme, en raison de leur gravité :
- relations entre le placenta et le paludisme (Brabin et al., 2004), - les molécules de cytoadhérence et leurs implications physiologiques (Wassmer
et al., 2004, Cockburn, 2004 #654), - rôle de l’activation immune dans la physiopathologie du neuropaludisme (Hunt
& Grau, 2003), - les facteurs moléculaires liés à la pathogénicité du neuropaludisme chez l’hôte
(Lopansri et al., 2003 ; Wassmer et al., 2003b), etc.
B.2.6- Etudes épidémiologiques
Les étude épidémiologiques ont trois grands objectifs dans le cadre du paludisme :
adapter les stratégies de lutte en dressant à un moment précis une carte du paludisme : répartition des différentes espèces plasmodiales et de leurs éventuelles résistances, rôle des différentes populations présentes dans la transmission (Erhart et al., 2004), etc. prévoir et déjouer les épidémies Avec des informations socio-économiques et
météorologiques (Thomas, 2004) et des données épidémiologiques locales, les épidémies de paludisme sont souvent prévisibles. En particulier, les épidémies dues à l'homme (en liaison avec des projets de développement, irrigation, alevinage, digues, etc.) peuvent être prévues avec une grande précision (Organisation Mondiale de la Santé, 2003). Précéder et optimiser les essais vaccinaux : le profil de la maladie dans la
population permet de dresser une sorte de tableau témoin avant la campagne de vaccination (Appawu et al., 2004).

Bilan et orientation de la recherche sur le paludisme
45
B.2.7- Etude de l’immunité liée au paludisme
B.2.7.1- objectif
L’immunologie est une science extrêmement complexe. Il serait difficile de lister ici tous les travaux réalisés sur le paludisme, mais l’enjeu principal de l’étude des interactions hôte-Plasmodium est le développement d’un vaccin. Actuellement les grands thèmes de recherche sont :
- l’étude des mécanismes d’échappement à la réponse immunitaire chez le parasite par variation antigénique (famille des gènes var) (Flick & Chen, 2004),
- rôle de l’immunité humorale (Garraud et al., 2003; John et al., 2005), - rôle de l’immunité cellulaire (Stevenson et al., 2004; Good et al., 2005), - les liens entre l’immunité et l’échappement à la réponse immunitaire (Hisaeda
et al., 2004; Hisaeda et al., 2005), - l’immunité impliquée dans le paludisme de la femme enceinte (Staalsoe et al.,
2004), - l’immunité impliquée dans le neuropaludisme (Aucan et al., 2003), - l’immunité interspécifique face à différentes espèces plasmodiales (Mayxay et
al., 2004), - l’efficacité et l’immunogénicité de différentes formulations d’antigènes pour
élaborer ou tester un vaccin (Haddad et al., 2004).
B.2.7.2- principaux résultats
Un des premiers constats est que l'expression clinique du paludisme peut varier en fonction des souches, de l'hôte et de facteurs environnementaux. Des individus soumis aux mêmes conditions ne développeront pas des accès de gravité semblable. Il existe au plan individuel un état de « prémunition ». Cette « prémunition » présente trois caractéristiques (Rosier et al., 1999) :
• elle est longue à apparaître : après plusieurs années seulement chez les sujets qui vivent en zone d'endémie, où ils sont régulièrement exposés à des piqûres de moustiques infectés ;
• cette protection n'est pas dirigée contre le parasite lui-même : celui-ci continue à infecter le sujet prémuni, mais il ne déclenche pas de maladie grave ;
• enfin, cette immunité reste précaire, puisqu'elle disparaît après quelques années passées hors de la zone d'endémie (Bouchaud et al., 2005).
L’immunité liée au paludisme est très complexe, faisant intervenir à la fois des facteurs humoraux (Garraud et al., 2003) et cellulaires en fonction des stades :
1. Les sporozoïtes : ce sont des anticorps qui bloquent l’invasion des hépatocytes. Pour une bonne protection, il faut de façon permanente un niveau élevé d’anticorps. Un seul sporozoïte qui échappe à ce système peut provoquer l’infection.
2. Les hépatocytes infectés : immunité essentiellement cellulaire faisant intervenir
principalement les lymphocytes T CD8+ et CD4+ et les cellules NK (Natural Killer).

Bilan et orientation de la recherche sur le paludisme
46
Le mécanisme le plus important, d’après des études sur souris, consiste en la production de monoxyde d’azote (NO) par les hépatocytes infectés. Cette production, aux propriétés antipaludiques, est induite par les IFN-γ (interférons) produits par les CD8+.
3. Les mérozoïtes : mécanisme essentiellement humoral, les anticorps bloquent l’invasion
des érythrocytes et interviennent dans les mécanismes d’opsonisation. 4. Les érythrocytes infectés : à la différence des hépatocytes, en raison de l’absence de
complexe majeur d’histocompatibilité (CMH) de classe 1 sur les globules rouges, la cytotoxité cellulaire n’est pas active. En revanche, les érythrocytes infectés peuvent exprimer à leur surface des antigènes parasitaires. Ainsi, la réponse immunitaire, complexe, fait à la fois intervenir des cytokines produites par les cellules T CD4+ (action antipaludique) et des anticorps (Hviid, 2005). Ces anticorps interviennent dans les mécanismes d’opsonisation (avec les monocytes), d’ADCC (antibody-dependant cellular cytotoxicity), de lyse cellulaire par le complément et de blocage de l’adhésion aux cellules endothéliales.
5. Les stades sexués : les anticorps bloquent la fécondation ou le passage de l’ookinète
dans les glandes salivaires du moustique. 6. Les toxines : elles sont libérées par les schizontes mâtures qui éclatent et seraient
responsables, par l’intermédiaire des TNF-α (tumor necrosis factor), de la fièvre et de certaines complications (anémie, neuropaludisme). Ce sont, là encore, des anticorps qui bloquent ces toxines.
B.2.8- Etude des apparitions de résistance aux anti-paludiques
B.2.8.1- objectifs et méthodes
Le développement de résistances est l’une des principales causes de l’accroissement de la morbidité et de la mortalité liées au paludisme en Afrique. Etudier ces résistances pour mieux les prévenir est donc crucial. Trois grandes techniques permettent d’étudier ces résistances :
la surveillance post-AMM (Autorisation de Mise sur le Marché) de sensibilité ou de résistance des souches sur le terrain par rapport à une ou plusieurs molécules (de Monbrison et al., 2003 ; Tanabe et al., 2004) les essais d’induction in vitro de ces résistances pour anticiper et donc contrer
leur apparition in vivo. Les propriétés pharmacocinétiques de la molécule (taux de parasites tués par cycle, demi-vie de la molécule, effets secondaires, etc.) sont étudiées, ainsi que la possibilité d’apparition de résistance de novo en fonction de leur cible (faut-il une simple mutation ou plusieurs pour obtenir un mutant résistant et viable ? etc.). les modèles mathématiques pour anticiper l’apparition et/ou l’expansion des
résistances (Hastings & D'Alessandro, 2000; Watkins et al., 2005).

Bilan et orientation de la recherche sur le paludisme
47
B.2.8.2- principaux résultats
Le premier événement de l’apparition de résistance est d’origine génétique. C’est un événement rare, spontané et indépendant de la molécule utilisée. Ainsi pour la chloroquine, il existe environ une chance sur 1020 multiplications parasitaires de voir apparaître une telle mutation car elle est multigénique. En revanche, pour l’apparition de résistance à l’atovaquone, un seul point de mutation est suffisant, rapportant ce taux à 1 pour 1012.
0
5
10
15
20
dans lemoustique
inoculum desporozoïtes
mérozoïtesdu foie
symptômesde paludisme
paludismesévère
dans lemonde
aujourd'hui
log
10 p
aras
ites
Figure 6 : nombre de parasites présents (échelle logarithmique) de l’inoculation par l’anophèle jusqu’à la fin du développement chez l’hôte et dans le monde. Ainsi, un adulte dont la parasitémie est de 2% enferme 1012 parasites. Il est évident qu’un individu à forte parasitémie présente plus de risques de création de résistance qu’un individu faiblement parasité. La résistance aurait donc, en suivant ce principe, plus de chances d’apparaître dans les régions à forte transmission où l’on rencontre des taux élevés de parasites (1010 à 1011 parasites) de façon asymptomatique. Mais ce raisonnement n’est valable que pour l’apparition des souches résistantes ; sa diffusion, d’abord au sein de l’hôte puis parmi d’autres hôtes repose sur d’autres conditions. Les facteurs qui favorisent ensuite l’expansion de la résistance sont assez proches de ceux qui concernent les antibio-résistances :
- la faible immunité de l’hôte (cas des zones de faible transmission, des enfants des zones à forte transmission, des personnes atteintes du SIDA, etc.),
- les molécules éliminées lentement qui restent alors à faible dose dans l’organisme (Hastings et al., 2002),
- les vomissements ou les mauvaises prises de traitement qui induisent aussi la persistance de doses sub-thérapeutiques dans l’organisme,
- la possibilité de résistances croisées à d’autres molécules (la souche n’est alors plus freinée par aucune molécule),
- l’augmentation du nombre de piqûres de moustiques (cas des femmes enceintes et des zones de forte transmission).
Ainsi, finalement, en tenant compte des risques d’apparitions mais aussi de diffusions, les résistances apparaissent préférentiellement chez les individus peu immuns, plutôt dans les zones de faible transmission (White, 2004).
A l’échelle d’un individu

Bilan et orientation de la recherche sur le paludisme
48
B.2.9- Développement de nouvelles stratégies thérapeutiques
Les chimiorésistances du plasmodium à un nombre croissant de molécules imposent de trouver continuellement de nouvelles solutions thérapeutiques (Danis, 2003). Deux options sont possibles : la première consiste en de nouvelles associations d’antipaludiques (récents ou anciens), la deuxième voie consiste à développer de nouvelles molécules.
B.2.9.1- développement de nouvelles associations thérapeutiques
L’émergence de trop nombreuses résistances durant ces dernières années a fait basculer la démarche du praticien : la prescription doit non seulement avoir une visée thérapeutique mais aussi s’inscrire dans un objectif de lutte contre l’apparition des résistances. Les associations de molécule permettent de remplir ce double rôle : si on utilise deux molécules possédant des modes d’actions différents (et par là un mécanisme de résistance différent), alors la probabilité de développer une résistance pour l’association est le produit de la probabilité d’apparition de chaque résistance. Par exemple, si une molécule A et une molécule B sont chacune associée à une probabilité de résistance de 1 sur 1012, la probabilité de voir apparaître une résistance à l’association A-B est de 1 sur 1024. Or le nombre de parasites présents dans le monde sur une année étant inférieur à 1020, cela ramènerait l’intervalle d’apparition d’une telle résistance à une tous les 10 000 ans (White, 2004). Pour les molécules ayant déjà des résistances, les associations conseillées sont :
• chlorproguanil-dapsone (Lang & Greenwood, 2003), • chlorproguanil-dapsone-artésunate, • chloroquinine- sulfadoxine-pyréthamine (Ndyomugyenyi et al., 2004), • atovaquone-proguanil (Danis & Bricaire, 2003), • atovaquone-proguanil-artésunate (Giao et al., 2003), • amodiaquine- sulfadoxine-pyréthamine (Dorsey et al., 2003).
Par ailleurs, parmi toutes les résistances rencontrées ces dernières années, aucune n’a pu vraiment être mise en évidence pour l’artémisinine ni pour ses dérivés, sauf cas isolé et controversé (Sahr et al., 2001) et même in vitro il n’a pas été possible d’en créer. Ainsi, l’artémisinine représente dans de nombreuses zones le dernier rempart contre les souches multirésistantes de P. falciparum (Van der Meersch, 2005) et il convient elle aussi de l’associer pour la protéger (Adjuik et al., 2004). Cinq associations sont proposées :
• artésunate-amodiaquine (Adjuik et al., 2002), • artésunate-sulfadoxine-pyréthamine (Dorsey et al., 2003), • artéméther-luméfantril (Omari et al., 2004)qui semble la plus efficace (Mutabingwa et
al., 2005), • dihydroartémisinine-pipéraquine (Denis et al., 2002), • artésunate-méfloquine (Suputtamongkol et al., 2003),
Le projet Roll Back Malaria s’est fixé deux objectifs à atteindre d’ici 2005 pour l’Afrique : un traitement efficace disponible immédiatement pour au moins 60% des malades et une prophylaxie intermittente systématique pour 60% des femmes enceintes. Les thérapies de

Bilan et orientation de la recherche sur le paludisme
49
première intention retenues par l’OMS dans le cadre de ce projet sont les associations contenant l’artémisinine. Ces associations semblent être un moyen simple et efficace de prévenir les résistances, mais sur le terrain elles se heurtent encore au problème du coût (Whitty et al., 2004). En effet, un tiers des décès dus au paludisme concerne des pays en guerre (Allan, 2001). Dans de tels pays, 75% de la population est incapable de se procurer un traitement complet de chloroquinine (Allan et al., 2001). Une simple association avec le sulfadoxine-pyréthamine multiplie ce coût par plus de 10 par traitement, ce qui n’est donc pas envisageable pour la plupart des malades.
0,059 0,067
1,75 1,84
4,41
1,97
0
1
2
3
4
5
chloroquine sulfadoxinepyréthamine
méfloquine artésunatesulfadoxinepyréthamine
artésunateméfloquine
artémétherluméfantril
Figure 7 : coût des différentes thérapeutiques antipaludiques
adapté d’après les données de l’OMS (Ondari, 2004) Enfin, ces associations thérapeutiques doivent s’accompagner de la prévention d’autres maladies liées au paludisme. La campagne de Roll Back Malaria (OMS) devrait être jumelée à celles d’autres maladies telles que les filarioses, onchocercoses, helminthoses et autres parasitoses qui suivent la même distribution géographique que le paludisme (Molyneux & Nantulya, 2004). Ces maladies touchent aussi les plus démunis et, responsables d’anémies, sont des facteurs aggravants du paludisme (Brabin et al., 2003). Sans aide supplémentaire, l’adoption d’une polythérapie en première intention risque donc d’avoir des conséquences désastreuses : l’exclusion simple des malades les plus démunis ou des traitements donnés à plus faible dose ou sur de plus courtes périodes, favorisant ainsi justement l’émergence de résistance (Liden & Nantulya, 2004). La solution semble donc plus politique que scientifique.
B.2.9.2- développement de nouvelles molécules préventives et/ou curatives
Les associations de traitements n’étant pas si simples à appliquer de façon systématique, il est indispensable de développer en parallèle de nouvelles armes thérapeutiques. Les récents
Coû
t moy
en d
’un
traite
men
t pou
r un
adul
te (e
uro)

Bilan et orientation de la recherche sur le paludisme
50
progrès dans le domaine de la biologie moléculaire ont permis de mieux connaître le fonctionnement intracellulaire du parasite. De nouvelles stratégies offensives à son encontre sont apparues, avec le développement de nouvelles molécules comme le MC(1) (Satayavivad et al., 2004), le 2-hydroxy-1-naphthylaldehyde-isonicotinoyl-hydrazone et le 2-hydroxy-1-naphthylaldehyde-4-phenyl-3-thiosemicarbazone (Walcourt et al., 2004) ou les trioxaquines (Dechy-Cabaret et al., 2004). Les principales nouvelles cibles parasitaires visées sont (Pradines et al., 2003) :
• le métabolisme des lipides → blocage du transporteur de choline en utilisant un analogue (Wengelnik et al., 2002),
• les protéases et la dégradation de l’hémoglobine → inhibiteurs des protéases (Nezami et al., 2003),
• les interactions avec le transport des molécules (glucose, acide lactique, etc.) → inhibition des transports (Desai et al., 2000),
• les interactions avec le métabolisme du fer → chélateurs de fer (Hammadi et al., 2003),
• l’apicoplaste → inhibiteurs de l’apicoplaste (Foth et al., 2003), • la transduction du signal → inhibiteurs enzymatiques (Syin et al., 2001), • les métallocènes → complexes métalliques capables d’augmenter l’activité de
composés biologiques (Pradines et al., 2002). Par ailleurs, si la plupart des antipaludiques actuels ont été développés par synthèse et par screening, cette méthode est assez coûteuse. Sur les 250 000 molécules synthétisées depuis les années 1970, seules la méfloquine et l’halofantrine sont passées au stade de développement comme antipaludiques pour un coût de plus de 120 millions d’euros (Oketch-Rabah & Mwangi, 1998). Enfin, les plantes médicinales sont considérées comme une source potentielle de nouveaux médicaments (Benoit-Vical et al., 2003). Certains des antipaludiques actuels les plus importants dérivent d’extraits de plante (Etkin, 2003) : la quinine provient du quinquina (Dobson, 1998) tandis que le dernier composé de la pharmacopée occidentale, l’artémisinine, dérive d’Artemisia annua, fébrifuge chinois traditionnel utilisé depuis 2000 ans (Rath et al., 2004). La médecine traditionnelle n’a pas fini de livrer ses secrets et elle représente une voie de recherche non négligeable (Caraballo et al., 2004).
B.2.10- Développement d’un vaccin antipaludique
Il n’existe actuellement aucune forme commercialisée de vaccin contre le paludisme, même si de nombreuses recherches sont menées depuis près d’un demi-siècle.
B.2.10.1- espoirs et contraintes
Il existe trois grandes raisons de croire en la faisabilité d’un vaccin anti-palustre, justifiant les efforts des recherches vaccinales menées jusqu’ici (Moorthy et al., 2004) :
o l’existence d’une immunité acquise dans les pays de forte endémie (Sergent & Parrot, 1935; Roberts, 2003). Cette immunité conduit d’abord à une réduction de la

Bilan et orientation de la recherche sur le paludisme
51
mortalité et des complications puis à une diminution des signes cliniques et enfin à une réduction de la parasitémie à des niveaux bas voire indétectables.
o Les travaux d’injection de sporozoïtes irradiés ont conduit à une certaine protection contre l’infection (Nussenzweig et al., 1972; Luke & Hoffman, 2003), que ce soit chez la souris (Nussenzweig et al., 1967), le singe (Collins & Contacos, 1972) ou l’homme (Rieckmann et al., 1979).
o Le transfert passif d’immunoglobulines à des individus naïfs infectés par P. falciparum permet d’éliminer le parasite du sang (Cohen et al., 1961). C’est ainsi que le transfert de sérum recueilli sur des enfants africains « immuns » a aidé des enfants thaïs en proie à une recrudescence de paludisme (Sabchareon et al., 1991).
Néanmoins les chercheurs se heurtent à plusieurs problèmes :
• l’existence de plusieurs espèces de Plasmodium dont chacune possède des souches génétiquement distinctes,
• l’existence de plusieurs stades de développement du parasite, • l’existence de stades intracellulaires, • l’extraordinaire capacité d’évasion immunitaire du parasite (par polymorphisme
notamment), • la complexité des réponses immunitaires de l’hôte, • les variations génétiques humaines qui conduisent à des réponses immunes
différentes, • la « fragilité » de l’immunité acquise naturellement : elle exige un contact
important, continu, de longue durée avec le parasite et elle n’est que partielle et brève. (Alors comment un vaccin peut-il faire mieux que ce que la nature « peine » à faire ?)
• L’absence de modèle animal tout à fait complet (sur le plan pathogène et immun) et à caractère prédictif,
• l’absence d’une solide corrélation entre la protection in vitro et in vivo, • le manque d’intérêt des industriels pharmaceutiques face à une maladie qui affecte
les populations défavorisées, la recherche étant essentiellement soutenue par des fonds publics.
Le choix du stade de développement constitue le premier enjeu pour créer un vaccin contre le paludisme. Mais la question du choix du ou des antigènes à incorporer à ce vaccin est aussi cruciale (Webster & Hill, 2003; Duffy et al., 2005). La séquence génomique du parasite est désormais disponible, offrant aux chercheurs un choix d’environ 5 300 gènes codant pour des protéines comme cible potentielle (Gardner et al., 2002).
B.2.10.2- étapes de développement d’un vaccin
Le développement du vaccin comporte trois phases : la recherche et le développement, les essais pré-cliniques, puis les essais cliniques.
La première étape consiste en l’identification, le clonage et la caractérisation d’un antigène ciblé par un mécanisme immun susceptible d’être protecteur (les modèles alors utilisés sont en général des modèles in vitro et des modèles murins). La seconde étape consiste en l’évaluation de l’innocuité, de l’immunogénicité et de
l’efficacité du futur vaccin (avec éventuellement différents dosages, différents adjuvants, différents calendriers vaccinaux, etc.) chez le modèle primate non-humain.

Bilan et orientation de la recherche sur le paludisme
52
La troisième étape, le passage à l’humain, se subdivise en quatre phases (voir tableau 2).
paramètre évalué sujets d’étude épreuve d’infection
méthode d’évaluation
Phase I innocuité immunogénicité volontaires non immuns aucune individuelle
Phase II a efficacité volontaires non immuns moustiques d’élevage
absence vs présence de
forme sanguine 1
Phase II b efficacité petit effectif de zone endémique naturelle
absence vs présence
d’infection, délai entre 2 infections
Phase III efficacité
effectif un peu plus large ; provenant de zones endémiques à
caractéristiques épidémiologiques
différentes
naturelle
zones de caractéristiques
épidémiologiques comparables, comparaison
groupe vacciné-groupe témoin :
attente d’une baisse de
l’incidence ou du nombre de cas
graves
Phase IV interactions vaccinales,
effet à long terme
large effectif, populations déjà dans
un programme vaccinalnaturelle
1 mise en place immédiate d’un traitement si apparition d’une forme sanguine (ce qui limite beaucoup les essais, en particulier le développement de vaccins dirigés contre les formes sanguines ; en effet, ce type de vaccin n’est en général effectif qu’après un léger développement sanguin du parasite).
Tableau 2 : différentes phases d’essais cliniques de vaccin antipaludique
Deux types de vaccins posent des problèmes particuliers pour leur passage à l’humain.
1- Les vaccins de stade érythrocytaire : il est impossible d’effectuer des infections d’épreuve chez des volontaires européens vaccinés avec des molécules appartenant au stade érythrocytaire. En effet, l’apparition des symptômes est concomitante au début de la détection des premiers parasites dans le sang en raison de l’absence d’immunité des européens. Dans ces conditions, les essais de protection, c’est-à-dire de phase II - III devront être conduites sur le terrain en Afrique ; cela nécessite donc de répéter les phases I chez des adultes africains sains puis de vacciner des populations exposées, principalement des grands enfants et des adolescents. Tout cela demande alors beaucoup de temps.

Bilan et orientation de la recherche sur le paludisme
53
2- Les vaccins bloquant la transmission : cela soulève certains problèmes éthiques particuliers. En effet, ces vaccins n’offrent aucun bénéfice direct à l’individu vacciné, mais ils sont destinés à empêcher que cette personne ne transmette le paludisme aux autres. Des bénéfices individuels sont attendus uniquement si une proportion importante de personnes dans la communauté est vaccinée. Ainsi, il existe un danger que les investigateurs soient enclins à exercer une pression sur les membres de la communauté afin d’assurer un taux élevé de participation. De plus, il sera nécessaire de vacciner les adultes, alors qu’ils peuvent n’être exposés qu’à un risque relativement faible de paludisme grave dans les zones de transmission intense. Il se peut qu’ils retirent peu de bénéfice de la vaccination, mais ils seront exposés à tout effet adverse du vaccin. Ce problème sera important si les effets secondaires de la vaccination ne sont pas mineurs.
Actuellement, aucun vaccin n’a encore atteint la phase IV. Seul le vaccin SPf66 a pu attendre la phase III (Carcaboso et al., 2003), les autres candidats sont en phase I et II.
B.2.10.3- les différents antigènes candidats
Si l’on considère l’ensemble du cycle effectué par le parasite, il existe, au sein des trois étapes, cinq types de cible pour un vaccin, auxquelles on peut ajouter la cible des toxines (tableau 3) :
1. étape pré-érythrocytaire : les cibles sont les sporozoïtes libres (le problème est que les sporozoïtes ne demeurent que quelques minutes dans le sang) et les hépatocytes infectés, afin de bloquer l’infection avant les symptômes. Les premiers travaux sont ceux de Nussenzweig dans les années 60, avec l’utilisation de sporozoïtes irradiés (Nussenzweig et al., 1967). Les principaux bénéficiaires de ce type de vaccin seraient les enfants des zones endémiques et les voyageurs et résidents non-immuns de zones de faible transmission.
2. Etape sanguine asexuée : les cibles sont les mérozoïtes (formes libres, certes
fugaces, mais qui reviennent plusieurs fois au cours d’un cycle) et les érythrocytes infectés. Les premiers travaux dans ce domaine sur les primates ont plus de 25 ans (Mitchell et al., 1975). Il s’agit là de réduire le niveau de parasitémie ; les principaux bénéficiaires de ce type de vaccin seraient les enfants des zones endémiques.
3. Etape sexuée : les cibles sont les formes sexuées du parasite, afin de bloquer la
transmission de la maladie après son expression chez l’hôte. Les principaux bénéficiaires de ce type de vaccin seraient les populations de zones de faible transmission. Un vaccin à ce stade ne protègerait donc pas l’individu vacciné, mais éviterait la propagation de la maladie (principe du vaccin dit « altruiste » qui est à associer avec des vaccins d’autres stades).
Pour chaque cible, les antigènes candidats à la vaccination sont choisis en fonction des connaissances de l’immunité de l’hôte.

Bilan et orientation de la recherche sur le paludisme
54
Cible Antigène candidat
Sporozoïtes
CircumSporozoite Protein (CSP) Thrombospondin-Relatited Adhesive Protein (TRAP)
Sporozoïte And Liver-Stage Antigen (SALSA) Sporozoïtes Threonine and Asparagines-Rich Protein (STARP)
Stades hépatiques
CSP Liver-Stage Antigen (LSA)-1 et –3
SALSA STARP
Mérozoïtes
Merozoite Surface Orotein (MSP)-1, -2, -3, -4 et –5 Erythrocyte-Binding Antigen (EBA)-175
Apical Membrane Antigen (AMA)-1 Rhoptry-Associated Protein (RAP)-1 et –2
Acidic-Basic Repeat Antigen (ABRA) Duffy-Binding Protein (DBP) (P. vivax)
Stades sanguins
Ring Erythrocyte Surface Antigen (RESA) Serine-Rich Protein (SERP)
Erythrocyte Membrane Protein (EMP)-1, -2 et –3 GLUtamate-Rich Protein (GLURP)
Toxine GlycosilPhosphatidylInositol (GPI) Stades sexués Ps25, Ps28, Ps28/45 et Ps230
Tableau 3 : liste des principaux antigènes candidats au vaccin contre le paludisme
(Carvalho et al., 2002)
B.2.10.4- les différentes formes vaccinales
La vaccinologie progresse très rapidement depuis quelques années et ces progrès profitent à la recherche sur le paludisme (Webster et al., 2003). Les vaccins peuvent se présenter sous plusieurs formes (Genton & Corradin, 2002) :
• vaccins contenant les parasites entiers, comme des sporozoïtes irradiés (très immunogènes mais difficiles à réaliser) ou déplétés (Menard, 2005; Mueller et al., 2005; Waters et al., 2005),
• vaccins contenant une protéine recombinante, exprimée par une bactérie : Escherichia coli (Planson et al., 2003), BCG de Mycobacterium bovis (Zheng et al., 2003), Pichia pastoris (Zou et al., 2003), etc.
• vaccins contenant un peptide de synthèse, • vaccins à ADN, peu immunogènes (March, 2004), en particulier lorsque l’on associe
plusieurs plasmides ensemble (Sedegah et al., 2004).
Les vaccins comportant des gènes peuvent être insérés dans un plasmide ou dans un vecteur vivant, ce qui rend alors les vaccins bien plus immunogènes. Ce vecteur est un virus modifié, capable d’exprimer les gènes correspondant aux antigènes plasmodiaux visés. Parmi les virus les plus courants on trouve : les Baculovirus (Yoshida et al., 2003), les Poxvirus, (Sedegah et al., 2003), les Ankara virus (Moorthy et al., 2003a; Moorthy et al., 2003b), les virus Lister défectifs (Ober et al., 2002), etc.
Le rôle de l’adjuvant semble lui aussi primordial : il s’agit de développer un adjuvant dénué de toxicité et en même temps capable de stimuler la réponse immunitaire.

Bilan et orientation de la recherche sur le paludisme
55
De l’historique adjuvant de Freund (extrêmement efficace mais mal toléré donc inapplicable aux humains, (Freund et al., 1945)), aux plus récents montanides ISA720 et ISA 51 (Hirunpetcharat et al., 2003), CpG oligodéoxynucleotides (ODN) (Kumar et al., 2004), QS 21 (Carcaboso et al., 2004) ou autre AS02 (Garcon et al., 2003), en passant par le classique alun (hydroxyde d’aluminium (Sattabongkot et al., 2003)), les adjuvants suscitent à eux seuls bon nombre de recherches dans le domaine vaccinal. Enfin, le mode et la voie d’administration vaccinale sont aussi importants (Carcaboso et al., 2004).
B.2.10.5- exemples de vaccins en cours de développement
Quatre vaccins sont très étudiés actuellement (Graves & Gelband, 2003) :
- le vaccin SPf66 (vaccin contre les stades sanguins asexués) C’est un peptide de synthèse linéaire comprenant plusieurs épitopes, dont l’adjuvant est en général l’alun. Les 10 essais cliniques de ce vaccin donnent des résultats assez hétérogènes : dans 4 essais africains, il n’a pas montré d’efficacité, mais dans 5 études hors Afrique, le nombre de primo-infections a diminué. Ce vaccin a une efficacité globale de 23% mais seulement de 2% chez les enfants africains, le groupe le plus sensible (Galindo et al., 2000). Ce vaccin devrait donc faire l’objet d’études plus poussées en Amérique du Sud ou en Afrique, mais alors avec une formulation différente.
- Le vaccin CSP-NANP (vaccin contre les stades pré-érythrocytaires) Il est de type protéine recombinante fusionnée avec l’antigène S du virus de l’hépatite B. Malheureusement, sur 3 essais, il n’a pas été démontré de réelle efficacité (Graves et al., 2003).
- Le vaccin Combination B (vaccin contre les stades sanguins asexués) Ce vaccin est dirigé contre les stades asexués du parasite, qui comprend les antigènes MSP-1, MSP-2 et RESA avec l’adjuvant Montanide ISA720. Il obtient une efficacité de 62 % dans la réduction de la parasitémie chez les enfants de Papouasie Nouvelle Guinée (Genton et al., 2000; Genton et al., 2003). Ces résultats variaient selon que les individus de l’étude avaient ou non été préalablement traités avec l’association sulfadoxine-pyriméthamine (diminution de l’efficacité vaccinale si traitement). D’autre part, la prévalence d’infection était réduite si le vaccin contenait l’antigène MSP-2 par rapport aux autres sous-types vaccinaux.
- Le vaccin RTS,S/AS02 (vaccin contre les stades pré-érythrocytaires) Ce vaccin prometteur (Ballou et al., 1999), dirigé contre les sporozoïtes, est une protéine recombinante contenant une large portion C-terminale de CSP fusionnée à l’antigène S de surface du virus de l’hépatite B et est adjuvé avec AS02 (Garcon et al., 2003). Lors d’un essai sur un petit nombre d’adultes non-immuns aux USA, RTS,S a permis une forte protection contre l’infection à P. falciparum. Lors d’un essai sur des adultes semi-immuns en zone endémique (Gambie), il y a eu une réduction du nombre de cas de paludisme la seconde année suivant la vaccination, avec une efficacité vaccinal de 34 % (Alloueche et al., 2003). La réponse immunitaire cellulaire était assez bonne (Epstein et al., 2004 ; Pinder et al., 2004). L’étude a été poursuivie dans une cohorte d’enfants gambiens pour lesquels l’immunogénicité et l’innocuité du

Bilan et orientation de la recherche sur le paludisme
56
vaccin semblent satisfaisantes (Bojang et al., 2005). La formulation est la propriété de la firme GlaxoSmithKline (Heppner et al., 2005).
B.2.10.6- vaccin contre Plasmodium vivax
Concernant le développement de vaccin contre Plasmodium vivax, bien peu de recherches sont menées en comparaison à P. falciparum, plus meurtrier. Pourtant c’est le plasmodium le plus répandu dans le monde et sa capacité à créer des rechutes en fait un ennemi du développement économique des zones touchées. En outre, de récentes résistances aux antipaludiques ont été décrites. En effet, les premiers cas de résistance de P. vivax à la chloroquine ont été décrits en 1989 (soit 32 ans après les premières résistances décrites chez P. falciparum) en Papouasie Nouvelle Guinée (Rieckmann et al., 1989), puis en Indonésie et en Inde. L’extension de ces zones de résistance est certes bien moins rapide que pour P. falciparum (en raison de leur différence de cycle), mais il convient d’être très vigilant et sans doute d’accorder plus d’importance à ce Plasmodium (Mendis et al., 2001). Les antigènes CSP, MSP-1, DBP, Pvs25 et Pvs28, des protéines de surface de l’ookinète, (Tsuboi et al., 2003) ont déjà été étudiés et leur immunogénicité est testée sur modèle primate (Arevalo-Herrera & Herrera, 2001). Le Pvs 25H est désormais en phase I d’étude avec des résultats d’innocuité satisfaisants et une production d’anticorps qui permettraient le blocage de la transmission du parasite (Malkin et al., 2005). Pour conclure, il apparaît que le vaccin idéal serait un vaccin de type combiné : plusieurs antigènes de stade sanguin et/ou plusieurs antigènes de différents stades. Par ailleurs, dans de nombreux pays, un vaccin à la fois protecteur contre P. falciparum et P. vivax serait très utile (Kieny, 2003 ; Tsuboi et al., 2003).

DDEEUUXXIIEEMMEE PPAARRTTIIEE :: LLEESS PPRRIIMMAATTEESS AAFFRRIICCAAIINNSS EETT LLAA RREECCHHEERRCCHHEE SSUURR LLEE PPAALLUUDDIISSMMEE


Les primates africains et la recherche sur le paludisme
59
A- UN POINT SUR L’UTILISATION DES PRIMATES NON-HUMAINS DANS LA RECHERCHE Afin de mieux cerner le cas particulier du paludisme, il est important de connaître les implications des primates dans l’ensemble de la recherche. La place des primates non-humains (PNH) dans la recherche a toujours été particulière, même si actuellement la tendance est à la diminution du nombre d’animaux utilisés sous la pression des associations de protection de l’animal. En effet, ils constituent souvent la ligne de frontière entre la recherche médicale et l’application médicale qui en résulte chez l’humain. Par ailleurs les réactions concernant leur utilisation sont aussi plus vives que pour les autres animaux. Les PNH présentent certaines caractéristiques essentielles à la recherche :
- des similarités génétiques et physiologiques par rapport aux humains ; le groupe des primates inclut l’humain. L’utilisation de primates non-humains permet d’explorer des domaines caractéristiques partagés uniquement par les humains et les autres primates.
- Des similarités de susceptibilité aux maladies infectieuses : des centaines de maladies touchant la population humaine sont partagées par les primates non-humains.
- Des similarités de profil dans les maladies chroniques : athérosclérose, ostéoporose, hypertension, etc.
- La possibilité de contrôle de la reproduction : cela permet de tester des hypothèses génétiques spécifiques, ce qui est impossible chez les humains.
- La possibilité de vivre en environnement contrôlé : les PNH peuvent être maintenus dans des conditions identiques et contrôlées toute leur vie, ce qui est bien sûr impossible dans les populations humaines.
Tout au long du siècle dernier, les découvertes et les avancées médicales permises grâce aux primates non-humains sont nombreuses :
- années 1900 : découverte de la composition du sang et du plasma ; - années 20 : diagnostic et traitement de la fièvre typhoïde ; - années 30 : développement de l’anesthésie moderne ; - années 40 : découverte des rhésus sanguins et du traitement de l’arthrite rhumatoïde ; - années 50 : développement des vaccins polio et fièvre jaune et développement de la
chimiothérapie anticancéreuse ; - années 60 : développement de la transplantation cornéenne et de l’utilisation
thérapeutique de la cortisone ; - années 70 : découverte du traitement de la lèpre ; - années 80 : développement de la cyclosporine et des molécules anti-rejet, premier
vaccin contre l’hépatite B, développement de systèmes d’aide à la survie des prématurés ;
- années 90 : développement du vaccin contre l’anthrax et des multitraitements contre le VIH.

Les primates africains et la recherche sur le paludisme
60
Les données présentées ici sont européennes et nord américaines. En effet, les Etats-Unis, le Canada et l’Europe publient régulièrement des rapports officiels concernant l’utilisation des animaux à des fins scientifiques, ce qui ne semble pas être le cas ailleurs.
A.1- IMPORTANCE QUANTITATIVE DES PRIMATES PAR RAPPORT AUX AUTRES ANIMAUX
A.1.1- Cas de l’Amérique du Nord
Selon le rapport annuel du Département de l’Agriculture américain (USDA), les USA utilisaient en 2001 1,2 millions d’animaux pour la recherche (U.S. Department of Agriculture, 2002). Mais ces chiffres ne regroupent pas les oiseaux ni les rongeurs qui doivent représenter environ 85% des animaux utilisés en recherche (Library of Congress, 2000). Ainsi, le nombre total d’animaux utilisés s’élèverait plutôt au moins à 10 millions et certaines associations de protection animale évaluent ce nombre à 23 millions (Laboratory Primate Advocacy Group, 2002). Sur les 10 à 20 millions d’animaux utilisés en 2001, près de 50 000 sont des primates non-humains (57 000 en 2000), soit 0,2 à 0,5% des animaux. Quel que soit le nombre total d’animaux estimé au départ, l’utilisation des primates aux USA est élevée par rapport au reste du monde. Ceci s’explique sans doute par le fait qu’une grande partie des industries pharmaceutiques sont localisées dans ce pays. Au Canada, les données sont celles récoltées par le Conseil Canadien de Protection des Animaux (CCPA). En 2001, il relevait 2,1 millions d’animaux utilisés pour la recherche, mais il n’est pas précisé quelles espèces sont inclues dans ce nombre. Dans ce pays, seulement 1 883 primates étaient utilisés sur les 2,1 millions d’animaux, soit à peine 0,1%.
A.1.2- Cas de l’Europe
Le nombre total d'animaux utilisés à des fins expérimentales et à d'autres fins scientifiques en 1999 en Europe est estimé à 9,8 millions. On observe une baisse de l’ordre de 15 % du nombre d’animaux utilisés par rapport à 1996 (Commission des Communautés Européennes, 2003). La France est devenue le premier utilisateur d’animaux devant le Royaume-Uni et l’Allemagne en 2001, malgré une baisse supplémentaire de 15 % depuis 1999. Selon certains, il n'est d’ailleurs pas impossible que le niveau atteint en 2001 soit proche de la limite incompressible en dessous de laquelle la recherche ne peut fonctionner.

Les primates africains et la recherche sur le paludisme
61
rongeurs et lagomorphes
86,9%
carnivores0,36%
artiodactyles et périssodactyles
1,26%
prosimiens et simiens0,09%autres
mammifères0,05%
oiseaux4,7%animaux à sang
froid6,6%
Avec 10 000 individus impliqués, les primates représentent à peine 0,1% des animaux utilisés en Europe et ce nombre est en légère diminution par rapport à une enquête de 1997. Les principaux états concernés sont la France, la Hollande, le Royaume Uni (3000 singes en 2000), l’Allemagne (2000 singes en 2000) et l’Italie. La France utilisait environ 3800 individus en 2001. Le nombre de primates utilisés a baissé de 26% entre 1990 et 1999.
A.2- UTILISATION DES PRIMATES PAR SECTEUR DE RECHERCHE
En Europe, 97% des primates sont utilisés pour la recherche, la mise au point de nouveaux traitements pour l'homme ainsi que pour s'assurer de la sécurité de leur utilisation.
production et qualité en médecine humaine
13%
autres3%
études en biologie
fondamentale18%
développement en médecine humaine et vétérinaire
13%essais
toxicologiques et autres
évaluations de sécurité
53%
Figure 9 : domaines d’utilisation des primates dans la recherche en Europe en 1999
(Commission des Communautés Européennes, 2003)
Figure 8 : catégories d’animaux utilisés dans la recherche en Europe en 1999 (Commission des Communautés Européennes, 2003)

Les primates africains et la recherche sur le paludisme
62
Actuellement les primates servent principalement :
- à l’élaboration de vaccin et de nouveaux traitements contre le VIH, l’hépatite C, le RSV ;
- à la meilleure compréhension de la biologie cérébrale, - au développement de traitements contre les leucémies et les lymphomes de l’adulte, - au développement de traitements contre les maladies d’Alzheimer et de Parkinson, - à l’élaboration de vaccin et de nouveaux traitements contre le paludisme.
Concernant les chimpanzés, ils sont désormais principalement utilisés pour les recherches sur l’hépatite C (Lanford et al., 2001), pour une meilleure connaissance de leur génome (Hacia, 2001) et pour des études cognitives.
A.3- REPARTITION DES DIFFERENTES ESPECES DE PRIMATES
Parmi les 200 espèces de primates, une trentaine est utilisée dans la recherche, mais seule une petite dizaine d’espèces est à l’origine de l’immense majorité des publications. Le choix d’un modèle parmi les primates résulte de la mise en balance de deux critères majeurs : son intérêt scientifique et sa disponibilité (aspects écologique, éthique et financier). Ainsi, parmi les 3 continents abritant des primates non-humains, les primates utilisés en recherche sont diversement représentés :
Le macaque est presque l’unique représentant asiatique dans les laboratoires et est de loin le primate le plus prisé en recherche (principalement les espèces rhésus et cynomolgus). Sur les 116 000 références obtenues avec le terme « primate » sur Pubmed, « macaca » en concerne 46 000, soit 40 % des publications. Ce primate a pour qualités principales sa petite taille (intérêt financier), sa facilité d’élevage (intérêt éthique et écologique) et sa sensibilité à de nombreuses maladies (SIDA, paludisme, troubles neurologiques liés au vieillissement, troubles articulaires, etc.).
Parmi les singes africains, le babouin, le vervet, le patas et le chimpanzé sont
les plus utilisés.
Les primates du continent américain ont six grands représentants dans la recherche : le singe écureuil, le singe de nuit, le singe araignée, le marmouset, le capucin et le tamarin.
A.3.1- Cas de l’Amérique du Nord
Au Canada, les 1 883 primates utilisés en 2001 se répartissaient comme tel : - macaque cynomolgus : 1 303 - macaque rhésus : 230 - singe écureuil : 166 - vervet : 35 - autres primates non-humains : 149

Les primates africains et la recherche sur le paludisme
63
Aux Etats-Unis, la politique actuelle consiste à diminuer les importations (19 000 singes en 2002 (California Biomedical Research Association, 2004)) en privilégiant les espèces élevées localement (National Center for Research Resources, 2002). Les USA se caractérisent également par l’utilisation de quelques 1500 chimpanzés, dont un grand nombre porteurs de HIV. Le Japon semble d’ailleurs être, avec les USA, l’un des derniers grands utilisateurs de singes anthropoïdes avec 107 chimpanzés répertoriés dans les laboratoires pharmaceutiques en 2002.
A.3.2- Cas de l’Europe
En Europe, seul le centre de primatologie de recherches biomédicales de Rikswijk en Hollande (BPRC) utilisait encore des chimpanzés en 2002. Seuls 6 chimpanzés étaient encore utilisés en 2004 pour 3 ans dans un protocole d’hépatite, mais une fois ce protocole terminé, ces animaux doivent être relâchés dans un sanctuaire en Espagne car la Hollande a récemment interdit l’utilisation de grands singes (Biomedical Primate Research Center, 2005). En Angleterre, les recherches invasives sur les chimpanzés ont été officiellement interdites depuis 1997. En France, en 2001, les primates représentaient 0,17 % des animaux utilisés avec :
- 3 072 Cercopithécidés (contre 2053 en 1993), - 589 Prosimiens (contre 392 en 1993), - 179 Cébidés (contre 240 en 1993).
Mais, comme en 1997, aucun singe anthropoïde n'a été utilisé (Commission des Communautés Européennes, 2003).
A.4- LES PRIMATES UTILISES POUR LA RECHERCHE SUR LE PALUDISME
Près de 5% des références concernant le paludisme impliquent des primates (sur les 37 000 références liées au terme « malaria », 1 600 incluent également le terme « primate » sur Pubmed). La répartition par espèce suit grossièrement celle des autres domaines de recherches avec quelques spécificités pour le paludisme.
Dans ce secteur de recherche aussi, le plus grand nombre de publications concernent le macaque (Macaca sp.), avec 45% des 1600 publications impliquant des primates référencées sur Pubmed. L’orang-outan (Pongo pygmaeus) et le gibbon (Hylobates sp.) sont aujourd’hui cités de façon anecdotique.
Le primate actuellement recommandé par l’OMS pour la recherche sur le paludisme,
est le singe de nuit, Aotus sp. (A. lemurinus, A. l. griseimembra, A. triviragtus, A. nancymai, A. vociferans) avec 25% des 1600 publications impliquant des primates référencées sur Pubmed. D’autres modèles sud-américains sont couramment utilisés, comme le saïmiri (Sciureus sp.), puis, de façon bien plus anecdotique, le tamarin (Saguinus sp.), le singe hurleur (Alouatta sp.), le singe araignée (Ateles sp.) et le capucin (Cebus sp.). Au total, les primates sud-américains sont impliqués dans 38% des références liées au paludisme et aux primates.

Les primates africains et la recherche sur le paludisme
64
Concernant les primates africains (17% des publications impliquant des primates sur Pubmed), le chimpanzé (Pan troglodytes) est, de loin, le plus fréquemment cité avec 164 références pour « pan + plasmodium ». Viennent ensuite les cercopithèques (Cercopithecus sp.), les babouins (Papio sp.), les mandrills et drills (Mandrillus sp.) et le groupe des lémuriens (Lemuridae).
B- LES PLASMODIUMS DES PRIMATES NON-HUMAINS Le genre Plasmodium, comme la plupart des protozoaires, présente un degré de spécificité d’hôte assez peu élevé. Il est donc présent largement dans l’ensemble du monde animal et a fortiori chez les primates non-humains (Kaup & Schwibbe, 2002). Sur les 200 espèces plasmodiales décrites, on en dénombre aujourd’hui plus de 25 chez les primates :
- 4 pour les grands singes, - 4 pour les gibbons, - 8 pour les cercopithèques, - 2 chez les cébidés, - 7 chez les lémuriens.
Ce nombre augmente encore avec les travaux les plus récents (de nouvelles sous-espèces mais aussi de nouvelles espèces de Plasmodium sont régulièrement isolées) mais certaines taxonomies demeurent incertaines.
B.1- HISTORIQUE
Les premières découvertes furent asiatiques, il y a près d’un siècle. Puis dans les années 20-30, beaucoup de travaux concernèrent les chimpanzés. En Asie, de nouvelles espèces plasmodiales furent mises en évidence dans les années 60. Enfin, depuis les années 80, la plupart des travaux de description de nouvelles espèces plasmodiales concernent les Lémuriens.
Plasmodium Hôte original Références de découverte
P. pitheci Pongo pygmaeus (orang-outan de Bornéo)
(Halberstaeder & Von Prowazek, 1907)
P. inui Macaca fascicularis (macaque crabier) (Halberstaeder et al., 1907)
P. cynomolgi Macaca fascicularis (macaque crabier) (Mayer, 1907)
P. brasilianum Cacajao calvus (ouakari)
(Gonder & Von Bernberg-Gossler, 1908)
P. reichenowi Pan troglodytes (chimpanzé) Gorilla gorilla (gorille) (Sluiter et al., 1922)
P. knowlesi Macaca fascicularis (macaque crabier)
(Sinton & Mulligan, 1932-1933)
P. gonderi Cercocebus torquatus atys (mangabé enfumé) (Sinton et al., 1932-1933)
P. schwetzi Pan troglodytes (chimpanzé) Gorilla gorilla (gorille) (Brumpt, 1939)

Les primates africains et la recherche sur le paludisme
65
P. hylobati Hylobates moloch (gibbon argenté de Java) (Rodhain, 1941)
P. simium
Alouatta fusca (singe hurleur brun) (Da Fonseca, 1951)
P. girardi Eulemur fulvus rufus (lémur à front roux) (Buck et al., 1952)
P. foleyi Eulemur fulvus rufus (lémur à front roux) (Buck et al., 1952)
P. coatneyi Anopheles hackeri * (Eyles et al., 1962a)
P. fieldi Macaca nemestrina (macaque à queue de cochon) (Eyles et al., 1962b)
P. lemuris Eulemur fulvus collaris (lémur à collier roux) (Huff & Oogstraal, 1963)
P. youngi Hylobates lar (gibbon à mains blanches) (Eyles et al., 1964)
P. cylesi Hylobates lar (gibbon à mains blanches) (Warren et al., 1965)
P. fragile Macaca sinica (macaque à toque) (Dissanaike et al., 1965b)
P. simiovale Macaca sinica (macaque à toque) (Dissanaike et al., 1965a)
P. jefferyi Hylobates lar (gibbon à mains blanches) (Warren et al., 1966)
P. silvaticum Pongo pygmaeus (orang-outan de Bornéo) (Garnham et al., 1972)
P. coulangesi Eulemur macaco macaco (lémur macaco) (Lepers et al., 1989)
P. percygarnhami Eulemur macaco macaco (lémur macaco) (Landau et al., 1989)
P. uilenbergi Eulemur fulvus fulvus (lémur brun) (Landau et al., 1989)
P. bucki Eulemur macaco macaco (lémur macaco) (Landau et al., 1989)
* seule espèce plasmodiale des primates découverte initialement dans un moustique Tableau 4 : liste des principales espèces plasmodiales de primates avec leur hôte original lors de leur découverte, adaptée d’après Collins (Collins & Aikawa, 1993)
Il semble que l’ancêtre du Plasmodium des primates soit Hepatocystis, un parasite ubiquiste et bénin des singes africains qui ne produit que des gamétocytes dans la circulation sanguine. Les premiers parasites sont donc vraisemblablement africains. L’infection des singes du Nouveau Monde viendrait des mouvements d’esclaves d’Afrique et d’Europe. D’ailleurs les deux plasmodiums des singes du Nouveau Monde, P. brasilianum et P. simium, sont d’un point de vue moléculaire très proches de P. malariae et P. vivax (Fandeur et al., 2000).

Les primates africains et la recherche sur le paludisme
66
B.2- LES PLASMODIUMS ET LEURS HOTES NATURELS
PRIMATES AFRICAINS Hôte naturel Plasmodium
Pan troglodytes (chimpanzé) P. schwetzi, P. reichenowi Gorilla gorilla (gorille) P. schwetzi, P. reichenowi Mandrillus leucophaeus (drill) P. gonderi Cercocebus torquatus atys (Cercocèbe enfumé) P. gonderi Cercocebus galeritus (Mangabey à ventre doré) P. gonderi
Tableau 5 a
PRIMATES ASIATIQUES
Hôte naturel Plasmodium Pongo pygmaeus (orang-outan de Bornéo) P. pitheci, P. silvaticum Hylobates lar (gibbon à mains blanches) P. eylesi, P. jeffreyi, P. youngi Hylobates moloch (gibbon argenté de Java) P. hylobati Macaca arctoides (macaque brun) P. cynomolgi Macaca cyclopsis (macaque de Formose) P. cynomolgi, P. inui Macaca fascicularis (macaque crabier) P. cynomolgi, P. coatneyi, P. fieldi, P.
inui, P. knowlesi Macaca mulatta (macaque rhésus) P. cynomolgi, P. inui Macaca nemestrina (macaque à queue de cochon) P. cynomolgi, P. fieldi, P. inui, P.
knowlesi Macaca radiata (macaque bonnet chinois) P. cynomolgi, P. fragile, P. inui Macaca sinica (macaque à toque) P. cynomolgi, P. fragile, P. simiovale
Tableau 5 b
PRIMATES AMERICAINS Hôte naturel Plasmodium
Alouatta fusca (singe hurleur brun) P. brasilianum, P. simiumAlouatta palliata (hurleur à manteau) P. brasilianum Alouatta seniculus straminea (hurleur rouge) P. brasilianum Alouatta villosa (hurleur noir) P. brasilianum Aotus sp.(singe de nuit) P. brasilianum Ateles fusciceps (singe araignée à tête brune) P. brasilianum Ateles geoffroyi (singe araignée de Panama à mains noires) P. brasilianum Ateles geoffroyi grisescens (singe araignée de Panama à mains noires)
P. brasilianum
Ateles paniscus (singe araignée noir) P. brasilianum Ateles chamek (singe araignée à tête noire) P. brasilianum Brachyteles arachnoides (muriqui) P. brasilianum, P. simiumCallicebus moloch ornatus (callicèbe gris) P. brasilianum Callicebus torquatus (callicèbe à fraise) P. brasilianum Cebus albifrons (sapajou à front blanc) P. brasilianum Cebus apella (capucin fauve) P. brasilianum Cebus capucinus (sapajou capucin) P. brasilianum Cebus capucinus capucinus (sapajou capucin) P. brasilianum

Les primates africains et la recherche sur le paludisme
67
Cebus capucinus imitator (sapajou capucin) P. brasilianum Chiropotes satanas chiropotes (saki capucin) P. brasilianum Lagothrix lagotricha cana (lagothrix d’Humboldt) P. brasilianum Lagothrix lagotricha infumata (lagothrix d’Humboldt) P. brasilianum Lagithrix lagotricha lagotricha (singe laineux gris) P. brasilianum Saimiri boliviensis (sapajou à tête noire) P. brasilianum Saimiri sciureus (singe écureuil) P. brasilianum Saguinus geoffroyi (tamarin) P. brasilianum
Tableau 5 c
Tableau 5 a, b, c : plasmodiums des primates simiens
Dans certaines classifications, on retrouve P. rodhaini, décrit chez le chimpanzé et le gorille et transmissible à l’homme. Il s’agirait en fait très probablement de P. malariae (Centre National de la Recherche Scientifique, 2004). Ainsi, si l’on excepte P. malariae, aucun primate ne semble naturellement sensible aux plasmodies humaines.
B.3- CYCLE DU PLASMODIUM CHEZ LES PRIMATES
Le cycle global du Plasmodium chez les primates est le même que celui des plasmodies humaines : passage de sporozoïtes de l’anophèle à l’hôte, développement dans le foie, invasion du sang par les mérozoïtes, transformation en gamétocytes et ingestion de ces derniers par le moustique. Comme P. vivax et P. ovale, P. cynomolgi, P. simiovale et P. fieldi produisent des stades hépatiques latents, les hypnozoïtes, responsables de rechutes. Les parasites des primates présentent en général une périodicité tierce, avec un cycle asexué toutes les 48h (comme P. falciparum, P. vivax et P. ovale) (Coatney et al., 1971).
C’est notamment le cas de tous les plasmodiums africains (sauf l’hypothétique P. rhodaini qui est quarte comme P. malariae). En Amérique, les cycles sont plutôt quartes (P. brasilianum) parfois tierces (P.
simium). En Asie, tous les cas existent : P. knowlesi a un cycle quotidien, P. inui est quarte, les
autres sont tierces.
B.4- CLINIQUE
A la grande différence de l’humain, les primates simiens autochtones ont des infections inapparentes ou de très légers troubles. Par exemple, P. schwetzi chez le chimpanzé provoque une infection légère avec parfois un peu de fièvre (Coatney et al., 1971). La maladie est de courte durée et la parasitémie est faible. Les rechutes sont rares et la guérison est spontanée. Une exception à cette règle concerne P. brasilianum, l’agent le plus commun de la malaria chez le saïmiri, qui provoque chez son hôte une anémie hémolytique bénigne, de l’apathie et une splénomégalie.

Les primates africains et la recherche sur le paludisme
68
Les symptômes peuvent cependant être plus marqués dans deux cas : - lorsque l’animal est immunodéprimé ou splénectomisé - lorsque l’on inocule expérimentalement un plasmodium chez un primate qui n’est pas l’hôte naturel (Centre National de la Recherche Scientifique, 2004) :
o P. knowlesi chez le macaque rhésus provoque une maladie grave parfois mortelle alors qu’il est asymptomatique chez son hôte, la macaque crabier.
o P. eylesi (du gibbon) entraîne une parasitémie élevée chez le mandrill. o P. knowlesi, P. coatneyi (2 plasmodiums du macaque crabier) et plus rarement
P. cynomolgi (commun au macaque crabier et au macaque rhésus) entraînent chez le macaque rhésus un ictère, une anorexie, de l’apathie, de la fièvre, une anémie et une splénomégalie chez l’animal non splénectomisé. Les femelles gestantes peuvent présenter des anémies sévères avec de sévères conséquences sur le foetus (Davison et al., 1998; Davison et al., 2000). On peut également fréquemment observer un oedème pulmonaire et/ou cérébral (Ibiwoye et al., 1993) ce qui n’est pas sans rappeler le neuropaludisme dû à P. falciparum.
En cas d’infection mortelle, les principales lésions retrouvées sont (Baskin, 1999) :
- poumons, foie et rate gris, - anémie, - présence dans les tissus de macrophages remplis de pigment plasmodial, - hémosidérose, - présence fréquente de thrombose et de coagulation intravasculaire avec accumulation
d’érythrocytes infectés, - oedème pulmonaire et/ou cérébral fréquent.
B.5- DIAGNOSTIC
Le diagnostic repose principalement sur le diagnostic direct (frottis et goutte épaisse) ; ce diagnostic est difficile lorsqu’il s’agit d’un Plasmodium naturel à cause de la faible parasitémie.
Une première attention doit être portée afin de ne pas confondre le genre Plasmodium avec d’autres parasites du groupe des Apicomplexa tels que Entopolypoides macaci ou Hepatocystis kochi qui sont aussi intra-érythrocytaires et fréquents chez le macaque rhésus, certains cercopithèques et le babouin (Ozwara et al., 2003b). Ces parasites sont plus petits que Plasmodium, remarquablement pléomorphiques, et ne produisent pas de pigment. L’infection qu’ils provoquent est toujours bénigne (Zeiss & Shomer, 2001), même chez des animaux splénectomisés, et ne répond pas aux antipaludiques. Ensuite, concernant le diagnostic d’espèce plasmodiale, il est parfois très délicat, comme par exemple dans le cas de P. brasilianum et P. simium qui sont respectivement morphologiquement similaires à P. malariae et P. vivax.
Les tests de recherche d’antigène (HRP2, LDH) peuvent être positifs pour
P. cynomolgi, P. coatneyi et P. knowlesi ; les autres espèces n’ont, à priori, pas été testées (Baskin, 1999).
La PCR permet, avec une extrême sensibilité, de détecter le parasite et d’exclure la
présence de plasmodiums humains. Cependant, assez peu d’amorces sont disponibles

Les primates africains et la recherche sur le paludisme
69
pour les plasmodiums des primates non-humains (principalement disponibles pour les plasmodiums de macaque, P. cynomolgi, P. knowlesi, et P. coatneyi).
La sérologie enfin s’applique bien aux chimpanzés, plus difficilement aux autres
espèces, et comporte les mêmes inconvénients que chez l’humain (long délai d’apparition des anticorps qui en fait un mauvais outil diagnostic).
B.6- THERAPEUTIQUE
La plupart des paludismes des primates peut être traitée à la chloroquine (7 mg/kg par voie intramusculaire ou per os pendant 5 jours). Cependant, le goût très amer des 4-aminoquinolines réduit leur usage aux voies intramusculaires ou aux gavages par sonde. De plus, la chloroquine agit sur les stades asexués sanguins mais pas sur les stades hépatiques ni sur les gamétocytes (Cogswell, 2000).
La méfloquine est aussi utilisable (20 mg/kg, une dose unique per os) et son emploi
est possible en cas de résistance à la chloroquine (Cogswell, 2000).
L’association pyriméthamine-sulfadoxine est pratique, car il suffit d’une injection unique pour obtenir une bonne clearance sanguine (utilisée au CIRMF chez le chimpanzé et le mandrill aux mêmes doses que chez l’humain).
Les stades hépatiques sont traités à la primaquine (3mg/kg, 7 jours). Ceci est
particulièrement important lors d’infection naturelle ou d’inoculation expérimentale par des sporozoïtes de P. cynomolgi, P. simiovale ou de P. fieldi qui possèdent des hypnozoïtes (Cogswell, 2000). En raison de leur toxicité augmentée lorsqu’elles sont administrées ensemble, la chloroquine et la primaquine doivent être données séparément. Ainsi, par exemple, le traitement classique des saïmiris d’expérimentation consiste en une série d’injections intramusculaires de chloroquinine (2,5 mg/kg) pendant 7 jours suivie par l’administration per os de primaquine (0,75 mg/kg) pendant 14 jours (Pingard, 1992).
Les thérapies plus récentes comme l’association atovaquone-proguanil ou artemether-
luméfantrine doivent être aussi peu utilisées que possible afin de limiter l’extension des résistances.
C- INTERETS DES PRIMATES AFRICAINS DANS LA RECHERCHE SUR LE PALUDISME
C.1- INTERET EN TANT QUE MODELE ANIMAL
Au cours des dernières décennies, des progrès remarquables ont permis de remplacer un grand nombre d’expériences sur les animaux par des méthodes dites alternatives. Les méthodes alternatives appartiennent à deux grandes catégories :
- des méthodes « synthétiques » : les méthodes de bio-informatique structurale et de criblage in silico ainsi que le recours à l'imagerie médicale ;

Les primates africains et la recherche sur le paludisme
70
- des méthodes cellulaires de type in vitro : le recours aux structures cellulaires ou subcellulaires, tissulaires et aux organes isolés, l'utilisation d'organismes moins évolués (invertébrés, plantes ou micro-organismes) ou de cellules souches embryonnaires.
C.1.1- Par rapport aux modèles bio-informatiques
Les progrès en informatique ont fait un immense bond durant ces dernières décennies. Les biochimistes ont développé des modèles mathématiques qui étudient assez précisément la cible des molécules recherchée pour en déduire la structure de la molécule ; en résumé, cela revient à mieux étudier la serrure, la modéliser, pour pouvoir en déduire la forme de la clé nécessaire (Volker & Brown, 2002). Cette technique est donc intéressante pour développer de nouvelles thérapies antipaludiques à partir des connaissances du modèle plasmodium ou de la cellule infectée par le plasmodium. Certains ont vu en l’apparition de ces logiciels la fin de l’utilisation de l’animal pour la recherche. C’est malheureusement impossible, pour deux raisons principales :
1- parce que ces logiciels ne fonctionnent que parce qu’ils sont programmés, en fonction de ce que l’on connaît de l’animal et de la molécule à tester. Pour exister, ces logiciels s’appuient donc sur les recherches sur l’animal.
2- parce que les prédictions basées sur ces modèles restent très imparfaites. Dans le cas du paludisme, aucun logiciel actuel ne peut intégrer la complexité et la dynamique des relations hôte-parasite.
La bio-informatique constitue donc un outil supplémentaire pour orienter les recherches sur le paludisme, mais en aucun cas les ordinateurs ne peuvent assumer seuls le rôle de modèle et se substituer à l’animal.
C.1.2- Par rapport aux modèles in vitro
De la même façon, le système d’étude in vitro reste sérieusement limité. Dans le cas du paludisme, la principale difficulté concerne la culture du plasmodium (Schuster, 2002). En effet, certaines espèces sont difficiles à entretenir, comme P. vivax (Golenda et al., 1997). Pour P. falciparum, si les souches de laboratoire sont faciles à mettre en culture, les souches de « terrain » sont bien plus délicates à faire pousser in vitro. Par ailleurs, ces parasites sont dans un environnement artificiel (sang ou hépatocytes), sans relation avec le reste de l’organisme qui les abrite. Or la biologie du plasmodium est très dépendante des interactions avec les autres cellules du corps (mécanismes immunitaires notamment). Les méthodes in vitro permettent donc seulement l’étude de secteurs bien limités. Ainsi, quelle que soit leur sophistication, ces méthodes ne pourront jamais simuler la relation hôte-parasite dans toute sa complexité ni garantir que tous ces systèmes et leurs interactions auront bien été pris en compte. Les mêmes cellules (plasmodies ou cellules de l’hôte) étudiées in vitro peuvent ne pas réagir de la même façon que celles évoluant au sein d’un organisme complet.

Les primates africains et la recherche sur le paludisme
71
Il sera donc toujours nécessaire à un moment donné de la recherche d'étudier l’hôte dans sa globalité, donc de recourir à l'expérimentation animale afin d'acquérir un maximum de garanties avant l'application à l'homme. Ainsi, par exemple, l’utilisation de l’animal est primordiale dans l’étude de la pathogénicité ou de la mise au point d’un vaccin : le vaccin n’agit pas seul, ni directement. Il nécessite de stimuler le système immunitaire pour lutter ensuite contre le plasmodium. Un vaccin doit donc être testé sur l’animal pour en vérifier l’innocuité sur l’ensemble des cellules et l’efficacité sur le système immunitaire.
C.1.3- Par rapport à une étude directe sur l’humain
Si les études sur les humains apportent de précieuses informations sur l’épidémiologie d’une maladie ou sur les maladies elles-mêmes, ces études sont avant tout limitées par des considérations éthiques. L'intérêt de la recherche chez le PNH est que l'on peut théoriquement mener les conditions expérimentales jusqu'à leur extrême, c'est-à-dire la mort. On peut ainsi :
- étudier les mécanismes immuno-pathogéniques en jeu au cours de l'accès pernicieux ; - avoir accès aux populations cellulaires spléniques, car l’immunité circulante
(accessible chez l’homme) ne suffit pas à expliquer les réactions immunitaires face au paludisme. La rate semble donner un bon reflet des mécanismes immunologiques mis en jeu par l'hôte contre le parasite ;
- avoir accès à des prélèvements post-mortem (cerveau, foie, rate, etc.) beaucoup plus facilement que chez l'homme ;
- tester des molécules (alors que chez l’homme il faut auparavant en démontrer l’innocuité) ;
- sélectionner les vaccins et ainsi éviter de les évaluer inutilement chez l’homme. En effet, les bonnes pratiques de réalisation d’essais vaccinaux dictées par l’OMS rendent ces essais difficiles, longs et très coûteux :
consentement éclairé des participants, structures de soins pour le traitement des réactions vaccinales possibles
et des accès palustres, comité indépendant de supervision des données et de l’innocuité, évaluation préalable de l’innocuité du candidat vaccin d’une part chez
des sujets de zone non-endémique, d’autre part chez des sujets de zone endémique, procédures de fabrication internationalement reconnues, durée minimale des essais supérieure à 5 ans, etc.

Les primates africains et la recherche sur le paludisme
72
C.2- INTERET PAR RAPPORT AUX MODELES NON PRIMATES
Dans le cadre du paludisme, le modèle animal le plus classiquement utilisé est le rongeur.
La première raison à cela est la même que pour tout domaine de recherche : les rongeurs sont des animaux biologiquement très bien définis, leur coût d’achat et d’entretien sont minimes et leur disponibilité est grande. La seconde raison est plus spécifique au paludisme : les plasmodiums de certains
rongeurs sont assez proches des plasmodiums humains (figure 10).
Figure 10 : phylogénie de quelques plasmodies établie à partir du gène CSP (Circumsporozoite protein) (Ayala et al., 1999) : 2 plasmodiums de rongeurs (P. yoelii et P. bergei) et 10 plasmodiums de primates Toutefois, les espèces plasmodiales des PNH sont plus proches des plasmodiums humains que ne le sont celles des rongeurs. De même, dans la phylogénie des espèces hôtes, les primates sont bien sûr bien plus proches des humains que ne le sont les rongeurs. Cette proximité des hôtes et des parasites a de nombreuses conséquences (immunitaires, métaboliques, etc.), qui aboutiront toujours à la possibilité d’extrapolation plus directe des observations à l'homme.
C.2.1- Sensibilité croisée des plasmodiums
Des études épidémiologiques couplées chez l’homme et le PNH ont montré la possibilité de passage de plasmodiums d’une espèce à l’autre (Volney et al., 2002) ; certains scientifiques avancent même que les PNH pourraient jouer le rôle de réservoir de paludisme (Fandeur et al., 2000).

Les primates africains et la recherche sur le paludisme
73
C.2.1.1- sensibilité des primates aux souches humaines
Il est primordial, dans le cadre du développement de vaccins ou de thérapies, de tester les nouvelles molécules sur des animaux pour en vérifier d’abord l’innocuité. Pour en mesurer l’efficacité, l’idéal serait donc que ces animaux soient porteurs du plasmodium humain que l’on souhaite combattre. Or il est impossible d’infecter un rongeur ou n’importe quel autre animal non primate avec les espèces plasmodiales humaines. Seuls quelques PNH présentent une sensibilité aux parasites humains. De façon naturelle, seul P. malariae infecterait les PNH, mais d’autres plasmodiums peuvent leur être inoculés de façon expérimentale (Centre National de la Recherche Scientifique, 2004).
• Espèces sensibles aux deux plasmodiums humains majeurs P. falciparum et P. vivax (Coatney et al., 1971) : singe nocturne : Aotus sp., ((Geiman & Meagher, 1967) pour P. falciparum ;
(Porter & Young, 1966; Collins et al., 2005b) pour P. vivax) singe hurleur : Alouatta fusca ((Taliaferro & Taliaferro, 1934) pour P. falciparum) tamarin : Saguinus geoffroyi ((Porter & Young, 1967) pour P. falciparum ; (Porter
et al., 1966) pour P. vivax) saïmiri : Saïmiri sciureus ((Young & Rossan, 1969c) pour P. falciparum ; (Deane
et al., 1966) pour P. vivax) singe capucin : Cebus capucinus ((Young & Baerg, 1969a) pour P. falciparum ;
(Young & Porter, 1969b) pour P. vivax) singe araignée : Ateles sp. ((Young et al., 1969b) pour P. vivax) chimpanzé : Pan troglodytes ((Bray, 1958a) pour P. falciparum ; (Mesnil &
Roubaud, 1920) pour P. vivax) gibbon : Hylobates lar, ((Ward et al., 1965) pour P. falciparum) macaque : Macaca sp. ((Cadigan et al., 1966) pour P. falciparum ; insensible à P.
vivax) • Espèces sensibles à P. malariae :
singe nocturne : Aotus sp. (Geiman & Siddiqui, 1969) chimpanzé : Pan troglodytes (Rodhain, 1948) saïmiri : Saïmiri sciureus (Collins et al., 1989b)
• Espèce sensible à P. ovale :
chimpanzee : Pan troglodytes (Bray, 1957; Collins & Jeffery, 2005a)
C.2.1.2- sensibilité des humains aux souches des primates
On a décrit des infections naturelles d’humains à des plasmodies d’origine simienne. Elles semblent assez rares et leurs manifestations cliniques sont bénignes.

Les primates africains et la recherche sur le paludisme
74
L’homme semble peu réceptif aux parasites des singes anthropoïdes et semble plus sensible aux plasmodiums des macaques du Sud-Est asiatique ou des primates sud-américains. Une petite dizaine de plasmodies est, de façon certaine, responsable d’infections humaines :
P. knowlesi (Singh et al., 2004) P. cynomolgi (Eyles et al., 1960) P. brazilianum (Deane, 1992) P. simium (Deane, 1992) P. inui (Collins, 2003) P. eylesi (Coatney et al., 1971) P. schwetzi (Rodhain & Dellaert, 1955b; Rodhain & Dellaert, 1955a)
En revanche, P. coatneyi, P. gonderi, P. fragile, P. fieldi, P. simiovale et, de façon plus étonnante, P. reichenowi (Rodhain, 1939) ne sont pas considérés comme infectieux pour l’homme. Ainsi, en général, cette sensibilité de l’homme aux plasmodiums simiens semble suffisamment anecdotique dans la nature pour minimiser son rôle dans le fléau du paludisme. Elle ne constitue pas pour l’instant un facteur limitant pour les programmes de prophylaxie et d’éradication de la maladie (Centre National de la Recherche Scientifique, 2004).
C.2.1.3- notion de réservoir
Dans le domaine des zoonoses, une idée répandue est celle que les primates ne peuvent avoir de rôle important dans le maintien de pathogènes dans la mesure où ils sont trop dispersés. Mais dans le cas du paludisme, la latence de la maladie permet même aux petites populations de maintenir le plasmodium. Le cas des orangs-outans en est un bon exemple : ils abritent deux espèces plasmodiales alors que leur densité n’est que de 2 individus par km2 (Peters, 1976). Suivant ce principe, l’existence d’hypnozoïtes de certaines espèces plasmodiales représenterait la condition extrême pour un maintien dans les plus petites populations. Par ailleurs, l’écologie des primates représente une autre condition favorable au maintien du paludisme :
- les primates vivent en forêt, en groupe, ce qui constitue un facteur attractif pour les moustiques ;
- leur régime frugivore leur impose une mobilité (terrestre ou arboricole) qui augmente leur exposition.
Deux éléments ont longtemps manqué pour définir le rôle exact des primates non-humains dans le maintien du paludisme :
• le peu de données concernant le statut sanitaire des primates non-humains sauvages, • le développement d’outils permettant un diagnostic d’espèces plus précis que le frottis
sanguin. Ainsi, le développement de la PCR pour le diagnostic d’espèces plasmodiales a été un élément décisif pour l’étude des interactions entre les parasites simiens, humains et leurs hôtes.

Les primates africains et la recherche sur le paludisme
75
Une étude dans le district de Kapit en Malaisie a indiqué que 20% des cas cliniques de paludisme humain ont une allure microscopique légèrement différente de la normale et sont négatifs en PCR pour P. malariae. Malgré ce doute, ces cas étaient tout de même jusque là déclarés comme paludisme à P. malariae. Des travaux sur cette population ont permis de mettre en évidence que 50% de ces cas douteux étaient liés à des infections à P. knowlesi (un plasmodium du macaque crabier), dont l’aspect microscopique est très similaire à P. malariae (Singh et al., 2004). Ainsi, dans cette région au moins, il semble que les infections à plasmodies d’origines simiennes soient largement sous-diagnostiquées.
Plusieurs études réalisées en Guyane française ont également soulevé la question du risque zoonotique pour le paludisme :
o Fandeur a d’abord démontré la forte prévalence (plus de 10% de PNH infectés) de P. brasilianum chez les primates sauvages (atèles, singes hurleurs et tamarins) (Fandeur et al., 2000).
o Ses résultats furent d’ailleurs confirmés par une autre étude épidémiologique concernant plusieurs espèces de mammifères et de reptiles (de Thoisy et al., 2000).
o Mais lorsque les séquences de ces parasites étaient alignées sur les séquences de P. malariae, rien ne permettait de distinguer les deux espèces plasmodiales.
o Une étude sérologique révéla que 73% des primates étaient positifs pour le peptide circumsporozoïte de P. malariae/P. brasilianum.
o Un certain nombre d’animaux réagirent également positivement aux séquences répétitives NANP de P. falciparum, suggérant ainsi que les primates étaient aussi piqués par des anophèles infectées par des parasites humains.
o Par ailleurs les populations humaines vivant à proximité des forêts eurent des sérologies à P. malariae/P. brasilianum assez élevées.
Ces résultats suggèrent que certains primates non-humains pourraient constituer un réservoir non seulement pour les autres espèces de singes, mais aussi pour les humains. Ils pourraient ainsi être responsables du maintien dans cette zone isolée de forêt amazonienne de P. malariae (Volney et al., 2002) car ils sont bien souvent utilisés comme animaux de compagnie par les populations (de Arruda et al., 1989).
Au Brésil, en Nouvelle-Guinée, en Indonésie, des études menées chez l’humain ont
permis de mettre en évidence une espèce plasmodiale très répandue, proche de P. vivax, appelée ainsi P. vivax-like, mais qui est en fait aussi très proche de P. simiovale des macaques asiatiques (Qari et al., 1993; Escalante et al., 1995).
Concernant l’Afrique, que dire des chimpanzés porteurs de P. ovale ? Ou de
P. rodhaini des chimpanzés et gorilles qui ne serait en fait que P. malariae ? Ou des suspicions de P. reichenowi chez des humains (Coatney et al., 1971) ? Ou des cas de P. schwetzi, proche de P. vivax, transmis du chimpanzé à l’humain via une piqûre de moustique (Contacos et al., 1970) ?
Cette liste n’est pas exhaustive car, bénéficiant des progrès PCR, les études sur ce sujet ne font que commencer.

Les primates africains et la recherche sur le paludisme
76
Les primates pourraient donc parfois jouer le rôle de réservoir du paludisme dans la mesure où ils ne sont pas malades avec leur parasite, mais avec une importance différente selon les régions du monde et le plasmodium considérés. Ainsi par exemple, P. reichenowi qui est pourtant le plus proche parent de P. falciparum, ne semble pourtant pas directement transmissible à l’homme (Coatney et al., 1971). Enfin, ce rôle de réservoir pourrait ne pas avoir que des inconvénients : on peut imaginer que les primates seraient aussi des réservoirs de souches sauvages, non résistantes aux antipaludiques. En effet, si un primate abrite deux souches, l’une sauvage, l’autre résistante, on peut penser que la compétition entre les deux souches aboutira à la disparition de la souche résistante. Ceci s’expliquerait par le fait que l’acquisition de la résistance a été coûteuse pour la souche (Wolfe et al., 1998). Un phénomène semblable a été décrit chez les babouins sauvages ; ils présentaient bien moins de souches de bactérie intestinales gram – résistantes que n’en possédaient les populations de babouins vivant auprès des humains (Rolland et al., 1985). Les études sur les primates sauvages doivent donc se poursuivre et se développer, afin de mieux cerner le rôle de ces derniers dans le maintien et la transmission du paludisme.
C.2.2- Proximité des tableaux cliniques
Les modèles rongeurs ne reproduisent que partiellement les symptômes se rapportant à l’homme : ils donnent un simple aperçu dans le cadre du neuropaludisme (Hunt et al., 2003) et par ailleurs les symptômes observés sont peu reproductibles.
C.2.2.1- les primates modèles cliniques
Les primates font d’excellents modèles pour tous les types de cliniques du paludisme :
- ils peuvent reproduire la variété des symptômes observés lors d’accès simples de paludisme, du simple syndrome grippal aux cas mortels. Cette diversité s’obtient aussi bien avec un parasite humain (ex : P. falciparum chez le saïmiri (Contamin et al., 2000)) qu’avec un parasite simien (ex : P. knowlesi chez le babouin (Ozwara et al., 2003b));
- ils peuvent développer un neuropaludisme, que ce soit avec un plasmodium humain (ex : P. falciparum chez le saïmiri (Gysin et al., 1992)) ou simien (ex : P. fragile chez le macaque rhésus) (Fujioka et al., 1994)) ;
- ils peuvent enfin être des modèles du paludisme de la femme enceinte (ex : P. coatneyi chez le macaque rhésus) (Davison et al., 1998)).
C.2.2.2- possibilité de tester l’efficacité de nouvelles molécules
La plupart des substances étant schizonticides, leur efficacité peut être testée avec de nombreux plasmodiums, c’est-à-dire aussi bien chez la souris (avec P. berghei et P. yoelii) que chez le primate (ex : P. knowlesi et P. cynomolgi) (Shi et al., 1999). Cependant, si les rongeurs possèdent un plasmodium « naturel » assez proche du plasmodium humain pour tester les nouvelles molécules, l’avantage du primate sur le rongeur est encore lié à sa proximité avec l’humain (notamment en ce qui concerne son métabolisme et donc la pharmacodynamique et la pharmacocinétique d’une molécule).

Les primates africains et la recherche sur le paludisme
77
Ainsi, on continue de tester les médicaments sur les rongeurs pour un premier aperçu de leur efficacité et de leur toxicité, mais ensuite les tests sur les primates permettent d’en déterminer plus précisément les doses thérapeutiques (Puri & Dutta, 2003). En effet, le passage du rongeur aux primates s’accompagne généralement d’une forte baisse de la dose thérapeutique efficace (Puri & Singh, 2000) ce qui est simplement lié au changement de poids de l’animal : la dose/kg augmente si le poids de l’animal diminue.
C.2.3- Proximité des systèmes immunitaires
C.2.3.1- les primates modèles d’immunité
Les PNH sont de bons modèles des mécanismes immunitaires qui ont lieu pendant le paludisme, avec, par rapport à l’homme, la possibilité de contrôler leur environnement. On peut ainsi suivre l’évolution du système immunitaire lors :
- d’accès simple comme avec P. cynomolgi (P. vivax-like) ou P. knowlesi (P. falciparum-like) (Praba-Egge et al., 2002)
- de neuropaludisme (de Souza & Riley, 2002) comme avec P. coatneyi chez le macaque rhésus (Tongren et al., 2000).
C.2.3.2- les primates modèles pour les essais vaccinaux
L’homologie des séquences vaccinales des plasmodies simiennes et humaines et la proximité des systèmes immunitaires des PNH et des humains placent évidemment les primates au centre des essais vaccinaux contre le paludisme. Un avantage des primates par rapport aux humains, outre les facilités éthiques, financières et législatives, est que l’on peut tester au départ des adjuvants assez puissants, même s’ils sont contre-indiqués chez l’humain (adjuvant de Freund par exemple). Cette proximité immunitaire implique également un contrôle sérologique des animaux avant chaque protocole puisque, comme chez l’humain, les réponses peuvent différer lorsque les primates sont déjà pré-immuns (Perraut et al., 2000). Comme dans le cadre des autres études, les primates peuvent être utilisés avec leur propre plasmodium ou avec du plasmodium humain : certains essais vaccinaux recherchent une protection contre des infections homologues, d’autres études recherchent une protection croisée (séquence vaccinale issue d’un plasmodium X pour protéger contre une espèce plasmodiale Y). Pour illustrer ces propos, parmi les très nombreux travaux réalisés dans ce domaine, l’exemple de la séquence AMA-1 (Apical Membrane Antigen, vaccin qui a pour cible le mérozoïte) est choisi. En effet, non seulement cette séquence se retrouve chez beaucoup de plasmodiums, mais l’immunité liée aux familles d’AMA-1 est assez proche entre les PNH et l’humain (Thomas et al., 1994a).

Les primates africains et la recherche sur le paludisme
78
a- essais vaccinaux avec du plasmodium de primates contre des infections homologues
La séquence AMA-1 a ainsi été utilisée pour vacciner le macaque rhésus, en choisissant AMA-1 issue de P. knowlesi pour lutter contre des infections homologues (Deans et al., 1988).
b- protection croisée
La séquence AMA-1 issue de P. vivax protège ainsi partiellement le macaque rhésus d’une infection à P. cynomolgi (Kocken et al., 1999) et AMA-1 de P. falciparum inhibe in vitro la croissance de P. reichenowi (Kocken et al., 2000). Ceci est également vrai lorsque cette séquence provient d’un plasmodium simien et cherche à protéger contre un plasmodium humain : AMA-1, avec une protéine recombinante de P. fragile protége partiellement le saïmiri d’une infection à P. falciparum (Collins et al., 1994b).
c- avec du plasmodium humain
Les primates pouvant être utilisés dans ce cadre sont déjà moins nombreux : il s’agit de la petite dizaine d’espèces permissives aux infections humaines (voir précédemment). Toutefois, obtenir une telle protection chez un primate représente l’étape ultime pour passer aux essais cliniques chez l’humain. Reprenons l’exemple d’un vaccin utilisant la séquence AMA-1, mais cette fois issue de P. falciparum :
- pas de protection du saïmiri contre une infection homologue (Pye et al., 1991), - développement d’une bonne immunité chez Aotus lemurinus lemurinus (Jones et al.,
2002). - le sérum obtenu de macaques rhésus vaccinés avec AMA-1 et MSP-1 (Merozoite
Surface Protein, même principe qu’AMA-1) issus de P. falciparum inhibe la croissance de P. falciparum in vitro (Pan et al., 2004).
Un autre exemple de protection in vivo concerne le développement du vaccin à base de LSA-3 (Liver Stage Antigen, dirigé contre les stades hépatiques) ; des essais de vaccination à l’aide de LSA-3 issu de P. falciparum ont en effet permis la protection contre des infections homologues chez Aotus lemurinus griseimembra (Perlaza et al., 2003) et chez le chimpanzé (Daubersies et al., 2000).
C.3- INTERET PAR RAPPORT AUX AUTRES PRIMATES
Les quelques 200 espèces de primates simiens (Haplorhiniens) se divisent pour moitié en Catarhiniens, singes de l’Ancien Monde (Afrique et Asie) et pour autre moitié en Platyrhiniens, singes dits du Nouveau Monde (Amérique). La recherche sur le paludisme utilise principalement les petits primates sud-américains et les macaques. Les travaux réalisés sur les primates africains sont plus rares et concernent alors à 80% les chimpanzés.

Les primates africains et la recherche sur le paludisme
79
Pourtant, l’intérêt des primates africains n’est pas négligeable dans le cas de la recherche sur le paludisme :
- ils sont, d’un point de vue phylogénétique et physiologique, plus proches de l’homme que leurs cousins américains ;
- certains sont sensibles à tous les parasites humains ; - certains sont permissifs aux deux types d’infection (sang ou sporozoïtes) ; - ils sont permissifs à d’autres plasmodiums simiens ; - certains de leurs plasmodiums naturels sont extrêmement proches de P. falciparum ; - certains de ces primates bénéficient d’une grande disponibilité et de facilité d’élevage.
C.3.1- Proximité phylogénétique des primates africains avec l’homme
Figure 11 : classification générale des primates (Dupuis, 2003)
L’homme est un primate simien, catarhinien, hominoïde dont la divergence avec les grands singes date d’environ 5 millions d’années. Les primates africains en général ont divergé il y a 15 à 20 millions d’années alors que le groupe des primates sud-américains a divergé il y a 30 millions d’années. Cette plus grande proximité des PNH africains avec l’humain par rapport aux PNH sud-américains se répercute notamment sur trois domaines qui concernent les recherches sur le paludisme :
1- leur taille et leur poids, 2- leur système immunitaire, 3- leur système reproducteur.

Les primates africains et la recherche sur le paludisme
80
C.3.1.1- conséquences des proximités de tailles et de poids
La plus grande taille des primates africains présente de réels avantages dans le domaine de la recherche sur le paludisme.
1- La possibilité de réaliser davantage de prélèvements durant le protocole : le volume total de sang d’un animal est d’environ 60mL/kg et l’on peut prélever 7,5% de ce volume par semaine (Diehl et al., 2001). Cela signifie que pour un saïmiri ou un singe de nuit de 800g, on ne peut prélever que 3,6 mL par semaine. Il semble difficile dans ces conditions de pouvoir réaliser des bilans parasitologiques, immnunologiques, biochimiques et hématologiques complets. Chez un babouin de 20 kg, ce sont en revanche 90 mL hebdomadaires qui peuvent être prélevés et pour un chimpanzé de 50 kg, 225mL. Cela permet alors de réaliser ces bilans, mais aussi de faire des cultures in vitro, des études plus poussées de l’immunité et/ou des congélations de souches.
2- Dans le cadre des recherches thérapeutiques, les doses efficaces obtenues sont plus proches de celles utilisées chez l’humain.
3- Ces primates peuvent abriter davantage de parasites, ce qui est intéressant
lorsqu’on cherche à produire un grand nombre de sporozoïtes. Ainsi, par exemple, le sang hépariné de 6 chimpanzés infectés permet de nourrir 45 000 moustiques (Collins et al., 1986) à la seule condition de respecter un délai inférieur à 45 minutes après le prélèvement pour nourrir les insectes (Ponnudurai et al., 1990). Le chimpanzé peut donc être utilisé comme une « usine à sporozoïtes » qui permet d’infecter d’autres espèces de PNH pour la recherche (Sullivan et al., 1996).
C.3.1.2- conséquences des proximités des système immunitaires
La grande proximité des systèmes immunitaires des primates africains et de l’homme est déterminante dans le cadre des recherches vaccinales (Kennedy & Walker, 1997). Ainsi par exemple, alors que les macaques ne présentent que 3 classes d’IgG (Martin, 1982), le babouin, les chimpanzés et l’humain en ont 4 (Damian et al., 1971). Les saïmiris possèdent quant à eux un système de groupes sanguins particulier avec des problèmes d’incompatibilité forte (production d’allo-anticorps anti-érythrocytaires) qui perturbe considérablement les parasitémies lors d’infections d’épreuve par les stades sanguins (Michel et al., 2005). Enfin au sein des primates africains, le chimpanzé appartient au petit groupe des singes supérieurs et est donc un modèle particulier. Les 99% d’homologie génétique avec l’homme (Pennisi, 2003) et son extraordinaire proximité du système immunitaire permettent entre autres :
de pouvoir utiliser des tests déjà développés pour l’homme ou de développer des tests
utilisables ensuite chez l’homme (Mucenski et al., 1986); des résultats de vaccination asses prédictibles (Thomas et al., 1994b ; BenMohamed et
al., 2004) ; de pouvoir tester des immunothérapies humaines, que ce soit par des anticorps
(Dziegiel, article soumis) ou par des IFNγ (Ferreira et al., 1986) ;

Les primates africains et la recherche sur le paludisme
81
de penser que les différences observées pourraient correspondre à la réelle capacité des primates africains à éliminer le parasite (et non pas seulement à la différence existant entre les 2 espèces hôtes). Cette hypothèse est déjà le principe des recherches sur le SIDA (le primate africain représente un bon modèle de non-pathogénicité). On a donc la possibilité de comparer, avec l’homme et le chimpanzé, deux modèles très proches, l’un pathogène, l’autre non (Nardin, 1990).
C.3.1.3- conséquences des proximités des systèmes reproducteurs
Ainsi, la placentation du macaque est double discoïde alors que celle de l’homme est simple, comme chez le babouin. L’étude des infections palustres de la femme enceinte serait donc plus indiquée sur des primates africains.
C.3.2- Sensibilité aux quatre plasmodiums humains
Le chimpanzé est le seul animal permissif aux quatre plasmodiums humains. • Même s’il peut se développer dans les hépatocytes de saïmiri (Millet et al., 1994), le
chimpanzé splénectomisé est le seul primate non-humain qui peut développer une parasitémie avec P. ovale (Morris et al., 1996; Collins et al., 2005a).
• P. malariae peut se développer chez le chimpanzé splénectomisé (Collins et al., 1990)
tout comme chez le singe de nuit splénectomisé ; mais les parasitémies obtenues chez le primate africain sont trois à dix fois plus élevées (jusqu’à 75 000 parasites/μL) que chez le modèle Aotus (Collins et al., 1994a).
• P. vivax se développe bien sur les chimpanzés splénectomisés, avec les mêmes
parasitémies que celles observées chez l’homme, contrairement aux faibles parasitémies (Collins et al., 1998) ou aux infections partielles (Collins et al., 1988) obtenues chez Aotus. En outre, le chimpanzé possède, comme chez l’homme, les formes hypnozoïtes de P. vivax (Krotoski et al., 1986). Enfin, le cycle complet peut être réalisé, quel que soit le mode d’infection, jusqu’à la transmission aux moustiques, comme pour les modèles sud-américains, mais avec un bien meilleur taux d’infection chez le chimpanzé. En effet, la transmission par sporozoïte chez Aotus n’aboutit qu’à 50% d’infection, ce qui est trop faible pour envisager de tester des vaccins ou des molécules dirigés contre les phases exo-érythrocytaires (Sullivan et al., 2001).
• Concernant P. falciparum enfin, il est possible d’obtenir une bonne parasitémie sur
animal non splénectomisé. Cependant, sur animal non splénectomisé, le développement parasitaire s’arrête de lui-même au bout de 1 ou 2 cycles (Bray, 1958b). Le recours à la splénectomie peut être également intéressant pour une comparaison avec les cas humains de paludisme sur personne splénectomisée (Grobusch et al., 2003).

Les primates africains et la recherche sur le paludisme
82
C.3.3- Sensibilité aux infections par les sporozoïtes de P. falciparum
Le chimpanzé est également le seul modèle animal sensible, de manière prévisible, à des infections par des sporozoïtes de P. falciparum. Il constitue donc l’un des meilleurs modèles pour tester la protection induite par des antigènes exprimés par les stades exo-érythrocytaires de P. falciparum. En effet, chez le chimpanzé, le taux de conversion des sporozoïtes injectés par le moustique en formes hépatiques est aussi élevé que chez l’homme (30-60%) et la durée du cycle hépatique est identique à celle de l’homme (Meis et al., 1990). Il est donc aussi sensible que l’homme au développement des formes hépatiques de P. falciparum (Meis et al., 1990). Ceci est un avantage sur les primates sud-américains qui doivent être en général splénectomisés (Collins et al., 1992a) et dont les parasitémies observées sont faibles, inférieures à 1000 parasites/μL quel que soit l’inoculum (Zapata et al., 2002).
C.3.4- Forte reproductibilité des résultats
D’une manière générale, la reproductibilité des infections d’épreuve chez le chimpanzé est aussi élevée que chez l’homme : le chimpanzé, comme l’homme, ne semble pas développer de réelle immunité à la suite de ces infections d’épreuves répétées. Cela diffère des primates sud-américains, dont la parasitémie est plus faible, dure moins longtemps (Collins et al., 2002) ou dont le niveau de protection vaccinale augmente s’ils ont déjà été en contact avec le parasite (Jones et al., 2000). Ainsi, du fait de la bonne reproductibilité à des infections d’épreuve successives, il est possible d’obtenir des résultats fiables à partir d’un faible nombre d’animaux. Cela permet de réduire les coûts, les durées de protocoles et de respecter le principe éthique de la réduction du nombre d’animaux utilisés en recherche. Les Aotus en revanche nécessitent souvent un plus grand nombre d’individus testés ce qui est problématique car il devient assez difficile de se les procurer (Herrera et al., 2002).
C.3.5- Sensibilité à d’autres plasmodiums simiens
Le babouin (Papio sp.) est naturellement infecté par P. gonderi mais cette infection est asymptomatique. En revanche, il représente un bon modèle de paludisme bénin ou même sévère lorsqu’il est infecté avec P. knowlesi, un plasmodium du macaque. Il peut alors, comme certains macaques, développer des infections aiguës ou chroniques et les symptômes aussi variés que chez l’homme (Ozwara et al., 2003b). L’avantage de ce modèle est double :
o sa taille est plus importante qu’un macaque (et a fortiori qu’un primate sud-américain) ;
o le plasmodium utilisé, P. knowlesi, est transmissible à l’homme, ce qui pourrait correspondre à une proximité biologique avec les plasmodiums humains.
Le mandrill (Mandrillus sphinx) quant à lui, peut développer de fortes parasitémies
avec une infection à P. eylesi, le plasmodium du gibbon (Centre National de la Recherche Scientifique, 2004).

Les primates africains et la recherche sur le paludisme
83
Utiliser des plasmodiums simiens sur des PNH africains permet conjointement : o de tester de nouvelles molécules antipaludiques sur un animal de grande taille, o d’éviter un risque de transmission à l’humain pendant le protocole.
C.3.6- Proximité phylogénétique de leurs plasmodiums naturels avec P. falciparum
Si les plasmodiums naturels des primates sud-américains, P. brasilianum et P. simium, sont respectivement très proches de P. malariae et P. vivax, ces primates ne sont presque utilisés que pour leur sensibilité à P. falciparum. Ils sont donc utilisés avec un parasite qui n’est pas naturel, dans le même principe que l’on utilise diverses souris de laboratoire avec des plasmodiums de rongeurs africains. On peut alors se demander quel est le biais induit par ces infections non naturelles en ce qui concerne les relations hôte-parasite. Le plasmodium naturel du chimpanzé, P. reichenowi, est quant à lui le plus proche parent de P. falciparum (voir figure 10) ; cette proximité concerne également son aspect microscopique et le cycle sanguin qui est de 48 heures pour les deux parasites. Ainsi, les séquences de l’antigène MSP-3 (Merozoite Surface Protein) de P. falciparum et de P. reichenowi présentent une quasi complète homologie (Okenu et al., 2000), ce qui implique que les anticorps induits par un vaccin MSP-3 dérivé de P. falciparum devraient avoir la même efficacité vis-à-vis de P. reichenowi. Il en va de même pour les antigènes vaccinaux AMA-1 (Kocken et al., 2000), MSP-2 (Dubbeld et al., 1998) et P48/45 (Milek et al., 1997). Il est donc possible pour de nombreux antigènes vaccinaux de les tester sur chimpanzés avec des infections par leur plasmodium naturel, P. reichenowi en utilisant des animaux non splénectomisés.
C.3.7- Disponibilité et facilité d’élevage
Les primates sud-américains comme Aotus ont été préférés dans le cadre des recherches sur le paludisme (Pico de Coana et al., 2003), mais le problème principal de ce modèle est sa disponibilité : protégé par la Convention de Washington, son usage est réglementé et malheureusement son élevage semble difficile. Cela n’est pas le cas du babouin, bien représenté en Afrique et dont l’élevage est assez facile (Ozwara et al., 2003b). L’élevage du mandrill et du chimpanzé est également assez facile, mais ces espèces sont quant à elles protégés.

Les primates africains et la recherche sur le paludisme
84
D- LIMITES D’UTILISATION DES PRIMATES AFRICAINS DANS LA RECHERCHE SUR LE PALUDISME
D.1- LIMITES BIOLOGIQUES
D.1.1- Intérêt de l’utilisation des techniques in vitro
Mis à part P. vivax, on maîtrise assez bien maintenant la culture in vitro des plasmodiums humains (Schuster, 2002) :
- P. falciparum (Trager & Jensen, 1976; Druilhe & Gentilini, 1982) et P. malariae (Siddiqui et al., 1972) se cultivent depuis longtemps ;
- quant à P. ovale, des stades hépatiques peuvent être obtenus à partir de sporozoïtes (Mazier et al., 1987; Millet et al., 1994).
Les domaines d’études in vitro sont donc de plus en plus importants (Trager & Jensen, 1997) :
- culture extracellulaire, - gamétocytogenèse, - recherche et caractérisation de gènes, - clonage, - variation antigénique, - apparition des résistances, - identification des récepteurs cellulaires, - mécanismes d’invasion cellulaire, - pathogenèse, - étude de cytoadhérence, - séquestration, - étude de nouvelles molécules antipaludiques (screening et toxicité), - production d’antigènes susceptibles de servir à des vaccins, etc.
D.1.2- Limites du modèle animal
Le développement d’un vaccin constitue parfois le point limite d’utilisation des primates. En effet, les variations inter-individuelles face au plasmodium sont importantes et le vaccin devrait donc être testé à très grande échelle pour connaître sa réelle efficacité. Or, pour des raisons éthiques, financières et écologiques, il est souvent difficile pour un laboratoire d’investir dans de grandes études vaccinales sur le modèle primate (d’autant qu’une fois le vaccin reconnu efficace, il faudra encore des millions d’euros et des années d’étude clinique chez des volontaires humains).
En outre, les essais cliniques des vaccins antipaludiques chez l’homme sont relativement aisés à réaliser dans les zones endémiques africaines ou asiatiques (ce qui n’est pas le cas des vaccins contre des maladies rares par exemple). Certains chercheurs proposent donc de n’utiliser le PNH que pour vérifier l’innocuité d’un vaccin contre le paludisme et non pour en tester son efficacité. En effet, dans le cadre du paludisme, beaucoup de vaccins prometteurs chez le primate s’avèrent inefficaces chez l’humain, l’inverse étant également possible.

Les primates africains et la recherche sur le paludisme
85
C’est le cas par exemple d’un vaccin MSP-3 (contre les stades sanguins) testé chez des volontaires européens. Cette étude testait deux adjuvants : l'alun et le Montanide ISA720. Alors qu'il était totalement inefficace chez les rongeurs et les primates, l'alun s'est avéré aussi efficace que le Montanide chez l’humain (Druilhe et al., 2002). Enfin, toujours dans le cadre des études vaccinales, certains gènes montrent une activité intéressante in vitro mais n’ont pas d’homologue chez les plasmodiums naturels des modèles animaux. Il n’existe par exemple aucun homologue pour l’antigène LSA-1 chez la souris ou le primate non-humain (Carvalho et al., 2002). Or il a été montré que LSA-1 possède une portion d’acides aminés en commun avec un des peptides qui entraîne une réponse spécifique HLA-B53, elle-même liée à une protection contre les cas de paludisme sévère en Gambie. Ainsi l’antigène LSA-1 est un bon candidat vaccinal mais pour lequel aucun modèle animal n’est intéressant. Dans le cadre de l’étude des résistances, les modèles animaux ne sont pas non plus indispensables :
- les études peuvent être conduites directement chez l’homme grâce à des tests de résistance ou des tests de caractérisation moléculaire ; pour les molécules dont les marqueurs de résistances sont identifiés, la PCR permet de les détecter dans le sang de cohortes humaines étudiées.
- Des tests in vitro d’exposition de souches plasmodiales à différentes molécules peuvent également être employés.
Enfin, certaines molécules comme l’artémisinine et ses dérivés, déclarées dangereuses pour l’animal, se sont avérées d’une utilisation peu risquée chez l’humain. Ceci pourrait s’expliquer par les voies d’administration différentes (intramusculaire à relargage lent chez l’animal, en général per os chez l’homme) ou par les doses, généralement assez élevées chez l’animal (Gordi & Lepist, 2004). Inversement, et plus dramatiquement, une molécule qui peut sembler inoffensive chez l’animal peut s’avérer nocive chez l’homme.
D.1.3- Avantages des autres modèles animaux par rapport aux primates africains
D.1.3.1- les rongeurs et leurs plasmodiums
Parmi les modèles animaux, les rongeurs permettent déjà de grandes avancées scientifiques avec leur plasmodium « naturel » et ceci à moindre coût. Les rongeurs possèdent en effet de nombreuses qualités pour la recherche sur le paludisme :
- la biologie de base des parasites des rongeurs et des humains est commune, - l’organisation génomique et la génétique sont conservées entre ces parasites, - les processus biochimiques sont conservés, - les principes moléculaires de sensibilité ou de résistance aux molécules ont des
caractéristiques communes, - la structure et la fonction des antigènes cibles pour les vaccins sont en général
conservées (comme TRAP et CSP des sporozoïtes, CTRP, P25 et P28 des ookinètes, AMA-1 et MSP-1 des mérozoïtes, P45/48 et P230 des gamètes, etc.),

Les primates africains et la recherche sur le paludisme
86
- le cycle complet incluant l’infection par l’anophèle est simple à obtenir et ne présente aucun danger ; ceci étant également intéressant pour faciliter le transport de prélèvements ;
- il est possible dans ce modèle d’étudier in vivo les interactions hôte-parasite et les effets de nouvelles molécules ;
- les rongeurs sont très bien définis génétiquement et il existe même plusieurs lignées qui forment d’excellents outils pour des études immunologiques.
En pratique on utilise des souris de laboratoire à qui on inocule un des plasmodiums de rats arboricoles africains du genre Thamnomys :
P. berghei, qui est analogue aux parasites humains pour les grands aspects de sa structure, de sa physiologie et de son cycle (Carter & Diggs, 1977). Ce plasmodium représente un très bon modèle de recherche sur le développement des parasites pour trois raisons principales :
o sa culture in vitro, sa large production et la purification de chaque stade sont bien maîtrisées ;
o la séquence et l’organisation de son génome sont connues et il existe même des possibilités de modifications génétiques du parasite ;
o des clones et des lignées de mutants génétiquement modifiés, incluant des parasites transgéniques sont disponibles.
P. chabaudi, qui est utilisé comme modèle d’étude d’apparition des résistances et de variation antigénique (Janssen et al., 2002). Ce parasite provoque chez le rongeur une infection non mortelle, de longue durée, avec une forte variation antigénique (ce type d’étude est impossible avec P. berghei qui est rapidement mortel).
P. yoelii, qui est utilisé dans le cadre des recherches sur les stades hépatiques et sur le rôle des antigènes de stades sanguins sur l’immunité. Par exemple, après avoir identifié les gènes intéressants du plasmodium en vue de la fabrication d’un vaccin, il est facile de faire un premier test de ces antigènes cibles sur des souris avec ce plasmodium naturel (Haddad et al., 2004).
Les études réalisées sur ces rongeurs ont donc permis de nombreuses découvertes sur les parasites humains, telles que :
- la morphologie ultra-structurale des différents stades du parasite (Sinden, 1978;
Aikawa & Seed, 1980) - la génétique des parasites (Walliker et al., 1971; Walliker, 1983), - la description de la méiose (Sinden & Hartley, 1985), - la description des télomères, des chromosomes et de la structure génomique du
parasite (Carlton et al., 2002), - la structure et la fonction de protéines spécifiques (Matuschewski et al., 2002), dont
certains antigènes candidats à la vaccination (Tomas et al., 2001), - la biologie des stades hépatiques (Carrolo et al., 2003), - des tests de molécules antipaludiques (Peters et al., 1987), - le développement de vaccins (Krzych et al., 2000), etc.
Enfin, le dernier grand apport des rongeurs par rapport aux primates est l’existence de souches de souris génétiquement modifiées extrêmement intéressantes pour la recherche.

Les primates africains et la recherche sur le paludisme
87
D.1.3.2- transgenèse
a- principe des souris transgéniques
Les mutations spontanées qui apparaissent dans les colonies de souris de laboratoires sont systématiquement conservées, étudiées et éventuellement utilisées si les modifications biologiques en résultant se rapprochent de maladies humaines identifiées. Mais ces mutations spontanées sont par essence relativement rares. Elles doivent donc être provoquées par l'addition ou l'inactivation de gènes dont les effets sont supposés importants. Des souris transgéniques peuvent ainsi être obtenues dans de multiples laboratoires (Houdebine, 1998). Il existe deux méthodes pour obtenir une souris transgénique :
i. la méthode « classique » par addition de gène dans un embryon de souris au stade d’une cellule ; l’addition de gènes n’est alors pas dirigée.
ii. La « recombinaison homologue » où ce sont des cellules souches embryonnaires transformées qui sont implantées dans des blastocytes d’embryons âgés de 3,5 jours. L’insertion du transgène est ici ciblée. Le souriceau obtenu est chimérique, à deux couleurs de pelage. Il n’est pas nécessaire de réaliser des tests pour vérifier si le transgène est bien inséré. On parle aussi de mutagénèse dirigée.
Encadré 5 : principes d’obtention de souris transgéniques (Arteb, 2002)
La possibilité d'ajouter, de muter ou de retirer un gène dans un animal entier offre aux expérimentateurs des possibilités sans précédent.
b- applications de la transgenèse
On peut utiliser l’animal transgénique seul ou pour réaliser des études comparatives entre des souris au génome modifié et des souris normales mais dont le génome est également connu. Ces études ont par exemple récemment permis :
- l’étude précise des mécanismes immunologiques : rôle des CD36 (Patel et al., 2004),
des cellules T (Tsuji et al., 2003), - l’étude des facteurs de résistance de l’hôte (Shear, 1993), - l’étude de l’apparition des résistances, que ce soit avec l’utilisation de souches
plasmodiales mutantes (Carlton et al., 2001) ou par l’explication des mécanismes mutationnels (Fohl & Roos, 2003),
- l’étude des gènes de prédisposition au paludisme en croisant des souches de souris sensibles et résistantes au paludisme (Kwiatkowski, 2000),
- l’évaluation rapide, in vivo, de l'intérêt protecteur de candidats vaccins recherche. Par exemple, la phase érythrocytaire de P. falciparum est reproduite en greffant des globules rouges et des monocytes humains à des souris mutantes immunodéprimées (Druilhe et al., 2002). Les chercheurs visent désormais à mettre au point un modèle souris reproduisant cette fois la phase hépatique du paludisme.
- La production de lait contenant un vaccin antipaludique par des souris transgéniques (Stowers et al., 2002b), etc.
Jusqu’à présent il était très difficile d’obtenir des PNH transgéniques en raison des faibles rendements des techniques de transfert de gène. Toutefois, un primate transgénique vient

Les primates africains et la recherche sur le paludisme
88
récemment d’être obtenu par recombinaison homologue : un macaque rhésus exprimant une protéine fluorescente au niveau des phanères est né en 2001 (Chan et al., 2001). L’accès aux primates transgéniques comme modèle d’étude à la compréhension du paludisme semble donc désormais possible et va sans nul doute permettre d’accélérer les découvertes dans le domaine fondamental et thérapeutique.
D.1.3.3- les autres modèles primates
Le modèle primate africain s’oppose principalement aux macaques (rhésus et cynomolgus) et aux primates sud-américains.
a- inconvénients des primates africains
Plusieurs éléments sont en défaveur de l’utilisation des primates africains :
- la faible nombre de modèles disponibles ; relativement peu d’espèces ont été étudiées dans le cadre du paludisme et beaucoup d’incertitudes demeurent, que ce soit sur leurs plasmodiums naturels (espèce plasmodiale et/ou vecteur inconnus) ou sur leur sensibilité aux autres plasmodiums.
- Le chimpanzé élimine assez rapidement le parasite (humain ou simien) ce n’est donc pas vraiment un bon modèle puisqu’il y a certainement des différences immunitaires qui participent à cette élimination (Taylor et al., 1985).
- La splénectomie est souvent indispensable à l’infection par des parasites humains ; or la rate est un organe clé des réactions immunitaires et la splénectomie induit un biais non négligeable dans une étude.
- Même splénectomisé la parasitémie d’un chimpanzé infecté avec P. ovale est transitoire et imprévisible (Morris et al., 1996).
- Même splénectomisé, le chimpanzé est un modèle dépourvu de pathogénicité. Ainsi, aucun essai clinique de thérapie contre les symptômes du paludisme (antipyrétiques ou autre) ni aucune recherche fondamentale sur la pathogénicité ne peuvent être envisagés sur le chimpanzé.
b- avantage des macaques
Tout d’abord, comme le macaque est extrêmement utilisé dans les autres domaines de recherche, ce modèle est très bien défini sur le plan biologique et il existe de nombreux outils associés (marqueurs immunologiques, amorces PCR, etc.). En outre, concernant les recherches sur le paludisme, les macaques possèdent certains atouts majeurs :
- ils sont sensibles à beaucoup d’espèces plasmodiales, - l’éventail des pathologies associées est large, - leur immunité est parfaitement connue, - ils font de très bons modèles de neuropaludisme, que ce soit avec leur plasmodiums
naturels (le macaque crabier est l’unique modèle primate naturel permettant l’étude de l’accès pernicieux) ou avec un plasmodium d’une autre espèce de macaque.

Les primates africains et la recherche sur le paludisme
89
Ce large panel de sensibilité permet de nombreuses études sur l’influence du plasmodium et de l’espèce de macaque considérée (selon si l’infection est naturelle ou artificielle par exemple) (Collins et al., 1992b). Par exemple, P. coatneyi peut être inoculé au macaque japonais (Macaca fuscata), au macaque rhésus (Macaca mulatta) et, bien sûr, au macaque crabier (Macaca fascicularis) qui est son hôte naturel :
- tandis que le macaque japonais présente des symptômes sévères proches du paludisme humain, le macaque crabier récupère rapidement après une légère anémie (Matsumoto et al., 2000);
- le macaque rhésus présente une pathologie également proche du neuropaludisme humain (Sein et al., 1993);
- enfin, plus étonnant, au sein même des macaques crabiers, P. coatneyi pourrait engendrer une mortalité chez les primates provenant de l’île Maurice, alors que ceux provenant des Philippines survivraient (Migot-Nabias et al., 1999).
c- avantage des primates américains
Le modèle Aotus est le modèle recommandé par l’OMS pour les études sur le paludisme (Pico de Coana et al., 2003) car ses avantages sont nombreux :
- éviter la splénectomie pour infecter les animaux, - parasitémies prévisibles, hautes et qui n’avortent pas au bout de 2-3 cycles, - possibilité de tester plus rapidement les vaccins tout en assurant une innocuité future
(Stowers & Miller, 2001); - excellent modèle (comme le saïmiri) pour les essais vaccinaux, que ce soit pour les
stades sanguins ou pré-érythrocytaires, dans la mesure où les souches plasmodiales sont bien sélectionnées (Collins, 1992);
- assez bonne corrélation avec les cellules B et T humaines (Herrera et al., 2002); - existence de réactions croisées avec les marqueurs de cytokines et de lymphocytes
humains (Herrera et al., 2002); - existence de similarités avec les CMH (complexes majeurs d’histocompatibilité)
humains (Nino-Vasquez et al., 2000); - bon modèle de paludisme sévère tel que celui des enfants africains (Egan et al., 2002),
etc.
D.2- CONTRAINTES FINANCIERES
La taille plus imposante des primates africains augmente considérablement les coûts d’entretien et d’anesthésie. Ainsi, au Centre de Primatologie du CIRMF :
• la ration journalière d’un chimpanzé de 50 kg revient à 1,5 euro, soit 10 fois plus que pour un macaque (les bananes et les autres fruits étant acheté localement à des prix assez bas) ;
• le coût de fabrication et d’entretien des cages est aussi bien plus élevé car la force et l’intelligence de ces derniers nécessitent des structures extrêmement solides ;
• les coûts des soins quotidiens délivrés par les animaliers sont augmentés : dans les conditions de notre centre, un chimpanzé requiert en moyenne 1/25ème du poste d’un animalier, contre 1/50ème pour un macaque ;

Les primates africains et la recherche sur le paludisme
90
• le coût des soins thérapeutiques et des anesthésiques est proportionnel au poids de l’animal. Il faut également ajouter le prix du matériel de télé-anesthésie, indispensable dans le cas des grands primates africains. Ainsi, pour un chimpanzé de 50 kg, le coût d’une simple anesthésie varie entre 30 et 50 euros alors qu’un macaque de 5-10 kg en cage individuelle à paroi coulissante peut être anesthésié pour moins d’un euro.
Un protocole lié au paludisme dure en général un à deux mois (une quinzaine de jours pour l’apparition des parasites puis encore souvent 1 mois pour l’étude de la parasitémie). Les primates subissent deux à trois prélèvements sanguins hebdomadaires, ce qui représente une vingtaine d’anesthésies par animal par protocole. Par ailleurs, pour obtenir des résultats exploitables, il est difficile d’utiliser moins de 4 animaux. Ainsi, l’utilisation de 5 chimpanzés adultes pendant 40 jours pour un protocole sur le paludisme coûte environ 4300 euros, juste en frais de nourriture et d’anesthésie, sans compter les frais d’entretien, d’analyses, ni les frais vétérinaires.
D.3- CONTRAINTES ECOLOGIQUES
D.3.1- Disponibilité du modèle
Selon les classifications données par la Convention de Washington (CITES) ou l’IUCN (International Union for Conservation of Nature and Natural Resources), respectivement 239 à 333 espèces de primates sont menacées. Parmi ces espèces, on retrouve la plupart des modèles utilisés pour la recherche sur le paludisme. Les primates du Nouveau-Monde couramment utilisés en recherche comme les singes de nuits (Aotus sp.), les singes hurleurs (Alouatta fusca) ou les tamarins (Saguinus sp.) sont classés dans l’appendice II de la liste CITES. Cela signifie qu’ils ne sont pas directement menacés actuellement mais qu’ils pourraient le devenir si aucune mesure n’est prise pour les protéger. Par ailleurs, alors qu’ils étaient autrefois menacés, les singes de nuit bénéficient aujourd’hui de structures d’élevage (Rieckmann et al., 1980) et les saïmiris sont eux aussi facilement disponibles grâce à la colonie formée en Guyane par l’Institut Pasteur (Perraut & Gerraud, 1998). En revanche, en Afrique la situation est différente : les chimpanzés (Pan troglodytes) ou les mandrills (Mandrillus sphinx) sont classés en annexe I de la Convention, ce qui signifie qu’ils sont déjà menacés d’extinction. Ce statut entraîne pour le chercheur l’interdiction de l’euthanasie à la fin des protocoles de recherche, induisant ainsi un surcoût important (entretien d’un animal pendant 40 à 50 ans dans le cas d’un chimpanzé).
D.3.2- Envoi des prélèvements
L’envoi d’échantillons (sang, sérum, etc.) de primates africains est complexe : lorsqu’une espèce est inscrite à l’annexe I de la convention de Washington, tout commerce international d’animaux ou même d’échantillons des animaux de cette espèce est très réglementé. Des circonstances exceptionnelles sont accordées dans le cadre de la recherche scientifique, mais l’envoi doit alors être accompagné conjointement d’un permis d’importation et d’un permis

Les primates africains et la recherche sur le paludisme
91
d’exportation (CITES, 2004). Ces démarches administratives sont lourdes et retardent souvent les envois. En revanche, seul le permis d’exportation est nécessaire dans le cadre de l’annexe II de la CITES. Quant à l’envoi international d’échantillons humains (frottis, gouttes épaisses par exemple), il ne nécessite qu’un certificat d’exportation délivré par les autorités locales. Celui-ci peut être obtenu rapidement (quelques jours au Gabon). Les mêmes échantillons, s’ils proviennent de chimpanzés, nécessitent 3 à 6 mois de procédures administratives avant leur envoi…
D.4- CONTRAINTES ETHIQUES ET LEGISLATIVES
D.4.1- Ethique et utilisation des primates
D.4.1.1- bases de la réflexion éthique
Le débat sur l’utilisation des primates non-humains est âpre (Goodman & Check, 2002). Il y a peu de sujets en science qui déclenchent autant de réactions que l’expérimentation sur nos cousins primates. En effet, le niveau cognitif des primates étant parmi les plus élevés, cela signifie qu’ils sont plus sensibles à leur environnement. Cela signifie-t-il qu’ils devraient être davantage protégés ? Pour le scientifique, le primate est souvent le seul modèle animal capable de reproduire les réponses humaines. Pour les activistes des droits de l’animal, ceux qui utilisent les primates sont les pires des « abuseurs » d’animaux. L’opinion publique diffère selon le pays concerné : en Grande-Bretagne par exemple, la pression des ligues de protection animale est extrêmement forte.
a- la règle des 3 R : Remplacer, Réduire, Raffiner.
Il y a cinquante ans, deux scientifiques britanniques définissaient les trois objectifs à atteindre dans le domaine de l'expérimentation sur le vivant (Russell & Burch, 1959) :
1) Remplacer les méthodes utilisant des vertébrés vivants conscients ; 2) Réduire le nombre d'animaux utilisés par :
a. la limitation aux seules expériences considérées comme absolument indispensables,
b. le rejet par un comité d'éthique d'investigations discutables, c. le fait d’éviter les répétitions inutiles.
3) Raffiner, c’est-à-dire diminuer les conséquences des procédures infligées aux animaux.
Toutefois, dans certains cas, il peut y avoir incompatibilité entre les trois objectifs de cette règle. En effet, la réduction du nombre d'animaux peut entraîner l'augmentation des contraintes et vice versa. Il est parfois question d'un quatrième voire d’un cinquième R, correspondant au R de respect dû à tout animal et au R de responsabilité des différents intervenants dans la recherche.

Les primates africains et la recherche sur le paludisme
92
b- comités d’éthique
La règle des 3 R correspond à un code de bonne pratique, mais elle ne permet pas de décider si les expérimentations sont recevables ou non. Afin d'aider l'évaluation éthique des protocoles, trois éléments majeurs doivent être pris en considération : les contraintes imposées aux animaux, l'intérêt de l'expérimentation pour la société et la qualité du plan expérimental. Un modèle de document de saisine du Comité Régional d’Ethique Ile de France Sud (auquel est rattaché le CIRMF) est présenté en annexe 7. La composition des comités d’éthique varie selon les pays : uniquement des scientifiques ou présence conjointe de membres de ligues de protection animale et/ou présence de représentants de la société civile (comités australiens) (Veissier, 1999). La valeur du comité d’éthique est nulle d’un point de vue légal mais cet avis est cependant devenu nécessaire pour pouvoir publier des résultats dans une revue scientifique.
D.4.1.2- cas particulier des grands singes
Les travaux sur les primates africains ne concernent quasiment que les chimpanzés. Ce sont les primates hominoïdes les plus utilisés en recherche. En effet, les gorilles et les orangs-outans ne sont pas utilisés en laboratoire, seuls quelques prélèvements chez des singes sauvages sont réalisés à des fins scientifiques (Verschoor et al., 2001). Les premières considérations particulières à l’utilisation des chimpanzés remontent aux années 60 (Cook et al., 1961). Dans les années 80, l’accent était mis sur la nécessité de préserver les populations sauvages (Cummins & Kalter, 1980) mais les chimpanzés étaient alors très utilisés pour les recherches concernant le SIDA (modèle non pathogène pour le HIV1) (Rutjens et al., 2003). C’est à la fin des années 80 que l’on commence sérieusement à se préoccuper de la « retraite » des animaux utilisés dans les protocoles (Eichberg & Speck, 1988) et du problème éthique lié à leur utilisation (Warshauer, 1985). Cette réflexion se poursuit dans les années 90 (van Akker et al., 1994) avec la création d’un cadre législatif pour les conditions de captivité des grands singes et leur devenir (Vogel, 2000 ; Goodman, 2001).
D.4.1.3- application à la recherche sur le paludisme
Les recherches sur le paludisme présentent plusieurs spécificités :
les protocoles sont plutôt courts (quelques mois) ; ils nécessitent souvent des anesthésies assez rapprochées pour suivre les parasitémies
(tous les 2-3 jours en moyenne) ; le développement de méthodes de coopération (Scott et al., 2003) est parfois
intéressant : en travaillant sur une technique d’apprentissage et de récompense, un primate peut rapidement (une heure au total pour un macaque adulte) apprendre à tendre la jambe pour subir une injection ou une prise de sang (Reinhardt, 2003). Cela permet d’une part d’éviter une anesthésie, d’autre part le stress lié à la contrainte, puisque c’est l’animal qui « choisit » d’être manipulé. Le degré de contrainte alors indiqué au comité d’éthique peut être « minime ».

Les primates africains et la recherche sur le paludisme
93
Ces techniques sont cependant trop dangereuses à mettre en pratique lorsque le primate est porteur d’agents infectieux dangereux pour l’humain (plasmodiums humains, HIV, SIV, etc.). 2 catégories de matières dangereuses, A et B, sont définies dans l’Accord Européen
relatif au transport international des marchandises dangereuses (J.O. du 27/12/2002) ; le plasmodium est par défaut classé parmi les matières de catégorie A car il peut potentiellement entraîner la mort (Institut Pasteur, 2005). Il convient donc de bien préciser la sensibilité du modèle utilisé pour moduler cette notion lors de la demande d’accord au comité d’éthique. Par ailleurs, les traitements humains pouvant être administrés au primate, rares sont les
infections mortelles (sauf bien sûr dans le cas des recherches menant volontairement à la mort). Les animaux doivent être placés dans des locaux isolés des moustiques. Les animaux peuvent être en groupe (sauf dans le cas d’étude d’infection par piqûre de
moustique), ce qui est primordial pour le bien-être des primates. Le degré de souffrance global de l’animal diffère considérablement d’un protocole à l’autre :
- le degré de souffrance physique : o les chimpanzés ne présentent presque pas de symptômes cliniques ; o les babouins, peuvent par contre développer un paludisme sévère avec certains
plasmodiums ; o les biopsies hépatiques et les splénectomies sont un élément important à
prendre en compte pour les risques secondaires de péritonites. - le degré de souffrance psychique :
o les chimpanzés sont des primates hominoïdes, ils possèdent un niveau cognitif très élevé et sont extrêmement sensibles à leur environnement ;
o les babouins sont moins sensibles.
Ainsi, dans le cadre de l’utilisation des primates africains, on est confronté : - soit à des animaux peu sensibles aux plasmodiums mais très sensibles à leur
environnement, - soit à des animaux aux capacités cognitives plus basses mais alors souvent utilisés
dans un contexte d’infection pathogène (comme les primates sud-américains). Les demandes d’accord du comité d’éthique comportent donc un degré de contrainte allant de « faible », pour les protocoles incluant des chimpanzés sans biopsie par exemple, à « moyenne » ou « forte » si le primate est très sensible au plasmodium et/ou que des biopsies sont prévues.
D.4.2- Ethique et législation
La règle des 3 R a reçu un large consensus. Elle a été adoptée par plusieurs institutions, dont le Conseil Canadien de Protection des Animaux (1993), le National Health and Medical Research Council australien (1997), le Département Américain chargé de l'Agriculture (1991) et le gouvernement britannique (1998) (Veissier, 1999). Par ailleurs, le Conseil de l’Europe s’est réuni dans le cadre de la convention européenne sur la protection des animaux vertébrés utilisés à des fins expérimentales ou à d'autres fins scientifiques.

Les primates africains et la recherche sur le paludisme
94
Les Etats signataires reconnaissent ainsi en préambule : - l'obligation morale de l’homme de respecter tous les animaux et de prendre dûment en
considération leur aptitude à souffrir et à se souvenir, - mais aussi que l'homme, dans sa quête de connaissance, de santé et de sécurité, a
besoin d'utiliser des animaux lorsqu'on peut raisonnablement espérer que cela fera progresser la connaissance, ou produira des résultats utiles d'une façon générale pour l'homme ou pour l'animal, au même titre qu'il utilise les animaux pour se nourrir, pour se vêtir et comme bêtes de somme (Conseil de l'Europe, 1986).
L’Union Européenne a ratifié la Directive n°86/609/CEE qui devrait bientôt être modifiée suite au rapport de Berlin (cf position commune du Parlement et du Conseil 2003/C 113 E/04). En France quatre textes sont relatifs à l’expérimentation animale :
- Décret n° 1987-848, - Arrêté du 19 avril 1988, - Décret n° 2001-464, - Décret n° 2001-486.
Depuis une vingtaine d’années, la législation impose des normes de plus en plus drastiques concernant le logement et les soins à apporter aux animaux, avec des mentions spécifiques aux primates. Cas particulier des chimpanzés : En Europe, l’utilisation de chimpanzés pour la recherche est presque partout interdite. Aux Etats-Unis elle est davantage réglementée depuis 2000. Certaines associations de protection animale veulent aller au delà de ces interdictions, comme le projet grands singes qui vise à élargir les droits accordés aux êtres humains à tous les grands singes (Cavalieri & Singer, 2004). Ainsi, si le chimpanzé représente d’un point de vue scientifique le modèle le plus intéressant pour la recherche sur le paludisme, les contraintes éthiques et écologiques associées à ce modèle le rendent très peu utilisable dans le cadre du laboratoire. Toutefois des recherches, aussi peu invasives que possible, sur les populations sauvages devraient s’avérer très précieuses sur la connaissance du paludisme. Quant aux autres primates africains, ils sont encore mal connus, mais ils semblent posséder des qualités qui sont encore sous-exploitées dans la recherche sur le paludisme.

Les p
rim
ates
afr
icai
ns e
t la
rech
erch
e su
r le
palu
dism
e
95
Mod
èle
in v
itro
étud
e di
rect
e ch
ez
l’hum
ain
(s
ans p
rim
ate)
ro
ngeu
rs
prim
ates
su
d-am
éric
ains
m
acaq
ues
prim
ates
afr
icai
ns
Ava
ntag
es
Coû
t Et
hiqu
e G
ain
de te
mps
Coû
t D
ispo
nibi
lité
Prox
imité
de
leur
s pl
asm
odiu
ms
Tran
sgen
èse
poss
ible
Coû
t G
rand
e se
nsib
ilité
à
P. fa
lcip
arum
Pr
oxim
ité d
es
syst
èmes
im
mun
itaire
s (m
ais
disc
utab
le se
lon
les
mod
èles
)
Coû
t D
ispo
nibi
lité
Sens
ibili
té à
pl
usie
urs
plas
mod
ium
s N
ombr
eux
outil
s et
conn
aiss
ance
s sur
ce
mod
èle
Sens
ibili
té a
ux 4
pl
asm
odiu
ms
hum
ains
Se
nsib
ilité
aux
sp
oroz
oïte
s Pr
oxim
ité d
u pl
asm
odiu
m n
atur
el
Rés
ulta
ts
préd
ictib
les
Nom
breu
x pr
élèv
emen
ts
poss
ible
s
Inco
nvén
ient
s N
e re
prod
uit p
as le
s in
tera
ctio
ns d
ans
l’org
anis
me
Ethi
que
Diff
éren
ces d
’hôt
es
Inse
nsib
ilité
aux
pl
asm
odiu
ms
hum
ains
Petit
e ta
ille
Peu
disp
onib
le
Peu
sens
ible
s aux
in
fect
ions
par
les
spor
ozoï
tes
Inse
nsib
ilité
aux
pl
asm
odiu
ms
hum
ains
Coû
t D
ispo
nibi
lité
Ethi
que
Prin
cipa
ux
dom
aine
s d’
utili
satio
n
Toxi
cité
cel
lula
ire
Bio
logi
e du
par
asite
Crib
lage
des
an
tigèn
es c
andi
dats
au
x va
ccin
s
Etud
e de
s ré
sist
ance
s Ep
idém
iolo
gie
Cer
tain
s ess
ais
vacc
inau
x
Bio
logi
e du
par
asite
To
xico
logi
e Im
mun
olog
ie
Pré-
essa
is
vacc
inau
x
Etud
e du
pal
udis
me
sévè
re
Vac
cins
stad
es
éryt
hroc
ytai
res
Essa
i clin
ique
d’
antip
alud
ique
Etud
e du
ne
urop
alud
ism
e Es
sai c
liniq
ue
d’an
tipal
udiq
ue
Essa
is v
acci
naux
Et
ude
du p
alud
ism
e sé
vère
M
odèl
e no
n pa
thog
ène
T
able
au 6
: ré
capi
tula
tif d
es d
iffér
ents
mod
èles
d’é
tude
du
palu
dism
e


TTRROOIISSIIEEMMEE PPAARRTTIIEE :: EEXXEEMMPPLLEESS DD’’UUTTIILLIISSAATTIIOONN DDEESS PPRRIIMMAATTEESS AAFFRRIICCAAIINNSS DDAANNSS LLAA
RREECCHHEERRCCHHEE SSUURR LLEE PPAALLUUDDIISSMMEE


Exemples d’utilisation des primates africains dans la recherche sur le paludisme
99
Quatre protocoles sont décrits ici, réalisés au CIRMF (Gabon) entre 1999 et 2004.
A- UTILISATIONS DE PRIMATES AFRICAINS AVEC LEUR PLASMODIUM NATUREL
A.1- ETUDE D’UNE NOUVELLE SOUCHE PLASMODIALE NATURELLE DU CHIMPANZE
Une nouvelle espèce plasmodiale, baptisée P. benjamini, a été découverte au CIRMF chez un chimpanzé. Or, comme démontré précédemment, le chimpanzé représente un modèle de choix pour les études sur le paludisme. En outre, il existe assez peu de modèles intéressants de plasmodiums naturels chez le primate. Du fait de la proximité des hôtes et de leurs plasmodiums déjà décrits, il semblait donc important d’étudier plus avant cette espèce plasmodiale.
A.1.1- Caractéristiques morphologiques
Les trophozoïtes ressemblent beaucoup à ceux de P. falciparum : ce sont de petits anneaux fins situés dans des érythrocytes non déformés. On peut observer plusieurs anneaux dans un même globule rouge et, comme dans le cas de P. falciparum, les anneaux sont plutôt en marge des globules. Toutefois, la particularité des trophozoïtes de P. benjamini est de posséder un noyau légèrement plus gros que P. falciparum (comme P. reichenowi). Les schizontes sont également proches de ceux de P. falciparum, même si certaines formes observées peuvent aussi évoquer l’aspect en marguerite de P. malariae. Les gamétocytes enfin, sont très proches de ceux de P. falciparum, en forme de « saucisse » assez épaisse.
A.1.2- Caractéristiques phylogénétiques
La caractérisation génétique s’appuie sur une PCR nichée classique utilisant comme amorces : - lors de la première PCR : rPLU5 et rPLU6, - puis à la deuxième PCR : rFAL1 et rFAL2 (P. falciparum) ou rMAL1 et rMAL2 (P.
malariae) ou rOVA1 et rOVA2 (P. ovale) ou rVIV1 et rVIV2 (P. vivax). Dans le cadre de P. benjamini, ces amorces ont été ajoutées aux primers spécifiques de P. reichenowi désignés à partir des séquences connues, soit 11 gènes (annexe 8) :
- gène EBP, - gène GLURP, - gène Prs48/45, - gène Cysteine proteinase, - gène Cytochrome b, - gène MSA-2, - gène Antigene 27 du gamète, - gène de l’antigène Sporozoïte,

Exemples d’utilisation des primates africains dans la recherche sur le paludisme
100
- gène CSP, - gène de l’antigène de l’ookinète 25 kDa, - gène MSP-3.
A.1.2.1- arbre phylogénétique
Les arbres ont été construits à partir du gène du cytochrome b, du gène de MSP-3, de CSP, de la cystéine protéinase et de la ssr ARN. Comme les caractéristiques morphologiques laissaient l’envisager, P. benjamini est très proche de P. falciparum et de P. reichenowi (figures 12 et 13).
Figure 12 : P. benjamini dans l’arbre phylogénétique de quelques espèces plasmodiales simiennes et humaines. Construction à partir du gène codant pour la cystéine protéase.
(Ollomo, 2004, non publié) P. benjamini possède pour le gène de la cystéine protéase :
o 91 à 98% d’homologie avec P. falciparum o 92 à 99% d’homologie avec P. reichenowi

Exemples d’utilisation des primates africains dans la recherche sur le paludisme
101
Figure 13 : P. benjamini dans l’arbre phylogénétique de quelques espèces plasmodiales simiennes et humaines. Construction à partir du gène codant pour le cytochrome b (1028 bp).
(Ollomo, 2004, non publié)
Le gène du cytochrome b de P. benjamini possède 82% d’homologie avec celui de P. reichenowi. Concernant les autres gènes, les homologies sont aussi importantes :
- gène de l’antigène ookinète o 94% d’homologie avec P. falciparum o 93% d’homologie avec P. reichenowi
- gène de la ssr ARN o 93 à 95% d’homologie avec P. falciparum o 98% d’homologie avec P. reichenowi
A.1.2.2- homologie des gènes utilisés en vaccination
Deux gènes ont été étudiés, MSP-3 et CSP. - MSP-3
o 84 à 96% d’homologie avec P. falciparum o 96% d’homologie avec P. reichenowi
- CSP o 77 à 96% d’homologie avec P. falciparum o 81% d’homologie avec P. reichenowi
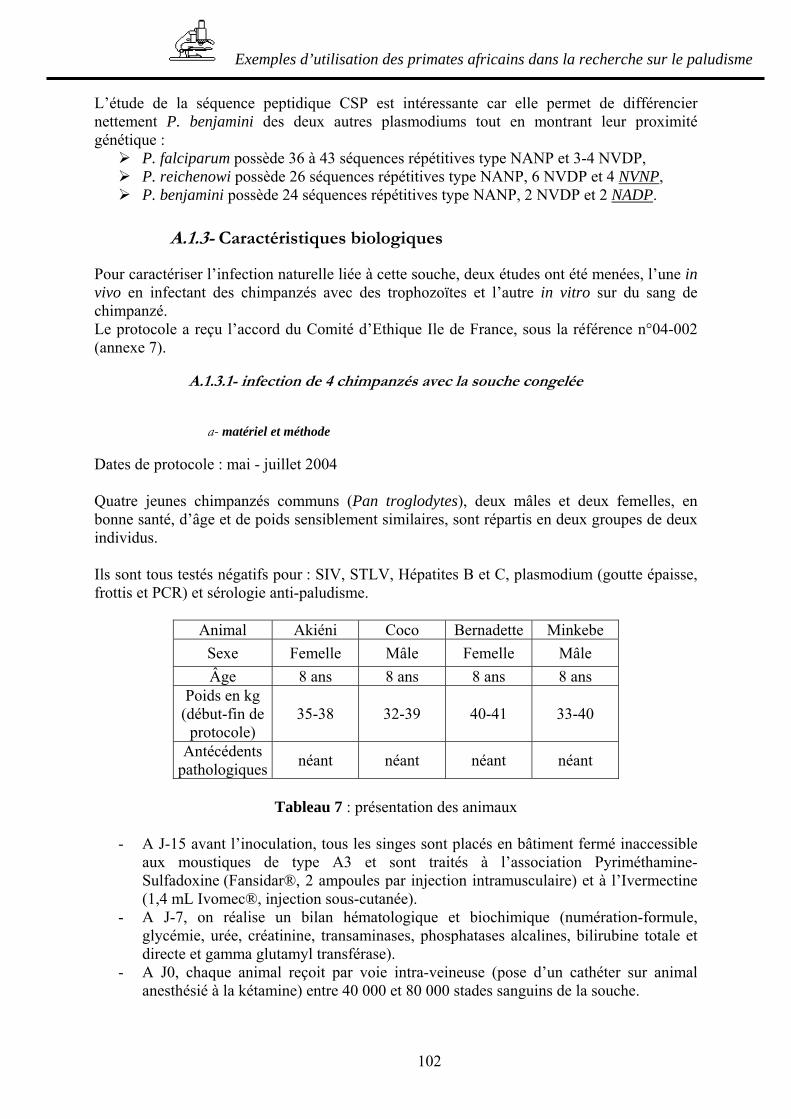
Exemples d’utilisation des primates africains dans la recherche sur le paludisme
102
L’étude de la séquence peptidique CSP est intéressante car elle permet de différencier nettement P. benjamini des deux autres plasmodiums tout en montrant leur proximité génétique :
P. falciparum possède 36 à 43 séquences répétitives type NANP et 3-4 NVDP, P. reichenowi possède 26 séquences répétitives type NANP, 6 NVDP et 4 NVNP, P. benjamini possède 24 séquences répétitives type NANP, 2 NVDP et 2 NADP.
A.1.3- Caractéristiques biologiques
Pour caractériser l’infection naturelle liée à cette souche, deux études ont été menées, l’une in vivo en infectant des chimpanzés avec des trophozoïtes et l’autre in vitro sur du sang de chimpanzé. Le protocole a reçu l’accord du Comité d’Ethique Ile de France, sous la référence n°04-002 (annexe 7).
A.1.3.1- infection de 4 chimpanzés avec la souche congelée
a- matériel et méthode
Dates de protocole : mai - juillet 2004 Quatre jeunes chimpanzés communs (Pan troglodytes), deux mâles et deux femelles, en bonne santé, d’âge et de poids sensiblement similaires, sont répartis en deux groupes de deux individus. Ils sont tous testés négatifs pour : SIV, STLV, Hépatites B et C, plasmodium (goutte épaisse, frottis et PCR) et sérologie anti-paludisme.
Animal Akiéni Coco Bernadette Minkebe Sexe Femelle Mâle Femelle Mâle Âge 8 ans 8 ans 8 ans 8 ans
Poids en kg (début-fin de
protocole) 35-38 32-39 40-41 33-40
Antécédents pathologiques néant néant néant néant
Tableau 7 : présentation des animaux
- A J-15 avant l’inoculation, tous les singes sont placés en bâtiment fermé inaccessible
aux moustiques de type A3 et sont traités à l’association Pyriméthamine-Sulfadoxine (Fansidar®, 2 ampoules par injection intramusculaire) et à l’Ivermectine (1,4 mL Ivomec®, injection sous-cutanée).
- A J-7, on réalise un bilan hématologique et biochimique (numération-formule, glycémie, urée, créatinine, transaminases, phosphatases alcalines, bilirubine totale et directe et gamma glutamyl transférase).
- A J0, chaque animal reçoit par voie intra-veineuse (pose d’un cathéter sur animal anesthésié à la kétamine) entre 40 000 et 80 000 stades sanguins de la souche.

Exemples d’utilisation des primates africains dans la recherche sur le paludisme
103
Akiéni
0 20000 40000 60000 80000
100000 120000
0 6 12 18 24 28 32 37 43 50 57 68
- La souche, conservée en azote liquide, provient d’un protocole d’étude réalisé au CIRMF (infection d’un chimpanzé du centre avec la souche « sauvage», prélevée sur un chimpanzé gabonais).
Akiéni Coco Bernadette Minkebe
Ordre d’inoculation 4 3 2 1 Nombre approximatif de
parasites inoculés 40 000 52 000 80 000 80 000
Tableau 8 : protocole d’infection des animaux
Des prélèvements sanguins sont effectués pour suivre la parasitémie (frottis sanguin, goutte épaisse et QBC) :
- tous les 3 à 4 jours lorsque la parasitémie est < 5000 parasites/μL, - tous les 2 jours lorsque la parasitémie est > 5000 parasites/μL, - toutes les 8 heures pendant 24 heures au moment du pic de parasitémie de l’animal qui
possède la plus forte parasitémie, afin de rechercher la présence de schizontes. Tous les 7 à 10 jours un bilan hématologique et biochimique complet est réalisé. La parasitémie est suivie pendant 2 mois. A J68 tous les animaux sont traités à l’association Pyriméthamine-Sulfadoxine (Fansidar®, 2 ampoules par voie intramusculaire). A J79 un contrôle est effectué avant de replacer les animaux dans leurs volières d’origine.
b- résultats
Les quatre animaux ont été réceptifs au parasite. Toutefois, à la suite du premier inoculum, Bernadette n’a développé aucune parasitémie jusqu’à J35. Elle a donc été ré-inoculée par voie intra-veineuse avec 0,47mL de sang frais de Coco à J35. Ce dernier présentait alors une parasitémie de 170 parasites/μL (sous forme de trophozoïtes sur le frottis), donc le volume injecté à Bernadette représentait environ 80 000 parasites.
Les premiers parasites sanguins ont été détectés à J9 pour un animal et J18 pour les
trois autres. Les pics parasitaires ont été observés à J18 pour un animal et J26 pour les trois autres. L’infection sanguine s’est prolongée jusqu’à la fin du protocole.
Figure 14 a

Exemples d’utilisation des primates africains dans la recherche sur le paludisme
104
Minkebe
0
5000
10000
15000
20000
0 6 12 18 23 28 32 37 43 50 57 68
Bernadette
0 20000 40000 60000 80000
100000 120000
0 2 5 8 10 12 14 16 18 19 20 22 26 33
Coco
0 10000 20000 30000 40000 50000
0 6 12 18 24 28 32 37 43 50 57 68
Figure 14 b
Figure 14 c
Figure 14 d
Figure 14 a, b, c, d : parasitémies (en nombre de parasites/μL) en fonction des jours post-inoculation
Les variations de température n’ont pas pu être directement corrélées à la parasitémie. Toutefois un pic suivi d’une chute de température a été observé autour de l’apparition des parasites en phase sanguine (J18 post inoculation pour Akiéni, Coco et Minkebe, J9 pour Bernadette) (figures 15 a à d).

Exemples d’utilisation des primates africains dans la recherche sur le paludisme
105
Akiéni
36,5
37
37,5
38
38,5
39
0 3 6 9 12 15 18 21 24 27 30 33 36 39 42 45 48 51 54 57 60
0
20000
40000
60000
80000
100000
120000
Coco
36,5
37
37,5
38
38,5
0 3 6 9 12 15 18 21 24 27 30 33 36 39 42 45 48 51 54 57 60
0
5000
10000
15000
20000
25000
30000
35000
40000
45000
Minkebe
37
37,5
38
38,5
0 3 6 9 12 15 18 21 24 27 30 33 36 39 42 45 48 51 54 57 60
0
2000
4000
6000
8000
10000
12000
14000
16000
18000
20000
Figure 15 a
Figure 15 b
Figure 15 c

Exemples d’utilisation des primates africains dans la recherche sur le paludisme
106
Bernadette
36,5
37
37,5
38
38,5
1 4 7 10 13 16 19 22 25 28 31 34
0
20000
40000
60000
80000
100000
120000
Figure 15 d
Figures 15 a, b, c, d : parasitémies ( ) (en nombre de parasites/μL) et températures (------) (en degrés Celsius) en fonction des jours post-inoculation (→ : apparition des parasites dans le sang)
Clinique : aucun symptôme n’a été observé.
Seules les données hématologiques et biochimiques apportent quelques éléments sur la pathogenèse de l’infection. Quatre paramètres sont principalement modifiés :
o le taux d’hémoglobine (normes : 11-16,5 g/dL), o le taux d’érythrocytes (normes : 3,8-5,8 106/mm3), o l’hématocrite (normes 35-50 %), o le pourcentage de lymphocytes dans la formule sanguine (normes 17-48 %).
Les données présentées ici (figures 16 à 19) concernent seulement les 3 chimpanzés qui ont développé l’infection à partir de la souche conservée en azote liquide. Bernadette n’a pu être retenue pour l’élaboration de ces courbes car elle n’a pas eu assez de prélèvements sanguins entre le moment où les parasites sont apparus en phase sanguine et la fin du protocole (3 prélèvements seulement). On peut toutefois signaler que ses paramètres ne sont pas contradictoires avec ceux relevés pour les trois autres animaux.
Chaque figure représente un paramètre sanguin (6 prélèvements par paramètre ont été effectués). Un paramètre donné est décrit par la courbe de tendance de chaque animal (3 courbes par figure) ; ces courbes sont des polynomiales de 3ème degré et R2 représente le coefficient de détermination de chaque courbe de tendance. Plus il est élevé, plus la courbe de tendance décrit fidèlement les données réelles des 6 prélèvements (R2 est compris entre 0 et 1).
L’échelle des abscisses représente les jours d’infection sanguine:
o J0 correspond à l’apparition du parasite en phase sanguine, o J8 correspond au pic de parasitémie, o J50 correspond à l’administration du Fansidar®.

Exemples d’utilisation des primates africains dans la recherche sur le paludisme
107
R2 = 0,99
R2 = 0,99
R2 = 0,99
8
9
10
11
12
13
14
15
0 20 40 60
Polynomial (Coco)Polynomial (Akiéni)Polynomial (Minkebe)
Figure 16 : variation du taux d’hémoglobine en fonction du temps (en jours, J0= apparition des parasites dans le sang), 11g/dL représente la borne inférieure de la norme.
Figure 17 : variation du taux d’érythrocytes en fonction du temps (en jours, J0= apparition des parasites dans le sang), 3,8 106/mm3 représente la borne inférieure de la norme.
R2 = 0,99
R2 = 0,98
R2 = 0,96
2,5
3
3,5
4
4,5
5
5,5
6
0 20 40 60
Polynomial (Coco)Polynomial (Akiéni)Polynomial (Minkebe)
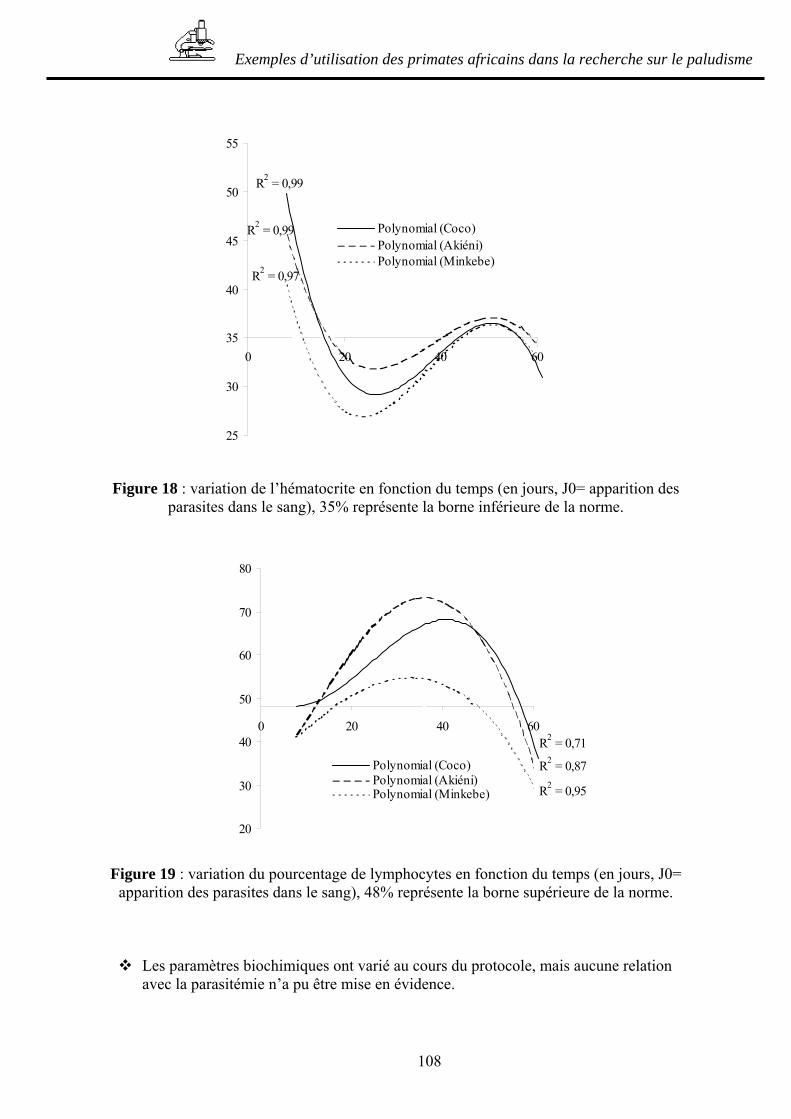
Exemples d’utilisation des primates africains dans la recherche sur le paludisme
108
R2 = 0,99
R2 = 0,99
R2 = 0,97
25
30
35
40
45
50
55
0 20 40 60
Polynomial (Coco)Polynomial (Akiéni)Polynomial (Minkebe)
R2 = 0,71
R2 = 0,95
R2 = 0,87
20
30
40
50
60
70
80
0 20 40 60
Polynomial (Coco)Polynomial (Akiéni)Polynomial (Minkebe)
Figure 18 : variation de l’hématocrite en fonction du temps (en jours, J0= apparition des parasites dans le sang), 35% représente la borne inférieure de la norme.
Figure 19 : variation du pourcentage de lymphocytes en fonction du temps (en jours, J0= apparition des parasites dans le sang), 48% représente la borne supérieure de la norme.
Les paramètres biochimiques ont varié au cours du protocole, mais aucune relation avec la parasitémie n’a pu être mise en évidence.

Exemples d’utilisation des primates africains dans la recherche sur le paludisme
109
c- discussion
• Le chimpanzé semble assez sensible à cette espèce plasmodiale, malgré un premier échec d’infection chez Bernadette. Cet échec est peut-être dû à de mauvaises conditions de stockage de la souche (problème de renouvellement de l’azote) et/ou à une immunité particulière de cet animal.
• Entre 40 000 et 80 000 parasites, la dose inoculée n’a pas d’influence sur la
parasitémie future (Bernadette avait reçu 80 000 parasites et n’a pas permis le développement du parasite, alors que Akiéni en avait reçu deux fois moins et a développé la plus forte parasitémie).
• Seuls des trophozoïtes et des gamétocytes (en fin d’infection, lorsque les parasitémies
étaient autour de 1000/μL) ont été observés in vivo. Aucun schizonte n’a pu être observé in vivo, même lors des 3 prélèvements effectués en 24h chez Akiéni au moment de son pic de parasitémie. Si ces schizontes n’apparaissent pas dans le sang alors que la parasitémie des chimpanzés dure longtemps (renouvellement assuré par les schizontes), c’est qu’il pourrait y avoir séquestration de ces schizontes, comme dans le cas de P. falciparum.
• On distingue 3 types de courbes de parasitémie en fonction de la hauteur du pic de
parasitémie et de la décroissance de cette parasitémie vers des valeurs inférieures à 1000/μL :
1- courbe présentant un pic élevé de parasites (de l’ordre de 100 000/μL), associé
à une très forte et rapide (1 à 4 jours) chute de la parasitémie à des valeurs inférieures à 1000/μL. (Akiéni et Bernadette)
2- Courbe présentant un pic moyen de parasites (de l’ordre de 35 000/μL), associé une chute de la parasitémie plus lente (7 jours) vers des valeurs inférieures à 1000/μL, et ceci en deux étapes (existence d’un second pic). (Coco)
3- Courbe présentant un pic plus faible de parasites (de l’ordre de 18 000/μL), associé à une chute lente de la parasitémie vers des valeurs inférieures à 1000/μL (20 jours) avec 3 pics secondaires. (Minkebe)
Il semblerait donc que la hauteur du pic parasitaire soit associée à la rapidité de la décroissance parasitaire et à l’éventuelle existence de pics secondaires de parasitémie.
• Les 3 animaux infectés avec la souche congelée présentent exactement les mêmes dates d’apparition du parasite dans le sang et de pic parasitaire, respectivement J18 et J26 post inoculation, ce qui est assez remarquable.
• Ces délais d’apparition du parasite dans le sang sont bien plus longs que ceux observés
avec P. falciparum chez l’homme ou qu’avec P. reichenowi chez le chimpanzé (moins d’une semaine) (Coatney et al., 1971). Cela peut être dû à l’espèce plasmodiale elle-même ou à ses conditions de stockage.
• Bernadette, ré-infectée à partir du sang de Coco, présente une avancée de la date
d’apparition du parasite de J18 à J9, du pic de parasitémie de J26 à J18 et une réduction de l’intervalle entre ces événements de 9 à 8 jours. Deux hypothèses peuvent être avancées : d’une part l’augmentation de la virulence de la souche après ce premier

Exemples d’utilisation des primates africains dans la recherche sur le paludisme
110
passage sur le sang de Coco, d’autre part des facteurs inhérents à Bernadette conduisant à sa plus grande sensibilité lors de la ré-inoculation.
• L’absence de réel pouvoir pathogène de cette espèce plasmodiale n’est pas
surprenante : ni P. falciparum, ni P. reichenowi ne sont pathogènes pour le chimpanzé, il est donc logique que cette espèce plasmodiale voisine suive le même schéma clinique.
• L’anémie (décrite par l’hématocrite, les taux d’hémoglobine et d’érythrocytes)
apparaît précocement (entre 10 et 18 jours après l’apparition des parasites dans le sang), est maximale au bout de 3 semaines à 1 mois, et disparaît rapidement lorsque les parasitémies sont stabilisées autour de 1000/μL.
• Cette anémie, causée par l’invasion des érythrocytes par le plasmodium, est cependant
loin de celle décrite dans les cas de paludisme grave (hématocrite < 15 % ou hémoglobine < 5 g/dL) (Saissy et al., 2003).
• Le pourcentage de lymphocytes augmente de façon plus tardive (20 jours environ
après la détection de parasites sanguins). Son maximum coïncide avec le maximum de l’anémie mais le retour à des valeurs normales est plus tardif (dans la semaine suivant l’administration de Fansidar®).
• L’augmentation du pourcentage de lymphocytes n’entraîne pas pour autant de
lymphocytose, c’est plutôt une inversion de la formule sanguine. Elle correspond sans doute à l’augmentation modérée des lymphocytes qui participe à la protection (Wipasa et al., 2002).
A.1.3.2- culture in vitro
Au moment des pics de parasitémie de Coco, Akiéni puis de Bernadette, des essais de mise en culture du parasite sur du sang de chimpanzé (celui de Bernadette avant la deuxième inoculation) ont été menés. La maturation de la souche s’obtient en 48 heures (comme P. falciparum) : de nombreux schizontes sont observés, ainsi que des gamétocytes. Cependant, au bout de quelques jours, la souche apparaît « en souffrance » (allure anormale des trophozoïtes et des schizontes) et meurt rapidement (entre 6 et 9 jours). Ce problème existe déjà avec P. falciparum : si la culture des souches dites « de laboratoire » (3D7 par exemple) est assez facile, la culture in vitro des souches dites « de terrain » est très délicate. En outre, le fait d’avoir utilisé pour ces cultures le sang d’un animal réfractaire à l’infection a peut-être eu une importance. En raison de l’absence de schizonte dans le sang et à la suspicion de phénomène de séquestration, quatre essais de cytoadhérence ont été menés sur des cellules d’endothélium pulmonaire humain. L’étude de cytoadhérence consiste à mesurer le pourcentage de cellules endothéliales sur lesquelles sont accrochés un ou plusieurs globules rouges parasités (par rapport aux cellules endothéliales sans érythrocytes parasités). La relation entre la séquestration et les formes graves du paludisme n’est pas clairement établie : si dans le cas de neuropaludisme il y a

Exemples d’utilisation des primates africains dans la recherche sur le paludisme
111
séquestration importante dans les capillaires cérébraux (Wassmer et al., 2003a), il existe également un phénomène de séquestration lors des paludismes bénins à P. falciparum. Notre souche de P. benjamini est chaque fois comparée à une souche de terrain de P. falciparum fortement cytoadhérente (92%) qui possède la même proportion de schizontes. Tous les essais ont été négatifs : 0% pour P. benjamini contre 16 % pour la souche de P. falciparum qui est le témoin positif (92 % était le score obtenu avant la mise en culture de la souche sauvage, mais au bout d’une semaine, cette souche ne cytoadhère plus qu’à 16% sans doute par une sorte de perte phénotypique). Cela peut signifier :
- que notre souche n’a pas de propriétés de cytoadhérence, - ou que cette cytoadhérence n’a pas lieu dans l’épithélium pulmonaire, - ou plus vraisemblablement que cette cytoadhérence ne peut s’exercer sur des
cellules humaines (ce qui n’exclurait donc pas la séquestration des schizontes chez le chimpanzé).
Il pourrait donc y avoir séquestration des schizontes de cette espèce plasmodiale, avec une cytoadhérence spécifique (spécifique au tissu ou à l’espèce hôte), sans toutefois entraîner de symptômes graves chez le chimpanzé.
A.1.4- Conclusion et applications possibles
Cette nouvelle espèce plasmodiale, tout comme P. reichenowi, n’est donc pas pathogène pour le chimpanzé, ce qui a plusieurs conséquences :
cette espèce ne possède aucun intérêt pour l’étude de la pathogenèse du paludisme. En revanche, comme pour P. reichenowi, l’étude précise des mécanismes d’immunité mis en jeu ainsi que des relations hôtes-parasites (séquestration et cytoadhérence notamment) sont intéressants en tant que modèle non pathogène. D’autres applications comme l’étude de nouvelles molécules antipaludiques ou
de recherche vaccinale pourraient être menées comme avec P. reichenowi. Sur le plan écologique enfin, il est intéressant de constater encore une fois
l'absence de pathogénicité de cette espèce plasmodiale pour le chimpanzé, en réponses aux récentes inquiétudes concernant l'impact éventuel du paludisme sur les grands singes.

Exemples d’utilisation des primates africains dans la recherche sur le paludisme
112
A.2- ESSSAI D’UN VACCIN ERYTHROCYTAIRE ET D’UNE THERAPIE PAR ANTICORPS RECOMBINANTS SUR DES CHIMPANZES INFECTES PAR PLASMODIUM REICHENOWI
A.2.1- Principe de l’étude
MSP-3 est une molécule vaccinale de P. falciparum appartenant au stade érythrocytaire, identifiée comme cible des mécanismes de défense acquis par l’adulte africain devenu immun au parasite. Cette étude a deux objectifs :
o l’étude prospective de l’intérêt d’une construction vaccinale donnée. Il faut tester l’innocuité et l’antigénicité de la protéine PfMSP3/210-380 (produite dans le système Lactococcus lactis), émulsifiée avec de l’adjuvant SBAS2 et injectée chez 3 chimpanzés.
o L’évaluation du pouvoir protecteur de ce vaccin lors d’une infection par le parasite naturel du chimpanzé commun, P. reichenowi (l’antigène P.r.MSP3 présentant 99% d’homologie avec P.f.MSP3).
RAM-1 est quant à lui un anticorps recombinant anti-MSP-3 obtenu par clonage des
ADN complémentaires de lymphocytes de sujets africains dans un phage filamenteux. Une sélection des recombinants exprimant l’anticorps MSP-3.b est ensuite réalisée car, étant la mieux conservée, elle constitue la cible principale. Il y a ensuite reconstruction d’une immunoglobuline complète en ajoutant les chaînes lourdes ad hoc et enfin expression de l’anticorps recombinant sous forme d’isotypes IgG1et IgG3. Ces versions IgG1 et IgG3 de RAM-1 ont prouvé leur efficacité :
o in vitro avec un puissant effet anti-parasitaire : dans les tests de coopération avec les monocytes ils exercent une forte action contre P. falciparum ;
o chez les souris SCID immunocompromises greffées en globules rouges humains et infectées par P. falciparum, ces anticorps recombinants induisent une chute de la parasitémie en présence de monocytes humains greffés chez ces mêmes souris. Cet effet est comparable à celui obtenu avec la chloroquine ou la quinine et conduit à la chute à zéro de la parasitémie en 3 jours.
Cette action anti-parasitaire est spécifique des anticorps dirigés contre MSP-3 : l’utilisation d’anticorps dirigés contre d’autres molécules est sans effet sur la parasitémie.
Enfin, la production de tels anticorps recombinants par des plantes est possible et ceci à un coût extrêmement attractif, de l’ordre de 0,8 à 1,5 euro par traitement pour un adulte. Une nouvelle voie thérapeutique du paludisme apparaît donc possible.
Le protocole a reçu l’accord du Comité d’Ethique Ile de France sous la référence n° 02-001.

Exemples d’utilisation des primates africains dans la recherche sur le paludisme
113
A.2.2- Matériel et méthode
A.2.2.1- présentation des animaux
Six chimpanzés communs (Pan troglodytes), deux mâles et quatre femelles adultes, en bonne santé, d’âge et de poids sensiblement similaires, sont répartis en deux groupe de trois individus :
- 1er groupe = groupe des vaccinés : Congo, Doumay et Nuria ; - 2ème groupe = groupe contrôle : Ondele, Mopia et Makata.
Animal Congo Doumay Nuria Sexe Mâle Femelle Femelle Âge 13 ans 14 ans 16 ans
Poids en kg (début-fin
de protocole) 56,7-55 37,2-40 47,5-47,5
Antécédents pathologiques
STLV+ Hepatite B Ac+ Ag-
Hepatite C Ac+
Participation à un protocole paludisme à P. falciparum en
1998
Hépatite C Ac+
Tableau 9 a : groupe des vaccinés
Animal Ondele Mopia Makata
Sexe Femelle Femelle Mâle Âge 23 ans 24 ans 21 ans
Poids en kg (début-fin
de protocole) 38,8-41 ,5 41,5-45 41,2-47
Antécédents pathologiques
Hépatite B Ac+ Ag- Participation à un
protocole paludisme à P. falciparum en
1997
Hepatite B Ag+ en 1997 puis juste Ac+
Hépatite B Ag et Ac+ en 1997 puis
juste Ac+ Participation à un
protocole paludisme à P. falciparum en
1997
Tableau 9 b : groupe contrôle Tableau 9 a et b : présentation des animaux
Trois semaines avant la première immunisation, tous les animaux sont placés en bâtiment de type A3, fermé inaccessible aux moustiques, sont prélevés pour QBC, goutte épaisse et frottis et sont traités à l’Ivermectine (Ivomec®, 1,8 à 2,5 mL en injection sous-cutanée).

Exemples d’utilisation des primates africains dans la recherche sur le paludisme
114
A.2.2.2- protocole d’immunisation des chimpanzés
Dates de protocole : avril 2002 - janvier 2003 Groupe de singes Nombre de
chimpanzés Doses d’antigène MSP-3 injectées
Adjuvant utilisé Volumes injectés à chaque fois
Lot vacciné Lot témoin
3 adultes 3 adultes
30 / 30 / 30 μg 0 / 0 / 0 μg
SBAS2 SBAS2
500 μL 500 μL
Tableau 10 : protocole d’immunisation des animaux
Le groupe vacciné reçoit 3 doses de protéine vaccinale émulsifiée dans l’adjuvant SBAS2 tandis que le lot contrôle reçoit le même volume mais contenant du NaCl 9 ‰ émulsifié dans le même adjuvant SBAS2. Pour tous les animaux les injections sont sous-cutanées et ont lieu à J0, J30 et J90. Préparation du mélange injecté : Le mélange antigène/adjuvant est réalisé extemporanément pour chaque injection. En condition stérile on ajoute soit 215 μL de la solution d’antigène pour les vaccinés (150 μL de solution de PfMSP3/210-380 à 0,2 mg/mL + 65 μL de PBS pH 7), soit 215 μL de NaCl pour le lot témoin, à 285 μl de SBAS2. On homogénéise par un bref coup de Vortex puis on laisse incuber 10 minutes sur un plateau agitant en position moyenne. Un ultime coup de Vortex est encore donné juste avant l’administration au primate afin de s’assurer de la parfaite homogénéité de la solution. Préparation des animaux : A chaque immunisation on prélève 20 mL de sang sur tube EDTA. A J0, J30, J90, 1 heure après chaque injection, on cherche des traces de réaction locale au point d’injection (érythème, induration, tuméfaction ou autre) et d’éventuelles limitations de mouvements de l’animal. A J2, J32 et J92 on cherche encore les symptômes locaux (mêmes symptômes mais aussi urticaire, exanthème) et les manifestations générales (fièvre, arthralgie, névralgie, asthme, syncope, nausées, palpitations) pour détecter les réactions retardées. A J0 et J110 un bilan complet hématologique et biochimique est effectué en plus de la mesure pondérale : numération-formule, sodium, potassium, transaminases, bilirubine totale, phosphatases alcalines, gamma glutamyl transférase, créatinine et glycémie.

Exemples d’utilisation des primates africains dans la recherche sur le paludisme
115
A.2.2.3- protocole d’infection d’épreuve par Plasmodium reichenowi et suivi de la parasitémie
Un seul animal est infecté avec les aliquots congelés de P. reichenowi car la souche est devenue très rare et il ne reste que quelques aliquots disponibles (pour cette étude, deux aliquots nous sont prêtés par le Biomedical Primate Research Centre - BPRC - en Hollande). L’infection d’épreuve a lieu à J 110 :
1- anesthésie à la kétamine de l’un des trois chimpanzés contrôle ; 2- pose d’un cathéter veineux et d’une perfusion NaCl 9‰ dans un des avant-bras ; 3- mise à 37°C de 5 mL de RPMI et d’un aliquot de la souche de P. reichenowi pendant
une minute ; 4- aspiration de 1 mL de RPMI chaud que l’on injecte dans le tube de la souche afin de
réchauffer rapidement le culot globulaire ; 5- aspiration de ce mélange que l’on injecte immédiatement dans la tubulure de perfusion
en accélérant le débit de celle-ci pour assurer une diffusion intraveineuse immédiate des globules rouges parasités.
Le suivi de la parasitémie du primate ainsi inoculé est réalisé tous les 2 jours (sous anesthésie générale on prélève 5 mL de sang sur tube EDTA). On effectue à partir de ce prélèvement un frottis sanguin, une goutte épaisse et un QBC. Puis on centrifuge le sang afin de séparer le sérum du culot qui sont stockés à –20°C afin d’exécuter ultérieurement une PCR et un test LDH. Lorsque la parasitémie atteint un niveau suffisant pour pouvoir injecter au moins 100 000 globules rouges parasités à chacun des 5 autres singes, on prélève le sang parasité sur tube EDTA que l’on réinjecte directement par cathéter aux receveurs.
A.2.2.4- protocole d’essai thérapeutique par l’anticorps recombinant RAM-1
Le choix de la posologie et du mode d’administration repose sur deux données : - chez la souris, la clearance de la parasitémie est obtenue avec des doses de 1mg/kg en
injection intrapéritonéale ; - par ailleurs, l’injection intraveineuse d’IgG totales africaines chez l’homme a induit
dans un cas une séquestration splénique des globules rouges parasités avec un accroissement rapide de la taille de la rate. Ce phénomène peut être évité en étalant le traitement sur 2 jours.
Ainsi, une dose totale de 30 mg, soit environ 0,5 mg/kg, est proposée pour les chimpanzés. On utilise la voie intraveineuse lente, en répartissant le traitement en 3 injections, chacune étant suivie de l’observation des signes cliniques et notamment des paramètres spléniques par échographie :
- Jour 1, 8h00 : 3 mg, soit environ un dixième de la dose, par voie intraveineuse lente, - Jour 1,12h00 : 7 mg, un peu plus d’un quart de la dose, par voie intraveineuse lente, - Jour 2, 8h00 : 20 mg, les deux derniers tiers de la dose, sont administrés en perfusion
NaCl 9‰. Ce traitement par anticorps est appliqué aux 3 chimpanzés contrôles, au moment de leur pic de parasitémie.
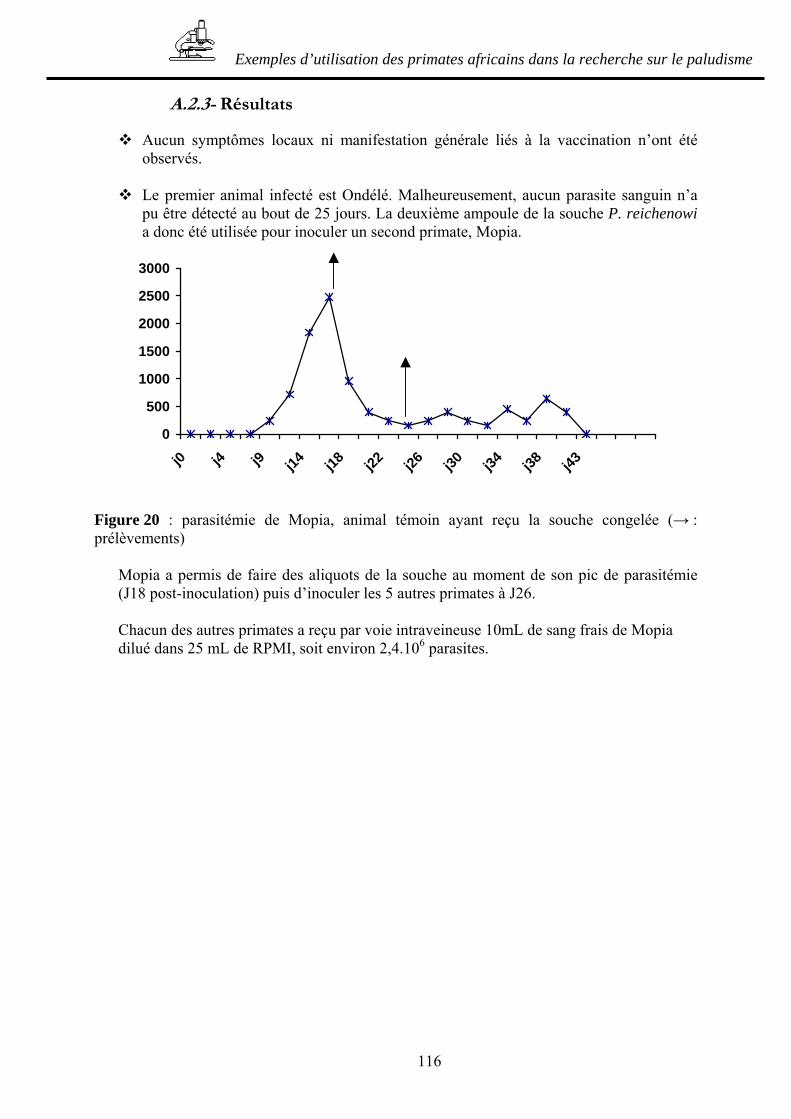
Exemples d’utilisation des primates africains dans la recherche sur le paludisme
116
A.2.3- Résultats
Aucun symptômes locaux ni manifestation générale liés à la vaccination n’ont été observés.
Le premier animal infecté est Ondélé. Malheureusement, aucun parasite sanguin n’a
pu être détecté au bout de 25 jours. La deuxième ampoule de la souche P. reichenowi a donc été utilisée pour inoculer un second primate, Mopia.
Figure 20 : parasitémie de Mopia, animal témoin ayant reçu la souche congelée (→ : prélèvements)
Mopia a permis de faire des aliquots de la souche au moment de son pic de parasitémie (J18 post-inoculation) puis d’inoculer les 5 autres primates à J26.
Chacun des autres primates a reçu par voie intraveineuse 10mL de sang frais de Mopia dilué dans 25 mL de RPMI, soit environ 2,4.106 parasites.
0
500
1000
1500
2000
2500
3000
j0 j4 j9 j14 j18 j22 j26 j30 j34 j38 j43

Exemples d’utilisation des primates africains dans la recherche sur le paludisme
117
0
20000
40000
60000
80000
100000
120000
140000
0 4 8 12 16 19 23
Ondele MopiaMakata
0
20000
40000
60000
80000
100000
120000
140000
0 4 8 12 16 21
CongoNuriaDoumay
Figures 21 et 22 : parasitémies (en nombre de parasites/μL) des animaux vaccinés (à gauche) et du groupe témoin (à droite) en fonction du nombre de jours post-inoculation.
Sur le plan clinique, aucun symptôme n’a pu être observé et les données
hématologiques n’ont pas révélé de modification importante liée à la parasitémie.
D’un point de vue immunologique, aucune réponse n’a été obtenue lors d’ELISA IgG sur les sérums des 6 animaux. Seules des réponses IgM sont détectables.
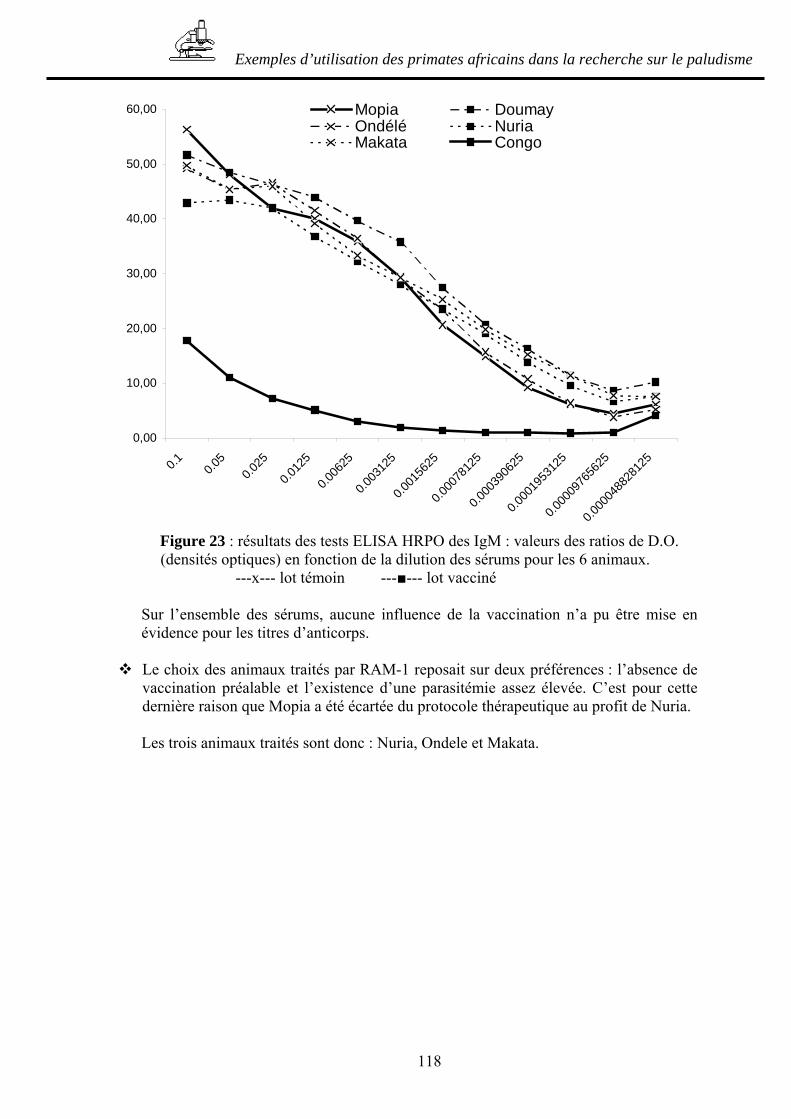
Exemples d’utilisation des primates africains dans la recherche sur le paludisme
118
0,00
10,00
20,00
30,00
40,00
50,00
60,00
0.1 0.05
0.025
0.012
5
0.006
25
0.003
125
0.001
5625
0.000
7812
5
0.000
3906
25
0.000
1953
125
0.000
0976
5625
0.000
0488
2812
5
Mopia DoumayOndélé NuriaMakata Congo
Figure 23 : résultats des tests ELISA HRPO des IgM : valeurs des ratios de D.O. (densités optiques) en fonction de la dilution des sérums pour les 6 animaux.
---x--- lot témoin ---■--- lot vacciné
Sur l’ensemble des sérums, aucune influence de la vaccination n’a pu être mise en évidence pour les titres d’anticorps.
Le choix des animaux traités par RAM-1 reposait sur deux préférences : l’absence de
vaccination préalable et l’existence d’une parasitémie assez élevée. C’est pour cette dernière raison que Mopia a été écartée du protocole thérapeutique au profit de Nuria. Les trois animaux traités sont donc : Nuria, Ondele et Makata.
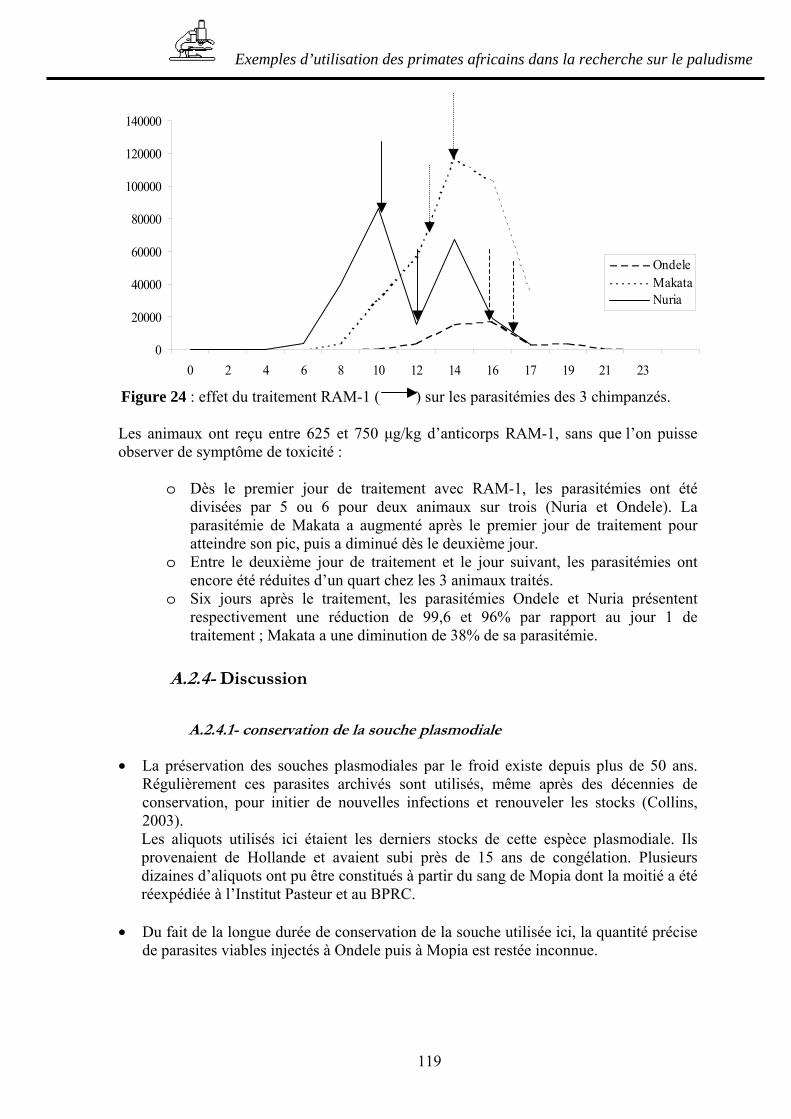
Exemples d’utilisation des primates africains dans la recherche sur le paludisme
119
0
20000
40000
60000
80000
100000
120000
140000
0 2 4 6 8 10 12 14 16 17 19 21 23
Ondele MakataNuria
Figure 24 : effet du traitement RAM-1 ( ) sur les parasitémies des 3 chimpanzés.
Les animaux ont reçu entre 625 et 750 μg/kg d’anticorps RAM-1, sans que l’on puisse observer de symptôme de toxicité :
o Dès le premier jour de traitement avec RAM-1, les parasitémies ont été
divisées par 5 ou 6 pour deux animaux sur trois (Nuria et Ondele). La parasitémie de Makata a augmenté après le premier jour de traitement pour atteindre son pic, puis a diminué dès le deuxième jour.
o Entre le deuxième jour de traitement et le jour suivant, les parasitémies ont encore été réduites d’un quart chez les 3 animaux traités.
o Six jours après le traitement, les parasitémies Ondele et Nuria présentent respectivement une réduction de 99,6 et 96% par rapport au jour 1 de traitement ; Makata a une diminution de 38% de sa parasitémie.
A.2.4- Discussion
A.2.4.1- conservation de la souche plasmodiale
• La préservation des souches plasmodiales par le froid existe depuis plus de 50 ans. Régulièrement ces parasites archivés sont utilisés, même après des décennies de conservation, pour initier de nouvelles infections et renouveler les stocks (Collins, 2003). Les aliquots utilisés ici étaient les derniers stocks de cette espèce plasmodiale. Ils provenaient de Hollande et avaient subi près de 15 ans de congélation. Plusieurs dizaines d’aliquots ont pu être constitués à partir du sang de Mopia dont la moitié a été réexpédiée à l’Institut Pasteur et au BPRC.
• Du fait de la longue durée de conservation de la souche utilisée ici, la quantité précise
de parasites viables injectés à Ondele puis à Mopia est restée inconnue.

Exemples d’utilisation des primates africains dans la recherche sur le paludisme
120
A.2.4.2- suivi des parasitémies
• Comme dans le précédent protocole, il est difficile de dire si l’échec d’infection d’Ondele est dû à la souche ou à l’animal.
• Concernant l’observation du parasite, seules des formes de type trophozoïte ont été
observées (sans doute parce que le protocole a été arrêté avant la phase de production des gamétocytes).
• Les infections ont des durées très variables : un animal (Mopia) avait encore des
parasites à J43, alors qu’un autre (Congo) n’avait plus de parasites à J21 post-infection.
• Si les pics de parasitémies sont proches de ceux obtenus avec P. benjamini (en général
104, parfois 105 parasites/μL), les courbes de parasitémies n’ont pas les mêmes caractéristiques :
o les courbes sont beaucoup moins homogènes, elles présentent de grandes variations individuelles, notamment en ce qui concerne les pics de parasitémies (de moins de 10 000 à plus de 120 000 parasites/μL) ;
o l’apparition des parasites dans le sang est plus précoce (entre J4 et J9) et correspond d’ailleurs aux données évoquées par Coatney pour P. reichenowi. (Coatney et al., 1971);
o si l’on fait abstraction de l’influence du traitement RAM-1, la hauteur du pic n’influence pas la décroissance parasitaire : la parasitémie de Nuria, avec un pic à près de 105 parasites/μL, décroît
assez lentement (7 jours) avec un pic secondaire. La parasitémie de Makata, malgré un pic de même hauteur, décroît en
12 jours. Mise à part Nuria, la parasitémie des animaux décroît toujours
progressivement, sans pic secondaire.
• Comme dans le cas de P. benjamini, le nombre de passage de la souche semble influencer la parasitémie : on observe une avancée de l’apparition des parasites dans le sang (J9 pour Mopia contre J6 en moyenne pour les autres animaux du 2ème passage).
• De façon moins marquée, le délai d’apparition du pic de parasitémie a aussi tendance à
diminuer avec le second passage : à J16 pour Mopia, à J10, J12, J12, J16 et J16 pour les cinq autres animaux.
• Le fait d’avoir déjà participé à des protocoles de paludisme, ou de posséder une
sérologie hépatite positive ne semble pas influencer la parasitémie.
A.2.4.3- vaccination
• La formule vaccinale testée ici est bien tolérée.
• En revanche, cette formule n’est pas immunogène. Seule une augmentation des IgM, anticorps non cytophiles, a été constatée post-infection d’épreuve. Elle est sans doute simplement liée à l’infection. On peut s’interroger sur la raison pour laquelle cette préparation vaccinale est restée sans effet alors qu’une préparation synthétique

Exemples d’utilisation des primates africains dans la recherche sur le paludisme
121
homologue à 70% donne d’excellents résultats d’immunogénicité chez l’humain et d’assez bons résultats chez Aotus et le rongeur (Hisaeda et al., 2002). Un problème de conservation du vaccin lors du transport de France au Gabon a été envisagé.
• La formule vaccinale n’est d’ailleurs pas protectrice (conséquence logique de
l’absence d’immunogénicité).
• Les essais vaccinaux menés chez l’homme à partir de début 2005 permettront sans doute de mieux comprendre l’échec de cette immunisation.
A.2.4.4- traitement par anticorps recombinants
• Le traitement par anticorps recombinants a été bien toléré, la légère et transitoire augmentation de taille de la rate constatée chez certains animaux était certainement due à l’infection paludique.
• Concernant le choix des animaux traités, il y a peut-être un biais à avoir utilisé Nuria
qui est « immunisée » MSP-3.
• Une autre critique qui peut être faite à ce protocole concerne l’absence de réelle situation témoin pour ce traitement. En effet, les 3 animaux non traités par RAM-1 étaient soit vaccinés (Congo et Doumay), soit infectés avec la souche congelée (Mopia). On peut tout de même signaler que la parasitémie de ces 3 animaux non traités chute spontanément de 85 à 92% dans les 6 jours suivant le pic (contre respectivement 99,6 et 96% pour Ondele et Nuria traitées avec RAM-1 mais seulement 38% pour Makata, pourtant lui aussi traité avec RAM-1).
• La réduction de la parasitémie des animaux traités avec RAM-1 est moins rapide que
celle observée chez la souris SCID et dans les études de transfert à l’homme. Ce retard à la décroissance est sans doute dû à un problème de coopération entre les immunoglobulines humaines tels que RAM-1 et les monocytes de chimpanzés, malgré leur proximité avec l’homme. On soupçonne fortement une interaction moléculaire sub-optimale entre les IgG humaines et les récepteurs Fcγ des chimpanzés ; d’une part la coopération monocyte-IgG semble moins forte qu’avec les récepteurs Fcγ des monocytes humains, d’autre part elle est très variable d’un chimpanzé à l’autre (Druilhe 2004, non publié).
• Par ailleurs, l’augmentation des IgM observée a peut-être entraîné une compétition
entre ces anticorps non cytophiles et les quelques IgG1 ajoutés sous forme de RAM-1. En effet, non seulement les IgM ne coopèrent pas avec les monocytes, mais ils peuvent entrer en compétition avec les anticorps cytophiles. Une dose plus importante de RAM-1 aurait alors peut-être permis d’éviter cette compétition. Un mapping épitopique des anticorps permettra de savoir s’ils sont dirigés vers le même épitope que celui ciblé par RAM-1.

Exemples d’utilisation des primates africains dans la recherche sur le paludisme
122
A.2.5- Conclusion
Ainsi, cette formule vaccinale, pourtant prometteuse chez la souris et les singes de nuit, n’a pas permis la protection attendue chez le chimpanzé. Il est donc intéressant ici de voir les limites du modèle chimpanzé, pourtant le plus proche parent de l’humain, lorsque l’on s’intéresse à des études immunologiques poussées. Les essais cliniques menés chez l’homme apporteront sans doute des éléments de réponses quant aux mécanismes exacts de protection de cette formule et quant aux différences homme-chimpanzé.

Exemples d’utilisation des primates africains dans la recherche sur le paludisme
123
B- UTILISATIONS DE PRIMATES AFRICAINS EN TANT QU’HOTES ARTIFICIELS DE PLASMODIUMS HUMAINS
B.1- RECHERCHE D’UN NOUVEAU MODELE POUR LE PALUDISME : ESSAI D’INFECTION DE QUATRE MANDRILLS PAR PLASMODIUM FALCIPARUM
B.1.1- Principe de l’étude
B.1.1.1- problématique
Le CIRMF abrite la plus grande colonie captive de mandrills du monde, avec près de 200 individus. Du fait de son mode de vie forestier et de la faible quantité d’individus captifs, le mandrill est encore peu étudié. Le CIRMF les utilise essentiellement pour des recherches concernant le SIV, puisque cette espèce présente l’intérêt de posséder 2 sous-types de SIVmd et d’être un modèle non pathogène pour le SIDA. Ce primate présente plusieurs avantages intéressants dans le cadre des recherches sur le paludisme ; d’un élevage aisé et de taille réduite par rapport au chimpanzé, son utilisation est bien moins coûteuse (réduction des coûts des structures de captivité, mais aussi d’alimentation et d’anesthésie) et son immunité a déjà été bien étudiée au CIRMF. Enfin, c’est une espèce qui, au niveau comportemental, semble assez bien supporter les conditions d’isolement liées aux protocoles. Cependant, les données concernant les infections palustres des mandrills étant très peu nombreuses, nous avons d’abord mené des études au sein de la colonie du Centre de Primatologie (CDP) du CIRMF.
- En 1995, une étude portant sur tous les primates du CDP n’avait rapporté aucun cas de paludisme sur les primates présents.
- Au cours de l'année 1999, l’analyse moléculaire des prélèvements sanguins de mandrills a montré la présence de P. falciparum chez deux mandrills de la colonie. Ce résultat a été obtenu par la technique PCR de Snounou (Snounou et al., 1993).
- En 2002, un contrôle de parasitologie systématique lors du contrôle sanitaire a été mis en place. Sur 176 mandrills testés, 57 % présentaient des gouttes épaisses positives. Les formes présentes sur les lames ne ressemblaient pas à P. falciparum. Il s’agirait peut-être d’une espèce plasmodiale propre au mandrill.
Par ailleurs, aucun signe clinique se rapportant au paludisme n’a jamais été observé chez les mandrills.

Exemples d’utilisation des primates africains dans la recherche sur le paludisme
124
Il y a alors deux possibilités :
Hypothèse 1 : l’espèce plasmodiale X observée chez les mandrills en 1999 et en 2002 est la même. Elle aurait des formes suffisamment proches de P. falciparum (du P. falciparum qui aurait muté ? Une espèce proche de P. falciparum ?) pour rendre une PCR positive (réaction croisée). Il s’agirait alors une mono infection.
Hypothèse 2 : il y a parfois réellement co-infection (ou en tout cas co-existence) de
P. falciparum et d’une espèce plasmodiale X (nouvelle espèce ou espèce déjà présente chez d’autres primates comme P. gonderi ?).
Dans la première hypothèse, c’est le séquençage de l’ADN extrait des deux mandrills de 1999 puis sa comparaison avec le génome de P. falciparum qui donnera une réponse plus précise. Dans la seconde hypothèse, il faut tenter de situer cette nouvelle espèce par rapport aux autres espèces existantes et se pencher sur la présence conjointe de P. falciparum. Dans quel état pouvait être P. falciparum ? Le fait de ne pas le voir sur les gouttes épaisses signifie t-il que ce parasite était en trop faible concentration dans le sang ? Ou que P. falciparum n’était présent que sous forme fragmentaire ? (mais alors ceci serait en contradiction avec la théorie selon laquelle une PCR positive signifie que le parasite est viable (Jarra et al., 1998)). Si c’est un fragment, d’où provient-il ? Au bout de combien de temps un mandrill peut-il « détruire » P. falciparum ? C’est donc l’analyse du génome des plasmodiums présents dans le sang des deux mandrills de 1999 et de ceux de 2002 qui permettrait d’apporter une orientation. Malheureusement jusque là les prélèvements présentaient trop peu de matériel génétique pour réaliser une telle étude. Cependant, quelle que soit l’hypothèse retenue, il est d’ores et déjà intéressant de se pencher sur la possibilité d’infecter le mandrill par P. falciparum. Le mandrill pourrait en effet être une espèce intermédiaire intéressante : plus proche de l’humain que les singes du Nouveau Monde, moins coûteuse que le chimpanzé et enfin bien étudiée sur le plan immunologique grâce aux travaux déjà réalisés en virologie. Le protocole a reçu l’accord du Comité d’Ethique Ile de France sous la référence n°03-004.
B.1.1.2- étude in vitro : culture sur sang de mandrill
Un prélèvement de 14 mL de sang sur tube EDTA a été effectué sur un mandrill indemne de paludisme, isolé en bâtiment depuis plusieurs années et SIV/STLV négatif. La souche 3D7 a été choisie pour des raisons pratiques (disponibilité, connaissance de la souche et culture aisée). La tentative d’infection des hématies de mandrill par cette souche de P. falciparum a échoué. Cet échec peut signifier :
- que P. falciparum n’est pas adaptable chez le mandrill, - que cette souche n’est pas adaptable chez le mandrill, - et/ou que cette culture est trop délicate.
Exemples d’utilisation des primates africains dans la recherche sur le paludisme
125
Une infection d’animaux est donc tout de même nécessaire pour définir la permissivité du mandrill vis-à-vis de ce plasmodium humain, ceci en parallèle de la caractérisation du parasite trouvé chez le mandrill.
B.1.2- Matériel et méthode
B.1.2.1- présentation des animaux
Quatre jeunes mandrills (Mandrillus sphinx) adultes, 2 femelles et 2 mâles, en bonne santé, d’âge et de poids sensiblement similaires, sont répartis en deux groupe de deux individus. Les quatre animaux sont testés négatifs pour SIVmd et STLV et leur sérologie anti-EBA-175 (antigène de mérozoïte) est négative.
Animal 17D3 17A6 17B6 6F Sexe Mâle Mâle Femelle Femelle Âge 5 ans 5 ans 5 ans 4 ans
Poids en kg (début-fin de protocole) 12,4-12 12,2-10 8,2-7,8 10,4-8,2
Antécédents pathologiques néant néant néant néant
Tableau 11 : présentation des animaux
B.1.2.2- protocole d’infection d’épreuve par Plasmodium falciparum et suivi de la parasitémie
Dates de protocole : octobre - novembre 2003
- J-14 avant inoculation : les primates sont placés dans des bâtiments de type A3, fermés non accessibles aux moustiques. Un QBC, une goutte épaisse et une PCR sont réalisés à J-14 et les animaux sont traités à l’association Pyriméthamine-Sulfadoxine (Fansidar®, 1 ampoule par voie intramusculaire pour le 17D3, ½ ampoule pour les 3 autres).
- J-7 : QBC, goutte épaisse et PCR sont réalisés pour vérification. - J0 : infection avec 600 μL d’hématies humaines parasitées à 20% par P. falciparum
souche 3D7, ce qui représente près de 600 000 parasites injectés par voie intraveineuse.
Pour le suivi de parasitémie, de J0 à J50, 3 mL EDTA sont prélevés sous anesthésie générale tous les 3 jours pour effectuer un QBC, un frottis et une goutte épaisse.
Un prélèvement hebdomadaire est effectué pour contrôle des paramètres hématologiques et biochimiques (urée, créatinine, transaminases, phosphatases alcalines, glucose, lactate).
B.1.3- Résultats
Aucun parasite sanguin n’a pu être décelé chez aucun des animaux.

Exemples d’utilisation des primates africains dans la recherche sur le paludisme
126
B.1.4- Discussion et conclusion
Plusieurs hypothèses peuvent être émises :
- le mandrill n’est pas permissif à l’infection par P. falciparum ; - le mandrill n’est pas permissif à l’infection par les stades érythrocytaires humains de
P. falciparum (il est tout de même surprenant de ne pas avoir vu un seul parasite) ; - la souche 3D7 n’est pas adaptée à l’infection chez le mandrill ; - une splénectomie est nécessaire.
Par conséquent, à moins que ce ne soit qu’un problème de souche, le mandrill est sans doute un mauvais modèle d’infection par P. falciparum. Par ailleurs, d’un point de vue écologique, le mandrill ne semble pas menacé par le paludisme humain.

Exemples d’utilisation des primates africains dans la recherche sur le paludisme
127
B.2- ESSAI D’UN VACCIN PRE-ERYTHROCYTAIRE AVEC DES CHIMPANZES INFECTES PAR PLASMODIUM FALCIPARUM
L’étude qui suit concerne une formule vaccinale en cours de développement ; ainsi, pour des raisons de confidentialité, une partie des données ne pourra pas être présentée (type de vaccin, formule, etc.).
B.2.1- Principe de l’étude
Le protocole dure 10 mois : après avoir immunisé les animaux, l’efficacité et la rémanence de la formule vaccinale sont testées avec une double infection par des sporozoïtes à 5 mois d’intervalle. Le choix du modèle animal s’est naturellement porté vers le chimpanzé pour deux raisons :
- il est l’un des seuls modèles réceptifs aux stades pré-érythrocytaires de P. falciparum ; - il constitue le modèle le plus proche de l’homme au stade pré-clinique du
développement vaccinal. Par ailleurs, il rend possible l’exploration détaillée des mécanismes immunitaires en employant des réactifs à usage humain.
Le protocole a reçu l’accord du Comité d’Ethique Ile de France sous la référence n°02-002.
B.2.2- Matériel et méthode
B.2.2.1- présentation des animaux
Du fait de l’utilisation de chimpanzé, un compromis entre les besoins statistiques et les contraintes financières et éthiques doit être envisagé. Ainsi, ce sont six jeunes adultes chimpanzés (Pan troglodytes), 2 mâles et 4 femelles en bonne santé, qui sont répartis en deux groupes :
- groupe témoin : Chico et Géraldine - groupe vacciné : Mayumba, Mickey, Belle et Doris.
Animal Chico Geraldine Mayumba Mickey Belle Doris
Sexe Mâle Femelle Femelle Mâle Femelle Femelle Âge 10 ans 10 ans 9 ans 10 ans 7 ans 9 ans
Poids en kg (début-fin de
protocole) 40-45,6 35-35,3 30-34,8 32-31,5 33-31,9 25-29
Antécédents pathologiques néant néant néant néant néant Hépatite B
Ac+ Ag-
Tableau 12 : présentation des animaux

Exemples d’utilisation des primates africains dans la recherche sur le paludisme
128
B.2.2.2- protocole d’immunisation
Un mois avant la première immunisation, les animaux sont placés en bâtiments expérimentaux de type A2 fermés inaccessibles aux moustiques ; une recherche de plasmodium est alors effectuée (QBC, goutte épaisse et frottis). A chaque jour d’immunisation, plusieurs prélèvements sont effectués avant la vaccination :
- 50 mL de sang sur tube EDTA pour l’analyse détaillée des réponses immunitaires préexistantes chez les chimpanzés (analyse en cytométrie de flux, tests ELISA et ELISpot) :
o anticorps de l’antigène vaccinal, o anticorps contre les stades érythrocytaires, o anticorps contre l’ensemble des antigènes de surface des sporozoïtes de
P. falciparum (souche NF54), o titres d’anticorps et tests de proliférations cellulaires avec les peptides
vaccinaux - 5 mL sur tube EDTA et 5 mL sur tube sec pour une numération-formule, une
biochimie classique et une recherche parasitaire.
Le groupe vacciné reçoit 3 doses vaccinales (composition confidentielle) à 1 mois d’intervalle. L’injection est faite dans le mollet, par voie intramusculaire. Des prélèvements (30 à 50 mL EDTA) sont effectués à J60, J75 et J105 après la dernière immunisation pour un suivi des paramètres immunologiques.
B.2.2.3- protocole d’infection d’épreuve par P. falciparum et suivi de la parasitémie
Des sporozoïtes sont obtenus par dissection et broyage des glandes salivaires de moustiques reçus du Biomedical Primate Research Centre (BPRC) en Hollande.
Chaque infection d’épreuve consiste en une infection à faible dose : 20 000 sporozoïtes (dans 0,7 mL de RPMI) sont injectés par voie intraveineuse à chaque animal. Deux infections sont réalisées, la première à J98, la seconde à J238 suivant le dernier rappel vaccinal. A chaque infection d’épreuve :
Du 6ème au 10ème jour post-infection, le suivi parasitaire est effectué (5 mL sur tube EDTA) par QBC, frottis et goutte épaisse, mais aussi PCR et test LDH. En parallèle, le plasma est prélevé pour la détection des antigènes solubles circulants. Du 10ème au 30ème jour post-infection, trois prélèvements hebdomadaires sont réalisés
pour QBC, frottis et goutte épaisse. A la fin du protocole, tous les animaux sont traités avec l’association Pyriméthamine-Sulfadoxine (Maloxine®, 2 comprimés en prise unique) avant d’être replacés dans leurs volières d’origine.
B.2.3- Résultats
Parasitémies
Trois animaux n’ont pas développé de parasitémie suite aux deux infections d’épreuve.

Exemples d’utilisation des primates africains dans la recherche sur le paludisme
129
�����������
������������
� � ��Belle
Jours
Par
asite
s/µl
�������
�����������
������������
� � ��Belle
Jours
Par
asite
s/µl
�� � �������� ���� ��������
5 6 7 8 9 10 11 12
0
40
80
120
5 6 7 8 9 10 11 12
0
20
40
60
80
238 jours post -immunisation98 jours post -immunisation
infection d’épreuve par une souche hétérologue
�������
Figures 25 et 26 : parasitémies des 4 animaux vaccinés (au premier plan) et des 2 témoins (en arrière-plan en italique) en fonction des jours post-inoculation des deux infections d’épreuve.
� Clinique : aucun symptôme n’a été observé à la suite des vaccinations ni à la suite des challenges.
� Résultats de la réponse lymphocytaire L’analyse par cytométrie en flux n’a pas décelé chez le lot vacciné d’augmentation significative du taux de lymphocytes T CD4+ ni CD8+ par rapport au lot contrôle. � Résultats de la réponse spécifique
o La réponse Antigène-spécifique mesurée par technique ELISA n’a donné que des réponses faibles en anticorps chez les individus vaccinés.
o ELISpot :
Des tests ELISpot sont réalisés juste avant la premiere infection d’épreuve, entre les deux infections et juste avant la deuxième infection ; ce type de test permet de mesurer le nombre de lymphocytes T (SFC : Spot Forming Cell) qui produisent de l’IFNγ après mise en contact avec un antigène donné. Contrairement à la cytométrie en flux qui quantifie le nombre de lymphocytes, cette technique contribue plutôt à l’évaluation de l’activation de ces lymphocytes par la formule vaccinale. L’IFN� peut être produit par trois populations de cellules dans le sang :
• les lymphocytes T8 (CMH I+), • les lymphocytes T4 (CMH II+), • les cellules NK (Natural Killer) (CMH I+), très minoritaires.

Exemples d’utilisation des primates africains dans la recherche sur le paludisme
130
La réponse est mesurée sur l’ensemble des populations (colonne de gauche des figures 27 à 36) ou sur les populations CMH II+ (colonne de droite des figures 27 à 36, en présence d’un anticorps anti-MHC class I restricted responses, clone W6-32). Les tests ont tous été réalisés en duplicats.
Antigènes choisis :
• Rec 1, 2 et 3 sont des protéines recombinantes qui couvrent
toute la formule vaccinale : o Rec 1 est l’extrémité N-terminale o Rec 2 est la partie centrale (zone de répétition) o Rec 3 est l’extrémité C-terminale
• Pep 1 est un peptide synthétique mimant la formule vaccinale, qui correspond à la région de Rec 3 ;
• GST (Glutathion S Transférase) est la protéine de fusion des recombinants 1, 2 et 3 utilisés, elle ne devrait pas être très immunogène ;
• HIV pep- est un peptide de la région Nef du VIH, c’est un témoin négatif car ces animaux sont séronégatifs pour le VIH-1 ;
• Med (medium) correspond au milieu seul, c’est le témoin négatif qui correspond au bruit de fond ;
• PPD (Purified Protein Derivates) est la tuberculine. En cas de contact des animaux avec des mycobactéries, la réponse à cette protéine est forte et persistante.
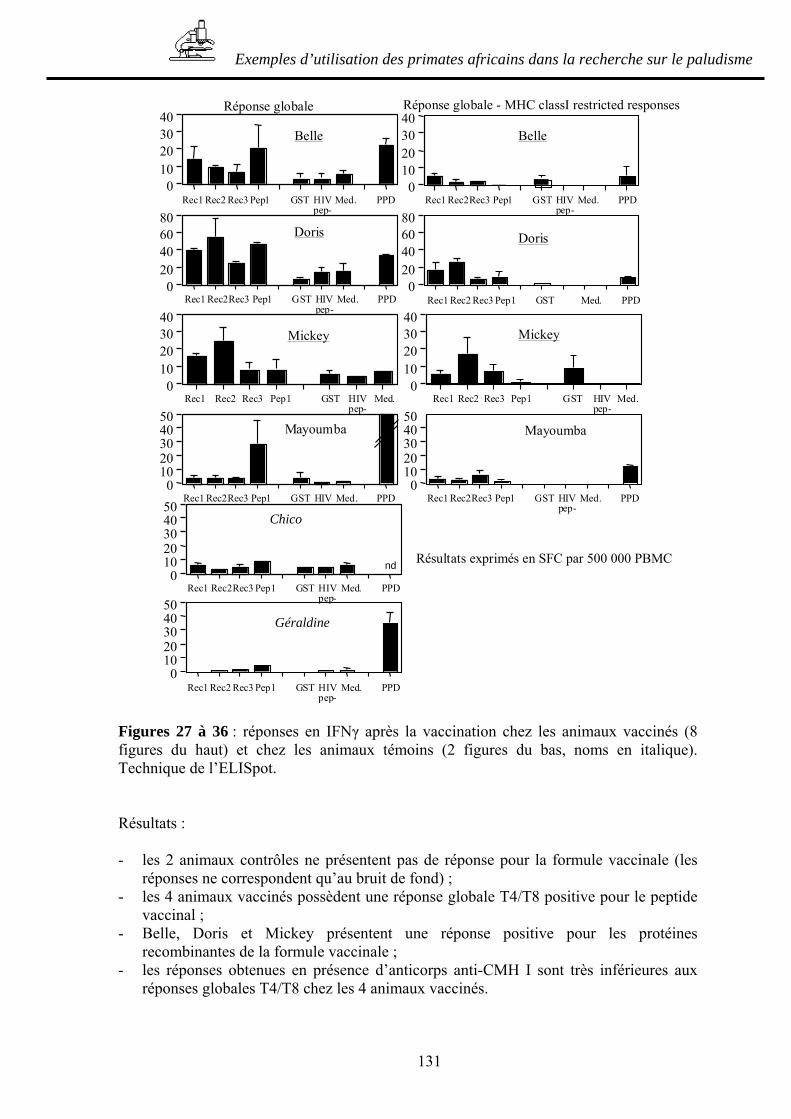
Exemples d’utilisation des primates africains dans la recherche sur le paludisme
131
010203040
Rec1 Rec2 Rec3 Pep1 GST HIVpep-
Med. PPD0
10203040
020406080
020406080
010203040
010203040
01020304050
01020304050
Mayoumba
Rec1 Rec2Rec3 Pep1 GST HIVpep-
Med. PPD
Rec1 Rec2 Rec3 Pep1 GST HIVpep-
Med.
Rec1 Rec2Rec3 Pep1 GST HIVpep-
Med. PPD Rec1 Rec2Rec3 Pep1 GST HIVpep-
Med. PPD
Rec1 Rec2 Rec3 Pep1 GST HIVpep-
Med.
Rec1 Rec2 Rec3 Pep1 GST Med. PPD
Rec1 Rec2Rec3 Pep1 GST Med. PPDHIVpep-
01020304050
Chico
01020304050
Rec1 Rec2Rec3 Pep1 GST HIVpep-
Med. PPD
Rec1 Rec2 Rec3 Pep1 GST HIVpep-
Med. PPD
nd
Réponse globale - MHC classI restricted responses
Résultats exprimés en SFC par 500 000 PBMC
Belle
Réponse globale
Géraldine
Mickey
Doris
Belle
Doris
Mickey
Mayoumba
Figures 27 à 36 : réponses en IFNγ après la vaccination chez les animaux vaccinés (8 figures du haut) et chez les animaux témoins (2 figures du bas, noms en italique). Technique de l’ELISpot. Résultats : - les 2 animaux contrôles ne présentent pas de réponse pour la formule vaccinale (les
réponses ne correspondent qu’au bruit de fond) ; - les 4 animaux vaccinés possèdent une réponse globale T4/T8 positive pour le peptide
vaccinal ; - Belle, Doris et Mickey présentent une réponse positive pour les protéines
recombinantes de la formule vaccinale ; - les réponses obtenues en présence d’anticorps anti-CMH I sont très inférieures aux
réponses globales T4/T8 chez les 4 animaux vaccinés.

Exemples d’utilisation des primates africains dans la recherche sur le paludisme
132
B.2.4- Discussion
B.2.4.1- inoculation d’épreuve
• La dose infectante de 20 000 sporozoïtes est qualifiée de faible par comparaison aux doses parfois utilisées dans d’autres protocoles.
• Malheureusement, le voyage infligé aux anophèles depuis les Pays-Bas aboutissait à
une forte mortalité, et limitait ainsi le nombre de sporozoïtes disponibles. • Cependant, par rapport à la dose naturellement reçue en cas de piqure d’anophèle
(quelques dizaines voire quelques centaines de sporozoïtes), cette « faible » infection d’épreuve reste 100 fois suffisante pour tester l’efficacité d’un vaccin.
B.2.4.2- suivi des parasitémies
• Les courbes de parasitémies obtenues chez les 3 chimpanzés infectés indiquent une détection du parasite dans le sang vers J5-J6 post infection, suivie d’un pic de parasitémie entre J8 et J10. Ces données sont tout à fait comparables à celles décrites chez l’homme dans le cadre d’une infection à P. falciparum.
• En revanche, on observe chez ces mêmes animaux une élimination du parasite dans un
délai de 12 à 15 jours suivant l’infection, ce qui n’est en général pas retrouvé chez l’homme. Le chimpanzé, s’il est le seul modèle sensible à l’infection sporozoïtaire à P. falciparum, présente la particularité de contrôler sa parasitémie rapidement.
• Les parasitémies des trois animaux sont assez faibles et similaires à celles observées
chez les humains. • La reproductibilité des parasitémies au cours des deux infections d’épreuve pour un
même animal ainsi que la similarité des courbes parasitaires de l’ensemble des animaux démontre encore une fois l’intérêt du modèle chimpanzé pour ce type d’étude, même lors d’infections d’épreuve à faibles doses ; l’Aotus, également utilisé pour les études pré-cliniques de vaccins pré-érythrocytaires présente des parasitémies bien moins reproductibles (Perlaza et al., 2003).
B.2.4.3- effet protecteur de la vaccination
• Mayoumba, vaccinée non protégée, développe une parasitémie presque deux fois inférieure à celle des animaux témoins. Cependant, dans la mesure où ces parasitémies sont faibles, où la parasitémie peut être extrêmement variable chez un même humain et enfin où le nombre d’animaux est restreint, il est impossible de conclure à une protection partielle chez cet individu.
• La protection offerte par ce vaccin est totale pour 3 des 4 animaux vaccinés, alors que
les 2 chimpanzés témoins développent immanquablement une infection, ce qui fait de cette formule un vaccin très prometteur. Certes le nombre d’animaux utilisés pour cette étude est trop faible pour une conclusion statistique significative ; cependant, la remarquable reproductibilité des résultats liée à l’utilisation du chimpanzé permet d’en tirer de fortes présomptions quant à son efficacité vaccinale.

Exemples d’utilisation des primates africains dans la recherche sur le paludisme
133
• Trois injections à un mois d’intervalle permettent une protection qui dure près d’un an.
Pour des raisons éthiques (captivité de chimpanzés en bâtiments classés A2 sans accès au milieu extérieur), l’étude de la protection à plus longue échéance n’a pu être réalisée. Les études cliniques sur les cohortes humaines apporteront des réponses plus précises quant à la durée de cette protection.
• ELISpot :
o La réponse obtenue chez les animaux vaccinés est très similaire à celle obtenue
chez l’homme après le même type d’immunisation, soulignant ainsi l’intérêt du chimpanzé comme modèle d’immunogénicité vaccinale, contrairement aux primates sud-américains dont les réponses lymphocytaires sont difficiles à cerner (Collins et al., 1989a; Perlaza et al., 2003).
o Réponse pour la tuberculine :
les 6 animaux utilisés ici n’ont jamais été vaccinés avec le BCG, ce n’est donc pas un témoin positif de la réponse ;
en revanche, ils peuvent avoir rencontré des mycobactéries, par l’intermédiaire de leurs animaliers par exemple (un cas de tuberculose chronique a été relevé chez l’un d’eux), ce qui pourrait être le cas de Mayoumba qui présente 138 SFC/500 000 PBMC ;
la réponse PPD perdure en présence d’anticorps anti-CMH I, ce qui semble logique, dans la mesure où la réponse à la tuberculine est de type mixte T4/T8 ;
par ailleurs il est connu que les tests tuberculiniques peuvent croiser avec d’autres antigènes, dont ceux de parasites intestinaux, ce qui pourrait expliquer les réponses des 5 autres individus.
o Les 3 animaux protégés par le vaccin présentent des réponses globales fortes
pour les protéines recombinantes, contrairement à Mayoumba, animal vacciné, non protégé.
o Diminution de la réponse en présence de l’anticorps anti-CMH I W6-32 :
on peut penser de prime abord que la réponse est principalement induite par des cellules CMH II+.
Cependant, dans la mesure où l’anticorps utilisé est d’usage humain, il est difficile de prédire son impact sur la réponse T4 de chimpanzé qu’il pourrait également diminuer (d’autant que les tests sont réalisés sur PBMC cryoconservés). Un nouveau protocole est prévu, en présence d’interleukines IL-7 et IL-15, qui augmentent les réponses T, pour mieux cerner les réponses de faibles amplitudes (Jennes et al., 2002).
L’efficacité de la déplétion en CMH I par le W6-32 semble confirmée par :
l’effondrement de la réponse au Pep1 chez les 4 animaux vaccinés ; en effet, les peptides étant toujours mieux processés in vivo par les lymphocytes T CD8+ CMH I+ que par les CD4+ CMH II+, la réponse des cellules T déplétées en CMH I correspond à la faible réponse des T CD4+ CMH II+.

Exemples d’utilisation des primates africains dans la recherche sur le paludisme
134
Une relative persistance de la réponse aux Rec, liée au fait que les lymphocytes T CD4+ CMH II+ répondent plutôt mieux aux protéines recombinantes ou aux lysats que les T CD8+ CMH I+.
Pour évaluer vraiment l’efficacité de l’anticorps anti-CMH I, une solution consisterait à passer les populations cellulaires en cytométrie de flux avant et après l’usage du W6-32 pour évaluer la déplétion des sous populations lymphocytaires.
o On peut s’interroger sur la différence de réponse globale (sans anticorps anti-
CMH I) des 3 femelles vaccinées vis-à-vis du Pep1 et de Rec3 qui correspondent pourtant à la même région vaccinale : Belle et Mayoumba présentent un écart-type important pour leur
réponse à Pep1, ce qui, dans le cas de Belle, rend cette réponse finalement comparable à celle pour Rec3.
Pour Mayoumba et Doris, une explication réside peut-être dans le fait que leur réponse globale au vaccin serait surtout CMH I dépendante, donc avec une meilleure réponse pour le peptide que pour les protéines recombinantes.
o Corrélation protection-immunité pour cette formule vaccinale :
La protection n’est pas associée à une augmentation du nombre de lymphocytes puisqu’aucune différence n’est observée entre les animaux vaccinés protégés et les autres.
Pour les mêmes raisons, la protection engendrée par cette formule ne passe pas par une réponse humorale, conformément à ce qui est observé pour ce type de vaccin (Belnoue et al., 2004).
Les profils de réponses après déplétion en CMH I sont les mêmes chez Belle (vaccinée protégée) et chez Mayoumba (vaccinée non protégée). La protection ne semble donc donc pas ici associée à la réponse CMH II.
Mayoumba ne présente pas de réponse aux Rec, à l’inverse des 3 individus vaccinés protégés. La protection serait donc associée à une réponse des cellules CMH I+ pour les protéines recombinantes (la partie la plus immunogène étant vraisemblablement Rec2, élément central de la protéine vaccinale) comme déjà constaté pour un autre vaccin pré-érythrocytaire (Sedegah et al., 2003).
o Enfin, si une bonne réponse en IFNγ est souvent associée à la protection contre
le paludisme, les discussions autour de ces réponses ELISpot démontrent encore une fois qu’elle ne suffit pas forcément (Wang et al., 2005). Il existerait d’autres marqueurs de protection contre l’infection, qui restent à définir pour améliorer les études vaccinales (Pinder et al., 2004).
B.2.5- Conclusion
Il s’agit donc ici d’une formule vaccinale prometteuse par son efficacité, dont d’autres formulations dérivées du même antigène ont également fait leurs preuves sur des chimpanzés (Druilhe, non publié).

Exemples d’utilisation des primates africains dans la recherche sur le paludisme
135
Cette formulation nécessitera peut-être quelques modifications pour améliorer la réponse Th1/Th2 et anticorps-spécifique (Carcaboso et al., 2004). En outre, si le mécanisme de protection n’est pas encore totalement élucidé, l’immunité du paludisme ne l’est pas non plus et l’utilisation du modèle chimpanzé a permis d’ouvrir de nombreuses pistes à vérifier en phase I sur des cohortes plus importantes.


137
Conclusion
Un petit unicellulaire du nom de Plasmodium continue de déjouer les stratégies les plus sophistiquées élaborées par l’homme. Son fléau est remarquable, d’une part par la complexité de la maladie qu’il engendre et d’autre part parce que la lutte doit intégrer les problèmes scientifiques aux données financières. Dans le domaine de la recherche, les scientifiques se heurtent à l’absence de modèle fiable pour cette maladie. Parmi tous les modèles existants en recherche sur le paludisme les techniques in vitro, les rongeurs et les primates non-humains sont les plus couramment utilisés. Ils ont déjà permis de grandes avancées mais ils ne constituent pas toujours des modèles assez prédictibles pour de solides extrapolations à l’homme. Parmi eux, ce sont bien sûr, comme dans les autres domaines scientifiques, les primates qui permettent de s’approcher le plus des données humaines. Cependant, au sein même de ce groupe, la prédictibilité est variable. En outre, d’autres facteurs rendent parfois leur utilisation délicate.
Ainsi, les primates du Nouveau Monde, sont actuellement les plus utilisés dans la recherche sur le paludisme ; ils sont certes sensibles aux plasmodiums humains, mais sont aussi les primates les plus éloignés de l’homme et leur disponibilité n’est pas toujours assurée.
Parmi les primates de l’Ancien Monde, les macaques, singes asiatiques, constituent un
modèle intéressant. Ils sont sensibles à un grand nombre de plasmodiums, leur coût est abordable et ils sont facilement disponibles. Cependant, leur biologie, et notamment leur immunité, est encore assez éloignée de l’homme.
Parmi les primates africains, les grands singes sont bien sûr les plus proches parents de
l’humain et représentent ainsi les meilleurs modèles en raison de leur prédictibilité. Mais leur coût d’utilisation et leur statut d’espèce menacée les rend souvent difficiles à utiliser à grande échelle. Leur utilisation en laboratoire doit donc se limiter à des études bien précises (vaccination pré-érythrocytaire par exemple).
Les autres primates africains comme le mandrill, les mangabeys, les singes verts ou les
babouins, encore très peu utilisés, pourraient alors constituer un bon intermédiaire : leur divergence avec l’humain est assez récente pour avoir conservé une biologie proche, leur disponibilité est assez grande et leur coût d’utilisation n’est pas beaucoup plus élevé que celui des macaques.
Enfin, comme les primates africains sont très peu utilisés dans le cadre du paludisme, il reste de nombreuses études à mener. En premier lieu, la sensibilité des humains et des primates non-humains aux différents plasmodiums des primates doit être mieux connue. Ces études de sensibilités croisées devraient aussi bien être menées en laboratoire grâce à des infections provoquées, qu’auprès des populations sauvages avec leurs infections naturelles. En effet, mieux connaître les plasmodiums naturels des primates africains doit permettre, en retraçant la co-évolution des plasmodiums et de leurs hôtes de mieux cerner le paludisme humain.

138
Quant à la connaissance des sensibilités de chaque primate, elle pourrait conduire d’une part à la découverte de nouveaux modèles d’infections (pathogènes ou non) et d’autre part à la meilleure définition du rôle de chacun de ces hôtes dans la composante zoonotique du paludisme.

139
BIBLIOGRAPHIE ABDEL-AZIZ I. Z., OSTER N., STICH A., COULIBALY B., GUIGEMDE W. A., WICKERT H., ANDREWS K. T., KOUYATE B., LANZER M. (2005) Association of Plasmodium falciparum isolates encoding the P. falciparum chloroquine resistance transporter gene K76T polymorphism with anemia and splenomegaly, but not with multiple infections. Am J Trop Med Hyg 72, 3, 252-255
ADJUIK M., AGNAMEY P., BABIKER A., BORRMANN S., BRASSEUR P., CISSE M., COBELENS F., DIALLO S., FAUCHER J. F., GARNER P., GIKUNDA S., KREMSNER P. G., KRISHNA S., LELL B., LOOLPAPIT M., MATSIEGUI P. B., MISSINOU M. A., MWANZA J., NTOUMI F., OLLIARO P., OSIMBO P., REZBACH P., SOME E., TAYLOR W. R. (2002) Amodiaquine-artesunate versus amodiaquine for uncomplicated Plasmodium falciparum malaria in African children: a randomised, multicentre trial. Lancet 359, 9315, 1365-1372
ADJUIK M., BABIKER A., GARNER P., OLLIARO P., TAYLOR W., WHITE N. (2004) Artesunate combinations for treatment of malaria: meta-analysis. Lancet 363, 9402, 9-17
AHMED A., BARARIA D., VINAYAK S., YAMEEN M., BISWAS S., DEV V., KUMAR A., ANSARI M. A., SHARMA Y. D. (2004) Plasmodium falciparum isolates in India exhibit a progressive increase in mutations associated with sulfadoxine-pyrimethamine resistance. Antimicrob Agents Chemother 48, 3, 879-889
AIKAWA M., SEED T. M. (1980) Morphology of Plasmodia in: Kreier J. P. Malaria. Academic Press, New York, 285-345
ALLAN R. (2001) Parasites and guns : waging awar on malaria. Africa Health (Medicine Digest) 24, 12-13
ALLAN R., DOULL L., VAN DER WIJK J. (2001) Cost recovery and malaria treatment in emergencies. The Health Exchange december 2001, 15-19
ALLEN S. J., O'DONNELL A., ALEXANDER N. D., ALPERS M. P., PETO T. E., CLEGG J. B., WEATHERALL D. J. (1997) alpha+-Thalassemia protects children against disease caused by other infections as well as malaria. Proc Natl Acad Sci U S A 94, 26, 14736-14741
ALLOUECHE A., MILLIGAN P., CONWAY D. J., PINDER M., BOJANG K., DOHERTY T., TORNIEPORTH N., COHEN J., GREENWOOD B. M. (2003) Protective efficacy of the RTS,S/AS02 Plasmodium falciparum malaria vaccine is not strain specific. Am J Trop Med Hyg 68, 1, 97-101

140
ANDREWS L., ANDERSEN R. F., WEBSTER D., DUNACHIE S., WALTHER R. M., BEJON P., HUNT-COOKE A., BERGSON G., SANDERSON F., HILL A. V., GILBERT S. C. (2005) Quantitative real-time polymerase chain reaction for malaria diagnosis and its use in malaria vaccine clinical trials. Am J Trop Med Hyg 73, 1, 191-198
APPAWU M., OWUSU-AGYEI S., DADZIE S., ASOALA V., ANTO F., KORAM K., ROGERS W., NKRUMAH F., HOFFMAN S. L., FRYAUFF D. J. (2004) Malaria transmission dynamics at a site in northern Ghana proposed for testing malaria vaccines. Trop Med Int Health 9, 1, 164-170
AREVALO-HERRERA M., HERRERA S. (2001) Plasmodium vivax malaria vaccine development. Mol Immunol 38, 6, 443-455
ARTEB (2002). (Page consultée le 17 mai 2004). Transgenese. [en ligne] Adresse: http://www.arteb.com/infos/Technoclef/transgenese.pdf
AUCAN C., WALLEY A. J., HENNIG B. J., FITNESS J., FRODSHAM A., ZHANG L., KWIATKOWSKI D., HILL A. V. (2003) Interferon-alpha receptor-1 (IFNAR1) variants are associated with protection against cerebral malaria in the Gambia. Genes Immun 4, 4, 275-282
AYALA F. J., ESCALANTE A. A., RICH S. M. (1999) Evolution of Plasmodium and the recent origin of the world populations of Plasmodium falciparum. Parassitologia 41, 1-3, 55-68
BALLOU W. R., KESTER K. E., STOUTE J. A., HEPPNER D. G. (1999) Malaria vaccines: triumphs or tribulations? Parassitologia 41, 1-3, 403-408
BASKIN G. B. (1999). (Page consultée le 21 janvier 2004). Patholoy of nonhuman primates. [en ligne] Adresse: http://www.afip.org/CLDavis/GrossCourse99/primates.htm
BELNOUE E., COSTA F. T., FRANKENBERG T., VIGARIO A. M., VOZA T., LEROY N., RODRIGUES M. M., LANDAU I., SNOUNOU G., RENIA L. (2004) Protective T cell immunity against malaria liver stage after vaccination with live sporozoites under chloroquine treatment. J Immunol 172, 4, 2487-2495
BENMOHAMED L., THOMAS A., DRUILHE P. (2004) Long-term multiepitopic cytotoxic-T-lymphocyte responses induced in chimpanzees by combinations of Plasmodium falciparum liver-stage peptides and lipopeptides. Infect Immun 72, 8, 4376-4384
BENOIT-VICAL F., VALENTIN A., DA B., DAKUYO Z., DESCAMPS L., MALLIE M. (2003) N'Dribala (Cochlospermum planchonii) versus chloroquine for treatment of uncomplicated Plasmodium falciparum malaria. J Ethnopharmacol 89, 1, 111-114
BIOMEDICAL PRIMATE RESEARCH CENTER (2005). (Page consultée le 15 juillet 2005). Relocation of Chimpanzees. [en ligne] Adresse: http://www.bprc.nl/BPRCE/L3/RelocChimps.html

141
BOJANG K. A., OLODUDE F., PINDER M., OFORI-ANYINAM O., VIGNERON L., FITZPATRICK S., NJIE F., KASSANGA A., LEACH A., MILMAN J., RABINOVICH R., MCADAM K. P., KESTER K. E., HEPPNER D. G., COHEN J. D., TORNIEPORTH N., MILLIGAN P. J. (2005) Safety and immunogenicty of RTS,S/AS02A candidate malaria vaccine in Gambian children. Vaccine 23, 32, 4148-4157
BOUCHAUD O., COT M., KONY S., DURAND R., SCHIEMANN R., RALAIMAZAVA P., COULAUD J. P., LE BRAS J., DELORON P. (2005) Do African immigrants living in France have long-term malarial immunity? Am J Trop Med Hyg 72, 1, 21-25
BOUGUERRA M. L. (1998) Insuffisamment rentable pour l'industrie pharmaceutique. le monde diplomatique juin 1998, 23
BRABIN B., PRINSEN-GEERLIGS P., VERHOEFF F., KAZEMBE P. (2003) Anaemia prevention for reduction of mortality in mothers and children. Trans R Soc Trop Med Hyg 97, 1, 36-38
BRABIN B. J., ROMAGOSA C., ABDELGALIL S., MENENDEZ C., VERHOEFF F. H., MCGREADY R., FLETCHER K. A., OWENS S., D'ALESSANDRO U., NOSTEN F., FISCHER P. R., ORDI J. (2004) The sick placenta-the role of malaria. Placenta 25, 5, 359-378
BRAY R. S. (1957) Studies on malaria in chimpanzees. IV. Plasmodium ovale. Am J Trop Med Hyg 6, 4, 638-645
BRAY R. S. (1958a) Studies on malaria in chimpanzees. V. The sporogonous cycle and mosquito transmission of Plasmodium vivax schwetzi. J Parasitol 44, 1, 46-51
BRAY R. S. (1958b) Studies on malaria in chimpanzees. VI. Laverania Falciparum. Am J Trop Med Hyg 7, 20-24
BRUMPT E. (1939) Les parasites du paludisme des chimpanzés. In: C. R. Séances Soc. Biol. Ses Fil., 837-840
BUCK G., COUDURIER J., QUESNIL J. J. (1952) Sur deux nouveaux plasmodium observes chez un lémurien de Madagascar splénectomisé. Arch. Inst. Pasteur Alger. 30, 240-243
CADIGAN F. C., JR., SPERTZEL R. O., CHAICUMPAN V., PUHOMCHAREON S. (1966) Plasmodium falciparum in non-human primates (macaque monkeys). Mil Med 131, 9, Suppl:959-960
CALIFORNIA BIOMEDICAL RESEARCH ASSOCIATION (2004). (Page consultée le 9 juin 2004). primates in biomedical research. [en ligne] Adresse: http://www.ca-biomed.org/pdf/mediakit/Primate.pdf

142
CARABALLO A., CARABALLO B., RODRIGUEZ-ACOSTA A. (2004) Preliminary assessment of medicinal plants used as antimalarials in the southeastern Venezuelan Amazon. Rev Soc Bras Med Trop 37, 2, 186-188
CARCABOSO A. M., HERNANDEZ R. M., IGARTUA M., GASCON A. R., ROSAS J. E., PATARROYO M. E., PEDRAZ J. L. (2003) Immune response after oral administration of the encapsulated malaria synthetic peptide SPf66. Int J Pharm 260, 2, 273-282
CARCABOSO A. M., HERNANDEZ R. M., IGARTUA M., ROSAS J. E., PATARROYO M. E., PEDRAZ J. L. (2004) Potent, long lasting systemic antibody levels and mixed Th1/Th2 immune response after nasal immunization with malaria antigen loaded PLGA microparticles. Vaccine 22, 11-12, 1423-1432
CARLTON J. M., ANGIUOLI S. V., SUH B. B., KOOIJ T. W., PERTEA M., SILVA J. C., ERMOLAEVA M. D., ALLEN J. E., SELENGUT J. D., KOO H. L., PETERSON J. D., POP M., KOSACK D. S., SHUMWAY M. F., BIDWELL S. L., SHALLOM S. J., VAN AKEN S. E., RIEDMULLER S. B., FELDBLYUM T. V., CHO J. K., QUACKENBUSH J., SEDEGAH M., SHOAIBI A., CUMMINGS L. M., FLORENS L., YATES J. R., RAINE J. D., SINDEN R. E., HARRIS M. A., CUNNINGHAM D. A., PREISER P. R., BERGMAN L. W., VAIDYA A. B., VAN LIN L. H., JANSE C. J., WATERS A. P., SMITH H. O., WHITE O. R., SALZBERG S. L., VENTER J. C., FRASER C. M., HOFFMAN S. L., GARDNER M. J., CARUCCI D. J. (2002) Genome sequence and comparative analysis of the model rodent malaria parasite Plasmodium yoelii yoelii. Nature 419, 6906, 512-519
CARLTON J. M., HAYTON K., CRAVO P. V., WALLIKER D. (2001) Of mice and malaria mutants: unravelling the genetics of drug resistance using rodent malaria models. Trends Parasitol 17, 5, 236-242
CARROLO M., GIORDANO S., CABRITA-SANTOS L., CORSO S., VIGARIO A. M., SILVA S., LEIRIAO P., CARAPAU D., ARMAS-PORTELA R., COMOGLIO P. M., RODRIGUEZ A., MOTA M. M. (2003) Hepatocyte growth factor and its receptor are required for malaria infection. Nat Med 9, 11, 1363-1369
CARTER R. (2003) Speculations on the origins of Plasmodium vivax malaria. Trends Parasitol 19, 5, 214-219
CARTER R., DIGGS C. L. (1977) Plamodia of rodents in: Kreier J. P. Parasitic Protozoa. Academic Press, New York, 359-465
CARUCCI D. J. (2005) Advances in malaria genomics since MIM Arusha, 2002. Acta Trop in press
CARVALHO L. J., DANIEL-RIBEIRO C. T., GOTO H. (2002) Malaria vaccine: candidate antigens, mechanisms, constraints and prospects. Scand J Immunol 56, 4, 327-343

143
CARVALHO T. G., MENARD R. (2005) Manipulating the Plasmodium genome. Curr Issues Mol Biol 7, 1, 39-55
CASTILLO-DAVIS C. I., BEDFORD T. B., HARTL D. L. (2004) Accelerated Rates of Intron Gain/Loss and Protein Evolution in Duplicate Genes in Human and Mouse Malaria Parasites. Mol Biol Evol 21, 7, 1422-1427
CATTERUCCIA F., GODFRAY H. C., CRISANTI A. (2003) Impact of genetic manipulation on the fitness of Anopheles stephensi mosquitoes. Science 299, 5610, 1225-1227
CAVALIERI P., SINGER P. (2004) The Great Ape Project : Equality beyond Humanity St. Martin's Griffin,, London, 312 pp.
CENTRE NATIONAL DE LA RECHERCHE SCIENTIFIQUE (2004). (Page consultée le 25 mai 2004). Paludisme. [en ligne] Adresse: http://www.cnrs.fr/SDV/Dept/paludisme.pdf
CHAN A. W., CHONG K. Y., MARTINOVICH C., SIMERLY C., SCHATTEN G. (2001) Transgenic monkeys produced by retroviral gene transfer into mature oocytes. Science 291, 5502, 309-312
CITES (2004). (Page consultée le 21 août 2004). Article III de la Convention de Washington. [en ligne] Adresse: http://www.cites.org/fra/disc/text.shtml#III
COATNEY G. R., COLLINS W. E., WARREN M., CONTACOS P. G. (1971) The Primate malaria US Government Printing Office, Washington DC, 366 pp.
COGSWELL F. B. (2000). (Page consultée le 3 janvier 2004). Malaria and Piroplasms of Non-Human Primates. [en ligne] Adresse: http://www.ivis.org/advances/Parasit_Bowman/cogswell_primate/chapter_frm.asp?LA=1
COHEN S., MC G. I., CARRINGTON S. (1961) Gamma-globulin and acquired immunity to human malaria. Nauchni Tr Vissh Med Inst Sofiia 192, 733-737
COLLINS W., AIKAWA M. (1993) Plasmodia of Nonhuman Primates. Parasitic Protozoa 5, 105-133
COLLINS W. E. (1992) South American monkeys in the development and testing of malarial vaccines--a review. Mem Inst Oswaldo Cruz 87 Suppl 3, 401-406
COLLINS W. E. (2003) CDC Program Reactivates Archived Parasites. MR4 Malaria Update 1-2
COLLINS W. E., CONTACOS P. G. (1972) Immunization of monkeys against Plasmodium cynomolgi by X-irradiated sporozoites. Nat New Biol 236, 67, 176-177

144
COLLINS W. E., JEFFERY G. M. (2005a) Plasmodium ovale: Parasite and Disease. Clin Microbiol Rev 18, 3, 570-581
COLLINS W. E., LOBEL H. O., MCCLURE H., STROBERT E., GALLAND G. G., TAYLOR F., BARRETO A. L., ROBERTO R. R., SKINNER J. C., ADAMS S., ET AL. (1994a) The China I/CDC strain of Plasmodium malariae in Aotus monkeys and chimpanzees. Am J Trop Med Hyg 50, 1, 28-32
COLLINS W. E., MCCLURE H. M., STROBERT E., FILIPSKI V., SKINNER J. C., STANFILL P. S., RICHARDSON B. B., MORRIS C. (1990) Infection of chimpanzees with the Uganda I/CDC strain of Plasmodium malariae. Am J Trop Med Hyg 42, 2, 99-103
COLLINS W. E., MCCLURE H. M., SWENSON R. B., MEHAFFEY P. C., SKINNER J. C. (1986) Infection of mosquitoes with Plasmodium vivax from chimpanzees using membrane feeding. Am J Trop Med Hyg 35, 1, 56-60
COLLINS W. E., NUSSENZWEIG R. S., BALLOU W. R., RUEBUSH T. K., 2ND, NARDIN E. H., CHULAY J. D., MAJARIAN W. R., YOUNG J. F., WASSERMAN G. F., BATHURST I., ET AL. (1989a) Immunization of Saimiri sciureus boliviensis with recombinant vaccines based on the circumsporozoite protein of Plasmodium vivax. Am J Trop Med Hyg 40, 5, 455-464
COLLINS W. E., PYE D., CREWTHER P. E., VANDENBERG K. L., GALLAND G. G., SULZER A. J., KEMP D. J., EDWARDS S. J., COPPEL R. L., SULLIVAN J. S., ET AL. (1994b) Protective immunity induced in squirrel monkeys with recombinant apical membrane antigen-1 of Plasmodium fragile. Am J Trop Med Hyg 51, 6, 711-719
COLLINS W. E., RICHARDSON B. B., MORRIS C. L., SULLIVAN J. S., GALLAND G. G. (1998) Salvador II strain of Plasmodium vivax in Aotus monkeys and mosquitoes for transmission-blocking vaccine trials. Am J Trop Med Hyg 59, 1, 29-34
COLLINS W. E., SATTABONGKOT J., WIRTZ R. A., SKINNER J. C., BRODERSON J. R., MILLET P. G., MORRIS C. L., RICHARDSON B. B., SULLIVAN J., FILIPSKI V. K. (1992a) Development of a polymorphic strain of Plasmodium vivax in monkeys. J Parasitol 78, 3, 485-491
COLLINS W. E., SKINNER J. C., BRODERSON J. R., FILIPSKI V. K., MORRIS C. M., STANFILL P. S., WARREN M. (1992b) Susceptibility of Macaca fascicularis monkeys from Mauritius to different species of Plasmodium. J Parasitol 78, 3, 505-511
COLLINS W. E., SKINNER J. C., BRODERSON J. R., RICHARDSON B. B., STANFILL P. S. (1989b) The Uganda I/CDC strain of Plasmodium malariae in Saimiri sciureus boliviensis. J Parasitol 75, 2, 310-313

145
COLLINS W. E., SKINNER J. C., PAPPAIOANOU M., BRODERSON J. R., MA N. S., FILIPSKI V., STANFILL P. S., ROGERS L. (1988) Infection of Peruvian Aotus nancymai monkeys with different strains of Plasmodium falciparum, P. vivax, and P. malariae. J Parasitol 74, 3, 392-398
COLLINS W. E., SULLIVAN J. S., GALLAND G. G., WILLIAMS A., NACE D., WILLIAMS T. (2002) Potential of the Panama strain of Plasmodium vivax for the testing of malarial vaccines in Aotus nancymai monkeys. Am J Trop Med Hyg 67, 5, 454-458
COLLINS W. E., SULLIVAN J. S., GALLAND G. G., WILLIAMS A., NACE D., WILLIAMS T., BARNWELL J. W. (2005b) Observations on the Vietnam Palo Alto strain of Plasmodium vivax in two species of Aotus monkeys. J Parasitol 91, 2, 461-463
COMMISSION DES COMMUNAUTÉS EUROPÉENNES (2003). (Page consultée le 21 mars 2004). Troisième rapport de la Commission au Conseil et au Parlement Européen sur les statistiques concernant le nombre d'animaux utilisés à des fins expérimentales et à d'autres fins scientifiques dans les Etats membres de l'Union Européenne. [en ligne] Adresse: http://europa.eu.int/eur-lex/fr/com/rpt/2003/com2003_0019fr01.pdf
CONSEIL DE L'EUROPE (1986). (Page consultée le 21 août 2004). Directive européenne 86/609 du 24/11/86. [en ligne] Adresse: http://www.ulg.ac.be/animal/loibelge.htm
CONTACOS P. G., COATNEY G. R., ORIHEL T. C., COLLINS W. E., CHIN W., JETER M. H. (1970) Transmission of Plasmodium schwetzi from the chimpanzee to man by mosquito bite. Am J Trop Med Hyg 19, 2, 190-195
CONTAMIN H., BEHR C., MERCEREAU-PUIJALON O., MICHEL J. (2000) Plasmodium falciparumin the squirrel monkey (Saimiri sciureus): infection of non-splenectomised animals as a model for exploring clinical manifestations of malaria. Microbes Infect 2, 8, 945-954
CONWAY D. J., BAUM J. (2002) In the blood--the remarkable ancestry of Plasmodium falciparum. Trends Parasitol 18, 8, 351-355
COOK J. E., FINEG J., MIKSCH E. D., BRITZ W. E. (1961) The chimpanzee. Procurement, standardization, and use in biomedical research. J Chronic Dis 138, 366-369
CUI L., MASCORRO C. N., FAN Q., RZOMP K. A., KHUNTIRAT B., ZHOU G., CHEN H., YAN G., SATTABONGKOT J. (2003) Genetic diversity and multiple infections of Plasmodium vivax malaria in Western Thailand. Am J Trop Med Hyg 68, 5, 613-619
CUMMINS L. B., KALTER S. S. (1980) Production and standardization of the chimpanzee for research. Dev Biol Stand 45, 67-72
DA FONSECA F. (1951) Plasmodio de primata do Brasil. Mem. Inst. Oswaldo Cruz 49, 543-551

146
DAMIAN R. T., GREENE N. D., KALTER S. S. (1971) IgG subclasses in the baboon (Papio cynocephalus). J Immunol 106, 1, 246-257
DANIS M. (2003) [Therapeutic advances against malaria in 2003]. Med Trop (Mars) 63, 3, 267-270
DANIS M., BRICAIRE F. (2003) The new drug combinations: their place in the treatment of uncomplicated Plasmodium falciparum malaria. Fundam Clin Pharmacol 17, 2, 155-160
DAUBERSIES P., THOMAS A. W., MILLET P., BRAHIMI K., LANGERMANS J. A., OLLOMO B., BENMOHAMED L., SLIERENDREGT B., ELING W., VAN BELKUM A., DUBREUIL G., MEIS J. F., GUERIN-MARCHAND C., CAYPHAS S., COHEN J., GRAS-MASSE H., DRUILHE P., MOHAMED L. B. (2000) Protection against Plasmodium falciparum malaria in chimpanzees by immunization with the conserved pre-erythrocytic liver-stage antigen 3. Nat Med 6, 11, 1258-1263
DAVISON B. B., COGSWELL F. B., BASKIN G. B., FALKENSTEIN K. P., HENSON E. W., KROGSTAD D. J. (2000) Placental changes associated with fetal outcome in the Plasmodium coatneyi/rhesus monkey model of malaria in pregnancy. Am J Trop Med Hyg 63, 3-4, 158-173
DAVISON B. B., COGSWELL F. B., BASKIN G. B., FALKENSTEIN K. P., HENSON E. W., TARANTAL A. F., KROGSTAD D. J. (1998) Plasmodium coatneyi in the rhesus monkey (Macaca mulatta) as a model of malaria in pregnancy. Am J Trop Med Hyg 59, 2, 189-201
DE ARRUDA M., NARDIN E. H., NUSSENZWEIG R. S., COCHRANE A. H. (1989) Sero-epidemiological studies of malaria in Indian tribes and monkeys of the Amazon Basin of Brazil. Am J Trop Med Hyg 41, 4, 379-385
DE MONBRISON F., RAYNAUD D., LATOUR-FONDANAICHE C., ANGEI C., KAISER K., PEYRON F., PICOT S. (2003) [Real-time PCR for detection of molecular markers of resistance in Plasmodium falciparum]. Pathol Biol (Paris) 51, 8-9, 528-533
DE SOUZA J. B., RILEY E. M. (2002) Cerebral malaria: the contribution of studies in animal models to our understanding of immunopathogenesis. Microbes Infect 4, 3, 291-300
DE THOISY B., MICHEL J. C., VOGEL I., VIE J. C. (2000) A survey of hemoparasite infections in free-ranging mammals and reptiles in French Guiana. J Parasitol 86, 5, 1035-1040
DEANE L. M. (1992) Simian malaria in Brazil. Mem Inst Oswaldo Cruz 87 Suppl 3, 1-20
DEANE L. M., FERREIRA NETO J., SILVEIRA I. P. S. (1966) Experimental infection of a splenectomized squirrel monkey, Saimiri sciureus, with Plasmodium vivax. Trans R Soc Trop Med Hyg 60, 811-812

147
DEANS J. A., KNIGHT A. M., JEAN W. C., WATERS A. P., COHEN S., MITCHELL G. H. (1988) Vaccination trials in rhesus monkeys with a minor, invariant, Plasmodium knowlesi 66 kD merozoite antigen. Parasite Immunol 10, 5, 535-552
DECHY-CABARET O., BENOIT-VICAL F., LOUP C., ROBERT A., GORNITZKA H., BONHOURE A., VIAL H., MAGNAVAL J. F., SEGUELA J. P., MEUNIER B. (2004) Synthesis and antimalarial activity of trioxaquine derivatives. Chemistry 10, 7, 1625-1636
DENIS M. B., DAVIS T. M., HEWITT S., INCARDONA S., NIMOL K., FANDEUR T., PORAVUTH Y., LIM C., SOCHEAT D. (2002) Efficacy and safety of dihydroartemisinin-piperaquine (Artekin) in Cambodian children and adults with uncomplicated falciparum malaria. Clin Infect Dis 35, 12, 1469-1476
DESAI S. A., BEZRUKOV S. M., ZIMMERBERG J. (2000) A voltage-dependent channel involved in nutrient uptake by red blood cells infected with the malaria parasite. Nature 406, 6799, 1001-1005
DIEHL K. H., HULL R., MORTON D., PFISTER R., RABEMAMPIANINA Y., SMITH D. L., VIDAL J., VAN DE VORSTENBOSCH C. (2001) A good practice guide to the administration of substances and removal of the blood, including routes and volumes. J Appl Toxicol 21, 15-23
DISSANAIKE A. S., NELSON P., GARNHAM P. C. C. (1965a) Plasmodium simiovale sp. nov., a new simian malaria parasite from Ceylon. Ceylon J. Med. Sci. 14, 27-32
DISSANAIKE A. S., NELSON P., GARNHAM P. C. C. (1965b) Two new malaria parasites, Plasmodium cynomolgi ceylonensis subsp. nov. and Plasmodium fragile sp. nov. from monkeys in Ceylon. Ceylon J. Med. Sci. 14, 1-9
DOBSON M. J. (1998) Bitter-sweet solutions for malaria: exploring natural remedies from the past. Parassitologia 40, 1-2, 69-81
DORSEY G., VLAHOS J., KAMYA M. R., STAEDKE S. G., ROSENTHAL P. J. (2003) Prevention of increasing rates of treatment failure by combining sulfadoxine-pyrimethamine with artesunate or amodiaquine for the sequential treatment of malaria. J Infect Dis 188, 8, 1231-1238
DROY J., J. L. (2003). (Page consultée le 4 mai 2004). Santé voyage Rouen, paludisme. [en ligne] Adresse: http://www.chu-rouen.fr/cap/paludisme
DRUILHE P., GENTILINI M. (1982) [In vitro cultivation of Plasmodium falciparum. Applications and limits.- Methodology]. Med Trop (Mars) 42, 4, 437-462
DRUILHE P., HAGAN P., ROOK G. A. (2002) The importance of models of infection in the study of disease resistance. Trends Microbiol 10, 10 Suppl, S38-46

148
DUBBELD M. A., KOCKEN C. H., THOMAS A. W. (1998) Merozoite surface protein 2 of Plasmodium reichenowi is a unique mosaic of Plasmodium falciparum allelic forms and species-specific elements. Mol Biochem Parasitol 92, 1, 187-192
DUFFY P. E., KRZYCH U., FRANCIS S., FRIED M. (2005) Malaria vaccines: using models of immunity and functional genomics tools to accelerate the development of vaccines against Plasmodium falciparum. Vaccine 23, 17-18, 2235-2242
DUPUIS M. (2003). (Page consultée le 22 août 2004). La classification générale des primates. [en ligne] Adresse: http://www.inrp.fr/acces/biotic/evolut/phylogene/documentation/html/prim.htm
DURIEZ T. (2002). (Page consultée le 20 mars 2003). Cours de parasitologie. [en ligne] Adresse: http://arachosia.univ-lille2.fr/labos/parasito/Internat/courspar/paludis.html
EGAN A. F., FABUCCI M. E., SAUL A., KASLOW D. C., MILLER L. H. (2002) Aotus New World monkeys: model for studying malaria-induced anemia. Blood 99, 10, 3863-3866
EICHBERG J. W., SPECK J. T., JR. (1988) Establishment of a chimpanzee retirement fund: maintenance after experimentation. J Med Primatol 17, 2, 71-76
EPSTEIN J. E., CHAROENVIT Y., KESTER K. E., WANG R., NEWCOMER R., FITZPATRICK S., RICHIE T. L., TORNIEPORTH N., HEPPNER D. G., OCKENHOUSE C., MAJAM V., HOLLAND C., ABOT E., GANESHAN H., BERZINS M., JONES T., FREYDBERG C. N., NG J., NORMAN J., CARUCCI D. J., COHEN J., HOFFMAN S. L. (2004) Safety, tolerability, and antibody responses in humans after sequential immunization with a PfCSP DNA vaccine followed by the recombinant protein vaccine RTS,S/AS02A. Vaccine 22, 13-14, 1592-1603
ERHART A., THANG N. D., HUNG N. Q., TOI LE V., HUNG LE X., TUY T. Q., CONG LE D., SPEYBROECK N., COOSEMANS M., D'ALESSANDRO U. (2004) Forest malaria in Vietnam: a challenge for control. Am J Trop Med Hyg 70, 2, 110-118
ESCALANTE A. A., AYALA F. J. (1994) Phylogeny of the malarial genus Plasmodium, derived from rRNA gene sequences. Proc Natl Acad Sci U S A 91, 24, 11373-11377
ESCALANTE A. A., BARRIO E., AYALA F. J. (1995) Evolutionary origin of human and primate malarias: evidence from the circumsporozoite protein gene. Mol Biol Evol 12, 4, 616-626
ESCALANTE A. A., CORNEJO O. E., FREELAND D. E., POE A. C., DURREGO E., COLLINS W. E., LAL A. A. (2005) A monkey's tale: the origin of Plasmodium vivax as a human malaria parasite. Proc Natl Acad Sci U S A 102, 6, 1980-1985
ESCALANTE A. A., FREELAND D. E., COLLINS W. E., LAL A. A. (1998) The evolution of primate malaria parasites based on the gene encoding cytochrome b from the linear mitochondrial genome. Proc Natl Acad Sci U S A 95, 14, 8124-8129

149
ESCALANTE A. A., GOLDMAN I. F., DE RIJK P., DE WACHTER R., COLLINS W. E., QARI S. H., LAL A. A. (1997) Phylogenetic study of the genus Plasmodium based on the secondary structure-based alignment of the small subunit ribosomal RNA. Mol Biochem Parasitol 90, 1, 317-321
ETKIN N. L. (2003) The co-evolution of people, plants, and parasites: biological and cultural adaptations to malaria. Proc Nutr Soc 62, 2, 311-317
EYLES D. E., COATNEY G. R., GETZ M. (1960) Vivax-type malaria parasite of macaques transmissible to man. Science 132, 1812-1813
EYLES D. E., FONG Y. L., DUNN F. L., GUINN E., WARREN M., SANDOSHAN A. A. (1964) Plasmodium youngi n. sp., a malaria parasite of the Malayan gibbon, Hylobates lar lar. Am. J. Trop. Med. Hyg. 13, 248-255
EYLES D. E., FONG Y. L., WARREN M., GUINN E., SANDOSHAN A. A., WHARTON R. H. (1962a) Plasmodium coatneyi, a nex species of primate malaria from Malaya. Am. J. Trop. Med. Hyg. 11, 597-604
EYLES D. E., LAING A. B. G., FONG Y. L. (1962b) Plasmodium fieldi sp. nov., a new species of malaria parasite from the pig-tailed macaque in Malaya. Ann. Trop. Med. Parasit. 56, 242-247
FANDEUR T., VOLNEY B., PENEAU C., DE THOISY B. (2000) Monkeys of the rainforest in French Guiana are natural reservoirs for P. brasilianum/P. malariae malaria. Parasitology 120 ( Pt 1), 11-21
FARCAS G. A., ZHONG K. J., LOVEGROVE F. E., GRAHAM C. M., KAIN K. C. (2003) Evaluation of the Binax NOW ICT test versus polymerase chain reaction and microscopy for the detection of malaria in returned travelers. Am J Trop Med Hyg 69, 6, 589-592
FERREIRA A., SCHOFIELD L., ENEA V., SCHELLEKENS H., VAN DER MEIDE P., COLLINS W. E., NUSSENZWEIG R. S., NUSSENZWEIG V. (1986) Inhibition of development of exoerythrocytic forms of malaria parasites by gamma-interferon. Science 232, 4752, 881-884
FINOT L. (2004). (Page consultée le 2 avril 2004). Paludisme. [en ligne] Adresse: http://membres.lycos.fr/lfinot/maladie/palu2.html
FLICK K., CHEN Q. (2004) var genes, PfEMP1 and the human host. Mol Biochem Parasitol 134, 1, 3-9
FOHL L. M., ROOS D. S. (2003) Fitness effects of DHFR-TS mutations associated with pyrimethamine resistance in apicomplexan parasites. Mol Microbiol 50, 4, 1319-1327

150
FOTH B. J., RALPH S. A., TONKIN C. J., STRUCK N. S., FRAUNHOLZ M., ROOS D. S., COWMAN A. F., MCFADDEN G. I. (2003) Dissecting apicoplast targeting in the malaria parasite Plasmodium falciparum. Science 299, 5607, 705-708
FREUND J., K. T., H. S., A. W., E. S. (1945) Immunization of rhesus monkeys against malarial infection (Plasmodium knowlesi) with killed parasites and adjuvants. Science 102, 202-204
FUJIOKA H., MILLET P., MAENO Y., NAKAZAWA S., ITO Y., HOWARD R. J., COLLINS W. E., AIKAWA M. (1994) A nonhuman primate model for human cerebral malaria: rhesus monkeys experimentally infected with Plasmodium fragile. Exp Parasitol 78, 4, 371-376
GACHOT B., BRUNEEL F., BEHR C. (2001) Paludisme grave. Rev Prat 51, 638-643
GACHOT B., PAYS J. F. (2002) Accès palustre simple. Méd Thérap 8, 115-123
GALINDO C. M., ACOSTA C. J., SCHELLENBERG D., APONTE J. J., ROCA A., OETTLI A., URASSA H., ARMSTRONG SCHELLENBERG J., KAHIGWA E., ASCASO C., MSHINDA H., LWILLA F., VIDAL J., MENENDEZ C., TANNER M., ALONSO P. L. (2000) Humoral immune responses during a malaria vaccine trial in Tanzanian infants. Parasite Immunol 22, 9, 437-443
GALLUP J., SACHS J. (1999). (Page consultée le 10 septembre 2003). Malaria, Climate, and Poverty. [en ligne] Adresse: http://www.cid.harvard.edu/caer2/htm/content/papers/paper48/paper48.htm
GARCON N., HEPPNER D. G., COHEN J. (2003) Development of RTS,S/AS02: a purified subunit-based malaria vaccine candidate formulated with a novel adjuvant. Expert Rev Vaccines 2, 2, 231-238
GARDNER M. J., GOLDMAN N., BARNETT P., MOORE P. W., RANGACHARI K., STRATH M., WHYTE A., WILLIAMSON D. H., WILSON R. J. (1994) Phylogenetic analysis of the rpoB gene from the plastid-like DNA of Plasmodium falciparum. Mol Biochem Parasitol 66, 2, 221-231
GARDNER M. J., HALL N., FUNG E., WHITE O., BERRIMAN M., HYMAN R. W., CARLTON J. M., PAIN A., NELSON K. E., BOWMAN S., PAULSEN I. T., JAMES K., EISEN J. A., RUTHERFORD K., SALZBERG S. L., CRAIG A., KYES S., CHAN M. S., NENE V., SHALLOM S. J., SUH B., PETERSON J., ANGIUOLI S., PERTEA M., ALLEN J., SELENGUT J., HAFT D., MATHER M. W., VAIDYA A. B., MARTIN D. M., FAIRLAMB A. H., FRAUNHOLZ M. J., ROOS D. S., RALPH S. A., MCFADDEN G. I., CUMMINGS L. M., SUBRAMANIAN G. M., MUNGALL C., VENTER J. C., CARUCCI D. J., HOFFMAN S. L., NEWBOLD C., DAVIS R. W., FRASER C. M., BARRELL B. (2002) Genome sequence of the human malaria parasite Plasmodium falciparum. Nature 419, 6906, 498-511

151
GARNHAM P. C., RAJAPAKSA N., PETERS W., KILLICK-KENDRICK R. (1972) Malaria parasites of the orang-utan (Pongo pygmaeus). Ann Trop Med Parasitol 66, 2, 287-294
GARRAUD O., MAHANTY S., PERRAUT R. (2003) Malaria-specific antibody subclasses in immune individuals: a key source of information for vaccine design. Trends Immunol 24, 1, 30-35
GASCOYNE P., SATAYAVIVAD J., RUCHIRAWAT M. (2004) Microfluidic approaches to malaria detection. Acta Trop 89, 3, 357-369
GEIMAN Q. M., MEAGHER M. J. (1967) Susceptibility of a New World monkey to Plasmodium falciparum from man. Nature 215, 99, 437-439
GEIMAN Q. M., SIDDIQUI W. A. (1969) Susceptibility of a New World monkey to Plasmodium malariae from aman. Am J Trop Med Hyg 18, 3, 351-354
GENTON B., AL-YAMAN F., ANDERS R., SAUL A., BROWN G., PYE D., IRVING D. O., BRIGGS W. R., MAI A., GINNY M., ADIGUMA T., RARE L., GIDDY A., REBER-LISKE R., STUERCHLER D., ALPERS M. P. (2000) Safety and immunogenicity of a three-component blood-stage malaria vaccine in adults living in an endemic area of Papua New Guinea. Vaccine 18, 23, 2504-2511
GENTON B., AL-YAMAN F., BETUELA I., ANDERS R. F., SAUL A., BAEA K., MELLOMBO M., TARAIKA J., BROWN G. V., PYE D., IRVING D. O., FELGER I., BECK H. P., SMITH T. A., ALPERS M. P. (2003) Safety and immunogenicity of a three-component blood-stage malaria vaccine (MSP1, MSP2, RESA) against Plasmodium falciparum in Papua New Guinean children. Vaccine 22, 1, 30-41
GENTON B., CORRADIN G. (2002) Malaria vaccines: from the laboratory to the field. Curr Drug Targets Immune Endocr Metabol Disord 2, 3, 255-267
GIAO P. T., DE VRIES P. J., HUNG L. Q., BINH T. Q., NAM N. V., KAGER P. A. (2003) Atovaquone-proguanil for recrudescent Plasmodium falciparum in Vietnam. Ann Trop Med Parasitol 97, 6, 575-580
GOLENDA C. F., LI J., ROSENBERG R. (1997) Continuous in vitro propagation of the malaria parasite Plasmodium vivax. Proc Natl Acad Sci U S A 94, 13, 6786-6791
GONDER R., VON BERNBERG-GOSSLER H. V. (1908) Untersuchungen über Malaria-plasmodien der Affen. Malaria-Inter. Arch. Leipzig 1, 47-56
GOOD M. F., XU H., WYKES M., ENGWERDA C. R. (2005) Development and regulation of cell-mediated immune responses to the blood stages of malaria: implications for vaccine research. Annu Rev Immunol 23, 69-99

152
GOODMAN S. (2001) Europe brings experiments on chimpanzees to an end. Nature 411, 6834, 123
GOODMAN S., CHECK E. (2002) The great primate debate. Nature 417, 6890, 684-687
GORDI T., LEPIST E. I. (2004) Artemisinin derivatives: toxic for laboratory animals, safe for humans? Toxicol Lett 147, 2, 99-107
GRASSE P. (1953) Traité de zoologie Masson et Cie, Paris, 1160 pp.
GRAVES P., GELBAND H. (2003) Vaccines for preventing malaria. Cochrane Database Syst Rev 1, CD000129
GRIMWADE K., FRENCH N., MBATHA D. D., ZUNGU D. D., DEDICOAT M., GILKS C. F. (2004) HIV infection as a cofactor for severe falciparum malaria in adults living in a region of unstable malaria transmission in South Africa. Aids 18, 3, 547-554
GROBUSCH M. P., BORRMANN S., OMVA J., ISSIFOU S., KREMSNER P. G. (2003) Severe malaria in a splenectomised Gabonese woman. Wien Klin Wochenschr 115, 1-2, 63-65
GYSIN J., AIKAWA M., TOURNEUR N., TEGOSHI T. (1992) Experimental Plasmodium falciparum cerebral malaria in the squirrel monkey Saimiri sciureus. Exp Parasitol 75, 4, 390-398
HACIA J. G. (2001) Genome of the apes. Trends Genet 17, 11, 637-645
HADDAD D., BILCIKOVA E., WITNEY A. A., CARLTON J. M., WHITE C. E., BLAIR P. L., CHATTOPADHYAY R., RUSSELL J., ABOT E., CHAROENVIT Y., AGUIAR J. C., CARUCCI D. J., WEISS W. R. (2004) Novel antigen identification method for discovery of protective malaria antigens by rapid testing of DNA vaccines encoding exons from the parasite genome. Infect Immun 72, 3, 1594-1602
HALBERSTAEDER L., VON PROWAZEK S. (1907) Untersuchungen über die Malaria-parasiten der Affen. Arb. Gesundheitsamte (Berlin) 26, 37-43
HAMMADI A., RAMIANDRASOA F., SINOU V., ROGIER C., FUSAI T., LE BRAS J., PARZY D., KUNESCH G., PRADINES B. (2003) Cellular uptake of a catechol iron chelator and chloroquine into Plasmodium falciparum infected erythrocytes. Biochem Pharmacol 65, 8, 1351-1360

153
HAPPI C. T., GBOTOSHO G. O., SOWUNMI A., FALADE C. O., AKINBOYE D. O., GERENA L., KYLE D. E., MILHOUS W., WIRTH D. F., ODUOLA A. M. (2004) Molecular analysis of Plasmodium falciparum recrudescent malaria infections in children treated with chloroquine in Nigeria. Am J Trop Med Hyg 70, 1, 20-26
HARLING R., CROOK P., LEWTHWAITE P., EVANS M., SCHMID M. L., BEECHING N. J. (2004) Burden and cost of imported infections admitted to infectious diseases units in England and Wales in 1998 and 1999. J Infect 48, 2, 139-144
HARTL D. L. (2004) The origin of malaria: mixed messages from genetic diversity. Nat Rev Microbiol 2, 1, 15-22
HASTINGS I. M., D'ALESSANDRO U. (2000) Modelling a predictable disaster: the rise and spread of drug-resistantmalaria. Parasitol Today 16, 8, 340-347
HASTINGS I. M., WATKINS W. M., WHITE N. J. (2002) The evolution of drug-resistant malaria: the role of drug elimination half-life. Philos Trans R Soc Lond B Biol Sci 357, 1420, 505-519
HAYTON K., SU X. Z. (2004) Genetic and biochemical aspects of drug resistance in malaria parasites. Curr Drug Targets Infect Disord 4, 1, 1-10
HEALER J., MURPHY V., HODDER A. N., MASCIANTONIO R., GEMMILL A. W., ANDERS R. F., COWMAN A. F., BATCHELOR A. (2004) Allelic polymorphisms in apical membrane antigen-1 are responsible for evasion of antibody-mediated inhibition in Plasmodium falciparum. Mol Microbiol 52, 1, 159-168
HEMINGWAY J., CRAIG A. (2004) Parasitology. New ways to control malaria. Science 303, 5666, 1984-1985
HEPPNER D. G., JR., KESTER K. E., OCKENHOUSE C. F., TORNIEPORTH N., OFORI O., LYON J. A., STEWART V. A., DUBOIS P., LANAR D. E., KRZYCH U., MORIS P., ANGOV E., CUMMINGS J. F., LEACH A., HALL B. T., DUTTA S., SCHWENK R., HILLIER C., BARBOSA A., WARE L. A., NAIR L., DARKO C. A., WITHERS M. R., OGUTU B., POLHEMUS M. E., FUKUDA M., PICHYANGKUL S., GETTYACAMIN M., DIGGS C., SOISSON L., MILMAN J., DUBOIS M. C., GARCON N., TUCKER K., WITTES J., PLOWE C. V., THERA M. A., DUOMBO O. K., PAU M. G., GOUDSMIT J., BALLOU W. R., COHEN J. (2005) Towards an RTS,S-based, multi-stage, multi-antigen vaccine against falciparum malaria: progress at the Walter Reed Army Institute of Research. Vaccine 23, 17-18, 2243-2250
HERRERA S., PERLAZA B. L., BONELO A., AREVALO-HERRERA M. (2002) Aotus monkeys: their great value for anti-malaria vaccines and drug testing. Int J Parasitol 32, 13, 1625-1635

154
HIRUNPETCHARAT C., WIPASA J., SAKKHACHORNPHOP S., NITKUMHAN T., ZHENG Y. Z., PICHYANGKUL S., KRIEG A. M., WALSH D. S., HEPPNER D. G., GOOD M. F. (2003) CpG oligodeoxynucleotide enhances immunity against blood-stage malaria infection in mice parenterally immunized with a yeast-expressed 19 kDa carboxyl-terminal fragment of Plasmodium yoelii merozoite surface protein-1 (MSP1(19)) formulated in oil-based Montanides. Vaccine 21, 21-22, 2923-2932
HISAEDA H., MAEKAWA Y., IWAKAWA D., OKADA H., HIMENO K., KISHIHARA K., TSUKUMO S., YASUTOMO K. (2004) Escape of malaria parasites from host immunity requires CD4+ CD25+ regulatory T cells. Nat Med 10, 1, 29-30
HISAEDA H., SAUL A., REECE J. J., KENNEDY M. C., LONG C. A., MILLER L. H., STOWERS A. W. (2002) Merozoite surface protein 3 and protection against malaria in Aotus nancymai monkeys. J Infect Dis 185, 5, 657-664
HISAEDA H., YASUTOMO K., HIMENO K. (2005) Malaria: immune evasion by parasites. Int J Biochem Cell Biol 37, 4, 700-706
HOFFMAN S. L., SUBRAMANIAN G. M., COLLINS F. H., VENTER J. C. (2002) Plasmodium, human and Anopheles genomics and malaria. Nature 415, 6872, 702-709
HOUDEBINE L. M. (1998). (Page consultée le 17 mai 2004). Les animaux transgéniques permettent-ils de faire progresser la recherche médicale ? [en ligne] Adresse: http://www.inra.fr/Internet/Directions/DIC/ACTUALITES/DOSSIERS/OGM/houd.htm
HUFF C. G., OOGSTRAAL H. (1963) Plasmodium lemuris n. sp. from Lemur collaris. J. Infect. Dis. 112, 233-236
HUNT N. H., GRAU G. E. (2003) Cytokines: accelerators and brakes in the pathogenesis of cerebral malaria. Trends Immunol 24, 9, 491-499
HVIID L. (2005) Naturally acquired immunity to Plasmodium falciparum malaria in Africa. Acta Trop in press
IBIWOYE M. O., HOWARD C. V., SIBBONS P., HASAN M., VAN VELZEN D. (1993) Cerebral malaria in the rhesus monkey (Macaca mulatta): observations on host pathology. J Comp Pathol 108, 3, 303-310
INSTITUT DE VEILLE SANITAIRE (2003). (Page consultée le 10 juillet 2003). Bulletin Epidémiologique Hebdomadaire. [en ligne] Adresse: http://www.invs.sante.fr/beh/2003/26_27/beh_26_27_2003.pdf
INSTITUT PASTEUR (2005). (Page consultée le 18 juillet 2005). Envois de matériel biologique aux Centres Nationaux de Référence. [en ligne] Adresse: http://www.pasteur.fr/sante/clre/chap/envois/a.html

155
JANSSEN C. S., BARRETT M. P., TURNER C. M., PHILLIPS R. S. (2002) A large gene family for putative variant antigens shared by human and rodent malaria parasites. Proc R Soc Lond B Biol Sci 269, 1489, 431-436
JARRA W., SNOUNOU G. (1998) Only viable parasites are detected by PCR following clearance of rodent malarial infections by drug treatment or immune responses. Infect Immun 66, 8, 3783-3787
JENNES W., KESTENS L., F. N. D., L. S. B. (2002) Enhanced ELISPOT detection of antigen-specific T cell responses from cryopreserved specimens with addition of both IL-7 and IL-15, the Amplispot assay. J Immunol Methods 270, 99-108
JOHN C. C., MOORMANN A. M., PREGIBON D. C., SUMBA P. O., MCHUGH M. M., NARUM D. L., LANAR D. E., SCHLUCHTER M. D., KAZURA J. W. (2005) Correlation of High Levels of Antibodies to Multiple Pre-Erythrocytic Plasmodium Falciparum Antigens and Protection from Infection. Am J Trop Med Hyg 73, 1, 222-228
JONES T. R., GRAMZINSKI R. A., AGUIAR J. C., SIM B. K., NARUM D. L., FUHRMANN S. R., KUMAR S., OBALDIA N., HOFFMAN S. L. (2002) Absence of antigenic competition in Aotus monkeys immunized with Plasmodium falciparum DNA vaccines delivered as a mixture. Vaccine 20, 11-12, 1675-1680
JONES T. R., OBALDIA N., 3RD, GRAMZINSKI R. A., HOFFMAN S. L. (2000) Repeated infection of Aotus monkeys with Plasmodium falciparum induces protection against subsequent challenge with homologous and heterologous strains of parasite. Am J Trop Med Hyg 62, 6, 675-680
JOY D. A., FENG X., MU J., FURUYA T., CHOTIVANICH K., KRETTLI A. U., HO M., WANG A., WHITE N. J., SUH E., BEERLI P., SU X. Z. (2003) Early origin and recent expansion of Plasmodium falciparum. Science 300, 5617, 318-321
KAUP F. J., SCHWIBBE M. (2002) [Primates as experimental animals]. Dtsch Tierarztl Wochenschr 109, 3, 104-108
KEDZIERSKI L., ESCALANTE A. A., ISEA R., BLACK C. G., BARNWELL J. W., COPPEL R. L. (2002) Phylogenetic analysis of the genus Plasmodium based on the gene encoding adenylosuccinate lyase. Infect Genet Evol 1, 4, 297-301
KENNEDY D., WALKER E. (1997) Malaria at the millenium. Scott Med J 42, 1, 3
KIENY M. P. (2003) [Malaria vaccines: state of advancement]. Med Trop (Mars) 63, 3, 245-246
KISSINGER J. C., COLLINS W. E., LI J., MCCUTCHAN T. F. (1998) Plasmodium inui is not closely related to other quartan Plasmodium species. J Parasitol 84, 2, 278-282

156
KOCKEN C. H., DUBBELD M. A., VAN DER WEL A., PRONK J. T., WATERS A. P., LANGERMANS J. A., THOMAS A. W. (1999) High-level expression of Plasmodium vivax apical membrane antigen 1 (AMA-1) in Pichia pastoris: strong immunogenicity in Macaca mulatta immunized with P. vivax AMA-1 and adjuvant SBAS2. Infect Immun 67, 1, 43-49
KOCKEN C. H., NARUM D. L., MASSOUGBODJI A., AYIVI B., DUBBELD M. A., VAN DER WEL A., CONWAY D. J., SANNI A., THOMAS A. W. (2000) Molecular characterisation of Plasmodium reichenowi apical membrane antigen-1 (AMA-1), comparison with P. falciparum AMA-1, and antibody-mediated inhibition of red cell invasion. Mol Biochem Parasitol 109, 2, 147-156
KORENROMP E. L., WILLIAMS B. G., GOUWS E., DYE C., SNOW R. W. (2003) Measurement of trends in childhood malaria mortality in Africa: an assessment of progress toward targets based on verbal autopsy. Lancet Infect Dis 3, 6, 349-358
KROTOSKI W. A., GARNHAM P. C., COGSWELL F. B., COLLINS W. E., BRAY R. S., GWASZ R. W., KILLICK-KENDRICK R., WOLF R. H., SINDEN R., HOLLINGDALE M., ET AL. (1986) Observations on early and late post-sporozoite tissue stages in primate malaria. IV. Pre-erythrocytic schizonts and/or hypnozoites of Chesson and North Korean strains of Plasmodium vivax in the chimpanzee. Am J Trop Med Hyg 35, 2, 263-274
KRZYCH U., SCHWENK R., GUEBRE-XABIER M., SUN P., PALMER D., WHITE K., CHALOM I. (2000) The role of intrahepatic lymphocytes in mediating protective immunity induced by attenuated Plasmodium berghei sporozoites. Immunol Rev 174, 123-134
KUMAR S., JONES T. R., OAKLEY M. S., ZHENG H., KUPPUSAMY S. P., TAYE A., KRIEG A. M., STOWERS A. W., KASLOW D. C., HOFFMAN S. L. (2004) CpG oligodeoxynucleotide and Montanide ISA 51 adjuvant combination enhanced the protective efficacy of a subunit malaria vaccine. Infect Immun 72, 2, 949-957
KWIATKOWSKI D. (2000) Genetic susceptibility to malaria getting complex. Curr Opin Genet Dev 10, 3, 320-324
KWIATKOWSKI D. P. (2005) How Malaria Has Affected the Human Genome and What Human Genetics Can Teach Us about Malaria. Am J Hum Genet 77, 2, 171-190
LAB.SHOKLO-UNIT (2002). (Page consultée le 27 juillet 2004). Rapid Diagnostic test for malaria. [en ligne] Adresse: http://lab.shoklo-unit.com/pc/rapid_e.html
LABORATORY PRIMATE ADVOCACY GROUP (2002). (Page consultée le 12 décembre 2003). Biomed for the Layperson. [en ligne] Adresse: http://www.lpag.org/biomedForTheLayperson.html
LANDAU I., LEPERS J. P., RABETAFIKA L., BACCAM D., PETERS W., COULANGES P. (1989) Plasmodies de lémuriens malgaches. Ann Parasitol Hum Comp 64, 3, 171-184

157
LANFORD R. E., BIGGER C., BASSETT S., KLIMPEL G. (2001) The chimpanzee model of hepatitis C virus infections. Ilar J 42, 2, 117-126
LANG T., GREENWOOD B. (2003) The development of Lapdap, an affordable new treatment for malaria. Lancet Infect Dis 3, 3, 162-168
LAVSTSEN T., SALANTI A., JENSEN A. T., ARNOT D. E., THEANDER T. G. (2003) Sub-grouping of Plasmodium falciparum 3D7 var genes based on sequence analysis of coding and non-coding regions. Malar J 2, 1, 27
LE BRAS J., DURAND R. (2003) The mechanisms of resistance to antimalarial drugs in Plasmodium falciparum. Fundam Clin Pharmacol 17, 2, 147-153
LEMA O. E., CARTER J. Y., NAGELKERKE N., WANGAI M. W., KITENGE P., GIKUNDA S. M., ARUBE P. A., MUNAFU C. G., MATERU S. F., ADHIAMBO C. A., MUKUNZA H. K. (1999) Comparison of five methods of malaria detection in the outpatient setting. Am J Trop Med Hyg 60, 2, 177-182
LEPERS J. P., RABETAFIKA L., LANDAU I., PETERS W. (1989) Une nouvelle espèce plasmodiale chez un lémurien Plasmodium coulangesi. Ann Parasitol Hum Comp 64, 3, 163-170
LIBRARY OF CONGRESS (2000). (Page consultée le 3 novembre 2003). Rats/Mice/and Birds Database : researchers, Breeders, Transporters and Exhibiters. [en ligne] Adresse: http://aphis.usda.gov/ac/locreport.html
LIDEN J., NANTULYA V. M. (2004) The Global Fund and effective malaria treatment. Lancet 363, 9418, 1396
LOPANSRI B. K., ANSTEY N. M., WEINBERG J. B., STODDARD G. J., HOBBS M. R., LEVESQUE M. C., MWAIKAMBO E. D., GRANGER D. L. (2003) Low plasma arginine concentrations in children with cerebral malaria and decreased nitric oxide production. Lancet 361, 9358, 676-678
LUKE T. C., HOFFMAN S. L. (2003) Rationale and plans for developing a non-replicating, metabolically active, radiation-attenuated Plasmodium falciparum sporozoite vaccine. J Exp Biol 206, Pt 21, 3803-3808
MALKIN E. M., DURBIN A. P., DIEMERT D. J., SATTABONGKOT J., WU Y., MIURA K., LONG C. A., LAMBERT L., MILES A. P., WANG J., STOWERS A., MILLER L. H., SAUL A. (2005) Phase 1 vaccine trial of Pvs25H: a transmission blocking vaccine for Plasmodium vivax malaria. Vaccine 23, 24, 3131-3138
MANGOLD K. A., MANSON R. U., KOAY E. S., STEPHENS L., REGNER M., THOMSON R. B., JR., PETERSON L. R., KAUL K. L. (2005) Real-time PCR for detection and identification of Plasmodium spp. J Clin Microbiol 43, 5, 2435-2440

158
MARCH J. B. (2004) The Viability of DNA Vaccines - marcus evans Conference. IDrugs 7, 3, 226-228
MARSH K. (1998) Malaria disaster in Africa. Lancet 352, 9132, 924
MARTIN L. N. (1982) Chromatographic fractionation of rhesus monkey (Macaca mulatta) IgG subclasses using deae cellulose and protein A-sepharose. J Immunol Methods 50, 3, 319-329
MARX A., PEWSNER D., EGGER M., NUESCH R., BUCHER H. C., GENTON B., HATZ C., JUNI P. (2005) Meta-analysis: accuracy of rapid tests for malaria in travelers returning from endemic areas. Ann Intern Med 142, 10, 836-846
MATSUMOTO J., KAWAI S., TERAO K., KIRINOKI M., YASUTOMI Y., AIKAWA M., MATSUDA H. (2000) Malaria infection induces rapid elevation of the soluble Fas ligand level in serum and subsequent T lymphocytopenia: possible factors responsible for the differences in susceptibility of two species of Macaca monkeys to Plasmodium coatneyi infection. Infect Immun 68, 3, 1183-1188
MATUSCHEWSKI K., NUNES A. C., NUSSENZWEIG V., MENARD R. (2002) Plasmodium sporozoite invasion into insect and mammalian cells is directed by the same dual binding system. Embo J 21, 7, 1597-1606
MAY J., MEYER C. G. (2003) Chemoresistance in falciparum malaria. Trends Parasitol 19, 10, 432-435; discussion 435-436
MAYER M. (1907) Uber Malaria beim Affen. Med. Klin. 3, 579-580
MAYXAY M., PUKRITTAYAKAMEE S., NEWTON P. N., WHITE N. J. (2004) Mixed-species malaria infections in humans. Trends Parasitol 20, 5, 233-240
MAZIER D., COLLINS W. E., MELLOUK S., PROCELL P. M., BERBIGUIER N., CAMPBELL G. H., MILTGEN F., BERTOLOTTI R., LANGLOIS P., GENTILINI M. (1987) Plasmodium ovale: in vitro development of hepatic stages. Exp Parasitol 64, 3, 393-400
MEIS J. F., PONNUDURAI T., MONS B., VAN BELKUM A., VAN EERD P. M., DRUILHE P., SCHELLEKENS H. (1990) Plasmodium falciparum: studies on mature exoerythrocytic forms in the liver of the chimpanzee, Pan troglodytes. Exp Parasitol 70, 1, 1-11
MENARD R. (2005) Medicine: knockout malaria vaccine? Nature 433, 7022, 113-114
MENDIS K., SINA B. J., MARCHESINI P., CARTER R. (2001) The neglected burden of Plasmodium vivax malaria. Am J Trop Med Hyg 64, 1-2 Suppl, 97-106

159
MESNIL F., ROUBAUD E. (1920) Essais d'inoculation du paludisme au chimpanzé. Ann Inst Pasteur, Paris 34, 466-480
MICHEL J. C., BEHR C., MORALES-BETOULLE M. E., JOUIN H., MERCEREAU-PUIJALON O., CONTAMIN H. (2005) Blood typing in Saimiri sciureus monkeys: influence of anti-red blood cell alloantibodies on Plasmodium falciparum parasitaemia in vivo. Microbes Infect 7, 983-989
MIGOT-NABIAS F., OLLOMO B., DUBREUIL G., MORELLI A., DOMARLE O., NABIAS R., GEORGES A. J., MILLET P. (1999) Plasmodium coatneyi: differential clinical and immune responses of two populations of Macaca fascicularis from different origins. Exp Parasitol 91, 1, 30-39
MILEK R. L., KOCKEN C. H., KAAN A. M., JANSEN J., MEIJERS H., KONINGS R. N. (1997) Plasmodium reichenowi: deduced amino acid sequence of sexual stage-specific surface antigen Prs48/45 and comparison with its homologue in Plasmodium falciparum. Exp Parasitol 87, 2, 150-152
MILLER L. H., MASON S. J., CLYDE D. F., MCGINNISS M. H. (1976) The resistance factor to Plasmodium vivax in blacks. The Duffy-blood-group genotype, FyFy. N Engl J Med 295, 6, 302-304
MILLET P., NELSON C., GALLAND G. G., SULLIVAN J. S., MORRIS C. L., RICHARDSON B. B., COLLINS W. E. (1994) Plasmodium ovale: observations on the parasite development in Saimiri monkey hepatocytes in vivo and in vitro in contrast with its inability to induce parasitemia. Exp Parasitol 78, 4, 394-399
MITCHELL G. H., BUTCHER G. A., COHEN S. (1975) Merozoite vaccination against Plasmodium knowlesi malaria. Immunology 29, 2, 397-407
MOCKENHAUPT F. P., EHRHARDT S., GELLERT S., OTCHWEMAH R. N., DIETZ E., ANEMANA S. D., BIENZLE U. (2004) {alpha}+-thalassemia protects from severe malaria in African children. Blood 104, 7, 2003-2006
MOLYNEUX D., NANTULYA V. (2004) Linking disease control programmes in rural Africa: a pro-poor strategy to reach Abuja targets and millennium development goals. Bmj 328, 1129-1132
MOORTHY V. S., GOOD M. F., HILL A. V. (2004) Malaria vaccine developments. Lancet 363, 9403, 150-156
MOORTHY V. S., MCCONKEY S., ROBERTS M., GOTHARD P., ARULANANTHAM N., DEGANO P., SCHNEIDER J., HANNAN C., ROY M., GILBERT S. C., PETO T. E., HILL A. V. (2003a) Safety of DNA and modified vaccinia virus Ankara vaccines against liver-stage P. falciparum malaria in non-immune volunteers. Vaccine 21, 17-18, 1995-2002

160
MOORTHY V. S., PINDER M., REECE W. H., WATKINS K., ATABANI S., HANNAN C., BOJANG K., MCADAM K. P., SCHNEIDER J., GILBERT S., HILL A. V. (2003b) Safety and immunogenicity of DNA/modified vaccinia virus ankara malaria vaccination in African adults. J Infect Dis 188, 8, 1239-1244
MOREIRA L. A., WANG J., COLLINS F. H., JACOBS-LORENA M. (2004) Fitness of anopheline mosquitoes expressing transgenes that inhibit Plasmodium development. Genetics 166, 3, 1337-1341
MORRIS C. L., SULLIVAN J. S., MCCLURE H. M., STROBERT E., RICHARDSON B. B., GALLAND G. G., GOLDMAN I. F., COLLINS W. E. (1996) The Nigerian I/CDC strain of Plasmodium ovale in chimpanzees. J Parasitol 82, 3, 444-448
MU J., DUAN J., MAKOVA K. D., JOY D. A., HUYNH C. Q., BRANCH O. H., LI W. H., SU X. Z. (2002) Chromosome-wide SNPs reveal an ancient origin for Plasmodium falciparum. Nature 418, 6895, 323-326
MUCENSKI C. M., GUERRY P., BUESING M., SZARFMAN A., TROSPER J., WALLIKER D., WATT G., SANGALANG R., RANOA C. P., TUAZON M., ET AL. (1986) Evaluation of a synthetic oligonucleotide probe for diagnosis of Plasmodium falciparum infections. Am J Trop Med Hyg 35, 5, 912-920
MUEHLEN M., SCHREIBER J., EHRHARDT S., OTCHWEMAH R., JELINEK T., BIENZLE U., MOCKENHAUPT F. P. (2004) Short communication: Prevalence of mutations associated with resistance to atovaquone and to the antifolate effect of proguanil in Plasmodium falciparum isolates from northern Ghana. Trop Med Int Health 9, 3, 361-363
MUELLER A. K., LABAIED M., KAPPE S. H., MATUSCHEWSKI K. (2005) Genetically modified Plasmodium parasites as a protective experimental malaria vaccine. Nature 433, 7022, 164-167
MUTABINGWA T. K., ANTHONY D., HELLER A., HALLETT R., AHMED J., DRAKELEY C., GREENWOOD B. M., WHITTY C. J. (2005) Amodiaquine alone, amodiaquine+sulfadoxine-pyrimethamine, amodiaquine+artesunate, and artemether-lumefantrine for outpatient treatment of malaria in Tanzanian children: a four-arm randomised effectiveness trial. Lancet 365, 9469, 1474-1480
NARASIMHAN V., ATTARAN A. (2003) Roll back malaria? The scarcity of international aid for malaria control. Malar J 2, 1, 8
NARDIN E. H. (1990) T cell responses in a sporozoite-immunized human volunteer and a chimpanzee. Immunol Lett 25, 1-3, 43-47
NATIONAL CENTER FOR RESEARCH RESOURCES (2002). (Page consultée le 9 juin 2004). Rhesus Monkey demands in biomedical research. [en ligne] Adresse: www.ncrr.nih.gov/compmed/rhesusworkshopreport.pdf

161
NDONG J. M., ATTEKE C., AUBOUY A., BAKARY M., LEBIBI J., DELORON P. (2003) In vitro activity of chloroquine, quinine, mefloquine and halofantrine against Gabonese isolates of Plasmodium falciparum. Trop Med Int Health 8, 1, 25-29
NDYOMUGYENYI R., MAGNUSSEN P., CLARKE S. (2004) The efficacy of chloroquine, sulfadoxine-pyrimethamine and a combination of both for the treatment of uncomplicated Plasmodium falciparum malaria in an area of low transmission in western Uganda. Trop Med Int Health 9, 1, 47-52
NEZAMI A., KIMURA T., HIDAKA K., KISO A., LIU J., KISO Y., GOLDBERG D. E., FREIRE E. (2003) High-affinity inhibition of a family of Plasmodium falciparum proteases by a designed adaptive inhibitor. Biochemistry 42, 28, 8459-8464
NINO-VASQUEZ J. J., VOGEL D., RODRIGUEZ R., MORENO A., PATARROYO M. E., PLUSCHKE G., DAUBENBERGER C. A. (2000) Sequence and diversity of DRB genes of Aotus nancymaae, a primate model for human malaria parasites. Immunogenetics 51, 3, 219-230
NOLAND G. S., BRIONES N., SULLIVAN D. J., JR. (2003) The shape and size of hemozoin crystals distinguishes diverse Plasmodium species. Mol Biochem Parasitol 130, 2, 91-99
NOSTEN F., VAN VUGT M., PRICE R., LUXEMBURGER C., THWAY K. L., BROCKMAN A., MCGREADY R., TER KUILE F., LOOAREESUWAN S., WHITE N. J. (2000) Effects of artesunate-mefloquine combination on incidence of Plasmodium falciparum malaria and mefloquine resistance in western Thailand: a prospective study. Lancet 356, 9226, 297-302
NOYA O., PATARROYO M. E., GUZMAN F., ALARCON DE NOYA B. (2003) Immunodiagnosis of parasitic diseases with synthetic peptides. Curr Protein Pept Sci 4, 4, 299-308
NUSSENZWEIG R. S., VANDERBERG J., MOST H., ORTON C. (1967) Protective immunity produced by the injection of x-irradiated sporozoites of plasmodium berghei. Nature 216, 111, 160-162
NUSSENZWEIG R. S., VANDERBERG J., SPITALNY G. L., RIVERA C. I., ORTON C., MOST H. (1972) Sporozoite-induced immunity in mammalian malaria. A review. Am J Trop Med Hyg 21, 5, 722-728
OBER B. T., BRUHL P., SCHMIDT M., WIESER V., GRITSCHENBERGER W., COULIBALY S., SAVIDIS-DACHO H., GERENCER M., FALKNER F. G. (2002) Immunogenicity and safety of defective vaccinia virus lister: comparison with modified vaccinia virus Ankara. J Virol 76, 15, 7713-7723
OKENU D. M., THOMAS A. W., CONWAY D. J. (2000) Allelic lineages of the merozoite surface protein 3 gene in Plasmodium reichenowi and Plasmodium falciparum. Mol Biochem Parasitol 109, 2, 185-188

162
OKETCH-RABAH H., MWANGI J. (1998) La Médecine traditionnelle et les plantes médicinales ont-elles une place dans la lutte antipaludique ? In: Malaria. 3ème conférence panafricaine sur le paludisme, Nairobi, 21-26 juin 1998, sujet 15
OMARI A. A., GAMBLE C., GARNER P. (2004) Artemether-lumefantrine for uncomplicated malaria: a systematic review. Trop Med Int Health 9, 2, 192-199
O'NEIL R. H., LILIEN R. H., DONALD B. R., STROUD R. M., ANDERSON A. C. (2003) Phylogenetic classification of protozoa based on the structure of the linker domain in the bifunctional enzyme, dihydrofolate reductase-thymidylate synthase. J Biol Chem 278, 52, 52980-52987
ORGANISATION MONDIALE DE LA SANTÉ (2002). (Page consultée le 21 mai 2004). Projet de budget et programme pour l'exercice 2004-2005. [en ligne] Adresse: http://www.who.int/gb/ebwha/pdf_files/EB111/fppb.pdf
ORGANISATION MONDIALE DE LA SANTÉ (2003). (Page consultée le 21 août 2003). voyages internationaux et santé, situation au 1er janvier 2003. [en ligne] Adresse: http://whqlibdoc.who.int/publications/2003/9242580287.pdf
ORGANISATION MONDIALE DE LA SANTÉ (2005). (Page consultée le 14 juillet 2005). World Malaria Report. [en ligne] Adresse: http://rbm.who.int/wmr2005/
OZWARA H., LANGERMANS J. A., KOCKEN C. H., VAN DER WEL A., VAN DER MEIDE P. H., VERVENNE R. A., MWENDA J. M., THOMAS A. W. (2003a) Transfected Plasmodium knowlesi produces bioactive host gamma interferon: a new perspective for modulating immune responses to malaria parasites. Infect Immun 71, 8, 4375-4381
OZWARA H., LANGERMANS J. A., MAAMUN J., FARAH I. O., YOLE D. S., MWENDA J. M., WEILER H., THOMAS A. W. (2003b) Experimental infection of the olive baboon (Paplio anubis) with Plasmodium knowlesi: severe disease accompanied by cerebral involvement. Am J Trop Med Hyg 69, 2, 188-194
PAN W., HUANG D., ZHANG Q., QU L., ZHANG D., ZHANG X., XUE X., QIAN F. (2004) Fusion of two malaria vaccine candidate antigens enhances product yield, immunogenicity, and antibody-mediated inhibition of parasite growth in vitro. J Immunol 172, 10, 6167-6174
PASVOL G., WEATHERALL D. J., WILSON R. J. (1978) Cellular mechanism for the protective effect of haemoglobin S against P. falciparum malaria. Nature 274, 5672, 701-703
PATEL S. N., SERGHIDES L., SMITH T. G., FEBBRAIO M., SILVERSTEIN R. L., KURTZ T. W., PRAVENEC M., KAIN K. C. (2004) CD36 mediates the phagocytosis of Plasmodium falciparum-infected erythrocytes by rodent macrophages. J Infect Dis 189, 2, 204-213

163
PATHIRANA S. L., ALLES H. K., BANDARA S., PHONE-KYAW M., PERERA M. K., WICKREMASINGHE A. R., MENDIS K. N., HANDUNNETTI S. M. (2005) ABO-blood-group types and protection against severe, Plasmodium falciparum malaria. Ann Trop Med Parasitol 99, 2, 119-124
PENNISI E. (2003) Evolution. Chimp genome draft online. Science 302, 5652, 1876
PERANDIN F., MANCA N., CALDERARO A., PICCOLO G., GALATI L., RICCI L., MEDICI M. C., ARCANGELETTI M. C., SNOUNOU G., DETTORI G., CHEZZI C. (2004) Development of a real-time PCR assay for detection of Plasmodium falciparum, Plasmodium vivax, and Plasmodium ovale for routine clinical diagnosis. J Clin Microbiol 42, 3, 1214-1219
PERKINS S. L. (2000) Species concepts and malaria parasites: detecting a cryptic species of Plasmodium. Proc R Soc Lond B Biol Sci 267, 1459, 2345-2350
PERLAZA B. L., ZAPATA C., VALENCIA A. Z., HURTADO S., QUINTERO G., SAUZET J. P., BRAHIMI K., BLANC C., AREVALO-HERRERA M., DRUILHE P., HERRERA S. (2003) Immunogenicity and protective efficacy of Plasmodium falciparum liver-stage Ag-3 in Aotus lemurinus griseimembra monkeys. Eur J Immunol 33, 5, 1321-1327
PERRAUT R., GERRAUD O. (1998) [Vaccination trials and immune response against Plasmodium falciparum in Saimiri monkeys]. Med Trop (Mars) 58, 1, 76-84
PERRAUT R., MORALES-BETOULLE S., LE SCANF C., BOURREAU E., GUILLOTTE M., BONNEFOY S., MICHEL J., MERCEREAU-PUIJALON O. (2000) Evaluation of immunogenicity and protective efficacy of carrier-free Plasmodium falciparum R23 antigen in pre-exposed saimiri sciureus monkeys. Vaccine 19, 1, 59-67
PETERS W. (1976) Malaria of the orang-utan (Pongo pygmaeus) in Borneo. Philos Trans R Soc Lond B Biol Sci 275, 941, 439-482
PETERS W., ROBINSON B. L., ELLIS D. S. (1987) The chemotherapy of rodent malaria. XLII. Halofantrine and halofantrine resistance. Ann Trop Med Parasitol 81, 5, 639-646
PICO DE COANA Y., RODRIGUEZ J., GUERRERO E., BARRERO C., RODRIGUEZ R., MENDOZA M., PATARROYO M. A. (2003) A highly infective Plasmodium vivax strain adapted to Aotus monkeys: quantitative haematological and molecular determinations useful for P. vivax malaria vaccine development. Vaccine 21, 25-26, 3930-3937
PINDER M., REECE W. H., PLEBANSKI M., AKINWUNMI P., FLANAGAN K. L., LEE E. A., DOHERTY T., MILLIGAN P., JAYE A., TORNIEPORTH N., BALLOU R., MCADAM K. P., COHEN J., HILL A. V. (2004) Cellular immunity induced by the recombinant Plasmodium falciparum malaria vaccine, RTS,S/AS02, in semi-immune adults in The Gambia. Clin Exp Immunol 135, 2, 286-293

164
PINGARD A. M. (1992) Le saïmiri : élevage et modèle d'étude du paludisme Thèse de Doctorat vétérinaire, Université Claude Bernard Lyon I, Lyon, 164 pp
PIZZI E., FRONTALI C. (1999) Molecular evolution of coding and non-coding regions in Plasmodium. Parassitologia 41, 1-3, 89-91
PLANSON A. G., GUIJARRO J. I., GOLDBERG M. E., CHAFFOTTE A. F. (2003) Assistance of maltose binding protein to the in vivo folding of the disulfide-rich C-terminal fragment from Plasmodium falciparum merozoite surface protein 1 expressed in Escherichia coli. Biochemistry 42, 45, 13202-13211
PONNUDURAI T., LENSEN A. H., VAN GEMERT G. J., BOLMER M., VAN BELKUM A., VAN EERD P., MONS B. (1990) Large-scale production of Plasmodium vivax sporozoites. Parasitology 101 Pt 3, 317-320
PORTER J. A., JR., YOUNG M. D. (1966) Susceptibility of Panamanian primates to Plasmodium vivax. Mil Med 131, 9, Suppl:952-958
PORTER J. A., JR., YOUNG M. D. (1967) The transfer of Plasmodium falciparum from man to the marmoset, Saguinus geoffroyi. J Parasitol 53, 4, 845-846
POVOA M. M., CONN J. E., SCHLICHTING C. D., AMARAL J. C., SEGURA M. N., DA SILVA A. N., DOS SANTOS C. C., LACERDA R. N., DE SOUZA R. T., GALIZA D., SANTA ROSA E. P., WIRTZ R. A. (2003) Malaria vectors, epidemiology, and the re-emergence of Anopheles darlingi in Belem, Para, Brazil. J Med Entomol 40, 4, 379-386
PRABA-EGGE A. D., MONTENEGRO S., COGSWELL F. B., HOPPER T., JAMES M. A. (2002) Cytokine responses during acute simian Plasmodium cynomolgi and Plasmodium knowlesi infections. Am J Trop Med Hyg 67, 6, 586-596
PRADINES B., TALL A., ROGIER C., SPIEGEL A., MOSNIER J., MARRAMA L., FUSAI T., MILLET P., PANCONI E., TRAPE J. F., PARZY D. (2002) In vitro activities of ferrochloroquine against 55 Senegalese isolates of Plasmodium falciparum in comparison with those of standard antimalarial drugs. Trop Med Int Health 7, 3, 265-270
PRADINES B., VIAL H., OLLIARO P. (2003) [Malaria prophylaxis and treatment: problems, recent developments and perspectives]. Med Trop (Mars) 63, 1, 79-98
PUKRITTAYAKAMEE S., IMWONG M., LOOAREESUWAN S., WHITE N. J. (2004) Therapeutic responses to antimalarial and antibacterial drugs in vivax malaria. Acta Trop 89, 3, 351-356
PURI S. K., DUTTA G. P. (2003) Blood schizontocidal activity of WR 238605 (Tafenoquine) against Plasmodium cynomolgi and Plasmodium fragile infections in rhesus monkeys. Acta Trop 86, 1, 35-40

165
PURI S. K., SINGH N. (2000) Azithromycin: antimalarial profile against blood- and sporozoite-induced infections in mice and monkeys. Exp Parasitol 94, 1, 8-14
PYE D., EDWARDS S. J., ANDERS R. F., O'BRIEN C. M., FRANCHINA P., CORCORAN L. N., MONGER C., PETERSON M. G., VANDENBERG K. L., SMYTHE J. A., ET AL. (1991) Failure of recombinant vaccinia viruses expressing Plasmodium falciparum antigens to protect Saimiri monkeys against malaria. Infect Immun 59, 7, 2403-2411
QARI S. H., SHI Y. P., POVOA M. M., ALPERS M. P., DELORON P., MURPHY G. S., HARJOSUWARNO S., LAL A. A. (1993) Global occurrence of Plasmodium vivax-like human malaria parasite. J Infect Dis 168, 6, 1485-1489
RAMHARTER M., WERNSDORFER W. H., KREMSNER P. G. (2004) In vitro activity of quinolines against Plasmodium falciparum in Gabon. Acta Trop 90, 1, 55-60
RATH K., TAXIS K., WALZ G., GLEITER C. H., LI S. M., HEIDE L. (2004) Pharmacokinetic study of artemisinin after oral intake of a traditional preparation of Artemisia annua L. (annual wormwood). Am J Trop Med Hyg 70, 2, 128-132
REINHARDT V. (2003) Working with rather than against macaques during blood collection. J Appl Anim Welf Sci 6, 3, 189-197
RICH S. M., AYALA F. J. (2000) Population structure and recent evolution of Plasmodium falciparum. Proc Natl Acad Sci U S A 97, 13, 6994-7001
RICH S. M., AYALA F. J. (2003) Progress in malaria research: the case for phylogenetics. Adv Parasitol 54, 255-280
RICH S. M., LICHT M. C., HUDSON R. R., AYALA F. J. (1998) Malaria's Eve: evidence of a recent population bottleneck throughout the world populations of Plasmodium falciparum. Proc Natl Acad Sci U S A 95, 8, 4425-4430
RIECKMANN K. H., BEAUDOIN R. L., CASSELLS J. S., SELL K. W. (1979) Use of attenuated sporozoites in the immunization of human volunteers against falciparum malaria. Bull World Health Organ 57 Suppl 1, 261-265
RIECKMANN K. H., DAVIS D. R., HUTTON D. C. (1989) Plasmodium vivax resistance to chloroquine? Lancet 2, 8673, 1183-1184
RIECKMANN K. H., MREMA J. E., MARSHALL P. H., HAFNER D. M. (1980) Breeding of Aotus monkeys for human malaria research. Bull Pan Am Health Organ 14, 3, 251-257
ROBERTS D. J. (2003) Understanding naturally acquired immunity to Plasmodium falciparum malaria. Infect Immun 71, 2, 589-590

166
RODHAIN J. (1939) Les plasmodiums des anthropoïdes de l'Afrique centrale et leurs relations avec les plasmodiums humains. Ann Soc Belg Med Trop 19, 563-572
RODHAIN J. (1941) Sur un Plasmodium du gibbon Hylobates lensciscus. Acta. Biol. Belg. 1, 118-123
RODHAIN J. (1948) Susceptibility of the chimpanzee to P. malariae of human origin. Am J Trop Med Hyg 28, 629-631
RODHAIN J., DELLAERT R. (1955a) [Studies on Plasmodium schwetzi E. Brumpt. III. Plasmodium schwetzi infection in humans.]. Ann Soc Belg Med Trop 35, 6, 757-775
RODHAIN J., DELLAERT R. (1955b) [Study of Plasmodium schwetzi E. Brumpt. II. Transmission of Plasmodium schwetzi to man.]. Ann Soc Belg Med Trop 35, 1, 73-76
ROLLAND R. M., HAUSFATER G., MARSHALL B., LEVY S. B. (1985) Antibiotic-resistant bacteria in wild primates: increased prevalence in baboons feeding on human refuse. Appl Environ Microbiol 49, 4, 791-794
ROSIER F., MAZIER D., LAVARDE V. (1999). (Page consultée le 2 février 2004). Qu'est-ce qu'un état de prémunition ? [en ligne] Adresse: http://www.frm.org/informez/info_ressources_dossiers_article_chapitre.php?id=13&type=10&numeroChapitre=6&act=
RUEBUSH T. K., 2ND, ZEGARRA J., CAIRO J., ANDERSEN E. M., GREEN M., PILLAI D. R., MARQUINO W., HUILCA M., AREVALO E., GARCIA C., SOLARY L., KAIN K. C. (2003) Chloroquine-resistant Plasmodium vivax malaria in Peru. Am J Trop Med Hyg 69, 5, 548-552
RUSSELL W. M. S., BURCH R. L. (1959) The principles of humane experimental technique Methuen, London, 238 pp.
RUTJENS E., BALLA-JHAGJHOORSINGH S., VERSCHOOR E., BOGERS W., KOOPMAN G., HEENEY J. (2003) Lentivirus infections and mechanisms of disease resistance in chimpanzees. Front Biosci 8, d1134-1145
SABCHAREON A., BURNOUF T., OUATTARA D., ATTANATH P., BOUHAROUN-TAYOUN H., CHANTAVANICH P., FOUCAULT C., CHONGSUPHAJAISIDDHI T., DRUILHE P. (1991) Parasitologic and clinical human response to immunoglobulin administration in falciparum malaria. Am J Trop Med Hyg 45, 3, 297-308
SACHS J., MALANEY P. (2002) The economic and social burden of malaria. Nature 415, 6872, 680-685

167
SAHR F., WILLOUGHBY V. R., GBAKIMA A. A., BOCKARIE M. J. (2001) Apparent drug failure following artesunate treatment of Plasmodium falciparum malaria in Freetown, Sierra Leone: four case reports. Ann Trop Med Parasitol 95, 5, 445-449
SAISSY J. M., ROUVIN B., KOULMANN P. (2003) [Severe malaria in intensive care units in 2003]. Med Trop (Mars) 63, 3, 258-266
SATAYAVIVAD J., WATCHARASIT P., KHAMKONG P., TUNTAWIROON J., PAVARO C., RUCHIRAWAT S. (2004) The pharmacodynamic study of a potent new antimalarial (MC1). Acta Trop 89, 3, 343-349
SATTABONGKOT J., TSUBOI T., HISAEDA H., TACHIBANA M., SUWANABUN N., RUNGRUANG T., CAO Y. M., STOWERS A. W., SIRICHAISINTHOP J., COLEMAN R. E., TORII M. (2003) Blocking of transmission to mosquitoes by antibody to Plasmodium vivax malaria vaccine candidates Pvs25 and Pvs28 despite antigenic polymorphism in field isolates. Am J Trop Med Hyg 69, 5, 536-541
SCHUSTER F. L. (2002) Cultivation of plasmodium spp. Clin Microbiol Rev 15, 3, 355-364
SCOTT L., PEARCE P., FAIRHALL S., MUGGLETON N., SMITH J. (2003) Training nonhuman primates to cooperate with scientific procedures in applied biomedical research. J Appl Anim Welf Sci 6, 3, 199-207
SEDEGAH M., BELMONTE M., EPSTEIN J. E., SIEGRIST C. A., WEISS W. R., JONES T. R., LU M., CARUCCI D. J., HOFFMAN S. L. (2003) Successful induction of CD8 T cell-dependent protection against malaria by sequential immunization with DNA and recombinant poxvirus of neonatal mice born to immune mothers. J Immunol 171, 6, 3148-3153
SEDEGAH M., CHAROENVIT Y., MINH L., BELMONTE M., MAJAM V. F., ABOT S., GANESHAN H., KUMAR S., BACON D. J., STOWERS A., NARUM D. L., CARUCCI D. J., ROGERS W. O. (2004) Reduced immunogenicity of DNA vaccine plasmids in mixtures. Gene Ther 11, 5, 448-456
SEIN K. K., BROWN A. E., MAENO Y., SMITH C. D., CORCORAN K. D., HANSUKJARIYA P., WEBSTER H. K., AIKAWA M. (1993) Sequestration pattern of parasitized erythrocytes in cerebrum, mid-brain, and cerebellum of Plasmodium coatneyi-infected rhesus monkeys (Macaca mulatta). Am J Trop Med Hyg 49, 4, 513-519
SERGENT E., PARROT L. (1935) L'immunité, la prémunition et la résistance innée. Arch Inst Pasteur Alger 3, 279
SHARMA V. P. (1996) Re-emergence of malaria in India. Indian J Med Res 103, 26-45
SHEAR H. L. (1993) Transgenic and mutant animal models to study mechanisms of protection of red cell genetic defects against malaria. Experientia 49, 1, 37-42

168
SHI Y. L., LI G. F., ZHAO J. H., YANG J. D., DING D. B. (1999) Schizontocidal effects of oral artesunate on Plasmodium berghei in mice and P knowlesi in monkeys. Zhongguo Yao Li Xue Bao 20, 8, 755-758
SIDDIQUI W. A., SCHNELL J. V., GEIMAN Q. M. (1972) In vitro cultivation of plasmodium malariae. J Parasitol 58, 4, 804
SINDEN R. (1978) Cell Biology in: Killick-Kendrick R. and Peters W. Rodent Malaria. Academic Press, London, 85-168
SINDEN R. E., HARTLEY R. H. (1985) Identification of the meiotic division of malarial parasites. J Protozool 32, 4, 742-744
SINGH B., KIM SUNG L., MATUSOP A., RADHAKRISHNAN A., SHAMSUL S. S., COX-SINGH J., THOMAS A., CONWAY D. J. (2004) A large focus of naturally acquired Plasmodium knowlesi infections in human beings. Lancet 363, 9414, 1017-1024
SINGH N., SHUKLA M. (2001) An assessment of the usefulness of a rapid immuno-chromatographic test, "Determine trade mark malaria pf" in evaluation of intervention measures in forest villages of central India. BMC Infect Dis 1, 1, 10
SINTON J. A., MULLIGAN H. W. (1932-1933) A critical review of the literature relating to the identification of the malarial parasites recorded from monkeys of the family Cercopithecidae and Colobidae. Rec. Malar. Surv. India 3, 357-380, 381-444
SITALAKSHMI S., SRIKRISHNA A., DEVI S., DAMODAR P., MATHEW T., VARGHESE J. (2003) Changing trends in malaria--a decade's experience at a referral hospital. Indian J Pathol Microbiol 46, 3, 399-401
SITE PORTAIL DE LA GUYANE (2004). (Page consultée le 26 avril 2004). Paludisme. [en ligne] Adresse: http://www.guyane-guide.com/pages/paludisme.htm
SLUITER C., SWELLENGREBEL N., IHLE J. (1922) De Dierlijke parasiten van den Mensch en van Onze Huisdieren Scheltema & Holkema's Boekhaendel, Amsterdam, 416 pp.
SNOUNOU G., VIRIYAKOSOL S., ZHU X. P., JARRA W., PINHEIRO L., DO ROSARIO V. E., THAITHONG S., BROWN K. N. (1993) High sensitivity of detection of human malaria parasites by the use of nested polymerase chain reaction. Mol Biochem Parasitol 61, 2, 315-320
SNOW R. W., GUERRA C. A., NOOR A. M., MYINT H. Y., HAY S. I. (2005) The global distribution of clinical episodes of Plasmodium falciparum malaria. Nature 434, 7030, 214-217

169
STAALSOE T., SHULMAN C. E., BULMER J. N., KAWUONDO K., MARSH K., HVIID L. (2004) Variant surface antigen-specific IgG and protection against clinical consequences of pregnancy-associated Plasmodium falciparum malaria. Lancet 363, 9405, 283-289
STAUFFER W., FISCHER P. R. (2003) Diagnosis and treatment of malaria in children. Clin Infect Dis 37, 10, 1340-1348
STEVENSON M. M., RILEY E. M. (2004) Innate immunity to malaria. Nat Rev Immunol 4, 3, 169-180
STOWERS A. W., CHEN LH L. H., ZHANG Y., KENNEDY M. C., ZOU L., LAMBERT L., RICE T. J., KASLOW D. C., SAUL A., LONG C. A., MEADE H., MILLER L. H. (2002a) A recombinant vaccine expressed in the milk of transgenic mice protects Aotus monkeys from a lethal challenge with Plasmodium falciparum. Proc Natl Acad Sci U S A 99, 1, 339-344
STOWERS A. W., KENNEDY M. C., KEEGAN B. P., SAUL A., LONG C. A., MILLER L. H. (2002b) Vaccination of monkeys with recombinant Plasmodium falciparum apical membrane antigen 1 confers protection against blood-stage malaria. Infect Immun 70, 12, 6961-6967
STOWERS A. W., MILLER L. H. (2001) Are trials in New World monkeys on the critical path for blood-stage malaria vaccine development? Trends Parasitol 17, 9, 415-419
SULLIVAN J. S., MORRIS C. L., MCCLURE H. M., STROBERT E., RICHARDSON B. B., GALLAND G. G., GOLDMAN I. F., COLLINS W. E. (1996) Plasmodium vivax infections in chimpanzees for sporozoite challenge studies in monkeys. Am J Trop Med Hyg 55, 3, 344-349
SULLIVAN J. S., STROBERT E., YANG C., MORRIS C. L., GALLAND G. G., RICHARDSON B. B., BOUNNGASENG A., KENDALL J., MCCLURE H., COLLINS W. E. (2001) Adaptation of a strain of Plasmodium vivax from India to New World monkeys, chimpanzees, and anopheline mosquitoes. J Parasitol 87, 6, 1398-1403
SUPUTTAMONGKOL Y., CHINDARAT S., SILPASAKORN S., CHAIKACHONPATD S., LIM K., CHANTHAPAKAJEE K., KAEWKAUKUL N., THAMLIKITKUL V. (2003) The efficacy of combined mefloquine-artesunate versus mefloquine-primaquine on subsequent development of Plasmodium falciparum gametocytemia. Am J Trop Med Hyg 68, 5, 620-623
SYIN C., PARZY D., TRAINCARD F., BOCCACCIO I., JOSHI M. B., LIN D. T., YANG X. M., ASSEMAT K., DOERIG C., LANGSLEY G. (2001) The H89 cAMP-dependent protein kinase inhibitor blocks Plasmodium falciparum development in infected erythrocytes. Eur J Biochem 268, 18, 4842-4849
TALIAFERRO W. H., TALIAFERRO L. G. (1934) The transmission of Plasmodium falciparum to the howler monkey Alouatta sp. I. General nature of the infections and morphology of the parasites. Am J Hyg. 19, 318-334

170
TANABE K., SAKIHAMA N., KANEKO A. (2004) Stable SNPs in malaria antigen genes in isolated populations. Science 303, 5657, 493
TAYLOR D. W., WELLS R. A., VERNES A., ROSENBERG Y. J., VOGEL S., DIGGS C. L. (1985) Parasitologic and immunologic studies of experimental Plasmodium falciparum infection in nonsplenectomized chimpanzees (Pan troglodytes). Am J Trop Med Hyg 34, 1, 36-44
TAYLOR W. R., WHITE N. J. (2004) Antimalarial drug toxicity: a review. Drug Saf 27, 1, 25-61
THOMAS A. W., NARUM D., WATERS A. P., TRAPE J. F., ROGIER C., GONCALVES A., ROSARIO V., DRUILHE P., MITCHELL G. H., DENNIS D. (1994a) Aspects of immunity for the AMA-1 family of molecules in humans and non-human primates malarias. Mem Inst Oswaldo Cruz 89 Suppl 2, 67-70
THOMAS A. W., SLIERENDREGT B., MONS B., DRUILHE P. (1994b) Chimpanzees and supporting models in the study of malaria pre-erythrocytic stages. Mem Inst Oswaldo Cruz 89 Suppl 2, 111-114
THOMAS C. (2004) Malaria: a changed climate in Africa? Nature 427, 6976, 690-691
TOMAS A. M., MARGOS G., DIMOPOULOS G., VAN LIN L. H., DE KONING-WARD T. F., SINHA R., LUPETTI P., BEETSMA A. L., RODRIGUEZ M. C., KARRAS M., HAGER A., MENDOZA J., BUTCHER G. A., KAFATOS F., JANSE C. J., WATERS A. P., SINDEN R. E. (2001) P25 and P28 proteins of the malaria ookinete surface have multiple and partially redundant functions. Embo J 20, 15, 3975-3983
TONGREN J. E., YANG C., COLLINS W. E., SULLIVAN J. S., LAL A. A., XIAO L. (2000) Expression of proinflammatory cytokines in four regions of the brain in Macaque mulatta (rhesus) monkeys infected with Plasmodium coatneyi. Am J Trop Med Hyg 62, 4, 530-534
TRAGER W., JENSEN J. B. (1976) Human malaria parasites in continuous culture. Science 193, 4254, 673-675
TRAGER W., JENSEN J. B. (1997) Continuous culture of Plasmodium falciparum: its impact on malaria research. Int J Parasitol 27, 9, 989-1006
TSUBOI T., TACHIBANA M., KANEKO O., TORII M. (2003) Transmission-blocking vaccine of vivax malaria. Parasitol Int 52, 1, 1-11
TSUJI M., ZAVALA F. (2003) T cells as mediators of protective immunity against liver stages of Plasmodium. Trends Parasitol 19, 2, 88-93
TYAGI B. K. (2004) A review of the emergence of Plasmodium falciparum-dominated malaria in irrigated areas of the Thar Desert, India. Acta Trop 89, 2, 227-239

171
VAN AKKER R., BALLS M., EICHBERG J. W., GOODALL J., HEENEY J. L., OSTERHAUS A. D., PRINCE A. M., SPRUIT I. (1994) Chimpanzees in AIDS research: a biomedical and bioethical perspective. J Med Primatol 23, 1, 49-51
VAN DER MEERSCH H. (2005) [Review of the use of artemisinin and its derivatives in the treatment of malaria]. J Pharm Belg 60, 1, 23-29
VANDERJAGT T. A., IKEH E. I., UJAH I. O., BELMONTE J., GLEW R. H., VANDERJAGT D. J. (2005) Comparison of the OptiMAL rapid test and microscopy for detection of malaria in pregnant women in Nigeria. Trop Med Int Health 10, 1, 39-41
VEISSIER I. (1999) Expérimentation animale : biologie, éthique, réglementation. Prod. Anim. 12, 365-375
VERSCHOOR E. J., WARREN K. S., LANGENHUIJZEN S., HERIYANTO, SWAN R. A., HEENEY J. L. (2001) Analysis of two genomic variants of orang-utan hepadnavirus and their relationship to other primate hepatitis B-like viruses. J Gen Virol 82, Pt 4, 893-897
VINKENOOG R., VELDHUISEN B., SPERANCA M. A., DEL PORTILLO H. A., JANSE C., WATERS A. P. (1995) Comparison of introns in a cdc2-homologous gene within a number of Plasmodium species. Mol Biochem Parasitol 71, 2, 233-241
VOGEL G. (2000) Laboratory animals. Congress OKs plan for retired chimps. Science 290, 5500, 2233
VOLKER C., BROWN J. R. (2002) Bioinformatics and the discovery of novel anti-microbial targets. Curr Drug Targets Infect Disord 2, 4, 279-290
VOLNEY B., POULIQUEN J. F., DE THOISY B., FANDEUR T. (2002) A sero-epidemiological study of malaria in human and monkey populations in French Guiana. Acta Trop 82, 1, 11-23
WALCOURT A., LOYEVSKY M., LOVEJOY D. B., GORDEUK V. R., RICHARDSON D. R. (2004) Novel aroylhydrazone and thiosemicarbazone iron chelators with anti-malarial activity against chloroquine-resistant and -sensitive parasites. Int J Biochem Cell Biol 36, 3, 401-407
WALLIKER D. (1983) The genetic basis of diversity in malaria parasites. Adv Parasitol 22, 217-259
WALLIKER D., CARTER R., MORGAN S. (1971) Genetic recombination in malaria parasites. Nature 232, 5312, 561-562

172
WANG R., RICHIE T. L., BARACEROS M. F., RAHARDJO N., GAY T., BANANIA J. G., CHAROENVIT Y., EPSTEIN J. E., LUKE T., FREILICH D. A., NORMAN J., HOFFMAN S. L. (2005) Boosting of DNA vaccine-elicited gamma interferon responses in humans by exposure to malaria parasites. Infect Immun 73, 5, 2863-2872
WARD R. A., MORRIS J. H., GOULD D. J., BOURKE A. T., CADIGAN F. C., JR. (1965) Susceptibility of the gibbon Hylobates lar to falciparum malaria. Science 150, 703, 1604-1605
WARREN M., BENNETT G. F., SANDOSHAN A. A., COATNEY G. R. (1965) Plasmodium eylesi sp. nov., a tertian malaria parasite from the white-handed gibbon, Hylobates lar. Ann. Trop. Med. Parasitol. 59, 500-508
WARREN M., COATNEY G. R., SKINNER J. C. (1966) Plasmodium jefferyi sp. n. from Hylobates lar in Malaya. J. Parasitol. 52, 9-13
WARSHAUER D. M. (1985) Primacy among the primates: killing chimpanzees to help humans. Jama 254, 3, 356
WASSMER S. C., COLTEL N., COMBES V., GRAU G. E. (2003a) [Pathogenesis of cerebral malaria: facts and hypotheses]. Med Trop (Mars) 63, 3, 254-257
WASSMER S. C., COMBES V., GRAU G. E. (2003b) Pathophysiology of cerebral malaria: role of host cells in the modulation of cytoadhesion. Ann N Y Acad Sci 992, 30-38
WASSMER S. C., LEPOLARD C., TRAORE B., POUVELLE B., GYSIN J., GRAU G. E. (2004) Platelets reorient Plasmodium falciparum-infected erythrocyte cytoadhesion to activated endothelial cells. J Infect Dis 189, 2, 180-189
WATERS A. P., MOTA M. M., VAN DIJK M. R., JANSE C. J. (2005) Parasitology. Malaria vaccines: back to the future? Science 307, 5709, 528-530
WATKINS W. M., SIBLEY C. H., HASTINGS I. M. (2005) The search for effective and sustainable treatments for Plasmodium falciparum malaria in Africa: a model of the selection of resistance by antifolate drugs and their combinations. Am J Trop Med Hyg 72, 2, 163-173
WEBSTER D., HILL A. V. (2003) Progress with new malaria vaccines. Bull World Health Organ 81, 12, 902-909
WENGELNIK K., VIDAL V., ANCELIN M. L., CATHIARD A. M., MORGAT J. L., KOCKEN C. H., CALAS M., HERRERA S., THOMAS A. W., VIAL H. J. (2002) A class of potent antimalarials and their specific accumulation in infected erythrocytes. Science 295, 5558, 1311-1314
WHITE N. J. (2004) Antimalarial drug resistance. the journal of clinical investigation 113, 8, 1084-1092

173
WHITE N. J., PONGTANORNPINYO W. (2003) The de-novo selection of drug resistance in malaria parasites. Philos. Trans. R. Soc. Lond. B Biol. Sci. 270, 545-554
WHITTY C., ALLAN R., WISEMAN V., OCHOLA S., NAKYANZI-MUGISHA M. V., VONHOM B., MWITA M., MIAKA C., OLOO A., PREMJ Z., BURGESS C., MUTABINGWA T. (2004) Averting a malaria disaster en Africa - where does the buck stop ? Bulletin of the World Health Organization 82, 5, 381-384
WILLIAMS T. N., MAITLAND K., BENNETT S., GANCZAKOWSKI M., PETO T. E., NEWBOLD C. I., BOWDEN D. K., WEATHERALL D. J., CLEGG J. B. (1996) High incidence of malaria in alpha-thalassaemic children. Nature 383, 6600, 522-525
WIPASA J., ELLIOTT S., XU H., GOOD M. F. (2002) Immunity to asexual blood stage malaria and vaccine approaches. Immunol Cell Biol 80, 5, 401-414
WOLFE N. D., ESCALANTE A. A., KARESH W. B., KILBOURN A., SPIELMAN A., LAL A. A. (1998) Wild primate populations in emerging infectious disease research: the missing link? Emerg Infect Dis 4, 2, 149-158
YOSHIDA S., KONDOH D., ARAI E., MATSUOKA H., SEKI C., TANAKA T., OKADA M., ISHII A. (2003) Baculovirus virions displaying Plasmodium berghei circumsporozoite protein protect mice against malaria sporozoite infection. Virology 316, 1, 161-170
YOUNG M. D., BAERG D. C. (1969a) Experimental infections of Plasmodium falciparum in Cebus capucinus (white faced capuchin) monkeys. Mil Med 134, 10, 767-771
YOUNG M. D., PORTER J. A., JR. (1969b) Susceptibility of Ateles fusciceps, Ateles geoffroyi and Cebus capucinus monkeys to Plasmodium vivax. Trans R Soc Trop Med Hyg 63, 2, 203-205
YOUNG M. D., ROSSAN R. N. (1969c) Plasmodium falciparum induced in the squirrel monkey, Saimiri sciureus. Trans R Soc Trop Med Hyg 63, 5, 686-687
ZAKAI H. A. (2003) Methods used in the diagnosis of malaria: where do we stand? J Egypt Soc Parasitol 33, 3, 979-990
ZAPATA J. C., PERLAZA B. L., HURTADO S., QUINTERO G. E., JURADO D., GONZALEZ I., DRUILHE P., AREVALO-HERRERA M., HERRERA S. (2002) Reproducible infection of intact Aotus lemurinus griseimembra monkeys by Plasmodium falciparum sporozoite inoculation. J Parasitol 88, 4, 723-729
ZEISS C. J., SHOMER N. (2001) Hepatocystosis in a baboon (Papio anubis). Contemp Top Lab Anim Sci 40, 1, 41-42

174
ZHENG C., XIE P., CHEN Y. (2003) Molecular cloning and sequencing of the merozoite surface antigen 2 gene from Plasmodium falciparum strain FCC-1/HN and expression of the gene in mycobacteria. J Eukaryot Microbiol 50, 2, 140-143
ZOU L., MILES A. P., WANG J., STOWERS A. W. (2003) Expression of malaria transmission-blocking vaccine antigen Pfs25 in Pichia pastoris for use in human clinical trials. Vaccine 21, 15, 1650-1657

175
ANNEXES
Annexe 1 : terminologie du paludisme Annexe 2 : liste des abréviations utilisées Annexe 3 : épidémiologie mondiale du paludisme Annexe 4 : stades sanguins de P. falciparum Annexe 5 : exemples de frottis sanguins humains Annexe 6 : liste des molécules antipaludiques et de quelques noms déposés Annexe 7 : document de saisine du comite d’éthique sur l’expérimentation animale Ile de France Sud Annexe 8 : amorces des 11 gènes utilises pour construire l’arbre de P. benjamini Annexe 9 : publications et participations à des congrès

176

177
ANNEXE 1 TERMINOLOGIE DU PALUDISME
AMA : antigène utilisé pour les vaccins contre les stades sanguins. Anophèle (Anopheles) : type de moustique dont la femelle est le vecteur du paludisme. Anthropozoonose : zoonose exclusivement transmise de l’animal à l’homme. Chimioprophylaxie : prévention d’une maladie par une substance chimique. Cryptozoïte : premier stade hépatique, lorsque le sporozoïte a pénétré dans un hépatocyte. CSP : protéine utilisée pour les vaccins contre les stades pré-érythrocytaires. Dixène : existence de 2 hôtes pour un même parasite. Erythrocytaire (stade) : stade sanguin qui s’oppose aux stades dans le foie dits exo-érythrocytaires ou pré-érythrocytaires. Gamète : cellule sexuelle mâle (microgamète) ou femelle (macrogamète) qui fusionne chez le moustique pour donner un ookinète. Gamétocyte : stade sanguin, ingéré par le moustique, qui aboutira à la production des gamètes (microgamétocyte mâle et macrogamétocyte femelle) Hématozoaire : parasite unicellulaire vivant dans le sang. Hôte définitif : hôte au sein duquel se déroule la reproduction sexuée du parasite (anophèle). Hôte intermédiaire : hôte au sein duquel ne se déroule pas la reproduction sexuée du parasite (par exemple l’homme pour P. falciparum). Hypnozoïte : stade hépatique latent responsable de rechutes. LSA : antigène utilisé pour les vaccins contre les stades exo-érythrocytaires. Macrogamète : voir gamète. Macrogamétocyte : voir gamétocyte. Mérozoïte : stade chez l’hôte intermédiaire, érythrocytaire, libre, issu de la schizogonie. Microgamète : voir gamète. Microgamétocyte : voir gamétocyte. MSP : protéine utilisée pour les vaccins contre les stades érythrocytaires. Neuropaludisme : forme mortelle de la malaria due à P. falciparum. Oocyste : stade parasitaire situé dans la paroi de l’estomac du moustique, dérivé du zygote, et donnant naissance aux sporozoïtes. Ookinète : oeuf mobile résultant de la fécondation. Paludisme bénin : forme classique du paludisme avec fièvre et divers symptômes sans pronostic vital. Paludisme grave ou pernicieux : neuropaludisme et/ou anémie sévère selon le malade. Paludisme viscéral : type chronique avec fièvre, splénomégalie, amaigrissement et anémie. Palustre : en rapport avec le paludisme. Périodicité ou fièvre quarte : alternance de fièvres liées au cycle sanguin du parasite en 72 heures. Périodicité ou fièvre tierce : alternance de fièvres liées au cycle sanguin du parasite en 48 heures. Plasmodie = plasmodium. Plasmodium (P. falciparum, P. vivax, P. ovale, P. malariae chez l’homme) : parasite véhiculant le paludisme. Prémunition (état de) : fragile protection, longuement acquise, contre les symptômes du paludisme. Rechute : poussée de parasitémie à partir d'hypnozoïtes hépatiques dans le cas de P. vivax et de P. ovale.

178
Recrudescence : augmentation de la parasitémie jusqu'à un niveau décelable cliniquement chez un sujet après traitement incomplet ou inefficace, ou chez un sujet semi-immun. Ré-Infection : nouvelle piqûre d'anophèle infectée. SALSA : antigène utilisé pour les vaccins contre les stades exo-érythrocytaires. Schizogonie : chez l’hôte intermédiaire, phase de multiplication asexuée du parasite qui consiste en une division des noyaux (dans le foie et dans le sang). Schizonte : stade hépatique ou sanguin abritant la schizogonie. Splénectomie : exérèse de la rate qui permet d’augmenter la susceptibilité de l’hôte au parasite. Splénomégalie : augmentation du volume de la rate. Sporogonie : multiplication asexuée des sporozoïtes dans l’oocyste chez l’anophèle. Sporozoïte : stade issu des oocystes qui, dans les glandes salivaires du moustique, sera injecté à l’hôte intermédiaire. STARP : protéine utilisée pour les vaccins contre les stades epré-érythrocytaires. Trophozoïte : stade sanguin intra-érythrocytaire issu de la pénétration d’un mérozoïte dans un globule rouge de l’hôte intermédiaire. Zoonose : maladie des animaux (en dehors de l'homme) qui peut être transmise à l'homme ou qui peut être transmise de l'homme aux animaux. Zygote : cellule formée chez l’anophèle par la fusion des cellules sexuelles mâle et femelle avant que les cellules fusionnées ne commencent à se diviser.

179
ANNEXE 2 LISTE DES ABREVIATIONS UTILISEES
ABRA : Acidic-Basic Repeat Antigen Ac : Anticorps ADCC : Antibody-Dependant Cellular Cytotoxicity ADCI : Antibody-Dependent Cellular Inhibition Ag : Antigène AMA : Apical Membrane Antigen AMM : Autorisation de Mise sur le Marché BEH : Bulletin Epidemiologique Hebdomadaire BPRC : Biomedical Primate Research Center CCPA : Conseil Canadien de Protection des Animaux CIRMF : Centre International de Recherches Médicales de Franceville CITES : Convention on International Trade in Endangered Species CMH : Complexe Majeur d’Histocompatibilité (voir HLA) CNREPIA : Centre National de Référence de l'Épidémiologie du Paludisme d'Importation et Autochtone CNRS : Centre National de Recherches Scientifiques CSP : CircumSporozoite Protein CTL : Cytotoxic T Lymphocyte DBP : Duffy-Binding Protein EBA : Erythrocyte-Binding Antigen ELISA : Enzyme Linked Immunosorbent Assay EMP : Erythrocyte Membrane Protein G6PDH : Glucose-6-Phosphate DesHydrogénase GLURP : Glutamate-Rich Protein GPI : GlycosilPhosphatidylInositol HIV : Human Immunodeficience Virus HLA : Human Leucocyte Antigen HRP2 : Histidine-Rich Protein-2 IFI : ImmunoFluorescence Indirecte IFN : InterFéroN Ig : Immunoglobuline IUCN : International Union for Conservation of Nature and Natural Resources LDH : Lactate DesHydrogénase LPAG : Laboratory Primate Advocacy Group LSA : Liver Stage Antigen MSP : Merozoite Surface Protein NK : Natural Killer ODN : OligoDéoxyNucleotides OMS : Organisation Mondiale de la Santé PBS : Phosphate Buffer Saline PCR : Polymerase Chain Reaction PNH : Primate Non-Humain QBC : Quantitative Buffy Coat RAM: Recombinant Antibody anti -MSP-3 RAP : Rhoptry-Associated Protein RESA : Ring Erythrocyte Surface Antigen

180
SALSA : Sporozoïte And Liver-Stage Antigen SCID : Severe Combined ImmunoDeficiency SERP : Serine-Rich Protein SIDA : Syndrome d’ImmunoDéficience Acquise SIV : Simian Immunodeficience Virus SNP : Single Nucleotide Polymorphism STARP : Sporozoïtes Threonine and Asparagines-Rich Protein STLV : Simian T-cell Leukemia Virus TNF : Tumor Necrosis Factor TRAP : Thrombospondin-Relatited Adhesive Protein USDA : United States Department of Agriculture VIH : Virus d’Immunodéficience Humaine WHO : World Health Organisation

181
ANNEXE 3 EPIDEMIOLOGIE MONDIALE
DU PALUDISME
Répartition mondiale des régions en fonction de leur risque paludique Adapté d’après l’Annexe du Bulletin Epidémiologique Hebdomadaire
(Institut de veille sanitaire, 2003) Classement des régions mondiales en quatre groupes:
Groupe 0 : zones sans paludisme Groupe 1 : zones sans chloroquinolorésistance Groupe 2 : zones de chloroquinolorésistance Groupe 3 : zones de prévalence élevée de chloroquinolorésistance ou multirésistance
REGION GROUPE 0 GROUPE 1 GROUPE 2 GROUPE 3
AFRIQUE
Ïle de la Réunion Île Sainte Hélène Lesotho Libye Seychelles Tunisie Algérie01 Cap-Vert0 Egypte0 île Maurice01 Maroc01
Burkina-Faso Côte d'Ivoire Gambie Guinée Guinée-Bissau Libéria Madagascar Mali Mauritanie Namibie Niger Sénégal Sierra Leone Tchad
Afr. du Sud (nord, sud4) Angola Bénin Botswana Burundi Cameroun Comores Congo Djibouti Ethiopie Gabon Ghana Guinée équatoriale Kenya Malawi Mozambique Nigeria Ouganda R. Démocratique du Congo République centrafricaine Rwanda São Tome & PrincipeSomalie Soudan Swaziland Tanzanie Togo Zaïre Zambie Zimbabwe

182
AMERIQUE
toutes les villes et Antigua Antilles néerlandaises Bahamas Barbade Barbuda Bermudes Canada Chili Cuba Dominique Etats Unis Grenade Guadeloupe Iles Caïmans Iles Malouines Iles Vierges Jamaïque Martinique Porto Rico Sainte Lucie Trinidad et Tobago Uruguay
Argentine (nord1, sud4) Belise1 Bolivie5 (sud1) Costa Rica1 El Salvador1 Équateur5 (ouest) Guatemala1 Haïti Honduras1 Mexique1 Nicaragua1 Panama1,5 (ouest3) Paraguay (est1, ouest4) Pérou5 (ouest1) République Dominicaine Venezuela5 (hors amazonie)
Colombie5 (hors amazonie)
Bolivie (amazonie, ailleurs4) Brésil (amazonie, ailleurs4) Colombie5 (amazonie) Équateur5 (est) Guyana Guyane Française (fleuves, z. cotière4) Panama5 (est) Pérou5 (amazonie, ailleurs1) Surinam Venezuela5 (amazonie)
ASIE
Toutes les villes et Brunei Georgie Guam Hong Kong Iles Christmas Iles de Cook Indonésie5 (Bali) Japon Kazakhstan Kirghizistan Macao Maldives Mongolie Singapour Taiwan Arménie01 Azerbaïdjan01 Corée du Nord01 Corée du Sud01 Ouzbékistan01 Turkménistan01
Chine5 (nord-est1) Thaïlande5 (sud1, centre4)
Bangladesh5 (hors sud-est) Bouthan Inde Indonésie5 (bali4,hors Irian Jaya) Malaisie5 (hors Sabah & Sarawak) Népal Pakistan Philippines Sri Lanka1 Thaïlande5 (sud-ouet, centre4)
Bangladesh5 (sud-est)Cambodge6 Chine5 (Yunnan & Hainan) Indonésie5 (Irian Jaya) Laos6 Malaisie5 (Sabah & Sarawak) Myanmar6 Thaïlande5 (z. frontalières6; centre4) Timor Oriental Vietnam (côtes & deltas4)

183
EUROPE
Tous les pays, dont: Açores Belarus Canaries Chypre Etats Baltes Fédération de Russie Turquie d'Europe Ukraine
OCEANIE
Toutes les villes et Australie Fidji Iles de Pâques Iles Hawaï Iles Mariannes Iles Marshall Micronésie Nouvelle Calédonie Nouvelle Zélande Polynésie française Samoa Tonga Tuvalu
Iles Salomon Vanuatu
Papouasie-Nouvelle Guinée
PROCHE ET MOYEN
ORIENT
Toutes les villes et Bahreïn Israël Jordanie Koweït Liban Qatar Emirats Arabes Unis0 Oman0 Syrie01 Turquie01
Irak1
Iran5 (nord et centre)
Afghanistan Arabie Saoudite (ouest; est & nord4) Iran (sud-est) Yemen
0 : pays ne nécessitant pas de traitement du fait de la faiblesse de la transmission quelle que soit la durée du séjour. 1 : essentiellement transmission de Plasmodium vivax. 2 : compte tenu de la faiblesse de la transmission du paludisme, il est admissible de ne pas prendre de traitement préventif quelle que soit la durée du séjour. 3 : dans ces régions il est licite de ne pas prendre de traitement préventif pour un séjour inférieur à 7 jours, à condition d'être en mesure dans les mois qui suivent le retour de consulter en urgence un médecin en cas de fièvre. 4 : pas de traitement préventif nécessaire. 5 : plusieurs zones de chloroquinoresistance dans ce pays. 6 : dans les zones frontalières de/avec la Thaïlande il est possible d'utiliser préventivement de la doxycycline au lieu de la méfloquine.

184
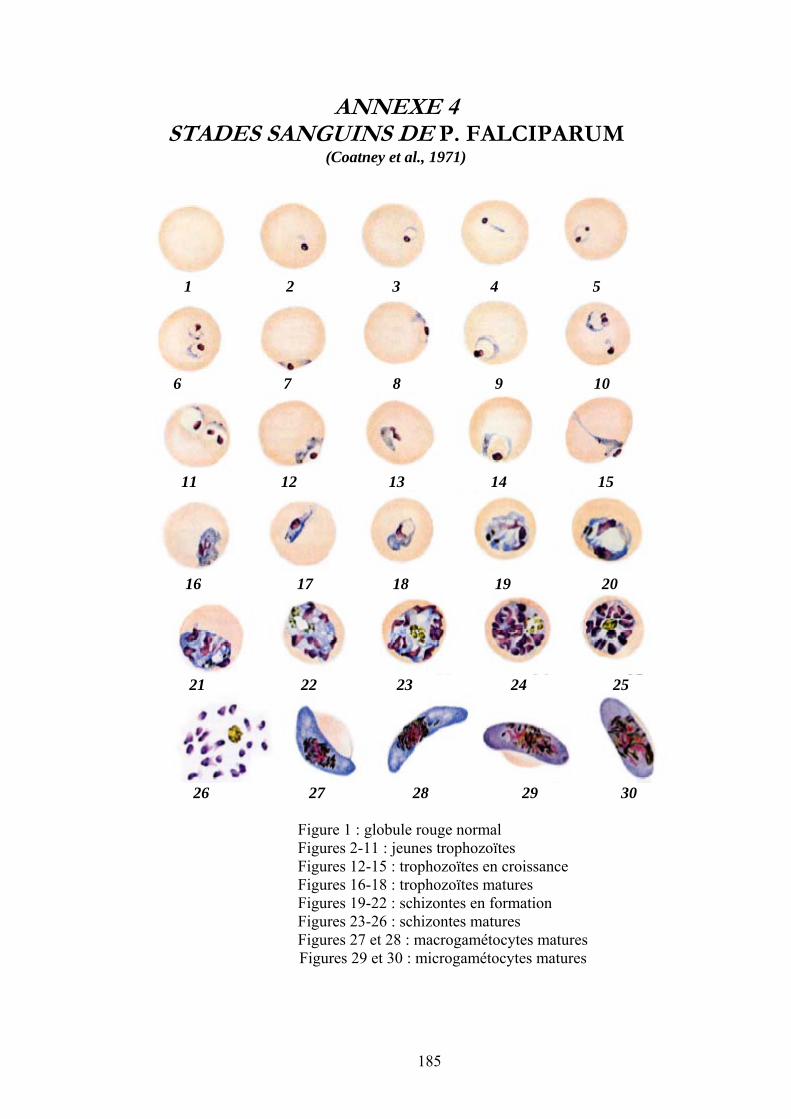
185
ANNEXE 4 STADES SANGUINS DE P. FALCIPARUM
(Coatney et al., 1971)
1 2 3 4 5
6 7 8 9 10
11 12 13 14 15
16 17 18 19 20
21 22 23 24 25
26 27 28 29 30
Figure 1 : globule rouge normal Figures 2-11 : jeunes trophozoïtes Figures 12-15 : trophozoïtes en croissance Figures 16-18 : trophozoïtes matures Figures 19-22 : schizontes en formation Figures 23-26 : schizontes matures Figures 27 et 28 : macrogamétocytes matures Figures 29 et 30 : microgamétocytes matures

186

187
ANNEXE 5 EXEMPLES DE FROTTIS SANGUINS HUMAINS
Frottis négatif : les globules rouges sont vides.
Trophozoïtes de P. falciparum : (exemples fléchés) c'est une infection classique à P. falciparum avec une densité parasitaire très élevée. Les hématies ne sont pas hypertrophiées (à la différence de certains autres plasmodiums humains), on peut voir des doubles grains de chromatine, de petites formes en anneau, des formes marginales et plusieurs parasites dans une même hématie (les hématies crénelées sont un artefact).

188
Macrogamétocyte de P. falciparum (flèche) Cette forme en croissant caractéristique de gamétocytes s’observe seulement chez P. falciparum. On peut noter l'aspect compact de la chromatine nucléaire.

189
ANNEXE 6
LISTE DES MOLECULES ANTIPALUDIQUES ET DE QUELQUES NOMS DEPOSES
Depuis janvier 1999, en France, tous les médicaments du paludisme sont inscrits aux listes I ou II. Le dictionnaire Vidal 2003 dresse la liste suivante d’antipaludiques :
• Amodiaquine : Flavoquine ® (cp) • Chloroquine : Nivaquine ® (cp pelliculé - cp sécable – sirop - sol inj) • Halofantrine : Halfan® (cp sec - sol buv) • Méfloquine : Lariam® (cp quadrisec) • Proguanil : Paludrine® (cp sec) • Proguanil-Atovaquone : Malarone® (cp enrobé) • Proguanil-Chloroquine : Savarine® (cp pelliculé) • Quinine : Quinimax® (cp pelliculé sec - sol inj), Quinine chlorhydrate Lafran®
(cp), Surquina® (cp pelliculé sec - sol pour perf), Quinoforme® (sol inj) • Pyriméthamine-Sulfadoxine : Fansidar® (cp quadrisec – sol inj) • Doxycycline : Doxypalu® (cp)
On peut également trouver d’autres molécules dans d’autres parties du monde :
• Dapsone : Disulone® • Sulfadiazine : Adiazine® • Proquanil : Paludrine® • Pyriméthamine : Malocide® • Pyriméthamine-Sulfadoxine-Méfloquine : Fansimef® • Artémisinine et ses dérivés : Paluther® • Pyronaridine : Malaridine®(extrême-orient uniquement) • Primaquine : ex Primaquin® en Allemagne (pays anglo-saxons uniquement) • Chlorproguanil-Dapsone : Lapdap® • Chlorproguanil-Dapsone-Artésunate : Lapdap Plus® • Dihydroartémisinine-Pipéraquine : Artékin ® • Artésunate-Amodiaquine : Arsucam ® • Artésunate-Sulfadoxine-Pyréthamine : Arsudar ®
Abréviations :
o cp : comprimé o sec : sécable o quadrisec : quadrisécable o sol : solution o buv : buvable o inj : injectable o perf : perfusion

190

191
ANNEXE 7 DOCUMENT DE SAISINE COMITE D’ETHIQUE
SUR L’EXPERIMENTATION ANIMALE ILE DE FRANCE SUD
Exemple : demande d’accord du comité pour le protocole d’étude clinique de l’infection de chimpanzés à P. benjamini.

192

193

194

195

196

197
ANNEXE 8 AMORCES DES 11 GENES UTILISES POUR CONSTRUIRE L’ARBRE DE P. BENJAMINI
EBP gene EBP-1 : 5' TTC CTT CTT TGT GTT ATA TTT TGC A 3' EBP-2 : 5' GAA ATT AAT TTT TTT TAT TGT TAC T 3' GLURP gene Glurp-1: 5' AAA ACT AAT ACA AGT GAG AAT AGA A 3' Glurp-2: 5' TGT AGG TAC CAC GGG TTC TTG TGG A 3' Prs48/45 gene A-1: 5' ATG ATG TTA TAT ATT TCT GCG AAA A 3' A-2: 5' CTA AAT ATA TAA TAA TAT TGC TAC A 3' Cysteine proteinase gene Cys-1: 5' AAA TTT ATG AAA GAA CAT AAT AAA G 3' Cys-2: 5' GAA AAC GTT GTC TCC ATT TTT ATT T 3' Cytochrome b gene Cytob-1: 5' TGG AAT TAC GGG TTC CTT TTA GGA A 3' Cytob-2: 5' GTA ATC ATA ATG TGT TGC TTT ATA A 3' MSA-2 gene Msa-21: 5' ATG AAG GTA ATT AAA ACA TTG TCT A 3' Msa-22: 5' TTA TAT GAA TAT GGC AAA AGC TAA A 3' Antigen 27 gamete gene A27-1: 5' ATG AGT AAG GTA CAA AAG GAT AGT G 3' A27-2: 5' AAT ATT GTT GTG ATG TGG TTC ATC A 3' Sporozoite antigen gene Sporo-1: 5' ATG AAT ATT CGA AAG TTC ATA CCA T 3' Sporo-2: 5' TTA AGA ATC ATC TCC TTC GTC TCC T 3' CSP gene Csp-1: 5' ATG ATG AGA AAA TTA GCT ATT TTA T 3' Csp-2: 5' CTA ATT AAG GAA CAA GAA GGA TAA T 3' 25 kDa ookinte antigen gene ooki-1: 5' ATG AAT AAA CTT TAC AGT TTG TTT C 3' ooki-2: 5' TTA CAT TAT AAA AAA GCA TAC TGA A 3' MSP-3 gene Msp-31: 5' TTA TTT TTG TTA CAT TTA TAT ATT T 3' Msp-32: 5' TTC TAC TAA ATC TTT TAA GGT AGA A 3'

198

199
ANNEXE 9 PUBLICATIONS ET PARTICIPATIONS A DES
CONGRES
DZIEGIEL M. H.L, LUNDQUIST R., MEJIA P., JAFARSHAD A., SOE-SOE, NIELSEN L.K., SALLE B., BOURRY O., ROUQUET P., DRUILHE P., 2005. Development of a recombinant antibody therapy against falciparum malaria. Soumis à Blood. SALLE B., 2005 Intérêt du primate africain dans la recherche sur le paludisme. 18éme Colloque de la Société Francophone De Primatologie, Besançon, France, 19-21 octobre 2005. Résumé à paraître dans Folia Primatologica. OLLOMO B. & SALLE B., 2004. A new model for primate malaria. COST B22 Congress on “Drugs development for parasitic diseases”, Anvers, Belgique, 22-26 novembre 2004. SALLE B., 2004. Les singes et la malaria. Interview radiophonique dans l’émission les Années Lumière de Pauline Vanasse sur Radio Canada du 27 Juin 2004. http://www.radio-canada.ca/radio/lumiere/.


SALLÉ BETTINA INTÉRÊT DES PRIMATES AFRICAINS DANS LA RECHERCHE SUR LE PALUDISME Thèse Vétérinaire : LYON, 3 février 2006
RESUME :
Le paludisme, causé par le plasmodium, tue chaque année plusieurs millions de personnes. Les recherches menées depuis un siècle sont nombreuses mais les scientifiques se heurtent toujours à l’absence d’un modèle fiable. Après un rappel sur le paludisme et sur les recherches liées à cette maladie, l’auteur compare les modèles déjà utilisés en orientant la discussion autour des primates africains. Quatre protocoles originaux utilisant ces primates sont enfin développés afin d’illustrer cette discussion.
MOTS CLES : - PALUDISME - PLASMODIUM - RECHERCHE BIOMEDICALE - PRIMATES
JURY :
Président : Monsieur le Professeur Stéphane PICOT 1er Assesseur : Monsieur le Professeur Gilles BOURDOISEAU 2ème Assesseur : Monsieur le Professeur Jean-Luc CADORE
DATE DE SOUTENANCE : 3 février 2006
ADRESSE DE L’AUTEUR : 11 rue Philippe de Beaumanoir ou CIRMF
60600 Clermont FRANCE BP 769 Franceville GABON



SALLÉ BETTINA INTÉRÊT DES PRIMATES AFRICAINS DANS LA RECHERCHE SUR LE PALUDISME Thèse Vétérinaire : LYON, 3 février 2006
RESUME :
Le paludisme, causé par le plasmodium, tue chaque année plusieurs millions de personnes. Les recherches menées depuis un siècle sont nombreuses mais les scientifiques se heurtent toujours à l’absence d’un modèle fiable. Après un rappel sur le paludisme et sur les recherches liées à cette maladie, l’auteur compare les modèles déjà utilisés en orientant la discussion autour des primates africains. Quatre protocoles originaux utilisant ces primates sont enfin développés afin d’illustrer cette discussion.
MOTS CLES : - PALUDISME - PLASMODIUM - RECHERCHE BIOMEDICALE - PRIMATES
JURY :
Président : Monsieur le Professeur Stéphane PICOT 1er Assesseur : Monsieur le Professeur Gilles BOURDOISEAU 2ème Assesseur : Monsieur le Professeur Jean-Luc CADORE
DATE DE SOUTENANCE : 3 février 2006
ADRESSE DE L’AUTEUR : 11 rue Philippe de Beaumanoir ou CIRMF
60600 Clermont FRANCE BP 769 Franceville GABON