Embed Size (px)
Citation preview

Promo : 2017-2018 Date : 28/02/2018
Ronéistes : Payet Charlotte Philippe Margaux
UE 8 : De l’agent infectieux à l’hôte
Plage horaire : 10h45 – 12h45 Enseignant : Jean Jacques Hoareau
TD DE BACTERIOLOGIE
Les bactéries ont une grande capacité à se multiplier, ce qui représente de ce fait un facteur de virulence. Mais dans nos corps elles ne peuvent pas croitre de la manière dont elles le voudraient, et heureusement, parce que ce serait vraiment préjudiciable pour nous. Cependant on a quand même du mal à empêcher les bactéries de croitre. C’est pourquoi on s’intéresse de plus en plus à l’anti- virulence thérapie. Ce type de traitement se développe depuis 10 ans déjà, mais il subsiste beaucoup de zones d’ombre: c’est pourquoi il est nécessaire que les nouvelles générations continuent les recherches afin d’acquérir plus de connaissances. La CONJUGAISON BACTERIENNE est l’élément central de l’anti-virulence thérapie. On utilise donc un plasmide, qui drive la conjugaison afin d’établir un pont entre les 2 bactéries. Il a ainsi pour rôle: le transfert du matériel plasmique et non pas chromosomique.
QCM 1 Les plasmides => CD A. Ce sont des molécules d’ADN extrachromosomiques présentes chez toutes les bactéries => non, pas
chez toutes. B. Ils sont nécessaires à la survie de l’hôte => confèrent un avantage sélectif. C. Ils confèrent un avantage sélectif dans certains milieux. D. Ils peuvent exister librement ou s’intégrer dans le génome de l’hôte. E. Les plasmides conjugatifs sont des plasmides qui ont pour fonction d’assurer le transfert du génome
d’une bactérie donneuse vers une bactérie receveuse => seulement pour les souches à haute fréquence de conjugaison
QCM 2: Le plasmide F impliqué dans la conjugaison bactérienne => ABCE A. Peut se répliquer de façon autonome. B. Peut être transféré d’une bactérie à une autre. C. Peut s’intégrer au chromosome de la bactérie => mais pas tous. D. Nécessite un bactériophage auxiliaire => pas besoin, même si in vitro on est capable de le faire. E. Modifie le sexe de la bactérie receveuse => plasmides F ont cette capacité.
QCM 6 La conjugaison => ABE A. Peut se faire entre bactéries de même espèce. B. Peut se faire entre bactéries d’espèces différentes. C. Ne concerne que les bactéries possédant un facteur de fertilité => pas seulement, mais
l’intégration au génome elle va être limitée. D. S’effectue uniquement entre bactérie fertiles (F+). E. Nécessite un contact direct entre bactéries = contact physique.

!

!

EXERCICE 1 ED 1) Quel est le marqueur de discrimination entre la souche Hfr et les recombinants ? Le marqueur de discrimination est un marqueur de résistance. En fait, il faut utiliser un marqueur pour éliminer la HFR parce qu'on ne veut garder que les recombinants. Ici il s’agit de la streptomycine car elle est présente partout.
2) L’ordre des gènes: on trouve l'ordre de gènes qui est le même que l'ordre de changement de phénotypes (ex : Thr -, Leu- est le premier à changer et à devenir Thr +, Leu+ donc c'est le 1er gène) . Ici l’ordre est : Thr, leucine, T1s, lac, gal
EXERCICE 2 ED On prend toujours un marqueur de sélection, ici c’est toujours la streptomycine.
Quel est l’ordre de passage des gènes ? Le maltose n’est pas un facteur discriminant, parce que les 2 bactéries sont capables de le produire.
Milieu 1 : niveau max de référence, car y a tout: Arg et gal ont été transférés très tôt car on obtient 60 colonies.
Milieu 3: on a 25 colonies. Absence de thr => la biotine est plus éloignée de l’origine de transfert que la thréonine car il lui faut plus de temps pour être transférée.
Milieu 4: il y a 20 colonies. Pas de Gal, ni de Biotine => soit se sont 2 marqueurs, soit l’un des 2 est responsable du faible taux car ils ont été transférés tardivement.
On comprend donc que moins on a de colonies sur un milieu, plus les gènes sont éloignés de l’origine: on n’a pas eu le temps de les transférer.
Thr se trouve avant la biotine qui elle, est plus éloignée de l’origine de transfert, puis c’est le galactose. ↪︎ DONC à partir de l’Origine de transfert on a: galactose/ arginine > thréonine > biotine
Mais on n’a pas d’informations pour discriminer le galactose et l’arginine (savoir qui est avant qui). Le transfert se fait de façon linéaire.
QCM 18 Concernant la croissance bactérienne => ABE A. Est fortement influencée par la nature physique et chimique de l’environnement. B. La majorité des bactéries sont neutrophiles et ne se développent que dans une gamme limitée de pH
comprise entre 5,5 et 8 => neutrophiles oui la majorité le sont, car le pH du cytoplasme est proche de 7 donc elles ont moins de difficultés à maintenir leur pH.
C. Pour chaque espèce bactérienne, entre les valeurs cardinales et maximales de températures, le métabolisme double à chaque augmentation d’environ 10°C => faux: c'est entre valeur minimale et valeur cardinale (= valeur optimale) que ceci est vrai ! Au contraire, quand on dépasse la valeur optimale le métabolisme (vitesse de croissance) diminue rapidement. De plus, la valeur cardinale est plus proche de la valeur max que de la valeur min (pas exactement entre les deux valeurs).
D. L’oxygène est l’accepteur d’électron indispensable au métabolisme des bactéries => oxygène pas chez toutes les bactéries, c’est pas absolu.
E. la croissance bactérienne est obtenu par scissiparité

QCM 19 La croissance bactérienne => BCE A. Les bactéries ne se divisent qu’en phase exponentielle de croissance => pas seulement. B. En milieu naturel, le développement bactérien est le plus souvent limité par des facteurs nutritifs ou
environnementaux. C. Le manque d’un seul facteur nutritif suffit à rendre la croissance bactérienne nulle => que ce soit un
macro ou un oligoélément. D. In vitro, la phase stationnaire de croissance correspond à un arrêt complet de la division bactérienne
due à un facteur limitant => nb division = nb de mort cellulaire => en phase stationnaire: autant de bactéries qui meurent que de bactéries qui apparaissent.
E. La diauxie peut permettre à certaines bactéries d’utiliser un métabolite alternatif lorsque son métabolisme préférentiel devient limitant => principe même de la diauxie.
EXERCICE 3 ED En phase exponentielle de croissance, chaque bactérie se divise à intervalle de temps constant. (donc doublement de population bactérienne à chaque intervalle de temps régulier, c’est ce qu’on appelle le temps de génération (tG) ou temps de doublement.)
1) Nt = N0 x 2^n => pour déterminer le nombre de bactéries obtenues après n générations, le développement de la population étant exponentiel . On aura n= ut et u= vitesse spé de croissance (nb de divisions / unité de temps)
2) Log Nt = Log N0 + nLog2 NB: Log2 = 0,301 alors donc n = (Log Nt - Log N0)/log 2 => pour déterminer le nombre de génération d’une population bactérienne à l’instant t
3) Supposons qu’une bactérie se divise toute les 20 minutes (No = 1), on a le tableau suivant car on a une division par scissiparité.
4) Déterminer le nombre théorique de bactéries obtenues après 12h, 24h et 48h de culture en phase exponentielle (N0 = 1).
Solution pour 12H On convertit 12H en minutes, que l’on divise par le temps de génération: 20 minutes. n=12*60/20 => n = 36 On applique ensuite la formule ci-dessus Nt = N0*2^n => Nt = 1 * 2^36 => Nt = 68 719 476 736 = 6,87 x10^10 bactéries

5) Sachant que le poids d’une bactérie de type E. Coli est estimé à 1 picogramme = 10^-12 - Discutez de la pertinence du résultat. En 12h, la masse de la bactérie s’élève à 69mg: c’est un résultat qu’il est possible d’obtenir. Par contre, à partir de 24h, la masse s’élève à 4720 tonnes ! Bien évidemment c’est impossible, ce qui montre bien que cette phase exponentielle de croissance n’est que passagère car les bactéries vont vite épuiser leur milieu. En condition naturelle, les bactéries sont rarement, ou alors très brièvement, en phase exponentielle de croissance. Dans leur milieu naturel, les bactéries sont toujours en conditions limitantes de nutriments et autres éléments indispensables à leur métabolisme: heureusement pour nous, même si elles n’ont pas besoin d’être au maximum pour nous envahir.
EXERCICE 4 ED
1) Combien de colonies devrait-on trouver sur cette gélose en supposant qu’elle soit composée de 10^7 bactéries ?
- 1 colonie = 10^7 batteries. - 1 colonie transférée dans 1L de bouillon => 1L à 10^7 bact/L.
- A T0 : 1mL de bouillon prélevé = 10^4 bactéries Diluées au 1/10 ème = 10mL à 10^3 bact/mL
- 100 microL prélevés = 10^2 bactéries Etalés sur gélose ordinaire à 37°C Chaque bactérie formant une colonie => 100 colonies (CFU) après culture.
2) Entre 0 et 2h ça ne bouge pas car on a une phase d’adaptation des bactéries. A partir de 3h ça augmente.
Il faut utiliser les valeurs entre la 2ème et la 6ème heure car avant le taux de croissance est nul.
Calculer le taux de croissance horaire (en h^-1) et le temps de génération (en minutes). Nt = No x 2^n -> Log Nt = log No + n Log 2 NB : Log 2 = 0,301
-> n= (Log Nt – Log No)/ log 2 avec n= u x t-> u x t = (log Nt – Log No)/ log 2
Determination de la vitesse spécifique de croissance entre 2 points de la phase exponentielle de croissance :
-> u = (Log Nt2 – log Nt2) / (t1-t2) x log 2= (7,2-6,4) / 0,301 = 2,67 divisions / h
ex : entre t5h et t6h Temps de génération t6 = 1 / u= 0,37h ( 22,5 min)

3) En une heure on sait qu’on a 2,67 divisions/h Nt = N0 x 2^n = N0 x 2^ut
Donc : N7h = N6h x 2^ut => N7h = 9, 95. 10^7
EXCERCICE 5
Dans une population de bactéries, on se rend bien compte que c’est une communauté mais on a aussi un certain nombre de mutants qui peuvent avoir un impact sur la croissance et sur la pathogénicité.
!

ETUDE 1 DE L’ED :
C’est une bactérie pouvant faire de la coopération cellulaire, ou former des biofilms. Puisque P.Aeruginosa fait du Quorum Sensing, on se demande si on ne pourrait pas établir une stratégie thérapeutique en ciblant ce mécanisme.
Le développement des biofilms est basé sur le Quorum Sensing (mais toutes les bactéries n'en sont pas capables) contrôlé par :
• ⁃ Gènes LAS I
• ⁃ Gènes RHL
L’expression constitutive de ces gènes va entrainer la formation des AHL (inducteurs), qui, en concentration suffisante, activent les gènes LAS R et induisent la sécrétion d’exo-produits qui auront une action dans le milieu :
- Enzymes de la virulence (—> dégradation tissu —> infection systémique)
• - De dégradation de substrat
• - De sporulation
Chez P. Aeruginosa, il existe naturellement 2 souches mutantes : - MUTANTS 1: Signal négatif: ils sont dépourvus du gène LAS I.
• - Détectent la densité cellulaire.
• - Ne produisent pas de AHL.
• - Mais libèrent des exoproduits (protéases) après stimulation par AHL.
- MUTANTS 2 = Signal blind: déficients en LAS R
• - Détectent la densité cellulaire.
• - Mais produisent AHL.
• - Ne produisent pas d’exo-produits (ø pathogénicité).
L’intérêt est donc de privilégier la souche « Signal Blind » pour réduire la pathogénicité.
Si AHL est présent il est possible d’agresser le tissu en produisant notamment de l’élastase.
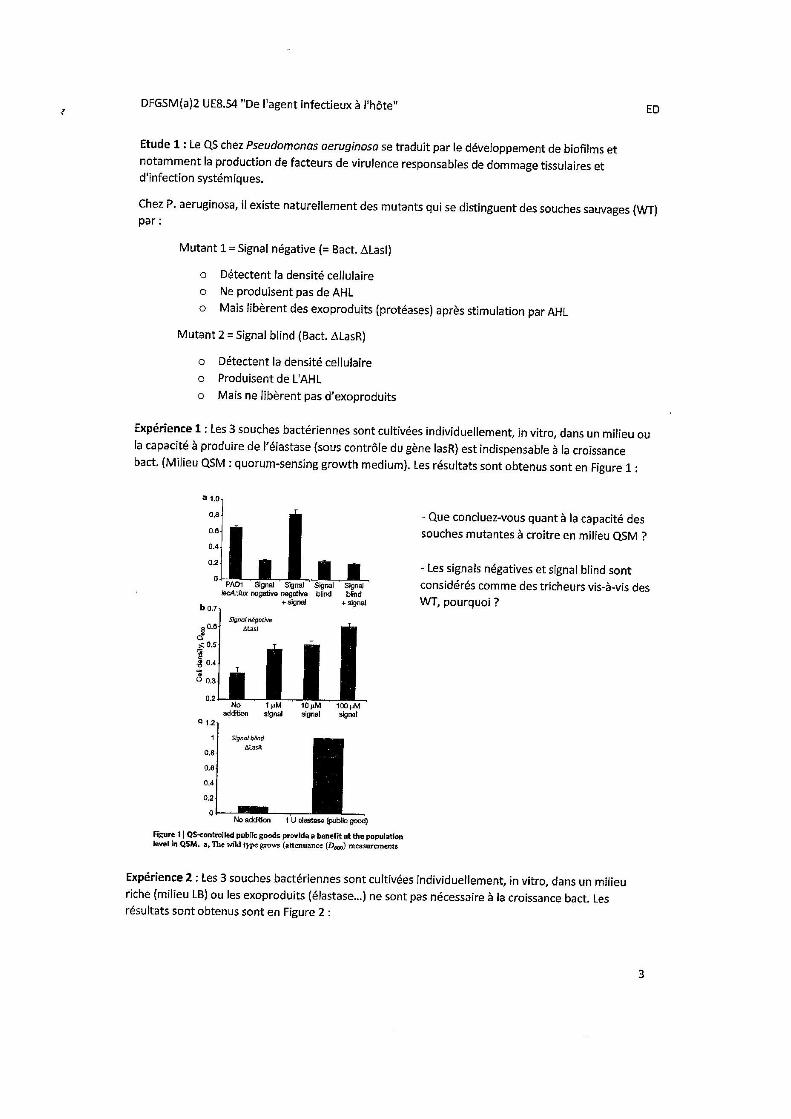
Expérience 1 : Les 3 souches bactériennes sont cultivées individuellement, in vitro, dans un milieu où la capacité à produire de l’élastase (sous contrôle du gène lasR) est indispensable à la croissance bactérienne. (Milieu QSM = quorum sensing growth médium). NB : La croissance bactérienne est d’autant plus importante que le signal est important. Ici, le facteur limitant est la production d’élastase. Sans elle, pas de croissance. Signal = AHL (c’est l’inducteur ici)

On compare la croissance de 3 différentes souches bactériennes, cultivées individuellement (in vitro) dans un milieu où la capacité à produire de l’élastase (sous contrôle du gène LasB) est indispensable à la croissance bactérienne.
• WT (Wild Type = souche naturelle) : souche de référence, ce qu’on devrait obtenir (produit AHL et exo- produits). • Signal négatif SEUL: ils sont seuls, or incapables de produire AHL => Pas de production d’exo-produits => pas de capacité de croissance dans le milieu.
• Signal blind SEUL: capables de produire AHL mais ne produisent pas d’exo-produits => ne peuvent dégrader le milieu à leur avantage => croissance lente. • Signal négatif + signal (HSL) : peuvent alors produire des exo-produits et dégrader le milieu : bonne croissance. Meilleure croissance que les WT (car les WT produisent eux-mêmes HSL donc dépensent plus d’énergie).
• Signal blind + signal : ne peuvent produire les exo-produits de toute façon donc pas de différence.
Souches tricheuses = signal négatif et blind car plus on ajoute de AHL plus la croissance est importante. Si on leur donne des inducteurs gratuitement ils vont croitre plus rapidement que ceux qui produisent les inducteurs eux mêmes.
Conclusion : La coopération bactérienne bénéficie à la communauté. Individuellement, les tricheurs sont défavorisés, mais en communauté ils sont favorisés par rapport aux WT.
EXPERIENCE 2

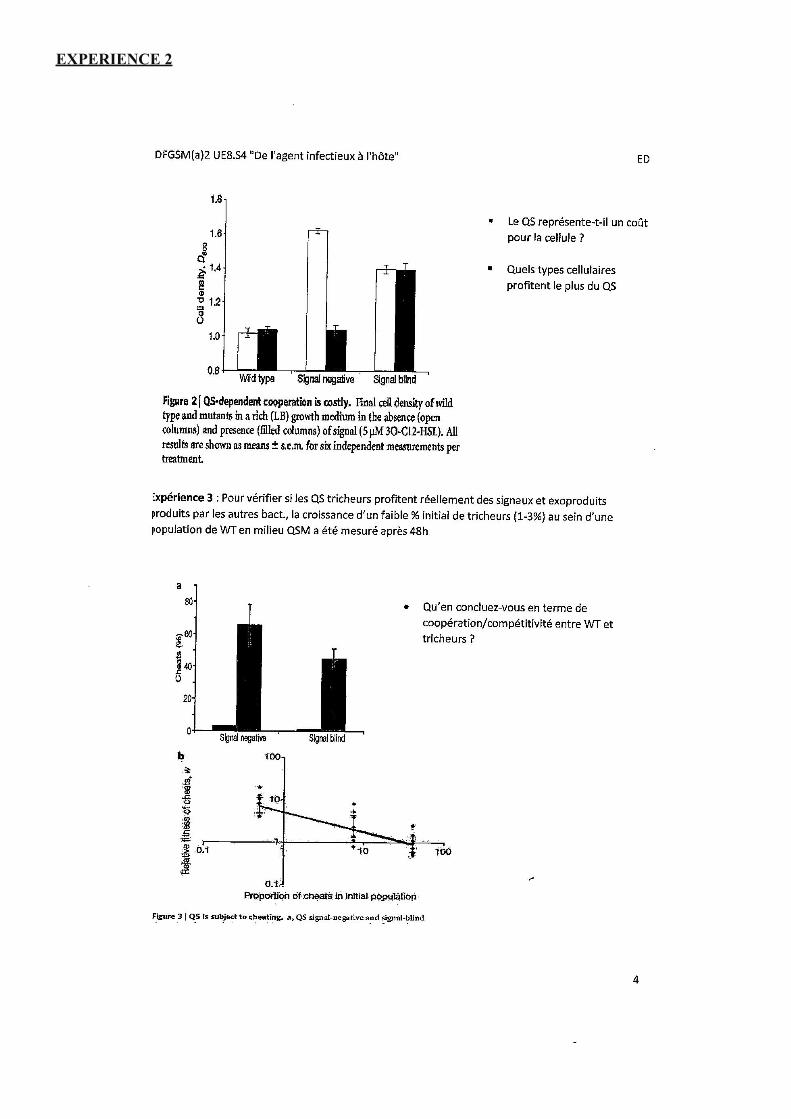
Expérience 2 : Les 3 souches bactériennes sont cultivées individuellement, in vitro, dans un milieu riche (LB) où les exo-produits (élastase) ne sont pas nécessaires à la croissance bactérienne. Les résultats sont en figure 2 :
Milieu où l’élastase n’est plus nécessaire à croissance bactérienne. En absence ou en présence de AHL.
• WT : ajout de AHL => différence non significative. • Signal négatif : croissance très importante en l’absence de AHL, quand on en rajoute, on observe une diminution de croissance.
• Quand on ne fournit pas de AHL, pas de production AHL, et pas de production d’élastase : peu de dépenses énergétiques.
• Quand on rajoute AHL, production d’élastase : dépenses énergétiques : la croissance diminue. • Signal blind: produit déjà AHL, et ne produit pas d’exo-produits : il n’y a donc pas d’influence lors du rajout de AHL.
Conclusion (cout de la coopération) : On note un fort coût énergétique pour les bactéries qui font du quorum sensing. En milieu riche (où il n’y a pas besoin d’élastase) les tricheurs produisent davantage de biomasse.
=> avantage des tricheurs !
NB : cout de production HSL< Elastase
Expérience 3 : On vérifie si les tricheurs profitent réellement des signaux et des exo-produits sécrétés par les autres bactéries, la croissance d’un faible % initial de tricheurs (1-3 %) au sein d’une population de WT en milieu QSM a été mesurée après 48h.
Ici, on regarde si, à partir d’un faible pourcentage de tricheurs, ces derniers deviennent majoritaires.
• - Signal négatif : + de 60 %
• - Signal blind :+ de 50 %
Au bout de 48h le signal négatif représente quasi 70%. Quand on regarde le taux de croissance en fonction de la proportion de tricheurs pour les WT : dès que le nombre de tricheurs augmente => taux de croissance des tricheurs diminue => moins le système marche.
Dans la figure B, on remarque un tricheur, ça a un avantage, mais à condition qu’il reste minoritaire. • Plus assez de producteurs de produits et du coup la croissance diminue.
En majorité on a des signal blinds, ils sont moins virulents que les souches sauvages (ø production d’exo- produits). • Lors d’une infection on pourrait agir sur le quorum sensing pour favoriser le développement des signal blind, car moins virulents ?

Qu’en concluez vous en terme de coopération entre WT et tricheurs ? => Les bactéries coopératrices sont en compétition avec les tricheuses.
- Les tricheuses envahissent les populations in vitro. - Mais la croissance des tricheuses est rapidement limitée par l’augmentation de leur % relatif au sein de la communauté.
!

ETUDE 2 DE L’ED: antivirulence thérapie
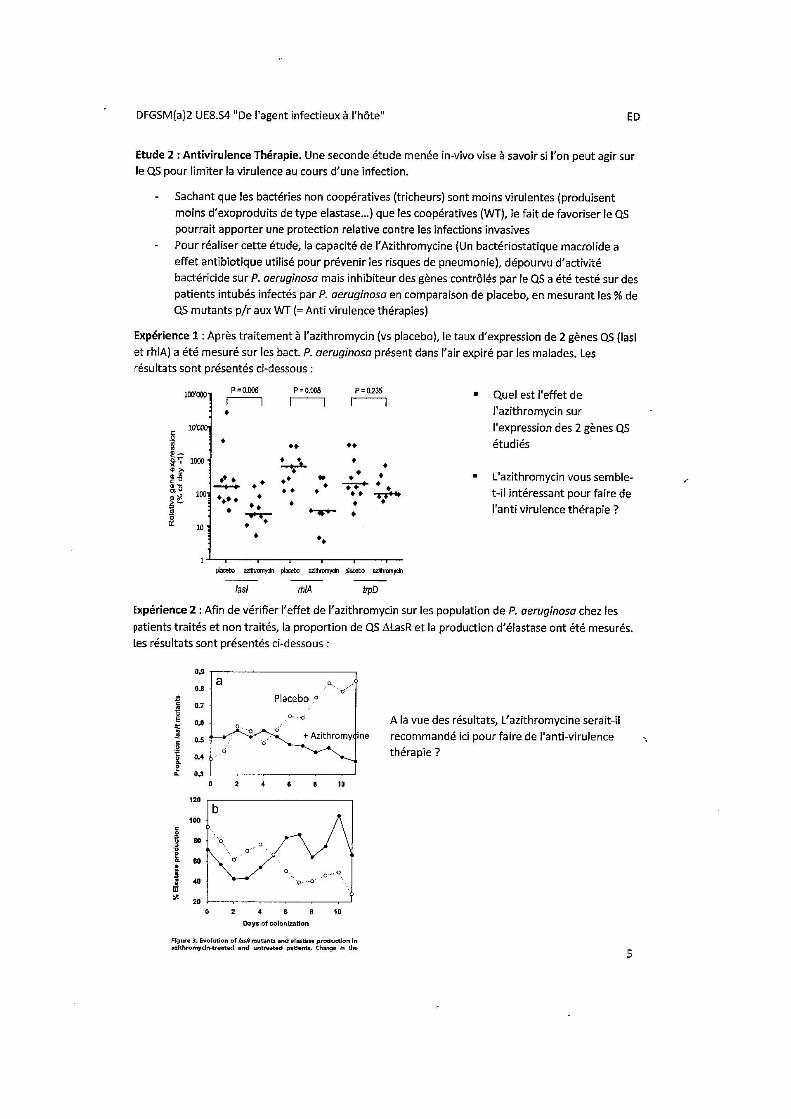
Azithromycine (A): inhibition des gènes contrôlés par le quorum sensing: ne tue pas les bactéries mais empêche multiplication bactérienne.
Effet de l’Azithromycine sur le mécanisme du quorum sensing mesuré par dosage de l’expression des gènes Las I et RHLa. Diminution de LAS I et RHL: l’azithromycine a un effet sur l’activité des gènes inducteurs du quorum sensing.
• A = Bactériostatique + agit sur les gènes du quorum sensing.
• Concept d’antivirulence thérapie.
L’azithromycine est capable d’inhiber l’expression des gènes QS => diminution du quorum sensing. Cette
expérience permet d’assurer que l’Azithromycine agit bien sur les 2 gènes en question.
- En l’absence d’azithromycine (A), les mutants QS (LasR = tricheurs), moins virulents, exploitent les ressources produites par les bactéries sauvages (coopérateurs), virulentes (LASl), et envahissent la population.
• En présence d’A, l’avantage sélectif des mutants LasR disparait et il n’y a pas d’augmentation du rapport LasR/wt.
• La drogue entraine une diminution du génotype avirulent LasR en milieu hospitalier.
• Avec le placebo, on voit une augmentation de la c° des tricheurs.
Cette expérience montre que, du fait de l’inhibition du gène LasI, la proportion de mutants LasR moins virulents diminue avec l’utilisation de l’Azithromycine, et la production d’élastase augmente.
En conclusion :
Arithromycine utilisée pour son pouvoir bactériostatique et non pas sur son effet anti-virulent, donc on ne remet pas en cause son traitement par Arithromycine mais cette augmentation de l'élastase qui est tout de même très légère reste à surveiller.





























