Embed Size (px)
Citation preview

Xylog6nkse chez les Dicotylkdones arborescentes. V. Formation du bois de tension et transport de I'acide indole adtique triti6 chez le Httre
SUZANNE LACHAUD Laboratoire de biologie et physiologie ve'ge'tales (Unite' associe'e 574), Station Biologique de Beau-Site, Faculte' des
sciences, 25, rue du Faubourg Saint-Cyprien, 86000 Poitiers, France
R e p le 2 avril 1986
LACHAUD, S. 1987. Xylogenkse chez les Dicotylkdones arborescentes. V. Formation du bois de tension et transport de l'acide indole acktique tritiC chez le HCtre. Can. J. Bot. 65 : 1253 - 1258.
En mai, les branches terminales de jeunes H&tres (Fagus silvatica L.) sont courbCes en anneau de Jaccard. Si elles restent en place sur l'arbre, elles produisent un bois de tension extkme, abondant, du c6tC supkrieur de la zone horizontale : nombreuses fibres gelatineuses, peu de vaisseaux et de parenchyme axial. Du c6tC inferieur de ces tiges, la xylogCnkse est pratiquement nulle. Si les branches tordues sont isolCes, la xylogCnkse est beaucoup moins dissymCtrique : le bois form6 du cBtC superieur comporte normalement vaisseaux et parenchyme axial mais aussi des fibres gelatineuses; du c6t6 infkrieur, le bois est normal. Le transport et le mCtabolisme de I'acide indole acCtique tritiC sont CtudiCs dans ces deux systkmes. Chez les branches restCes en place sur les arbres, le transport lateral de l'auxine s'effectue preferentiellement vers la moitiC inferieure des tiges horizon- tales. Chez les branches isolCes, on ne dCckle pas de sens preferentiel au transport lateral de l'auxine. La differenciation de bois de tension extrsme, dans la moitiC supCrieure d'une branche inclinCe, necessite donc l'intervention de facteurs correlatifs pro- venant non seulement des bourgeons mais aussi de la base de l'arbre, condition dans laquelle s'effectue en particulier le transport lateral descendant de l'auxine.
LACHAUD, S. 1987. XylogCnkse chez les Dicotyl6dones arborescentes. V. Formation du bois de tension et transport de l'acide indole acCtique tritiC chez le HCtre. Can. J. Bot. 65: 1253- 1258.
Upper stems of young beeches (Fagus silvatica L.) were bent into Jaccard loops in May. If they stayed on the tree, they pro- duced a wide arc of extreme tension wood on the upper side of the loop, which contained numerous gelatinous fibers, few vessels, and little axial parenchyma. On the lower side of these loops, xylogenesis was nearly stopped. If bent stems were removed from the tree, xylogenesis was far less asymmetric: the wood formed on the upper side contained normal vessels and axial parenchyma, but also gelatinous fibers; on the lower side, the wood was normal. Transport and metabolism of %-labelled indole acetic acid was studied in both models. In intact loops left on the trees, lateral auxin transport took place mainly towards the lower half of the stem. In isolated loops, no preferential direction could be detected for lateral auxin trans- port. Extreme tension wood differentiation on the upper side of a bent stem requires the intervention of correlative factors, not only from buds, but also from the base of the tree, which particularly influence lateral downward transport of auxin.
Introduction De nombreux travaux ont t t t consacrts B l'ttude de la
formation du bois de tension chez les Dicotyltdones arbores- centes; plusieurs articles de synthkse sont parus depuis une vingtaine d'anntes sur ce sujet (Wardrop 1964, 1965; Hugues 1965; Robards 1969; Scurfield 1973; Wilson et Archer 1977; Wicker 1979). Cette xylogtnkse particulikre se produit, en gtntral, du c6tt suptrieur de branches inclintes. Elle se carac- ttrise surtout par la difftrenciation de trks nombreuses fibres gtlatineuses, dont la paroi secondaire comporte une tpaisse couche peu lignifite, et par la rartfaction des vaisseaux, dont le diamktre est diminut; les cellules de parenchyme du bois sont gtntralement peu abondantes. Le bois de tension ne peut Ctre considtrt comme un phtnomkne de "tout ou rien" (Robards 1969); tous les intermtdiaires sont possibles entre le bois normal, qui peut contenir des groupes sporadiques de fibres gtlatineuses (Hugues 1965) et le bois de tension "extrCme" (Wardrop 1964; Wilson et Archer 1977) chez lequel le pourcentage de fibres gtlatineuses est trks tlevt. La formation du bois de tension est en gtntral associte B une activitt cambiale accrue du c6tt suptrieur, entrainant une croissance excentrique.
La participation de substances de croissance B cette xylo- g tnbe asymttrique est envisagte par de nombreux auteurs, dans l'optique d'une modification de leur rtpartition lattrale. Seul, le cas de l'auxine, le plus ttudiC, sera examint dans le cadre de cet article.
Le transport de l'acide indole acttique (AIA) dans les tiges horizontales a t t t assez peu Ctudit chez les arbres. Chez les jeunes tiges herbactes, qui manifestent un gtotropisme
Printed in Canada / lrnprirnC au Canada
ntgatif, l'auxine est prtsente prtftrentiellement B 60% dans la moitit inftrieure, vers laquelle peut s'effectuer un transport lattral (Goldsmith et Wilkins 1962; Gillespie et Thimann 1963; Lyon 1965). Ce phtnomkne semble se produire tgale- ment chez des tiges ligneuses inclintes situtes sur arbres ou arbustes entiers (NeEesany 1958; El Antably 1975); les rtsul- tats ttant parfois contradictoires chez des segments de tige : l'auxine y serait abondante du c6tt suptrieur (Wareing et al. 1964), ou du cat6 inftrieur (Leach et Wareing 1967; Lepp et Peel 1971). L'Ctude du mttabolisme de l'auxine exogkne dans les tiges horizontales montre que le transport lattral conceme essentiellement 17AIA lui-mCme; elle a surtout tt6 faite chez des plantes herbactes (Lyon 1965; Shaw et al. 1973; Mig- liaccio et Rayle 1984), rarement chez des arbres (Leach et Wareing 1967).
Chez des Dicotyltdones arborescentes, des auteurs (Netesany 1958; Casperson 1965; Kennedy et Farrar 1965; Cronshaw et Morey 1965, 1968) ont modifit ou provoqut la formation de bois de tension par des applications d'AIA ou d'acide triiodobenzoi'que sur des tiges horizontales ou verti- cales en place sur des arbres entiers. 11s ttablissent tous une relation entre le gradient lattral d'auxine et la formation de bois de tension du c6tt suptrieur des tiges inclinks. Cette interprttation est reprise par les ouvrages de synthbe (Robards 1969; Zimmermann et Brown 1971; Philipson et al. 1971; Wilson et Archer 1977; Wicker 1979). Plus rtcemment, Fisher (1985) observe la formation de bois de tension chez Terrninalia sous une anntlation et reprend l'hypothkse de la dtficience auxinique, alors que pour Neville et Woltz (1980), la forma- tion de ce bois chez Gleditsia ne rtsulte pas d'une simple redis- tribution radiale de l'auxine. Chez les plantes herbactes, de
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y Si
mon
Fra
ser
Uni
vers
ity o
n 11
/10/
14Fo
r pe
rson
al u
se o
nly.

1254 CAN. J . BOT. VOL. 65, 1987
chromatographies 74d' \h
comptages
FIG. 1. Application d'AIA [3H] sur des branches de Hetre tordues : localisation des zones analysees. (A) DCpBt de l'hormone du cBtC supCrieur s; (B) dCpBt de l'hormone du cGtC infkrieur i ; (C) dCtail de la zone analysCe, situCe de 7 B 16 cm du point d'application, vers la base de la tige.
nombreuses expkriences rkcentes remettent en cause l'ancienne hypothkse de Cholodny et Went (Hart et MacDonald 1984) attribuant 1'Clongation asymktrique des jeunes tiges horizontales B la formation d'un gradient latkral d'auxine. L'accent est mis davantage actuellement sur l'extm- sion de protons, plus active dansl'apoplaste pour la moitik infkrieure de ces tiges (Wright et Rayle 1983) ou celle du Ca2+ pour la moitiC supkrieure (Slocum et Roux 1983); toutefois le trans~ort l a t h 1 descendant de I'AIA. liC aux deux phknomknes prkckdents, est toujours considirk par ces auteurs comme faisant partie de la kponse de la plante B la gravid.
Pour tenter de ~rkciser les conditions dans lesauelles se forme le bois de tension, nous avons d'abord Ctabli chez le H&tre des systkmes expkrimentaux produisant ou non un bois de tension exteme, puis nous avons Ctudik le transport et le mktabolisme de l'auxine dans ces systkmes.
Matdriel et mdthodes Des Hetres (Fagus silvatica L.) de 2 B 4 m de hauteur, BgCs de 10 21
15 ans, ont CtC utilisCs. 11s sont plant& sous-couvert en for& domaniale de Moulikre arterne ern en the la Vienne). Pour provoquer de f a ~ o n certaine la diffkrenciation de bois de tension typique, des branches terminales ou sub-terminales sont courbCes et attachCes en "anneau de Jaccard" de 20 B 30 cm de diamktre (fig. 1). Les rameaux latCraux prisents au niveau de la torsion sont CliminCs. Plusieurs series d'anneaux sont rCalisCes pendant la pkriode de forte activitk cambiale (mai-juin), au cours de 3 annCes successives.
Une Ctude cytologique du bois form6 est effectuCe sur deux ensembles de rameaux tordus : les uns sont laissCs en place sur les arbres (13 exemplaires), les autres (14 exemplaires) sont coupCs le jour meme de la torsion, 70 cm environ sous l'anneau, et transportCs au laboratoire. 11s sont alors placCs en plein air, B l'ombre; leur base est plongCe dans un vase contenant de l'eau, la position originale de
en attente pendant 3 B 4 semaines; au bout de ce dClai, des segments de tige sont prklevks dans les zones horizontales des torsions. 11s sont fixCs B I'alcool Cthylique B 70%. Des coupes transversales faites au microtome sont colorCes au carmin alunC et vert d'iode, ou au bleu de toluidine pour mettre en Cvidence la couche G (Fisher 1985); elles sont alors examinCes au microscope.
Les applications d'AIA [3H] ont lieu en mai et juin sur des rameaux tordus depuis 3 B 4 semaines et restCs en place sur les arbres. Ces rameaux sont divisCs en deux groupes la veille de l'application de I'auxine : les uns (1 1 exemplaires) restent en for&, les autres (22 exemplaires) sont coupCs, apportCs au laboratoire et aliment& en eau comme dans 1'expCrience prkcidente. L'AIA [3H] (activitC spici- fique, 20 Ci par mmol, ou 740 GBq par mmol, CEA, Gif-sur-Yvette, France), contenu dans une goutte gClosCe de 20 pL (0,5 nmol, ou 92 ng, soit 370 000 Bq par rameau) est dCposC aprks grattage de l'icorce du cBtC supCrieur ou infirieur du rameau (fig. 1). Les gouttes sont protCgCes de la lumibre et de la dessication par une feuille d'aluminium. Ce dispositif est laissC en place pendant 3 jours, durant lesquels la plus grande partie des molCcules d'auxine s'Ccoule vers le bas du rameau B la vitesse moyenne de 13 B 15 cmljour B 20°C (Lachaud et Bonnemain 1982). La radioactivite que nous analyserons ensuite dans la zone horizontale de la torsion conceme donc les molC- cules retenues par les tissus aprks le passage de la vague basipke. Au bout de 3 jours, les rameaux sont sectionnCs, congelCs par de la carbo- glace, puis dCbitCs en centimktres B la semi-obscuritk dans un conge- lateur selon le schCma de la figure 1 ; chaque centimktre est fendu en moitiCs supCrieure et infirieure.
Pour d~f in i r la rkpartition globale de la radioactivitC dans les deux moitiCs de chaque centimktre, les segments destines au comptage sont IyophilisCs, pesCs, puis brCllCs dans un oxymat (IN 4101, Intertech- nique); la mdioactivite extraite est indiquCe par un compteur 21 scintil- lation liquide SL 33 (Intertechnique); le bruit de fond et le quenching sont comgCs. Pour prkciser la nature des moltcules marquees immo- bilisCes dans les deux moitiCs, les segments destinCs 21 la chroma- tographie sont regroup& pour plusieurs (2 B 6) demi-torsions, dCbitCs en copeaux, broyCs au mortier, puis extraits par le mCthanol B 80% B 4°C pendant 48 h B 1'obscuritC. Aprks prklkvement d'Cchantillons destines au comptage, des chromatographies de l'extrait concentrk sont effectuCes, sur 40 cm de papier Whatman no 1 (solvant : isopro- panol -ammoniaque-eau, 8:0,2: 1,8). La radioactivitC contenue dans cheque centimktre de papier sec est solubilisCe (Lumasolve, puis Lipoluma-Kontron), puis lue au compteur B scintillation liquide. La combustion des rCsidus solides indique la radioactiviti insoluble.
Influence de l'isolement du rameau tordu sur la formation du bois de tension
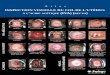
Chez les rameaux tordus intacts, laissks en place sur les arbres, un bois de tension exteme s'est formk en abondance du cBtC supCrieur des zones horizontales de tous les anneaux. Une petite couche de bois initial, dans lequel des vaisseaux se sont diffkrencib avant la date de la torsion, est prksente au dCbut du cerne (fig. 2A); cette couche est plus importante si la torsion est plus tardive. Le bois de tension dkveloppk ensuite prksente tous les critkres classiques dkcrits par les auteurs. Une zone cambiale large, trks active (fig. 2B) produit des sCries de fibres
chaque torsion Ctant conservie. L'ensemble de cis branchesest laissC B diffkrenciation lente, dans lesquelles apparait progressive-
FIG. 2. tat de la xylogCnkse le 11 juin chez des rameaux tordus le 17 mai. (A-F) Rameau laissC en place sur I'arbre. (A) SchCma du cBtC supirieur; (B) zone cambiale et bois de tension en formation; (C) partie supirieure du ceme montrant le dCpBt progressif de la couche G; (D) partie mCdiane du ceme avec bois de tension diffCrenciC; (E) schema du cBtC infkrieur; (F) bois form6 du cBtC infkrieur. (G-L) Rameau isole. (G) SchCma du cBtC supCrieur; (H et I) bois form6 du cBtC supirieur; (J) schema du cBtC infkrieur; (K et L) bois form6 du cBtC infkrieur. La longueur du trait de repkre est indiquCe en pm. Les hachures verticales reprksentent le bois de tension extreme sur le schCma A. Le cambium est situC vers le haut de la page pour les details C, D, I, vers le bas pour le dCtail K. La Keche horizontale indique le dkbut du demier ceme pour les details F et L. c, zone cambiale; f, fibres normales; g, fibres gelatinewes diffCrenciCes; gd, fibres gklatineuses en diffkrenciation; p, parenchyme axial; r, rayon; v, vaisseau.
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y Si
mon
Fra
ser
Uni
vers
ity o
n 11
/10/
14Fo
r pe
rson
al u
se o
nly.

LACHAUD
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y Si
mon
Fra
ser
Uni
vers
ity o
n 11
/10/
14Fo
r pe
rson
al u
se o
nly.

1256 CAN. J. BOT. VOL. 65. 1987
ment la couche gklatineuse : d'aspect plissk au dCbut de la diffkrenciation, elle s'kpaissit et rkgularise ensuite (fig. 2C et 2D). Les vaisseaux sont trks rares, voire absents. Le paren- chyme horizontal semble peu modifiC; le parenchyme axial, de type apotrachCal dans le bois de H&tre normal, n'est pas visible sur les coupes. Le pourcentage de fibres gklatineuses est ainsi trks ClevC dans ce bois de tension ccextr&mes.
Du c6te infkrieur de la tige, le bois est normal (fig. 2E et 2F), mais le cerne est trks rCduit 2 cause de la forte excentri- citC de croissance : la xylogtnbse est pratiquement nulle 2 partir de la date de la torsion.
Chez les rameaux isolCs, le bois qui s'est form6 du c6tC supCrieur des zones horizontales chez tous les rameaux n'est pas du bois de tension caractkristique (fig. 2G-21). La zone cambiale est active (fig. 2H); mais la proliferation des cellules est moins importante que chez les -rameaux tordus intacts (fig. 2B); la zone de diffkrenciation du xylkme est donc plus proche du cambium. Une couche gClatineuse se depose pour- tant dans les fibres (fig. 21), mais leur differenciation ne se fait pas aux dCpens de celle des autres types cellulaires. De nom- breux vaisseaux sont en effet prksents, ainsi que du paren- chyme axial, comme dans un bois initial normal; le diamktre des vaisseaux semble toutefois un peu rkduit.
I1 se forme du bois du c6tC infkrieur des rameaux isolCs (fig. 2J-2L) et celui-ci est normal; ses fibres n'ont pas de couche gelatinewe. La croissance du xylkme est encore excentrique, mais beaucoup moins que chez les rameaux tordus intacts : ainsi, pour les exemples reprCsentCs figure 2, 1'Cpaisseur totale du cerne de bois form6 est deux fois moins grande du c6tC infC- rieur que du c6tC supCrieur chez le rameau isolC, 8 2 9 fois moins grande chez le rameau tordu intact.
Transport et mktabolisme de L'AIA 13H] duns des rameaux tordus intacts et isolks
Les comptages et chromatographies sont effectuCs sur quatre lots expCrimentaux : arbres intacts, application d7auxine sur le c6tC infkrieur (lot 1) ou supCrieur (lot 2) du rameau; tiges isolCes, application d'auxine sur le c6tC inftrieur (lot 3 ) o u supkrieur (lot 4) du rameau.
Rkpartition de la radioactivitk duns le 10' cm en dessous du point d'application de L'AIA 13H]
Les molCcules marquCes sont retenues de f a ~ o n relativement uniforme dans la zone analysCe (du 7e au 16e cm en-dessous du point d'application de l'auxine, voir figure 1); on prend comme exemple le loe cm. La figure 3A indique la quantitC moyenne de radioactivitk retenue dans chaque moitiC du loe cm, en dpm (60 dpm = 1 Bq) par mg de poids sec, pour chacun des quatre lots expCrimentaux. La variabilitC des rCsul- tats est assez importante; on peut remarquer toutefois que la quantitC globale de radioactivite retenue semble plus grande dans le lot 4 que dans les trois autres lots. Cette observation se trouve d'ailleurs confirmCe ulterieurement Dar 1'Cvaluation. au cours des chromatographies, de la quantitC totale de radio- activitC extraite dans les segments 8, 11 et 14. L'exarnen de la proportion de molCcules marqutes retenues dans la moitit opposCe 2 l'application, par rapport 2 la quantitt totale de radioactivitk pksente au loe cm (fig. 3B), fait apparaitre que le transfert lattral s'est effectuC surtout dans le lot 2 (application du c6tC supkrieur de branches intactes, formant donc du bois de tension caractCristique). Dans ce lot, 18% en moyenne des molCcules marquCes feraient donc l'objet d'un transport lateral vers le bas. Ce transfert de radioactivit~ ne semble pas imputable 2 une rktention prtfkrentielle des radioClCments dans
. . . . 100 * cote d 'appl i -
cation de I 'AIA[~H]
S moyenne
mdtabolites 0 @@ insolubles
M J M J M M J
FIG. 3. RCpartition de la radioactivitk et mCtabolisme de I'AIA [3H] dans les moitiCs supCrieure et inferieure de rameaux horizontaux, 3 jours aprbs I'application. Arbres intacts : lot 1, application du cGtC inferieur; lot 2, application du cGtC supCrieur. Branches isolCes : lot 3, application du cGtC infkrieur; lot 4, application du cGtC supCrieur. (A) RadioactivitC moyenne en dpm par mg de matibre sbche, dans les moitiCs supCrieure (s) et infCrieure (i) des rameaux, au loc centimbtre suivant le point d'application en direction de la base. Les intervalles de confiance des moyennes sont indiquCs avec 5 % de risques; la limite supCrieure de l'erreur est seule repr6sentCe. (B) Transport lateral : pourcentages de la radioactivite retenue, du c6tC opposC a l'application, par rapport a la quantitC totale de radioactivite prksente au loc centimbtre, et exprimCe par mg de matibre skche. Chaque trait horizontal indique la valeur trouvCe pour un individu; le cercle noir reprksente la moyenne de ces valeurs pour chaque lot. (C) Chromato- graphies : pourcentage de m o l ~ c u l e s ~ a ~ a n t le R , de I'AIA, celui de l'AIAsp, ainsi que d'autres mCtabolites solubles et insolubles, par rapport 2 la radioactivite totale, dans les moitiCs supCrieure (s) et infC- rieure (i) de groupes de rameaux, en mai (M) et juin (J). Les nombres de rameaux sont respectivement de gauche 2 droite pour chaque sCrie : lot 1, 3 (mai) et 2 uuin); lot 2, 3 (mai) et 3 @in); lot 3, 6 (mai); lot 4, 6 (mai) et 5 @in). Les chiffres indiquks reprksentent les pourcentages correspondant 2 I'AIA.
la moitit inferieure du rameau, Ctant donnt que le marquage n'augmente pas lorsque l'application a lieu du c6tC infkrieur (fig. 3A7 lot 1). Dans ce dernier cas, le passage de la radio- activitk en sens inverse (du cbtC infirieur vers le cbtt supCrieur) ayant concern6 en moyenne 4 3 % des molkcules (fig. 3B7 lot I), on peut en dCduire que 13% environ des traceurs ont CtC transportCs prkfkrentiellement du cbtC sup& rieur vers le cbtt infkrieur des branches tordues intactes (lot 2).
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y Si
mon
Fra
ser
Uni
vers
ity o
n 11
/10/
14Fo
r pe
rson
al u
se o
nly.

LACHA
Chez les branches tordues isoltes, qui forment un bois relative- ment comparable des deux c6tts, il n'y a pas de sens prtftren- tie1 au transport lattral (fig. 3B, lots 3 et 4).
Mktabolisme de 1'AIA r3H] dans les moitiks supkrieure et in- fkrieure des tiges courbkes, au-dessous du point d'appli- cation (ge, 11 et 1 4e cm)
Les chromatographies sont rtsumtes figure 3C. Les rtsultats obtenus en mai et juin ont t t t figurts stpartment parce que le mttabolisme de 1'AIA [3H] chez les rameaux isolts du lot 4 s'est rtvtlt assez difftrent pour ces deux ptriodes (alors que la quantitt totale de radioactivitt retenue ne vane pas de mai 2 juin). Les principales substances reptrtes sont situtes aux Rf de 1'AIA et de l'acide indole acttyl aspartique (AIAsp); chaque diagramme indique les pourcentages de ces deux sub- stances par rapport 2 la radioactivitt totale. La radioactivitt insoluble, tvalute par combustion des rtsidus solides, reprt- sente en moyenne 14% de la radioactivitt totale; ses variations ne semblent pas likes 2 la nature des exptriences.
Chez les arbres intacts, les plus forts pourcentages d'AIA [3H] (45 et 50%) sont prtsents dans la moitit inftrieure des branches inclintes quand l'application a lieu du c6tt suptrieur (lot 2). Le transport lattral prtftrentiel vers le bas, dtcelt par comptages, concernerait donc surtout 1'AIA libre. Les plus faibles pourcentages d'AIA [3H] (28 et 28 %) sont obtenus pour la moitit inftrieure quand l'application a lieu de ce mCme c6tt (lot 1). On ne note pas de difftrences dans le mttabolisme de 1'AIA [3H] chez les rameaux intacts entre mai et juin.
Chez les branches inclintes isoltes, les pourcentages d'AIA [3H] sont trbs voisins dans les moitits suptrieure et inftrieure pour chaque lot : cette similitude est peut-&tre like 2 l'absence de transport lattral pkftrentiel de l'hormone dans ce systbme.
Dans le cas du lot 4, pour lequel la rttention globale de radioactivitt dtcelte par comptage est particulibrement impor- tante, le pourcentage d'AIA [3H] est plus tlevt dans les extraits de mai que dans ceux de juin. La forte ktention con- cernerait donc surtout 1'AIA en mai, quand le bois initial se difftrencie; par contre l'auxine serait davantage mttaboliste en juin, quand la couche de bois initial est pratiquement achevte et que l'on tvolue vers le bois final.
Discussion et conclusions Les observations effectutes permettent donc de dtfinir deux
systbmes exptrimentaux constituts par des tiges de H&tre tordues en anneau de Jaccard, pour lesquels la xylogtnbse est difftrente dans la moitit suptrieure : les rameaux en place sur l'arbre forment du bois de tension <<extr&me~, les rameaux isolts forment un bois de structure intermtdiaire entre bois normal et bois de tension. Dans ce dernier cas, le bois possbde, certes, des fibres 2 couche gtlatineuse, mais les proportions des divers types cellulaires ne semblent pas modifites. Rappe- lons ici que la plupart des auteurs ne caracttrisent pas unique- ment le bois de tension par la presence d'une couche G dans les fibres (cela peut se produire chez le H&tre dans certaines zones du bois "normal" formt sur des tiges verticales (Hugues 1965)) mais par la frtquence tlevte de ces fibres (Wilson et Archer 1977; Wicker 1979), qui implique une diminution de la frtquence des autres types cellulaires, en particulier des vais- seaux. Si la ntcessitt de la prtsence de l'apex pour la forma- tion du bois de tension a t t t montrte depuis les travaux dYOnaka et de Wardrop (citts par Wardrop 1964 et Hugues 1965), impliquant l'intervention de rtgulateurs de croissance 2 circulation basipbte (Fisher 1985), celle de la partie basale de
l'arbre ne semble pas encore avoir t t t signalte par les auteurs. Dans la moitit suptrieure d'une branche inclinte, posstdant son bourgeon terminal, la gravitt favorise peut-&tre le dtp6t d'une couche gtlatineuse dans les fibres, mais elle est insuffi- sante pour modifier la determination des modbles cellulaires. Une influence de la base de l'arbre, peut Ctre en provenance du systbme racinaire, serait donc responsable d'un dtrbglement asymttrique du processus de xylogtnbse chez les cellules issues des initiales fusiformes. Elle aboutirait, dans la moitit suptrieure de la branche 2 la multiplication du modble fibre gtlatineuse et 2 la rartfaction des modbles vaisseau et paren- chyme, dans la moitit inftrieure 2 un arr&t presque total de la xylogtnbse.
Nos exptriences sur le transport et le mttabolisme de l'auxine dans les branches inclintes de H&tre intactes montrent que 13% environ de 1'AIA [3H] retenu se sont dtplacts lattralement vers la moitit inftrieure de la tige. Cette propor- tion correspond aux donntes de la litttrature sur la rtpartition lattrale de 1'AIA dans les tiges horizontales de plantes her- bactes, ou de ligneux (NeEesany 1958; El Antably 1975). En revanche, dans les branches horizontales de HCtre isoltes, o i ~ un bois presque normal s'est formt dans la moitit suptrieure, la rtpartition lattrale de 1'AIA retenu est symttrique : le trans- port lattral prtftrentiel vers le bas de la phytohormone n'a apparemment pas eu lieu. I1 semble donc y avoir un lien entre l'existence d'un transport lattral descendant de l'auxine et la formation d'un bois de tension .extreme* dans les branches inclintes de HCtre. Nos rtsultats concordent avec les remarques de plusieur auteurs (NeEesany 1958; Casperson 1965; Cronshaw et Morey 1968; Wilson et Archer 1977; Fisher 1985) sur le fait que le bois de tension se formerait chez les arbres entiers dans une zone dtficiente en auxine. Toute- fois, ils n'expliquent pas pourquoi le transport lattral de l'auxine, qui n'a pas lieu dans les branches isoltes du H&tre, se produit dans les tiges isoltes de plantes herbactes : chez les arbres, seuls vtgttaux B former du bois de tension, le transport lattral de l'AIA, ou du moins sa rttention prtftrentielle du c6tt inftrieur des branches, pourrait dtpendre d'un ensemble plus complexe de corrtlations. Par ailleurs, notre travail ne prttend pas ttablir, chez les branches tordues intactes, une relation de causalitt directe entre les difftrents tvbnements : inclinaison de la tige, descente d'auxine, difftrenciation du bois de tension extsme. La rtponse gtotropique peut com- mencer effectivement par une redistribution lattrale d'AIA (Migliaccio et Rayle 1984), mais d'autres facteurs sont probablement impliquts dans la regulation asymttrique de la xylogtnbse.
Remerciements
Ce travail a btntficit de l'aide du Centre National de la Recherche Scientifique (Unit6 associte 574 et Action Thtma- tique Prograrnrnte ~Physiologie de la croissance et du dtveloppement des vtgttaux ligneuxn). Nous remercions vive- ment l'Office National des For& pour sa collaboration, ainsi que A. Potonier pour son assistance technique.
CASPERSON, G. 1965. ~ b e r endogene Faktoren der Reaktionsholzbil- dung. I. Wuchsstoffapplikation an Kastanienepikotylen. Planta, 64 : 224 - 240.
CRONSHAW, J . , et MOREY, P. R. 1965. Induction of tension wood by 2,3,5-triiodobenzoic acid. Nature (London), 205 : 816 -818.
1968. The effect of plant growth substances on the develop- ment of tension wood in horizontally inclined stems of Acer rubrum seedlings. Protoplasma, 65 : 379-391.
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y Si
mon
Fra
ser
Uni
vers
ity o
n 11
/10/
14Fo
r pe
rson
al u
se o
nly.

1258 CAN. J. BOT. VOL. 65, 1987
EL ANTABLY, H. M. M. 1975. Redistribution of endogenous indole acetic acid, abscisic acid and gibberellin in geotropically stimulated Ribes nigrurn shoots. Physiol. Plant. 34 : 167 - 170.
FISHER, J. B. 1985. Induction of reaction wood in Terminalia (Com- bretaceae) : role of gravity and stress. Ann. Bot. (London), 55 : 237-248.
GILLESPIE, B., et THIMANN, K. V. 1963. Transport and distribution of auxin during tropistic response. I. The lateral migration of auxin in geotropism. Plant Physiol. 38 : 214-255.
GOLDSMITH, M. H. M., et WILKINS, M. B. 1962. Movement of auxin in coleoptiles of Zea mays during geotropic stimulation. Plant Physiol. 37 : 492-505.
HART, J. W., et MACDONALD, I. R. 1984. Is there a role for apex in shoot gravitropism? Plant Physiol. 74 : 272-277.
HUGUES, F. E. 1965. Tension wood : a review of literature. Part I. Occurrence and development of tension wood. For. Abstr. 26 : 1-16.
KENNEDY, R. W., et FARRAR, J. L. 1965. Induction of tension wood with the antiauxin 2,3,5-triiodobenzoic acid. Nature (London), 208 : 406-407.
LACHAUD, S., et BONNEMAIN, J. L. 1982. Xylogtnbse chez les DicotylCdones arborescentes. 111. Transport de l'auxine et activitt cambiale dans les jeunes tiges de HCtre. Can. J. Bot. 60 : 869- 876.
LEACH, R. W., et WAREING, P. F. 1967. Distribution of auxin in horizontal woody stems in relation to gravimorphism. Nature (London), 214 : 1025 - 1027.
LEPP, N. W., et PEEL, A. J. 1971. Distribution of growth regulators and sugars by the tangential and radial transport system of stem segments of willow. Planta, 99 : 275-282.
LYON, C. J. 1965. Auxin transport in geotropic curvatures of a branched plant. Plant Physiol. 40 : 18-24.
MIGLIACCIO, F., et RAYLE, D. L. 1984. Sequence of key events in shoot gravitropism. Plant Physiol. 75 : 78-81.
NEEESANY, V. 1958. Effect of /3-indole acetic acid on the formation of reaction wood. Phyton (Buenos Aires), 11 : 117 - 127.
NEVILLE, P., et WOLTZ, P. 1980. Recherches sur le systbme d'inter- actions internes contr6lant la formation et le fonctionnement du
cambium caulinaire chez Gleditsia et Quercus. Action concert&. : materiau-bois. Dtltgation gCnCrale A la recherche scientifique et technique (D.G.R.S.T.) 78 7.3133.
PHILIPSON, J. J. , WARD, J. M., et BUTTERFIELD, B. G. 1971. The vascular cambium, its development and activity. Chapman & Hall, London.
ROBARDS, A. W. 1969. The effect of gravity on the formation of wood. Sci. Progr. Oxford 57 : 513-532.
SCURFIELD, G. 1973. Reaction wood : its structure and function. Science (Washington, D.C.), 179 : 647 -655.
SHAW, S., GARNER, G., et WILKINS, M. B. 1973. The lateral transport of IAA in intact coleoptiles of Avena sativa L. and Zea mays L. during geotropic stimulation. Planta, 115 : 97- 11 1.
SLOCUM, R. D., et Roux, S. J. 1983. Cellular and subcellular locali- zation of calcium in gravistimulated oat coleoptiles and its possible significance in the establishment of tropic curvature. Planta, 157 : 481 -492.
WARDROP, A. B. 1964. The reaction anatomy of arborescent Angio- sperms. Dans The formation of wood in forest trees. Editeur : M. H. Zimmermann. Academic Press, New York. pp. 405 -456.
1965. Formation and function pf reaction wood. Dans Cel- lular ultrastructure of woody plants. Editeur : J. A. CotC. Syracuse University Press, Syracruse. pp. 371 -390.
WAREING, P. F., HANNEY, C. E. A., et DIGBY, J. 1964. The role of endogenous hormones in cambial activity and xylem differentia- tion. Dans The formation of wood in forest trees. Editeur : M. H . Zimmermann. Academic Press, New York. pp. 323 -344.
WICKER, M. 1979. Le bois de tension : acquisitions rtcentes. Ann. Biol. 18 : 221 -254.
WILSON, B. F., et ARCHER, R. R. 1977. Reaction wood : induction and mechanical action. Annu. Rev. Plant Physiol. 28 : 23-43.
WRIGHT, L. Z., et RAYLE, D. L. 1983. Evidence for a relationship between H+ excretion and auxin in shoot gravitropism. Plant Physiol. 72 : 99-104.
ZIMMERMANN, M. H., et BROWN, C. L.. 1971. Trees, structure and function. Springer-Verlag, New York.
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y Si
mon
Fra
ser
Uni
vers
ity o
n 11
/10/
14Fo
r pe
rson
al u
se o
nly.