Embed Size (px)
Citation preview

VOL. 21 (I956) SHORT COMMUNICATIONS 385
The dis t r ibut ion of the isotopic carbon in the glucose from liver glycogen was found to be essentially the same whether the glucose-I-l*C was administered by s tomach tube or by intra- peri toneal injection. In both cases most of the radioact ivi ty was found in C-z and the activity present in the other carbon a toms was dis t r ibuted almost equally among them. None of the carbon a toms 2 th rough 6 contained significantly more radioactivi ty than the others. As expected, there was slightly more randomizat ion in the 9o-minute exper iments than in the 6o-minute experiments , bu t the distr ibut ion of isotope th roughou t the carbon chain was essentially the same.
Ttlese data indicate tha t cleavage of the six carbon chain of glucose during the process of intestinal absorpt ion and t r anspor t to the liver is of minor quant i ta t ive significance. In these in vivo exper iments over 9o per cent of the absorbed glucose found in the liver glycogen mus t have been absorbed by the intestine and t ranspor ted to the liver as a six carbon unit. This is in agreement with the recently reported perfusion studies of KiYASU A N D C H A I K O F F 7.
Fur the r studies are being conducted to determine the nature of the six carbon uni t t ha t is t ranspor ted across the intestinal cells to the portal blood.
This work was suppor ted in pa r t by a grant (H-I731-C2) from the U.S. Public Health Service.
Department o/ Physiological Chemistry, The Johns Hopkins VV. R. TAYLOR University School o/Medicine, Baltimore, :]ld.( U.S.,4 .) ROBERT G. LANGDON *
1 M. P. HELE, Biochem. J., 55 (1953) 857, 864. 2 A. SOLS, Bioehim. Biophys. Acta, 19 (1956) 144. 3 S. HESTRIN-LERNER AND B. SHAPIRO, Biochim. Biophys. Acta, 12(1953) 533; 13 (1954) 54- 4 T. H. WILSON AND G. WISEMAN, J. Physiol., 123 (1954) 116. 5 \¥. R. TAYLOR AND R. G. I,ANGDON, Federation Proc., 15 (1956) 369 . 6 p. A. MARKS AND B. L. HORECKER, J. Biol. Chem., 218 (1956 ) 327 • 7 j . y . KIYASU AND I. L. CHAIKOFF, Federation Proc., 15 (I956) i i i .
Received May 23rd, 1956
• This work was (lone during the tenure of a Lederle Medical Facul ty Award.
Assimilation de I'ad~nine par la levure
L'ad6nine-8-14C est rap idement assimil6e par la levure en milieu glucos6 d6pourvu d 'a l iment azot4. Ire 14C se re t rouve dans les deux purines des acides nucl6iques et dans divers compos6s pur iques "acido-solubles"l , 2.
Nous avons 6tudi6 les variations, au cours du temps, de la dis tr ibut ion du 14C dans l 'ensemble
des d4riv6s pur iques de la levure. Exp4rience: Le m u t a n t "pet i tes colonies ''3 est cultiv6 2 4 heures £ 28 ° en milieu liquide
/~ l 'abri de l 'air 4. La levure rdcoltde est lav6e et remise en suspension (3 mg poids sec par ml) dans une solution contenant 4 ° g de glucose, 2oo nlg de sulfate de magn6sium et 45 ° nlg de phos- phate monopotass ique par litre. Cette suspension, maintenue pendant 75 minutes ~ 3 o°, manifeste une fermentat ion tr6s active. A 5 ° ml de cette suspension de levure "au repos" sont ajoutds 0.5 nil d 'une solution de sulfate d'ad6nine-8-t~C contenant 2/zmoles d 'ad6nine et 2 ttcuries de 14C par nil. Des 6chantillons de 5 illl sont pr61ev6s ~ divers moments , m6lang6s h 5 ml d 'une solution 2 M d'acide perchlorique contenant des quanti t~s bien d6termin6es (et voisines de o. 3 mg par ml) de xanth ine et d ' hypoxan th ine qui serviront d 'ent ra lneurs (carriers). Le m61ange est chauff6 3 ° minutes ~ IOO ° pour lib6rer les purines de leurs compos6s. Apr~s centrifugation et 61imination du s6diment, les purines sont pr6cipit6es de l 'extrai t selon ABRAMS 5, isol6es par chromatographie sur papier (solvants: butanol tertiaire-HC1 puis butanol normal -ammoniaque) et leur radioactivitd sp6cifique est d6termin6e selon une technique que nous avons d6crite ailleurs 2.
L'ad6nine et la guanine totales sont dos6es par la m6thode de KERR et al. 6, l ' hypoxanth ine par dilution isotopique. La quant i t6 totale tie xanth ine contenue dans la levure est n6gligeable devant celle de l 'entra ineur ajout6.
L'hypoxanthine-8-14C utilis6e pour le dosage par dilution isotopique a 6t6 pr6par6e de la fa~on suivante : A 2 mg d'ad6nine-8-14C dissoute dans o. 5 ml d'acide ac6tique glacial, on ajoute o. 5 ml d 'une solution aqueuse /~ 2o % de ni tr i te de sodium. Le m61ange est laiss6 4 ° heures ~ la temp6ra ture du laboratoire, puis 4vapor6 ~ sec sur un bain marie bouillant ; le r6sidu est redissous dans 2 ml d'acide perchlorique i ]~'/ et les purines sont pr6cipit6es par l 'argent selon ABRAMS 5. L 'hypoxan th ine est s6par6e de petites quant i t6s d 'ad6nine par chromatographie sur une courte colonne de r6sine sulfon6e Dowex-5o; une solution 2 M d'HC1 est utilis6e comme 61uant. Le rendement de cette prepara t ion est voisin de 9o %, l ' hypoxanth ine isol6e ne donne qu 'une seule tache radioactive lorsqu 'on la soumet ~ la chromatographie sur papier, et son spectre d 'absorpt ion dans l 'ultra-violet est correct.
386 SHORT COMMUNICATIONS VOL. 21 (I956)
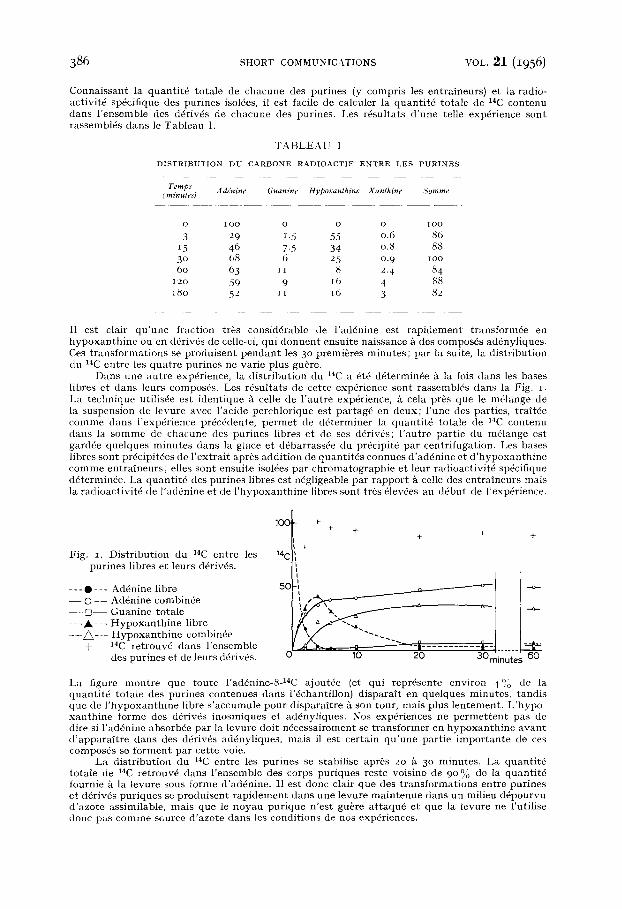
Connaissant la quanti t0 totale de chacune des purines (y compris les entraineurs) et ta radio- activit6 sp6cifique des purines isoMes, il est facile de calculer la quanti t~ totale de 14C contenu dans l 'ensemble des d6rivds de chacune des purines. Les r6sultats d 'une telle exp6rience sont rassembl~s dans le Tableau I.
T A B L E A U I
D I S T R I B U T I O N D U C A R B O N E R A D I O A C T I F E N T R E L E S P U R I N E S
Temps (minutes) Addnine Guanine Hypoxanthine Xanthine Somme
O 1OO O O O I O O
3 ~9 L5 55 0.6 86 15 46 7.5 34 0.8 88 3 ° 68 6 25 0.9 ioo 60 63 I1 8 2. 4 84
12o 59 9 16 4 88 I80 52 I I 16 3 82
I1 est clair qu 'une fraction tr6s consid6rable de l 'ad6nine est rap idement transform6e en hypoxan th ine ou en d6rivds de celle-ci, qui donnent ensuite naissance A des compos~s ad6nyliques. Ces t rans format ions se produisent pendan t les 3 ° premi6res minutes; par la suite, la distr ibution du 14C entre les quat re purines ne varie plus gu~re.
Dans une autre exp6rience, ia dis tr ibut ion du 14C a 6t6 ddtermin6e /t la fois dans les bases libres et dans leurs compos6s. Les r6sultats de cette experience sont rassemblds dans la Fig. 1. La technique utilis6e est identique A celle de l 'autre experience, A cela pros que le mdlange de la suspension de levure avec l'acide perchlorique est partag6 en deux; l 'une des parties, trait6e comme dans l 'exp~rience pr6cddente, perniet de ddterminer la quanti t6 totale de laC contenu dans la somme de chacune des purines libres et de ses d6rivds; l 'autre partie du mdlange est gard6e quelques minutes dans la glace et d6barrass6e du pr~cipit6 par centrifugation. Les bases libres sont p%cipitdes tie l 'extrai t apr~s addition de quanti t0s connues d'ad6nine et d 'hypoxan th ine comme entra ineurs ; elles sont ensuite isol6es par chromatographic et leur radioactivit6 sp6cifique d6termin~e. La quanti t6 des purines libres est n6gligeable par rappor t A celle des entraineurs mais la radioactivit0 de l 'addnine et de l ' hypoxanth ine libres sont tr6s 61ev6es au d6but de l'exp6rience.
Fig. i. Dis t r ibut ion du 14C entre les purines libres et leurs d6riv6s.
- -- • - -- Ad6nine libre - - O - Ad6nine combinde
- - ~ - - Guanine totale - - - l k - - - Hypoxan th ine libre - - ~ - Hypoxan th ine combinde
+ 14C retrouvd dans l 'ensemble des purines et de leurs ddriv6s.
100~ r +
14C1 ~ + -t- + + + H
5011 , -o-
10 20 30rninutes 60
La figure mont re que toute l'ad6nine-8-1aC ajout6e (et qui repr6sente environ 4% de la quanti t6 totale des purines contenues dans l'6chantillon) disparait en quelques minutes, tandis que de l 'hypoxanth ine libre s 'accumule pour disparaitre/~ son tour, mais plus lentement. L 'hypo- xanth ine forme des d6riv6s inosiniques et addnyliques. Nos exp6riences ne pe rmet ten t pas de dire si l 'ad6nine absorb6e par la levure doit ndcessairement se t ransformer en hypoxan th ine avan t d 'appara i t re dans des d6riv6s ad6nyliques, mais il est certain qu 'une partie impor tan te de ces compos6s se forment par cette voie.
La distr ibut ion du 14C entre les purines se stabilise apr~s 2o ~ 3 ° minutes. La quanti td totale de 14C retrouv~ dans l 'ensemble des corps puriques reste voisine de 9o% de la quanti t6 fournie A la levure sous forme d'ad0nine. I1 est donc clair que des t ransformat ions entre purines et d6riv0s pur iques se produisent rap idement dans une levure maintenue dans un milieu d6pourvu d 'azote assimilable, mais que le noyau pur ique n 'es t gu~re a t taqu6 et que la levure ne l'utilise donc pas comme sGurce d 'azote dans les conditions de nos exp6riences.

VOL. 21 (I956) SHORT COMMUNICATIONS 387
L'hypoxan th ine est un pr6curseur impor t an t des nucl6otides ad6nyliques chez la levure; dans une 6tude sur la synth~se de l'acide nncl6ique chez divers embryons, STEINERT 7 6tait arriv6
une conclusion semblable, il avait montr6 que dans les premiers stades du d6veloppement la synth~se des nucl6otides ad6nyliques se fait largement aux d6pens d 'hypoxanth ine .
Laboratoire de Chimie biologique de la Faculti des Sciences, Universitd libre de Bruxelles (Belgique)
H. CHANTRENNE S. DEVREUX
l S. KERR, K. SERAIDARIAN ET G. B. BROWN, J . Biol. Chem., 188 (1951) 207. 2 H. CHANTRENNE, zJ rch. Biochem. Biophys. (sous presse). g B. EPHRUSSI, a . I~IoTTINGUER ET A.-M. CHIM~kNES, Ann. Inst. Pasteur, 76 (]949) 351. 4 H. CHANTRENNE ETC. COURTOIS, Biochim. Biophys. Acta, 14 (1954) 397. 5 R. ABRAMS, ~4rch. Biochem., 3 ° (I951) 44. 6 S. KERR, K. SERAIDARIAN ET M. VVTARGON, J . Biol. Chem., 181 (I949) 76I. 7 M. STEINERT, Biochim. Biophys. Acta, 18 (1955) 511.
Re~u le 31 mai 1956
The presence of uracil, uridine and hypoxanthine in ether-ethanol extracts of Tumor 755 and other biological material
During a s tudy of the ul traviolet- l ight-absorbing compounds present in ether extracts of t rans- plantable adenocarcinoma 755 and in the organs of C57BL mice carrying these tumors as well as in those of normal rabbi ts paper ch romatograms consistently showed the presence of uracil. When larger quant i t ies of tissue were used uridine was also found. Subsequent shaking with ethanol extracted hypoxan th ine from these tissues, from the milk of R I I I mice carrying the agent of spontaneous m a m m a r y t u m o r and from frogs' eggs. Neither uracil nor uridine was detected in extracts from the last two sources.
The organs were dropped into liquid nitrogen immediately after sacrificing the animals. Later they were rapidly disintegrated at low tempera tu re and dried from the frozen state. The intact eggs and milk were freeze-dried, the lat ter after centrifugation to remove fat. The desiccated materials were extracted with anhydrous ether in a Soxhlet appara tus for 5 clays, the extract was concentrated, an equal volume of water and 0.2 volume, or more, of lO% trichloroacetic acid added and the mixture stirred mechanically for i to 1 ~2 hours. The water layer was removed, concentra ted to a very small volume under reduced pressure and subjected to chromatography . The ether-extracted materials were shaken wi th absolute ethanol for 6 to 7 hours a day for 5 days using fresh solvent daily. The pooled extracts were centrifuged, the supe rna tan t fluid taken to dryness under reduced pressure and the residue dissolved in a little water and analyzed. In some cases, e.g. liver extracts , the residue contained lipides which had escaped removal by the ether. They were dissolved in ether; at the same t ime the hypoxan th ine went into solution as shown by its presence in the water layer when this ether solution was t reated as the initial ext rac t with this solvent had been. I t is not known why this purine dissolved in ether at this point and failed to do so (luring the initial extract ion with this solvent. Uracil was sometinles found in the alcohol extracts in addition to being present in the original ether solution.
After the ch romatograms were developed the dark areas seen under a n.y. l amp were cut out, eluted, and the solution concentrated and again analyzed, a different solvent being used for the new chromatogram. Light absorpt ion measurements were then made on eluates of the dark areas seen under the lamp using eluates of equal areas from unused par ts of the sheet for control solutions in the cuvette.
Uracil. This compound, found in ether extracts of mouse t u m o r 755, spleen, kidney, liver and lung as well as in extracts of rabbi t liver, kidney and lung (mixed) had RF values of 0.28, o.37 and 0.85, respectively, on ch romatograms developed with n-butanol and ammonia (60, io of 0. 7 M); n-butanol, formic acid and water (77, io, 13) ; and 72 % phenol (solvents i, 2 and 3). These values were in agreement with those of an authent ic sample of uracil as were the wave lengths at which maximal and minimal absorpt ion occurred by solutions adjusted to p H 2, 7 and 13.
Uridine. This compound was present in the ether extracts of tumor , in those of mouse spleen and kidney (mixed) and in those of rabbi t liver and lung (mixed). The R/; values, when solvents z and 2 were used, were o.13 and o.2o. They were the same for a known sample of uridine. Light absorpt ion measurements were made at p H 2 and 13. The posit ions of maximal and minimal absorpt ion were the same for the tissue compound and the known sample. On heat ing the tissue compound with HCIO 4 at IOO ° for 2 hours uracil was formed.
25