Embed Size (px)
Citation preview

i
N° d’ordre ...................
UNIVERSITE DE OUAGADOUGOU ---------------
École Doctorale Sciences et Technologies
--------------- Laboratoire de Biologie et de Génétique moléculaire
(LABIOGENE)
Thèse Présentée
Par METUOR DABIRE Amana
Pour obtenir le grade de
Docteur de l’Université de Ouagadougou
Option Sciences Appliquées
Spécialité : Biologie Moléculaire/Enzymologie
THEME :
Caractérisations moléculaire et cinétique des types de β-lactamases à spectre élargi
(BLSE) de souches bactériennes collectées au Centre Hospitalier Universitaire
Pédiatrique Charles De Gaulle (CHUP-CDG) de Ouagadougou
Soutenue le 30 Juin 2014 devant le jury composé de :
Président: - Pr Mondher EL JAZIRI, professeur titulaire de Biologie des organismes, Université
Libre de Bruxelles (Rapporteur)
Membres: - Pr Rasmata TRAORE/OUEDRAOGO, Professeur titulaire de Bactériologie/Virologie,
Université de Ouagadougou (Rapporteur)
- Pr Jacques SIMPORE, professeur titulaire de Génétique et de Biologie Moléculaire,
Université de Ouagadougou (Co-Directeur de thèse)
- Dr Boukaré ZEBA, Maître de conférences, Université de 0uagadougou (Directeur de thèse)
- Dr D. Simplice KAROU, Maître de conférences, Université de Lomé (Rapporteur)

ii
DEDICACES
A
Mon père SOME Donbéyir
A
Ma mère DABIRE Yâomèyèro
A
Mes enfants DABIRE Sawdetouo Francis et METUOR DABIRE Gnowtar Lise-Maelle
A
Ma chérie SORE Cathérine
A
La mémoire de mon oncle SOMDA Nurukyor Claude
A
Mes frères et soeurs : SOME Sankoubéiyel, DABIRE Kô-samè, SOME Ayouo, DABIRE
Kouoritew, SOME Nanwniatoulou Franceline, SOMDA Dogfounianalou et SOMDA
Debloukoun.

iii
REMERCIEMENTS
Ce travail a été réalisé au Laboratoire d’Enzymologie de la Chimio-Résistance Bactérienne
(L.E.C.R.B) du département de Biochimie-Microbiologie de l’Université de Ouagadougou, au
laboratoire de Biologie Moléculaire du Centre National de Recherche et de Formation sur le
Paludisme (CNRFP) à Ouagadougou et au Laboratoire de Biotechnologie Végétale (LBV) à
l’Université libre de Bruxelles en Belgique.
Cette formation a été rendue possible grâce au financement de la Commission Universitaire
pour le Développement (CUD). Ce travail n’aurait pas vu le jour sans la franche collaboration
et le soutien de plusieurs personnes auxquelles nous exprimons une profonde gratitude.
Nous tenons à remercier particulièrement :
Le professeur Boukaré ZEBA, promoteur et Directeur de cette Thèse et Directeur du
Laboratoire d’Enzymologie de la Chimio-Résistance Bactérienne (L.E.C.R.B.). Merci de
m’avoir accueilli dans votre laboratoire pour m’initier aux techniques de recherches
scientifiques. Merci professeur pour les multiples corrections que vous avez apporté à mes
écrits.
Le professeur Mondher EL JAZIRI responsable academique de la CUD en Belgique,
Directeur du Laboratoire de Biotechnologie Végétale (LBV) à l’Université libre de Bruxelles
en Belgique qui a d’emblée porté un interêt tout particulier à ce travail et a tenu à sa
réalisation. Merci pour les soutiens de tout genre et les précieux conseils qu’il n’a cessé de me
prodiguer. Sa rigueur scientifique et son amour du travail bien fait ont été déterminant dans la
finalisation de ce travail. Il m’a fait l’honneur de m’accueillir dans son laboratoire en
Belgique. Ces séjours m’ont permis d’acquérir de nouvelles connaissances et d’approfondir
certains aspects de la recherche qui, pour des raisons logistiques n’étaient pas réalisables à
Ouagadougou. Merci infiniment au Pr Mondher pour toutes les facilités qu’il a toujours
veillées à m’offrir tant dans les locaux de son laboratoire qu’au niveau du logement lors de

iv
mes séjours en Belgique. Qu’il me soit permis de lui témoigner ici ma profonde
reconnaissance.
Le Pr Jacques SIMPORE, Professeur titulaire, Responsable du Laboratoire de Biologie
Moléculaire et de Génétique (LABIOGENE); Coordonnateur du Master de Biologie
Moléculaire et de Génétique Moléculaire Appliquées (BioGeMA), Université de
Ouagadougou; Directeur du Centre de Recherche Biomoléculaire Pietro Annigoni (CERBA);
Recteur de l’Université Saint Thomas d’Aquin (USTA); Membre de l’Académie Nationale
des Sciences du Burkina (ANSB) et notre Co-Directeur de thèse pour avoir accepté notre
encadrement. Il n’a ménagé aucun effort pour la correction de notre document de thèse malgré
ses multiples occupations.
Le professeur Rasmata TRAORE/OUEDRAOGO, Chef de service des laboratoires au
Centre Hospitalier Universitaire Pédiatrique Charles De Gaulle (CHUP-CDG) qui a bien
voulu m’autoriser à faire un stage en bactériologie. Merci encore professeur pour m’avoir
permis de collecter mes échantillons dans votre laboratoire et pour avoir accepté de juger
mon mémoire de DEA et présider le Jury.
Nous remercions Madame Marie BAUCHER pour les multiples corrections apportées à nos
articles et ses encouragements. Merci encore Marie pour votre grande disponiblité et votre
grande gentillesse.
Nous adressons nos vifs remerciements à M. Pascal SOME pour ses soutiens de tout genre et
pour ses précieux conseils et encouragements.
Un très grand merci à SANOU Antoine et à MOUSSAWI Jihad pour leur bonne
collaboration et leurs assistances techniques sur les paillasses respectivements au CNRFP à
Ouagadougou et au LBV en Belgique.
Grand merci au Dr COMPAORE Moussa pour la correction de mes écrits et ses
encouragements.

v
Nous adressons nos vifs remerciements au Dr SIRIMA, le Directeur du CNRFP et au Dr
SOULAMA, le responsable du laboratoire de la Biologie Moleculaire du CNRFP qui nous
ont acordé un séjour dans ledit centre pour les analyses de nos échantillons.
Nous remercions Dr Martin TIENDREBEOGO pour ses conseils et encouragements et Dr
D Simplice KAROU pour avoir accepté juger notre travail et participer au jury.
Grand merci à tout le personnel du Centre Hospitalier Universitaire Pédiatrique Charles De
Gaulle (CHUP-CDG), pour leur bonne collaboration. Particulièrement à M. TAMBOURA
Mamadou et à M. KAMBIRE Dianibè. Nous remercions également tout le personnel du
CNRFP à Ougadougou et tout le personnel du LBV en Belgique pour leur bonne
collaboration.
Infiniment merci à tout le personnel administratif de la CUD au Burkina Faso et en
Belgique.
Merci à M. Basile DARGA pour ses encouragements.
Nous remercions tous les enseignants qui ont contribué chacun dans son domaine à notre
formation. Grand merci à mon collègue ZONGO K. Jacob pour son soutien de tout genre et
sa bonne collaboration et à Mlle SORE Cathérine pour son soutien de tout genre et surtout
pour sa grande patience. Nous remercions nos frères et amis particulièrement
OUEDRAOGO Nicolas, DABIRE Ives, SOME Kouhinir, SAWADOGO Adama,
GOMINA G. Cyrille, Mme SOMDA /SOME Bèlouniakoun, DABIRE Isidore, BIMBILE
SOMDA N. Séverin, KABORE Awa, GOULLA Gauthier, KABORE Boukaré, ZEBA
Harouna et SAWADOGO Harouna pour leurs soutiens et encouragements. Grand merci à
M. NIKIEMA Fulbert, à M. HIEN Christian et à M. COMPAORE Muller pour leur
appui technique et encouragement. Grand merci également à M. Roland MEDA et à M.
Alphonse SOMDA pour leurs appuis informatiques et encouragements.

vi
Nous témoignons ici notre reconnaissance à M. Zoulyandine NAPON, mon maître de l’école
primaire qui m’a hébergé durant mon cycle primaire et participé activement à mon éducation
depuis mon bas âge.
Mention spéciale à mes parents pour les sacrifices à mon éducation, mon instruction, tout
simplement à ma vie.
Nous remercions l’UEMOA à travers son programme PACER2 et la CEI pour leurs soutiens
financiers.

vii
TABLE DES MATIERES
INTRODUCTION .................................................................................................................... 1
CHAPITRE I : SYNTHESE BIBLIOGRAPHIQUE ............................................................ 5
I- Bactéries ............................................................................................................................. 5
I-1-Définition .......................................................................................................................... 5
I-2-Paroi bactérienne .............................................................................................................. 5
I-2-1-Structure ..................................................................................................................... 5
I-2-2-Propriétés de la paroi ................................................................................................. 8
I-2-3-Biosynthèse de la paroi .............................................................................................. 8
I-3-protéines-liant-pénicilline (PLP) ...................................................................................... 9
II-Antibiotiques ...................................................................................................................... 10
II-1- Généralités .................................................................................................................... 10
II-2-Effet antibactérien ......................................................................................................... 12
II-3-Sites d’action et propriétés d’activité ............................................................................ 13
II-3-1-Antibiotiques agissant sur la synthèse du peptidoglycane ......................................... 13
II-3-2-Antibiotiques agissant sur les membranes (externe et cytoplasmique) ..................... 14
II-3-3-Antibiotiques agissant sur l’appareil nucléaire .......................................................... 14
II-3-4-Antibiotiques agissant sur les ribosomes ................................................................... 15
II-3-5-Antibiotiques agissant sur les acides mycoliques (antituberculeux) .......................... 16
II-4-Méthodes d’étude « in vitro » de l’activité des antibiotiques ....................................... 16
II-4-1-Etude de la bactériostase ............................................................................................ 17
II-4-2-Etude de la bactéricidie .............................................................................................. 18
II-5- β-lactamines ................................................................................................................. 19
II-5-1- Définition et structure chimique ............................................................................ 19

viii
II-5-2- Mode d’action sur les bactéries ............................................................................. 22
II-6-inhibiteurs des β-lactamases .......................................................................................... 23
III-Résistances bactériennes aux β-lactamines .................................................................... 24
III-1-Résistance par production de β-lactamases .................................................................. 26
III-1-1-Définition des β-lactamases .................................................................................. 26
III-1-2-Classification des β-lactamases ............................................................................. 28
III-2- β-lactamases à spectre élargi ....................................................................................... 30
III-2-1-Généralités ............................................................................................................ 30
III-2-2-Répartition mondiale ............................................................................................. 31
III-2-2-1-techniques microbiologiques ............................................................................. 32
III-2-2-2-techniques moléculaires ..................................................................................... 36
III-2-3-diversité des types de BLSE ................................................................................. 37
III-2-4-Epidémiologie des BLSE ...................................................................................... 39
III-2-4-1-Apparition et propagation des BLSE en Europe ............................................ 39
III-2-4-2-Apparition et propagation des BLSE en Asie .................................................... 40
III-2-4-3-Apparition et propagation des BLSE en Amérique .......................................... 41
III-2-4-4-Apparition et propagation des BLSE en Afrique .............................................. 41
CHAPITRE II : MATERIEL ET METHODES ................................................................. 44
I-Cadre de l’étude ................................................................................................................... 44
II-Type d’étude ....................................................................................................................... 44
III-Population d’étude ou Echantillonage ............................................................................ 45
III-1-Critères d’inclusion ...................................................................................................... 45
III-2-Critères de non inclusion ............................................................................................. 45
IV-Matériel ............................................................................................................................. 45

ix
IV-1-Les produits chimiques ................................................................................................ 45
IV-1-1- les produits chimiques fournis par la maison Sigma ........................................... 45
IV-1-2- les produits chimiques fournis par la maison Vel ................................................ 45
IV-1-3- les produits chimiques fournis par la maison Difco ............................................ 46
IV-2-Les souches bactériennes ............................................................................................. 46
V-Méthodes ............................................................................................................................. 46
V-1- Isolement et identification des souches bactériennes d’intérêt médical ...................... 46
V-1-2 - Examen à l’état frais ................................................................................................ 47
V-1-3 – Identification des souches bactériennes .................................................................. 47
V-2-Sensibilité aux antibiotiques et détection des BLSE ..................................................... 48
V-3-Condition de culture des souches .................................................................................. 50
V-4-Extraction des BLSE ..................................................................................................... 50
V-5-Hydrolyse des β-lactamines par les extraits bruts de BLSE ........................................ 51
V-6-Extraction des ADNs .................................................................................................... 52
V-7-Amplification des gènes de BLSE ............................................................................... 52
V-8-Electrophorèse sur gel d’agarose .................................................................................. 54
V-9-Séquençage des gènes de BLSE .................................................................................... 54
V-10-Numéros d’accession des séquences ........................................................................... 55
III-10-Traitement des données .............................................................................................. 55
CHAPITRE III : RESULTATS ............................................................................................ 57
I- Souches bactériennes ...................................................................................................... 57
II- Les BLSE détectées ......................................................................................................... 58
III- Les BLSE caractérisées .................................................................................................. 65

x
III-1- Le profil hydrolytique des céphalosporines de troisième génération (C3G) par les
BLSEs ................................................................................................................................... 65
III-2-Caractérisation des gènes codant pour la production des BLSEs ................................ 69
III-3- Séquençage des gènes bla ........................................................................................... 73
CHAPITRE IV : DISCUSSION ............................................................................................ 85
CONCLUSION ET PERSPECTIVES ................................................................................. 92
REFERENCES BIBLIOGRAPHIQUES ............................................................................. 94
ANNEXE A ........................................................................................................................... 117
ANNEXE B ........................................................................................................................... 125

xi
Liste des figures
Figure 1: paroi des bactéries à Gram-positif et à Gram-négatif ................................................ 7
Figure 2: Acide 7-aminocéphalosporanique ............................................................................ 20
Figure 3: Comparaison de la structure des pénicillines et des céphalosporines ....................... 21
Figure 4: Structure de quelques β-lactamines ......................................................................... 21
Figure 5: Voie catalytique générale des β-lactamases à sérine active ...................................... 27
Figure 6: Mécanisme d'hydrolyse de la pénicilline par la β-lactamase .................................... 28
Figure 7: Phénotype de résistance sur boîte de pétri des souches présumées productrices de
BLSE ........................................................................................................................................ 58
Figure 8: Effet de synergie d’action montrant le profil BLSE sur boîte de pétri ..................... 59
Figure 9: Répartition des pathogènes producteurs de BLSE isolés en fonction des espèces ... 60
Figure 10: Pourcentage des souches productrices de BLSE en fonction des services ............. 61
Figure 11: Courbe d’hydrolyse de la nitrocéfine (CPR) par l’extrait de la Klebsiella
pneumoniae 736. ...................................................................................................................... 66
Figure 12: Courbe d’hydrolyse du céfotaxime (CTX) par l’extrait de la Klebsiella pneumoniae
736. ........................................................................................................................................... 66
Figure 13: Courbe d’hydrolyse de la céfuroxime (CXM) par l’extrait de la Klebsiella
pneumoniae 736. ...................................................................................................................... 67
Figure 14: Courbe d’hydrolyse du céfépime (FEP) par l’extrait de la Klebsiella pneumoniae
736. ........................................................................................................................................... 67
Figure 15: Courbe d’hydrolyse de la ceftazidine (CTX) par l’extrait du Pseudomonas sp. 555.
.................................................................................................................................................. 68
Figure 16: Courbe d’hydrolyse de la ceftriaxone (CRO) par l’extrait de l’ E.coli 408 ........... 68
Figure 17: Courbe d’hydrolyse de l’imipénème (IPM) par l’extrait de l’E.coli 154. .............. 69
Figure 18: Gel d’agarose des produits PCR de CTX-M ......................................................... 70

xii
Figure 19: Gel d’agarose des produits PCR de
TEM…………………………………………………………………………………………..71
Figure 20: Gel d’agarose des produits PCR de SHV .............................................................. 71
Figure 21: Comparaison entre la protéine CTX-M-1 et la protéine CTX-M-15 de nos souches
.................................................................................................................................................. 76
Figure 22: Comparaison entre les séquences nucléotidiques des gènes SHV de nos isolats et
la séquence standard du gène SHV-1. ...................................................................................... 82
Figure 23: Comparaison entre la séquence de TEM-1 et la séquence de KPC-2 de nos
souches ..................................................................................................................................... 84

xiii
Liste des tableaux
Tableau I: Comparaison des points critiques nationaux des antibiotiques pour les
Entérobactéries ...................................................................................................................... 323
Tableau II: Protocoles de PCR suivant le type de gène ......................................................... 534
Tableau III: Séquences des amorces utilisées au niveau de cette étude ................................. 535
Tableau IV: Répartition des souches cliniques par espèces bactériennes ................................ 58
Tableau V: Répartition des souches cliniques par produits pathologiques ............................ 601
Tableau VI: Fréquence des souches productrices de BLSE selon le genre des bactéries
étudiées………………………………………………………………………………………………………....62
3
Tableau VII: Répartition des bactéries présentant le phénotype BLSE par produits
pathologiques et les CMI des antibiotiques. ................................. Erreur ! Signet non défini.4
Tableau VIII: Nombre et pourcentage des extraits en fonction de substrats hydrolysés ....... 656
Tableau IX: Répartition des gènes en fonction des échantillons............................................ 722
Tableau X: Répartition des gènes en fonction des services ................................................... 723
Tableau XI: Répartition des gènes en fonction des espèces ................................................... 733
Tableau XII: Caractéristiques moléculaires des gènes hébergés par les isolats traités .......... 744

xiv
Liste des abréviations
µM: micro molaire
AMC : Amoxicilline + acide clavulanique
AMX : Amoxicilline
AN : Amikacine
BLSE : β-lactamases à spectre élargi
C : Chloramphénicol
C2G : céphalosporines de deuxième génération
C3G : céphalosporines de troisième génération
CAZ : Ceftazidime
CF : céfalotine
Chir : chirurgie
CIP : Ciprofloxacine
CMI: concentration minimale inhibitrice
CO : Cotrimoxazole
CPR : Nitrocéfine
CRO: Ceftriaxone
CTX : céfotaxime
CTX-M : céfotaximase
CXM : Céfuroxime
E.coli : Escherichia coli
ESBL : Extented-spectral- β-lactamase
EXT : extérieur
FEP : Céfépime
FT : Furanes

xv
GE : Grand Enfant
GM (G) : Gentamicine
IPM : Imipénème
K.pneumoniae : Klebsiella pneumoniae
Kb : Kilo base
MBL : Metallo- β-lactamases
MI : Maladie Infectieuse
mL : mililitre
mM : milimolaire
Min : Minute
MONOD : Monodose
NA : Acide nalidixique
NET : Netilmicine
NOR : Norfloxacine
Nrs : nourrisson
Pb : paire de base
PBP : Penicillin-Binding Protein
PEF : Péfloxacine
PIP : Pipéracilline
Réa : Réanimation
SHV : Sulfhydril Variable
SXT : Triméthoprime-sulfamides
TEM : Temoniera
TIC : Ticarcilline
TS : Cotrimoxazole
UC : Urgence Chirurgicale

xvi
UM : Urgence Médicale
Vo : Vitesse initiale (à l’instant proche de zéro)
ΔA : Variation d’Absorbance

xvii
Résumé
Introduction : les bactéries productrices de β-lactamases à spectre élargi (BLSE) ont été
rapportées dans plusieurs pays, mais aucune information n’est disponible sur les différents
types de BLSE produites par les entérobactéries au Burkina Faso. Pour palier au déficit
d’informations scientifiques d’une extrême importance, notre objectif principal a été de
caractériser les différents types de β-lactamases à spectre élargi (BLSE) produites par les
bactéries à Gram-négatif isolées des patients au Centre Hospitalier Universitaire Pédiatrique
Charles De Gaulle (CHUP-CDG) de Ouagadougou.
Méthodologie : les méthodes de disque et de dilution ont permis l’étude de la sensibilité in
vitro à des antibiotiques ciblés de Cent quatre vingts(180) souches de bactéries à gram négatif
résistantes à au moins une céphalosporine de troisième génération entre 2010-2012. En vue de
détecter les espèces bactériennes productrices de BLSE, les souches ont été soumises au test
de double synergie puis à des analyses cinétiques. Les extraits enzymatiques bruts des
souches ont été obtenus par le cycle de congélation/décongélation. Les génomes des
différentes souches ont été obtenus par extraction au DNAZOL. La caractérisation
moléculaire des BLSE s’est basée sur la technique de la PCR classique et du séquençage à
l’aide d’amorces spécifiques des gènes CTX-M, SHV et TEM.
Résultats : l’imipénème, la ceftazidime et le céphépime sont les antibiotiques les plus actifs
sur les différentes bactéries. Les différentes souches présentent un niveau de résistance élevé à
la céfuroxime, au céfotaxime et à la cetriaxone. Cent vingt et un (121) souches ont montré un
test positif à la production de BLSE. La prévalence des souches productrices de BLSE est de
67,22%. Ces souches présentant un phénotype BLSE sont observées chez toutes les espèces
de bactéries étudiées avec une prévalence de 69,38% (Klebsiella spp.), 75% (Enterobacter
spp.), 65,88% (E. coli), 33,33% (Proteus spp.), 58,33% (Pseudomonas spp.) et de 66,66%
(Salmonella spp.).

xvii
i
Les extraits d’ADN des 121 souches soumis à la technique de PCR classique, en utilisant les
amorces spécifiques des gènes blaSHV, blaCTX-M et blaTEM ont montré qu’ils contiennent les
gènes des β-lactamases du type SHV (sulfhydryl variable), CTX-M (cefotaximase) et TEM
(Temoniera). L’analyse des séquences des différents produits PCR a permis de montrer que
nos souches hébergent des β-lactamases de type: TEM-1, OKP-B, SHV-1, SHV-11, SHV-12,
SHV-28, SHV-32, SHV-38, SHV-76, SHV-99, CTX-M-15 et une carbapénèmase de type
KPC-2 (hébergée par le plasmide pKPN101-IT).
Conclusion : il a été mis en évidence des β-lactamases de types CTX-M-15, SHV-1,-11,-12,-
28,-32,-38,-76 et -99, TEM-1et KPC-2 au Centre Hospitalier Universitaire Pédiatrique
Charles De Gaulle (CHUP-CDG) de Ouagadougou, Burkina Faso. Vu les résultats obtenus,
des mesures doivent être prises pour la prevension et la lutte contre les bactéries productrices
de ces enzymes.
Mots clés : β-lactamase, β-lactamase à spectre élargi, bactéries à gram négatif et Centre
Hospitalier Universitaire Pédiatrique Charles De Gaulle.

xix
ABSTRACT
Introduction: Bacteria producing extended-spectrum β-lactamase (ESBL) have been reported
in many countries, but there is lack of data on different type of ESBL-producing
enterobacteria in Burkina Faso. For stage with the deficit of scientific information of extreme
importance, our principal objective was to characterize the various types of extended-
spectrum-β-lactamase (ESBL) produced by the negative Gram bacteria isolated from the
patients in the University Hospital complex Paediatric Charles De Gaulle of Ouagadougou.
A total of 180 strains of bacteria with negative gram were isolated between 2010-2012 from
pathological products (urines, pus, blood, saddles and céphalo-rachidian liquid) of patients at
the University Hospital complex Paediatric Charles De Gaulle (CHUP-CDG) in Burkina
Faso.
Methodology : agar disk diffusion and dilution methods permitted the study of in vitro
susceptibility of the bacteria to antibiotics. To identify ESBL -producing bacterial species,
the strains were subjected to the test of double synergy and then kinetic analyzes. The crude
enzyme extracts of the strains were obtained from the cycle of freezing / thawing. The
genomes of different strains were obtained by extraction with DNAzol. Molecular
characterization of ESBL was based on the conventional PCR technique and sequencing using
primers specific for CTX -M, SHV and TEM genes.
Results: imipenem, ceftazidim and cephepim are the most active antibiotics on different
bacteria. Different strains have a high level of resistance to cefuroxim, cefotaxim and
cetriaxon. One hundred twenty-one (121) strains showed a positive test of ESBL production .
The prevalence of ESBL-producing strains is 67.22 %. These strains with ESBL phenotype
observed in all species studied bacteria with a prevalence of 69.38 % ( Klebsiella spp. ), 75 %
( Enterobacter spp.) , 65.88 % (E. coli) , 33.33 % ( Proteus spp.) , 58.33% (Pseudomonas
spp.) and 66.66% (Salmonella spp.). DNA extracted from 121 strains subjected to the
standard PCR technique using specific primers of genes blaSHV , blaCTX-M and blaTEM

xx
showed that they contain the β-lactamase gene SHV type ( variable sulfhydryl ) CTX -M (
cefotaximase ) and TEM ( Temoniera ). Sequence analysis of the different PCR products
showed that our strains harboring β-lactamases type TEM -1, OKP -B , SHV- 1 , SHV- 11 ,
SHV- 12 , SHV- 28 , SHV- 32, SHV- 38, SHV- 76, SHV- 99 , CTX -M- 15 and a
carbapenemase KPC- 2 type ( hosted by the plasmid pKPN101-IT) .
Conclusion: It has been revealed β-lactamase types CTX- M-15 , SHV- 1 , -11 , -12, -28 , -32
, -38 , -76 and -99 , TEM- 1 and KPC- 2 in the University Hospital complex Paediatric
Charles De Gaulle of Ouagadougou/BF. Considering the results obtained, measures must be
taken for the prevension and the fight against the producing bacteria of these enzymes.
Key words: antibiotic, β-lactamase, extended-spectrum-β-lactamase, bacteria, CTX-M, SHV,
TEM and University Hospital complex Paediatric Charles De Gaulle.

Introduction
1
INTRODUCTION
La résistance se définit par la propriété qu’a une cellule bactérienne de se développer
en présence d’une concentration élevée d’un antibiotique. La résistance aux antibiotiques est
un problème mondial mais qui se manifeste différemment selon les pays. À tous les niveaux,
national et international, recherche et prévention s’organisent (Kunin et al., 1990). Les β-
lactamases sont des enzymes produites par certaines bactéries et sont responsables de leurs
résistances aux antibiotiques appartenant à la famille des β-lactamines.
Les β-lactamines représentent la principale famille d’antibiotiques la plus développée et la
plus utilisée dans le monde. Cette large utilisation est due à leur large spectre d’action, leur
faible toxicité, leur efficacité et à leur faible coût pour certaines molécules (Livermore, 1995).
Les bactéries ont développé différents mécanismes pour contrecarrer l'action des β-
lactamines, entre autre :
- la modification de la cible (Protéines liants Pénicillines) qui les rend moins sensibles
aux β-lactamines mais permet de maintenir son activité physiologique normale ;
- la synthèse des enzymes (β-lactamases) qui inactivent les β-lactamines par
modification chimique ;
- l’acquisition ou la surproduction des pompes efflux qui peuvent expulser l’antibiotique
hors de la cellule même contre le gradient de concentration
- et la modification des porines chez les bactérie à Gram-négatif, ayant pour résultat la
diffusion plus lente des β-lactamines à travers la membrane externe (Galleni et al.,
1995 ; Nikaido, 1998 ; Lakaye et al., 1999 ; Walsh, 2003).
Les β-lactamases catalysent de manière efficace et irréversible l’hydrolyse de la liaison amide
du cycle β-lactame des antibiotiques de la famille des β–lactamines, donnant un produit
biologiquement inactif qui perd totalement son activité antimicrobienne (Matagne et al.,
1998). Les gènes qui codent pour ces enzymes sont d'origine chromosomique ou plasmidique.

Introduction
2
Ces gènes ont aussi été détectés sur des transposons et des integrons facilitant ainsi leur
transfert horizontal entre espèces phylogénétiquement éloignées. Ces enzymes sont exportées
dans le milieu extracellulaire (bactéries à Gram positif) ou périplasmique (bactéries à Gram
négatif). La production des β-lactamases est le mécanisme de résistance le plus répandu et le
plus important des bactéries vis-à-vis des β-lactamines (Livermore, 1995).
Plus de 290 types de β-lactamases sont décrits (www.lahey.org/studies/inc_webt.html) et
classés suivant leur sutructure primaire en 4 classes (A-D) (Ambler, 1980). Une autre
classification basée sur les propriétés fonctionnelles (profil du substrat et susceptibilité aux
inhibiteurs des β-lactamases) a été donnée (BUSH et al., 1995). La plus grande partie des β-
lactamases a été mise en évidence chez les bacilles à Gram négatif qui sont par ailleurs
responsables de la majorité des infections hospitalières (60%) et sont de plus en plus
multirésistants (Bergone-Berezin et al., 1993 ; Bush, 1995 ; Archibald et al., 1997). La
dissémination des β-lactamases communes et l'apparition de nouvelles enzymes ou d'enzymes
mutants sont reliées directement à l'utilisation abusive des β-lactamines dans le domaine de la
santé humaine et animale et en agriculture (Bibbal, 2008 ; Faure, 2009).
Les BLSE constituent un groupe d'enzymes qui ont la propriété commune de conférer la
résistance aux pénicillines, à la 1ère, 2ème, 3ème et 4ème
génération de céphalosporines, à
l’aztréoname (mais non aux céphamycines et carbapénème) par hydrolyse de ces
antibiotiques, et qui sont inhibées par les inhibiteurs des β-lactamases tel l’acide clavulanique
(Bush et al., 1995).
La présence des BLSE a été décrite pour la première fois chez Klebsiella ozaenae, en 1983 en
République Fédérale d'Allemagne, puis en 1984 chez K. pneumoniae et E. coli en France et en
Tunisie (Knothe et al., 1983; Kliebe et al, 1985). Des travaux portant sur la production des
BLSE par les Entérobactéries en Afrique sub-saharienne sont rares et éparses. Au Burkina
Faso, la majorité de la population n’a pas accès aux médicaments appropriés pour les soins.

Introduction
3
Outre ce déficit de couverture sanitaire, la résistance des bactéries aux antibiotiques constitue
un facteur aggravant. A l’heure actuelle, les données locales sur la résistance bactérienne sont
peu nombreuses d’où l’insuffisance d’informations pouvant orienter la conduite à tenir en
présence de ces microorganismes dans les établissements de soins de santé dans notre pays.
Sur le plan bibliographique, on note qu’à l’échelle du Burkina, les travaux d’investigation sur
les β-lactamases sont relativement récents (Zeba et al., 2003 ; Zeba et al., 2004 ; Zeba et al.,
2005a ; Zeba et al., 2005b ; Zeba et al, 2007a ; Zeba et al., 2007b ; Mètuor-Dabiré et al.,
2013).
L’acquisition des données sur la résistance bactérienne aux antibiotiques est nécessaire pour
une meilleure prise en charge thérapeutique des infections et pour élaborer une stratégie de
contrôle de la résistance antimicrobienne. Pour palier au déficit d’informations scientifiques
d’une extrême importance, les présentes investigations ont été entreprises.
Dans cette étude, notre objectif principal a été de caractériser les différents types de β-
lactamases à spectre élargi (BLSE) produites par les bactéries à Gram-négatif isolées des
patients au Centre Hospitalier Universitaire Pédiatrique Charles De Gaulle (CHUP-CDG) de
Ouagadougou.
Les objectifs spécifiques ont consisté à:
- Décrire les phénotypes de résistance ;
- Déterminer les souches de bactéries à Gram-négatif productrices de BLSE ;
- Identifier les grands groupes de BLSE ;
- Identifier les sous-groupes de BLSE
Ce travail est structuré en quatre chapitres : le premier chapitre consiste en une synthèse
bibliographique, le deuxième chapitre concerne le matériel et les méthodes utilisés, le
troisième chapitre présente l’ensemble des résultats obtenus et le quatrième chapitre traite les

Introduction
4
discussions qui en découlent. Nous terminons ce travail par une conclusion et des
perspectives de recherches à venir.

Synthèse bibliographique
5
CHAPITRE I : SYNTHESE BIBLIOGRAPHIQUE
I- Bactéries
I-1-Définition
La bactérie est un micro-organisme ubiquiste, unicellulaire et sans noyau (procaryote)
dont le génome est constitué d'ADN. Celui-ci consiste en un seul chromosome et on
note éventuellement la présence de plasmides. L'ensemble des bactéries forme le règne des
protistes inférieurs et le domaine des Bacteria.
I-2-paroi bactérienne
I-2-1-Structure
La paroi bactérienne est une structure unique qui entoure la membrane cellulaire. Bien
que n’étant pas présente chez toutes les espèces bactériennes, la paroi est une structure
cellulaire très importante. Elle joue plusieurs rôles (Nanninga, 1998) :
- maintient la forme de la cellule ;
- préserve l’intégrité de la cellule en contrant l’effet de la pression osmotique
interne ;
- intervient dans la division cellulaire ;
- offre les sites de fixation pour les virus comme les bactériophages ;
- sert enfin de support pour les extensions de surface comme les pili, les flagelles
et les fimbriae qui émanent de la paroi et se prolongent au-delà d’elle.
La structure de la paroi de toutes les bactéries n’est pas identique. En fait sa
composition constitue un critère très important dans l’analyse et la différentiation des espèces
bactériennes. On distingue deux types majeurs de paroi bactérienne : la paroi des bactéries à
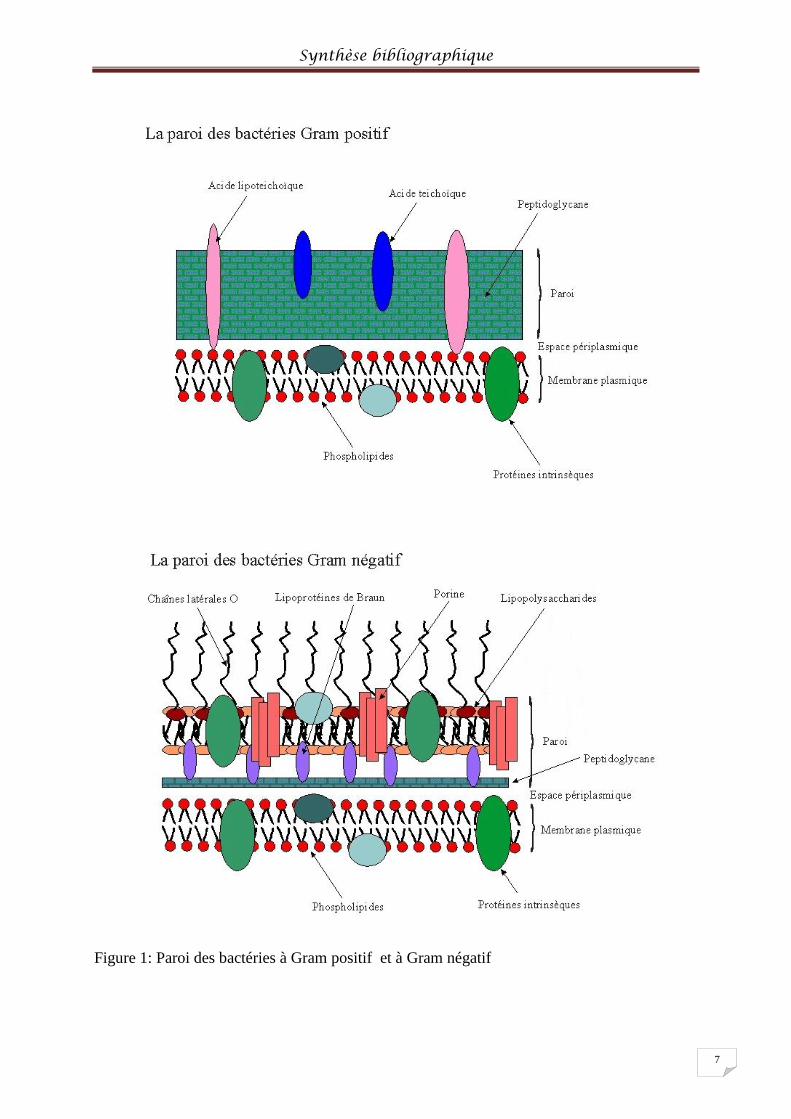
Gram positif et celle des bactéries à Gram négatif. La paroi des bactéries à Gram-positif est

Synthèse bibliographique
6
composée par une épaisse couche de peptidoglycane (muréine) sur laquelle sont ancrés les
polysaccharides acides (acide téchoïque). La paroi des bactéries à Gram-négatif quant à elle,
est caractérisée par une couche de muréine qui est située entre la membrane externe et la
membrane cytoplasmique, donc dans le périplasme (espace entre la membrane externe et la
membrane cytoplasmique). Le peptidoglycane est relié à la membrane externe par des
lipoprotéines. Ainsi, la membrane externe (sur laquelle sont fixés les polysaccharides)
possède à sa face interne les lipoprotéines et à sa surface externe les lipopolysaccharides (Van
Hoof, 2001) (Figure1)
Synthèse bibliographique
7
Figure 1: Paroi des bactéries à Gram positif et à Gram négatif

Synthèse bibliographique
8
Source : (http://www.bacterio.cict.fr/bacdico/bacteriogene/structure.html#morphologie)
(10 /03/2013)
I-2-2-Propriétés de la paroi
Le peptidoglycane fonctionne comme une passoire moléculaire et ne représente pas de
véritable barrière de perméabilité pour la plupart des antibiotiques mais la membrane
plasmique est une barrière importante. La paroi des bactéries à Gram-négatif du fait de la
présence de la membrane externe (bicouche lipidique, présence des porines) est moins
perméable que celle des bactéries à Gram-positif (Van Hoof, 2001).
Bien que la composition chimique du peptidoglycane soit variable entre les groupes
bactériens, son architecture de base est conservée. Le peptidoglycane est un réseau des
chaînes glycanes non ramifiées composés de sous-unités de saccharides réticulées avec des
courts peptides (tétrapeptides). Contrairement à la chitine, le peptidoglycane n’est pas une
structure cristalline, mais un polymère dont les propriétés élastiques résident dans la liberté
conformationelle des liaisons peptidiques (Van Hoof, 2001).
I-2-3-Biosynthèse de la paroi
La biosynthèse du pepdidoglycane se déroule en deux étapes (Van Heijenoort, 2001) :
La première étape consiste en la synthèse des monomères grâce aux enzymes localisées dans
le cytoplasme ou sur la face interne de la membrane cytoplasmique. Le produit final de cette
étape est un intermédiaire lipidique undecaprenol (Lipide II). Ce produit est transporté à
travers la membrane plasmique par un mécanisme non encore élucidé vers la surface externe
de la membrane où il sert de substrat initial aux réactions de polymérisation qui auront lieu
pendant la deuxième étape. La deuxième étape de synthèse du peptidoglycane se résume en la
polymérisation des monomères et l’incorporation du nouveau matériel dans le pepdidoglycane
préexistant. L’assemblage final des monomères pendant la seconde phase, peut se diviser en 5

Synthèse bibliographique
9
groupes de réaction : formations de l'uridine 5'-pyrophosphate- N – acétylglucosamine (UDP-
GlcNAc), de l’uridine 5'-pyrophosphate-N - acide acétyl-muramique (UDPMurNAc), de
l’UDP-MurNAc-peptides et des intermédiaires lipidiques. La polymérisation du résidu C55-
disaccharide-pentapeptide pour constituer le pepdidoglycane fait intervenir deux types
majeurs de réactions enzymatiques :
- les glycosyltransférases qui catalysent la formation de la chaîne polysaccharidique linéaire;
- les transpeptidases qui catalysent la formation des liaisons peptidiques croisées assurant
ainsi la réticulation de l’hétéropolymère. Cette étape de la synthèse du peptidoglycane
représente une cible privilégiée pour de nombreux antibiotiques (β-lactamines, glycopeptides,
bacitracine). Ces agents antimicrobiens sont capables d’inhiber la biosynthèse de la muréine
en inhibant l’étape 2 qui est plus facilement accessible car se réalisant à la surface externe de
la membrane.
La transpeptidation et la transglycosylation sont effectuées par des protéines bactériennes
uniques appelées les Protéines-Liant-Pénicilline (PLP) ou tout simplement Penicilline-
Binding-Proteins (PBPs). Ce sont des protéines qui appartiennent à la famille des protéines à
sérine active. Elles sont encore appelées DD-pepdidases (Frère et Joris, 1985).
I-3-Protéines-Liant-Pénicilline (PLP)
Les cibles des β-lactamines sont des protéines enzymatiques insérées sur la face externe de la
membrane cytoplasmique, dénommées Protéines-Liant-Pénicilline (PLP). Les PLP
interviennent dans la synthèse et le remodelage du peptidoglycane, constituant principal de la
paroi bactérienne (Ghuysen, 1991 ; Nanninga, 1991).
Le nombre et la nature des PLP varient selon les espèces bactériennes. Les travaux de Spratt
et Pardee (1975) ont permis de séparer six de ces protéines et de les classer en deux grands
groupes selon leur poids moléculaire. Ainsi on distingue les PLP de haut poids moléculaire

Synthèse bibliographique
10
(PLP 1 à 3) et celles de faible poids moléculaire (PLP 4 à 6). Les PLP de haut poids
moléculaire sont des molécules multidomaines, constituées d’une courte région
cytoplasmique se résumant au domaine transmembranaire et d’une longue partie
périplasmique. Ces PLP de fort poids moléculaire se subdivisent en deux groupes de
molécules, le sous-groupe A (possède les activités de transglycosylation et de
transpeptidation) et le sous-groupe B qui contient un domaine transpeptidase ajouté au
domaine N-terminal à fonction inconnue. Les PLP de faible poids moléculaire jouent un rôle
dans la catalyse de l’hydrolyse du peptidoglycane (carboxypeptidases) régulant ainsi le
nombre de liaisons dans la muréine (Ghuysen, 1991 ; 1994). Chez Escherichia coli, la PLP1
intervient dans l’élongation de la cellule, la PLP2 dans la forme de la cellule et la PLP3 dans
la formation des septa (Spratt, 1975). Chaque espèce bactérienne possède au moins trois et
jusqu’à plus de huit PLPs (Hakenbeck et Coyette, 1998). Les PLPs sont des cibles
physiologiques des antibiotiques de la famille des β-lactamines. Ces cibles létales des
bactéries sont des enzymes à sérine active, qui réalisent leur cycle de catalyse par des
mécanismes d’acylation/désacylation avec des intermédiaires acylenzymes. Ce mécanisme est
schématiquement représenté part un modèle à trois étapes (Ghuysen et al., 1986) :
1.1
E est l’enzyme, D le substrat donneur du carbonyl (D-alanyl-D-alanine), HY accepteur
nucléophile, P1 et P2 les produits de la réaction de catalyse, E•D le complexe non covalent de
Henri-Michaelis, E-D* complexe covalent acyl-enzyme.
II-Antibiotiques
II-1- Généralités

Synthèse bibliographique
11
Les antibiotiques (du grec anti : contre, et bios : la vie) sont des substances chimiques
qui ont une action spécifique avec un pouvoir destructeur sur les micro-organismes. Elles sont
dépourvues de toxicité pour les autres cellules. En général, le terme antibiotique est employé
pour désigner une molécule qui a une action négative sur les bactéries. Ces molécules peuvent
avoir une action drastique, c’est-à-dire bactéricide où leur efficacité peut être limitée à
empêcher le développement des micro-organismes (action bactériostatique). Lorsque l’action
négative des molécules est exercée sur les champignons, on emploie le terme d’antifongiques
et lorsqu’elle est exercée sur les virus on parle d’antivirales. L’action antibiotique est
normalement ciblée. Elle se distingue d’un antiseptique qui détruit tout germe et parfois
même la cellule hôte, de manière non ciblée. Le premier antibiotique identifié fut la
pénicilline découverte par hasard (Fleming, 1929). Ce premier antibiotique a ouvert une voie
nouvelle dans la lutte contre de nombreuses maladies qui étaient considérées comme
incurables auparavant. Suite à la découverte de la pénicilline, de nombreuses autres
générations d’antibiotiques ont vu le jour et continuent d’apparaître dans le but de combattre
les microorganismes qui résistent aux précédents.
Les antibiotiques ont des effets à faibles concentration et ont un métabolisme relativement
lent (5 à 6 heures).
Ils peuvent être d’origine naturelle, hémi-synthétique, ou purement synthétique. Ils sont
soumis dans l’organisme humain à un ensemble de processus d’absorption, de diffusion et
d’élimination : c’est la pharmacocinétique. Cet ensemble de processus confère des
caractéristiques pharmaco-cinétiques (pic sérique, temps de ½ vie, voie d’élimination, etc…)
particulières à chacun des antibiotiques.
Un antibiotique a un spectre d’activité théorique (naturel) : avant tout emploi en thérapeutique
il est d’une part actif sur un ensemble d’espèces bactériennes (souches sauvages sensibles) et
d’autre part inactif sur un certain nombre d’espèces (souches sauvages résistantes). Tous les
antibiotiques n’ont pas le même spectre d’activité théorique : certains agissent sur un grand

Synthèse bibliographique
12
nombre d’espèces bactériennes, leur spectre est dit « large » ; d’autres agissent sur un nombre
restreint d’espèces bactériennes, leur spectre est dit « étroit ».
Au cours de leur utilisation, les antibiotiques sélectionnent les souches bactériennes qui leur
sont résistantes : les unes leur sont naturellement résistantes (souches sauvages résistantes
naturelles) et les autres leur sont devenues résistantes secondairement par acquisition de gènes
de résistance (souches résistantes acquises) à la suite de modifications de leur génome par
mutation ou transfert de gènes ; cet effet sélectif modifie le spectre d’activité des antibiotiques
: le spectre d’activité théorique d’un antibiotique est remplacé au cours des années par un
spectre d’activité actualisé (celui qui est effectif au moment de la prescription) ;
Les antibiotiques sont répartis en familles qui réunissent les molécules qui ont une
structure chimique de base identique et possèdent un même mécanisme d’action antibactérien.
Les antibiotiques d’une même famille peuvent se différencier par leur spectre
d’activité ; ils sont alors réunis par groupes de spectre identique quelquefois subdivisés en
sous-groupes ; les antibiotiques d’un même groupe ou d’un même sous-groupe peuvent
différer par leurs propriétés pharmaco-cinétiques.
II-2-Effet antibactérien
L’action d’un antibiotique sur la population de bactéries d’une souche bactérienne se
manifeste, pour une concentration déterminée, par un effet anti-bactérien qui peut être soit nul
soit une bactériostase soit une bactéricidie.
Un effet anti-bactérien nul ne provoque aucun changement de la croissance bactérienne.
La bactériostase consiste en un ralentissement de la croissance de la population bactérienne
pouvant aller jusqu’à une absence de croissance.
L’activité bactériostatique d’un antibiotique sur la population d’une souche bactérienne est
indiquée par la détermination de la mesure de la CMI (concentration minimale inhibitrice).

Synthèse bibliographique
13
Cette CMI entraîne un ralentissement de la croissance de la population bactérienne jusqu’à
atteindre une absence de croissance.
La CMI est définie comme la plus faible concentration d’un antibiotique donné capable
d’interrompre, dans un milieu et à des conditions parfaitement définis, toute croissance visible
d’une souche bactérienne donnée.
La bactéricidie consiste en la destruction d’une partie de la population bactérienne d’une
souche bactérienne.
L’activité bactéricide d’un antibiotique sur une souche bactérienne est indiquée par la
détermination de la mesure de la CMB (concentration minimale bactéricide).
La CMB d’un antibiotique pour une souche bactérienne donnée est définie comme la plus
faible concentration de cet antibiotique permettant une réduction du nombre de survivants de
la population de cette souche au moins égale à 10-4
bactéries/ml d’un inoculum de 106
bactéries/ml (soit 100 survivants/ml sur 1.000.000 de bactéries ensemencées/ml c’est à dire 1
survivant sur 10.000 bactéries de l’inoculum) après 18h de culture à 37° de cette souche en
présence de l’antibiotique.
L’activité bactéricide d’un antibiotique peut être dépendante du temps (antibiotiques
bactéricides temps dépendant) ou être dépendante de la concentration (antibiotiques
bactéricides concentration dépendant).
L’effet post-antibiotique est l’effet anti-bactérien observé après suppression du contact de la
population bactérienne avec l’antibiotique.
La tolérance est un état acquis par une partie de la population d’une même souche bactérienne
dans laquelle un antibiotique réputé bactéricide reste bactériostatique mais perd son effet
bactéricide.
II-3-Sites d’action et propriétés d’activité
Un antibiotique ne peut être actif que si ses molécules atteignent leur cible.
II-3-1-Antibiotiques agissant sur la synthese du peptidoglycane

Synthèse bibliographique
14
Les bêta-lactamines
Elles sont réparties en 4 sous familles : les pénicillines, les céphalosporines, les
monobactames et les carbapénèmes.
Elles se fixent préférentiellement sur certaines des protéines de liaison aux pénicillines (PLP)
qui sont des enzymes de la phase terminale de la synthèse du petidoglycane (transpeptidases,
carboxypeptidases) catalysant les liaisons entre les chaînes peptidiques dans la paroi des
bactéries. Les bêta-lactamines jouent le rôle d’un substrat formant une liaison stable avec
certaines PLP et bloquent l’action de ces dernières. Ce sont des produits bactéricides temps
dépendants.
Les glycopeptides
Ils ont pour cible : l’undécaprényl-phosphate (UDP), qui est un transporteur trans-
membranaire des précurseurs du peptido-glycane : la chaîne de peptido-glycane en formation,
les peptides de la paroi non encore couplés.
Ils sont bactéricides temps dépendants lents.
La fosfomycine
Elle inhibe une des phases cytoplasmiques de la synthèse de la paroi en bloquant une pyruvyl-
transférase ; Elle est bactéricide
II-3-2-Antibiotiques agissant sur les membranes (externe et cytoplasmique)
Les polymyxines
Elles se fixent sur les phospholipides membranaires ; les membranes ne peuvent plus se
remanier, se déforment et deviennent perméables.
Elles sont bactéricides mais diffusent mal dans les tissus.
II-3-3-Antibiotiques agissant sur l’appareil nucleaire
Les sulfamides et le triméthoprime
Ils agissent sur des enzymes de la voie de synthèse de l’acide folique et des folates, qui sont
des cofacteurs de la synthèse des acides nucléiques ; les sulfamides agissent sur la

Synthèse bibliographique
15
dihydroptéroate-synthétase ; le triméthoprime agit sur la dihydrofolate réductase. Ils sont
bactéricides.
Les quinolones
Elles sont réparties en deux groupes : les quinolones et les fluoroquinolones larges. Elles
agissent sur des enzymes réglant la conformation de l’ADN, les topo-isomérases
(essentiellement les topo-isomérases II ou ADN gyrases). Elles sont bactéricides. Elles
atteignent de bonnes concentrations intracellulaires dans les cellules eucaryotes.
Les rifamycines
Ce sont des produits inhibant la synthèse des ARN messager par inhibition de l’ARN
polymérase ADN dépendante. Elles sont bactéricides et surtout utilisées pour traiter la
tuberculose. Elles atteignent de bonnes concentrations intracellulaires dans les cellules
eucaryotes.
Les nitro-imidazolés
Réduits en dérivés actifs en atmosphère strictement anaérobie, ils forment un complexe avec
un brin d’ADN provoquant une coupure de ce dernier ; Ils sont bactéricides.
II-3-4-Antibiotiques agissant sur les ribosomes
Les phénicols
Ils se fixent sur le ribosome au niveau du site amino-acyl et inhibent l’élongation de la chaine
peptidique. Ils sont bactériostatiques ; actuellement ils sont très peu employés car ils sont
toxiques sur la moelle osseuse.
Les tétracyclines
Elles se fixent sur le ribosome au niveau du site aminoacyl mais aussi au niveau du site
peptidyl quand les molécules d’acyl-tARN fixées antérieurement sont nombreuses.
Elles sont bactériostatiques et ont de bonnes concentrations intracellulaires dans les cellules
eucaryotes.

Synthèse bibliographique
16
Les macrolides, lincosamides et synergistines
Ces produits se fixent sur la sous-unité 50S du ribosome. Les macrolides et les lincosamides
sont bactériostatiques ; les synergistines sont bactéricides. Ils atteignent de bonnes
concentrations intracellulaires dans les cellules eucaryotes.
L’acide fusidique
Il se fixe sur le site aminoacyl et bloque la translocation de la chaine peptidique en formation.
Il est bactériostatique.
Les aminosides
Ils se fixent irréversiblement au niveau de la sous-unité 30S du ribosome ; ce sont des
inhibiteurs de la traduction : ils provoquent des erreurs de lecture du message porté par l’ARN
messager. Les aminosides sont de puissants bactéricides concentration-dépendants.
II-3-5-Antibiotiques agissant sur les acides mycoliques (antituberculeux)
L’isoniazide
Il inhibe la synthèse des acides mycoliques, constituants essentiels de la paroi des
mycobactéries. Il est bactéricide sur les bacilles à multiplication active et sur les bacilles
phagocytées.
L’ethambutol
Il inhibe la fixation à la paroi des acides mycoliques nouvellement constitués. Il est
bactériostatique sur les bacilles à multiplication active et sur les bacilles phagocytées.
Le pyrazinamide
Il est actif uniquement sur les bacilles phagocytées.
II-4-Méthodes d’étude « in vitro » de l’activité des antibiotiques
Le choix d’un traitement antibiotique ne peut être fait uniquement sur la base du
spectre théorique des antibiotiques en raison de la possibilité d’acquisition de résistance aux
antibiotiques par les espèces bactériennes.

Synthèse bibliographique
17
Le laboratoire peut préciser in vitro quels sont, aux doses utilisables in vivo, les antibiotiques
actifs sur la souche bactérienne responsable de l’infection.
II-4-1-Etude de la bacteriostase
Détermination de la CMI
Des méthodes biologiques permettent de déterminer cette valeur pour chacun des
antibiotiques vis à vis d’une souche bactérienne donnée ; la détermination de la CMI par la
méthode de dilution en milieu liquide est la plus facile ; une méthode par diffusion en milieu
solide utilisant des bandelettes imprégnées de gradients de concentration d’antibiotique
(epsilomètre ou E-test) existe maintenant.
La confrontation entre la CMI d’un antibiotique mesurée pour une souche bactérienne donnée
et les concentrations habituelles connues de cet antibiotique dans le sérum de l’organisme
humain lors des traitements antibiotiques permet de qualifier la souche bactérienne étudiée de
« sensible » ou de « résistante » ou de « intermédiaire » à l’antibiotique étudié:
Une souche bactérienne est dite sensible (S) à un antibiotique quand la CMI de cet
antibiotique pour cette souche est nettement inférieure aux concentrations usuelles de cet
antibiotique obtenues in vivo lors d’un traitement par cet antibiotique administré aux doses
usuelles.
Une souche bactérienne est dite résistante (R) à un antibiotique quand la CMI de cet
antibiotique pour cette souche est supérieure aux concentrations maximales de cet antibiotique
qui peuvent être atteintes in vivo lors d’un traitement par cet antibiotique administré aux doses
maximales autorisées.
Une souche bactérienne est dite de sensibilité intermédiaire (I) à un antibiotique quand la CMI
de cet antibiotique pour cette souche est inférieure mais proche des concentrations usuelles de
cet antibiotique obtenues in vivo lors d’un traitement par cet antibiotique administré aux doses
usuelles.

Synthèse bibliographique
18
En thérapeutique une marge de sécurité entre la CMI et la concentration in vivo est
indispensable pour envisager un succès thérapeutique ; elle est appréciée par le quotient
inhibiteur : résultat du rapport entre la concentration de l’antibiotique dans le sérum sur la
valeur de la CMI de la souche bactérienne responsable de l’infection ; il est admis
empiriquement qu’un traitement antibiotique assure un succès thérapeutique si le quotient
inhibiteur est égal ou supérieur à 8.
Antibiogramme
L’antibiogramme est une technique simplifiée d’appréciation de l’activité bactériostatique des
antibiotiques sur une souche bactérienne.
Il peut être réalisé par différentes techniques manuelles ou semi-automatisées ; le
résultat peut être obtenu dans un délai de 4 à 48 h selon la technique employée ; la technique
de diffusion en milieu gélosé à l’aide de disques d’antibiotiques encore appelée « méthode des
disques » est la méthode de référence; Quelle que soit la technique utilisée, elle a l’avantage
de permettre de tester plusieurs antibiotiques simultanément avec un minimum de
manipulations ; elle ne permet pas de déterminer des valeurs précises de CMI mais fournit le
même type d’interprétation que celle de la détermination de la CMI : souche sensible (S ),
souche résistante (R), souche intermédiaire (I)
II-4-2-Etude de la bactericidie
Détermination de la CMB
Elle est possible mais rarement effectuée ;
Elle est réalisée à partir de chacun des tubes qui ont permis de déterminer la CMI ; elle
consiste, juste après la détermination de la CMI, à ensemencer un aliquote de chacun des
tubes de la détermination de la CMI sur un milieu solide sous forme d’une strie individuelle et
à faire incuber le milieu portant les différentes stries à 37° pendant 18 heures ; puis à
comparer le nombre de colonies apparues sur chacune des stries au nombre de colonies
observées sur des stries obtenues de façon identique à partir de chacun des aliquotes de 5

Synthèse bibliographique
19
tubes de dilution de raison 10 (100
, 10-1
, 10-2
, 10 -3
, 10- 4
) d’une gamme de référence établie à
partir de la suspension bactérienne utilisée comme inoculum pour la détermination de la CMI.
La CMB d’un antibiotique pour une souche bactérienne donnée est définie comme la
plus faible concentration de cet antibiotique ne laissant subsister qu’un nombre de survivants
inférieur ou égal de 10-4
bactéries/ml d’un inoculum initial de 106
bactéries/ml de cette souche
( soit 1 survivant sur 10.000 bactéries ensemencées ) après 18 heures de culture à 37° de cette
souche en présence de l’antibiotique.
Détermination du rapport CMB/CMI
Ce rapport est utilisé pour distinguer les antibiotiques bactéricides (CMB/CMI < 2) des
antibiotiques bactériostatiques (CMB très éloignée de la CMI). Il permet de définir également
la tolérance d’une souche bactérienne à un antibiotique bactéricide (CMB/CMI > 32).
II-5- β-lactamines
II-5-1- Définition et structure chimique
Les β-lactamines sont des antibiotiques qui possèdent un noyau β-lactame qui est la
partie efficace de la molécule. Des variations au niveau de la chaîne latérale permettent de
modifier les propriétés de la molécule antibiotique. Par exemple, une modification des chaînes
latérales entraîne une résistance aux acides (suc gastrique) et permet l'administration de
l'antibiotique par voie orale. Les β-lactamines ou antibiotiques à noyau β-lactame sont une
large classe d'antibiotiques qui comprennent les pénames, les pénèmes, les céphèmes, les
monobactames et les oxapènames, en bref, toute substance qui contient un noyau β-lactame
dans sa structure moléculaire. Les céphalosporines constituent les molécules d’intérêt de notre
étude. Leur structure chimique de base est constituée d'un noyau β-lactame accolé à un noyau
dihydrothiazine, l’ensemble formant un noyau unique appelé l'acide 7-
aminocéphalosporanique (figure2).

Synthèse bibliographique
20
Figure 2: Acide 7-aminocéphalosporanique (Glupczynski et al., 2002).
On distingue à l'heure actuelle 4 générations de céphalosporines. Les céphalosporines de 1ère
génération ont un spectre essentiellement dirigé vers les bactéries à Gram (+), et ce spectre
s'élargi vers lesbactéries à Gram (-) dans les générations suivantes. Les céphalosporines de
1ère génération constituent un premier choix dans la prophylaxie chirurgicale. Les
céphalosporines de 2è génération sont utilisées dans un grand nombre d'infections, notamment
respiratoires, urinaires, ostéoarticulaires. Les céphalosporines de troisième et de quatrième
génération sont utilisées dans le traitement d'infections sévères à bactéries à Gram négatif.
L'émergence des résistances aux céphalosporines de troisième génération pose actuellement
un problème en milieu hospitalier (Glupczynski et al., 2002). Comparé à l'acide 6-
aminopénicillanique, l'acide 7-aminocéphalosporanique possède un carbone supplémentaire
(figure 3). Un élément important est la possibilité de substitution en C3 par des groupements
électrocapteurs. Ceux-ci permettent une meilleure délocalisation des électrons au niveau du
cycle β-lactame, rendant en principe les céphalosporines plus actives vis-à-vis des
transpeptidases en comparaison des pénicillines (Glupczynski et al., 2002).
Synthèse bibliographique
21
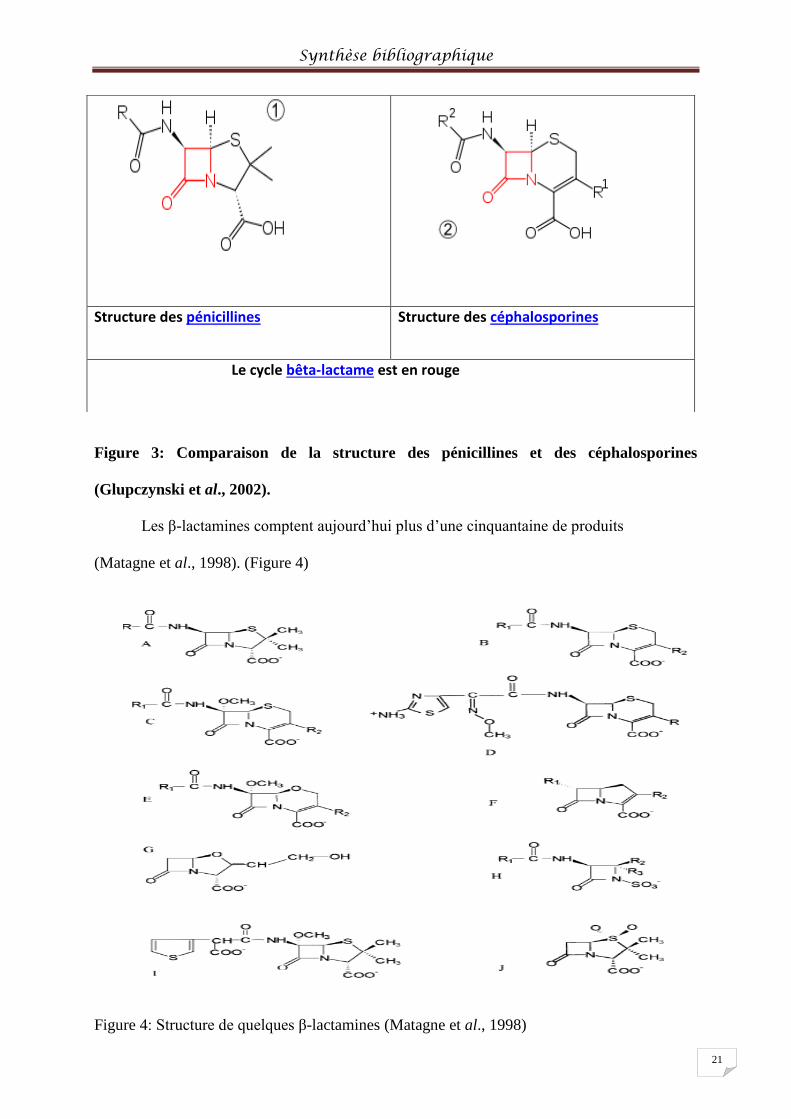
Figure 3: Comparaison de la structure des pénicillines et des céphalosporines
(Glupczynski et al., 2002).
Les β-lactamines comptent aujourd’hui plus d’une cinquantaine de produits
(Matagne et al., 1998). (Figure 4)
Figure 4: Structure de quelques β-lactamines (Matagne et al., 1998)
Structure des pénicillines Structure des céphalosporines
Le cycle bêta-lactame est en rouge

Synthèse bibliographique
22
(A) Penams (e.g. benzylpenicillin, ampicillin, amoxicillin), (B) cephems (cephalosporins), (C)
cephamycins (e.g. cefoxitin), (D) cefotaxime (oxyimino cephalosporin; R= CH2-O-CO-CH3)
(E) oxacephamycines (e.g. moxalactam), (F) carbapenems (e.g. imipenem), (G) clavulanate
(oxapenam), (H) monobactams (e.g. aztreonam), (I) temocillin (6-a-methoxy penam) et (J)
sulbactam (penam sulphone).
II-5-2- Mode d’action sur les bactéries
La diversité de choix des β-lactamines telles que les pénicillines de type amino-, carboxy-,
acyluréido- et amidino-, céphalosporines (1re
, 2e et 3
e génération, voire de 4
e génération),
céphamycines (céfoxitine, céfotétan) et enfin carbapénèmes, illustre la complexité de leur
mode d'action avec au moins sept cibles dénommées protéines liant la pénicilline (PLP).
(Fontana et al.,1983; Al-Obeid et al.,1990). L'action antibactérienne des β-lactamines
provient de leur liaison aux PLP et surtout, de l'inhibition de l’activité des trans-peptidases
impliquées dans la synthèse de la paroi bactérienne. Cette inhibition est due à la parenté
structurale entre les β-lactamines et le dipeptide D-alanyl-D-alanine (substrat des PLPs),
constitutif de la paroi bactérienne. Ces protéines sont des enzymes qui catalysent les liaisons
entre les chaînes peptidiques dans la paroi ou assurent le remaniement de ces chaînons.
Certaines PLP sont des D, D-peptidases qui appartiennent à la super famille des enzymes à
sérine active reconnaissant les pénicillines (Joris et al., 1988). L’interaction entre ces
protéines et les β-lactamines entraîne la formation d’un acyl-enzyme (E-S*) où l’antibiotique
est covalentiellement lié au résidu de la serine active (Gilliane et al., 1996).
1 2 3*
1
k K k
kE S ES ES E P
2.2
Équation 1: Interaction entre les PLPs et les β-lactamines (Gilliane et al., 1996).

Synthèse bibliographique
23
Toutefois la valeur de k3 est très faible comparée a celle de la réaction 1.1, ce qui entraîne une
accumulation de l’acylenzyme. Le complexe covalent ES* résulte de l’attaque nucléophile du
groupement carbonyl du carbone du noyau β-lactame par le groupement hydroxyle de la
chaîne latérale de la sérine active. Á l’issue d’une attaque nucléophile par une molécule d’eau
ou d’un composé aminé, l’enzyme est régénérée et le produit libéré (Frère et al., 1975 ; Fuad
et al., 1976).
II-6-inhibiteurs des β-lactamases
Afin d’assurer une stabilité vis à vis des bêtalactamases, l’une des acquisitions les plus
spectaculaires au sein de la famille des β-lactamines fut la découverte des inhibiteurs de
bêtalactamases. Doués d’une faible activité intrinsèque, les inhibiteurs de bêtalactamases
doivent être associés à d’autres β-lactamines ayant une bonne activité antibactérienne mais
hydrolysables par les β-lactamases (Gutmann, 1989). Cette nouvelle approche permet de
restituer l’activité de certaines bêtalactamines devenues inefficaces vis-à-vis des bactéries
productrices de bêtalactamases (Cartier et al.,1988). Trois inhibiteurs de bêtalactamases sont
actuellement utilisés en thérapeutiques : l’acide clavulanique, le sulbactam et le tazobactam.
L’acide clavulanique inhibe seulement les pénicillinases alors que le tazobactam et le
sulbactam peuvent inhiber les pénicillinases mais surtout les céphalosporinases (Gutmann,
1989). Ces inhibiteurs de bêtalactamases ont été associés à l’amoxicilline, à la ticarcilline et
aux ureïdo pénicillines. L’association amoxicilline + acide clavulanique commercialisée en
1984 sous le nom d’Augmentin est la mieux connue et la plus utilisée. Son spectre d’activité
associe à celui de l’amoxicilline, les bactéries productrices des pénicillinases soit d’origine
chromosomique (Klebsiella, Branhamella catarrhalis, Bactéroïdes fragilis) soit d’origine
plasmidique (Staphylococcus aureus, Haemophilus influenzae, Neisseria gonorrhoeae,
certaines entérobactéries : Escherichia coli, Proteus, Salmonella…) (Gutmann, 1989).

Synthèse bibliographique
24
III-Résistances bactériennes aux β-lactamines
Les bactéries ont développé différents mécanismes pour contrecarrer l'action des β-
lactamines (Galleni et al., 1995 ; Nikaido, 1998 ; Lakaye et al., 1999 ; Walsh, 2003).
Parmi les armes de la chimiothérapie anti-infectieuses, les antibiotiques comportant le noyau
-lactame ou -lactamines connaissent un usage massif à travers le monde et plus
particulièrement dans les pays en développement en raison de leur activité élevée, de leur
absence presque totale d’effets collatéraux et de leur accessibilité. Les bactéries ont développé
toutefois des mécanismes de résistance efficaces reposant sur plusieurs facteurs
génétiquement indépendants.
La création d’une barrière d’imperméabilité (absence ou réduction du nombre de porines) qui
s’oppose à la diffusion de l’antibiotique jusqu’aux cibles physiologiques qui sont les PBP
ancrées sur la surface externe de la membrane cytoplasmique, la modification de la structure
même des cibles conduisant à une baisse de leur affinité pour les antibiotiques, l’acquisition
ou surproduction des pompes efflux qui peuvent expulser l’antibiotique hors de la cellule
même contre le gradient de concentration et enfin la production d’enzymes qui inactive
l’antibiotique. Pour être efficace, un antibiotique doit parvenir au contact de la bactérie,
ensuite y pénétrer, sans être détruit ni modifié, et se fixé à une cible (PBP) pour perturber
ainsi la physiologie bactérienne. Si l’une de ces conditions n’est pas remplie, l’antibiotique,
même correctement administré, se révèle inefficace. Ce phénomène appelé résistance est
lourd de conséquences et doit être, si possible, dépisté au laboratoire. D’un point de vu
bactériologique, on dit qu’une souche est résistante lorsqu’elle peut croitre en présence d’une
concentration d’antibiotique plus élevée que la concentration qui inhibe la majorité des
souches de la même espèce. Il faut donc tenir compte d’un effet dose. On parle de bas niveau
de résistance si la croissance est stoppée par de faible concentration d’antibiotiques et de haut
niveau de résistance si de fortes concentrations sont nécessaires.

Synthèse bibliographique
25
Il existe des résistances naturelles, programmées sur le génome bactérien, donc fixes et
croissantes à l’intérieur du taxon. A ce titre, elles constituent un critère d’identification.
Il existe également des résistances acquises, consécutives à des modifications de l’équipement
génétique chromosomique ou plasmidien. Elles ne concernent que quelques souches d’une
même espèce mais peuvent s’étendre : leur fréquence varie dans le temps mais aussi dans
l’espace – région, ville, hôpital ou même service. Elles constituent un marqueur
épidémiologique. Les modifications génétiques responsables de résistance acquise sont
chromosomiques, secondaires à une mutation portant sur le chromosome ou extra-
chromosomique par acquisition de gènes. Les résistances mutationnelles ou chromosomiques
sont spontanées et stables. Elles préexistent à l’usage de l’antibiotique, et se transmettent
verticalement dans le clone bactérien. Il semble que la résistance par mutation soit peu
rependue en clinique (moins de 20% des résistances acquises). L’usage de l’antibiotique
sélectionne des souches résistantes et la parade consiste donc à associer les antibiotiques. Ce
type de résistance est observé, entre autres, chez les mycobactéries. Les résistances extra-
chromosomiques sont celles dont le support génétique est un plasmide ou un transposon
acquis par conjugaison ou plus rarement par transduction. Elles sont fréquentes (plus de 80%
des résistances acquises), elles sont contagieuses et se transmettent horizontalement entre
bactéries cohabitant, même d’espèces différentes. Elles peuvent concerner plusieurs
antibiotiques, voire plusieurs familles d’antibiotiques, conduisant à une multi-résistance.
Toutes les espèces bactériennes sont ou peuvent être de ce type de résistance. Au fil des
années la pénicilline et ses dérivées se sont avérées utiles de divers façon, surtout en médecine
humaine et vétérinaire, pour soigner les maladies infectieuses, de même qu’en zootechnie
pour favoriser la croissance et prévenir les maladies chez les animaux destinés à
l’alimentation. Grâce aux médicaments basés sur le noyau -lactame, la médecine humaine a
pu faire des percées remarquables aux doubles plan de la santé humaines et de l’espérance de
vie. Mais il faut se garder d’ignorer ou de sous-estimer la combativité et l’ingéniosité des

Synthèse bibliographique
26
bactéries. La production d’enzyme hydrolytiques du noyau -lactame a été, et reste une arme
particulièrement meurtrière pour les hommes et les animaux d’autant plus que son spectre
d’action est constamment révisé et donc capable de s’adapter aux arsenaux chimiques mis au
point par l’homme.
III-1-Résistance par production de β-lactamases
III-1-1-définition des β-lactamases
Les β-lactamases sont des enzymes produites par certaines bactéries et sont
responsables de leurs résistances aux β-lactamines comme les pénicillines, les
céphalosporines , les céphamycines, et les carbapénèmes. Ce sont des enzymes dont l’action
hydrolytique sur les β-lactamines conduit à leur inactivation. Le mécanisme le plus répandu
de la résistance bactérienne aux antibiotiques à noyau β-lactame est donc la production des β-
lactamases (Livermore, 1995).
La première mise en évidence des β-lactamases remonte au début des années 40
(Abraham et Chain, 1940). Elles ont été décrites pour la première fois sur des souches de
E.coli et de Klebsiella pneumoniae (Datta et Kontomichalou, 1965 ; Medeiros, 1984 ;
Bradford, 2001). Les gènes qui codent pour ces enzymes sont d'origine chromosomique ou
plasmidique. Ces gènes ont aussi été détectés sur des transposons et des intégrons facilitant
ainsi le transfert horizontal de ces gènes entre espèces phylogénétiquement éloignées. Ces
enzymes sont exportées dans le milieu extracellulaire (bactéries à Gram positif) ou
périplasmique (bactéries à Gram négatif). L’expression de ces protéines enzymatiques est
constitutive chez certaines espèces bactériennes et inductible chez d’autres. La dénomination
des enzymes se rapporte entre autres, soit au nom du patient: TEM pour Temoneira (Bradford,
2001), soit au pays d’origine ou encore au gène codant: SHV pour sulfhydryl-variable. A la
suite des types TEM et SHV les phénomènes de résistances bactériennes ont été renforcés par
la naissance d’autres types d’enzymes dont les types CTX-M, OXA et bien d’autres. Plus de

Synthèse bibliographique
27
290 types de β-lactamases sont décrits (www.lahey.org/studies/inc_webt.html). Les β-
lactamases catalysent de manière efficace et irréversible l’hydrolyse de la liaison amide du
cycle β-lactame donnant un produit biologiquement inactif qui perd totalement son activité
antimicrobienne (Figure 5)
(Matagne et al., 1998). La dissémination des β-lactamases communes et l'apparition de
nouvelles enzymes ou d'enzymes mutantes sont reliées directement à l'utilisation abusive des
β-lactamines dans le domaine de la santé humaine et animale et en agriculture (Kunin et al,
1990 ; Witte, 1997 ; Normark and Normark, 2002).
Figure 5: Voie catalytique générale des β-lactamases à sérine active (Matagne et al.,
1998)
En général, l’acylation (k+2) et l’hydrolyse de l’acylenzyme (k+3) sont rapides.

Synthèse bibliographique
28
D’une autre façon catalytique générale des β-lactamases, on a le schéma réactionnel proposé
par Bush en 1988. (Figure 6)
N
S
O
NR
O
COOH
H
HN
S
OH
NH
R
COOH
O
O
HN
S
NH
R
COOH
O
b-lactamase decarboxylationspontanee
Figure 6: Mécanisme d'hydrolyse de la pénicilline par la β-lactamase (Bush, 1988)
III-1-2-classification des β-lactamases
Généralement, Les β-lactamases sont classées suivant deux schémas : La classification
moléculaire de Ambler (Ambler et al. 1980) et la classification fonctionnelle de Bush-Jacoby-
Medeiros (Bush et al., 1995). La classification moléculaire tient compte de la structure
primaire des différentes β-lactamases et les divisent en quatre classes (A à D). Pour la
classification fonctionnelle, les auteurs tiennent compte de la fonctionnalité des β-lactamases
(substrat, profil d’inhibition) et divisent aussi ces enzymes en quatre groupes (1 à 4) avec
plusieurs sous-groupes. Quoique différents de par le principe, il existe une correspondance
entre les deux schémas (Bush et al., 1995). Les β-lactamases des classes A, C et D font partie
des enzymes à sérine active, c'est-àdire qui possèdent dans leur site actif une sérine qui
intervient dans le mécanisme d’acylation au cours de l’hydrolyse des β-lactamines. Par contre
la classe B inclut les métallo- β-lactamases dont l'activité nécessite la présence d'ions
métalliques.
La classe A est la plus diversifiée, on y retrouve les pénicillinases des bactéries à Gram
positif, les β-lactamases plasmidiques à large spectre qui hydrolysent les céphalosporines avec
autant d'efficacité que les pénicillines, les β-lactamases à spectre élargi qui hydrolysent les

Synthèse bibliographique
29
céphalosporines de 3éme génération et les monobactames. La majorité de ces enzymes est
sensible aux inhibiteurs suicides (acide clavulanique, sulbactame et tazobactame) utilisés en
médecine. Les principaux représentants de ce groupe sont les β-lactamases du type TEM,
SHV et très récemment le type CTX-M. Dans la classe C, on retrouve les céphalosporinases
qui sont des enzymes résistantes à l’action de l’acide clavulanique et le sulbactame ; toutefois
certaines sont faiblement inhibées par le tazobactame (Doi et al., 2004). Leur hyperproduction
est associée au phénotype de multirésistance observé chez certains bacilles à Gram négatif.
Au départ à médiation chromosomique, les β-lactamases de la classe C sont aussi aujourd’hui
à médiation plasmidique (Philippon et al., 2002). Les représentants de ce groupe sont les
enzymes du type, AmpC, FOX, ACT, CMY.
La classe D regroupe les β-lactamases qui hydrolysent les isoxazolylpénicillines comme la
cloxacilline et l'oxacilline. Ces dernières sont réfractaires à l'hydrolyse par les autres classes
de β-lactamases et possèdent une certaine activité inhibitrice. On les appelle des oxacillinases
et elles sont représentées par les β-lactamases du type OXA. Ces enzymes sont plus ou moins
résistantes à l’action de l’acide clavulanique, mais sont bien inhibées par le tazobactame
(Livermore, 1995). Contrairement aux sérine-β-lactamases, les β-lactamases de la classe B
nécessitent la présence d'un ou deux ions zinc pour être actives. Leur importance clinique est
liée au fait qu’elles hydrolysent les carbapenèmes, composés qui échappent à l’activité des β-
lactamases à sérine active. La plupart des métallo-β-lactamases hydrolysent une variété de
pénicillines et de céphalosporines, et sont insensibles aux inhibiteurs classiques (acide
clavulanique, sulbactame, tazobactame). Elles sont inhibées par les agents chelatants comme
l’éthylènediaminetétracétique (EDTA) et l’acide dipicolinique. A partir de la séquence des
enzymes, cette classe est subdivisée en trois sous classes, B1, B2, et B3 (Galleni et al, 2001).
La production de ces enzymes par les bactéries pathogènes telles Klebsiella pneumoniae,
Escherichia coli, Serratia marcescens, Pseudomonas aeruginosa, Bacteroides fragilis,

Synthèse bibliographique
30
Stenotrophomonas maltophilia, Chrysobacterium meningosepticum, Acinetobacter
baumannii, rend difficile le choix d'une antibiothérapie efficace.
III-2- β-lactamases à spectre élargi
III-2-1-généralités
Les β-lactamases à spectre élargi (BLSE) constituent un groupe d'enzymes qui ont la
propriété commune de conférer la résistance aux pénicillines, à la 1ère, 2nde et 3ème
génération de céphalosporines, à l’aztréonam (mais non aux céphamycines et carbapenèmes)
par hydrolyse de ces antibiotiques, et qui sont inhibées par les inhibiteurs des β-lactamases tel
l’acide clavulanique (BUSH et al., 1995). La présence des BLSE a été décrite pour la
première fois chez Klebsiella ozaenae, en 1983 en République Fédérale d'Allemagne, puis en
1984 chez K. pneumoniae et E. coli en France et en Tunisie (Knothe et al., 1983; Kliebe et al,
1985). Ces enzymes avaient été désignées céfotaximase et ceftazidimase car elles conféraient
aux bactéries qui les produisaient une résistance préférentielle à la céfotaxime ou à la
ceftazidime. Elles inactivaient aussi d'autres oxyimino-β-lactamines telles que la ceftriaxone
et l’aztréoname. La répartition de ces enzymes est aujourd’hui mondiale (Bradford, 2001 ;
Paterson et Bonomo, 2005). L'utilisation des techniques moléculaires et la détermination des
séquences peptidiques ont permis de démontrer que la plupart des BLSE dérivent des
pénicillinases déjà connues (TEM-1, TEM-2, SHV-1) par substitution d’un ou de plusieurs
acides aminés qui leur permet d’hydrolyser les oxyimino-β-lactamines et les monobactames
(Bush et al., 1995).
Aujourd’hui on assiste à une émergence des BLSE non TEM et non SHV incluant les
types CTX-M, PER, VEB, GES and BES (Bradford, 2001). Les BLSE sont à médiation
plasmidique et appartiennent à la classe moléculaire A de Ambler et au groupe 2be de la
classification de Bush-Jacoby- Medeiros à l’exception des BLSE OXA (Classe D de Ambler
et 2d de Bush-Jacoby-Meideros) (Bush et al., 1995). De par leur propriété d’être inhibées par

Synthèse bibliographique
31
l’acide clavulanique elles se différentient des β-lactamases du groupe C produites par les
bacilles à Gram-négatif tel Enterobacter qui peuvent posséder un spectre élargi par
surproduction mais qui sont résistantes à l’acide clavulanique et sont sensibles aux
céphalosporines de 4ème génération (céfépime, cefpirome) (Cosgrove et al., 2002). Elles
diffèrent aussi des métallo-β-lactamases qui, quoique hydrolysant toutes les β-lactamines
(pénicillines, céphalosporines et carbapenèmes) ne sont inhibées que par l’EDTA (Walsh et
al. 2005).
III-2-2-Repartition mondiale
Depuis leur apparition en Europe en 1983, les BLSE ont aujourd’hui une répartition
mondiale. On assiste à une augmentation de plus en plus élevée de la prévalence et une
émergence grandissante de nouveaux types. Bon nombre de laboratoires de microbiologie
clinique se limitent à utiliser les points critiques des antibiotiques et ne détectent pas les
souches productrices de BLSE ou en sont incapables (Tenover et al., 1999). Ces points
critiques sont variables de pays en pays (Tableau I) et certaines souches productrices de BLSE
apparaissent sensibles aux céphalosporines de 3e génération.
Des travaux sur l’évaluation d’une collection de souches productrices de BLSE en
utilisant les points critiques du « Clinical Laboratory Standards Institute » (ancien « National
Committee for Clinical Laboratory Standards ») ont montré que 13 à 49% des souches testées
sont sensibles au cefotaxime, 36 à 79% au ceftriaxone, 11 à 52% au ceftazidime et 10 à 67% à
l’aztréoname.
Approximativement 40% des souches testées sont sensibles à au moins une oxyimino-β-
lactamine et 20% à toutes les oxyimino-β-lactamines. Cette apparente sensibilité peut
s’expliquer par le fait que les BLSE hydrolysent différemment les céphalosporines et la
perméabilité envers les différentes oxyimino-β-lactamines n’est pas la même (Paterson &
Bonomo, 2005). Il est donc nécessaire d’avoir des méthodes adéquates qui permettent de
détecter les BLSE. Différentes approches ont été développées pour accroître la détection en
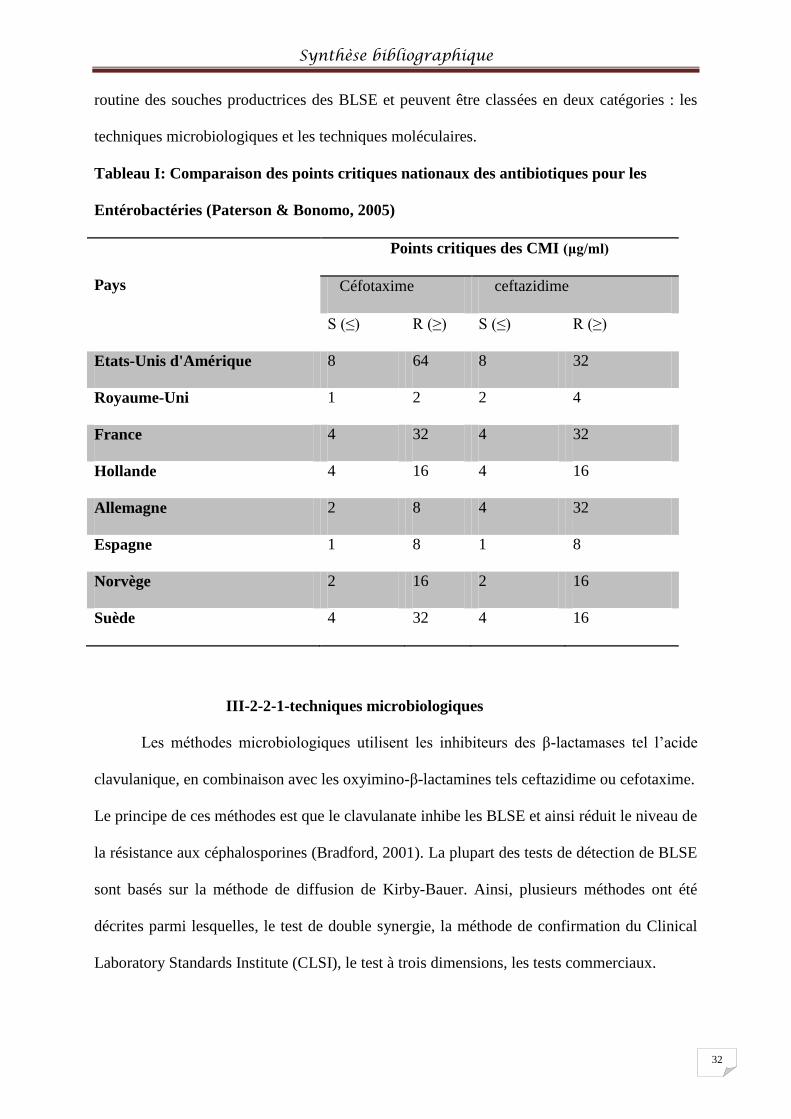
Synthèse bibliographique
32
routine des souches productrices des BLSE et peuvent être classées en deux catégories : les
techniques microbiologiques et les techniques moléculaires.
Tableau I: Comparaison des points critiques nationaux des antibiotiques pour les
Entérobactéries (Paterson & Bonomo, 2005)
Pays
Points critiques des CMI (μg/ml)
Céfotaxime ceftazidime
S (≤) R (≥) S (≤) R (≥)
Etats-Unis d'Amérique 8 64 8 32
Royaume-Uni 1 2 2 4
France 4 32 4 32
Hollande 4 16 4 16
Allemagne 2 8 4 32
Espagne 1 8 1 8
Norvège 2 16 2 16
Suède 4 32 4 16
III-2-2-1-techniques microbiologiques
Les méthodes microbiologiques utilisent les inhibiteurs des β-lactamases tel l’acide
clavulanique, en combinaison avec les oxyimino-β-lactamines tels ceftazidime ou cefotaxime.
Le principe de ces méthodes est que le clavulanate inhibe les BLSE et ainsi réduit le niveau de
la résistance aux céphalosporines (Bradford, 2001). La plupart des tests de détection de BLSE
sont basés sur la méthode de diffusion de Kirby-Bauer. Ainsi, plusieurs méthodes ont été
décrites parmi lesquelles, le test de double synergie, la méthode de confirmation du Clinical
Laboratory Standards Institute (CLSI), le test à trois dimensions, les tests commerciaux.

Synthèse bibliographique
33
Le test de double synergie (Jarlier et al., 1988) entre un disque de céphalosporine de
troisième génération ou une monobactame (aztréonam) et un inhibiteur de β-
lactamase tel l'acide clavulanique est l’un des premiers tests à être décrit. C'est le test
le plus utilisé et le plus recommandé. Ce test possède plusieurs variantes et est basé
sur la mise en évidence d'une restauration de l'activité des céphalosporines de
troisième génération ou de l’aztréonam en présence d'un inhibiteur enzymatique. Le
test de synergie peut être insensible pour plusieurs raisons :
la distance entre les disques ;
l'inaptitude des inhibiteurs à inhiber toutes les β-lactamases
(céphamycinases qui ne sont pas inhibées par exemple) ;
l'inaptitude du test à détecter la présence des BLSE (montrer l'image de
synergie « tire-bouchon » chez les souches productrices des
céphalosporinases chromosomiques).
Malgré ces défaillances, c'est une méthode assez utilisée en routine car elle ne nécessite pas
beaucoup de matériel et est de réalisation facile.
L’interprétation de la faible diminution de la sensibilité des souches vis-à-vis des
céphalosporines de troisième génération et de l’aztréonam n'est pas une méthode assez
fiable car la diminution de la sensibilité peut impliquer un mécanisme de résistance
autre que la production des BLSE (Thomson & Sanders, 1992).
Le test à trois dimensions (Thomson et Sanders, 1992) permet non seulement d’avoir
les informations sur le profil des β-lactamases, mais aussi sur la sensibilité de la
souche vis-à-vis des antibiotiques utilisés. Cette méthode a une sensibilité élevée, mais
n'est pas de réalisation facile.
L'utilisation du disque de cefpodoxine (30 μg) est une méthode qui permet de
détecter les BLSE chez E. coli et K. pneumoniae. Ce disque n'est pas beaucoup utilisé
par les laboratoires de microbiologie clinique (Thomson, 1995) ;

Synthèse bibliographique
34
L'utilisation des disques de ceftazidime de 5 μg est proposée par Jacoby & Han
(1996) pour la détection des BLSE chez E. coli et K. pneumoniae. Les BLSE sont
suspectées lorsque le diamètre d'inhibition est inférieur à 18 mm. Ces auteurs
proposent d’ajouter 20 μg de sulbactame sur un disque d’oxyimino-céphalosporine.
L’augmentation du diamètre d’inhibition de 5 mm comparée au disque sans inhibiteur
est signe de la production d’une BLSE ;
Le CLSI propose l’utilisation de la méthode des disques pour rechercher la production de la
BLSE par Klebsiella, Escherichia coli et Proteus mirabilis. L’obtention d’un diamètre
d’inhibition spécifique à un des oxyimino-β-lactamines ou à l’aztréonam [aztréonam (<28
mm), cefotaxime (< 26 mm), ceftriaxone (<26 mm), ceftazidime (<23 mm), cefpodoxine
(<18 mm)] permet de suspecter la production de BLSE. Toutefois la sensibilité de détection
est grande si on teste plus d’une de ces cinq β-lactamines. Dès lors on fait appel à un test de
confirmation pour confirmer le diagnostic (NCCLS, 2005). Cet institut propose de réaliser les
tests de dilution en mettant dans le milieu de culture 1μg/ml d’une des cinq β-lactamines à
spectre élargi. Les souches présentant des CMI ≥ 2μg/ml seront suspectées comme
productrices de BLSE. Les tests de confirmation consistent en ce qui concerne la méthode des
disques à tester le disque de cefotaxime (30μg) ou de ceftazidime (30μg) seul et contenant
10μg d’acide clavulanique. L’augmentation du diamètre d’inhibition d’au moins 5 mm du
disque contenant l’inhibiteur comparé à celui du disque sans inhibiteur confirme la production
de BLSE par la souche. Pour les méthodes en dilution, le cefotaxime (0,25 -64 μg/ml) ou la
ceftazidime (0,25-128 μg/ml) sera testé en présence de l’acide clavulanique (4μg/ml). La
diminution de la concentration minimale inhibitrice (CMI) d’au moins trois fois de la série de
double dilution en présence de l’acide clavulanique est indicatrice de la production de BLSE
par la souche qui peut être considérée comme résistante à toutes les oxyimino-β-lactamines et
l’aztréoname (NCCLS, 2005).

Synthèse bibliographique
35
Plusieurs industries ont développé des tests de détection de BLSE qui peuvent être utilisés en
même temps que les tests de détection des CMI dans les laboratoires de microbiologie
clinique et sont commercialisables.
Le test de confirmation par les disques utilisant les disques BBL Sensi-Disk (BD
Biosciences, Sparks, Md.). Ce test utilise la méthode de diffusion avec les disques de
cefotaxime et de ceftazidime seuls et contenant de l’acide clavulanique. La BLSE est
détectée lorsque le diamètre d’inhibition de la combinaison céphalosporine/acide
clavulanique est supérieur d’au moins 5 mm à celui de la céphalosporine seule.
Les bandelettes E-test pour la détection des BLSE (AB Biodisk, Solna, Sweden). Ces
bandelettes renferment sur un bout un gradient de ceftazidime ou de cefotaxime et sur
l’autre bout la ceftazidime ou cefotaxime avec l’acide clavulanique. Le test est positif
lorsqu’on observe une réduction d’au moins 3 fois de CMI de la ceftazidime en
présence de l’acide clavulanique. Ce test est assez sensible (87-100%) et spécifique
(95-100%) (Cormican et al.1996 ; Brown et al. 2000). Ce test est de réalisation facile
mais il arrive que l’acide clavulanique diffuse du coté où la ceftazidime est seule,
rendant difficile la lecture (Vercauteren et al. 1997). Ce test coûte cher et n’est pas à la
portée des laboratoires des pays en développement.
Le test Vitek pour BLSE (bioMerieux Vitek, Hazelton, Missouri). La carte Vitek pour
BLSE utilise la ceftazidime ou le cefotaxime seul (0,5μg/ml) et en combinaison avec
l’acide clavulanique (4μg/ml). La réduction de la croissance dans les puits contenant
l’acide clavulanique comparée à ceux contenant les céphalosporines seules indique la
présence de BLSE. La sensibilité et la spécificité de ce test sont supérieures à 90%
(Sanders et al., 1996).
L’avantage certain de ce test est qu’il peut être facilement intégré dans les différents
processus des laboratoires qui utilisent déjà le système Vitek. Le coût de l’équipement pour la
réalisation de ce test dans les laboratoires des pays en développement est très élevé.

Synthèse bibliographique
36
D’autres systèmes automatisés tels Dade Behring MicroScan (Sacramento, Calif.) et le
système microbiologique automatisé BD Phoenix de Becton Dickinson Biosciences
(Sparks, Md) sont aussi capables de détecter les BLSE. Sur ces tests commerciaux,
Linscott & Brown (2005) ont montré que leur sensibilité variait de 96% (BBL sensi-
Disk) à 100% (MicroScan) et leur spécificité de 94% (E-test) à 100% (BBL sensi-
Disk). La plupart de ces tests visent à détecter la production des BLSE par E. coli,
Proteus mirabilis et Klebsiella.
La détection de ces enzymes chez les souches productrices des céphalosporinases reste un
véritable problème. Toutefois, Pitout et al. (2003) proposent une modification du test de
double synergie pour détecter la production des BLSE par ces bactéries. Ils remplacent le
disque de ceftriaxone par celui de céfépime (30μg) et placent le disque de
pipéracilline/tazobactame (100/10μg) à 25 mm de ce dernier. A ces techniques basées sur
l'antibiogramme, il existe d'autres utilisant les méthodes moléculaires.
III-2-2-2-techniques moléculaires
Les tests phénotypiques permettent de manière présomptive d’identifier la présence de
BLSE. Pour connaître le type de BLSE que produisent les souches cliniques, il faut avoir
recours aux techniques moléculaires qui sont assez complexes et laborieuses. Ces techniques
requièrent des personnes hautement qualifiées et un laboratoire bien équipé. Elles ne sont pas
facilement réalisables en routine surtout dans les pays en développement. Ces techniques ont
le mérite d’être spécifiques et permettent de connaître le type de BLSE produit par les isolats
cliniques. Plusieurs de ces techniques sont utilisées entre autres : l’utilisation des sondes
d’acides désoxyribonucléiques, la « Polymerase Chain Reaction » (PCR) avec plusieurs
variantes, l’oligotypage, la « Ligase Chain Reaction », et le séquençage des nucléotides
(Bradford, 2001). Ces techniques en plus de la détection permettent aussi de caractériser les
BLSE.

Synthèse bibliographique
37
III-2-3-diversité des types de BLSE
On dénombre jusqu’à ce jour environ 200 BLSE appartenant à plusieurs types parmi lesquels
les types TEM et SHV sont prédominants.
TEM
Les BLSE de types TEM dérivent de TEM-1 et TEM-2 par substitution d’un ou de plusieurs
acides aminés. Plus de cent types existent et leurs points isoélectriques varient entre 5,4 - 6,5.
(http://www.lahey.org/studies/temtable.htm). Bien que les BLSE de ce type soient le plus
souvent produites par E. coli et K. pneumoniae, ces enzymes ont été aussi mises en évidence
chez les autres espèces d’Entérobactéries (Enterobacter aerogenes, Morganella morganii,
Proteus mirabilis, Proteus rettgeri et Salmonella spp.), chez Pseudomonas aeruginosa (TEM-
42) et Capnocytophaga ochraceae (TEM-17) (Bradford, 2001).
SHV
La β-lactamase SHV-1 qui est l’enzyme à partir de laquelle les BLSE de type SHV dérivent
par substitution d’un ou de plusieurs acides aminés est très fréquente chez K.pneumoniae.
Cette enzyme hydrolyse les pénicillines, les céphalosporines mais pas les oxyimino-
céphalosporines et l’aztréoname. La transformation du phénotype non-BLSE en phénotype
BLSE est pratiquement due toujours au remplacement de la glycine par la sérine en position
238. La mutation en position 240 qui remplace le glutamate en lysine ne fait qu’augmenter
l’activité de l’enzyme. La présence de la séquence d’insertion IS26 sur le gène SHV
faciliterait l’acquisition du phénotype BLSE (Hammond et al., 2005). Environ 70 BLSE du
type SHV ont été décrites et leurs points isoélectriques varient de 7,0 à 8,2
(http://www.lahey.org/studies/webt.html#SHV). La majorité de ces enzymes ont été décrites
chez les souches de K. pneumoniae. Toutefois, ces enzymes ont été trouvées chez Citrobacter
freundii, C. diversus, E. coli, Enterobacter cloacae et P. aeruginosa (Gangoué Piéboji, 2000;
Bradford, 2001 ; Gangoué-Piéboji et al., 2005b).
CTX-M

Synthèse bibliographique
38
Les BLSE du type CTX-M font partis des BLSE non-TEM et SHV et possèdent moins de
40% d’homologie avec les BLSE TEM et SHV. Elles sont plasmidiques et confèrent une forte
résistance au céfotaxime et au ceftriaxone et ont juste un effet marginal sur la CMI de la
ceftazidime (Tzouvelekis et al. 2000). Raportées pour la première fois au milieu des années
1980, elles montrent un niveau de dissémination dans les bactéries et partout dans le monde
qui a augmenté dramatiquement depuis 1995. Les enzymes de la famille CTX-M
comprennent aujourd’hui une cinquantaine d’enzymes isolées à partir de nombreuses espèces
d’Entérobactéries et qui, sur la base de leur séquences protéiques sont divisées en cinq
groupes, CTX-M-1, CTX-M-2, CTX-M-8, CTX-M-9 et CTX-M-25) (Bonnet, 2004,
http://www.lahey.org/studies/other.html#table1). Des travaux ont montré que les gènes
plasmidiques des CTX-M ont des progéniteurs (parents) des enzymes naturelles
chromosomiques de classe A de Kluyvera ascorbata (CTX-M-2), Kluyvera georgiana (CTX-
M-8), Kluyvera cryocresens (CTX-M-1) (Bonnet, 2004).
OXA
Les BLSE OXA diffèrent des types TEM et SHV par le fait qu’elles appartiennent à la classe
D et au groupe 2d (Bush et al. 1995). Contrairement à ce qui est observé dans les BLSE de la
classe A qui sont produites le plus souvent par les Entérobactéries, les BLSE OXA sont en
grande majorité détectée chez P. aeruginosa. Plus d’une dizaine de ces enzymes ont été mises
en évidence et la majorité provient de OXA-10 par mutation d’un ou de plusieurs acides
aminés. A la différence des autres types, les BLSE OXA renferment plus d’une vingtaine de
représentant non-BLSE (Bradford, 2001 ; http://www.lahey.org/studies/other.html#table1).
Les autres types de BLSE
En plus des BLSE connues, de nouvelles familles classées parmi les BLSE non-TEM et non-
SHV sont apparues. Ces BLSE appartiennent à la classe A de Ambler et ont été isolées non
seulement de P. aeruginosa et Acinetobacter baumannii mais aussi des Entérobactéries. Parmi
ces nouvelles familles, le type GES (Guiana Extended Spectrum beta-lactamase) est le plus

Synthèse bibliographique
39
développé avec environ 9 représentants. Les autres types, PER (P. Normann, E., Ronco, R.
Labia beta-lactamase) et VEB (Vietnamasse Extended Spectrum beta-lactamase) n’ont que
trois représentants pour chacun (Poirel & Nordmann, 2005 ;
http://www.lahey.org/studies/other.html#table1).
III-2-4-Epidémiologie des BLSE
III-2-4-1-Apparition et propagation des BLSE en Europe
La première β-lactamase capable d’hydrolyser les céphalosporines à spectre élargi a
été décrite en 1983 en Allemagne dans les services de réanimation. Cette enzyme a été isolée
d’une souche de klebsiella pneumoniae qui hébergeait le gène SHV-2 mutant ponctuel de
SHV-1 (Knothe et al., 1983). En outre une BLSE de type CTX-M-1 a été décrite pour la
première fois en 1989 produite par E.coli. CTX-M-23 provenant d’une souche de la même
espèce a également été détectée (Bonnet, 2004). En France, des BLSE de type VEB-1/3 ont
été décrites sur E.coli, K. Pneumoniae, Proteus mirabilis (Alert, 2003). Les BLSE de type
GES sont de plus en plus rapportées chez les bactéries à gram négatif (BGN), notamment P.
aeruginosa, E. coli et K. pneumoniae. L’enzyme GES-1 a été décrite chez une souche de K.
pneumoniae en 1998 (Naas et al, 2008). En 2003, des BLSE dont le type est non précisé ont
été identifiées sur des souches de Salmonella enterica Newport (Weill et al., 2004). En
Turquie, l’enzyme PER-1, initialement découverte en 1993 chez P. aeruginosa est fréquente
chez P. aeruginosa et Acinetobacter spp. Mais a aussi été détectée chez S. enterica sérovar
Typhimurium, Providencia spp Proteus mirabilis et Alcaligenes faecalis (Waldhagen et al,
2003). La découverte de BLSE de type OXA a été rapportée pour la première fois en Turquie
(Nass et al., 1999). En outre une BLSE de type CTX-M-3 a été décrite en 2000 chez
Salmonella sonnei (Bonnet, 2004). CTX-M-9 a été détecté sur P.mirabilis et K. pneumoniae
en 2003 (Bonnet, 2004). En outre une BLSE de type CTX-M-8 a été décrite chez Salmonella
enterica en 2005 (Rossolini, 2008). En Grèce les BLSE de type CTX-M-6 et CTX-M-7

Synthèse bibliographique
40
produites par Salmonella. typhimirium ont été respectivement identifiées en 1996 et en 1997
(Bonnet, 2004). Il faut signaler qu’en Europe, un réseau s’est constitué pour harmoniser et
renforcer la surveillance des résistances bactériennes (Réseau Européen de Surveillance de la
Résistance Bactérienne aux Antibiotiques: EARSS). Récemment, un dérivé de GES-1, GES-2
identifié à partir de souches sud-africaines a été également retrouvé en Grèce. Le potentiel
hydrolytique de cette enzyme s’étend à toutes les ß-lactamines. (Poirel, 2007). CTX-M-3 a
été identifié chez E.coli et C. freundii en 1996 et chez S. enterica en 1999 (Bonnet, 2004). En
Italie, l’enzyme PER-1 a été détectée en 1997 sur P. mirabilis (Waldhagen, 2003). En
Europe, la prévalence des BLSE chez les entérobactéries varie d’un pays à un autre. Une
étude portant sur E. coli, K. pneumoniae et Enterobacter spp. a montré que la prévalence de
BLSE est de 4,7% en Europe du Nord contre 13,5% au Sud de l’Europe (Bouchillon et al.,
2004).
III-2-4-2-Apparition et propagation des BLSE en Asie
Au Japon, GES-2, variant de GES-1 identifié à partir de souches sud-africaines a été
également décrit (Poirel, 2007). Toujours au Japon, les dérivés CTX-M-2 et CTX-M-11 ont
été détectés en 2000 respectivement sur E.coli et K. pneumoniae (Philippon, 2006). En Corée
CTX-M-3 et CTX-M-9 ont été décrites en 2003 chez E. coli (Philippon, 2006). La BLSE du
type PER-1 identifié à partir d’un isolat prélevé sur un porteur turc en 1993 à Paris est
actuellement très répandue en Asie (Corée du Sud notamment) (Naas et al, 2OO8). Au
Vietnam l’enzyme VEB-1 (38 % d’identité avec PER-1) a été retrouvée en 1996 dans une
souche de E. coli puis chez P. aeruginosa (Bonnet, 2004). Les BLSE de type CTX-M-17,
CTX-M-18 et CTX-M-19 ont été détectées respectivement en 1996, 1997 et 1999 sur des
souches de E.coli et K.pneumonaie (Philippon, 2006). En Inde CTX-M-15 a été mise en
évidence sur des souches de E. coli et Enterobacter cloaceae (Bonnet, 2004). Au Japon, le
pourcentage de résistance aux β-lactamines par production de BLSE chez E. coli et K.
pneumoniae est assez faible. Les travaux de Yagi et al. (2000) ont montré que ce pourcentage

Synthèse bibliographique
41
est <1% pour E. coli et 0, 3% pour K. pneumoniae. Dans d’autres régions de l’Asie, la
proportion de E. coli et K. pneumoniae productrices de BLSE varie de 4,8% en Corée (Pai et
al., 1999) à 8,5% au Taiwan (Yan et al., 2000) et supérieur à 12% à Hong Kong (Ho et al,
2000).
III-2-4-3-Apparition et propagation des BLSE en Amérique
Les BLSE de type GES initialement décrites chez une souche de K. pneumoniae
isolée en 1998 en France ont été aussi décrite en Argentine, au Brésil (Naas et al., 2008). Au
Mexique l’enzyme TLA-1 (Tlahuicas - tribu indienne) hébergée par une souche de E. coli a
été rapportée en 1993. Depuis, plusieurs cas de bactériémies et d’infections urinaires
nosocomiales dues à K. pneumoniae produisant à la fois SHV-5 et TLA-1 ont également été
rapportés au Mexique (Naas et al., 2008). A noter que depuis lors, TLA-1 n’a été identifié
qu’au Mexique (Naas et al., 2008) . L’enzyme PER-2 apparentée à la BLSE du type PER-1
identifiée à partir d’un isolat prélevé sur un patient turc en 1993 à Paris a été décrite
également en Amérique du Sud. (Naas et al, 2008). Au Canada, une étude entre 2000 et
2002 menée dans les hôpitaux des régions de Durham, York, Toronto et Ontario a rapporté
des souches de E.coli et K.pneumoniae sécrétrices de BLSE (Muller et al., 2002) sans en
préciser les types. Aux Etats Unis, la prévalence des Entérobactéries productrices des BLSEs
varie de 0 à 25% avec une moyenne nationale de 3% (Centers for Disease Control and
Prevention, National Nosocomial Infectious surveillance,
(http://www.cdc.gov/ncidod/hip/SURVEILL/NNIS.HTM). Au Canada, la prévalence de
production des BLSE est faible et est de l’ordre de 0,25% pour E. coli et 0,8% pour Klebsiella
sp. (Mulvey et al., 2004).
III-2-4-4-Apparition et propagation des BLSE en Afrique

Synthèse bibliographique
42
Les BLSE ont diffusé à travers l’Afrique et ont fait l’objet de plusieurs travaux. Ainsi
concernant la partie Nord de l’Afrique, d’après Bauernfeind et al. en 1992, des BLSE auraient
été rapportées en Tunisie en 1988 et en Algérie en 1990. En outre des BLSE de types SHV-
11, CTX-M-15, CTX-M-27, CTX-M-28 et TEM-164 ont été isolées et décrites en Tunisie
(LARABI et al., 2003 ; ACHOUR et al., 2008; ACHOUR et al.,2009 ; Réjiba et al., 2011). Il
a été également rapporté des BLSE de type CTX-M en Algérie (Touati et al., 2006 ; Messai et
al., 2008) tandis qu’au Maroc ce sont des BLSE de types SHV-2, SHV-5, SHV-12, TEM-21,
TEM-27et CTX-M2 qui ont été mises en évidence (Mhand et al., 1999 ; ‘Mahmal et al.,
2004). En Égypte la première description d’une enzyme CTX-M a été faite à partir d’un isolat
d’E. coli (Wiegand et Al-Agamy, 2003).
Pour les pays de l’Afrique de l’Est, l’existence des BLSE a été rapportée en Éthiopie
et au Kenya sans précision (Seid et Asrat, 2005). Concernant la partie centrale de l’Afrique, il
a été rapporté au Cameroun des BLSE de types SHV-12, CTX-M-1, CTX-M-15, et OXA-30
(Gangoue-Pieboji et al., 2005a, 2005b; Gangoue-Pieboji, 2007; Gangoue-Pieboji et al.,
2009). De même en République Centrafricaine, la prévalence de BLSE chez les
Entérobactéries est de 4 % (Frank et al., 2006).
Pour la partie Sud africaine, l’existence des BLSE a été rapportée en Afrique du Sud et
ce sont les BLSE de types GES-2 qui ont été décrites (Derby et Berens, 1997; Wadula et al.,
2006; Naas et al., 2008). Il a été également fait cas des BLSE en Tanzanie et ce sont des
BLSE de types CTX-M-15 et SHV-12 qui ont été mises en évidence (Blomberg et al., 2005).
A Madagascar des BLSE de types SHV-2 et CTX-M-15 ont été principalement
rencontrées (Vedy et al., 2009).
Pour la région Ouest Africaine, l’existence des BLSE a été rapportée en Côte D’Ivoire
sans précision (N’guessan et al., 2007). Tandis qu’au Bénin, des BLSE de types SHV et TEM
ont été décrites (Ahoyo et al., 2007). Au Sénégal ce sont des BLSE de type CTX-M-15 qui
ont été rapportées (Ruppe et al., 2009). Au Mali, il a été retrouvé des BLSE de type CTX-M

Synthèse bibliographique
43
produites par les entérobactéries (Duval et al., 2009). Il a été également fait cas des BLSE au
Niger mais sans précision. Ainsi il a été isolé 29 souches productrices de BLSE à partir de
prélèvements d’enfants malades au Niger (Nguyen et al., 2009). Au Burkina Faso, à notre
connaissance aucun type de BLSE n’a été décrit. Néanmoins une étude sur la fréquence des
souches productrices de BLSE impliquées dans les infections urinaires a été faite à
Ouagadougou (Karou et al., 2009).

Matériel et Méthodes
44
CHAPITRE II : MATERIEL ET METHODES
I-Cadre de l’étude
La récolte de nos isolats s’est effectuée au Centre Hospitalier Universitaire
Pédiatrique Charles De Gaulle (CHUP-CDG) de Ouagadougou qui abrite un Laboratoire
d’analyse médicale. Le CHUP-CDG est situé au croisement du boulevard Charles de Gaulles
et du boulevard de la jeunesse (boulevard des Tensoba) au secteur 28 de Ouagadougou. Il
comprend un laboratoire sectionné comme suit : la Bactériologie où s’est déroulée la collecte
de nos isolats, la Parasitologie, l’Hématologie, la Biochimie, l’Immuno-Sérologie et, la
biologie moléculaire
Des études approfondies sur les différents isolats récoltés ont été poursuivies au Laboratoire
d’Enzymologie de la Chimio-Résistance Bactérienne (LECRB) du département de Biochimie-
Microbiologie situé dans l’Unité de Formation et de Recherche en Sciences de la Vie et de la
Terre (UFR-SVT) à l’Université de Ouagadougou.
La recherche des gènes de BLSE par la technique de la PCR classique a été réalisée au Centre
National de Recherche et de Formation sur le Paludisme (CNRFP).
La suite de la recherche des gènes par la PCR et le séquençage des différents produits PCR se
sont réalisés au Laboratoire de Biotechnologie Végétale (LBV) à l’Université libre de
Bruxelles.
II-Type d’étude
Notre étude est une étude prospective analytique réalisée sur diverses espèces
bactériennes productrices de BLSE isolées dans le Centre Hospitalier Universitaire
Pédiatrique Charles De Gaulle (CHUP-CDG) de Ouagadougou.

Matériel et Méthodes
45
III-Population d’étude ou Echantillonage
III-1-Critères d’inclusion
Les échantillons retenus sont ceux en provenance des enfants malades hospitalisés ou non
hospitalisés au CHUP-CDG de Ouagadougou. Quelques critères de qualité étaient de mise. Il
s’agit de la conformité du bulletin d’examen qui doit comporter entre autre l’identité du client,
la nature du prélèvement, le diagnostic ou le bilan clinique, le service de consultation et/ou
d’hospitalisation.
III-2-Critères de non inclusion
Les échantillons recueillis dans du matériel non stérile ou soupçonnés de toute marque de
souillures extérieures sont exclus.
IV-Matériel
IV-1-Les produits chimiques
IV-1-1- les produits chimiques fournis par la maison Sigma
Ces produits sont constitués en grande partie par les β-lactamines : benzylpénicilline,
ampicilline, amoxicilline, carbénicilline, pipéracilline, oxacilline, céfalotine, céfaloridine,
céfalexine, céfuroxine, céfotaxime, ceftazidime, imipénème, nitrocéfine, ceftriaxone,
ticarcilline. Les β-lactamines en poudre sont utilisées comme substrats, tandisque en disques
elles sont utilisées pour les antibiogrammes. Le tampon phosphate à pH 7 est utilisé à 100mM
pour l’extraction des enzymes et à 50mM pour les expériences de cinétique enzymatique.
IV-1-2- les produits chimiques fournis par la maison Vel
Ce sont des composés essentiellement minéraux. Les différentes formes du phosphate
(mono et disodique, mono et dipotassique). Ces composés sont utilisés pour la préparation des

Matériel et Méthodes
46
solutions tampons. Vel est aussi le fournisseur de chlorure de potassium (KCl), du chlorure de
sodium (NaCl), de l’hydroxyde de sodium (NaOH).
IV-1-3- les produits chimiques fournis par la maison Difco
Ces produits regroupent principalement les milieux « Luria-Bertani » (L.B) pour la
préparation de bouillons de culture et des géloses de soja pour la préparation des milieux
solides. Les milieux sont préparés en dissolvant 25g de poudre par litre d’eau distillée de
« Luria-Bertani » et 40g de poudre par litre d’eau distillée de gélose de soja tryptique. Dans
tous les cas le pH du milieu est ajusté à 7, puis les milieux sont stérilisés à 121ºC pendant
15mn.
IV-2-Les souches bactériennes
Cent quatre vingts (180) souches de bactéries à Gram négatif résistantes au moins à
une céphalosporine de troisième génération (C3G) ont été isolées entre juillet 2010 et mars
2012 au Centre Hospitalier Universitaire Pédiatrique Charles De Gaulle (CHUP-CDG) de
Ouagadougou au Burkina Faso. Ces souches ont été isolées de différents services de l’hôpital
et sont issues de divers prélèvements incluant principalement pus et urines provenant de
patients de zéro (0) à quatorze (14) ans. L’identification bactérienne a été réalisée par les
techniques conventionnelles (Farmer, 1999).
V-Méthodes
V-1-L’isolement et identification des souches bactériennes d’intérêt
Les travaux ayant conduit à l’isolement des souches se sont déroulés dans les laboratoires
d’analyse médicale (service de Bactériologie, …) où sont traités des échantillons biologiques
que sont entre autres les selles, les urines, le sang, le liquide céphalo-rachidien et les liquides
d’épanchement (pus). L’isolement comporte plusieurs étapes.

Matériel et Méthodes
47
V-1-2 - L’examen à l’état frais
Les échantillons sont examinés en deux phases :
Examen macroscopique
Examen microscopique
L’examen macroscopique s’intéresse à l’état de l’échantillon biologique. Selon le type
d’échantillon, les paramètres visés sont : couleur, turbidité, aspect (liquide, semi liquide,
pâteux…). Les échantillons suspects de souillure extérieure sont éliminés.
L’examen microscopique s’intéresse à l’état frais et à la coloration du frottis. A cette étape on
recherche la morphologie et le Gram des bactéries. Après l’examen macroscopique, un
étalement de l’échantillon entre lame et lamelle est examiné au microscope afin d’évaluer la
densité et la morphologie des éléments cellulaires et autres éléments associés. Après l’examen
à l’état frais, l’échantillon biologique est ensemencé sur un milieu de culture approprié et
incubé à 37°C pendant 18-24 heures. Les colonies obtenues sur milieu solide et présentant des
caractéristiques de souches d’intérêt sont réisolées sur milieu solide afin d’être identifiées.
V-1-3 – Identification des souches bactériennes
Après réisolement, les bactéries obtenues sont identifiées. D’abord un examen à l’état frais
permet la détermination de la morphologie et de la mobilité des bactéries présentes dans
l’échantillon biologique. Ensuite on étudie les caractères biochimiques afin de confirmer
l’identité de ces germes. Pour les tests biochimiques, des galeries minimales et le système API
20 (bioMérieux France) sont utilisés. Une galerie minimale renferme cinq milieux :
Le milieu KLIGLER HAJNA qui permet d’étudier la productivité de gaz, de
H2S, l’utilisation du glucose et de lactose par les bactéries.
Le milieu mannitol mobilité qui permet d’étudier la fermentation du mannitol
par les bactéries et leur mobilité.

Matériel et Méthodes
48
Le milieu citrate de Simmons qui permet d’étudier l’utilisation du carbone par
les bactéries.
Le milieu urée -indole qui permet d’étudier l’utilisation de l’urée et de l’indole
Eau peptonée (EP) qui permet de rechercher la production d’indole
Certains milieux ont besoin d’ajoût de révélateur pour la lecture. Par exemple le réactif de
Kovacks permet la révélation de l’indole dans l’urée indole ou dans l’eau peptonée. L’ajout de
ce réactif dans ces milieux présente un anneau rouge à la surface quand le test est positif.
L’urée initialement orange vire au rouge quand le test est positif.
Le citrate initialement vert vire au bleu quand le test est positif.
Le système API 20 consiste à préparer une suspension d’une colonie bactérienne pure dans
2ml de liquide physiologique. Cette suspension est utilisée pour inoculer les puits de la
galerie API. Après 24heures d’incubation à 37°C, la lecture des caractères biochimiques qui
nécessite pour certains des réactifs de révélation permet de déterminer habituellement le genre
et l’espèce de la souche.
Des fiches techniques d’identification biochimique ont permis d’identifier les germes en
fonction des résultats obtenus des tests biochimiques.
V-2-Sensibilité aux antibiotiques et détection des BLSE
La sensibilité aux antibiotiques a été déterminée par antibiogramme qui a été réalisé
par la méthode de diffusion en milieu gélosé de Muller-Hinton (MH) vis-à-vis des β-
lactamines selon les recommandations du Comité de l’antibiogramme de la Société française
de microbiologie.
Pour cela, on réalise une première suspension d’une colonie bactérienne pure (à l’aide
d’une solution physiologique sérum salé isotonique) dont la turbidité doit être équivalente à
celle du Mac Farland 0,5 qui est une solution de chlorure de baryum. Ensuite on procède à

Matériel et Méthodes
49
une dilution au 1/10è ou au 1/100è selon le type d’ensemencement qu’on aura à faire. Ainsi
un ensemencement par écouvillonnage nécessite une dilution au 1/10è tandis qu’un
ensemencement par inondation nécessite une dilution au 1/100è. L’ensemencement se réalise
sur le milieu MH coulé dans des boites de Pétri et dont l’épaisseur doit être de 4mm. Après un
temps de séchage (5-10mn), des disques d’antibiotiques sont déposés sur le milieu avec une
distance minimale de 20mm. Après une incubation de 18 à 24 heures à 37°, on mesure le
diamètre des zones d’inhibition autour des antibiotiques qui traduit la susceptibilité des
souches bactériennes à ces antibiotiques. En comparant ce diamètre à des standards (norme
française CA-SFM) on détermine le phénotype sensible(S), intermédiaire(I), ou résistant (R)
de la souche bactérienne vis-à-vis de l’antibiotique. Ainsi toutes les souches bactériennes
ayant présenté une résistance à au moins une céphalosporine de troisième génération sont
présumées productrices de BLSE. Ces souches ont été retenues pour la réalisation des tests de
synergie dont la positivité rend plus probant le profil BLSE. Mais retenons que le test de
synergie peut conduire à des faux négatifs ou faux positifs (Rashed et al., 1997) d’où l’intérêt
d’autres méthodes analytiques.
Les antibiotiques testés ont été l’amoxicilline, l’amoxicilline/acide clavulanique,
ticarcilline, ticarcilline/acide clavulanique, pipéracilline, céfazoline, céfoxitine, céfotaxime,
ceftriaxone, ceftazidime, cefpirome, cefépime, aztréoname, imipénème, amikacine,
gentamicine, ciprofloxacine, norfoxacine, ofloxacine, triméthoprime/sulfaméthoxazole.
Pour la détermination des concentrations minimales inhibitrices (CMI) des
antibiotiques ciblés (céfuroxime, céfotaxime, ceftriaxone, ceftazidime, cefépime, imipénème)
des concentrations croissantes de ces antibiotiques ont été mélangées à un milieu solide
(milieu TSA) à son état liquide dans différents flacons. Les mélanges ont été réalisés à 55°C
et ont été coulés dans des boîtes de pétri. Après solidification des milieux coulés, les
différentes boîtes de Pétri ont été ensuite ensemencées à partir d’une même suspension de

Matériel et Méthodes
50
culture d’une souche pure en milieu liquide. Les boîtes ont été ensuite incubées à 37°C à
l’étuve GALLENKAMP et les résultats ont été lus 24 heures après.
V-3-condition de culture des souches
Pour les identifications et les purifications, les bactéries ont été cultivées sur du milieu
MH (Mueller-Hinton) et sur du milieu TSA (Triptyque soy agar) en boîtes de pétri et incubées
à 37°C pendant 24h. Pour les extractions, les bactéries ont été cultivées dans un bouillon de
culture Luria-Bertani tamponné à pH 7, sous agitation à 250 tours par minute et à 37°C à
l’incubateur agitateur Aquatron. Les inductions ont été réalisées au moyen de la cefoxitine.
V-4-Extraction des BLSE
Chez les bactéries à gram-négatif, les β-lactamases sont normalement sécrétées et
maintenues dans le périplasme. En pratique, les extraits bruts effectués à des fins analytiques
(mise en évidence et mesure de l’activité β-lactamases) ont toujours porté sur 4 mL de
suspension cellulaires à 108 UFC/mL. Le culot bactérien obtenu après centrifugation à
3000tours par minutes pendant 30minutes à 4°C à la centrifugeuse (BR4i) a été mis en
suspension par 300µl de tampon phosphate 100mM pH 7. Les contenus de l’espace
périplasmique ont été libérés par la méthode physique des cycles de
congélation /décongélation (Bidwell et Reeves, 1980 ; Simpson et James, 1982). Le principe
de la méthode consiste à préparer un bain froid en refroidissant de l’acétone par la neige
carbonique. On obtient un milieu liquide qui se trouve à une température allant de -65°C à -
75°C. Les cellules sont immergées pendant 8mn dans ce milieu ce qui entraîne une
congélation instantanée. Ensuite elles sont transférées dans un Bain-marie à 37°C pour
décongélation également instantanée. Ces changements brusques d’état (état congelé - état
décongelé et inversement) provoquent une pulvérisation des parois bactériennes avec
libération des BLSE qui sont localisées dans le compartiment periplasmique des bactéries. Les
échantillons sont ensuite centrifugés à 8000 tours par minute pendant 20mn à la micro

Matériel et Méthodes
51
centrifugeuse MSE. Les débris cellulaires sont ensuite éliminés et les surnageants récupérés et
testés pour l’activité β-lactamase.
V-5-Hydrolyse des β-lactamines par les extraits bruts de BLSE
Les différents extraits bactériens obtenus ont été testés en vue de doser leur activité β-
lactamase avec la nitrocéfine. Cette β-lactamine chromogénique est un substrat passe-partout
de la quasi-totalité des β-lactamases. Un milieu réactionnel constitué de tampon phosphate
50mM pH7, d’enzyme (5 à 10µl) et de nitrocéfine en concentration finale de 100µM est
réalisé dans une cuvette en quartz. Les extraits présentant une activité β-lactamase ont été
testés avec différents β-lactamines dans le but de déterminer leur profil hydrolytique. A cet
effet, des solutions d’antibiotiques (β-lactamines) à une concentration de 2mM ont été
préparées dans des tubes ependorf. Ces solutions sont utilisées comme substrat dans un milieu
réactionnel où la concentration finale d’antibiotique varie de façon croissante (25µM ; 50µM
et 100µM).
Les activités enzymatiques ont été suivies sur un spectrophotomètre UV/VIS double
faisceau (Uvikon 923). Les courbes cinétiques d’hydrolyse des noyaux β-lactames par les
différents extraits ont été enregistrées aux longueurs d’ondes appropriées, à 482 nm (lorsque
la nitrocefine est utilisée comme substrat), à 260 nm (pour le céfotaxime, la ceftazidime, la
céfuroxine, la ceftriaxone, le cefépime). L’hydrolyse de l’imipénème est suivie à 300 nm.
Toutes ces expériences ont été effectuées à 30°C dans du tampon phosphate 50mM pH 7. La
vitesse initiale (Vo) d’hydrolyse de chaque composé (exprimée en unités d’absorbance par
minute) a été évaluée selon la relation Vo = ΔAλ / Δt dans laquelle ΔAλ / Δt représente la
pente à l’origine de la courbe de variation d’absorbance à la longueur d’onde λ en fonction
du temps. La Vo obtenue et exprimée en unités d’absorbances/mn est convertible en µM de
substrats transformés ou de produits libérés par la relation Vo = ΔAλ / Δt x 1 / Δελ dans
laquelle Δελ est la différence entre l’extinction molaire du produit et du substrat à la longueur

Matériel et Méthodes
52
d’onde λ. Le spectrophotomètre Uvikon visualise l’absorbance/temps en temps quasi réel. Le
programme « Time drive » à vocation essentiellement cinétique a été utilisé. Dans ce cas
l’appareil calcule l’activité (pente) par la régression linéaire et donne la valeur chiffrée en
unité d’absorbance par minute. Si le tronçon de la courbe correspondant aux premiers points
(proches de zéro) est sélectionné et si les conditions : [S]o>>[E]o et [S] ≈[S]o sont satisfaites,
l’activité mesurée correspond à la vitesse dite initiale (Vo) (Zeba, 2005).
V-6-Extraction des ADNs
Les génomes des différentes souches ont été obtenus par extraction au DNAZOL dont les
grandes étapes sont : désorganisation des membranes, élimination des débrits membranaires,
précipitation des ADNs, lavage et séchage des ADNs. L’extraction des ADNs bactériens a porté
sur 1mL de suspension bactérienne dont la turbidité est comparée au Mac Farland 0.5 équivalente
à 108 UFC/mL en suivant les instructions du fabriquant du DNAZOL.
V-7-Amplification des gènes de BLSE
La PCR a été faite dans un volume réactionnel de 25 μL. Trois programmes d’amplification
ont été utilisés pour la recherche des trois gènes (Tableaux II).
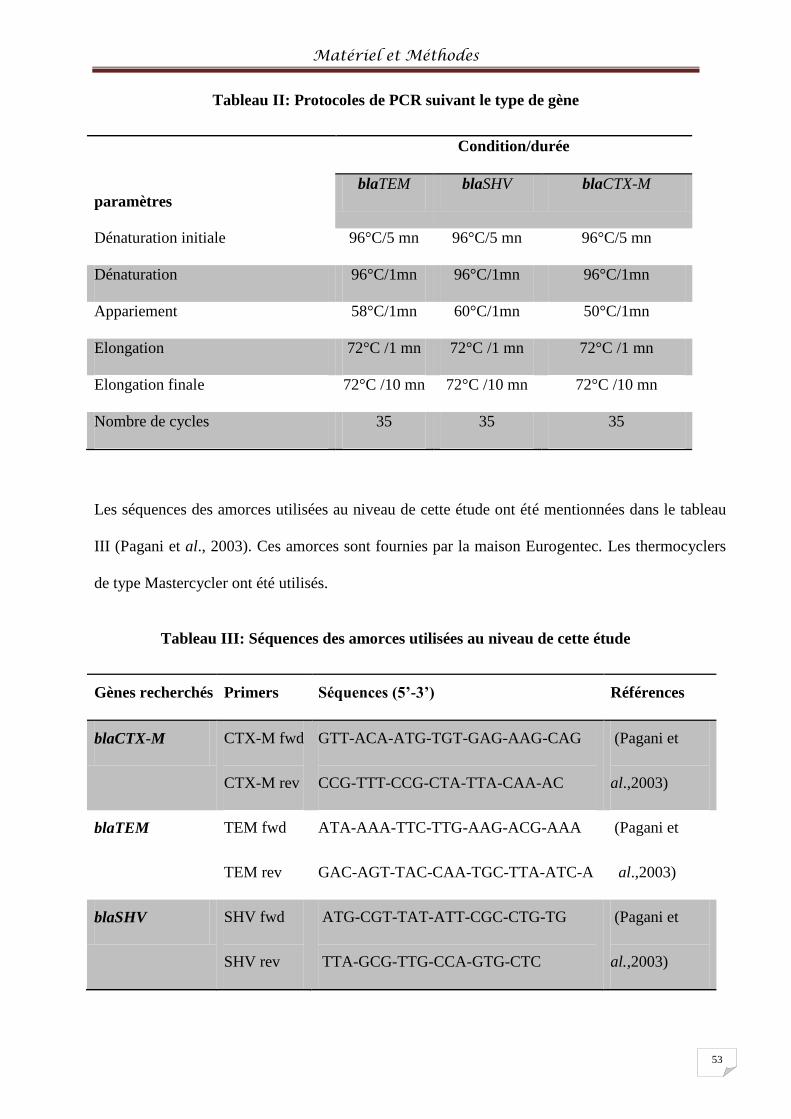
Matériel et Méthodes
53
Tableau II: Protocoles de PCR suivant le type de gène
paramètres
Condition/durée
blaTEM blaSHV blaCTX-M
Dénaturation initiale 96°C/5 mn 96°C/5 mn 96°C/5 mn
Dénaturation 96°C/1mn 96°C/1mn 96°C/1mn
Appariement 58°C/1mn 60°C/1mn 50°C/1mn
Elongation 72°C /1 mn 72°C /1 mn 72°C /1 mn
Elongation finale 72°C /10 mn 72°C /10 mn 72°C /10 mn
Nombre de cycles 35 35 35
Les séquences des amorces utilisées au niveau de cette étude ont été mentionnées dans le tableau
III (Pagani et al., 2003). Ces amorces sont fournies par la maison Eurogentec. Les thermocyclers
de type Mastercycler ont été utilisés.
Tableau III: Séquences des amorces utilisées au niveau de cette étude
Gènes recherchés Primers Séquences (5’-3’) Références
blaCTX-M CTX-M fwd
CTX-M rev
GTT-ACA-ATG-TGT-GAG-AAG-CAG
CCG-TTT-CCG-CTA-TTA-CAA-AC
(Pagani et
al.,2003)
blaTEM TEM fwd
TEM rev
ATA-AAA-TTC-TTG-AAG-ACG-AAA
GAC-AGT-TAC-CAA-TGC-TTA-ATC-A
(Pagani et
al.,2003)
blaSHV SHV fwd
SHV rev
ATG-CGT-TAT-ATT-CGC-CTG-TG
TTA-GCG-TTG-CCA-GTG-CTC
(Pagani et
al.,2003)

Matériel et Méthodes
54
V-8-Electrophorèse sur gel d’agarose
Les fragments d’ADN amplifiés par PCR étaient séparés par électrophorèse sur un gel
d’agarose à 1,5 % préparé dans une solution de tris base - borate – EDTA 0,5X et contenant
du bromure d’éthidium. La migration s’est réalisée sous une tension de 110 millivolts (mV).
Le tampon de migration utilisé a été le Tris-EDTA -Acétate (TEA) (Tris-HCL, 1,6mM, pH 8 ;
EDTA, 4 μM ; Acétate de sodium, 1,6mM). La migration a été suivie grâce à une solution de
dépôt [bleu de bromophénol 0,25% (p/v) ; xylène-cyanol 0,25% (p/v) ; EDTA 25 mM ;
glycérol 50% (v/v)] ajoutée à la solution d’ADN à environ 20% du volume. Les tailles des
différents fragments ont été déterminées en déposant à coté des puits contenant l’ADN des
marqueurs de masses moléculaires. Ces fragments étaient visualisés sous lumière UV. Des
fragments d’ADN de 1000 paires de base (pb) étaient obtenus en cas de présence de blaCTX-
M ou de blaTEM. Un fragment d’ADN de 875 paires de base (pb) était obtenu en cas de
présence de blaSHV.
V-9-Séquençage des gènes de BLSE
Les produits PCR comprenant les régions codantes des différents gènes ont été
séquencés en utilisant les amorces suivantes : TEM-F et TEM-R pour blaTEM, SHV-F et
SHV-R pour blaSHV, CTX-M-F et CTX-M-R pour blaCTX-M. Les amorces « R »
s’apparient à l’extrémité 5’, à l’intérieur de la région codante et les amorces « F » s’apparient
à l’extrémité 3’ à l’intérieur de la région codante de chaque gène. Ainsi le séquençage dans les
deux sens a permis d’obtenir la séquence complète de chaque gène et de corriger les erreurs
de séquençage. Le séquençage des différents gènes a été réalisé par la société GATC Biotech
(http://www.gatc-biotech.com/fr/home) qui est le prestataire leader de séquençage d’ADN et
de bioinformatique pour l’industrie et la recherche académique. Toutes les amorces utilisées
dans cette étude sont présentées au tableau III. Les différentes séquences obtenues ont été
analysées avec le logiciel BLAST (Basic Local Alignment Search Tool) disponible sur le site
Internet du National Center for Biotechnology Information (http://wwww.ncbi.nih.gov), le

Matériel et Méthodes
55
logiciel ‘Reverse Complement’ disponible sur le site Internet
http://www.bioinformatics.org/sms/rev_comp.html, le logiciel Clustalw2 disponible sur le site
Internet http://www.ebi.ac.uk/Tools/msa/clustalw2/ et avec ApE (A plasmid editor Etreprise)
version 8.5.2.0
V-10-Numéros d’accession des séquences
Les numéros d’accession des séquences d’identification et de comparaison de nos gènes sont :
-CTX-M-1: Sequence ID: |emb|X92506.1|
-CTX-M-15: Sequence ID: gb|JQ686199.1|
-TEM-1: Sequence ID: gb|JQ941741.1|
- pKPN101-IT: Sequence ID: gb|JX283456.1|
-SHV-1: Sequence ID: gb|GU064389.1|
- SHV-11: Sequence ID: gb|GU064384.1|
-SHV-12: Sequence ID: gb|GU732834.1|
-SHV-28: Sequence ID: gb|HQ877609.1|
-SHV-32: Sequence ID: gb|AY037778.1|
-SHV-38: Sequence ID: gb|GQ407125.1|
-SHV-76: Sequence ID: emb|AM176551.2|
- SHV-99: Sequence ID: emb|AM922305.1|
- OKP-B: Sequence ID: emb|AM850919.2|
III-10-Traitement des données
Les données ont été analysées par le logiciel Standard Statistical Package for Social

Matériel et Méthodes
56
Sciences (SPSS) version 17.0 pour Windows et par Excel version 2007.
Résultats
57
CHAPITRE III : RESULTATS
I- Souches bactériennes
Parmi les 180 souches bactériennes à Gram négatif résistantes à au moins une céphalosporine
de troisième génération (C3G), les différentes espèces qui ont été étudiées sont E. coli (n=85),
Klebsiella pneumoniae (n=28), K. oxytoca (n=6), Proteus mirabilis (n=3), Klebsiella sp
(n=15), Pseudomonas sp (n=9), Pseudomonas aeruginosa (n=3), Enterobacter sp (n=27),
Enterobacter agglomerans (n=1), Salmonella sp (n=2), Salmonella groupe B (n=1). (Tableau
IV)
Tableau IV: Fréquence des souches bactériennes
Espèces Nombre Pourcentage (%)
E.coli 85 47,22
K.pneumoniae 28 15,55
K.oxytoca 6 3,33
Klebsiella sp 15 8,33
Pseudomonas sp 9 5
Pseudomonas 3 1,67
Enterobacter sp 27 15
Enterobacter agglomerans 1 0,56
Proteus mirabilis 3 1,67
Salmonella sp 2 1,11
Salmonella groupe B 1 0,56
TOTAL 180 100

Résultats
58
II- Les BLSE détectées
La résistance des bactéries cibles aux C3G peut être un indice de la présence de BLSE
(Figure7).
Figure 7: Phénotype de résistance sur boîte de Pétri des souches présumées productrices
de BLSE : Klebsiella pneumoniae en (a) et E.coli en (b) (Mètuor-Dabiré et al., 2010)
En considérant comme souche résistante une souche non sensible, c'est-à-dire une souche
catégorisée résistante ou intermédiaire, il ressort que la sensibilité globale des souches
bactériennes isolées montre une forte résistance au céfuroxime (92,8%), au céfotaxime
(87,8%) et au ceftriaxone (68,9%). Les antibiotiques les plus actifs sur ces souches ont été
l’imipénème (100% de sensibilité), le céfépime (97,2% de sensibilité) et la ceftazidime (94,4
% de sensibilité).
La présomption de BLSE est renforcée par le test de synergie d’action entre l’augmentin
(AMC) et une céphalosporine de troisième génération (C3G) qui donne une image de synergie
d’action en cas de présomption positive de BLSE (Figure 8).
CTX
CRO
AMC
SXT
CIP
AN IPM
NA
GM
CF
PIP
NOR
CN
CTX IPM
AN
CRO
GM
AMC
CIP
NOR SXT
GM CF
PIP
a b
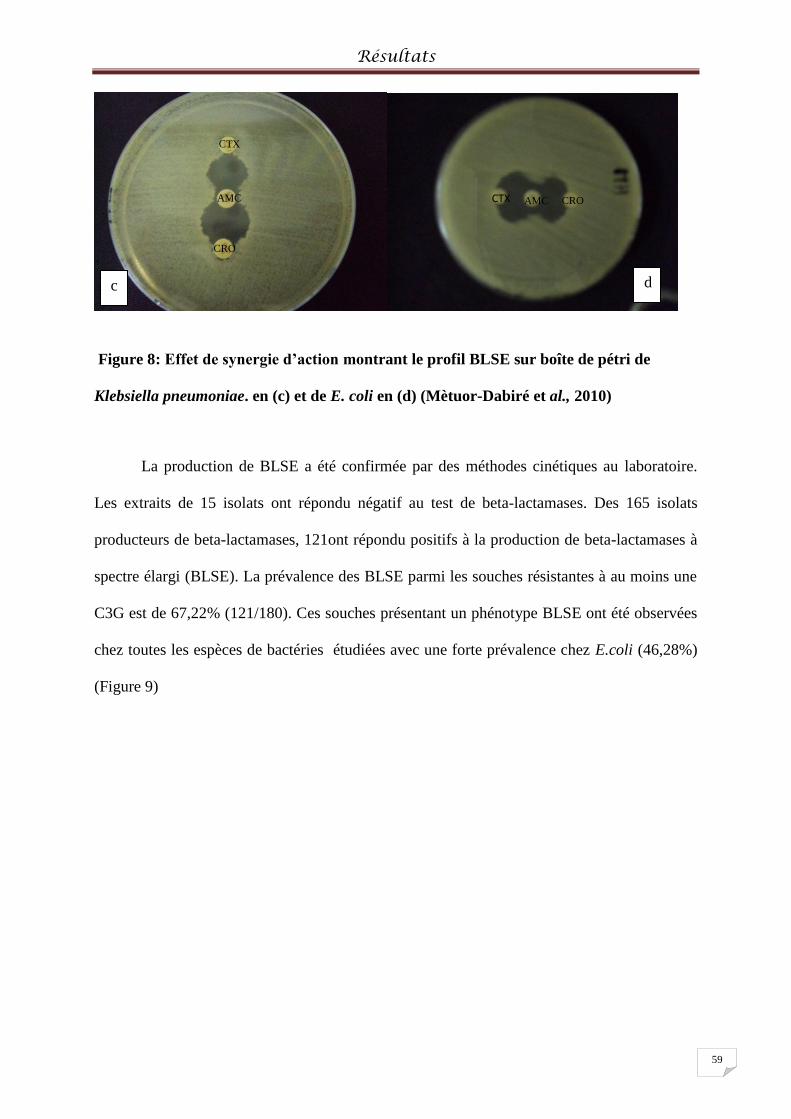
Résultats
59
Figure 8: Effet de synergie d’action montrant le profil BLSE sur boîte de pétri de
Klebsiella pneumoniae. en (c) et de E. coli en (d) (Mètuor-Dabiré et al., 2010)
La production de BLSE a été confirmée par des méthodes cinétiques au laboratoire.
Les extraits de 15 isolats ont répondu négatif au test de beta-lactamases. Des 165 isolats
producteurs de beta-lactamases, 121ont répondu positifs à la production de beta-lactamases à
spectre élargi (BLSE). La prévalence des BLSE parmi les souches résistantes à au moins une
C3G est de 67,22% (121/180). Ces souches présentant un phénotype BLSE ont été observées
chez toutes les espèces de bactéries étudiées avec une forte prévalence chez E.coli (46,28%)
(Figure 9)
CTX
AMC
CRO
CTX AMC CRO
c d
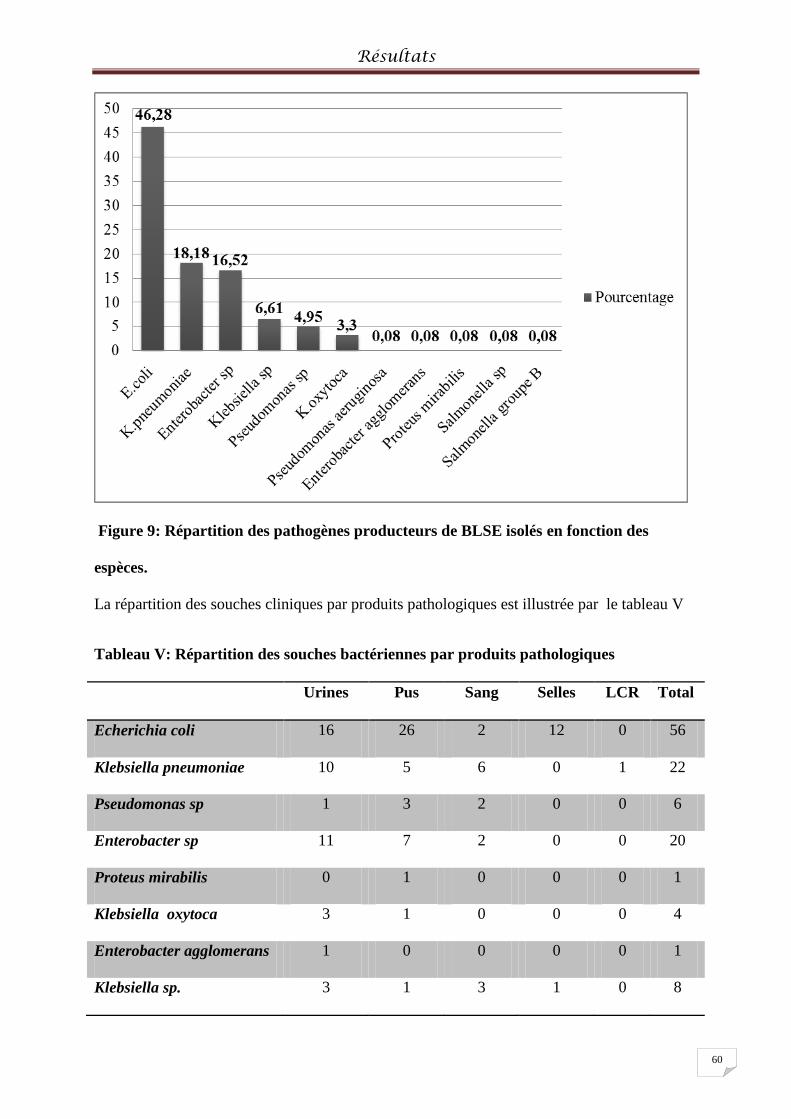
Résultats
60
Figure 9: Répartition des pathogènes producteurs de BLSE isolés en fonction des
espèces.
La répartition des souches cliniques par produits pathologiques est illustrée par le tableau V
Tableau V: Répartition des souches bactériennes par produits pathologiques
Urines Pus Sang Selles LCR Total
Echerichia coli 16 26 2 12 0 56
Klebsiella pneumoniae 10 5 6 0 1 22
Pseudomonas sp 1 3 2 0 0 6
Enterobacter sp 11 7 2 0 0 20
Proteus mirabilis 0 1 0 0 0 1
Klebsiella oxytoca 3 1 0 0 0 4
Enterobacter agglomerans 1 0 0 0 0 1
Klebsiella sp. 3 1 3 1 0 8
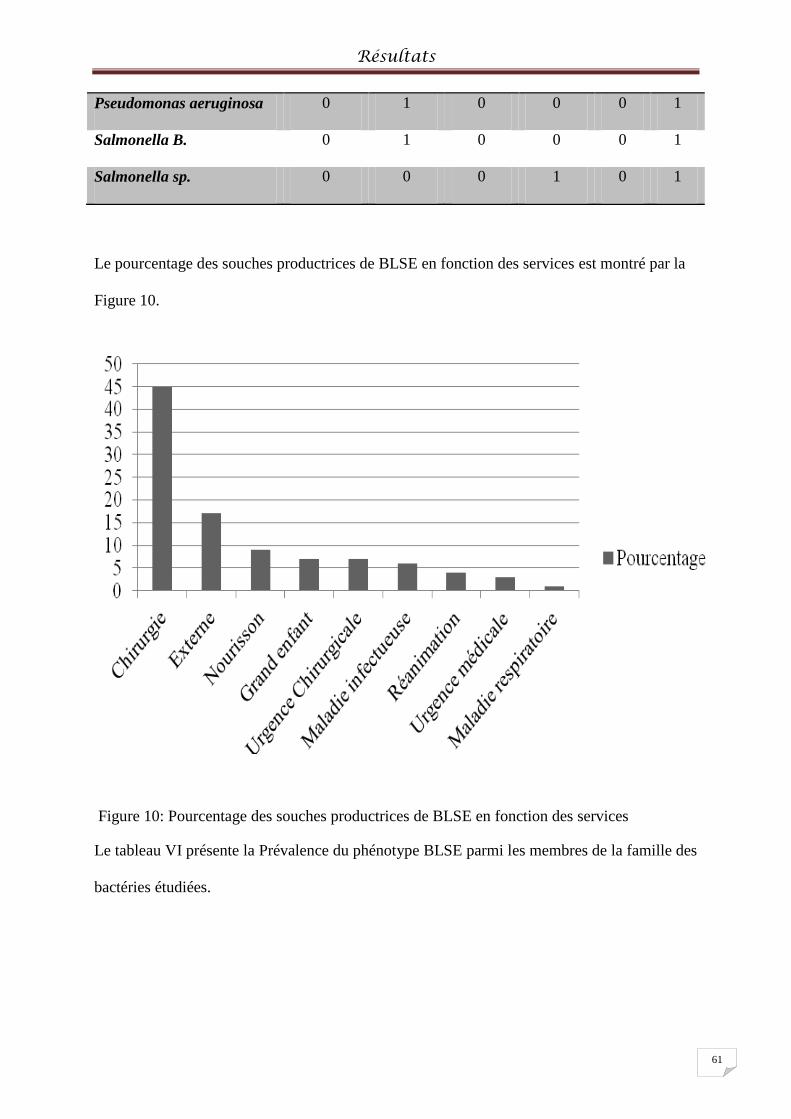
Résultats
61
Pseudomonas aeruginosa 0 1 0 0 0 1
Salmonella B. 0 1 0 0 0 1
Salmonella sp. 0 0 0 1 0 1
Le pourcentage des souches productrices de BLSE en fonction des services est montré par la
Figure 10.
Figure 10: Pourcentage des souches productrices de BLSE en fonction des services
Le tableau VI présente la Prévalence du phénotype BLSE parmi les membres de la famille des
bactéries étudiées.
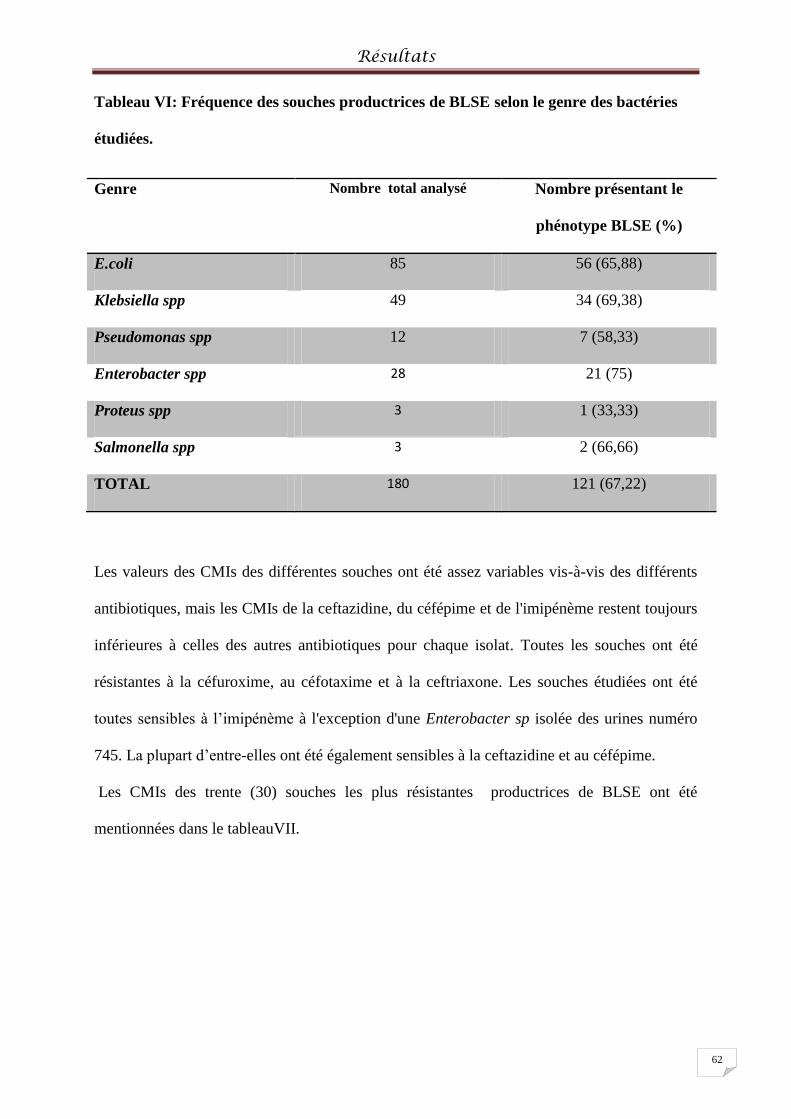
Résultats
62
Tableau VI: Fréquence des souches productrices de BLSE selon le genre des bactéries
étudiées.
Genre Nombre total analysé Nombre présentant le
phénotype BLSE (%)
E.coli 85 56 (65,88)
Klebsiella spp 49 34 (69,38)
Pseudomonas spp 12 7 (58,33)
Enterobacter spp 28 21 (75)
Proteus spp 3 1 (33,33)
Salmonella spp 3 2 (66,66)
TOTAL 180 121 (67,22)
Les valeurs des CMIs des différentes souches ont été assez variables vis-à-vis des différents
antibiotiques, mais les CMIs de la ceftazidine, du céfépime et de l'imipénème restent toujours
inférieures à celles des autres antibiotiques pour chaque isolat. Toutes les souches ont été
résistantes à la céfuroxime, au céfotaxime et à la ceftriaxone. Les souches étudiées ont été
toutes sensibles à l’imipénème à l'exception d'une Enterobacter sp isolée des urines numéro
745. La plupart d’entre-elles ont été également sensibles à la ceftazidine et au céfépime.
Les CMIs des trente (30) souches les plus résistantes productrices de BLSE ont été
mentionnées dans le tableauVII.
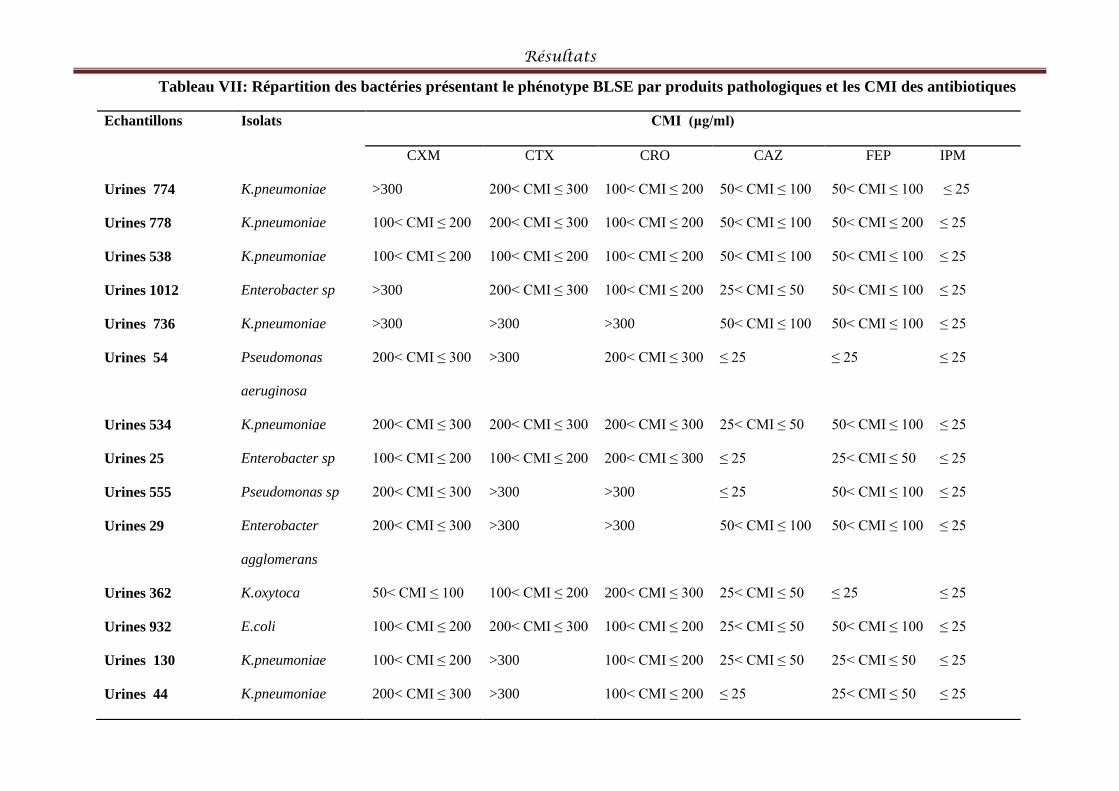
Résultats
Echantillons Isolats CMI (μg/ml)
CXM CTX CRO CAZ FEP IPM
Urines 774 K.pneumoniae >300 200< CMI ≤ 300 100< CMI ≤ 200 50< CMI ≤ 100 50< CMI ≤ 100 ≤ 25
Urines 778 K.pneumoniae 100< CMI ≤ 200 200< CMI ≤ 300 100< CMI ≤ 200 50< CMI ≤ 100 50< CMI ≤ 200 ≤ 25
Urines 538 K.pneumoniae 100< CMI ≤ 200 100< CMI ≤ 200 100< CMI ≤ 200 50< CMI ≤ 100 50< CMI ≤ 100 ≤ 25
Urines 1012 Enterobacter sp >300 200< CMI ≤ 300 100< CMI ≤ 200 25< CMI ≤ 50 50< CMI ≤ 100 ≤ 25
Urines 736 K.pneumoniae >300 >300 >300 50< CMI ≤ 100 50< CMI ≤ 100 ≤ 25
Urines 54 Pseudomonas
aeruginosa
200< CMI ≤ 300 >300 200< CMI ≤ 300 ≤ 25 ≤ 25 ≤ 25
Urines 534 K.pneumoniae 200< CMI ≤ 300 200< CMI ≤ 300 200< CMI ≤ 300 25< CMI ≤ 50 50< CMI ≤ 100 ≤ 25
Urines 25 Enterobacter sp 100< CMI ≤ 200 100< CMI ≤ 200 200< CMI ≤ 300 ≤ 25 25< CMI ≤ 50 ≤ 25
Urines 555 Pseudomonas sp 200< CMI ≤ 300 >300 >300 ≤ 25 50< CMI ≤ 100 ≤ 25
Urines 29 Enterobacter
agglomerans
200< CMI ≤ 300 >300 >300 50< CMI ≤ 100 50< CMI ≤ 100 ≤ 25
Urines 362 K.oxytoca 50< CMI ≤ 100 100< CMI ≤ 200 200< CMI ≤ 300 25< CMI ≤ 50 ≤ 25 ≤ 25
Urines 932 E.coli 100< CMI ≤ 200 200< CMI ≤ 300 100< CMI ≤ 200 25< CMI ≤ 50 50< CMI ≤ 100 ≤ 25
Urines 130 K.pneumoniae 100< CMI ≤ 200 >300 100< CMI ≤ 200 25< CMI ≤ 50 25< CMI ≤ 50 ≤ 25
Urines 44 K.pneumoniae 200< CMI ≤ 300 >300 100< CMI ≤ 200 ≤ 25 25< CMI ≤ 50 ≤ 25
Tableau VII: Répartition des bactéries présentant le phénotype BLSE par produits pathologiques et les CMI des antibiotiques
ciblés.
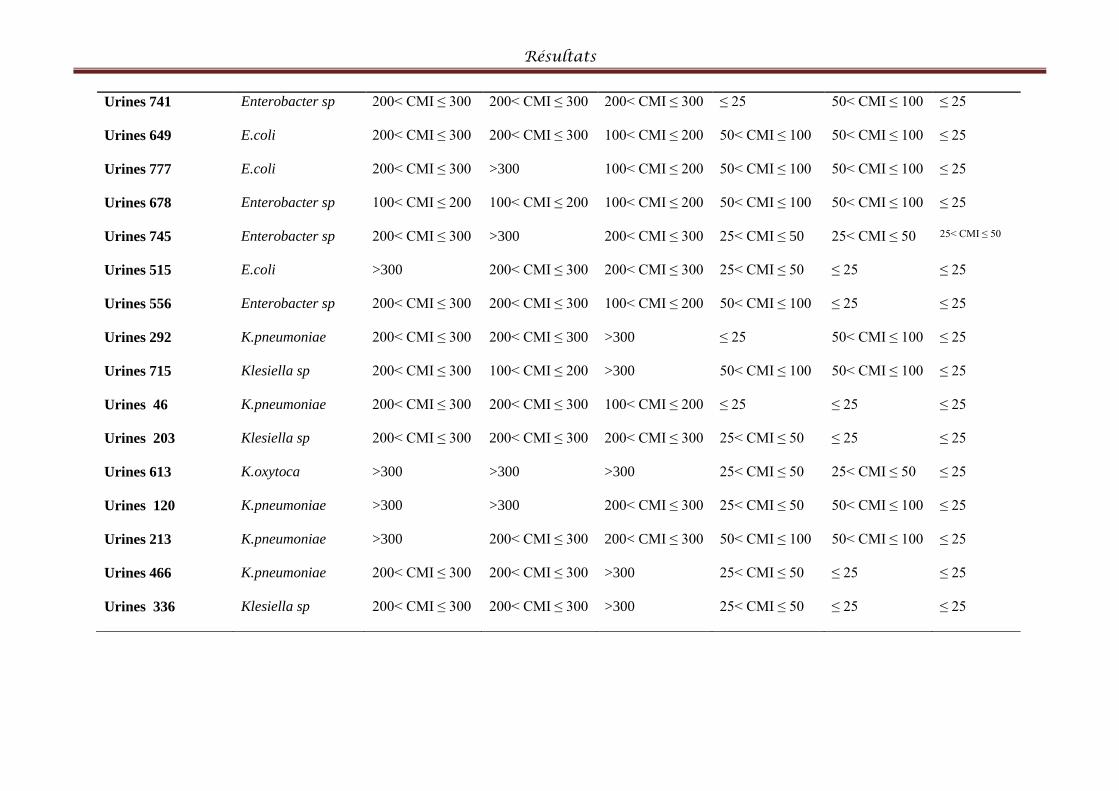
Résultats
Urines 741 Enterobacter sp 200< CMI ≤ 300 200< CMI ≤ 300 200< CMI ≤ 300 ≤ 25 50< CMI ≤ 100 ≤ 25
Urines 649 E.coli 200< CMI ≤ 300 200< CMI ≤ 300 100< CMI ≤ 200 50< CMI ≤ 100 50< CMI ≤ 100 ≤ 25
Urines 777 E.coli 200< CMI ≤ 300 >300 100< CMI ≤ 200 50< CMI ≤ 100 50< CMI ≤ 100 ≤ 25
Urines 678 Enterobacter sp 100< CMI ≤ 200 100< CMI ≤ 200 100< CMI ≤ 200 50< CMI ≤ 100 50< CMI ≤ 100 ≤ 25
Urines 745 Enterobacter sp 200< CMI ≤ 300 >300 200< CMI ≤ 300 25< CMI ≤ 50 25< CMI ≤ 50 25< CMI ≤ 50
Urines 515 E.coli >300 200< CMI ≤ 300 200< CMI ≤ 300 25< CMI ≤ 50 ≤ 25 ≤ 25
Urines 556 Enterobacter sp 200< CMI ≤ 300 200< CMI ≤ 300 100< CMI ≤ 200 50< CMI ≤ 100 ≤ 25 ≤ 25
Urines 292 K.pneumoniae 200< CMI ≤ 300 200< CMI ≤ 300 >300 ≤ 25 50< CMI ≤ 100 ≤ 25
Urines 715 Klesiella sp 200< CMI ≤ 300 100< CMI ≤ 200 >300 50< CMI ≤ 100 50< CMI ≤ 100 ≤ 25
Urines 46 K.pneumoniae 200< CMI ≤ 300 200< CMI ≤ 300 100< CMI ≤ 200 ≤ 25 ≤ 25 ≤ 25
Urines 203 Klesiella sp 200< CMI ≤ 300 200< CMI ≤ 300 200< CMI ≤ 300 25< CMI ≤ 50 ≤ 25 ≤ 25
Urines 613 K.oxytoca >300 >300 >300 25< CMI ≤ 50 25< CMI ≤ 50 ≤ 25
Urines 120 K.pneumoniae >300 >300 200< CMI ≤ 300 25< CMI ≤ 50 50< CMI ≤ 100 ≤ 25
Urines 213 K.pneumoniae >300 200< CMI ≤ 300 200< CMI ≤ 300 50< CMI ≤ 100 50< CMI ≤ 100 ≤ 25
Urines 466 K.pneumoniae 200< CMI ≤ 300 200< CMI ≤ 300 >300 25< CMI ≤ 50 ≤ 25 ≤ 25
Urines 336 Klesiella sp 200< CMI ≤ 300 200< CMI ≤ 300 >300 25< CMI ≤ 50 ≤ 25 ≤ 25

Résultats
65
III- Les BLSE caractérisées
III-1- Le profil hydrolytique des céphalosporines de troisième génération
(C3G) par les BLSEs
Les enzymes de toutes les cent quatre vingts (180) souches étudiées ont été extraites et
analysées au spectrophotomètre. Les substrats utilisés sont : la nitrocéfine qui est un substrat
test de beta-lactamases, le Céfuroxime (CXM) qui est une puissante C2G, le céfotaxime
(CTX), la Ceftriaxone (CRO) et la ceftazidine (CAZ) qui sont des C3G (les substrats cibles
des BLSE), le Céfépime (FEP) qui est une C4G et l’imipénème (IPM) qui est une
carbapenème.
Les extraits de 15 isolats ont répondu négatifs au test de beta-lactamases. Des 165 isolats
producteurs de beta-lactamases, 121ont répondu positifs à la production de beta-lactamases à
spectre élargi (BLSE) c’est-à-dire hydrolysent au moins une C3G. Ces extraits hydrolysent
fortement le céfuroxime, le céfotaxime(CTX) et la ceftriaxone (CRO) et hydrolysent à un
moindre dégrée la ceftazidime (CAZ) et le céfépime (CFM). Dans nos conditions
expérimentales aucun extrait de nos isolats n’hydrolyse l’imipenème (IPM) (Tableau VIII)
Tableau VIII: Nombre et pourcentage des extraits en fonction des substrats hydrolysés
Substrats
Nitrocéfine
(CPR)
Céfuroxime
(CXM)
Céfotaxime
(CTX)
Ceftriaxone
(CRO)
Céfépime
(FEP)
Ceftazidime
(CAZ)
Imipenème
(IPM)
Nombre
D’extraits
121 121 121 23 1 1 0
Les courbes d’hydrolyse des différents substrats sont représentées par les figures suivantes :
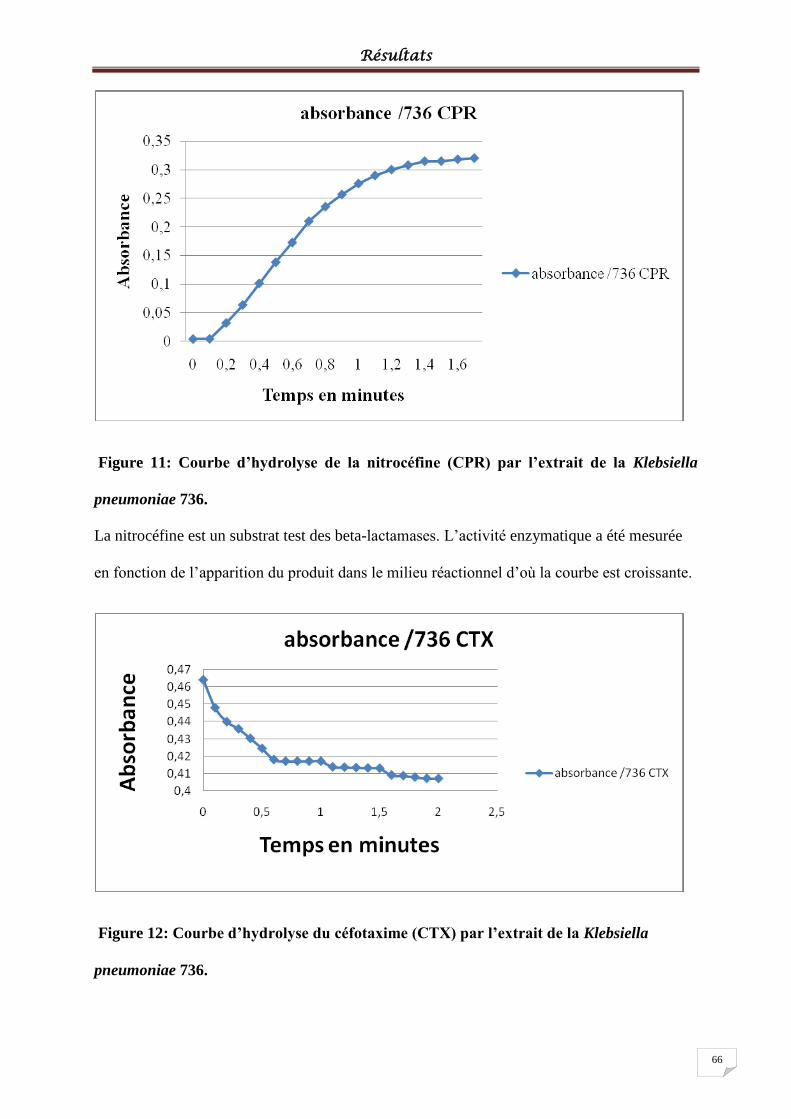
Résultats
66
Figure 11: Courbe d’hydrolyse de la nitrocéfine (CPR) par l’extrait de la Klebsiella
pneumoniae 736.
La nitrocéfine est un substrat test des beta-lactamases. L’activité enzymatique a été mesurée
en fonction de l’apparition du produit dans le milieu réactionnel d’où la courbe est croissante.
Figure 12: Courbe d’hydrolyse du céfotaxime (CTX) par l’extrait de la Klebsiella
pneumoniae 736.
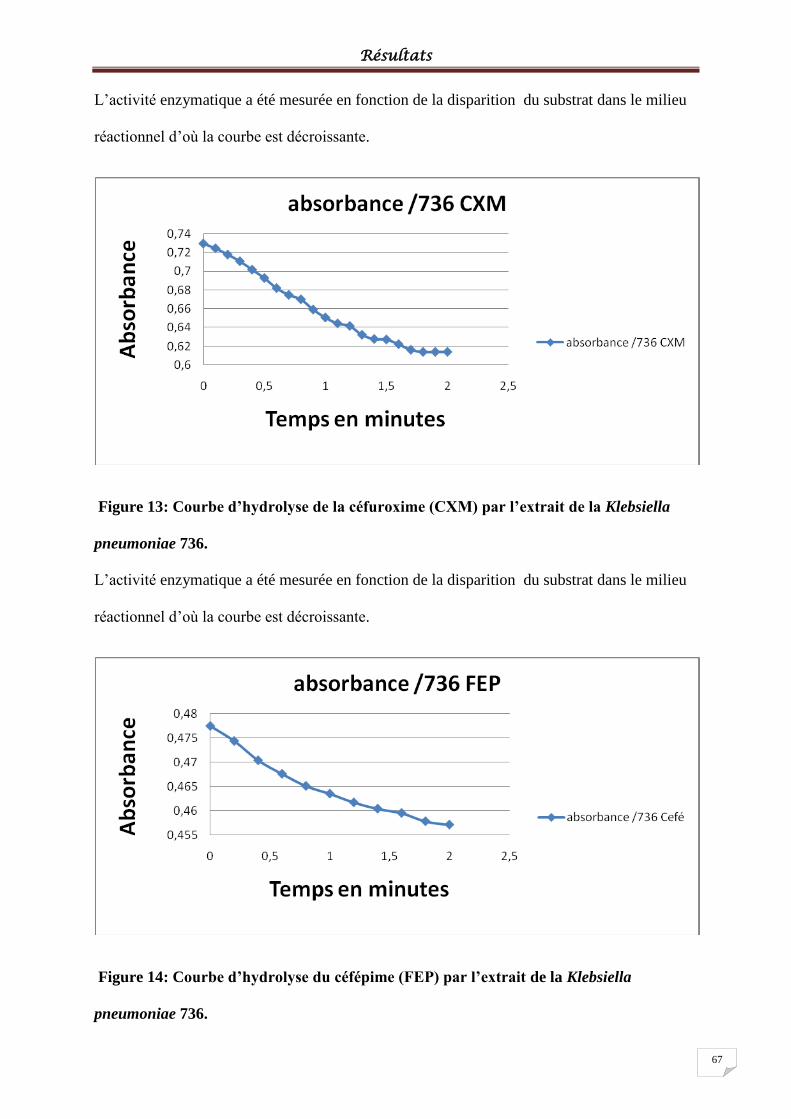
Résultats
67
L’activité enzymatique a été mesurée en fonction de la disparition du substrat dans le milieu
réactionnel d’où la courbe est décroissante.
Figure 13: Courbe d’hydrolyse de la céfuroxime (CXM) par l’extrait de la Klebsiella
pneumoniae 736.
L’activité enzymatique a été mesurée en fonction de la disparition du substrat dans le milieu
réactionnel d’où la courbe est décroissante.
Figure 14: Courbe d’hydrolyse du céfépime (FEP) par l’extrait de la Klebsiella
pneumoniae 736.
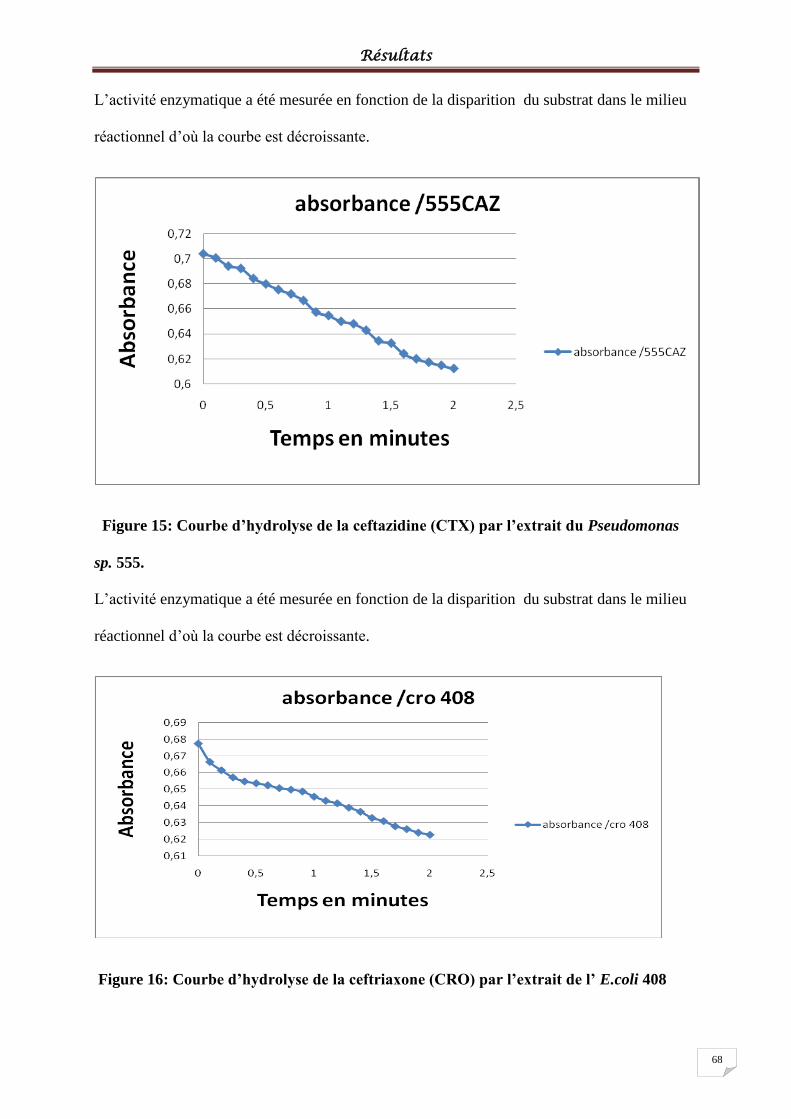
Résultats
68
L’activité enzymatique a été mesurée en fonction de la disparition du substrat dans le milieu
réactionnel d’où la courbe est décroissante.
Figure 15: Courbe d’hydrolyse de la ceftazidine (CTX) par l’extrait du Pseudomonas
sp. 555.
L’activité enzymatique a été mesurée en fonction de la disparition du substrat dans le milieu
réactionnel d’où la courbe est décroissante.
Figure 16: Courbe d’hydrolyse de la ceftriaxone (CRO) par l’extrait de l’ E.coli 408
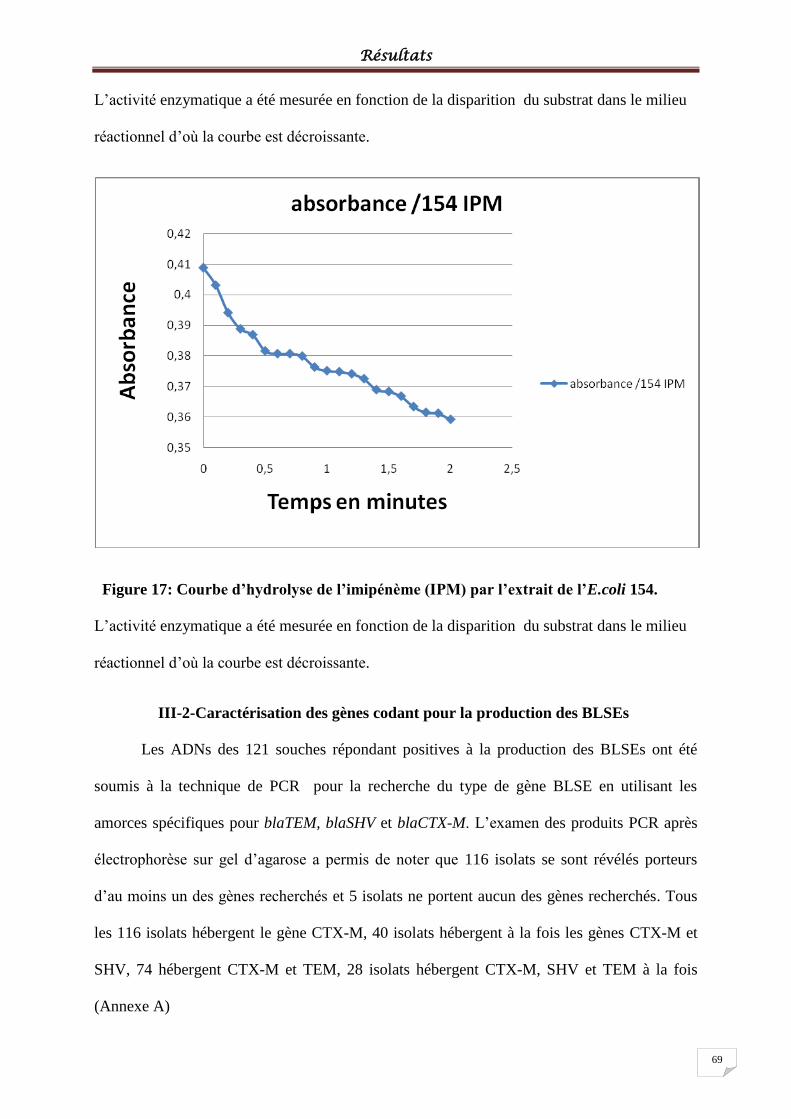
Résultats
69
L’activité enzymatique a été mesurée en fonction de la disparition du substrat dans le milieu
réactionnel d’où la courbe est décroissante.
Figure 17: Courbe d’hydrolyse de l’imipénème (IPM) par l’extrait de l’E.coli 154.
L’activité enzymatique a été mesurée en fonction de la disparition du substrat dans le milieu
réactionnel d’où la courbe est décroissante.
III-2-Caractérisation des gènes codant pour la production des BLSEs
Les ADNs des 121 souches répondant positives à la production des BLSEs ont été
soumis à la technique de PCR pour la recherche du type de gène BLSE en utilisant les
amorces spécifiques pour blaTEM, blaSHV et blaCTX-M. L’examen des produits PCR après
électrophorèse sur gel d’agarose a permis de noter que 116 isolats se sont révélés porteurs
d’au moins un des gènes recherchés et 5 isolats ne portent aucun des gènes recherchés. Tous
les 116 isolats hébergent le gène CTX-M, 40 isolats hébergent à la fois les gènes CTX-M et
SHV, 74 hébergent CTX-M et TEM, 28 isolats hébergent CTX-M, SHV et TEM à la fois
(Annexe A)

Résultats
70
Des amplicons de 1000 paires de bases (pb) étaient obtenus en cas de présence de blaCTX-M
ou de blaTEM (Figure 18et 19) et des amplicons de 875 paires de bases (pb) étaient obtenus
en cas de présence de blaSHV (Figure 20)
Figure 18: Gel d’agarose des produits PCR de CTX-M
M1, Marqueur de poids moléculaire (GeneRuler 100pb DNA Ladder), M2, Marqueur de
poids moléculaire (GeneRuler 1Kb DNA Ladder), les numéros 1-9 représentent les
échantillons: 1=Uro774; 2=Uro778; 3=Uro538; 4=Uro1012; 5=LCR736; 6=Uro534;
7=Uro25; 8=Copro290; 9=Pus54. Le sens de migration de l’électrophorèse est du haut vers le
bas
Figure 19 : M1, Marqueur de poids moléculaire (GeneRuler 100pb DNA Ladder), les
numéros 1-5 représentent les échantillons: 1=Uro362; 2=Uro932; 3=Hemo44; 4=Uro745;
5=Pus130. Le sens de migration de l’électrophorèse est du haut vers le bas.
1000 Pb
Figure 19: Gel d’agarose des produits PCR de TEM

Résultats
71
Figure 20 : M1, Marqueur de poids moléculaire (GeneRuler 100pb DNA Ladder), les numéros
1-7 représentent les échantillons: 1=Uro556; 2=Uro292; 3=Uro715; 4=Hemo46; 5=Uro613;
6=Uro466; 7=Hemo120. Le sens de migration de l’électrophorèse est du haut vers le bas.
Les répartitions des gènes en fonction des échantillons, des services et en fonction des
espèces sont mentionnées dans les tableaux ci-dessous (Tableau IX, Tableau X et Tableau
XI).
Au niveau des échantillons biologiques ; les pus hébergent plus de souches produisant de
BLSE de type CTX-M suivis des urines, du sang et des selles. Les isolats qui produisent les
BLSE de type TEM sont beaucoup plus rencontrés et à nombre égal dans les urines et dans les
pus. Ce nombre est suivi de celui du sang, puis des selles. Le grand nombre de souches
productrices de BLSE de type SHV se retrouve dans les urines. Ce nombre est suivi de celui
des souches hébergées par les pus puis celui du sang.
Selon les services, c’est le service de la chirurgie qui possède le plus grand nombre de
souches productrices des trois types de BLSE rencontrées. Au niveau des espèces, CTX-M
est beaucoup plus retrouvé chez E. coli suivie de K.pneumoniae et d’Enterobacter sp. Le
gène TEM est plus hébergé par E. coli suivie simultanément de K. pneumoniae et
875 Pb
1000 Pb
Figure 20: Gel d’agarose des produits PCR de SHV
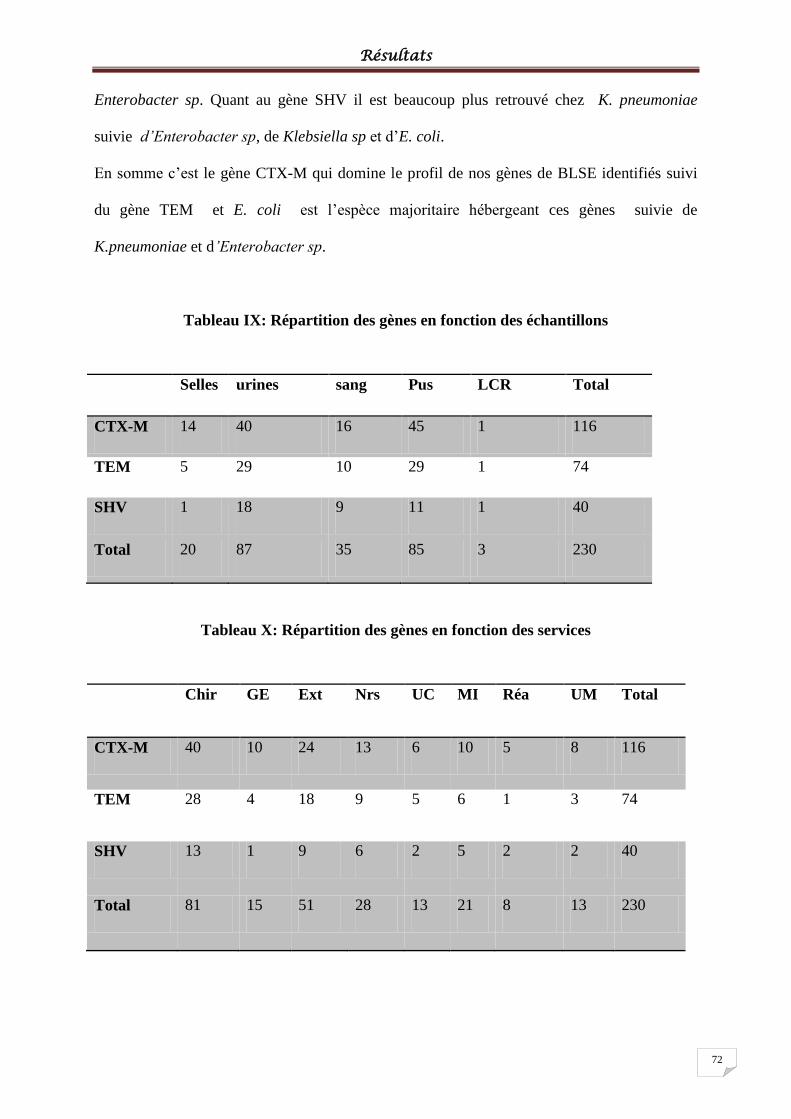
Résultats
72
Enterobacter sp. Quant au gène SHV il est beaucoup plus retrouvé chez K. pneumoniae
suivie d’Enterobacter sp, de Klebsiella sp et d’E. coli.
En somme c’est le gène CTX-M qui domine le profil de nos gènes de BLSE identifiés suivi
du gène TEM et E. coli est l’espèce majoritaire hébergeant ces gènes suivie de
K.pneumoniae et d’Enterobacter sp.
Tableau IX: Répartition des gènes en fonction des échantillons
Selles urines sang Pus LCR Total
CTX-M 14 40 16 45 1 116
TEM 5 29 10 29 1 74
SHV 1 18 9 11 1 40
Total 20 87 35 85 3 230
Tableau X: Répartition des gènes en fonction des services
Chir GE Ext Nrs UC MI Réa UM Total
CTX-M 40 10 24 13 6 10 5 8 116
TEM 28 4 18 9 5 6 1 3 74
SHV 13 1 9 6 2 5 2 2 40
Total 81 15 51 28 13 21 8 13 230
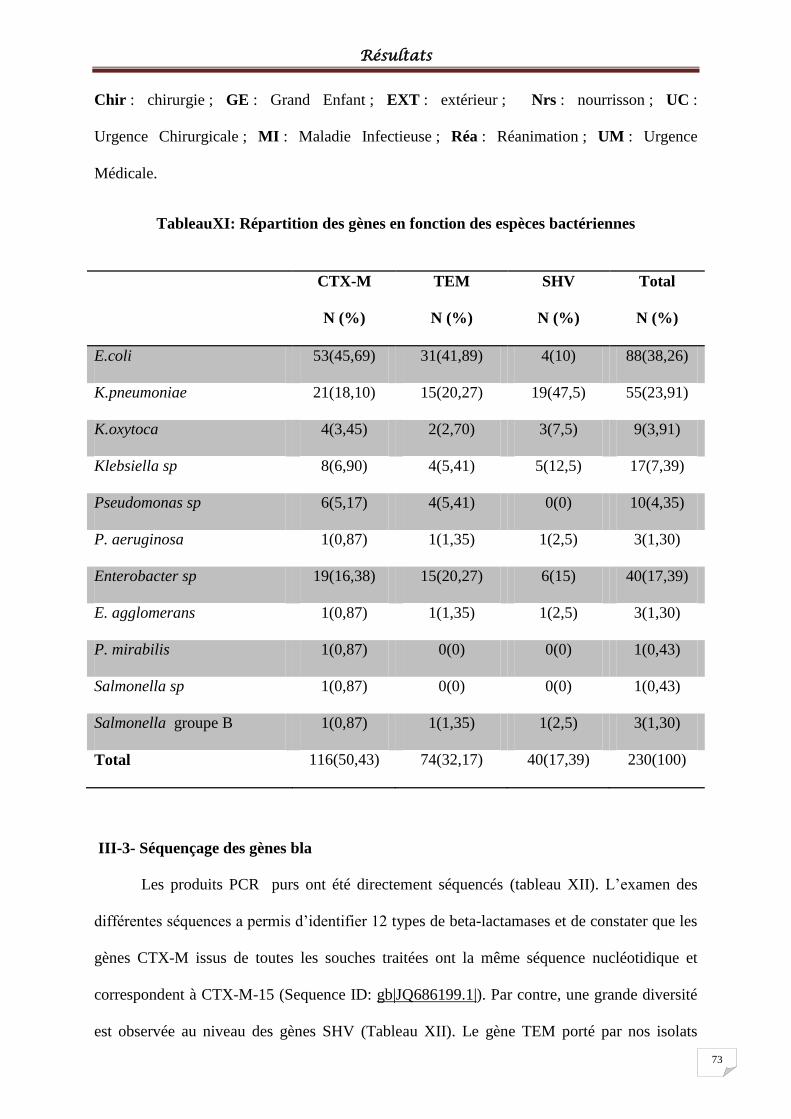
Résultats
73
Chir : chirurgie ; GE : Grand Enfant ; EXT : extérieur ; Nrs : nourrisson ; UC :
Urgence Chirurgicale ; MI : Maladie Infectieuse ; Réa : Réanimation ; UM : Urgence
Médicale.
TableauXI: Répartition des gènes en fonction des espèces bactériennes
CTX-M
N (%)
TEM
N (%)
SHV
N (%)
Total
N (%)
E.coli 53(45,69) 31(41,89) 4(10) 88(38,26)
K.pneumoniae 21(18,10) 15(20,27) 19(47,5) 55(23,91)
K.oxytoca 4(3,45) 2(2,70) 3(7,5) 9(3,91)
Klebsiella sp 8(6,90) 4(5,41) 5(12,5) 17(7,39)
Pseudomonas sp 6(5,17) 4(5,41) 0(0) 10(4,35)
P. aeruginosa 1(0,87) 1(1,35) 1(2,5) 3(1,30)
Enterobacter sp 19(16,38) 15(20,27) 6(15) 40(17,39)
E. agglomerans 1(0,87) 1(1,35) 1(2,5) 3(1,30)
P. mirabilis 1(0,87) 0(0) 0(0) 1(0,43)
Salmonella sp 1(0,87) 0(0) 0(0) 1(0,43)
Salmonella groupe B 1(0,87) 1(1,35) 1(2,5) 3(1,30)
Total 116(50,43) 74(32,17) 40(17,39) 230(100)
III-3- Séquençage des gènes bla
Les produits PCR purs ont été directement séquencés (tableau XII). L’examen des
différentes séquences a permis d’identifier 12 types de beta-lactamases et de constater que les
gènes CTX-M issus de toutes les souches traitées ont la même séquence nucléotidique et
correspondent à CTX-M-15 (Sequence ID: gb|JQ686199.1|). Par contre, une grande diversité
est observée au niveau des gènes SHV (Tableau XII). Le gène TEM porté par nos isolats

Résultats
74
analysés est le type TEM-1. Il a été également mis en évidence au sein de deux souches
bactériennes par le séquençage des amplicons de TEM, un grand plasmide dénommé
pKPN101-IT hébergeant blaKPC-2 (blaKlebsiella pneumoniae carbapenèmase). Le gène
OKP-B, un variant de la famille du blaOKP a été retrouvé chez une Klebsiella sp en
séquençant les amplicons de SHV.
Tableau XII: Caractéristiques moléculaires des gènes hébergés par les isolats traités
Isolats
gène Bla
Types de beta-lactamases CTX-M TEM SHV
K.pneumoniae 774 + + + CTX-M-15 ; TEM-1 ; SHV-12
K.pneumoniae778 + + + CTX-M-15 ; TEM-1 ; SHV-32
K.pneumoniae 538 + + + CTX-M-15 ; TEM-1 ; SHV-76
Enterobacter sp1012 + + + CTX-M-15 ; TEM-1 ; SHV-11
K.pneumoniae736 + + + CTX-M-15 ; pKPN101-IT ; SHV-1
Enterobacter sp25 + + - CTX-M-15 ; TEM-1
K.oxytoca362 + + - CTX-M-15 ; TEM-1
E.coli 932 + + - CTX-M-15 ; TEM-1
K.pneumoniae130 + + - CTX-M-15 ; TEM-1
K.pneumoniae44 + + - CTX-M-15 ; TEM-1
Enterobacter sp741 + + - CTX-M-15 ; TEM-1
E.coli649 + + - CTX-M-15 ; TEM-1
E.coli777 + + - CTX-M-15 ; TEM-1
Enterobacter sp 678 + + - CTX-M-15 ; TEM-1
Enterobacter sp 745 + + - CTX-M-15 ; pKPN101-IT

Résultats
75
E.coli515 + + - CTX-M-15 ; TEM-1
K.pneumoniae534 + - + CTX-M-15 ; SHV-1
Enterobacter sp556 + - + CTX-M-15 ; SHV-11
K.pneumoniae292 + - + CTX-M-15 ; SHV-1
Klesiella sp715 + - + CTX-M-15 ; SHV-11
K.pneumoniae46 + - + CTX-M-15 ; SHV-99
Klesiella sp203 + - + CTX-M-15 ; OKP-B
K.oxytoca613 + - + CTX-M-15 ; SHV-1 1
K.pneumoniae 120 + - + CTX-M-15 ; SHV-38
K.pneumoniae 213 + - + CTX-M-15 ; SHV-11
K.pneumoniae466 + - + CTX-M-15 ; SHV-28
Klesiella sp336 + - + CTX-M-15 ; SHV-11
Pseudomonas sp555 + - - CTX-M-15
Enterobacter agglomerans29 + - - CTX-M-15
Pseudomonas aeruginosa54 +
-
-
CTX-M-15
La comparaison entre la séquence nucléotidique du gène CTX-M-15 de nos souches et la
séquence standard du gène CTX-M-1 (GenBank accession No. X92506) (figure 21) nous a
permis de noter cinq (05) mutations qui entraînent des substitutions d’acides aminés. Par
rapport au gène CTX-M-1, les différentes mutations après la numérotation des acides aminés
suivant Ambler et al. (1981) ont été observées aux positions :
- 80 [Valine (CTX-M-1) Alanine (CTX-M-15)] ;
- 117 (Aspartate Asparagine) ;
- 143 (Sérine Alanine),
-242 (Aspartate Glycine) ;
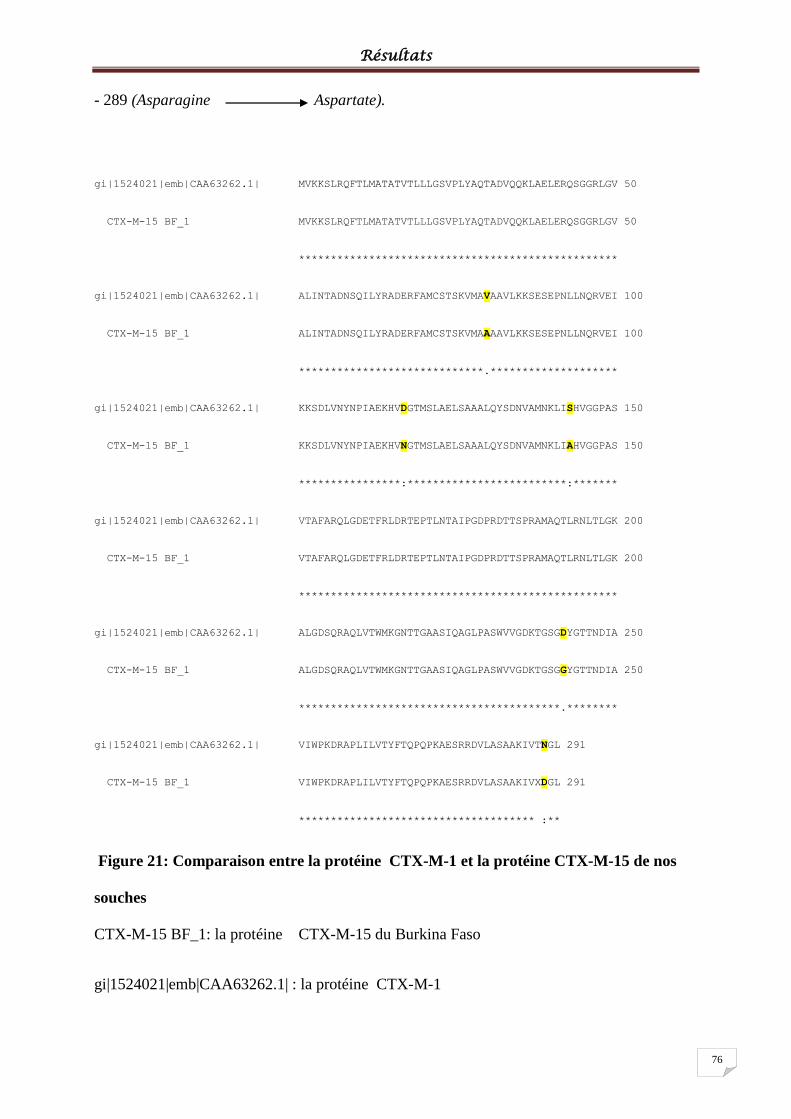
Résultats
76
- 289 (Asparagine Aspartate).
gi|1524021|emb|CAA63262.1| MVKKSLRQFTLMATATVTLLLGSVPLYAQTADVQQKLAELERQSGGRLGV 50
CTX-M-15 BF_1 MVKKSLRQFTLMATATVTLLLGSVPLYAQTADVQQKLAELERQSGGRLGV 50
**************************************************
gi|1524021|emb|CAA63262.1| ALINTADNSQILYRADERFAMCSTSKVMAVAAVLKKSESEPNLLNQRVEI 100
CTX-M-15 BF_1 ALINTADNSQILYRADERFAMCSTSKVMAAAAVLKKSESEPNLLNQRVEI 100
*****************************.********************
gi|1524021|emb|CAA63262.1| KKSDLVNYNPIAEKHVDGTMSLAELSAAALQYSDNVAMNKLISHVGGPAS 150
CTX-M-15 BF_1 KKSDLVNYNPIAEKHVNGTMSLAELSAAALQYSDNVAMNKLIAHVGGPAS 150
****************:*************************:*******
gi|1524021|emb|CAA63262.1| VTAFARQLGDETFRLDRTEPTLNTAIPGDPRDTTSPRAMAQTLRNLTLGK 200
CTX-M-15 BF_1 VTAFARQLGDETFRLDRTEPTLNTAIPGDPRDTTSPRAMAQTLRNLTLGK 200
**************************************************
gi|1524021|emb|CAA63262.1| ALGDSQRAQLVTWMKGNTTGAASIQAGLPASWVVGDKTGSGDYGTTNDIA 250
CTX-M-15 BF_1 ALGDSQRAQLVTWMKGNTTGAASIQAGLPASWVVGDKTGSGGYGTTNDIA 250
*****************************************.********
gi|1524021|emb|CAA63262.1| VIWPKDRAPLILVTYFTQPQPKAESRRDVLASAAKIVTNGL 291
CTX-M-15 BF_1 VIWPKDRAPLILVTYFTQPQPKAESRRDVLASAAKIVXDGL 291
************************************* :**
Figure 21: Comparaison entre la protéine CTX-M-1 et la protéine CTX-M-15 de nos
souches
CTX-M-15 BF_1: la protéine CTX-M-15 du Burkina Faso
gi|1524021|emb|CAA63262.1| : la protéine CTX-M-1

Résultats
77
La comparaison entre les séquences nucléotidiques des gènes SHV de nos isolats et la
séquence standard du gène SHV-1 (Bradford, 1999) nous a permis de noter plusieurs
différences.
uro213 ---------------GTTAA-----AATNNCGCCTGTGTATTATCTCCCTGTTAGCCACC 40
URO715 ------------------------------CGCCTGTGTATTATCTCCCTGTTAGCCACC 30
URO613 -------------------------------GCCTGTGTATTATCTCCCTGTTAGCCACC 29
uro336 ------------------------------CGCCTGTGTATTATCTCCCTGTTAGCCACC 30
URO46 -------------------------------GNCTGTGTATTATCTCCCTGTTAGCCACC 29
uro120 ------------------------------CGNCTGTGTATTATCTCCCTGTTAGCCACC 30
URO292 ------------------------------CGCCTGTGTATTATCTCCCTGTTAGCCACC 30
uro466 ---------------------------------CTGTGTATTATCTCCCTGTTAGCCACC 27
URO556 ---------------------------------CTGTGTATTATCTCCCTGTTAGCCACC 27
SHV-1 CAAGGATGTATTGTGGTTATGCGTTATATTCGCCTGTGTATTATCTCCCTGTTAGCCACC 240
uro203 ------------------------------CGCCTGTGCCTTATCTCCCTGATTGCCGCC 30
***** .***********:*:***.**
uro213 CTGCCGCTGGCGGTACACGCCAG--CCCGCA-GCCGCTTGAGCAAATTAAACAAAGCGAA 97
URO715 CTGCCGCTGGCGGTACACGCCAG--CCCGCA-GCCGCTTGAGCAAATTAAACAAAGCGAA 87
URO613 CTGCCGCTGGCGGTACACGCCAG--CCCGCA-GCCGCTTGAGCAAATTAAACAAAGCGAA 86
uro336 CTGCCGCTGGCGGTACACGCCAGGCCCCGCA-GCCGCTTGAGCAAATTAAACAAAGCGAA 89
URO46 CTGCCGCTGGCGGTACACGCCAG--CCCGCA-GCCGCTTGAGCAAATTAAACTAAGCGAA 86
uro120 CTGCCGCTGGCGGTACACGCCAGC-CCCGCA-GCCGCTTGAGCAAATTAAACTAAGCGAA 88
URO292 CTGCCGCTGGCGGTACACGCCAG--CCCGCA-GCCGCTTGAGCAAATTAAACTAAGCGAA 87
uro466 CTGCCGCTGGCGGTACACGCCAG--CCCGCA-GCCGCTTGAGCAAATTAAACTAAGCGAA 84
URO556 CTGCCGCTGGCGGTACACGCCAG--CCCGCA-GCCGCTTGAGCAAATTAAACAAAGCGAA 84
SHV-1 CTGCCGCTGGCGGTACACGCCAG--CCCGCA-GCCGCTTGAGCAAATTAAACTAAGCGAA 297
uro203 CTGCCACTGGCGGTATTCGCCAG-CCCCTCA-GCCGCTTGAGCAGATTAAAATCAGCGAA 88
*****.********* :****** *** ** ************.******.:.******
uro213 AGCCAGCT-GTCGGGCCGCGTAGGCATGATAGAAATGGATCTGGCCAGCGGCCGCACGCT 156
URO715 AGCCAGCT-GTCGGGCCGCGTAGGCATGATAGAAATGGATCTGGCCAGCGGCCGCACGCT 146
URO613 AGCCAGCT-GTCGGGCCGCGTAGGCATGATAGAAATGGATCTGGCCAGCGGCCGCACGCT 145
uro336 AGCCAGCT-GTCGGGCCGCGTAGGCATGATAGAAATGGATCTGGCCAGCGGCCGCACGCT 148
URO46 AGCCAGCT-GTCGGGCCGCGTAGGCATGATAGAAATGGATCTGGCCAGCGGCCGCACGCT 145
uro120 AGCCAGCT-GTCGGGCCGCGTAGGCATGATAGAAATGGATCTGGCCAGCGGCCGCACGCT 147
URO292 AGCCAGCATGTCGGGCCGCGTAGGCATGATAGAAATGGATCTGGCCAGCGGCCGCACGCT 147
uro466 AGCCAGCT-GTCGGGCCGCGTAGGCATGATAGAAATGGATCTGGCCAGCGGCCGCACGCT 143
URO556 AGCCAGCT-GTCGGGCCGCGTAGGCATGATAGAAATGGATCTGGCCAGCGGCCGCACGCT 143

Résultats
78
SHV-1 AGCCAGCT-GTCGGGCCGCGTAGGCATGATAGAAATGGATCTGGCCAGCGGCCGCACGCT 356
uro203 GGTCAGCT-GGCGGGCCGGGTGGGCTATGTTGAAATGGATCTGGCCAGCGGCCGCACGCT 147
.* ****: * ******* **.***:: .*:*****************************
uro213 GACCGCCTGGCGCGCCGATGAACGCTTTCCCATGATGAGCACCTTTAAAGTAGTGCTCTG 216
URO715 GACCGCCTGGCGCGCCGATGAACGCTTTCCCATGATGAGCACCTTTAAAGTAGTGCTCTG 206
URO613 GACCGCCTGGCGCGCCGATGAACGCTTTCCCATGATGAGCACCTTTAAAGTAGTGCTCTG 205
uro336 GACCGCCTGGCGCGCCGATGAACGCTTTCCCATGATGAGCACCTTTAAAGTAGTGCTCTG 208
URO46 GACCGCCTGGCGCGCCGATGAACGCTTTCCCATGATGAGCACCTTTAAAGTAGTGCTCTG 205
uro120 GACCGCCTGGCGCGCCGATGAACGCTTTCCCATGATGAGCACCTTTAAAGTAGTGCTCTG 207
URO292 GACCGCCTGGCGCGCCGATGAACGCTTTCCCATGATGAGCACCTTTAAAGTAGTGCTCTG 207
uro466 GACCGCCTGGCGCGCCGATGAACGCTTTCCCATGATGAGCACCTTTAAAGTAGTGCTCTG 203
URO556 GACCGCCTGGCGCGCCGATGAACGCTTTCCCATGATGAGCACCTTTAAAGTAGTGCTCTG 203
SHV-1 GACCGCCTGGCGCGCCGATGAACGCTTTCCCATGATGAGCACCTTTAAAGTAGTGCTCTG 416
uro203 GGCCGCCTGGCGCGCCAGTGAGCGCTTTCCGCTGATGAGCACCTTTAAAGTGCTGCTCTG 207
*.**************..***.******** .*******************. *******
uro213 CGGCGCAGTGCTGGCGCGGGTGGATGCCGGTGACGAACAGCTGGAGCGAAAGATCCACTA 276
URO715 CGGCGCAGTGCTGGCGCGGGTGGATGCCGGTGACGAACAGCTGGAGCGAAAGATCCACTA 266
URO613 CGGCGCAGTGCTGGCGCGGGTGGATGCCGGTGACGAACAGCTGGAGCGAAAGATCCACTA 265
uro336 CGGCGCAGTGCTGGCGCGGGTGGATGCCGGTGACGAACAGCTGGAGCGAAAGATCCACTA 268
URO46 CGGCGCAGTGCTGGCGCGGGTGGATGCCGGTGACGAACAGCTGGAGCGAAAGATCCACTA 265
uro120 CGGCGCAGTGCTGGCGCGGGTGGATGCCGGTGACGAACAGCTGGAGCGAAAGATCCACTA 267
URO292 CGGCGCAGTGCTGGCGCGGGTGGATGCCGGTGACGAACAGCTGGAGCGAAAGATCCACTA 267
uro466 CGGCGCAGTGCTGGCGCGGGTGGATGCCGGTGACGAACAGCTGGAGCGAAAGATCCACTA 263
URO556 CGGCGCAGTGCTGGCGCGGGTGGATGCCGGTGACGAACAGCTGGAGCGAAAGATCCACTA 263
SHV-1 CGGCGCAGTGCTGGCGCGGGTGGATGCCGGTGACGAACAGCTGGAGCGAAAGATCCACTA 476
uro203 CGGCGCGGTGCTGGCCCGGGTGGATGCCGGCGACGAACAGCTGGATCGGCGGATCCACTA 267
******.******** ************** ************** **...*********
uro213 TCGCCAGCAGGATCTGGTGGACTACTCGCCGGTCAGCGAAAAACACCTTGCCGACGGCAT 336
URO715 TCGCCAGCAGGATCTGGTGGACTACTCGCCGGTCAGCGAAAAACACCTTGCCGACGGCAT 326
URO613 TCGCCAGCAGGATCTGGTGGACTACTCGCCGGTCAGCGAAAAACACCTTGCCGACGGCAT 325
uro336 TCGCCAGCAGGATCTGGTGGACTACTCGCCGGTCAGCGAAAAACACCTTGCCGACGGCAT 328
URO46 TCGCCAGCAGGATCTGGTGGACTACTCGCCGGTCAGCGAAAAACATCTTGCCGACGGCAT 325
uro120 TCGCCAGCAGGATCTGGTGGACTACTCGCCGGTCAGCGAAAAACACCTTGCCGACGGCAT 327
URO292 TCGCCAGCAGGATCTGGTGGACTACTCGCCGGTCAGCGAAAAACATCTTGCCGACGGCAT 327
uro466 TCGCCAGCAGGATCTGGTGGACTACTCGCCGGTCAGCGAAAAACATCTTGCCGACGGCAT 323
URO556 TCGCCAGCAGGATCTGGTGGACTACTCGCCGGTCAGCGAAAAACACCTTGCCGACGGCAT 323

Résultats
79
SHV-1 TCGCCAGCAGGATCTGGTGGACTACTCGCCGGTCAGCGAAAAACACCTTGCCGACGGCAT 536
uro203 CCGCCAGCAGGATCTGGTGGACTACTCCCCGGTCAGCGAAAAACACCTTGCCGACGGGAT 327
************************** ***************** *********** **
uro213 GACGGTCGGCGAACTCTGCGCCGCCGCCATTACCATGAGCGATAACAGCGCCGCCAATCT 396
URO715 GACGGTCGGCGAACTCTGCGCCGCCGCCATTACCATGAGCGATAACAGCGCCGCCAATCT 386
URO613 GACGGTCGGCGAACTCTGCGCCGCCGCCATTACCATGAGCGATAACAGCGCCGCCAATCT 385
uro336 GACGGTCGGCGAACTCTGCGCCGCCGCCATTACCATGAGCGATAACAGCGCCGCCAATCT 388
URO46 GACGGTCGGCGAACTCTGTGCCGCCGCCATTACCATGAGCGATAACAGCGCCGCCAATCT 385
uro120 GACGGTCGGCGAACTCTGTGCCGCCGCCATTACCATGAGCGATAACAGCGCCGCCAATCT 387
URO292 GACGGTCGGCGAACTCTGCGCCGCCGCCATTACCATGAGCGATAACAGCGCCGCCAATCT 387
uro466 GACGGTCGGCGAACTCTGCGCCGCCGCCATTACCATGAGCGATAACAGCGCCGCCAATCT 383
URO556 GACGGTCGGCGAACTCTGCGCCGCCGCCATTACCATGAGCGATAACAGCGCCGCCAATCT 383
SHV-1 GACGGTCGGCGAACTCTGCGCCGCCGCCATTACCATGAGCGATAACAGCGCCGCCAATCT 596
uro203 GACCGTTGGCGAACTCTGCGCCGCCGCCATCACCATGAGCGACAACAGCGCCGGCAATCT 387
*** ** *********** *********** *********** ********** ******
uro213 GCTGCTGGCCACCGTCGGCGGCCCCGCAGGATTGACTGCCTTTTTGCGCCAGATCGGCGA 456
URO715 GCTGCTGGCCACCGTCGGCGGCCCCGCAGGATTGACTGCCTTTTTGCGCCAGATCGGCGA 446
URO613 GCTGCTGGCCACCGTCGGCGGCCCCGCAGGATTGACTGCCTTTTTGCGCCAGATCGGCGA 445
uro336 GCTGCTGGCCACCGTCGGCGGCCCCGCAGGATTGACTGCCTTTTTGCGCCAGATCGGCGA 448
URO46 GCTGCTGGCCACCGTCGGCGGCCCCGCAGGATTGACTGCCTTTTTGCGCCAGATCGGCGA 445
uro120 GCTGCTGGCCACCGTCGGCGGCCCCGTAGGATTGACTGCCTTTTTGCGCCAGATCGGCGA 447
URO292 GCTGCTGGCCACCGTCGGCGGCCCCGCAGGATTGACTGCCTTTTTGCGCCAGATCGGCGA 447
uro466 GCTGCTGGCCACCGTCGGCGGCCCCGCAGGATTGACTGCCTTTTTGCGCCAGATCGGCGA 443
URO556 GCTGCTGGCCACCGTCGGCGGCCCCGCAGGATTGACTGCCTTTTTGCGCCAGATCGGCGA 443
SHV-1 GCTACTGGCCACCGTCGGCGGCCCCGCAGGATTGACTGCCTTTTTGCGCCAGATCGGCGA 656
uro203 GCTGTTGAAGAGCGTCGGCGGCCCCGCGGGATTGACCGCTTTTCTGCGCCAGATCGGTGA 447
***. **.. * ************** .******** ** *** ************* **
uro213 CAACGTCACCCGCCTTGACCGCTGGGAAACGGAACTGAATGAGGCGCTTCCCGGCGATGC 516
URO715 CAACGTCACCCGCCTTGACCGCTGGGAAACGGAACTGAATGAGGCGCTTCCCGGCGATGC 506
URO613 CAACGTCACCCGCCTTGACCGCTGGGAAACGGAACTGAATGAGGCGCTTCCCGGCGATGC 505
uro336 CAACGTCACCCGCCTTGACCGCTGGGAAACGGAACTGAATGAGGCGCTTCCCGGCGACGC 508
URO46 CAACGTCACCCGCCTTGACCGCTGGGAAACGGAACTGAATGAGGCGCTTCCCGGCGACGC 505
uro120 CAACGTCACCCGCCTTGACCGCTGGGAAACGGAACTGAATGAGGCGCTTCCCGGCGACGC 507
URO292 CAACGTCACCCGCCTTGACCGCTGGGAAACGGAACTGAATGAGGCGCTTCCCGGCGACGC 507
uro466 CAACGTCACCCGCCTTGACCGCTGGGAAACGGAACTGAATGAGGCGCTTCCCGGCGACGC 503
URO556 CAACGTCACCCGCCTTGACCGCTGGGAAACGGAACTGAATGAGGCGCTTCCCGGCGACGC 503

Résultats
80
SHV-1 CAACGTCACCCGCCTTGACCGCTGGGAAACGGAACTGAATGAGGCGCTTCCCGGCGACGC 716
uro203 CAACGTCACCCGCCTTGACCGCTGGGAAACGGAACTCAATGAGGCGCTTCCCGGCGACGT 507
************************************ ******************** *
uro213 CCGCGACACCACTACCCCGGCCAGCATGGCCGCGACCCTGCGCAAGCTGCTGACCAGCCA 576
URO715 CCGCGACACCACTACCCCGGCCAGCATGGCCGCGACCCTGCGCAAGCTGCTGACCAGCCA 566
URO613 CCGCGACACCACTACCCCGGCCAGCATGGCCGCGACCCTGCGCAAGCTGCTGACCAGCCA 565
uro336 CCGCGACACCACTACCCCGGCCAGCATGGCCGCGACCCTGCGCAAGCTGCTGACCAGCCA 568
URO46 CCGCGACACCACTACCCCGGCCAGCATGGCCGCGACCCTGCGCAAGCTGCTGACCAGCCA 565
uro120 CCGCGACACCACTACCCCGGCCAGCATGGCCGCGACCCTGCGCAAGCTGCTGACCAGCCA 567
URO292 CCGCGACACCACTACCCCGGCCAGCATGGCCGCGACCCTGCGCAAGCTGCTGACCAGCCA 567
uro466 CCGCGACACCACTACCCCGGCCAGCATGGCCGCGACCCTGCGCAAGCTGCTGACCAGCCA 563
URO556 CCGCGACACCACTACCCCGGCCAGCATGGCCGCGACCCTGCGCAAGCTGCTGACCAGCCA 563
SHV-1 CCGCGACACCACTACCCCGGCCAGCATGGCCGCGACCCTGCGCAAGCTGCTGACCAGCCA 776
uro203 GCGCGACACCACCACCCCGGCCAGCATGGCCACCACCCTGCGCAAGCTGCTAACCACCCC 567
*********** ******************.* *****************.**** **.
uro213 GCGTCTGAGCGCCCGTTCGCAACGGCAGCTGCTGCAGTGGATGGTGGACGATCGGGTCGC 636
URO715 GCGTCTGAGCGCCCGTTCGCAACGGCAGCTGCTGCAGTGGATGGTGGACGATCGGGTCGC 626
URO613 GCGTCTGAGCGCCCGTTCGCAACGGCAGCTGCTGCAGTGGATGGTGGACGATCGGGTCGC 625
uro336 GCGTCTGAGCGCCCGTTCGCAACGGCAGCTGCTGCAGTGGATGGTGGACGATCGGGTCGC 628
URO46 GCGTCTGAGCGCCCGTTCGCAACGGCAGCTGCTGCAGTGGATGGTGGACGATCGGGTCGC 625
uro120 GCGTCTGAGCGCCCGTTCGCAACGGCAGCTGCTGCAGTGGATGGTGGACGATCGGGTCGC 627
URO292 GCGTCTGAGCGCCCGTTCGCAACGGCAGCTGCTGCAGTGGATGGTGGACGATCGAGTCGC 627
uro466 GCGTCTGAGCGCCCGTTCGCAACGGCAGCTGCTGCAGTGGATGGTGGACGATCGAGTCGC 623
URO556 GCGTCTGAGCGCCCGTTCGCAACGGCAGCTGCTGCAGTGGATGGTGGACGATCGGGTCGC 623
SHV-1 GCGTCTGAGCGCCCGTTCGCAACGGCAGCTGCTGCAGTGGATGGTGGACGATCGGGTCGC 836
uro203 CTCTCTGAGCGCCCGTTCGCAGCAGCAGCTGCTGCAGTGGATGGTGGACGACCGGGTGGC 627
******************.*.*************************** **.** **
uro213 CGGACCGTTGATCCGCTCCGTGCTGCCGGCGGGCTGGTTTATCGCCGATAAGACCGGAGC 696
URO715 CGGACCGTTGATCCGCTCCGTGCTGCCGGCGGGCTGGTTTATCGCCGATAAGACCGGAGC 686
URO613 CGGACCGTTGATCCGCTCCGTGCTGCCGGCGGGCTGGTTTATCGCCGATAAGACCGGAGC 685
uro336 CGGACCGTTGATCCGCTCCGTGCTGCCGGCGGGCTGGTTTATCGCCGATAAGACCGGAGC 688
URO46 CGGACCGTTGATCCGCTCCGTGCTGCCGGCGGGCTGGTTTATCGCCGATAAGACCGGAGC 685
uro120 CGGACCGTTGATCCGCTCCGTGCTGCCGGCGGGCTGGTTTATCGCCGATAAGACCGGAGC 687
URO292 CGGACCGTTGATCCGCTCCGTGCTGCCGGCGGGCTGGTTTATCGCCGATAAGACCGGAGC 687
uro466 CGGACCGTTGATCCGCTCCGTGCTGCCGGCGGGCTGGTTTATCGCCGATAAGACCGGAGC 683
URO556 CGGACCGTTGATCCGCTCCGTGCTGCCGGCGGGCTGGTTTATCGCCGATAAGACCGGAGC 683

Résultats
81
SHV-1 CGGACCGTTGATCCGCTCCGTGCTGCCGGCGGGCTGGTTTATCGCCGATAAGACCGGAGC 896
uro203 CGGCCCGTTGATCCGCGCCGTGCTGCCGGCGGGCTGGTTTATCGCCGATAAAACCGGGGC 687
***.************ **********************************.*****.**
uro213 TGGCGAACGGGGTGCGCGCGGGATTGTCGCCCTGCTTGGCCCGAATAACAAAGCAGAGCG 756
URO715 TGGCGAACGGGGTGCGCGCGGGATTGTCGCCCTGCTTGGCCCGAATAACAAAGCAGAGCG 746
URO613 TGGCGAACGGGGTGCGCGCGGGATTGTCGCCCTGCTTGGCCCGAATAACAAAGCAGAGCG 745
uro336 TGGCGAGCGGGGTGCGCGCGGAATTGTCGCCCTGCTTGGCCCGAATAACAAAGCAGAGCG 748
URO46 TGGCGAACGGGGTGCGCGCGGGATTGTCGCCCTGCTTGGCCCGAATAACAAAGCAGAGCG 745
uro120 TGGCGAACGGGGTGCGCGCGGGATTGTCGCCCTGCTTGGCCCGAATAACAAAGCAGAGCG 747
URO292 TGGCGAACGGGGTGCGCGCGGGATTGTCGCCCTGCTTGGCCCGAATAACAAAGCAGAGCG 747
uro466 TGGCGAACGGGGTGCGCGCGGGATTGTCGCCCTGCTTGGCCCGAATAACAAAGCAGAGCG 743
URO556 TGGCGAGCGGGGTGCGCGCGGGATTGTCGCCCTGCTTGGCCCGAATAACAAAGCAGAGCG 743
SHV-1 TGGCGAGCGGGGTGCGCGCGGGATTGTCGCCCTGCTTGGCCCGAATAACAAAGCAGAGCG 956
uro203 CGGTGAGCGGGGCTCACGCGGCATTGTCGCCCTGCTCGGCCCGGACGGCAAAGCGGAGCG 747
** **.***** *.***** ************** ******.* ..******.*****
uro213 CATTGTGGTGATTTATCTGCGGGATACCCCGGCGAGCATGGCCG-AGCGAAAT-CAGCAA 814
URO715 CATTGTGGTGATTTATCTGCGGGATACCCCGGCGAGCATGGCCG-AGCGAAANNCAGCA- 804
URO613 CATTGTGGTGATTTATCTGCGGGATACCCCGGCGAGCATGGCCG-AGCGAAAT-CAGCAA 803
uro336 CATTGTGGTGATTTATCTGCGGGATACCCCGGCGAGCATGGCCG-AGCGAAA-TCAGCAA 806
URO46 CATCGTGGTGATTTATCTGCGGGATACGCCGGCGAGCATGGCCG-AGCGAAAT-CAGCAA 803
uro120 GATTGTGGTGATTTATCTGCGGGATACGCCGGCGAGCATGGCCG-AGCGAAAT-CAGCAA 805
URO292 CATCGTGGTGATTTATCTGCGGGATACCCCGGCGAGCATGGCCG-AGCGAAAT-CAGCAA 805
uro466 CATCGTGGTGATTTATCTGCGGGATACCCCGGCGAGCATGGCCG-AGCGAAAT-CAGCAA 801
URO556 CATTGTGGTGATTTATCTGCGGGATACGCCGGCGAGCATGGCCG-AGCGAAAT-CAGCAA 801
SHV-1 CATTGTGGTGATTTATCTGCGGGATACCCCGGCGAGCATGGCCG-AGCGAAAT-CAGCAA 1014
uro203 TATCGTGGTGATCTATCTGCGTGATACCCCGGCGACCATGGTCG-AGCGTAAC-CAGCAG 805
** ******** ******** ***** ******* ***** ** ****:** *****
uro213 ATCGCCGGGATCGGCGCGGCGCTGATCGAG-CCNG------CNAA--------------- 852
URO715 ------------------------------------------------------------
URO613 ATCGCCGGGATCGGCGCGGCGCTGATCGA------------------------------- 832
uro336 ATCGCCGGNATCGGCGCGGCGCTGATCGAG-CA--------CTG---------------- 841
URO46 ATCGCCGGGATCGGCGCGGCGCTGATCGAGCACTG------GNNAA-------------- 843
uro120 ATCGCCGGGATCGGCGCGGCGCTGATCGAGCACTG------CNNAACGC----------- 848
URO292 ATCGCCGGGATCGGCGCGGCGCTGATCGAGCACTG------GNAAACGCTAAA------- 852
uro466 ATCGCCGGGATCGGC--------------------------------------------- 816
URO556 ATCGCCGGGATCGGCGCGGCGCTGATCGAGCATGG------------------------- 836

Résultats
82
SHV-1 ATCGCCGGGATCGGCGCGGCGCTGATCGAGCACTGGCAACGCTAACCCGGCGGTGGCCGC 1074
uro203 ATCGCCGGGATAGGCGCGGCGCTGATCGAGCACTG------C------------------ 841
uro213: SHV-11; URO715: SHV-11; URO613: SHV-11; uro336: SHV-11; URO46: SHV-99; uro120: SHV-
38; URO292: SHV-1; uro466: SHV-28; URO556: SHV-11; uro203:OKP-B
Figure 22: comparaison entre les séquences nucléotidiques des gènes SHV de nos isolats
et la séquence standard du gène SHV-1(Bradford, 1999).
La comparaison entre la séquence nucléotidique du gène KPC-2 hébergé par le plasmide
pKP101-IT retrouvé dans nos isolats et la séquence standard du gène TEM-1 (Sequence ID:
gb|JQ941741.1|) (figure 23) nous a permis également de noter des différences notament la
substitution de C par T et celle de T par C.
TEM-1 -------------------------GAAAGGGCCTCGTGATACGCCTATTTTTATAGGTT 240
uro745 -------------------------GAAAGGGCCTCGTGATACGCTTATTTTTATAGGTT 35
******************** **************
TEM-1 AATGTCATGATAATAATGGTTTCTTAGACGTCAGGTGGCACTTTTCGGGGAAATGTGCGC 300
uro745 AATGTCATGATAATAATGGTTTCTTAGACGTCAGGTGGCACTTTTCGGGGAAATGTGCGC 95
************************************************************
TEM-1 GGAACCCCTATTTGTTTATTTTTCTAAATACATTCAAATATGTATCCGCTCATGAGACAA 360
uro745 GGAACCCCTATTTGTTTATTTTTCTAAATACATTCAAATATGTATCCGCTCATGAGACAA 155
************************************************************
TEM-1 TAACCCTGGTAAATGCTTCAATAATATTGAAAAAGGAAGAGTATGAGTATTCAACATTTT 420
uro745 TAACCCTGATAAATGCTTCAATAATATTGAAAAAGGAAGAGTATGAGTATTCAACATTTC 215
********.**************************************************
TEM-1 CGTGTCGCCCTTATTCCCTTTTTTGCGGCATTTTGCCTTCCTGTTTTTGCTCACCCAGAA 480
uro745 CGTGTCGCCCTTATTCCCTTTTTTGCGGCATTTTGCCTTCCTGTTTTTGCTCACCCAGAA 275
************************************************************
TEM-1 ACGCTGGTGAAAGTAAAAGATGCTGAAGATCAGTTGGGTGCACGAGTGGGTTACATCGAA 540
uro745 ACGCTGGTGAAAGTAAAAGATGCTGAAGATCAGTTGGGTGCACGAGTGGGTTACATCGAA 335
************************************************************

Résultats
83
TEM-1 CTGGATCTCAACAGCGGTAAGATCCTTGAGAGTTTTCGCCCCGAAGAACGTTTTCCAATG 600
uro745 CTGGATCTCAACAGCGGTAAGATCCTTGAGAGTTTTCGCCCCGAAGAACGTTTTCCAATG 395
************************************************************
TEM-1 ATGAGCACTTTTAAAGTTCTGCTATGTGGTGCGGTATTATCCCGTGTTGACGCCGGGCAA 660
uro745 ATGAGCACTTTTAAAGTTCTGCTATGTGGCGCGGTATTATCCCGTGTTGACGCCGGGCAA 455
***************************** ******************************
TEM-1 GAGCAACTCGGTCGCCGCATACACTATTCTCAGAATGACTTGGTTGAGTACTCACCAGTC 720
uro745 GAGCAACTCGGTCGCCGCATACACTATTCTCAGAATGACTTGGTTGAGTACTCACCAGTC 515
************************************************************
TEM-1 ACAGAAAAGCATCTTACGGATGGCATGACAGTAAGAGAATTATGCAGTGCTGCCATAACC 780
uro745 ACAGAAAAGCATCTTACGGATGGCATGACAGTAAGAGAATTATGCAGTGCTGCCATAACC 575
************************************************************
TEM-1 ATGAGTGATAACACTGCTGCCAACTTACTTCTGACAACGATCGGAGGACCGAAGGAGCTA 840
uro745 ATGAGTGATAACACTGCTGCCAACTTACTTCTGACAACGATCGGAGGACCGAAGGAGCTA 635
************************************************************
TEM-1 ACCGCTTTTTTGCACAACATGGGGGATCATGTAACTCGCCTTGATCGTTGGGAACCGGAG 900
uro745 ACCGCTTTTTTGCACAACATGGGGGATCATGTAACTCGCCTTGATCGTTGGGAACCGGAG 695
************************************************************
TEM-1 CTGAATGAAGCCATACCAAACGACGAGCGTGACACCACGATGCCTGCAGCAATGGCAACA 960
uro745 CTGAATGAAGCCATACCAAACGACGAGCGTGACACCACGATGCCTGCAGCAATGGCAACA 755
************************************************************
TEM-1 ACGTTGCGCAAACTATTAACTGGCGAACTACTTACTCTAGCTTCCCGGCAACAATTAATA 1020
uro745 ACGTTGCGCAAACTATTAACTGGCGAACTACTTACTCTAGCTTCCCGGCAACAATTAATA 815
************************************************************
TEM-1 GACTGGATGGAGGCGGATAAAGTTGCAGGACCACTTCTGCGCTCGGCCCTTCCGGCTGGC 1080
uro745 GACTGGATGGAGGCGGATAAAGTTGCAGGACCACTTCTGCGCTCGGCCCTTCCGGCTGGC 875
************************************************************
TEM-1 TGGTTTATTGCTGATAAATCTGGAGCCGGTGAG-CGTGGGTCTCGCGGTATCATTGCAGC 1139
uro745 TGGTTTATTGCTGATAAATCTGGAGCCGGTGAG-CGTGGGTCTCGCGGTATCATTGCAGC 934
********************************* **************************

Résultats
84
TEM-1 ACTGGGGCCAGATGGTAAGCCCTCCCGTATCGTAGTTATCTACACGACGGGG-AGTCAGG 1198
uro745 ACTGGGGCCAGATGGTAAGCCCTCCCGTATCGTAGTTATCTACACGACGGGG-AGTCAGG 993
**************************************************** *******
Figure 23: Comparaison entre la séquence de TEM-1 et la séquence de KPC-2 de nos
souches
TEM-1: Gene bank TEM-1 (Sequence ID: gb|JQ941741.1|) ; uro745: KPC-2 de nos souches

Discussion
85
CHAPITRE IV : DISCUSSION
L’objectif de cette étude était de caractériser les types des BLSEs produites par les
souches de bactéries à gram négatif isolées des enfants malades au Centre Hospitalier
Universitaire Pédiatrique Charles De Gaulle (CHUP-CDG) de Ouagadougou au Burkina Faso.
Nous avons observé le phénotype BLSE chez tous les genres bactériens étudiés. La
prévalence de BLSE a été retrouvée chez 67,22% des 180 souches de bactéries isolées dans
ledit centre. Elle a été de 69,38% chez Klebsiella spp, 65,88% chez E.coli, 58,33% chez
Pseudomonas spp, 75% chez Enterobacter spp, 33,33 % chez Proteus spp et de 66,66 % chez
Salmonella spp.
La caractérisation de ces souches a permis de mettre en évidence la production des
BLSE des types SHV (SHV-12, SHV-28, SHV-32, SHV-38, SHV-76 et SHV-99), CTX-M
(CTX-M-15) et une carbapénèmase de type KPC-2 (hébergée par le plasmide pKPN101-IT)
par les espèces de bactéries isolées. A notre connaissance, cette étude constitue la première
qui caractérise les BLSE au Burkina Faso.
Les résultats de la sensibilité aux antibiotiques ont montré en général que les bactéries
présentent un taux de résistance aux antibiotiques communément utilisés. Toutes les bactéries
étudiées ont montré un haut niveau de résistance vis-à-vis de certains antibiotiques
(céfuroxime, céfotaxime et ceftriaxone). Plusieurs souches multirésistantes ont été observées.
Cette multirésistance peut s’expliquer par le fait que les gènes responsables de ces résistances
peuvent être portés par le même plasmide, par la coexistence de plusieurs mécanismes de
résistance, ou par la production de plusieurs types enzymatiques (Harbottle et al, 2006;
Boerlin et Reid-Smith, 2008 ; Muylaert et Mainil, 2012).
Les taux de résistance que nous avons observés sont très élevés pour certains
antibiotiques [céfuroxime (92,8%), céfotaxime (87,8%), ceftriaxone (68,9%) et ceftazidime
(5,6 %)] par rapport à ceux observés dans les pays développés mais se rapprochent des taux

Discussion
86
de résistance rapportés dans les pays en développement (Philippon et al., 2006 ; Gangoué
Piéboji, 2007; Xiao et al., 2011; Zhuo et al.,2013). Ce niveau de résistance élevé est une
conséquence de l’acquisition des facteurs de résistance aux antibiotiques qui est généralement
secondaire à l’utilisation abusive et incontrôlée de ces antibiotiques (Harbottle et al, 2006;
Boerlin et Reid-Smith, 2008 ; Muylaert et Mainil, 2012).
En Afrique, des bactéries résistantes au triméthoprime/sulfaméthoxazole ont été mises
en évidence dans des selles d’enfants qui n’ont jamais été exposés aux antibiotiques
(Huovinen et al., 1995), ce qui témoigne de l’émergence et de la dissémination des germes
résistants aux antibiotiques à cause généralement des mauvaises conditions de vie et
d’hygiène observées dans les pays en développement. Ces résultats sur la sensibilité aux
antibiotiques [céfuroxime (92,8% de résistance), céfotaxime (87,8% de résistance),
ceftriaxone (68,9% de résistance), ceftazidime (5,6 % de résistance) et céfépime (2,8% de
résistance)] confirment les résultats des travaux déjà rapportés sur la prévalence élevée de la
résistance des entérobactéries aux β-lactamines dans les pays en développement, qui est plus
élevée que celles des pays développés.
Dans cette étude, la majorité des souches productrices des BLSE (45%) ont été isolées
des patients hospitalisés dans les services de chirurgie. Dans ces services, les bactéries sont
soumises à une pression élevée des antibiotiques. Plusieurs de ces patients sont
particulièrement vulnérables aux infections à cause de leur immunodéficience ou ont des
portes d’accès facile aux bactéries dues aux traitements invasifs (Xiong et al., 2002). Les
BLSE sont généralement portées par de grands plasmides qui, en plus portent aussi les gènes
de résistances aux autres antibiotiques non-β-lactamines tels les aminoglycosides, le
triméthoprime-sulfaméthoxazole, les quinolones et le métronidazole. Il a été montré que
l’utilisation de ces antibiotiques contribue à la sélection des souches productrices de BLSE
(Paterson & Bonomo, 2005 ; Jacoby, 1997 ; Xiong et al., 2002). Les résultats de cette
sensibilité aux antibiotiques ont aussi montré que toutes les souches sont sensibles à

Discussion
87
l’imipénème (100% de sensibilité), et que la plupart est sensible à la ceftazidime (94,4 % de
sensibilité) et au céfépime (97,2% de sensibilité). Les carbapénèmes sont les antibiotiques de
premier choix chez un patient infecté par une souche productrice de BLSE car ils ne sont pas
hydrolysés par ces enzymes. Toutefois, les quinolones peuvent aussi être utilisées seules ou en
combinaison avec le métronidazole suivant le site de l’infection (Paterson & Bonomo, 2005).
Toutes nos souches traitées portent le gène CTX-M-15 qui coexiste le plus souvent
avec d’autres types de beta-lactamases. Cela pourrait s’expliquer par le fait que ce gène soit
porté par un grand plasmide, de taille approximative variant entre 50 et 90 Kb (Guangoué,
2007) et qui est facilement transferable entre bactéries. Ce plasmide porte en plus d’autres
gènes de résistances aux antibiotiques et de production d’autres types de β-lactamases (TEM-
1, SHV). La présence de tous ces gènes explique bien la multirésistance des souches qui les
portent.
Des 180 souches ayant présenté une résistance à au moins une céphalosporine de 3ème
génération, le test de double synergie n’a été positif que pour certaines souches. De plus, nous
avons noté une très grande variation des CMI des différentes souches vis-à-vis des
céphalosporines de 3ème génération. Le profil BLSE de nos isolats a été élargi et confirmé
par des méthodes cinétiques. En effet, toute image de synergie d’action n’implique pas
obligatoirement la production d’une BLSE par la souche. Une hyperproduction des β-
lactamases naturelles produirait aussi une image de synergie. Dans ce cas on parle de faux
positif (Yang et Guglielmo, 2007). En exemple nous avons une hyperproduction de la
pénicillinase naturelle dénommée CKO chez Citrobacter koseri qui produit une image de
synergie d’action. Ces « faux phénotypes de BLSE » sont également observés pour d’autres
espèces comme Klebsiella oxytoca (hyperproduction de la pénicillinase naturelle OXY) ou
Proteus vulgaris (hyperproduction de la céfuroximase naturelle CumA) (Yang et Guglielmo,
2007). Par ailleurs, l’image de synergie d’action entre l’acide clavulanique et une C3G
(céfotaxime) n’est pas observée sur boîte de pétri en présence de disques d’antibiotiques avec

Discussion
88
certaines souches. Un résultat qui n’atteste pas la présence de BLSE. Il se trouve que les
mêmes souches ont réagi positivement à la production de BLSE par approches cinétiques. Ce
qui prouve que les disques d’antibiotiques présentent souvent des insuffisancees pour la
détection des BLSE (Yang et Guglielmo, 2007). Cette conclusion montre que le test de double
synergie et autres tests physiologiques ont une sensibilité limitée dans la détection des
souches productrices de BLSE, fait qui avait déjà été signalé en 1995 par Livermore (1995) et
confirmé ensuite par des travaux de mutagenèse dirigée (Stapleton et al., 1999 ; Randegger et
al., 2000).
Un grand pourcentage (34,5%) des souches productrices de BLSE a été mis en
évidence dans les urines. Ce résultat corrobore ceux de Philippon et al. (1993), de Bermudes
et al. (1997) et de Gangoué (2007) qui rapportent que la majorité des souches BLSE
proviennent de l’urine. La vessie est un site de l’organisme humain où se trouvent des
concentrations élevées en certains antibiotiques (quinolones et β-lactamines) (NCCLS, 1999).
Les BLSE constituent aujourd’hui un problème thérapeutique mondial chez les patients
hospitalisés. La prévalence des Entérobactéries productrices de BLSE varie d’un pays à un
autre, d’une institution à l’autre (Bradford, 2001 ; Winokur et al., 2001 ; Paterson & Bonomo,
2005). La prévalence de 67,22% que nous avons observée dans cette étude est supérieure à
celle observée en Hollande (2,0%), Allemagne (2,6%), France (5,2%), Egypte (38,5%), Grèce
(27,4%), Afrique du Sud (15,0%) (Bouchillon et al., 2004) et au Cameroun (12%) . En
Hollande, la prévalence des souches de E. coli et K. pneumoniae productrice de BLSE est
<1% ; Cependant, plus de 40% de souches de K. pneumoniae sont résistantes à la ceftazidime
en France. Au Japon, le pourcentage de production de BLSE chez E. coli et K. pneumoniae
est <0,1 et 0,3% respectivement, comparé à 4,8% (Corée), 8,5% (Taiwan) et plus de 12,0% à
Hong Kong (Bradford, 2001). Les prévalences de production de BLSE chez Klebsiella et E.
coli que nous avons observé dans cette étude ont été de 69,38% et 65,88% respectivement.
Ces prévalences sont supérieures pour les espèces de Klebsiella et pour E. coli à celles

Discussion
89
rapportées par le programme de surveillance mondiale de la résistance aux antibiotiques
(SENTRY) qui sont respectivement de 45,0% et 8,5 % (Amérique latine), 25,0% et 7,9%
(Pacifique occidental), 23,0% et 5,3% (Europe) (Winokurr et al., 2001). La prévalence de
BLSE chez nos souches de Klebsiella spp.est supérieure à celle rapportée en Ethiopie (33,3%)
(Seid et Astrat, 2005), en Tanzanie (25%) par Blonblerg et al., (2005), au Cameroun(18,8%)
par Gangoué ( 2007) et au Maroc (7,5%) par Abouddihaj et al., (2012) . Cette prévalence reste
supérieure à celle (5,2%) rapportée dans plusieurs pays de l’Europe du Nord (France,
Allemagne, Hollande, Belgique, Suisse, Autriche) (Bouchillon et al., 2004). La prévalence de
BLSE dans nos souches de E.coli est inférieure à celle (76,0%) rapportée en Arabie Saoudite
(Bindayna et al., 2010).
La forte prévalence observée dans nos souches BLSE s’expliquerait par le fait que nous avons
utilisé des méthodes cinétiques qui nous ont permis d’identifier au maximum possible les
souches productrices de BLSE. Contrairement à notre étude, bon nombre d’études ce sont
limité au profil BLSE sur boîte de pétri par l’image de synergie d’action laissant ainsi
s’échapper les faux négatifs.
Le séquençage des produits PCR de 30 souches a permis de mettre en évidence la
production de huit types de BLSE et d’une carbapénèmase par les bactéries isolées au CHUP-
CDG. Il s’agit de SHV-12, 28, 32, 38, 76 et 99 et de CTX-M-15 qui est elle, produite par
toutes les 30 souches. Cette production de CTX-M-15 par toutes ces souches vient confirmer
les résultats de la cinétique qui montrent une hydrolyse du céfotaxime par tous les extraits
bruts de ces souches. La carbapénèmase est du type KPC-2 hébergée par un grand plasmide
dénommé pKP101-IT. La BLSE SHV-12 a été isolée pour la première fois en 1997 en Suisse
chez une souche de K. pneumoniae isolée d’un patient venant de l’Afrique du Nord (Nüesch-
Inderbinen et al., 1997). Depuis lors, cette BLSE a une répartition mondiale : Australie
(Howard et al., 2002), Canada (Mulvey et al., 2004) ; Chine (Chanawong et al., 2002),
Croatie (Bedenic et al., 2001), France (Weill et al., 2004), Grèce (Neonakis et al., 2003), Inde

Discussion
90
(Dhawan et al., 2003), Italie (Perilli et al., 2002), Japon (Yagi et al., 2000), Corée (Kim et al.,
1998), Espagne (Briñas et al., 2003), Taiwan (Yan et al, 2003), Thaillande (Chanawong et al.,
2001), Etats-Unis (Wong-Beringer et al., 2002), Afrique du Sud (Kruger et al., 2004),
Tanzanie (Blomberg et al., 2005), Cameroun (Gangoué Piéboji, 2000), RCA (Frank et al.,
2006), Tunisie (Ben-Hamouda et al., 2004), Sénégal (Cardinale et al., 2000) et Mali (Weill et
al., 2004). La BLSE SHV-12 apparaît alors être un gène particulièrement migratoire. Le fait
que l’on ait obtenu 100% d’homologie entre la SHV-12 du Burkina et la SHV-12 originale
(isolée pour la première fois en 1997 en Suisse chez une souche de K. pneumoniae isolée d’un
patient venant de l’Afrique du Nord (Nüesch-Inderbinen et al., 1997)) montrerait une origine
africaine de cette BLSE. Les BLSE de type TEM et particulièrement du type SHV sont très
rarement trouvées chez certains genres d’Entérobactéries tels Enterobacter, Serratia,
Citrobacter, chez lesquels la céphalosporinase chromosomique est prédominante (Jacoby,
1994; Livermore, 1995). Néanmoins, la BLSE SHV-4 a été mise en évidence chez Serratia
marcescens et Enterobacter cloacae (Philippon et al., 1989), la BLSE SHV-5 chez S.
marcescens (Gianneli et al., 1994), E. cloacae et Citrobacter freundii (Venezia et al., 1995),
la BLSE SHV-12 chez C. freundii et E. cloacae (Gangoué Piéboji, 2000).
La BLSE du type CTX-M a été rapportée pour la première fois en Europe Occidentale
en 1989 et c’était la CTX-M-1. Cette enzyme a été détectée en Allemagne, en Italie et en
France chez des espèces variées d’Entérobactéries (Bauernfeind et al., 1990, Bernard et al.,
1992 ; Pagani et al., 2003). La BLSE CTX-M-15 comme les BLSE CTX-M-16 et 19
confèrent une forte résistance à la ceftazidime que d’autres BLSE du type CTX-M (Poiret et
al., 2001 ; Bonnet, 2004). Toutefois, nous avons observé que les souches productrices de cette
BLSE ont été sensibles ou modérément résistantes à la ceftazidime. La CTX-M-15 qui diffère
de la CTX-M-1 par la mutation de cinq acides aminés a été identifiée pour la première fois en
Inde et au Japon et a été par la suite mise en évidence dans plusieurs autres pays (Canada,
Russie, Bulgarie, Pologne, France, Royaume uni, Turquie, Taiwan, Cameroun, Tunisie) (YU

Discussion
91
et al, 2003, Musthtaq et al., 2003 ; Latrigue et al.,2003 ; Bonnet, 2004 ; Gangoué, 2007 et
Samia et al., 2011). Nous avons décrit pour la première fois cette enzyme au Burkina. Cette
enzyme fait parti du groupe de CTX-M-1 qui possède 86% d’identité avec les acides aminés
de la β-lactamase chromosomique KLUC-1 de Kluyvera cryocrescens. D’autres types de
CTX-M ont été rapportés en Afrique, la CTX-M-2 (Afrique du Sud) la CTX-M-3 (RCA et
Cameroun) (Frank et al., 2006 ; Gangoué, 2007), la CTX-M-12 (Kenya) (Kariuki et al., 2001
; Paterson et al., 2005), la CTX-M-28 (Tunisie) (Achour, 2008). Les gènes blaCTX-M-15 que
nous avons décrit ont été soit associés aux gènes blaTEM-1 et/ou à un des autres types de
SHV comme c’est souvent le cas dans les autres études (Bonnet, 2004 ; Gangoué, 2007). Les
gènes blaCTX-M seraient portés par des plasmides différents qui faciliteraient leurs
disséminations. Des groupes géographiques et temporels des gènes blaCTX-M identiques et
portés par des plasmides apparemment différents ont été rapportés dans bon nombre d’études
dont Bonnet (2004), Gangoué (2007), Coque (2008), Carattoli (2009), Chouchani (2012) et
Zhuo (2013).
L’information sur les BLSEs dans l’espace et dans le temps serait un point de départ pour le
dévéloppement des stratégies de lutte contre les bactéries productrices de ces enzymes.

Conclusion et Perspectives
92
CONCLUSION ET PERSPECTIVES
Ce travail avait pour objectif de caractériser les BLSE produites par les bactéries à gram
négatif isolées des enfants malades au CHUP-CDG de Ouagadougou. Les résultats exposés
permettent de dégager un certains nombre d’éléments. Les bactéries isolées présentent des
niveaux de résistance élevés aux bêta-lactamines testées. Toutefois, le carbapénème
(imipénème) suivi de la céphalosporine de 3ème génération (ceftazidime) et de la
céphalosporine de 4ème génération (cefépime) sont les antibiotiques les plus actifs sur ces
bactéries.
La prévalence des bactéries productrices de BLSE est de 67,22% répartie comme suit :
Klebsiella spp(69,38%), E.coli(65,88%), Pseudomonas spp(58,33%), Enterobacter spp(75%),
Proteus spp(33,33%) et Salmonella spp(66,66 %). L’utilisation des méthodes cinétiques en
plus du test de double synergie permet une meilleure détection des BLSE. Les BLSE étudiées
proviennent des souches isolées en majorité des urines et des pus provenant des patients
hospitalisés en chirurgie (45,0%). Les BLSE que nous avons étudiées sont du groupe 2be de
Bush-Jacoby-Medeiros et de la classe A de Ambler car elles sont inhibées par l’acide
clavulanique. Les différents types de BLSE que nous avons mis en évidence sont SHV-12,
SHV-28, SHV-32, SHV-38, SHV-76, SHV-99, CTX-M-15 et une carbapénèmase de type
KPC-2 (hébergée par le plasmide pKPN101-IT). Ces différentes BLSE sont rapportées au
Burkina pour la première fois.
A la lumière de toutes ces informations, il apparaît que le péril BLSE existe au Centre
Hospitalier Universitaire Pédiatrique Charles De Gaulle (CHUP-CDG). Notre travail a donc
le mérite de sonner l’alerte sur la résistance bactérienne par production de BLSE. Toutefois, la
variabilité des problèmes posés par les bactéries productrices de BLSE d'un pays à un autre
implique la mise en place dans chaque pays un réseau national de surveillance. En effet, il est
primordial de bien cerner l'étendue et le développement de la résistance aux antibiotiques
dans le temps pour arriver à contrôler ce problème. Pour maîtriser la résistance aux

Conclusion et Perspectives
93
antibiotiques, il est clair qu'on ne peut plus compter actuellement sur le seul développement
de nouvelles molécules dont les perspectives restent très limitées, mais il est possible de
ralentir le développement des résistances par des stratégies de prévention. Ainsi, il est
actuellement impératif pour freiner le développement des résistances et maîtriser les coûts de
santé de changer nos comportements, et ne prescrire les antibiotiques que si nécessaire et de
façon raisonnée. Cela doit passer par une éducation des patients, la formation et la
sensibilisation des prescripteurs, l’application des procédures d’hygiène, et la mise en place
d’un réseau de surveillance des résistances bactériennes. Ce n’est qu’à travers les efforts
consentis par les différents acteurs que la recrudescence actuelle de la résistance bactérienne
par voie enzymatique peut être maîtrisée. Ainsi, il serait possible de maintenir ou prolonger
l’efficacité des antibiotiques existants.
A la lumière des présentes investigations il est hors de doute que les BLSE jouent un
rôle crucial dans la résistance bactérienne et il est à craindre que cela ne s’accentue en raison
du risque d’apparition de mutants souvent plus redoutables. Ces enzymes compromettent les
soins de santé.
C’est pourquoi les présentes investigations à caractère prospectif devront se
poursuivre. L’expérience acquise au cours de ce travail permettra de mieux cibler les
objectifs. Vu les résultats obtenus, il serait intéressant de connaître :
- les supports géniques des BLSE,
- la prévalence et les types de BLSE dans tous les hôpitaux de Ouagadougou voire du
Burkina Faso afin de mieux assurer la prise en charge des infections causées par des
souches productrices de ces enzymes.
- Les plantes médicinales constitueraient un axe de recherche des inhibiteurs de ces β-
lactamases.

Références Bibliographiques
94
REFFERENCES BIBLIOGRAPHIQUES
1. Abouddihaj B., Fatima E.O., Mustapha T., Ahmed R., Loubna J., Khalid Z. and
Mohammed T. 2012. Prevalence and genotypic analysis of plasmidmediated b-
lactamases among urinary Klebsiella pneumoniae isolates in Moroccan community.
The Journal of Antibiotics; 66: 11–16
2. Abraham, E.P., Chain, E., 1940. An enzyme from bacteria able to Destroy Penicillin.
Nature (London) 146: 837
3. Achour Ben N., P.S. Mercuri, P. Power, C. Belhadj, M. Ben Moussa , M. Galleni ,
O. Belhadj, 2008. Première détection de la CTX-M-28 dans un hôpital tunisien à partir
d’une souche de Klebsiella pneumoniae résistante au céfotaxime ; Patbio; 57: 343-348.
4. Achour Ben Nahed, Paola Sandra Mercuri, Chrifa Belhadj, Mohamed Ben
Moussa, Moreno Galleni and Omrane Belhadj. 2008. Cefotaxime and Ceftriaxon
resistant Klebsiella pneumoniae associated with SHV-11 hyperproduction. Annals of
Microbiology.58(3): 727-730
5. Achour Ben Nahed, Paola Sandra Mercuri, Mohamed Ben Moussa, Moreno
Galleni and Omrane Belhadj. 2009. Characterization of a novel Extended-Spectrum
TEM-Type beta-lactamase, TEM-164, in a clinical Strain of Klebsiella pneumoniae in
Tunisia. Microbial drug resistance 15: 195-199
6. Ahoyo, A.T., Baba-Moussa, L., Anago, A.E., Avogbe, P., Missihoun, T.D., Loko,
F., Prévost, G., Sanni, A. and Dramane, K., 2007. Incidence d'infections liées
à Escherichia coli producteur de bêta lactamase à spectre élargi au Centre hospitalier
départemental du Zou et Collines au Bénin . 37(11): 746-752
7. Al-Obeid, S., Gutmann L. and Williamson, R., 1990. Modification of penicillin-
binding proteins of penicillin-resistant mutants of different species of enterococci.
Journal of Antimicrobial Chemotherapy. 26 : 613-618

Références Bibliographiques
95
8. Ambler RP, Coulson AFW, Frere JM, Ghuysen JM, Joris B, Forsman M, et al.
1991. A standard numbering scheme for the class A b-lactamases. Biochem J; 276:
269–70.
9. Ambler, R. P. 1980. The structure of β-lactamases. Philos. Trans. R. Soc. Lond. (Biol.)
289: 321-331.
10. Ambler, R. P. et K. Scott, 1978. Partial aminoacid sequence of penicillinase coded by
Escherichia coli plasmid R6K. Proc.Natl. Acad. Sci. USA; 75: 3732-3736
11. Arlet G, Philippon A, 2003. Les nouvelles ß-lactamases à l’aube du troisième
millénaire. Rev Franç Lab; 352: 41-55
12. Bauerfeind A, Grimm H, Schweighart S., 1990. A new plasmidic cefotaximase in a
clinical isolate of Escherichia coli. Infection; 18: 294–8.
13. Bauernfeind, A., Casellas, JM., Goldberg, M., et al., 1992. A new plasmidic
cefotaximase from patients infected with Salmonella typhimurium. Infection; 20: 158-
163.
14. Bauernfeind, A., Grimm, H. & S. Schweighart. 1990. A new plasmidic cefotaximase
in clinical isolate of Escherichia coli. Infection 18: 294-298.
15. Bedenic, B., Randegger, C. C., Stobberingh, E., & H. Hächler. 2001. Molecular
epidemiology of extended-spectrum beta-lactamases from Klebsiella pneumonia strains
isolated in Zagreb, Croatia. Eur. J. Clin. Microbiol. Infect. Dis. 20: 505-508.
16. Ben Hassen, A., Fournie, G. r, Kechrid, A., Fendri ,C., Ben Redjeb, S. & A.
Philippon. 1990. Résistance enzymatique au céfotaxime chez cinquante-six souches de
Klebsiella spp., Escherichia coli et Salmonella spp. dans un hopital tunisien (1984–
1988). Pathol. Biol. 38:464–469.
17. Ben Redjeb, S., Fournie G., Mabilat, C., Ben Hassen ,A. & A. Philippon. 1998.
Two novel transferable extended-spectrum beta-lactamases from Klebsiella
pneumoniae in Tunisia. FEMS Microbiol. Lett., 55:33–38.

Références Bibliographiques
96
18. Ben-Hamouda, T., Foulon,T. & K. Ben-Mahrez. 2004. Involvement of SHV-12 and
SHV-2a encoding plasmids in outbreaks of extended-spectrum beta-
lactamaseproducing Klebsiella pneumoniae in a Tunisian neonatal ward. Microb. Drug
Resist.10:132–138.
19. Bermudes, H., Arpin, C., Inde, F. El Harrif, Z., Bebear, C., & C.Quentin. 1997.
Molecular epidemiology of an outbreak due to expanded spectrum betalactamase
producing Enterobacteriaceae in French hospital. Eur. J. Microbiol. Infect. Dis. 16:
523-259.
20. Bernard, H., Tancrede, C., Livrelli, V., Morand, A., Barthelemy, M. & R. Labia.
1992. A novel plasmid-mediated extended-spectrum β-lactamase not derived from
TEM- or SHV-type enzymes. J. Antimicrob. Agents Chemother. 29: 590-592.
21. Bibbal Delphine, 2008. Impact des beta-lactamines sur l’émergence d’entérobactéries
résistantes dans la flore digestive chez le porc : caractérisation et stratégie de
prévention. Thèse de doctorat, Université Toulouse III-Paul Sabatier, 152p.
22. Bidwell, J.L. and Reeves, D. S., 1980. Isoelectric focusing of β-lactamases. Journal of
Antimicrobial Chemotherapy, 6:793-798.
23. Bindayna, K., H.S. Khanfar, A.C. Senok and G.A. Botta. 2010. Predominance of
CTX-M genotype among Extended Spectrum beta-lactamase isolates in a tertiary
hospital in Saudi Arabia. Saudi. Med. J. 30: 859-863.
24. Blomberg, B., Juren, R., Manji, K.P., Tamin, B.S., Mwakagile, D.S.M., Urassa, W.
K., Fataki, M., Msangi, V., Tellevik, M.G., Maselle, S.Y. & Y. Langeland. 2005.
High rate of fatal cases of pediatric septicemia caused by Gram-negative bacteria with
extended-spectrum beta-lactamases in Dar Es Salaam, Tanzania. J. clin. Microb., 43 :
745-749.
25. Boerlin P., Reid-Smith R.J. 2008. Antimicrobial resistance: its emergence and
transmission. Anim. Health Res. Rev., 9, 115-126.

Références Bibliographiques
97
26. Bonnet R., 2004. Growing group of extended-spectrum β-lactamases: the CTX-M
enzymes. AntimicrobAgents Chemother; 48: 1-14
27. Bouchillon, S.K., Johnson, B.M., Hobana, D.J., Johnson, J.L., Dowzicky, M.J.,
Wu, D.H., Visallic, M.A. & P.A. Bradford. 2004. Determining incidence of extended
spectrum β -lactamase producing Enterobacteriaceae, vancomycin-resistant
Enterococcus faecium and methicillin-resistant Staphylococcus aureus in 38 centres
from 17 countries: the PEARLS study 2001–2002. J. Antimicrob. Agents, 24: 119-124.
28. Bradford PA., 2001. Extended-spectrum β-lactamases in the 21st century:
characterization, epidemiology, and detection of this important resistance threat. Clin
Microbiol Rev; 14: 933-951
29. Bradford, P. A. 1999. Automated thermal cycling is superior to traditional methods
for nucleotide sequencing of bla (SHV) genes. Antimicrob. Agents Chemother. 43:
2960-2963.
30. Brinas, L., Moreno, M. A., Teshager, T., Zarazaga, M., Sáenz, Y., Porrero, C.,
Dominguez, L. & C. Torres. 2003. Beta-lactamase characterization in Escherichia
coli isolates with diminished susceptibility or resistance to extended-spectrum
cephalosporins recovered from sick animals in Spain. Microb. Drug Resist. 9: 201-09.
31. Brown, D. F., Andrews, J., King, A., & A. P. MacGowan. 2000. Detection of
extended-spectrum beta-lactamases with E-test and double-disc potentiation methods.
J. Antimicrob. Chemother. 46:327–328.
32. Bush,K., Jacob, G.A., Medeiros , A.A., 1995. A functional classificcation scheme for
β-lactamases and its correlation with molecular structure. Antimicrob Agents
Chemother. 39:1211-1233
33. Carattoli Alessandra. 2009. Resistance Plasmids Families in Enterobacteriacae.
Microbial agent and Chemotherapy; 53: 2227-2238.

Références Bibliographiques
98
34. Cardinale, E., P. Colbachini, J. D. Perrier-Gros-Claude, A. Gassama, & A.
Aidara-Kane. 2000. Dual emergence in food and humans of a novel multiresistant
serotype of Salmonella in Senegal: Salmonella enterica susbp. enterica serotype 35: C:
12. J. Clin. Microbiol. 39: 2373-2374.
35. Cartier F., Michelet C., Camus C. 1988. Les inhibiteurs de bêtalactamases :
applications cliniques. Lettre de l’infect., 18, 687-697.
36. Chanawong, A., M'Zali, F. H., Heritage, J., Lulitanond, A. & P. M. Hawkey. 2001.
SHV-12, SHV-5, SHV-2a and VEB-1 extended-spectrum beta-lactamases in Gram
negative bacteria isolated in a university hospital in Thailand. J.
Antimicrob.Chemother. 48: 839-852.
37. Chanawong, A., M'Zali, F. H., Heritage, J., Xiong, J. H. & P. M. Hawkey. 2002.
Three cefotaximases, CTX-M-9, CTX-M-13, and CTX-M-14,
amongEnterobacteriaceae in the People's Republic of China. Antimicrob. Agents
Chemother. 46: 630-637.
38. Chouchani C, El Salabi A, Marrakchi R, Abouelkacem and Walsh T. R, 2012.
Occurrence of clinical isolates of Klebsiella pneumonia harboring chromosomally
mediated CTX-M-15 beta-lactamase in a Tunisian hospital; Can. J. Microbiol; 58:
1099-1103.
39. CLSI. 2007. Development of In Vitro Susceptibility Testing Criteria and Quality
Control Parameters for Veterinary Antimicrobial Agents; Approved Guideline-Third
Edition. M37-A3Vol. 0 No. 0 Replaces M37-A2; 22 : 7.
40. Coque TM, Novais A, Carattoli A, Poirel L, Pitout J, Peixe L, et al., 2008.
Dissemination of clonally related Escherichia coli strains expressing extended-
spectrum blactamase CTX-M-15. Emerg Infect Dis; 14:195–200.

Références Bibliographiques
99
41. Cormican, M. G., Marshall, S. A. & R. N. Jones. 1996. Detection of
extendedspectrum β-lactamase (ESBL)-producing strains by the Etest ESBL screen. J.
Clin. Microbiol. 34:1880–1884.
42. Cosgrove, S. E., Kaye, K. S., Eliopoulous, G. M. & Y. Carmeli. 2002. Health and
economic outcomes of the emergence of third-generation cephalosporin resistance in
Enterobacter species. Arch. Intern. Med. 162: 185–190.
43. Datta, N., Kontomichalou, P., 1965. Penicillinase synthesis controlled by infectious R
factors in Enterobacteriaceae. Nature. 208: 239-244
44. Derby, P.; Berens, J., 1997. ESBL-producing organisms and cephalosporins. samj.
South African medical journal ISSN : 0256-9574
45. Dhawan, B., Bonnet, R., Shukla, N. K., Mathur, P., Das, B. K. & A. Kapil. 2003.
Infection with an extended-spectrum beta-lactamase-producing strain of Serratia
marcescens following tongue reconstruction. J. Clin. Microbiol. 41: 2233-2234.
46. Doi, Y., Wachino, J-I., Ishiguro, M., Kurokawa, H., Yamane, K., Shibata, N.,
Shibayama K., Yokoyama, K., Kato, H., Yagi, T. & Arakawa, Y. 2004.
Inhibitorsensitive AmpC β-lactamase variant produced by an Escherichia coli clinical
isolate resistant to oxyiminocephalosporins and cephamycins. Antimicrob. Agents
Chemother., 48: 2652-2658.
47. Duval, V., Maiga, I., Maiga, A., Guillard, T., Brasme, L., Forte, D., Madoux, J.,
Vernet-Garnier, V. and Christophe DE Champs. 2009. High Prevalence of CTX-M-
Type β-Lactamases among Clinical Isolates of Enterobacteriaceae in Bamako, Mali.
Antimicrob. Agents Chemother. 53: 4957-4958
48. Farmer, J. J., III 1999. Enterobacteriaceae: introduction and identification, 442–458.
In

Références Bibliographiques
100
49. Faure Stéphanie. 2009. Transfert d’un gène de résistance aux β-lactamines blaCTX-
M-9 entre Salmonella et les entérobactéries de la flore intestinale humaine : impact
d’une antibiothérapie. Thèse de doctorat, Université de rennes1, 190p.
50. Fleming, A., 1929. On the antibacterial action of cultures of a penicillium, with special
reference to their use in the isolation of b. influenzae. Brit. J. Exp. Path. 10: 226-236
51. Fontana, R., CaneparI, P., Lleò, M. M. and Satta, G., 1983. Mechanisms of
resistance of Enterococci to beta-lactam antibiotics. European journal of clinical
microbiology and infectious deaseases. 9:103-105
52. Frank, T., Arlet, G., Gautier, V., Talarmin, A. & R. Bercion. 2006. Extended
Spectrum β-Lactamase–producing Enterobacteriaceae, Central African Republic.
Emerging Infect. Dis. 12: 863-865
53. Frank, T., Arlet, G., Gautier, V., Talarmin, A. and Bercion, R., 2006. Extended-
spectrum β-Lactamase–producing Enterobacteriaceae. Central African Republic. 12,
(5): 211-217.
54. Frère, J.-M. & B. Joris. 1985. Penicillin-sensitive enzymes in peptidoglycan
synthesis, CRC Crit. Rev. Microbiol., 11: 299-396.
55. Frère, J.-M., Ghuysen, J.-M. & M.Iwatsubo. 1975. Kinetics of the interaction
between the exocellular DD-carboxypeptidase-transpeptidase from Streptomyces R39,
Eur. J. Biochem. 57: 343- 351.
56. Fuad, N., Frère, J.-M., Ghuysen, J.-M., Duez, C. & M., Iwatsubo. 1976. Mode of
interaction between beta-lactam antibiotics and the exocellular DD-
carboxypeptidasetranspeptidase from Streptomyces R39 and beta-lactam antibiotics. A
choice of models, Biochem. J. 155 : 623-629
57. Galleni, M., Lamotte-Brasseur, J., Raquet, X., Dubus, A., Monnaie, D., Knox, J.R.
& J.- M.Frère. 1995. The enigmatic catalytic mechanism of active-site serine β-
lactamases, Biochem. Pharmacol. 49: 1171-1178.

Références Bibliographiques
101
58. Galleni, M., Lamotte-Brasseur, J., Rossolini, G.M., Spencer, J., Dideberg, O., J.M.
Frère & the metallo- β-lactamase working group. 2001. Standard numbering
scheme for class B β-lactamases. Antimicrob. Agents Chemother., 45 : 660-663.
59. Gangoué Piéboji J. 2007. Caractérisation des beta-lactamases et leur inhibition par les
extraits de plantes médicinales. Thèse de doctorat, Centre d’Ingénierie des protéines,
127 p.
60. Gangoué Piéboji J. 2000. Résistance des bacilles à Gram négatif aux antibiotiques:
prévalence et caractérisation des β -lactamases à spectre élargi à l’Hôpital Central de
Yaoundé. Thèse de Doctorat de 3e cycle, Université de Yaoundé I, 210 p.
61. Gangoué-Piéboji, J., Bedenic, B., Koulla-Shiro S., Randegger, C., Adiogo, D.,
Ngassam, P., Ndumbé, P., & H. Hächler. 2005. Extebnded-Spectrum-β-
lactamaseproducing Enterobacteriaceae in Yaounde, Cameroon. J. Clin. Microb., 43:
3273-3277.
62. gangoue-Pieboji, J., Eze N., Djintchui A., Ngameni B., Tsabang, N., Pegnyemb, D.
E., Biyiti, L., Ngassam, P. , Koulla-Shiro, S. and Galleni, M., 2009. The in-vitro
antimicrobial activity of some traditionally used medicinal plants against beta-lactam-
resistant bacteria., Institute of Medical Research and Medicinal Plant Studies, PO Box
8404, 87: 865-879
63. Gangoue-Pieboji, J., Miriagou, V., Vourli, S., Tzelepi, E., Ngassam, P., and
Tzouvelekis, L. S., 2005. Emergence of CTX-M-15-Producing Enterobacteria in
Cameroon and Characterization of a blaCTX-M-15-Carrying Element. Journal List
Antimicrob Agents Chemother. 49: 510-517
64. Ghuysen, J.-M. 1991. Serine beta-lactamases and penicillin-binding proteins. Annu.
Rev. Microbiol. 45: 37-67.
65. Ghuysen, J.-M. 1994. Molecular structures of penicillin-binding proteins and β-
lactamases, Trends Microbiol. 2: 372-380.

Références Bibliographiques
102
66. Ghuysen, J.-M., Frère, J.-M., Leyh-Bouille, M., Nguyen-Distèche, M. & J.
Coyette. 1986. Active-siteserine D-alanyl-D-alanine-cleaving-peptidase-catalysed
acyl-transfer reactions, Biochem. J. 235: 159-165.
67. Gianneli, D., Tzelepi, E., Tzouvelekis, L. S. Mentis, A. F. & C. Nikolopoulou. 1994.
Dissemination of cephalosporin-resistant Serratia marcescens strains producing a
plasmidic SHV type β-lactamase in Greek hospitals. Eur. J. Clin. Microbiol. Infect.
Dis. 13 : 764-767
68. Gilliane Guillaume, Marc Vanhove, Josette Lamotte-Brasseur, Philippe Leden,
Marc Jamin, Bernard Joris and Jean Marie frère, 1996. Site-directed Mutagenesis
of Glutamate 166 in two ß-lactamases. Journal of biological chemistry, 272: 5438-
5444
69. Glupczynski ,Y., Lambert, D., Spinewine, A., Tulkens, P. M., Van Bambeke, F.,
Van Laethem, Y., Van Landuyt ,H., 2002. Pharmacologie et pharmacothérapie des
anti-infectieux. Antimicrob Agents Chemoter. 42 : 13-26.
70. Gniadkowski M. 2001. Evolution and epidemiology of Extended Spectrum-lactamases
and ESBL- producing microorganisms. Clin Microbiol infect; 7: 597–608.
71. Gutmann L. 1989. Spectre des inhibiteurs de bêtalactamases. Med. Mal. Inf., Hors
série, Mai, 52-56.
72. Hakenbeck, R. & J. Coyette. 1998. Resistant penicillin-binding proteins. Cell. Mol.
Life Sci. 54: 332-340.
73. Hammond, D.S. Schooneveldt, J.M., Nimmo, G.R., Huygens, F. and P.M. Giffard.
2005. baSHV genes in Klebsiella pneumoniae: different allele distributions are
associated with different paromoters within individual isolates. Antimicrob. Agents
Chemother., 49: 256-263.
74. Harbottle H., Thakur S., Zhao S., White D.G. 2006. Genetics of antimicrobial
resistance. Anim. Biotechnol., 17: 111-124.

Références Bibliographiques
103
75. Ho, P. L., Tsang, D. N. C., Que, T. L., Ho, M. & K. Y. Yuen. 2000. Comparison of
screening methods for detection of extended-spectrum β -lactamases and their
prevalence among Escherichia coli and Klebsiella species in Hong Kong. APMIS, 108:
237–240.
76. Howard, C., Daal, A., Kelly, G., Schooneveld, J., Nimmo, G. & M. P. Giffard.
2002. Identification and minisequencing-based discrimination of SHV β-lactamases in
nosocomial infection-associated Klebsiella pneumoniae in Brisbane, Australia.
Antimicrob. Agents Chemother. 46: 659-664
77. Huovinen P. 1997. Increases in rates of resistance to trimethoprim. Clin. Infect. Dis.
24:S63-S66.
78. Huovinen, P., Sundström, L., Swedberg, G. & O.Sköd. 1995. Trimethoprim and
sulfonamide resistance. Antimicrob Agents Chemother, 39: 279-89.
79. Huovinen, P., Huovinen, S., Jacoby, G. A. 1988. Sequence of PSE-2 β-lactamase.
Antimicrob AgentsChemoter; 32:134-136
80. Jacoby GA, Munoz-price, L S., 2005. The new β-lactamases. N Engl J Med 352: 380-
391
81. Jacoby, G. A. & P. Han. 1996. Detection of extended-spectrum β-lactamases in
clinical isolates of Klebsiella pneumoniae and Escherichia coli. J. Clin. Microbiol. 34:
908-911.
82. Jacoby, G. A. 1994. The genetics of extended-spectrum beta-lactamases. Eur. J.Clin.
Microbiol. Infect. Dis. 13: 2-11
83. Jacoby, G. A. 1997. Extended-spectrum β-lactamases and other enzymes providing
resistance to oxyimino-β-lactams. Infect. Dis. Clin. North America. 11 : 875-887
84. Jarlier, V., Nicolas, M. -H., Fournier, G. & A. Philippon. 1988. Extended broad -
spectrum β-lactamases confering transferable resistance to newer β-lactam agents in

Références Bibliographiques
104
Enterobacteriaceae: hospital prevalence and susceptibility patterns. Rev. Infect. Dis.
10: 867-878.
85. Jaurin, B., Grundstrom, T. 1982. AmpC cephalosporinase of Escherichia coli K-12
has a different evolutionary origin from that of β-lactamases of penicillinase type.
Proc. Natl. Acad.Sci.USA; 78: 4897-4901
86. Jones, R. N., Pfaller M. A., Doern G. V., Erwin, M. E., & R. J. Hollis. 1998.
Antimicrobial activity and spectrum investigation of eight broad spectrum betalactam
drugs : a surveillance trial in 102 medical centres in the United states. Diagn.
Microbiol. Infect. Dis. 30: 215-228.
87. Kariuki, S., Corkill, J. E., Revathi, G., Musoke, R. & C. A. Hart. 2001. Molecular
characterization of a novel plasmid-encoded cefotaximase (CTXM-12) found in
clinical Klebsiella pneumoniae isolates from Kenya. Antimicrob. Agents Chemother.
45:2141–2143.
88. Karou, S. D., Ilboudo, D. P. , Nadembega, W. M.C. , Ameyapoh, Y., Ouermi, D.,
Pignatelli, S., Pietra, V., Traore, A. S., Comla DE Souza, and Simpore, J., 2009.
Antibiotic resistance in urinary tract bacteria in Ouagadougou. Pakistan Journal of
biological Science ISSN1028-8880. 12(9): 712-716
89. Kim, J., Kwon Y., Pai, H., Kim, J. W. & D. T. Cho. 1998. Survey of Klebsiella
pneumoniae strains producing extended-spectrum beta-lactamases: prevalence of SHV-
12 and SHV-2a in Korea. J. clin. Microbiol. 36: 1446-1449.
90. Kliebe, C., B. A. Nies, J. F. & R.M Meyer,. Tolxdorff-Neutzling, and B.
wiedemann. 1985. Evolution of plasmid-coded resistance to broad-spectrum
cephalosporins. Antimicrob. Agents Chemother. 28: 302-307.
91. Knothe, H. P. Shah, V., Kreméry, M. & S. Mitsuhashi. 1983. Transferable
resistance to cefotaxime, cefoxitin, cefamandole and cefuroxime in clinical isolates of
Klebsiella pneumoniae and Serratia marcescens. Infection 11: 315-317.

Références Bibliographiques
105
92. Kolar, M., Urbanek, K. & T.Latal. 2001. Antibiotic selective pressure and
development of bacterial resistance. Int J Antimicrob Agents, 17: 357-63.
93. Koulla-Shiro, S., Boye, C.S., Dosso, M. & the members of the Palm Project. 1998.
Surveillance of antimicrobial susceptibility of Gram negative pathogens responsible for
nosocomial infections in West Africa (Abstract number E-95). San Diego, California:
42nd ICAAC; p. 197.
94. Kruger, T., Szabo, D., Keddy, K.H., Deeley, K., Marsh, J.W., Hujer A.M.,
Bonomo, R.A. &D.L. Paterson. 2004. Infections with Nontyphoidal Salmonella
Species Producing TEM-63 or a Novel TEM Enzyme, TEM-131, in South Africa.
Antimicrob. Agents Chemother., 48: 4263-4270
95. Kunin, C. M., K. S. Johansen, A. M. Worning, & F. D. Daschner. 1990. Report of a
symposium on use and abuse of antibiotics worldwide. Rev. Infect. Dis. 12: 12- 19.
96. Lahlou Amine I., Chegri M., L’kassmi H., 2009. Épidémiologie et résistance aux
antibiotiques des entérobactéries isolées d’infections urinaires à l’hôpital militaire
Moulay-Ismail de Meknès. Issue 2.11, pp. 90-96
97. Lakaye, B., Dubus, A., Lepage, S., Groslambert, S. & J. M. Frère. 1999. When
drug inactivation renders the target irrelevant to antibiotic resistance: a case story with
β-lactams. Mol. Microbiol. 31: 89–101.
98. Lamikamra, A & R.B. Ndep. 1989. Trimethoprim resistance in urinary tract
pathogens in two Nigerian hospitals. J Antimicrob Chemother., 23:151-154.
99. Larabi, K., Masmoudi, A. and Fendri, C., 2003. Étude bactériologique et phénotypes
de résistance des germes responsables d’infections urinaires dans un CHU de Tunis : à
propos de 1930 casBacteriological and susceptibility study of 1.930 strains isolated
from UTIs in a Tunis university hospital. Issue 7. 33: 348-352

Références Bibliographiques
106
100. Latrigue, M.F., Poiret, L., Heritier, C., Tolűn, V. & P. Nordmann. 2003. First
description of CTX-M-15-producing Klebsiella pneumoniae in Turkey. J. Antimicrob.
Chemother. 52: 315-316.
101. Lesch ,C.A., Itokazu, G.S., Danziger, L.H. & R.A. Weinstein. 2001. Multi-hospital
analysis of antimicrobial usage and resistance trends. Diagn. Microbiol. Infect. Dis.,
41:149-54.
102. Linscott, A.J., & W.J. Brown. 2005. Evaluation of four commercially available
extende-spectrum β-lactamase phenotypic confirmation tests. J. Clinic. Microbiol.,
43:1081-1085.
103. Livermore, D., 1995. Beta-lactamases in laboratory and clinical resistance. Clin
Microbiol Rev; 8: 557-584.
104. Livermore, D. M. 1995. β-lactamase mediated resistance: past, present and future. J.
Infect. Dis. Soc. 6: 75-83.
105. Mahmal, L., Loukili, A., Harif, M., Quessar, A., Benbachir, M., Benchekroun,
S., 2004. Les germes producteurs de beta lactamases a spectre etendu: epidemie dans
un service d’Hemato-Oncologie. Tunis Med, ISSN 0041-4131 82(11): 1006-1011.
106. Matagne, A., Lamotte-Brasseur J. & J-M. Frère. 1998. Catalytic properties of class
A β-lactamases: efficiency and diversity. Biochem. J., 330: 581-598.
107. Medeiros AA., 1984. Betalactamase. Br. Bull.40: 18-27
108. Messai, Y., Iabadene, H., Benhassine, T., Alouache, S., Tazir, M., Gautier, V.,
Arlet, G. bakour, R., 2008. Prevalence and characterization of extended-spectrum β-
lactamases in Klebsiella pneumoniae in Algiers hospitals (Algeria). Pathologie
Biologie, Issue 5, 56: 319-325.
109. Mètuor Dabiré A., Zongo K. J., Zeba B., Moussawi J., Baucher M. and El Jaziri
M., 2013. Resistances to the oxyimino-cephalosporins by ctx-m-15 producing
klebsiella isolated from the urines samples of patients in the university hospital

Références Bibliographiques
107
complex paediatric charles de gaulle (chup-cdg) of ouagadougou in burkina faso.
Journal of Asian Scientific Research, 3(9): 882-890.
110. Mhand, R. A., Soukri, A., Amarouch, A., El Mdaghri, N., Benbachir, M., 1999.
Typage de souches de Salmonella Typhimurium productrices de bêta-lactamase à
spectre élargi isolées dans une unité pediatrique. Cahier de santé ; 9 : 341-400
111. Monnet, D.L., Archibald, L.K., Phillips, L., Tenover, F.C., McGowan, J.E. & R.P.
Gaynes. 1998. Antimicrobial use and resistance in eight US hospitals: complexities of
analysis and modeling. Infect Control Hosp Epidemiol;19: 388-394.
112. Muller, A MC Geer, D Reynolds, Sylveman, MA Green, M Culf., 2000. Eclosions
attribuables à Escherichia coli multirésistant aux antibiotique dans les établissements
de soins de longue durée des régions de Durham,York, Toronto et Ontario. Relevé des
maladies transmissibles au Canada .ISSN 1188-4169, 28 : 113- 130
113. Mulvey, R.M., Bryce, E., Boyd, D., Ofner-Agostini, M., Christianson, S., Simor,
E.S., Paton, S., & the Canadian Hospital Epidemiology Committee of The
Canadian Nosocomial Infection Surveillance Program, Health Canada. 2004.
Ambler Class A Extended-Spectrum Beta-Lactamase-Producing Escherichia coli and
Klebsiella spp. In Canadian Hospitals. Antimicrob Agents Chemoter., 48: 1204-1214.
114. Mushtaq, S., Woodford, N., Potzand, N. & D. M. Livermore. 2003. Detection of
CTX-M-15 extended-spectrum β-lactamase in the United Kingdom. J.
Antimicrob.Chemother. 52:528-529.
115. Muylaert A., Mainil J. G. 2012. Résistances bactériennes aux
antibiotiques : les mécanismes et leur « contagiosité » Ann. Méd.
Vét., 156, 109- 123.
116. N’guessan, J D., Dinzedi, MR. , Guessennd, N. , Coulibaly, A. , Dosso, M. ,
Djaman, AJ. , Guede-Guina, F., 2007. Antibacterial activity of the aqueous extract of

Références Bibliographiques
108
Thonningia sanguinea against Extended-Spectrum-β-Lactamases (ESBL) producing
Escherichia coli and Klebsiella pneumoniae strains. Tropical Journal of
Pharmaceutical Research, 6(3): 779-783.
117. Naas T, Lezzar A, Bentchouala C, et al. 2005. Multidrug-resistant Salmonella
enterica serotype Senftenberg isolates producing CTX-M (-lactamases from
Constantine, Algeria. J Antimicrob Chemother; 56: 439–40.
118. Naas T, Poirel L, Nordmann P., 2008. Minor extended-spectrum β-lactamases. Clin
Microbiol Infect;14: 42-52.
119. Naas, T., Nordmann, P., 1999. OXA type β-lactamases. Curr. Pharm. Design .5:
865-879
120. Nanninga, N. 1991. Cell division and peptidoglycan assembly in Escherichia coli.
Mol. Microbiol. 5:791–795.
121. Nanninga, N., 1998. Morphogenesis of Escherichia coli Microbiol Mol Biol Rev, 62,
110-129.
122. National Committee for Clinical Laboratory Standards. 1999. Performance
standards for antimicrobial susceptibility testing; ninth informational supplement.
Approved standard M100-S9. NCCLS, Wayne, Pa.
123. Neonakis, I.K., Scoulica, E. V., Dimitrio, S. K., Gikas, A. I. & Y. J. Tselentis.
2003. Molecular epidemiology of extended-spectrum beta-lactamases produced by
clinicalisolates in a university hospital in Greece: detection of SHV-5 in Pseudomonas
aeruginosa and prevalence of SHV-12. Microb. Drug Resist. 9: 161-165.
124. Nguyen, M. C. P, Woerther, P-L., Bouvet, M., Andremont, A., Leclercq, R., and
Canu, A., 2009. Escherichia coli as Reservoir for Macrolide Resistance Genes.
Emerging Infectious Diseases • www.cdc.gov/eid . 15(10): 239-244
125. Nikaido, H. 1998. Multiple antibiotic resistance and efflux. Curr. Opin. Microbiol.
1:516–523.

Références Bibliographiques
109
126. Normark, B. H. & S. Normark. 2002. Evolution and spread of antibiotic resistance. J
Intern. Med., 252: 91-106.
127. Nüesch-Inderbinen, M. T., Kayser, F. H. & H. Hächler. 1997. Survey and
molecular genetics of SHV β-lactamases en enterobacteriaceae in Switzerland: two
novel enzymes, SHV-11 and SHV-12. Antimicrob. Agents. Chemother. 41: 943-949.
128. Okeke, I.N., Lamikanra, A, & R. Edelman. 1999. Socio-economic and behavioral
factors leading to acquired bacterial resistance to antibiotics in developing countries.
Emerg Infect Dis.5:18-27.
129. P. R. Murray, E. J. Baron, M. A. Pfaller, F. C. Tenover, and R. H. Yolken (ed.),
Manual of clinical microbiology, 7th ed. American Society for
Microbiology,Washington, D.C.
130. Pagani L, Dell’Amico E, Migliavacca R et al., 2003. Multiple CTX-M-type extended
spectrum beta-lactamases in nosocomial isolates of Enterobacteriaceae from a hospital
in northern Italy. J Clin Microbiol; 41: 4264-9
131. Pai, H., Lyu, S., Lee, J. H., Kim, J., Kwon, Y., Kim, J.-W., & K. W. Choe. 1999.
Survey of extended-spectrum β -lactamases in clinical isolates of Escherichia coli and
Klebsiella pneumoniae: prevalence of TEM-52 in Korea. J. Clin. Microbiol. 37:1758–
1763.
132. Paterson, D.L. & R.A. Bonomo. 2005. Extended-spectrum β-lactamases: a clinical
update. Clinic. Microbiol. Rev., 18: 657-686.
133. Perilli, M., Dell'Amico, E., Segatore, B., de Massis, M. R., Bianchi, C., Luzzaro,
F., Rossolini, G. M., Toniolo, A., Nicolletti, G. & G. Amicosante. 2002. Molecular
characterization of extended-spectrum beta-lactamases produced by nosocomial
isolates of Enterobacteriaceae from an Italian nationwide survey. J. Clin. Microbiol.
40: 611-614.

Références Bibliographiques
110
134. Pfaller, M.A., Jones, R.N., Doern, G.V. & K. Kugler. 1998. Bacterial pathogens
isolated from patients with blood stream infection: frequencies of occurrence and
antimicrobial susceptibility patterns from the SENTRY antimicrobial surveillance
program (United States and Canada, 1997). Antimicrob Agents Chemother., 42:1762-
1770.
135. Philippon A, Arlet G, Lagrange PH. 1994. Origin and impact of plasmid-mediated
of extended-spectrum-beta-lactamases. Eur J Clin Microbiol Infect Dis; 13: 17–29.
136. Philippon A, Arlet G. 2006. Beta-lactamases of Gram negative bacteria: neverending
clockwork! Ann Biol Clin; 64:37–51.
137. Philippon, A., Arlet G. & P. H. Lagrange. 1993. β-lactamses à Spectre élargi. Rev.
Prat. 43: 2387-2395.
138. Philippon, A., Arlet, G. & G.A. Jacoby. 2002. Plasmid-determined AmpC-type β-
lactamases. Antimicrob. Agents Chemother., 46: 1-11.
139. Philippon, A., Labia, R. & J. A. Jacoby. 1989. Extended-spectrum β-lactamases.
Antimicrob. Agents Chemother. 33: 1131-1136.
140. Pitout, J. D. D., Thomson, K. S., Hanson, N. D., Ehrhardt, A. F., Coudron, P., &
C.C. Sanders. 1998. Plasmid mediated resistance to expanded-spectrum
cephalosporins among Enterobacter aerogenes strains. Antimicrob. Agents Chemother.
42: 596-600.
141. Pitout, J.D.D., Reisbig, M.D., Venter E.C., Church, D.L. & N.D. Hanson. 2003.
Modification of the Double-disk test for detection of Enterobacteriaceae produicing
extended-spectrum and AmpC β-lactamases. J. Clinic. Microbiol., 41: 3933-3935.
142. Poirel, L. and P Nordmann. 2005. Nomenclature of GES-type extended-spectrum β-
lactamases. Antimicrob. Agents Chemother., 49: 2148-2150.

Références Bibliographiques
111
143. Poirel, L., L. Villa, A. Bertini, J. D. Pitout, P. Nordmann, and A. Carattoli. 2007.
Expanded-spectrum beta-lactamase and plasmid-mediated quinolone resistance. Emerg
Infect Dis 13: 803-5.
144. Poiret, L., Naas, T., Le Thomas, I., Karim, A., Bingen, E., & P. Nordman. 2001.
CTX-M-type extended-spectrum β-lactamase that hydrolyses ceftazidime through
asingle amino acid substitution in the Omega loop. Antimicrob. Agents Chemother.45:
3355-3361.
145. Randegger, C. C., Keller, A., Irla, M., Wada, A.&H. Hächler. 2000. Contribution
of natural amino acid substitutions in SHV extended-spectrum β-lactamases to
resistance against various β-lactams. Antimicrob. Agents Chemother. 44: 2759-2763.
146. Rossolini GM, D’andrea MM, Mugnaioli, C., 2008. The spread of CTX-M-type
extended-spectrum β-lactamases. Clin Microbiol Infect;14: 33-4115
147. Ruppé E., Woerther, P-L., Diop, A., Sene, A-M., Costa, A. D., Arlet, G.,
Andremont, A. and Rouveix, B., 2009. Carriage of CTX-M-15-Producing
Escherichia coli Isolates among Children Living in a Remote Village in Senegal.
Antimicrobial Agents and Chemotherapy, 53(7): 3135-3137
148. Samia Réjiba, Paola Sandra Mercuri, Pablo power and Amel Kechrid, 2011.
Emergence and Dominance of CTX-M-15 Extended Spectrum Beta-Lactamase Among
Escherichia coli Isolats from Children; MICROBIAL DRUG RESISTANCE; 00: 1-6
149. Sanders, C. C. & W. E. Jr Sanders. 1992. β-lactam resistance in gram negative
bacteria : global trends and clinical impact. Clin. Infect. Dis. 15: 824-839.
150. Sanders, C. C., Barry, A. L., Washington, J. A., Shubert, C., Moland, E. S.,
Traczewski, M. M., Knapp, C. & R. Mulder. 1996. Detection of extended spectrum
beta-lactamase-producing members of the family Enterobacteriaceae with Vitek ESBL
test. J. Clin. Microbiol. 34: 2997–3001.

Références Bibliographiques
112
151. Seid, J. & D. Asrat. 2005. Occurrence of extended spectrum beta-lactamase enzymes
in clinical isolates of Klebsiella species from Harar region, eastern Ethiopia. Acta Trop.
95: 143-148.
152. Simpson, I. N., James, M., 1982. Comparaison of routine techniques for cell
breakage and release of β-lactamase activity. Journal of Antimicrobial chemotherapy.
9: 119-123
153. Sirot, D. 1995. Extended-spectrum plasmid-mediated beta-lactamases. J. Antimicrob.
Chemother. 36: 19-34.
154. Sougakoff W., S. Goussarg, G. Gerbaud and P. Courvalin. 1988. The TM-3 β-
lactamase, which hydrolyses broad-spectrum cephalosporins is derived from the TEM-
2 penicillinase by two aminoacid substitutions.FEMS Microbiol. Lett. 56: 3343-348
155. Spratt, B.G.& A.B Pardee. 1975. Penicillin-binding proteins and cell-shape in
Escherichia coli. Nature, 254: 516-517.
156. Stapleton, P. D., Shannon, K. P. & G. L. French. 1999. Construction and
characterization of mutants of the TEM-1 β-lactamase containing amino acid
substitutions associated with both extended-spectrum resistance and resistance to β-
lactamase inhibitors. Antimicrob. Agents Chemother. 43: 1881-1887.
157. Su-Fei Tian, Bai YI, C., Yun Zhuo, C. , Shuang, W., 2002. Prevalence of rectal
carriage of extended-spectrum β-lactamase-producing Escherichia coli among elderly
people in community settings in China Canadian. journal of microbiology
ISSN 0008-4166 CODEN CJMIAZ, 54(9): 781-785
158. Tenover, F. C., Mohammed, M. J., Gorton, T. S., & Z. F. Dembek. 1999.
Detection and reporting of organisms producing extended-spectrum betalactamases:
survey of laboratories in Connecticut. J. Clin. Microbiol. 37: 4065–4070

Références Bibliographiques
113
159. Thomson, K. S. & C. C. Sanders. 1992. Detection of extended-spectrum β-
lactamases in members of the family Enterobacteriaceae : comparison of the
doubledisk and three-dimensional tests. Antimicob. Agents Chemother. 36: 1877-1882.
160. Thomson, K. S. 1995. Improved detection of extended-spectrum β-lactamases from
disk diffusion susceptibility test (Abst. C-87). The general meeting of the American
Society for Microbiology. American Society for Microbiology, Washington, D. C.
161. Touati, A., Benallaoua, S., Forte, D., Madoux, J., Brasme, L., and DE Champs C.,
2006. First report of CTX-M-15 and CTX-M-3 β-lactamases among clinical isolates of
Enterobacteriaceae in Béjaia, Algeria. International Journal of Antimicrobial Agents.
Issue5.27: 397-402
162. Tzouvelekis, L., Tzelepi, E., Tassios, P. T. & N. J. Legakis. 2000. CTX-M-type β-
lactamases: an emerging group of extended-spectrum enzymes. Inter. J. Antimicrob.
Agents 14: 137-142.
163. Van Hoof P.A.M. 2001. Molecular modelling studies with regar to the observed
resistance profile of streptococcus pneumoniae PBP2X resulting from some naturally
occurring active site mutations.Ph.D.Thesis, Heinrich-Hein-Universität dussel dorf
(Germany). 238p.
164. Vedy, S., Rakotovao, D., Ramarokoto, C., Ratsitohaina, H., Carod, J., Ratsima,
E., Morillon M. et Talarmin, M., 2009. Rôle of contaminated aspirations tubes in
nosocomiales outbreak of Klebsiella pneumoniae producing SHV-2, CTX-M-15
extended spectrum bêta-lactamases. Journal of hospital infection, 72(1): 23-29.
165. Venezia, R. A., Scarano, F. J., Preston, K. E., Steele, L. M., Root, T.P.,
Limberger, R., Archinal, W. & M.A. Kacica. 1995. Molecular epidemiology of an
SHV-5 extended-spectrum beta-lactamase in Enterobacteriaceae isolated from infants
in a neonatal intensive care unit. Clin. Infect. Dis. 21: 915-923

Références Bibliographiques
114
166. Vercauteren, E., P. Descheemaeker, M. Leven, C. C. Sanders, & H. Goossens.
1997. Comparison of screening methods for the detection of extended-spectrum β-
lactamases and their prevalence among blood isolates of Escherichia coli and
Klebsiella spp. in a Belgian teaching hospital. J. Clin. Microbiol., 35:2191–2197.
167. Wadula, J., Von Gottberg, A., Kilner, D., DE Jong, G., Cohen, C., Khoosal, M.,
Keddy, K., Crewe-Brown, H., 2006. Nosocomial outbreak of extended-spectrum β-
lactamase-producing Salmonella isangi in pediatric wards. The Pediatric infectious
disease journal ISSN 0891-3668 CODEN PIDJEV, 25(9): 843-844
168. Waldhagen GF, Poirel L, Nordmann P. 2003. Ambler class A extended-spectrum
beta-lactamases in Pseudomonas aeruginosa: novel developments and clinical impact.
Antimicrob Agents Chemother; 47: 2385–92.
169. Walsh, C. 2003. Antibiotics: actions, origins, resistance. Washington, D.C., ASM
Press.
170. Walsh, T. R., Toleman, M. A., Poirel, L., & P. Nordmann. 2005. Metallo- β-
lactamases: the quiet before the storm? Clin. Microbiol. Rev. 18:306–325.
171. Weill, F. X., M. Demartin, D. Tandé, E. Espié, I. Rakotoarivony & P. D. A.
Grimont. 2004. SHV-12-like extended-spectrum-β-lactamase-producing strains of 19
Salmonella enterica serotypes Babelsberg and Enteridis isolated in France among
infants adopted from Mali. J. Clin. Microbiol. 42: 2432-2437.
172. Weill, F. X., R. Lailler, K. Praud, A. Kerouanton, L. Fabre, A. Brisabois, P. A.
Grimont, and A. Cloeckaert. 2004. Emergence of extended-spectrum-ß-lactamase
(CTX-M- 9)producing multiresistant strains of Salmonella enterica serotype Virchow
in poultry and humans in France. J Clin Microbiol. 42:5767-73.
173. Wiegand, I., Al-Agamy, MH., 2003. First Description of CTX-M Enzymes in
Clinical E. coli Strains from Egypt. Interscience Conference on Antimicrobial Agents
and Chemotherapy. Nature. 208: 239-244

Références Bibliographiques
115
174. Winokur, P. L., Canton, R., Casellas, J. M. & N Legakis. 2001. Variations in the
prevalence of strains expressing an extended-spectrum b-lactamase phenotype and
characterization of isolates from Europe, the Americas, and the Western Pacific region.
Clin. Infect. Dis. 32(2): S94-S103.
175. Witte, W. 1997. Impact of antibiotic use in animal feeding on resistance of bacterial
pathogens in humans. Ciba Found Symp 207: 61-71; discussion 71-5.
176. Wong-Beringer, A., Hindler, J., Loeloff, M., Queena, A. M., Lee, N., Pegues, D.
A., Quinn, J. P., and K. Bush. 2002. Molecular correlation for the treatment outcomes
in bloodstream infections caused by Escherichia coli and Klebsiella pneumoniae with
reduced susceptibility to ceftazidime. Clin. Infect. Dis. 34: 135-146.
177. Xiong, Z., Zhu, D., Wang, F., Zhang, Okamoto, Y. R. & A. Inoue. 2002.
Investigation of extended-spectrum β -lactamase in Klebsiella pneumoniae and
Escherichia coli from China. Diagn. Microbiol. Infect. Dis. 44: 195-200.
178. Yagi, T., Kruokawa, H., Shibata, N., Shibayama, K., & Y. Arakawa. 2000. A
preliminary survey of extended-spectrum β -lactamases (ESBLs) in clinical isolates of
Klebsiella pneumoniae and Escherichia coli in Japan. FEMS Microbiol. Lett., 184:53–
56.
179. Yan, J.-J., Wu, S.-M., Tsai, S.-H., Wu, J.-J. & I.-J. Su. 2003. Prevalence of SHV-
12 among clinical isolates of Klebsiella pneumoniae producing extended-spectrum β -
lactamases and identification of a novel AmpC enzyme (CMY-8) in southern Taiwan.
Antimicrob. Agents Chemother. 44:1438–1442.
180. Yang K. et Guglielmo,
B.J., 2007. Le Traitement des Bactéries Bêtalactamases à
Spectre Étendue et de Type AmpC. The Annals of Pharmacotherapy; 41( 9): 1427-
1435.

Références Bibliographiques
116
181. Yu, L. W., Wu, L. T., Pfaller, M. A., Winokur, P. L. & R. N. Jones. 2003.
Emergence of two Klebsiella Isolates harboring plasmid-mediated CTX-M-15 β-
lactamase in Tawan. Antimicrob. Agents Chemother. 48: 362-363.
182. Zeba, B., 2005. Overview of β-lactamases incidence on bacterial drug resistance.
African journal of Biotechnology. 4:1559-1562
183. Zeba, B., Kiendrebeogo, M., Lamien, A., Docquier, J. D., Simporé, J., Nacoulma
O G., 2007. Major enzymatic factors involved in bacterial penicillin resistance in
Burkina Faso. Pak J Biol sci.10(3): 506-10
184. Zeba, B., Simporé, J., Nacoulma O G., 2004. Production périplasmique de β-
lactamases par des isolats cliniques.Antibiotiques ; 7 : 183-190
185. Zeba, B., Simporé, J., Nacoulma O G., 2007. β-lactamase Molecular classes Linked
to production ways from clinical isolates in Burkina Faso/West Africa. Journal of
Applied sciences7 ISSN (1), 98-102: 1812-5654
186. Zeba, B., Simporé, J., Nacoulma O G., Frère, J.M., 2005. Identification of metallo-
β-lactamases from a clinical isolate at Saint Camille medical center of
Ouagadougou/Burkina Faso. African journal of Biotechnology. 4(3): 286-288
187. Zeba, B., Simporé, J., R. P. J., and Nacoulma, O G. 2003. Profils des substrats des
β-lactamases extraites d’isolats cliniques au Centre Hospitalier Saint Camille de
Ouagadougou. West Afr. J. Pharmacol. Drug Res. 19: 28-32.
188. Zhuo C, Li X-q, Zong Z-y, Zhong N-S, 2013. Epidemic Plasmid Carrying blaCTX-
M-15 in Klebsiella penumoniae in China. PLoS ONE 8(1): e52222.
doi:10.1371/journal.pone.0052222
Annexes
Annexe A
N°
d'ordre
de PCR Echantillons numéro Espèces Sexe des malades
Age des
malades services CTX-M SHV TEM
1 Urines 538 K. pneumoniae F ≤ 2 ans MI 1 1 1
2 selles 376 E. coli F ≤ 2 ans MI 1 0 1
3 Pus 329 E. coli F > 2 ans CHIR 1 0 1
4 Pus 130 K. pneumoniae M > 2 ans UC 1 1 1
5 LCR 736 K. pneumoniae M ≤ 2 ans REA 1 1 1
6 Urines 515 E. coli F ≤ 2 ans MI 1 0 1
7 sang 55 K. pneumoniae F ≤ 2 ans NRS 1 1 0
8 Pus 252 Pseudomonas sp M > 2 ans EXT 1 0 0
9 Pus 44 E. coli F > 2 ans CHIR 1 0 1
10 Urines 694 E. coli F > 2 ans EXT 1 0 1
11 sang 44 K. pneumoniae M ≤ 2 ans NRS 1 1 1
12 Pus 221 K. pneumoniae M ≤ 2 ans UC 1 0 1
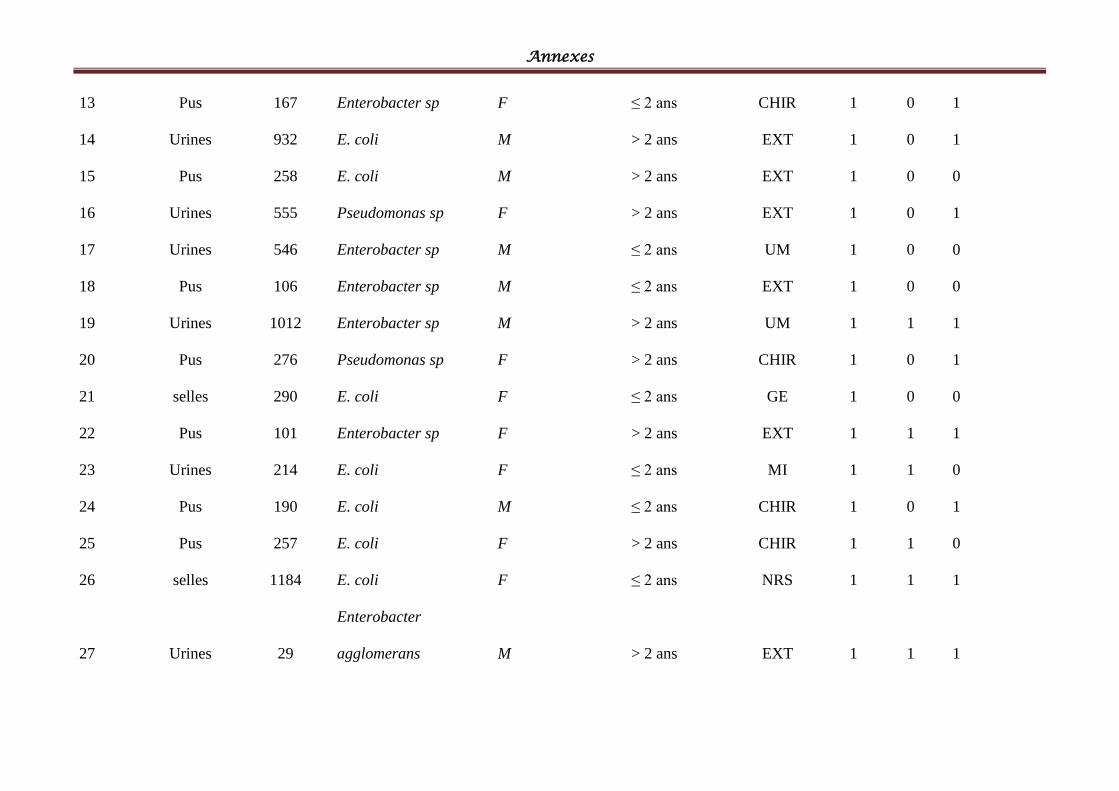
Annexes
13 Pus 167 Enterobacter sp F ≤ 2 ans CHIR 1 0 1
14 Urines 932 E. coli M > 2 ans EXT 1 0 1
15 Pus 258 E. coli M > 2 ans EXT 1 0 0
16 Urines 555 Pseudomonas sp F > 2 ans EXT 1 0 1
17 Urines 546 Enterobacter sp M ≤ 2 ans UM 1 0 0
18 Pus 106 Enterobacter sp M ≤ 2 ans EXT 1 0 0
19 Urines 1012 Enterobacter sp M > 2 ans UM 1 1 1
20 Pus 276 Pseudomonas sp F > 2 ans CHIR 1 0 1
21 selles 290 E. coli F ≤ 2 ans GE 1 0 0
22 Pus 101 Enterobacter sp F > 2 ans EXT 1 1 1
23 Urines 214 E. coli F ≤ 2 ans MI 1 1 0
24 Pus 190 E. coli M ≤ 2 ans CHIR 1 0 1
25 Pus 257 E. coli F > 2 ans CHIR 1 1 0
26 selles 1184 E. coli F ≤ 2 ans NRS 1 1 1
27 Urines 29
Enterobacter
agglomerans M > 2 ans EXT 1 1 1
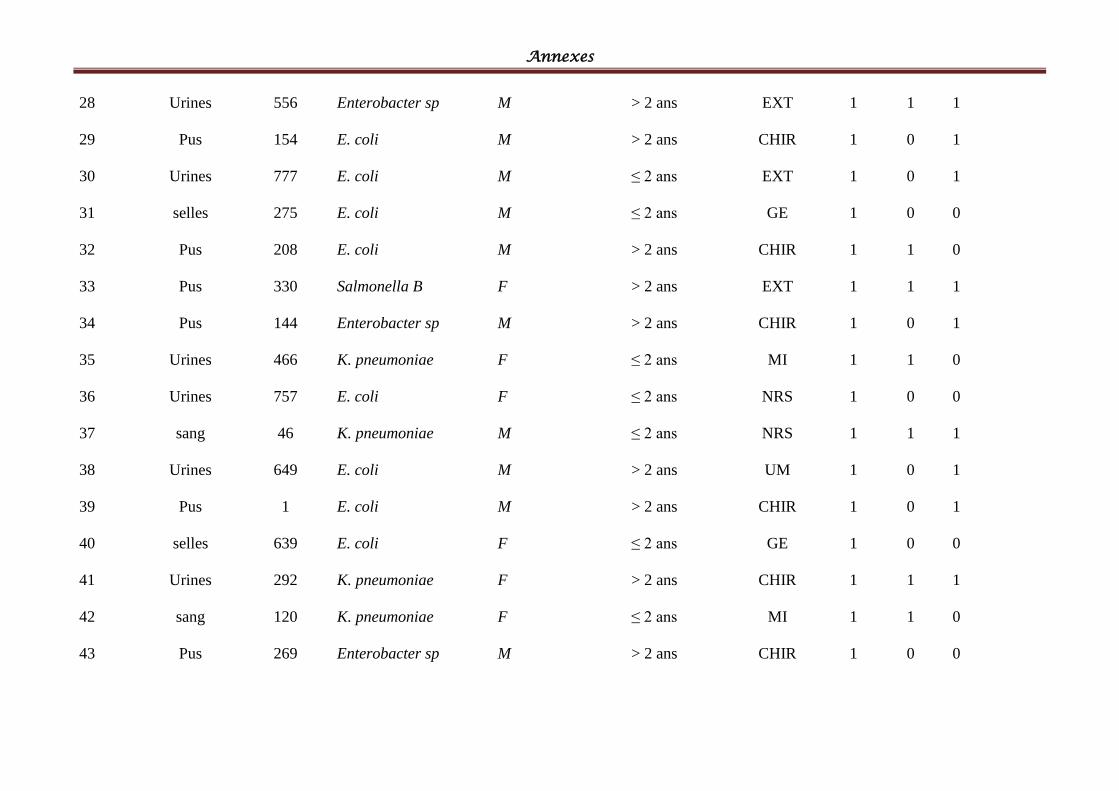
Annexes
28 Urines 556 Enterobacter sp M > 2 ans EXT 1 1 1
29 Pus 154 E. coli M > 2 ans CHIR 1 0 1
30 Urines 777 E. coli M ≤ 2 ans EXT 1 0 1
31 selles 275 E. coli M ≤ 2 ans GE 1 0 0
32 Pus 208 E. coli M > 2 ans CHIR 1 1 0
33 Pus 330 Salmonella B F > 2 ans EXT 1 1 1
34 Pus 144 Enterobacter sp M > 2 ans CHIR 1 0 1
35 Urines 466 K. pneumoniae F ≤ 2 ans MI 1 1 0
36 Urines 757 E. coli F ≤ 2 ans NRS 1 0 0
37 sang 46 K. pneumoniae M ≤ 2 ans NRS 1 1 1
38 Urines 649 E. coli M > 2 ans UM 1 0 1
39 Pus 1 E. coli M > 2 ans CHIR 1 0 1
40 selles 639 E. coli F ≤ 2 ans GE 1 0 0
41 Urines 292 K. pneumoniae F > 2 ans CHIR 1 1 1
42 sang 120 K. pneumoniae F ≤ 2 ans MI 1 1 0
43 Pus 269 Enterobacter sp M > 2 ans CHIR 1 0 0
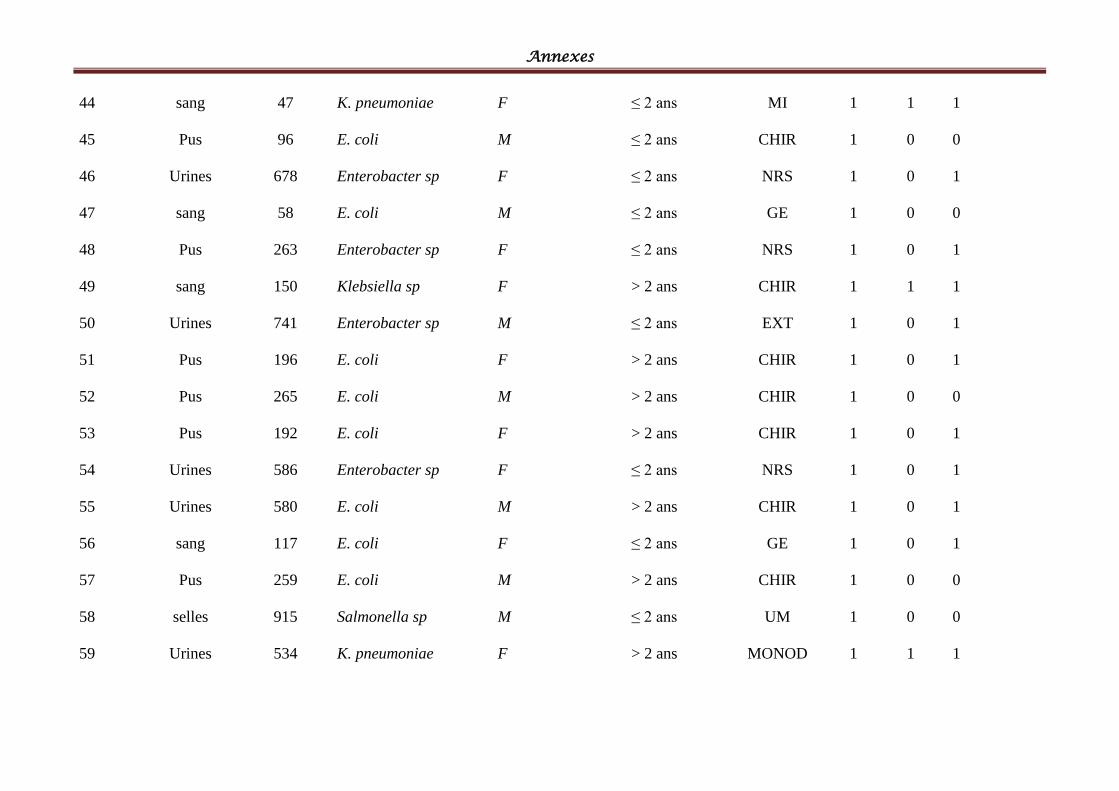
Annexes
44 sang 47 K. pneumoniae F ≤ 2 ans MI 1 1 1
45 Pus 96 E. coli M ≤ 2 ans CHIR 1 0 0
46 Urines 678 Enterobacter sp F ≤ 2 ans NRS 1 0 1
47 sang 58 E. coli M ≤ 2 ans GE 1 0 0
48 Pus 263 Enterobacter sp F ≤ 2 ans NRS 1 0 1
49 sang 150 Klebsiella sp F > 2 ans CHIR 1 1 1
50 Urines 741 Enterobacter sp M ≤ 2 ans EXT 1 0 1
51 Pus 196 E. coli F > 2 ans CHIR 1 0 1
52 Pus 265 E. coli M > 2 ans CHIR 1 0 0
53 Pus 192 E. coli F > 2 ans CHIR 1 0 1
54 Urines 586 Enterobacter sp F ≤ 2 ans NRS 1 0 1
55 Urines 580 E. coli M > 2 ans CHIR 1 0 1
56 sang 117 E. coli F ≤ 2 ans GE 1 0 1
57 Pus 259 E. coli M > 2 ans CHIR 1 0 0
58 selles 915 Salmonella sp M ≤ 2 ans UM 1 0 0
59 Urines 534 K. pneumoniae F > 2 ans MONOD 1 1 1
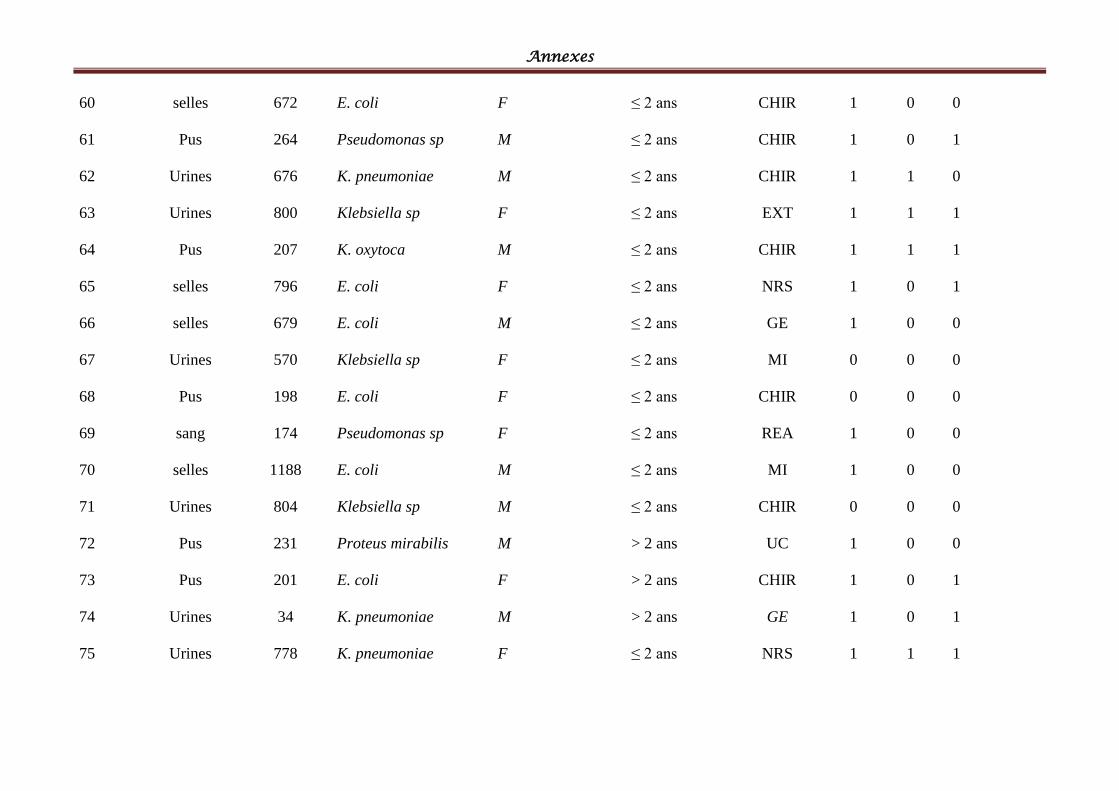
Annexes
60 selles 672 E. coli F ≤ 2 ans CHIR 1 0 0
61 Pus 264 Pseudomonas sp M ≤ 2 ans CHIR 1 0 1
62 Urines 676 K. pneumoniae M ≤ 2 ans CHIR 1 1 0
63 Urines 800 Klebsiella sp F ≤ 2 ans EXT 1 1 1
64 Pus 207 K. oxytoca M ≤ 2 ans CHIR 1 1 1
65 selles 796 E. coli F ≤ 2 ans NRS 1 0 1
66 selles 679 E. coli M ≤ 2 ans GE 1 0 0
67 Urines 570 Klebsiella sp F ≤ 2 ans MI 0 0 0
68 Pus 198 E. coli F ≤ 2 ans CHIR 0 0 0
69 sang 174 Pseudomonas sp F ≤ 2 ans REA 1 0 0
70 selles 1188 E. coli M ≤ 2 ans MI 1 0 0
71 Urines 804 Klebsiella sp M ≤ 2 ans CHIR 0 0 0
72 Pus 231 Proteus mirabilis M > 2 ans UC 1 0 0
73 Pus 201 E. coli F > 2 ans CHIR 1 0 1
74 Urines 34 K. pneumoniae M > 2 ans GE 1 0 1
75 Urines 778 K. pneumoniae F ≤ 2 ans NRS 1 1 1
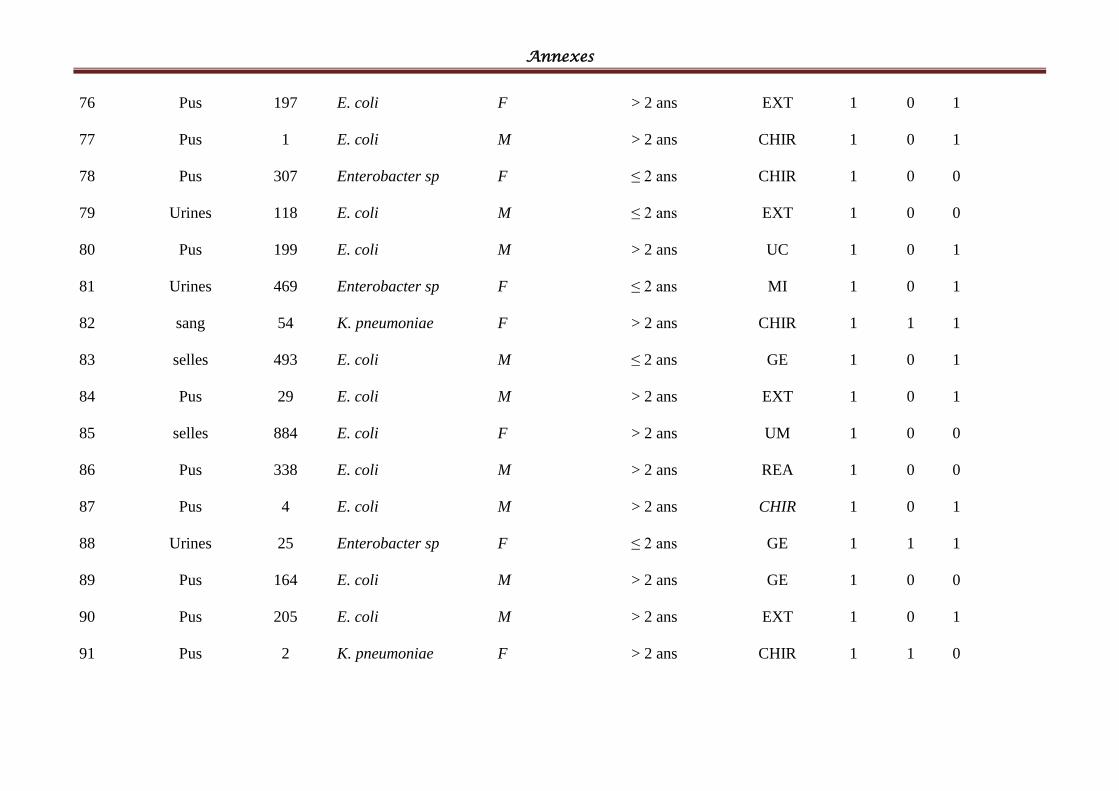
Annexes
76 Pus 197 E. coli F > 2 ans EXT 1 0 1
77 Pus 1 E. coli M > 2 ans CHIR 1 0 1
78 Pus 307 Enterobacter sp F ≤ 2 ans CHIR 1 0 0
79 Urines 118 E. coli M ≤ 2 ans EXT 1 0 0
80 Pus 199 E. coli M > 2 ans UC 1 0 1
81 Urines 469 Enterobacter sp F ≤ 2 ans MI 1 0 1
82 sang 54 K. pneumoniae F > 2 ans CHIR 1 1 1
83 selles 493 E. coli M ≤ 2 ans GE 1 0 1
84 Pus 29 E. coli M > 2 ans EXT 1 0 1
85 selles 884 E. coli F > 2 ans UM 1 0 0
86 Pus 338 E. coli M > 2 ans REA 1 0 0
87 Pus 4 E. coli M > 2 ans CHIR 1 0 1
88 Urines 25 Enterobacter sp F ≤ 2 ans GE 1 1 1
89 Pus 164 E. coli M > 2 ans GE 1 0 0
90 Pus 205 E. coli M > 2 ans EXT 1 0 1
91 Pus 2 K. pneumoniae F > 2 ans CHIR 1 1 0
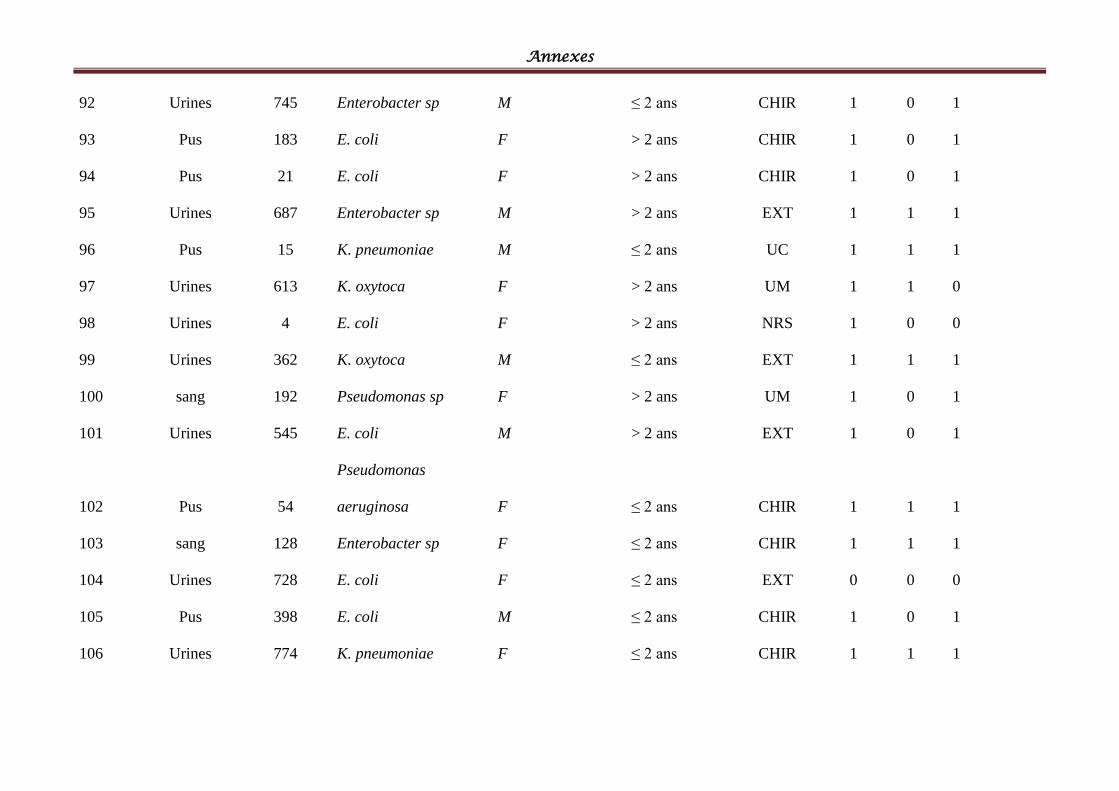
Annexes
92 Urines 745 Enterobacter sp M ≤ 2 ans CHIR 1 0 1
93 Pus 183 E. coli F > 2 ans CHIR 1 0 1
94 Pus 21 E. coli F > 2 ans CHIR 1 0 1
95 Urines 687 Enterobacter sp M > 2 ans EXT 1 1 1
96 Pus 15 K. pneumoniae M ≤ 2 ans UC 1 1 1
97 Urines 613 K. oxytoca F > 2 ans UM 1 1 0
98 Urines 4 E. coli F > 2 ans NRS 1 0 0
99 Urines 362 K. oxytoca M ≤ 2 ans EXT 1 1 1
100 sang 192 Pseudomonas sp F > 2 ans UM 1 0 1
101 Urines 545 E. coli M > 2 ans EXT 1 0 1
102 Pus 54
Pseudomonas
aeruginosa F ≤ 2 ans CHIR 1 1 1
103 sang 128 Enterobacter sp F ≤ 2 ans CHIR 1 1 1
104 Urines 728 E. coli F ≤ 2 ans EXT 0 0 0
105 Pus 398 E. coli M ≤ 2 ans CHIR 1 0 1
106 Urines 774 K. pneumoniae F ≤ 2 ans CHIR 1 1 1
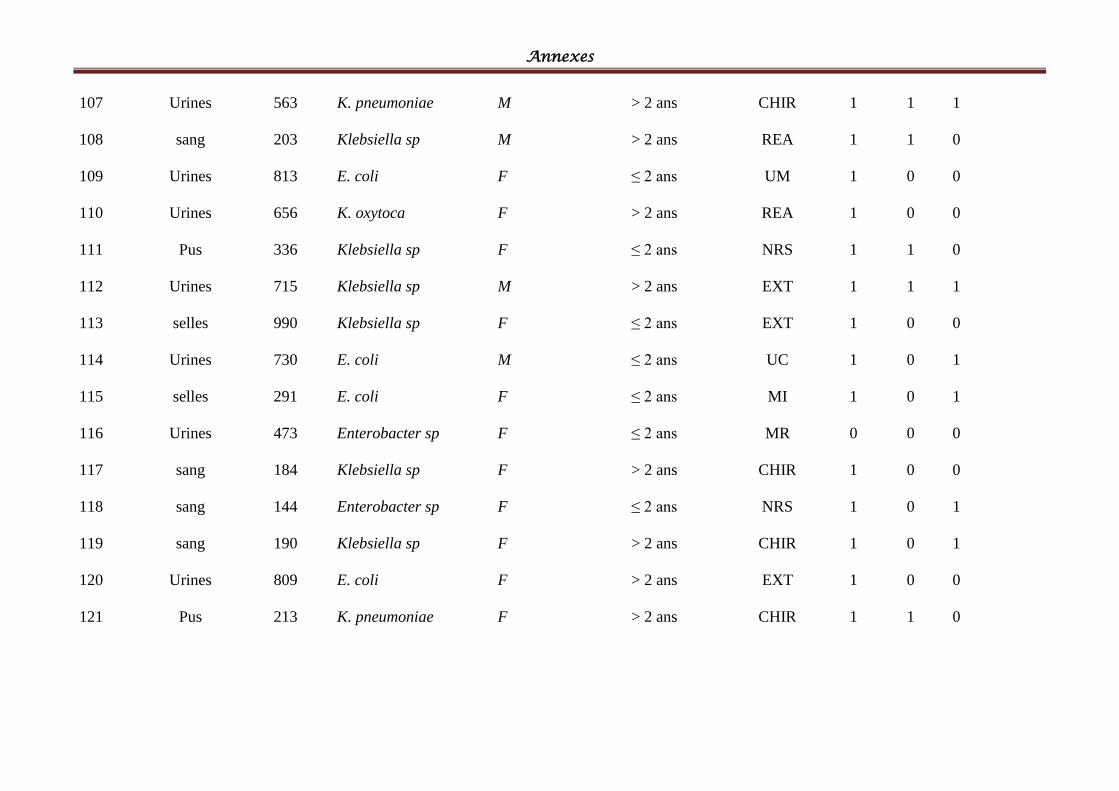
Annexes
107 Urines 563 K. pneumoniae M > 2 ans CHIR 1 1 1
108 sang 203 Klebsiella sp M > 2 ans REA 1 1 0
109 Urines 813 E. coli F ≤ 2 ans UM 1 0 0
110 Urines 656 K. oxytoca F > 2 ans REA 1 0 0
111 Pus 336 Klebsiella sp F ≤ 2 ans NRS 1 1 0
112 Urines 715 Klebsiella sp M > 2 ans EXT 1 1 1
113 selles 990 Klebsiella sp F ≤ 2 ans EXT 1 0 0
114 Urines 730 E. coli M ≤ 2 ans UC 1 0 1
115 selles 291 E. coli F ≤ 2 ans MI 1 0 1
116 Urines 473 Enterobacter sp F ≤ 2 ans MR 0 0 0
117 sang 184 Klebsiella sp F > 2 ans CHIR 1 0 0
118 sang 144 Enterobacter sp F ≤ 2 ans NRS 1 0 1
119 sang 190 Klebsiella sp F > 2 ans CHIR 1 0 1
120 Urines 809 E. coli F > 2 ans EXT 1 0 0
121 Pus 213 K. pneumoniae F > 2 ans CHIR 1 1 0

Annexes
125
Annexe B
Publications:
1. RESISTANCES TO THE OXYIMINO-CEPHALOSPORINS BY CTX-M-15
PRODUCING KLEBSIELLA ISOLATED FROM THE URINES SAMPLES OF
PATIENTS IN THE UNIVERSITY HOSPITAL COMPLEX PAEDIATRIC CHARLES
DE GAULLE (CHUP-CDG) OF OUAGADOUGOU IN BURKINA FASO
METUOR DABIRE Amana
Laboratoire d’Enzymologie de la Chimio-Résistance Bactérienne, Université de
Ouagadougou (U.O), Burkina Faso, Ouagadougou.
ZONGO K Jacob
Laboratoire d’Enzymologie de la Chimio-Résistance Bactérienne, Université de
Ouagadougou (U.O), Burkina Faso, Ouagadougou.
ZEBA Boukaré
Laboratoire d’Enzymologie de la Chimio-Résistance Bactérienne, Université de
Ouagadougou (U.O), Burkina Faso, Ouagadougou.
MOUSSAWI Jihad
Laboratoire de Biotechnologie Végétale, Université libre de Bruxelles, 12 rue des Professeurs
Jeneer et Brachet, Gosselies, Belgium
BAUCHER Marie
Laboratoire de Biotechnologie Végétale, Université libre de Bruxelles, 12 rue des Professeurs
Jeneer et Brachet, Gosselies, Belgium
EL JAZIRI Mondher
Laboratoire de Biotechnologie Végétale, Université libre de Bruxelles, 12 rue des Professeurs
Jeneer et Brachet, Gosselies, Belgium

Annexes
126
2-Title: First detection of SHV-type Extended Spectrum β-lactamases in the
University Hospital complex Paediatric Charles De Gaulle (CHUP-CDG) of
Ouagadougou in Burkina Faso.
Authors/Affiliation: METUOR DABIRE Amana1, ZONGO K Jacob
1, ZEBA Boukaré
1,
TRAORE/OUEDRAOGO Rasmata3, MOUSSAWI Jihad
2, BAUCHER Marie
2 and EL
JAZIRI Mondher 2
1 Laboratoire d’Enzymologie de la Chimio-Résistance Bactérienne (L. E. C. R. B), Université
de Ouagadougou (U.O), Burkina Faso ; B.P. : 03 B.P. 7021 Ouagadougou 03 ; Tél : (226) 50
30 70 64/30 70 65 ; Fax. (226) 50 30 72 42 ; Télex 5270 BF
2 Laboratoire de Biotechnologie Végétale, Université libre de Bruxelles, 12 rue des Professeurs
Jeneer et Brachet, B-6041 Gosselies, Belgium ; Téléphone : +32 2 650 95 72
3 Laboratoire d’analyse médicale du Centre Hospitalier Universitaire Pédiatrique Charles De
Gaulle (CHUP-CDG) de Ouagadougou.
Corresponding author : METUOR DABIRE Amana ([email protected])
Articles soummis :
1- Résistance aux β-lactamines par des bactéries gram négatif, productrices de
plusieurs types d’enzymes, isolées des urines au Centre Hospitalier Universitaire
Pédiatrique Charles De Gaulle de Ouagadougou au Burkina Faso
Resistance to β-lactamines by negative gram bacteria, producing several types of
enzymes, insulated from urines in University Hospital complex Paediatric Charles De
Gaulle of Ouagadougou in Burkina Faso

Annexes
127
Auteurs/Affiliation: METUOR DABIRE Amana1, ZONGO K Jacob
1, ZEBA Boukaré
1,
TRAORE/OUEDRAOGO Rasmata3, MOUSSAWI Jihad
2, BAUCHER Marie
2 and EL
JAZIRI Mondher 2
1 Laboratoire d’Enzymologie de la Chimio-Résistance Bactérienne (L. E. C. R. B), Université
de Ouagadougou (U.O), Burkina Faso ; B.P. : 03 B.P. 7021 Ouagadougou 03 ; Tél : (226) 50
30 70 64/30 70 65 ; Fax. (226) 50 30 72 42 ; Télex 5270 BF
2 Laboratoire de Biotechnologie Végétale, Université libre de Bruxelles, 12 rue des Professeurs
Jeneer et Brachet, B-6041 Gosselies, Belgium ; Téléphone : +32 2 650 95 72
3 Laboratoire d’analyse médicale du Centre Hospitalier Universitaire Pédiatrique Charles De
Gaulle (CHUP-CDG) de Ouagadougou.
Auteur correspondant : METUOR DABIRE Amana ([email protected])
Poster :
METUOR DABIRE Amana; ZONGO K. Jacob; ZEBA Boukaré. Dépistage des
bactéries productrices de β-lactamases a spectre élargi (BLSE) en milieu pédiatrique.
Journées portes ouvertes sur les activités des doctorants, à l’université de Ouagadougou, 2010.
Communication orale :
METUOR DABIRE Amana; ZONGO K. Jacob; ZEBA Boukaré. Résistance des bactéries
Gram-négatif aux oxyimino-céphalosporines par production de β-lactamases a spectre élargi
(BLSE) en milieu pédiatrique.
Journées portes ouvertes sur les activités des doctorants, à l’université de Ouagadougou, 2011.

Annexes
128
Annexe C
COLORATION DIFFERENTIELLE
La coloration la plus courante est celle de Gram (Gram, 1884) car fondamentale en
bactériologie.
Réactifs nécessaires : solution de cristal violet, solution de lugol, alcool 95°, solution de
safranine.
Conditions opératoires :
Préparation du frottis
Suspendre un peu de culture bactérienne dans une goutte d’eau, sécher l’étalement en agitant
la lame dans le courant d’air chaud, provoqué par la flamme d’un bec bunsen. Fixer à la
chaleur en écrasant deux fois la lame avec la flamme, ou fixer à l’alcool 95°C en mouillant le
frottis et laisser reposer 5 minutes.
Étapes de la coloration et observation du frottis coloré
Recouvrir le frottis de cristal violet et laisser agir 1 min
Rincer à l’eau pour enlever l’excès de colorant en veillant à ne pas détacher le frottis
Recouvrir avec quelques gouttes de lugol et laisser agir 1 min
Rincer à l’eau, égoutter
Traiter la préparation avec l’alcool goutte à goutte jusqu’à ce que l’alcool ajouté
n’entraîne plus le cristal violet,
Rincer à l’eau
Recolorer à la safranine pendant 30s

Annexes
129
Rincer à l’eau, égoutter, sécher doucement entre deux feuilles de papier joseph (filtre)
Observer à l’aide d’huile à immersion à l’objectif X100. Noter les observations
NB : Le principe de la coloration est basé sur la nature de la paroi des bactéries. Cela permet
de distinguer les bactéries à GRAM+ (couleur violette) des bactéries à GRAM- (couleur rose
ou rouge).