Embed Size (px)
Citation preview

ENVIRONNEMENTS ET SPECIES DU PARCO DEL BEIGUAET DES SITES DE LA RETE NATURA 2000 FONCTIONNELLEMENT LIES
par Sergio G. Fasano, Renato Cottalasso, Massimo Campora, Luca Baghino, Roberto Toffoli et Antonio Aluigi.

Realizzato da Il Piviere S.r.l.
Via G. Mameli 16 r - 15066 Gavi (AL)
tel e fax +39 0143346427
[email protected] www.ilpiviere.com
Coordinamento editoriale: A. Aluigi, M. Burlando, M. Campora,
R. Cottalasso, S. G. Fasano
Testi: S. G. Fasano, M. Campora, R. Cottalasso, L. Baghino, R. Toffoli
Foto in copertina: Codirossone di R. Cottalasso
Le foto, salvo differenti indicazioni, sono degli autori
Elaborazioni grafi che e GIS: S. G. Fasano
Progetto grafi co ed impaginazione: Elisa Dolcino
Stampa: Grafi che G7
Finito di stampare nel mese di marzo 2013
© 2013 Ente Parco del Beigua
Proprietà artistica e letteraria riservata. È vietata la riproduzione anche parziale di testi, fotografi e, disegni,
illustrazioni, in ogni forma e con qualsiasi mezzo, compresi microfi lm e copie fotostatiche.
Realisé par Il Piviere S.r.l. Via G. Mameli 16 r - 15066 Gavi (AL) tel et fax +39 0143346427 [email protected] www.ilpiviere.com
Coordination éditoriale: A. Aluigi, M. Burlando, M. Campora, R. Cottalasso, S. G. Fasano Textes: S. G. Fasano, M. Campora, R. Cottalasso, L. Baghino, R. Toffoli Photo de couverture: Monticole de roche de R. Cottalasso Les photos, sauf indication contraire, sont des auteurs Designs graphiques et GIS: S. G. Fasano Conception graphique et mise en page: Elisa Dolcino Traduction: Decarolis Traduzioni di Dott.ssa Eliana Decarolis
Imprimé en Mars 2013 © 2013 Ente Parco del Beigua
Propriété littéraire et artistique réservés. Il est interdit la reproduction même partielle des textes, des photographies et des dessins, illustrations, sous quelque forme ou par quelque moyen, y compris les microfilms et la photocopie.

AMBIENTI E SPECIEDEL PARCO DEL BEIGUA
E DEI SITI DELLA RETE NATURA 2000 FUNZIONALMENTE CONNESSI
a cura di Sergio G. Fasano, Renato Cottalasso, Massimo Campora, Luca Baghino, Roberto Toffoli e Antonio Aluigi
ENVIRONNEMENTS ET SPECIES DU PARCO DEL BEIGUAET DES SITES DE LA RETE NATURA 2000 FONCTIONNELLEMENT LIES
par Sergio G. Fasano, Renato Cottalasso, Massimo Campora, Luca Baghino, Roberto Toffoli et Antonio Aluigi.

4 AMBIENTI E SPECIE444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444 AAMAMAMMBAMAMBAMBMBAAMAAMMBBMBMBAMBMBMAMMAMBAMBMMBMAMMMBAAMAMMMMBBMBMBBBBBAMBAMBBBAAAAAAMAMAAAAAAAMAMAMBAMBAAMAMAMMBMBMAMBMBMMMMAMMMBMAMBMBBBBBBMBBMBBBAAAAAAMAMAMAAAAMAAMAAAAMMMMBAMMBMAMMAMBBBBBAMBAMBMBBBBBAAAAAAAMAAAAAAAAAAAAAAAMBAMBMBMBMBMMBMAMAMMMMMBMMBMMBMBMAMBBMBBMBBBBBBBBAAAAMBAAAAAAAAAAAAAMAMBMBMBMMBMMMMMMMAMBAMBMBMBBBBBAAAAAAMAAAAMBAAAAAAMBAAAAMMBMBAMMAMMMAMAMMBMBMBMBMBAMBMBBAMBAMBMBMAAAAAAAMAAAAAAAAAAAAMAAMBMMBMAMMAMBBAMBAMBBBAMBAMBAMBAAAAAAMAAAAAAAAAAAAAMBMMMMAMAMBMBBBBBAMBBBAMBBAMBMAMAAAAAAAAMAAAAAAAAAAAAMBMMMMBAMAMAMMMMBMBBBAMBBAMBAMBAAAAAAAAAAAMBAAAAAAAMBMMMMMBAMMMMBMBBBAMBBBBBBAAAAAAAAAAAAAMBAAAAAAAAMBMMMBMMMMMMMMMMBBBBBBBBAAAAAAAMAAMBAMBAAAAAAMMMMMMMMMMMMMMMAMBBBAAAAAAAAMBAAAAMBAAAMMMMMAMBMAMMMMMMMBBAMBAAAAAAAAAAMMMMMMMMMMMBBBAAAAAAAAMAAAMBMMBMMBMMMMMMMBMBBBBBAMBAAAAAAAAMBAAMAAMMMMAMAMMMMMMAMBAAAMAAAAAAAAAAMBMMMAMAMAMMMMMBBBMBMBMBBAMBBBBBAAAMMMMMMMMMMMAMBMBBBBBMBAAAAAAAAAAAMMMMMMAMMMMMMMBBBBBBAAAAAAAAAMMMMMMMMMMAMMMMMMMBBBBAAAAAAAAAAAMBAMMMMMMMMMMMMMMMMMMBBBBBAAAAAAAAAAAMMMMMMMAMMMMMMMMMMMMAMBBAAAAAAAAAAAAAMAAMMMMMMMMMMMMMMMMMAMMBAAAAAAAAAAMMMMMMMMMMMMMMMMMMBBBBBAAAAAAAAAMAMMMMMMMMMMMMMMMMMMMBBB EI NIENIIENIIENENIENENENENEEEEEENENIENI NIENNIENENIEIIENIENIIIENEEIEEIEEEEEEENNNNNNNNNNNENNNNNNNNNENNIIENIENIENENIEENEIENEEEEEENNNNNNNNNNNENNNNNNNIENIENIIIIIIIIIENEENENIEENENENEEENENEENENEENNNNNNNNNNIENNNNENNIENNNNIENIENIEIIIIIIIENENEENEEIENEENENEEENNNNNNNNNNNNNIENIENNNNNNNIEIENIIIIIIIEIEENEEEEEEENIENNNNNNNNNENENENNNNIENNNNIEIENIIIIIEIENENEEEENNNNNIENNNNNNIENENNENNNIIIIIIEIEIEEENEEEEENNNNNNNNIENENNNIENENENNIENIENIIIIIIIIIENIENEENEEEEENENENNNNNNNNNNNNNNNNNIENIENIIIIIIIIEIENENENEENENENNENNENNNIENNNIENNIENNENIIIIIIEENEEENIENNENNNNNIENIIIIIIIIEEENENNENNNIENNNNIENNNNIIIIIIIEEEEENENNNNNNNNNNIENNNNIENIIIIIIENEEEEEENNNIENNNNNIIIIIIEEENENEENEEEEENENNNNNNNNNNNNIENIIIIEEEEEEEEEENNNNNNNIIIIIENIEEEEENEEEEEIEIENNNNNIENNNNNNNNNIIIIENIIENIEEEEIENIENNNNNNNIIIIEENNNNNIIEEENNNNNNNIIEENENNENNNNNNNIIIIIENNNNIIIIEENENNNNNNIIIIEEEEEENNNNNNNIIIIENNNTTIITITTITTTTIIIITITTTITITITTTTTTITITIIIIITTTTTTTITTITTTITITIIIIITTTTTTTTTTITITITTTTTITIIIITIITIIITTTTTTTTTTITTTTTTTIIIIIITTTTTTTTTTTTITTIIIIIITIIITTTTTTTTTTTITTTTTIITIIIIITTTTTTITTITTTTTTTITIITIITITTTTTTTTTTTTTITTTTIITIITIIIITTTTTTTITTTTTTTTIIIIIIITTTTTTTTTITTTITIIITITTTTTTTITTTIIIIITTTTTTITITITTTIIIITIITTTTTTTTTITITIIITTTTTTTTTTTTIIIIITIITIITTTTTTTTTTTIIIIIIITTTTTTTTTTTTTIIIIIIIITTTTTTTTTTTIIIIIITITITTTTTTTTTTTIIIIIITTTTTTTTTTTTTTIIIIITIIITTTTTTTTTTTTTTTTIIITITITTTTTTTTIIIIITTTTTTTTTTTTIIIIIITTTTTTTIIIIIITTTTTTTTTTTTTTIIIIIII SE SE SSE SEEEE SEE SEE SSSSE SSSSSE SE SE SEEEE SE SEEEEE SEEE SE SSSSSSSE SSSSE SSE SE SEEE SE SEEEEEE SSSSSSE SSSSSE SE SEEEEE SEEEEEEEEEEE SE SSSSSE SSE SSSSSSE SSSEE SEEEEEEEEEE SE SSSSE SSSSE SSEE SEE SEEEEEEEEEEEEE SSSE SE S SSSSSSSSSSSEE SE SEEEE SE SSE SSSSSSE SEEEEEEEEEEEEEE SSSSSSSSSSE SEEEEEEEEEEEEEEEE SSSSSSSSSSE SEEEEEEEEEEEEEE SSE SE SSE SE SE SE SEEEEEEEEEEE SSSSSSE SE SEEEEEEEEE SSSSSEEEEEEEEEEEEE SSSSSEEEEEEEEE SE SEEEEEEE SE SSSEEEEEEEE SSSSEEE SEEEEEEE SSSSSEE SEEEEE SEEEEE SSSSSSSSE SE SEEEEEEEE SSSE SSSE SEEEE SSSSSSEEEEEE SSEEEEEEE SSSSEEEEEEEE SSSSSSSEEEEEEEE E EEE SSEEEEEEEEEEEE SSSS EPECPECPECPECPECCPPECCPEPECPECPEPEPEECPECPECPECPECECECPECECECPECPECPECPECPECCEPECPECECECPECPECPECPECPEPEPEPEPECPPECPPECPPPPEPEECEEECEPEPECEPECPECCECECCPECPECPPECPECPECPPECPPPEPEECEPEEEPECPECCPECCECPECPECPPPPEPECPPPPPEPPPPEEECECPECEEEPECECPEPEEPECCCCCCPECCCCCPECECPECCCECPECPEPPPPPPPPPPECEEPECECPECCCCCCPECECPECPECPECPPEPPECPEPPPEPPPECECPEEEEECCCCCCCECCPECPPPECPPPPPPPPPEECPEPEEPECCCECPECPECPEPEPPECPPPECPPPEPECPECEPECPECECCCCCCPECPECPPEPPPPPPPEECEECECCCCCCCPEPPEPPPPPPEEEPEECECCPECECCCCPPPPPPPPPEEEEECCCCCCPECPEPEPPPPPPEPECEPECPEPECCPECCCCPPPPPPPPEPEEECCCCCPPPPPPPPPEEPECCCCPPPPPPPPPPPPPEEEEECPECCPPPPPECPPPEEEEECCCCPECCPPECPPPPPPPEEECCCCCCCCCCCCPPPPEPPPPPEEEECCCCCCCPPPPEEEECCCCPPPECPPPECEPEECCCPECCCPECPEECCCECPEEEEECCCCCCPPPPPPPPPPPEEEEEECCCCPPPPPPPPEEEEECCCCCCCCCCC EIEIEEIIIEIEIEIEIEEEEEIEEIEIIIIEEEEEEEEEEEEEEEEIEIIEIIEIEEEEEEEEEIEIEIIEIEIEIIEIEIIIIIEEEEEIEEEEEIEEEIEEIEIEIIEIEIIIEEEEEEEIEIEIEIIIIEEEEEEEIEEIEIEIEIEIIIEEEEEEIEIEIEIEIIIEIIEEEEEEEEEEEEEEIEIEIEIEIIIIEEEEEEEEEEIEIEIIIIEEEEEEEEEEIEIEIIEEEEEEEEIEIEIIIIIEIEEEEEEEEEIEIEIIIIIEEEEEEEIEEEIEIIIIIEEEEEIIIEEEEEEEEEEEIEEEEEEEEIIIEIIEEEEEEEIEIIIIIEEEEIIEIIEEEEEEIIEEIEEEEEIIIIIEEEEEEEEEEIIIIEEEEEEEEIIIEEEEEEEIIIIEEEEEEEIIIIIEEEEEEEEE
INDICE
Prefazione ........................................................................................... pag. 7
Introduzione ........................................................................................... pag. 8
La rete ecologica europea ....................................................................... pag. 9
Breve guida alla lettura .......................................................................... pag. 10
Il Progetto di “Indagini ambientali e divulgazione dei risultati
delle attività di ricerca condotte nel Parco del Beigua e nei settori
della Rete Natura 2000 funzionalmente connessi” ............................... pag. 12
IL PARCO DEL BEIGUA .................................................................. pag. 14
ATTIVITÀ DI RICERCA, CENSIMENTO E MONITORAGGIO .......................................................................... pag. 18
GLI UCCELLI ................................................................................. pag. 20
L’AVIFAUNA NIDIFICANTE .............................................................. pag. 26
LA MIGRAZIONE ........................................................................... pag. 31
LA MIGRAZIONE DEI RAPACI DIURNI ................................................ pag. 32
L’ATTIVITÀ DI INANELLAMENTO A SCOPO SCIENTIFICO ...................... pag. 39
I CHIROTTERI ............................................................................... pag. 41
IL TRAPPOLAGGIO FOTOGRAFICO .......................................... pag. 48
IL LUPO .......................................................................................... pag. 51
INDEX
Préface pag. 7
Introduction pag. 8
Le réseau écologique Europeén pag. 9
Petite guide à la lecture pag. 10
Le projet des “Indagini ambientali e divulgazione dei risultati delle attività di ricerca condotte nel Parco del Beigua e nei settori della Rete Natura 2000 funzionalmente connessi” pag. 12
LE PARCO DEL BEIGUA pag. 14
ACTIVITES DE RECHERCHE, RECENSEMENT ET CONTROLE pag. 18
LES OISEAUX pag. 20
La communauté des oiseaux nidifiants pag. 26
La migration pag. 31
La migration des Rapaces diurnes pag. 32
L’activité de baguement à but scientifique pag. 39
LES CHIROPTERES pag. 41
LES TRAPPES PHOTOGRAPHIQUES pag. 48
LE LOUP pag. 51

AMBIENTI E SPECIE ....................................................................... pag. 59
BOSCHI ........................................................................................... pag. 60
Falco pecchiaiolo ................................................................. pag. 62
Astore ................................................................................. pag. 63
Picchio nero ......................................................................... pag. 64
Cincia dal ciuffo................................................................... pag. 65
AREE APERTE ............................................................................... pag. 66
LE PRATERIE ................................................................................ pag. 68
Pernice rossa ........................................................................ pag. 69
Biancone ............................................................................. pag. 70
Succiacapre ......................................................................... pag. 71
Tottavilla ............................................................................. pag. 72
Calandro .............................................................................. pag. 73
Ortolano .............................................................................. pag. 74
GLI ARBUSTETI ............................................................................. pag. 75
Magnanina comune .............................................................. pag. 76
GLI AMBIENTI ROCCIOSI ................................................................ pag. 77
Aquila reale ......................................................................... pag. 78
Falco pellegrino ................................................................... pag. 79
Gufo reale ........................................................................... pag. 80
Codirossone ......................................................................... pag. 81
I RETTILI ..................................................................................... pag. 82
CORSI D’ACQUA E ZONE UMIDE ............................................... pag. 83
GLI ODONATI ................................................................................ pag. 84
I PESCI ........................................................................................ pag. 85
GLI ANFIBI ................................................................................... pag 86
Merlo acquaiolo ................................................................... pag. 87
AREE ANTROPIZZATE E ZONE AGRICOLE .............................. pag. 88
Averla piccola ...................................................................... pag. 89
Bibliografi a ........................................................................................... pag. 90
ENVIRONNEMENTS ET SPECIES pag. 59
LES BOIS pag. 60
Bondrée apivore pag. 62
Autour des palombes pag. 63
Pic noir pag. 64
Mésange huppée pag. 65
ZONES OUVERTES pag. 66
LES PRAIRIES pag. 68
Perdrix rouge pag. 69
Circaète Jean-le-blanc pag. 70
Engoulevent d’Europe pag. 71
Alouette lulu pag. 72
Pipit rousseline pag. 73
Bruant ortolan pag. 74
LES ARBRISSEAUX pag. 75
Fauvette pitchou pag. 76
LES ENVIRONNEMENTS ROCHEUX pag. 77
Aigle royal pag. 78
Faucon pélerin pag. 79
Hibou grand-duc pag. 80
Monticole de roche pag. 81
LES REPTILES pag. 82
COURSES D’EAU ET ZONES HUMIDES pag. 83
LES ODONATES pag. 84
LES POISSONS pag. 85
LES AMPHIBIENS pag. 86
Cincle plongeur pag. 87
ZONES ANTHROPIQUES ET ZONES AGRICOLES pag. 88
Pie-grièche écorcheur pag. 89
Bibliographie pag. 90

6

7
PREFAZIONE
Il modello di gestione del Parco naturale regionale del Beigua, scaturito nel biennio 2000-2001 nella complessa fase di redazione del piano dell’area protetta, era fondato sui concetti base della conoscenza, della divulgazione delle conoscenze acquisite e della conservazione-valorizzazione. Oltre dieci anni di costante presenza sul territorio, unita-mente ad azioni mirate di ricerca approfondita e metodologicamente appropriata, hanno prodotto un ragguardevole bagaglio di dati e conoscenze negli ambiti delle piante, degli artropodi, dei funghi, dei pipistrelli, con la pubblicazione di guide tematiche: Uccelli 2006; Chirotteri 2006; Funghi 2007. A partire dal 2006 l’azione del parco ha riservato particolare attenzione alle attività di monitoraggio, soprattutto degli uccelli, sia nidifi can-ti che migratori; dal 2011 le indagini si sono incentrate sul lupo, integrando ed appro-fondendo quanto si sta già ottenendo con il progetto regionale “Il Lupo in Liguria”, ma hanno riguardato anche diverse specie di libellule, farfalle, anfi bi e rettili. Nel recente biennio, l’adesione del Parco del Beigua al progetto di Cooperazione delle Reti Ecolo-giche del Mediterraneo (Co.R.E.M.; in partnership con il Parco di Portofi no, il Parco della Maremma e la Provincia di Oristano), ha impresso una nuova e vigorosa sferzata di energie ai monitoraggi delle specie viventi nel territorio del Parco e dei siti della Rete Natura 2000 (Siti di Importanza Comunitaria - SIC, Zone di Protezione Speciale – ZPS) ad esso connessi. Il risultato è costituito da una mole decisamente signifi cativa di nuovi dati e conoscenze, ma anche dal coinvolgimento di molteplici realtà, dagli enti locali, al mondo scolastico e dell’università, dall’associazionismo, alle forze di pubblica tutela, nei progetti di divulgazione e di conservazione-valorizzazione.
Il presente volume, che porta le fi rme dei migliori esperti di settore che hanno colla-borato in questi anni attorno al vasto e complesso “progetto biodiversità” del Parco del Beigua, rappresenta la migliore sintesi dei risultati conseguiti che, come si evince dal titolo, sono stati organizzati secondo lo schema ampiamente collaudato di ambienti e specie. Il compito che attende il Parco nel prossimo futuro sarà indubbiamente quello di mantenere costantemente aggiornata una tale m ole di dati e conoscenze, nonché di sapersi misurare con effi cacia sugli obiettivi prefi gurati dallo stesso progetto Co.R.E.M.: tutela e valorizzazione del patrimonio naturalistico e della biodiversità della Rete Eco-
logica, mirando a ridurre la pressione e le minacce sulle risorse ambientali e a favorire una fruizione sociale ed economica sostenibile delle stesse, grazie al coinvolgimento edalla sensibilizzazione dei cittadini e delle imprese.
Prof. Dario FranchelloPresidente del Parco naturale regionale del Beigua
PRÉFACE
Le modèle de gestion du Parco naturale regionale del Beigua, émergé en 2000-2001 dans le processus complexe d'élaboration du plan de la zone protégée, était fondé sur les concepts base de la connaissance, de la divulgation des connaissances achevées et de la conservation - valorisation. Plus de dix ans de présence continue dans le territoire, ainsi que des actions spécifiques de recherche approfondie et méthodologiquement appropriée, ont produit une remarquable richesse de données et des connaissances dans les domaines des plantes, des arthropodes, des champignons, des chauves-souris, avec la publication de guides thématiques: Uccelli (Oiseaux) 2006; Chirotteri (Chiroptères) 2006; Funghi (Champignons) 2007. À partir de l’année 2006, l'action du Parc a accordé une attention particulière aux activités de surveillance, en particulier des oiseaux, soit nicheurs soit migrateurs ; depuis 2011, les recherches ont étés concentrées sur le loup, en intégrant et approfondissent ce qui est déjà en train d’être obtenu par le projet régional “Il Lupo in Liguria” (Le Loup en Ligurie), mais ont aussi concerné plusieurs espèces de libellules, papillons, amphibiens et reptiles. Au cours des récentes deux années, l’adhésion du Parco del Beigua au projet de Coopération des Réseaux Écologiques dans la Méditerranée (Co.R.E.M.; en partnership avec le Parco di Portofino, le Parco della Maremma et la Provincia di Oristano), a donné une nouvelle et vigoureuse énergie aux monitorages des espèces vivantes dans le territoire du Parc et des sites de la Rete Natura 2000 (Sites d’Importance Communautaire - SIC, Zones de Protection Spéciale – ZPS) à lui reliés. Le résultat est constitué par une significative quantité de nouvelles données et connaissances, ainsi que par l’implication des réalités multiples, des collectivités locales, du monde des écoles et de l’université, des associations, des forces publiques, des projets de divulgation et de conservation et valorisation. Ce volume, qui porte les signatures des experts les meilleurs du secteur qui ont contribué dans ces années au grand et complexe “progetto biodiversità” du Parco del Beigua, représente la synthèse la meilleure des résultats achevés qui, comme on peut évincer par le titre, ont étés organisés selon le schéma amplement éprouvé des environnements et espèces. Le tâche qui attends le Parc dans le futur prochain sera sans doute cela de maintenir constamment mise à jour cette quantité de données et connaissances, ainsi que de se mesurer avec efficacité sur les objectifs préfigurés par le projet Co.R.E.M. (Co.R.ET.M): la protection et la valorisation du patrimoine naturel et de la biodiversité du Réseau écologique,en visant à réduire la pression et les menaces sur les ressources environnementales et à favoriser une fruition sociale et économique soutenable de ces dernières, grace à la participation et à la sensibilisation des citoyens et des entreprises.
Prof. Dario Franchello President du Parco naturale regionale del Beigua

8
INTRODUZIONE
Il sesto programma di azione europeo “Ambiente 2010: il nostro futuro, la nostra scel-ta”, aveva individuato nell’uso razionale delle risorse ambientali e nella salvaguardia degli ecosistemi, i presupposti essenziali dello sviluppo sostenibile, integrandoli con gli obiettivi socio-economici.
La rete ecologica europea Natura 2000, assieme al progetto di rete Smeraldo per i pa-esi non ancora membri, si propone di identifi care un’infrastruttura naturale e ambientale fi nalizzata a mettere in relazione e a connettere aree dotate di una elavata presenza di naturalità. Nell’ambito di questo complesso di elementi le aree protette assumono il ruolo di nodi da connettere tra loro e con altre aree di rilevante interesse naturalistico (Siti di Importanza Comunitaria - SIC, Zone di Protezione Speciale - ZPS, Aree Importanti per l’Avifauna - IBA, Siti Smeraldo) attraverso corridoi ecologici e zone di transizione (o “cuscinetto”).
In questa prospettiva si riconoscono alle aree protette forti responsabilità nell’avviare e incrementare processi di miglioramento e valorizzazione ambientale, di tutela della bio-diversità e nel fungere da laboratori di sperimentazione di forme innovative di sviluppo sostenibile, che siano poi esportabili al contesto territoriale circostante.
Coscienti di questa responsabilità, gli enti preposti alla gestione sono chiamati a ca-pitalizzare gli investimenti effettuati dalla collettività preservando e incrementando il valore naturalistico dei settori sottoposti alla loro tutela. In questo ambito è rilevante porre le basi per una collaborazione e una sinergia d’azione riferita alle stesse aree bioge-ografi che, indipendentemente dai confi ni amministrativi e nazionali.
Gli enti gestori hanno quindi la responsabilità della conservazione a lungo termine dei valori naturalistici presenti sul territorio protetto. La catalogazione di questi valori e la capacità di seguirne l’evoluzione nel tempo risultano quindi determinanti e preliminari per poter agire con cognizione di causa. Questo principio, sicuramente valido e rico-nosciuto teoricamente, nella realtà si scontra con esigenze e limitazioni di tipo pratico. Prima di tutto: a che livello considerare la conoscenza della propria area suffi ciente per un agire responsabile? E in seguito, vista la limitazione dei mezzi a disposizione, quale priorità d’azione attuare?
Il lavoro svolto nel Parco del Beigua, prendendo spunto e sviluppando quanto già realizzato nell’ambito del programma comunitario Interreg III A Italia – Confederazione Elvetica con il progetto “Indagine naturalistica e variabilità ambientale – Impostazionedi una piattaforma comune di lavoro per la verifi ca degli obiettivi di conservazione e per la realizzazione di programmi di ricerca e di monitoraggio nelle aree protette”, vorrebbe porre le basi per una gestione comune dell’informazione e per la messa a punto di stru-menti di aiuto alla decisione che permettano ad ogni Ente di agire localmente ponderando in maniera oggettiva i processi decisionali che sfociano in azioni concrete sul territorio. Si intendono in questo senso sia azioni a livello gestionale, sia azioni riferite a indirizzi di studio e ricerca, al completamento di lacune conoscitive, alla defi nizione di controlli ripetuti nel tempo, alla classifi cazione dei valori in scale di priorità, alle defi nizioni voca-zionali di comparti e pianifi cazioni ecologiche, ma anche, non ultime per importanza, le azioni volte alla divulgazione delle informazioni acquisite.
INTRODUCTION
Le sixième programme-cadre européen “Environnement 2010: notre avenir, notre choix”, avait individué dans l’utilisation rational des ressources environnementales et dans la sauvegarde des écosystèmes, les conditions essentielles du développement soutenable, en intégrant-les avec des objectifs socio-économiques. Le réseau écologique Européenne Natura 2000, avec le projet de réseau Emeraude pour les pays ne pas encore membres, propose d’identifier une infrastructure naturelle et d’environnement finalisée à mettre en relation et à connecter les zones ayant haute présence de naturalité. Dans le cadre de cet ensemble d’éléments, les zones protégées prennent le rôle de nœuds à relier l’un avec l’autre et avec des autre zones de relevant intérêt naturel (Sites d’Importance Communautaire - SIC, Zones de Protection Spéciale - ZPS, Aires Importantes pour les Oiseaux - IBA, Sites Emeraude) au moyen des couloirs écologiques et de zones de transition (ou « tampon »). Dans cette perspective, on reconnaît aux aires protégées des grands responsabilités dans le déclenchement et l’incrément des processus d'amélioration et valorisation de l'environnement, de la protection de la biodiversité et dans l'agir comme des laboratoires pour tester des formes innovantes de développement soutenable, qui soient ensuite exportables au contexte territorial local.En ayant conscience de cette responsabilité, les organismes préposés à la gestion sont donc chargés à la capitalisation des investissements effectués par la collectivité en préservant et augmentant la valeur naturaliste des secteurs soumis à leur protection. Dans ce cadre il est important de mettre des bases pour une collaboration et synergie d’action référée aux mêmes aires biogéographiques, indépendamment des frontières administratives et nationales. Les organismes de gestion ont donc la responsabilité de la conservation à long terme des valeurs naturelles qui sont présentes sur le territoire protégé. La classification de ces valeurs et la capacité d’en suivre l’évolution au fil du temps, sont donc déterminantes et préliminaires à fin de pouvoir agir en connaissance de cause. Ce principe, bien sûr valide et reconnu par la théorie, dans la réalité se heurte avec exigences et limitations pratiques. Tout d’abord : à quel niveau faut-il considérer la connaissance de sa propre aire comme suffisante pour un agir responsable ? Et en suite, vu la limitation des moyens disponibles, quelle priorité d’action faut-il effectuer ? L’activité développée dans le Parco del Beigua, en inspirant et développant ce qui a été déjà réalisé dans le cadre du programme communautaire Interreg III A Italie – Confederation Helvetique avec le projet “Indagine naturalistica e variabilità ambientale – Impostazione di una piattaforma comune di lavoro per la verifica degli obiettivi di conservazione e per la realizzazione di programmi di ricerca e di monitoraggio nelle aree protette”, voudrait mettre les bases pour une gestion commune de l’information et pour la mise au point des instruments d’aide à la décision qui permettaient à chaque organisme d’agir localement avec une pondération objective des processus de décision qui aboutissent en actions concrètes dans le territoire. En ce sens on entend actions soit à niveau de gestion, soit action référées à des adresses d’étude et recherche, au complètement des lacunes de connaissance, à la définition de contrôles répétés au fil du temps, à la classification des valeurs en échelles de priorité, aux définitions de vocation de compartiments et planifications écologiques, mais aussi, ne pas dernières par importance, les actions adressées à la divulgation des informations obtenues.

9
LA RETE ECOLOGICA EUROPEA
Dal 1992 l’Unione Europea si è impegnata nella conservazione della biodiversità pro-gettando la realizzazione di una rete di ambienti da tutelare, la cosiddetta Rete Natura 2000. Al fi ne di una corretta individuazione dei siti che costituiranno la rete di aree de-stinate alla conservazione della biodiversità (i Siti di Importanza Comunitaria – SIC, individuati dagli Stati membri), essa ha emanato la Direttiva 92/43/CEE (detta Direttiva ‘Habitat’; D.P.R. 357/97 e s. m. i.), che intende “salvaguardare la biodiversità mediante la conservazione degli habitat naturali, nonché della fl ora e della fauna selvatiche nel territorio europeo degli Stati membri”. La Rete Natura 2000 comprende anche le zone di protezione speciale (ZPS) classifi cate dagli Stati membri a norma della direttiva 79/409/CEE (detta Direttiva ‘Uccelli’, ora aggiornata dalla Direttiva 2009/147/CE; L. 157/92 e s. m. i.), che ha come scopo “la conservazione di tutte le specie di uccelli viventi natural-mente allo stato selvatico nel territorio europeo degli Stati membri”.
Le Direttive Habitat e Uccelli costituiscono attualmente i più importanti strumenti nor-mativi rivolti alla conservazione delle specie animali e vegetali, in quanto, oltre a tutelare gli individui delle specie segnalate, sanciscono contemporaneamente la protezione degli habitat in cui tali specie vivono, assumendo inoltre notevoli risvolti pratici sulla piani-fi cazione e la gestione del territorio. In precedenza, infatti, la tutela legale riguardava esclusivamente gli individui appartenenti ad una data specie (che pertanto non potevano essere raccolti o cacciati), e non gli habitat in cui questi individui vivono; ne risultava il paradosso che era possibile estinguere intere popolazioni di anfi bi o di piante acquatiche bonifi cando un singolo stagno, mentre era vietato raccogliere anche un singolo individuo di pianta o anfi bio protetti.
Le due direttive sono accompagnate da una serie di allegati relativi a specie e tipi di habitat di particolare interesse ai fi ni della loro salvaguardia.
La Direttiva Habitat comprende 7 allegati, dei quali i seguenti interessano la tutela di ambienti e specie:
Allegato I - Tipi di habitat di interesse comunitario la cui conservazione richiede la designazione di aree speciali di conservazione.Allegato II - Specie animali e vegetali d’interesse comunitario la cui conservazione richiede la designazione di zone speciali di conservazione.Allegato IV - Specie animali e vegetali di interesse comunitario che richiedono una protezione rigorosa.La Direttiva Uccelli comprende 5 allegati; per le specie elencate nell’allegato I sono
previste misure speciali di conservazione per quanto riguarda l’habitat, per garantire la sopravvivenza e la riproduzione di dette specie nella loro area di distribuzione.
Purtroppo gli allegati che segnalano gli habitat e le specie oggetto di tutela, per le persone non esperte, rappresentano unicamente un lungo elenco di nomi astrusi. Cercare di rendere comprensibile ad un maggior numero di persone il signifi cato degli elenchi di nomi contenuti negli allegati delle Direttive Habitat e Uccelli è una delle fi nalità che si propone questa pubblicazione, unitamente a quella di divulgare informazioni su alcuni aspetti del patrimonio naturale ancora poco conosciuti, e fornire alle persone interessate uno strumento che permetta loro di acquisire nuove conoscenze sulle specie e sugli am-bienti naturali presenti sul territorio del Parco del Beigua la cui conservazione è conside-rata una delle fi nalità dell’Unione Europea.
LE RÉSEAU ÉCOLOGIQUE EUROPEÉN
Depuis 1992 l’Union Européenne est engagée dans la conservation de la diversité biologique avec l’étude de la réalisation d’un réseau à sauvegarder, soi-disant Rete Natura 2000. Afin d’une correcte individuation des sites qui constitueront le réseau d’aires adressées à la conservation de la diversité biologique (les Sites d’Importance Communautaire – SIC, identifiés par les Etats membres), elle à promulgué la Directive 92/43/CEE (dite Directive ‘Habitat’; D.P.R. 357/97 et s. m. i.), qui s’adresse à « sauvegarder la diversité biologique via la conservation des habitats naturels, ainsi que de la faune et de la flore sauvages dans le territoire Européen des Etats membres ». La Rete Natura 2000 comprends aussi les zones de protection spéciale (ZPS) classifiées par les Etats membres à norme de la Directive 79/409/CEE (dite Directive ‘Oiseaux’, aujourd’hui mise à jour par la Directive 2009/147/CE; L. 157/92 et s. m. i.), qui a comme objectif « la conservation de toutes espèces de oiseaux vivants naturellement à l’état sauvage dans le territoire européen des Etats membres » .Les directives Habitat et Oiseaux constituent actuellement les instruments normatifs les plus importants qui soient destinés à la conservation des espèces animales et végétales, parce que, en plus de protéger les individus des espèces signalisées, établissent au même temps la protection des habitats dont ils vivent, en assumant par ailleurs des effets pratiques significatifs sur la planification et gestion du territoire. Auparavant, en effet, la protection légale, concernait exclusivement les individus appartenant à une particulière espèce (qui donc ne pouvaient pas être chassés ou rassemblées), et ne concernait pas les habitats dont ces individus vivent ; ça résultait en un paradoxe pour lequel était possible éteindre populations entières d’amphibiens ou des plantes aquatiques en bonifiant un single étang, mais il était interdit de cueillir même un single individu de plante ou d’amphibien protégé. Les deux directives sont accompagnées par une pluralité d’annexes concernant espèces et typologies d’habitat de notable intérêt en ce qui concerne leur sauvegarde. La Directive Habitat comprend 7 annexes, dont ceux qui suivent concernent la protection des environnements et espèces: Annexe I – Typologie d’habitat d’intérêt communautaire dont la conservation nécessite la désignation d’aires spéciales de conservation.Annexe II – Espèces animales et végétales d’intérêt communautaire dont la conservation nécessite la désignation de zones spéciales de conservation. Annexe IV – Espèces animales et végétales d’intérêt communautaire qui nécessitent une rigoureuse protection. La Directive Oiseaux comprend 5 annexes; pour les espèces listées dans l’Annexe I sont prévues des mesures spéciales de conservation en ce qui concerne l’habitat, afin de garantir survie et reproduction de ces espèces dans leur aire de distribution. Malheureusement, les annexes qui signalisent les habitats et espèces objectif de la protection, pour des personnes qui ne sont pas expertes, représentent uniquement une longue liste de noms obscurs. Chercher donc de rendre compréhensible à la plus partie des personnes le significat des listes de noms contenus dans les annexes des Directives Habitat et Oiseaux, est une des finalités que la présente publication se propose, ainsi que divulguer des informations sur certains aspects du patrimoine naturel qui sont encore peu connus, et fournir aux personnes intéressées un instrument qui puisse permettre leur d’apprendre nouvelles connaissances sur les espèces et sur les environnements naturels qui sont présents sur le territoire du Parco del Beigua dont la conservation est considérée une des finalités de l’Union Européenne.

10
BREVE GUIDA ALLA LETTURA
Nell’impostazione data alla guida si è cercato di mettere in relazione le specie con gli ambienti da esse preferiti. Va però considerato che tali preferenze non sono sempre ben defi nite, oppure gli ambienti scelti per la riproduzione differiscono da quelli frequentati per la ricerca del cibo (ad es. nel caso dei Chirotteri ed alcune specie di Rapaci); in alcuni di questi casi si è quindi preferito dedicare uno spazio a sé alle specie o ai gruppi di spe-cie in questione, in altri casi le specie sono state invece trattate in relazione alla tipologia ambientale, scelta per la riproduzione e/o la ricerca del cibo.
Lo spazio dedicato ai singoli elementi che caratterizzano la biodiversità del Parco del Beigua, è primariamente in relazione alla quantità di informazioni disponibili per ognuno di essi relativamente all’area di studio, dando comunque maggior rilevanza a quelli di particolare interesse nel contesto della rete ecologica europea. Vengono quindi eviden-ziati gli ambienti (all. I; il segno * indica i tipi di habitat prioritari) e le specie animali (all. II e IV) individuati dalla Direttiva Habitat (DH) e, per quanto riguarda l’avifauna, le specie incluse nell’allegato I della Direttiva Uccelli (DU); oltre a queste, si sono poi trattate anche le specie tipiche delle principali tipologie ambientali e/o individuate quali target dal Parco del Beigua.
In alcuni casi si è poi ritenuto necessario dedicare approfondimenti su singoli aspetti dei temi trattati, o sulle metodologie di indagine adottate.
Nei grafi ci relativi ad alcuni gruppi, vie-ne indicata, in rapporto all’intero territorio interessato dal Parco del Beigua ed ai setto-ri ad esso connessi, la percentuale di unità territoriali (individuate da ‘celle’ di 500 per 500 metri di lato) per le quali sono attual-mente note segnalazioni della singola specie. Questo valore (indicato nei grafi ci come ‘Di-stribuzione (%)’), è ovviamente infl uenza-to dalla tipologia delle indagini sino ad ora condotte per i singoli taxa; va inoltre con-siderato che, talvolta, le specie più comuni possono essere state penalizzate, in quanto, la catalogazione e l’archiviazione di tutte le osservazioni effettuate nel corso delle attivi-tà di campo è stata effettuata solo a partire dai tempi più recenti.
I testi evidenziati con il carattere blu de-scrivono una specifi ca fi gura (ad es., ‘[a]’), identificata da una lettera riportata solitamen-te nel suo angolo superiore sinistro.
Le mappe possono raffi gurate l’arealedell’elemento considerato (generalmente cal-colato in base ai risultati ottenuti con l’analisi dei vicini più prossimi; Newton 1976) [a] oppure indicazioni sulla sua abbondanza o im-portanza (resi graficamente con tonalità di colore crescenti dal giallo al rosso in relazione all’incremento dei valori rappresentati) [b].
[a]
[b]
PÈTITE GUIDE À LA LECTURE
Dans le cadre fourni à la guide, on a cherché de mettre en liaison les espèces avec les environnements par eux préférés. Toutefois, il faut considérer que dites préférences ne sont pas toujours bien définies, ou que les environnements choisis pour la reproduction diffèrents par ces qui sont fréquentés lors de la recherche de la nourriture (par exemple dans le cas des Chiroptères et de quelque espèce de oiseau de proie); en certains de ces cases, on a donc préféré dédier un espace en soi aux espèces ou aux groupes des espèces en question, en autres cases les espèces ont étés au contraire traitées en relation à la typologie d’environnement, choisie pour la reproduction et/ou la recherche de nourriture. L’espace dédié aux éléments individuels qui caractérisent la biodiversité du Parco del Beigua, est primairement en relation à la quantité d’informations disponibles pour chacun d’eux en relation à l’aire objectif d’étude, en donnant toujours une plus grande relevance à ceux qui ont particulier intérêt dans le contexte du réseau écologique Européen. On marque donc les environnements (ann. I; le marque * montre typologies d’habitat prioritaires) et les espèces animales (ann. II et IV) individuées par la Directive (DH) et, en ce qui concerne les oiseaux, les espèces incluses dans l’Annexe I de la Directive Oiseaux (DU); en plus de ces-ci, sont aussi traitées les espèces typiques des principales typologies d’environnement et/ou individuées comme objectif par le Parco del Beigua. En certains cases, on à puis retenu nécessaire de dédier des recherches ultérieures sur certains aspects ou thèmes traités, ou sur les méthodologies d’enquête adoptées. Dans les graphiques concernant certains groupes, il est indiqué, en rapport à l’entier territoire intéressé par le Parco del Beigua et aux secteurs à lui annexés, le pourcentage d’unités territoriales (individuées par cellules de 500x500 mètres en coté) pour lesquelles sont actuellement connues des signalisations de la single espèce. Cette valeur (indiquée dans les graphiques comme Distribution (%)’), est bien sûr influencée par la typologie des enquêtes jusqu’à aujourd’hui conduites par les taxa; if faut aussi considérer que, quelque fois, les espèces plus communes peuvent avoir étés pénalisées, parce que le catalogage et la mise en archive des toutes observations effectuées pendant l’activité sur le champ, ont été effectués seulement dans le temps le plus récent. Les textes marqués par un caractère bleu décrivent une spécifique figure (par ex., ‘[a]’), identifiée par une lettre montrée typiquement dans l’angle supérieur gauche. Les plans peuvent représenter l’aire de l’élément considéré (généralement calculé sur les résultats obtenus par l’analyse des voisins les plus prochains; Newton 1976) [a] ou indications sur sa abondance ou importance (rendus graphiquement avec tonalités de couleur croissantes à partir du jaune vers le rouge en relation à l’incrément des valeurs représentées) [b].

11
Avifauna
Le informazioni riguardanti la distribuzione, la riproduzione, le caratteristiche migra-torie ed ecologiche, lo status europeo ed italiano e gli andamenti delle popolazioni sono stati desunti principalmente dai lavori di Cramp et al. (1977-1994), BirdLife Internatio-nal (2004), Brichetti & Fracasso (2003-2011), Fornasari et al. (2002b, 2010), Hagemeijer e Blair (1997), Gariboldi & Ambrogio (2006), Meschini e Frugis (1993), Rete Rurale Nazionale & LIPU (2011), Rossi (2012) e Spina & Volponi (2008a, 2008b). L’ordine sistematico e la nomenclatura utilizzate sono quelle indicate da Fracasso et al. (2009). Inoltre, per una valutazione della situazione regionale e locale, si sono consultati princi-palmente gli atlanti degli uccelli nidifi canti e svernanti in Liguria (Aa. Vv. 1989, Spanò et al. 1998), la check-list regionale (Baghino et al. 2012), la Guida agli uccelli della ZPS “Beigua-Turchino” e del Parco del Beigua (Galli et al. 2006) e le relazioni tecniche re-lative agli studi condotti dall’Ente Parco del Beigua. Nei testi a seguire, al fi ne di evitare un’eccessiva frammentazione della lettura, la citazione dei suddetti lavori è stata omessa.
Nelle schede delle singole specie, in relazione allo stato di conservazione ed all’inte-resse europeo sono stati associati ai seguenti simboli:
Specie incluse nell’allegato I della Direttiva Uccelli (DU).
Le specie indicate con numero da 1 a 3 sono quelle la cui conservazione risulta di parti-colare importanza per l’Europa (BirdLife International 2004). La priorità decresce da 1 a 3 secondo il seguente schema: Categoria 1 - specie globalmente minacciate e quindi di particolare importanza conserva-zionistica a livello globale.Categoria 2 - specie che non hanno uno stato di conservazione favorevole e le cui popo-lazioni sono concentrate in Europa.Categoria 3 - specie che non hanno uno stato di conservazione favorevole in Europa, ma le cui popolazioni non sono concentrate in Europa.Le specie contrassegnate dal simbolo ‘*’ presentano popolazioni o areali concentrati in Europa e sono caratterizzate da un favorevole stato di conservazione.
Categoria di minaccia attribuita dalla Lista Rossa italiana (Peronace et al. 2012) secondo il seguente schema: in pericolo critico (CR), in pericolo (EN), vulnerabile (VU), quasi minacciato (NT), carente di dati (DD), a minor preoccupazione (LC).
Sono state inoltre adottate le seguenti abbreviazioni: ind. = individuo/i; cp = coppia/e; PC = punto d’ascolto.
Oiseaux
Les informations concernant la distribution, la reproduction, les caractéristiques migratoires et écologiques, le status europèen ou italien et les changements de population ont étés déduits principalement par les travaux de Cramp et al. (1977-1994), BirdLife International (2004), Brichetti & Fracasso (2003-2011), Fornasari et al. (2002b, 2010), Hagemeijer & Blair (1997), Gariboldi & Ambrogio (2006), Meschini & Frugis (1993), Rete Rurale Nazionale & LIPU (2011), Rossi (2012) et Spina & Volponi (2008a, 2008b). L’ordre systématique et la nomenclature qui ont étés utilisées sont celles indiquées par Fracasso et al. (2009). En outre, pour une évaluation de la situation régionale et locale, on a consulté principalement les atlas des oiseaux nidifiants et qui passent l’hiver en Ligurie (Aa. Vv. 1989, Spanò et al. 1998), la check-list régionale (Baghino et al. 2012), la Guide aux oiseaux de la ZPS “Beigua-Turchino” et du Parco del Beigua (Galli et al. 2006) et les relations techniques concernant les études effectués par Ente Parco del Beigua. Dans les textes à suivre, afin d’éviter une excessive fragmentation de la lecture, la citation de dites travaux a été omise.
Dans les fichiers concernant chacune espèce, en relation à l’état de conservation et à l’interet européen ont étés associés les symboles suivants :
Espèces incluses dans l’Annexe I de la Directive Oiseaux (DU).
Les espèces indiquées avec le numéro de 1 à 3 sont celles dont la conservation résulte ayant particulière importance pour l’Europe (BirdLife International 2004). La priorité décroît à partir de 1 à 3 selon le schéma qui suit: Catégorie 1 – espèces globalement menacés et donc de particulière importance de conservation à niveau global. Catégorie 2 – espèces qui n’ont pas un état de conservation favorable et dont les populations sont concentrées en Europe. Catégorie 3 – espèces qui n’ont pas un état de conservation favorable en Europe, mais dont les populations ne sont pas concentrées en Europe. Les espèces marquées par le symbole ‘*’ indiquent des populations ou aires concentrées en Europe et sont caractérisées par un favorable état de conservation.
Catégorie de menace attribuée par la Liste Rouge Italienne (Peronace et al. 2012) selon le schéma qui suit: en danger critique (CR), en danger (EN), vulnérable (VU), quasi menacé (NT), pauvre de données (DD), a moindre préoccupation (LC).
En outre, les suivantes abréviations ont étés adoptées: ind. = individu/s; cp = couple/s; PC = point d’écoute.

12
IL PROGETTO DI “INDAGINI AMBIENTALI E DIVULGAZIONE DEI
RISULTATI DELLE ATTIVITÀ DI RICERCA CONDOTTE NEL PARCO
DEL BEIGUA E NEI SETTORI DELLA RETE NATURA 2000
FUNZIONALMENTE CONNESSI”
Finanziato dal primo bando dei Progetti Strategici del Programma Operativo Italia-Francia “Marittimo”, nel quadro dell’Asse III “Ambiente e produzioni rurali e marine”, il Progetto di Cooperazione delle Reti Ecologiche del Mediterraneo (Co.R.E.M.) vede la partecipazione di un ampio partenariato, di cui è capofi la l’Assessorato della Difesa dell’Ambiente della Regione Autonoma della Sardegna, composto dalle principali am-ministrazioni pubbliche locali, da primarie agenzie di protezione ambientale, importanti parchi nazionali e regionali ed aree marine protette, unitamente a prestigiose università e centri di ricerca. Il Progetto Co.R.E.M. si propone di tutelare e valorizzare il patrimonio naturalistico e la biodiversità della Rete Ecologica, mirando a ridurre la pressione e le minacce sulle risorse ambientali e a favorire una fruizione sociale ed economica soste-nibile delle stesse, grazie al coinvolgimento ed alla sensibilizzazione dei cittadini e delle imprese.
Il Parco del Beigua è coinvolto nell’Azione di Sistema H “Disseminazione del Model-lo Natura 2000”, in partnership con il Parco di Portofi no, il Parco della Maremma e la Provincia di Oristano. Le azioni mirano alla valorizzazione delle risorse naturali (anche in forme congiunte alle risorse culturali e in funzione di un loro miglioramento e qua-lifi cazione) e a promuovere la disseminazione del modello Rete Natura 2000 e dei suoi contenuti a livello transfrontaliero, per favorire una comune capacità di governance per la tutela della biodiversità e la gestione delle aree naturali della Rete Natura 2000, anche in relazione organica con le altre forme di tutela operanti sul territorio.
Gli obiettivi del progetto si possono quindi così riassumere:• valorizzare le risorse naturali attraverso la disseminazione del modello Natura 2000 a
livello transfrontaliero;• favorire forme di governance orientate alla tutela della biodiversità ed alla gestione
delle aree naturali della Rete Natura 2000, in coerenza rispetto alle altre forme di tutela operanti sul territorio;
• promuovere la salvaguardia e la sensibilità ambientale;• favorire la valorizzazione economica e la fruibilità sostenibile delle aree natura-
li, attraverso una gestione partecipativa che coinvolga, in particolare, gli operatori dell’agricoltura, della pesca, del turismo e dei servizi ambientali;
tali obiettivi si potranno raggiungere realizzando i seguenti interventi:• campagne di comunicazione;• eventi informativi per operatori turistici;• eventi culturali e pubblici seminari per cittadini e fruitori;• Piano di Azione per la promozione della comunicazione e della partecipazione degli
stakeholders (disseminazione modello Natura 2000 in AL21).
LE PROJET DES “ INDAGINI AMBIENTALI E DIVULGAZIONE DEI RISULTATI DELLE ATTIVITÀ DI RICERCA CONDOTTE NEL PARCO
DEL BEIGUA E NEI SETTORI DELLA RETE NATURA 2000 FUNZIONALMENTE CONNESSI”
Financé par le premier avis de Projets Strategiques du Programme Operatif Italie France “Maritime”, dans le cadre de l’Axe III “Environnement et productions rurales et marines ”, le projet de Coopération des Réseaux Écologiques dans la Méditerranée (Co.R.E.M.) concerne la participation d’un grand partenariat, dont la tête est l’Assessorato della Difesa dell’Ambiente de la Regione Autonoma della Sardegna, constitué par les principales administrations publiques locales, par des primaires agences de protection de l’environnement, importants parcs nationaux et régionaux et aires maritimes protégées, ainsi que prestigieuses universités et centres de recherche. Le projet Co.R.E.M. se propose de protéger et valoriser le patrimoine naturel et la biodiversité du Réseau Ecologique, en s’adressant à réduire la pression et les menaces sur les ressources de l’environnement et à favoriser une fruition sociale et économique soutenable de ces dernières, grâce à l’engagement et à la sensibilisation des citoyens et entreprises. Le Parco del Beigua est impliqué dans l’Action de Système H “Dissémination du Modèle Natura 2000 ”, en partnership avec le Parco di Portofino, le Parco della Maremma et la Provincia di Oristano. Les actions sont adressées à la valorisation des ressources naturelles (ainsi en formes conjointes aux ressources culturales et en fonction d’une leur amélioration et qualification) et à promouvoir une dissémination du modèle Rete Natura 2000 et des ses contenus à niveau transfrontalier, pour favoriser une capacité commune de governance pour la protection de la biodiversité et la gestion des aires naturelles de la Rete Natura 2000, aussi en relation organique avec des autres formes de protection qui opèrent sur le territoire. Les objectifs du projet se résument donc comme suit: • valoriser les ressources naturelles avec la dissémination du modèle Natura 2000 à niveau transfrontalier ; • favoriser les formes de governance orientées à la protection de la biodiversité et à la gestion des aires naturelles de la Rete Natura 2000, en cohérence par rapport aux autres formes de protection qui opèrent sur le territoire; • promouvoir la sauvegarde et sensibilité pour l’environnement; • favoriser la valorisation économique et l’accessibilité soutenable des aires naturelles, avec une gestion participative qui engage, notamment, les opérateurs agricoles, de la pêche, du tourisme et des services d’environnement; Ces objectifs pourront être achevés en réalisant ces interventions: • campagnes de communication; • évents informatifs pour les opérateurs touristiques; • évents culturales et séminaires publiques pour citoyens et utilisateurs ; • Plan d’Action pour promouvoir la communication et la participation des stakeholders (dissemination du modèle Natura 2000 en AL21).

13
È quindi in questo contesto che il Parco del Beigua ha commissionato un’indagine volta all’acquisizione di informazioni scientifi che relative alla distribuzione delle specie e degli habitat più signifi cativi, con lo scopo di attivare effi caci canali di informazione e di comunicazione sia verso gli operatori della vigilanza (Corpo Forestale dello Stato e Polizia Provinciale di Genova e Savona), sia verso il più vasto pubblico di appassionati e visitatori dell’area protetta, stimolando ed indirizzando le energie nel modo più utile per la messa a punto delle strategie di conservazione di volta in volta migliori. Le azioni previste da questa indagine si possono così riassumere:• Azione 1: Analisi preliminare dei materiali già disponibili, volta all’individuazione
di aree campione e specie target sulle quali condurre sperimentazioni in merito alle strategie ed alle tecniche di disseminazione dei dati naturalistici, producendo un re-port che identifi chi le aree campione da sottoporre ad indagine.
• Azione 2: Conduzione di indagini pilota per la raccolta di dati e informazioni sulle specie target fi nalizzate alla conservazione della biodiversità e funzionali all’attività di disseminazione / informazione. Nel corso del 2012, sono state quindi condotte indagini sperimentali fi nalizzate alla conservazione della biodiversità e funzionali all’attività di divulgazione, e che, in sintesi, prevedono l’acquisizione di informa-zioni sulla presenza del Lupo mediante la tecnica del trappolaggio fotografi co (ad integrazione ed approfondimento di quanto si sta già ottenendo con il progetto re-gionale “Il Lupo in Liguria”), e censimenti mirati a colmare lacune conoscitive su uccelli, anfi bi, rettili, libellule ed alcune specie di farfalle (Euplagia quadripunctaria,Euphydryas (aurinia) provincialis e Zerynthia polyxena).
• Azione 3: Organizzazione e predisposizione di materiali informativi mediante la mes-sa a punto di format fi nalizzati alla disseminazione / divulgazione / comunicazione dei risultati ottenuti con le attività di ricerca e monitoraggio condotte, ed utilizzabili su prodotti cartacei e/o supporti informatici.
Per l’attuazione di queste azioni si è quindi adottata un’impostazione della gestione dell’informazione che considera tre tappe principali:1. Defi nire il punto della situazione relativamente alle conoscenze acquisite fi no ad ora
(CONOSCERE);2. Codifi care l’informazione raccolta in un sistema di gestione gerarchizzato, mediante
strumenti oggettivi (DARE SIGNIFICATO ALL’INFORMAZIONE);3. Defi nire delle priorità di interesse e di azione ottimizzando e “personalizzando” le
possibilità di intervento (CONSERVARE).Al completamento del processo conoscitivo, l’integrazione delle informazioni dispo-
nibili permetterà quindi di istituire strumenti di aiuto alla decisione che rappresenteranno il mezzo base per l’organizzazione, il controllo e la divulgazione del continuo fl usso di informazione.
Il est donc en ce contexte que le Parco del Beigua a commissionné une enquête pour l’acquisition d’informations scientifiques concernant la distribution des espèces et des habitats les plus significatifs, au but d’activer chaînes efficaces pour l’information et la communication soit vers les opérateurs de vigilance (Corpo Forestale dello Stato et Polizia Provinciale de Genova et Savona), soit vers le publique le plus vaste de passionnés et visiteurs de l’aire protégée, en stimulant et adressant les énergies avec la façon la plus utile pour la mise au point des stratégies de conservation chaque fois améliorées. Les action prévues par cette enquête peuvent être résumées comme suit: • Action 1: Analyse préliminaire des données déjà disponibles, adressée à l’individuation de aires objet et espèces target sur lesquelles effectuer des expérimentations en ce qui concerne les stratégies et techniques de dissémination des données naturelles, en produisant en suite un report qui identifie les aires objet à soumettre à l’enquête. • Action 2: Conduire des enquêtes-pilote pour rassembler des données et informations sur les espèces cible (target) adressées à la conservation de la biodiversité et qui soient fonctionnelles à l’activité de dissémination et information. Pendant l’année 2012, ont donc étés effectuées des enquêtes expérimentales finalisées à la conservation de la biodiversité et fonctionnelles à l’activité de divulgation, et que, en résumé, nécessitent l’acquisition d’information sur la présence du Loup avec la technique des trappes photographiques (comme intégration et renseignement ultérieur de ce qui est déjà en cours d’obtention avec le projet régional “Il Lupo in Liguria”), et recensements adressés à remplir les lacunes de connaissance sur les oiseaux, amphibiens, reptiles, libellules et certaines espèces de papillons (Euplagia quadripunctaria,Euphydryas (aurinia) provincialis et Zerynthia polyxena).• Action 3: Organisation et prédisposition de documents d’information avec la mise au point de format de dissémination / divulgation / communication des résultats obtenus avec les activités de recherche et contrôle effectuées, et qui soient utilisables sur des produits en papier et/ou produits informatiques. Pour l’actuation de ces actions, on a adopté une structure de gestion de l’information qui considère trois étapes principales: 1. Définir la situation actuelle en relation aux connaissances apprises jusqu’à aujourd’hui (CONNAITRE);2. Codifier l’information rassemblée dans un système de gestion hiérarchique, au moyen d’instruments objectifs (DONNER UN SIGNIFICAT À L’INFORMATION);3. Définir les priorités d’intérêt et d’action en optimisant et en “personnalisant” les possibilités d’intervention (CONSERVATION).Au complètement du processus d’apprentissage, l’intégration des informations disponibles permettra donc la création d’instruments d’aide à la décision qui représenteront le moyen de base pour l’organisation, le contrôle et la divulgation du flux continu des informations.

14 IL PARCO DEL BEIGUA14
IL PARCO DEL BEIGUA
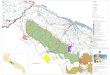
Il Parco Naturale Regionale del Beigua copre una superfi cie di 8.715 ettari che si estende su parte del territorio provinciale di Savona e Genova; ad esso sono connesse cinque aree della Rete Natura 2000: i SIC IT1331402 Beigua – Monte Dente – Gargassa – Pavaglione (16.922 ettari), IT1321313 Foresta della Deiva – Torrente Erro (886 ettari), IT1330620 Pian della Badia (249 ettari), IT1331501 Praglia – Pracaban – Monte Leco – Punta Martin (6.958 ettari) e la ZPS IT1331578 Beigua – Turchino (9.914 ettari) [a].Complessivamente, l’insieme delle Aree Protette e della Rete Natura 2000, che risultano tra loro in gran parte sovrapposte, si estendono per circa 19.479 ettari ed interessano 14 Comuni. Di seguito, tenendo anche conto del fatto che animali e piante non rispettano le delimitazioni ‘umane’, l’indicazione ‘Parco del Beigua’ (o ‘Beigua’) indicherà l’insieme dei settori compresi tra la Foresta della Deiva (ad Ovest), Punta Martin (ad Est), la Val Gargassa (a Nord) e la linea di costa del Mar Mediterraneo (a Sud).
Il gruppo montuoso del Beigua è formato da una successione di rilievi rappresentati da una dorsale di circa 26 chilometri, che si sviluppa dal Colle del Giovo al Passo del Turchino con andamento parallelo alla costa, ed una diramazione che dal Monte Dente si dirige per un’altra decina di chilometri verso Nord sino alla Val Gargassa, con quote massime comprese tra i 1.000 ed i 1.300 metri s.l.m..
Anche se l’elevazione è, tutto sommato, modesta, visto dal mare il complesso montuo-so si presenta come una barriera di una certa imponenza la cui linea di cresta dista non più di cinque o sei chilometri dal litorale. In effetti tale dorsale montuosa presenta una notevole asimmetria tra il versante tirrenico, particolarmente acclive, e quello padano, dalla pendenza assai meno accentuata e contrassegnato da lunghe vallate.
Zone umide Mosaici agrari
Zone edificate
Boschidi latifoglie
Corsid’acqua
Boschi mistidi conifere e latifoglie
LE PARCO DEL BEIGUA
Le Parco Naturale Regionale del Beigua couvre une surface de 8.715 hectares qui étends sur partie du territoire provincial de Savona et Genova; à celui-ci sont connectées cinq aires de la Rete Natura 2000 : les SIC IT1331402 Beigua – Monte Dente – Gargassa – Pavaglione (16.922 hectares), IT1321313 Foresta della Deiva – Torrente Erro (886 hectares), IT1330620 Pian della Badia (249 hectares), IT1331501 Praglia – Pracaban – Monte Leco – Punta Martin (6.958 hectares) et la ZPS IT1331578 Beigua – Turchino (9.914 hectares) [a]. Globalement, l’ensemble des Aires Protégées et de la Rete Natura 2000, qui résultent entre eux substantiellement superposées, étendent pour environ 19.479 hectares et concernent 14 Municipalités. Ci-dessous, en comptant que les animaux et plantes ne respectent pas les délimitations humaines, l’indication ‘Parco del Beigua’ (ou ‘Beigua’) indiquera l’ensemble des secteurs comprises entre la Foresta della Deiva (à Ouest), Punta Martin (à Est), la Val Gargassa (à Nord) et la ligne costière de la Mer Méditerranée (à Sud). Le groupe de montagnes du Beigua comprend une succession de reliefs qui identifie une dorsale d’environ 26 kilomètres, qui se développe à partir du Colle del Giovo jusqu’au Passo del Turchino avec une évolution parallèle à la côte, et une diramation que à partir de Monte Dente dirige pour une autre dizaine de kilomètres à Nord, jusqu’ù la Val Gargassa, avec des hauteurs maximales comprises entre 1.000 et 1.300 mètres au dessus du niveau de la mer. Bien que l’hauteur soit tout à fait modeste, lors que observé par la mer, le complexe montagneux se présente comme une barrière d’une certaine importance dont la ligne de crête est non plus que cinq ou six kilomètre loin de littoral. En effet, cette dorsale montagneuse présente une significative asymétrie entre le versant du Tyrrhénienne, particulièrement en pente, et celui padane, dont la pente est beaucoup moins accentuée et qui est contresigné par des longues vallées.

15
PraterieAree
rocciose Arbusteti
Boschidi conifere
Macchia mediterranea
Allegato I: n° di habitat
Allegati II/IV: n° di specie animali e vegetali
Direttiva Uccelli Allegato I: n° di specie
Direttiva Habitat
In basso: sezione semplifi cata del Parco del Beigua, nella quale vengono visualizzate le principali tipologie ambientali e la loro importanza conservazionistica, defi nita in base al numero di ambienti e/o specie ad esse legati che siano inclusi negli allegati delle direttive Habitat e Uccelli.
�
�
�
�
�
�
�
�
�
�
�
�
�
�
�
�
�
�
�
�
�
�
�
��
�
�
���������
������� ���
�������
��������
�����
�� �����
��������� �������
������ �� ��
�������
������
�������
�����
������� ���
����� �����
�� �����
��������
���������
��� �!���
������"��
����������#$"��
������#�
������
�������
������%���
���
�&''&()*
�&''&+)&
�&''),*)
�&'*&'&'
� �-��� �&''&+./�0�� �1����%��������#���0�� �
[a] En bas : section simplifiée du Parco del Beigua, dont ils sont représentées les principales typologies d’environnement et leur importance de conservation, définie en accord au numéro d’environnements et/ou espèces à leur reliées qui soient incluses dans les annexes des directives Habitat et Oiseaux.

16 IL PARCO DEL BEIGUA
Aree edificate
Mosaici agrari0,8%
Boschi di latifoglie48,1%
Boschi misti di conifere e latifoglie
10,8%
0,1%Boschi di conifere
9,4%
Praterie13,6%Arbusteti
12,4%
Aree rocciose o con vegetazione rada
4,2%
Zone umide0,1%
Corsi d'acqua0,5%
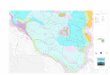
Le aree protette del Beigua sono caratterizzate da ampie superfi ci boscate, aree a pascolo naturale e praterie, arbusteti (brughiere, cespuglieti, macchia mediterranea) ed aree rocciose o con vegetazione rada. Anche se meno rappresentate in quanto a superfi cie, zone umide, corsi d’acqua e mosaici agrari rivestono però notevole importanza per ciò che concerne biodiversità e valore conservazionistico [a, b].
[a]
[b]
Les aires protégées du Beigua sont caractérisées par amples surfaces de bois, aires à pâturage naturel et prairies, arbrisseaux (landes et broussailles, forêts de la méditerranée) et aires rocheuses ou avec peu de végétation. Bien que moins représentées en ce qui concerne le montant de surface, les zones humides, les courses d’eau et les mosaïques agraires revêtent toutefois une significative importance en ce qui concerne la biodiversité et la valeur de conservation [a, b].

17
Decisamente marcate sono le differenze climatiche tra i due versanti, con inverni miti ma talora contrassegnati da violenti rovesci e lunghe estati siccitose quello tirrenico, con estati calde ed inverni anche assai freddi quello padano (con temperature medie del mese più freddo prossime ai -2 °C e prolungata permanenza al suolo del manto nevoso).
L’area del Beigua è quindi caratterizzata da evidenti contrasti fra i versanti marittimi e quelli settentrionali; le quote relativamente elevate e la presenza di microclimi freddi consentono la vita di specie boreali in vicinanza del Mar Mediterraneo, ed il substrato, per gran parte ofi olitico, condiziona la fl ora offrendo opportunità di rifugio ad interes-santi serpentinofi te. Considerando gli elementi contemplati negli allegati della direttiva Habitat troviamo ben 31 tipologie ambientali censite come habitat dell’allegato I e che interessano gran parte dell’area [b], tra le specie vegetali incluse negli allegati II o IV, sono da segnalare Gladiolus palustris e Spiranthes aestivalis; di notevole importanza è poi la presenza di specie vegetali endemiche, ad elevato valore fi togeografico o mi-nacciate di scomparsa (Anagallis tenella, Cerastium utriense, Cheilanthes marantae,Crocus ligusticus, Minuartia laricifolia ssp. ophiolitica, Ophrys sp., Osmunda regalis,Robertia taraxacoides, Viola bertolonii). Altrettanto importanti sono le specie animali, 43 di esse sono incluse negli allegati II o IV della direttiva ‘Habitat’[a], due delle quali (Callimorpha quadripunctaria e Lupo) sono di interesse prioritario; numerose altre sono poi rilevanti dal punto di vista scientifico o conservazionistico (Cicindela maroccanapseudomaroccana, Carabus italicus italicus, Carabus vagans, Carabus solieri ligura-nus, Nebria tibialis tibialis, Haptoderus apenninus, Philorhizus liguricus). L’avifauna che caratterizza l’area risulta poi particolarmente importante, sia in periodo riproduttivo che durante le migrazioni; delle 204 specie di uccelli attualmente censite ben 51 sono infatti incluse nell’allegato I della Direttiva Uccelli e 13 di esse nidifi cano nell’area [a].
12141618
Numero di specie incluse in all. II/IV Direttiva Habitat e all. I Direttiva Uccelli
02468
10
Piante Invertebrati Pesci Anfibi Rettili Uccelli MammiferiPiante Invertebrati Pesci Anfibi Rettili Uccelli (nidificanti)
Mammiferi
[a]
[b]Distribuzione delle tipologie ambientali incluse nell’all. I della Direttiva Habitat
Bien marquées sont les différences de climat entre les deux versants, avec des hivers doux mais quelquefois contresignés par des averses violentes et longues étés séchés le versant Tyrrhénien, et avec des chaudes étés et très froid hivers le versant padan (avec températures moyennes du mois le plus froid prochaines à -2 °C et ayant une permanence prolongée du manteau neigeux au sol). L’aire du Beigua est donc caractérisée par des évidents contrastes entre les versants maritimes et du nord ; les hauteurs modestement élevées et la présence de microclimats froids permettent la vie d’espèces boréales, proche de la mer Méditerranée et le substrat, pour sa majorité ophiolitique, conditionne la fleure en offrant opportunités de refuge pour des intéressants serpentinephiles. En considération des éléments contemplés dans les annexes de la Directive Habitat nous retrouvons bien 31 typologies d’environnement comme habitats de l’Annexe I et qui intéressent la plus grande partie de l’aire [b], parmi les espèces végétales incluses dans les annexes II ou IV, sont à mentionner Gladiolus palustris et Spiranthes aestivalis; de grande importance est en outre la présence d’espèces végétales endémiques, avec grande valeur phytogéographique ou à risque de disparition (Anagallis tenella, Cerastium utriense, Cheilanthes marantae, Crocus ligusticus,Minuartia laricifolia ssp. ophiolitica, Ophrys sp., Osmunda regalis, Robertia taraxacoides, Viola bertolonii). Autant importantes sont les espèces animales, dont 43 sont incluses dans les annexes II ou IV de la Directive ‘Habitat’[a], dont deux (Callimorpha quadripunctaria et Loup) sont d’intérêt prioritaire; plusieurs d’autres sont puis relevantes par le point de vue scientifique ou de conservation (Cicindela maroccana pseudomaroccana, Carabus italicus italicus, Carabus vagans, Carabus solieri liguranus, Nebria tibialis tibialis, Haptoderus apenninus, Philorhizus liguricus). Les oiseaux qui caractérisent l’aire sont particulièrement importants, soit pendant la période de reproduction, soit pendant les migrations; des 204 espèces de oiseaux actuellement recensées, même 51 sono tout à fait incluses dans l’Annexe I de la Directive Oiseaux, dont 13 d’eux nidifient dans l’aire [a].

18
Funghi; 150Anfibi; 9
Rettili; 14
Uccelli; 204 Mammiferi; 35
Numero di specie
Briofite; 1
Pteridofite; 8
Spermatofite; 173Invertebrati; 198
Pesci; 12
Anfibi; 9
p
Ambienti24,3%
Uccelli
Mammiferi6,9%
Percentuale di dati in archivio
Funghi0,4%
Briofite0,0%
Pteridofite0,2%
S t fit
60,3%
0,2%Spermatofite
3,7%Invertebrati2,0%
Pesci0,4%
Anfibi1,1%
Rettili0,6%
[a]
[b]
ATTIVITÀ DI RICERCA, CENSIMENTO E MONITORAGGIO
ATTIVITÀ DI RICERCA, CENSIMENTOE MONITORAGGIO
Negli ultimi anni si è sempre più affermata la necessità di svolgere attività di controllo con le quali verifi care lo “stato di salute” di specie ed habitat meritevoli di tutela e po-tenzialmente minacciati da un’ampia gamma di fattori limitanti, sia di origine umana che naturali. Tale fi ne è perseguibile attuando attività di monitoraggio, le quali permettono di documentare lo stato di popolazioni e specie, fornendo le informazioni di base necessarie per identifi care diffi coltà, collegarle a fattori ad impatto negativo e, ove possibile, impo-stare iniziative di rimedio.
Stante la necessità di conseguire dati aggiornati, funzionali ad ottenere un soddisfa-cente stato delle conoscenze fi nalizzate alla gestione della Rete Natura 2000, si deve riconoscere che il livello qualitativo e quantitativo delle informazioni disponibili solo in pochi casi si può defi nire suffi ciente e adeguato. È quindi indispensabile acquisire tutta una serie di elementi conoscitivi e/o mantenere il loro continuo aggiornamento annua-le tramite rilievi atti a monitorare le effettive presenze, effettuando censimenti condotti su base pluriennale con metodologie e protocolli standardizzati (indispensabili per la ripetibilità ed il confronto con altri studi). Gli studi volti, ad esempio, al monitoraggio dell’avifauna, individuata dalla Commissione Europea quale indicatore di biodiversità ed integrità degli ecosistemi, possono quindi servire a proteggere, gestire, regolare ed eventualmente sfruttare queste risorse.
Le attività di ricerca naturalistica condotte nel Parco del Beigua, hanno spaziato, nel corso degli anni, in numerosi ambiti, indagando dalle piante agli artropodi e dai funghi ai pipistrelli, portando anche alla pubblicazione di guide tematiche (Uccelli, Galli et al.2006; Chirotteri, Calvini 2006; Funghi, Zotti et al. 2007).
Particolare attenzione si è riservata, in particolare dal 2006 in avanti, al monitoraggio degli uccelli, sia nidifi canti che migratori. Con il 2012 si sono poi colmate alcune lacune conoscitive sulla distribuzione e/o consistenza delle popolazioni di libellule, alcune spe-cie di farfalle, anfi bi, rettili e Lupo.
L’organizzazione di tutte le informazioni ambientali, botaniche e faunistiche dispo-nibili al gennaio 2013, ha portato alla realizzazione di una banca dati che, al termine di questa fase, era costituita da oltre 41.000 record relativi a più di 800 specie animali e vegetali [a, b].
ACTIVITÉS DE RECHERCHE, RECENSEMENT ET CONTRÔLE
Dans les dernières années s’est toujours plus affirmée la nécessité d’effectuer des activités de contrôle avec lesquelles on puisse vérifier l’ « état de santé » des espèces et des habitats qui méritent la protection et qui sont potentiellement menacés par une vaste gamme de facteurs de limitation, soient eux d’origine humaine que naturelle. Ce but peut être poursuit en effectuant des activités de contrôle, qui permettent de documenter l’état des populations et des espèces, et fournissant les informations de base nécessaires pour identifier les difficultés, les connecter à des facteurs à impact négatif et, où possible, exécuter des initiatives de remède. Vue la nécessité d’achever des données mises à jour, qui soient fonctionnelles pour obtenir un satisfaisant état des connaissances finalisées à la gestion de la Rete Natura 2000, il faut reconnaître que le niveau qualitatif et quantitatif des informations disponibles seulement sur peu cases peut être défini comme suffisant et adéquat. Il est donc indispensable acquérir une pluralité des éléments de connaissance et/ou maintenir leur mise à jour continue annuelle avec des relèves aptes à contrôler les effectives présences, en effectuant des recensions sur base pluriannuelle avec des méthodologies et protocoles standardisées (indispensables pour la répétabilité et la confrontation avec des autres études). Les études adressées, par exemple, au monitorage des oiseaux, individués par la Commission Européenne comme indicateur de biodiversité et d’intégrité des écosystèmes, peuvent donc servir à protéger, gérer, réguler et éventuellement exploiter ces ressources. Les activités de recherche naturelle effectuées dans le Parco del Beigua, ont variait, au fil des années, en plusieurs secteurs, avec le contrôle des plantes, des arthropodes, et à partir des champignons jusqu’à les chauves-souris, pour arriver à la parution des guides thématiques (Uccelli, Galli et al. 2006; Chirotteri, Calvini 2006; Funghi, Zotti et al. 2007). Une attention particulière est réservée, notamment à partir de 2006 en avant, au monitorage des oiseaux, soient eux nidifiants ou migratoires. Avec le 2012 certaines lacunes de connaissance sur la distribution et/ou la composition des populations des libellules, et des certaines espèces de papillons, amphibiens, reptiles et loups ont étés remplies. L’organisation de toutes informations d’environnement, botanique et faunistique disponibles en janvier 2013, a emmené à la réalisation d’une banque de données qui, au terme de cette étape, était constituée de plus des records concernant plus de 800 espèces animales et végétales [a, b].

19
L’analisi delle informazioni raccolte evi-denzia inoltre come, anche in un’area ben caratterizzata come quella del Parco del Bei-gua, indagini integrative possano portare a notevoli incrementi qualitativi e quantitativi delle conoscenze disponibili. Possiamo infatti osservare come, per tutti i gruppi animali og-getto di azioni specifiche nel corso del 2012 (comprendenti sia censimenti che reperimen-to di informazioni pregresse), il contributo conoscitivo sia stato notevole [a], con signifi -cativi incrementi all’archivio dei dati disponi-bili. Anche per gli uccelli, già oggetto di mo-nitoraggi pluriennali, la costante ripetizione dei censimenti ha permesso, dal 2005 ad oggi, di incrementare di ben 23 nuove specie l’e-lenco di quelle note per l’area [b]; le indagini integrative svolte nell’ultimo anno, hanno poi consentito di completare l’indagine prelimi-nare di caratterizzazione avifaunistica di Par-co, ZPS e SIC, nonché dei corridoi ecologici di connessione tra i differenti settori [c, d].
Le conoscenze raccolte, sono state quindi analizzate in funzione dell’utilità che rive-stono ai fini della definizione di una strategia di conservazione, realizzando un approccio multilivello che mettesse in relazione le spe-cie presenti nelle diverse aree con gli ambienti da esse utilizzati, e consentisse, unendo le in-formazioni relative alla loro distribuzione, diindividuare settori prioritari dal punto di vi-sta gestionale [e], ottenendo così strumenti di aiuto alla decisione basati su criteri oggettivi.
0% 20% 40% 60% 80% 100%
Libellule
F f ll 't t'
Contributo all'archivio dati apportato dalle azioni attuate nel 2012
Farfalle 'target'
Anfibi
Rettili
Uccelli
Mammiferi
Numero di nuove specie di uccelli contattate tra il 2005 e il 2012.
2323
70%
80%
90%
100%
Km
Uccelli: copertura progressiva dell'area di studio.
10%
20%
30%
40%
50%
60%
cent
uale
par
tice
lle 1
x 1
0%
10%
2005 2006 2007 2008 2009 2010 2011 2012
Per
c
Anni
��
����
�
������
�������
��
��������
�
����������
�������������
������������
������������
������������
�����������
�������������
�����������������
�������������������
�������������������
�������������������
��
�����������������
�
�������������������
�������������������
�������������������
������������������
��������������
������������
�
����������
�
���������
�
�
���
�
��
���
�
�����
���
����
�
����
��
�
��
� � � � � ���� � �
� ����� �� �� �������� �� �
������� �������� �
�� ���������� ����� ���������� � � ��
� � ���� ���������� �� � ����� ����� ���������� � �
� ������ ������������ ����� ���� ��������� � ����
� � � �� ���������� � ��� � � ����������
��������� � ����������� ��� �
� ������������� � ������ � ����������� ����������
����� � � � � � ������������ ������������ ������� ������� ���� ���������������������������
� ������� �������������� ������������������ ������������������ ����������������� ���������������� �������������� ���������������� ���������������� ��������������� ���������������� � ���������������� ��
���������� ��������������������������� ���� ��� ����� � �������������������������������� ��
����� ������������������������������ �����������������������������������
�������������������������������������� �������������������������������������
�������������������������������������������������������������������������������
� ���������������������������������������� � ��������������������������������������� �� �����������������������������������
� ������������������������������������ ������������������������� �� �
���� � �������������� �� � �� ��� � � ��������� � ��
���������� ��������� �
������������� ����� ��� ��
[a] [b]
[c]
[d]
[e]
L’analyse des informations rassemblées montre en outre que, même dans une aire bien caractérisée comme est le Parco del Beigua, des enquêtes intégratives puissent conduire à des incrémentations significatives de la qualité et quantité des connaissances disponibles. On peut en effet observer comme, pour tous les groupes d’animaux faut objet d’actions spécifiques pendant l’année 2012 (comprenant soit des recensions soit l’acquisition d’informations déjà connues) , la contribution à la connaissance soit été remarquable [a], avec des significatives incrémentations d’archive des données disponibles. Même pour les oiseaux, déjà fait objet de monitorages pluriannuels, la constante répétition des recensions a permis, à faire compte de 2005 jusqu’à aujourd’hui, d’incrémenter de 23 nouvelles espèces la liste de celles connues dans l’aire [b]; les enquêtes intégratives, effectuées dans la dernière année, ont donc permis le complètement de l’enquête préliminaire de caractérisation des oiseaux du Parc, ZPS et SIC, ainsi que des couloirs écologiques de lien entre secteurs diverses [c, d]. Les connaissances rassemblées, ont donc été analysées en fonction de l’utilité qui revêtent au fin de la définition d’une stratégie de conservation, en constituant une approche multi niveau qui met en relation les espèces présentes dans les plusieurs aires avec les environnements par eux utilisés et qui permet, en rassemblent les informations qui concernent leur distribution, d’identifier des secteurs prioritaires en point de vue de la gérance [e], obtenant donc des instruments d’aide à la décision basés sur des critères objectifs.

20 ATTIVITÀ DI RICERCA, CENSIMENTO E MONITORAGGIO
GLI UCCELLI
Il valore ornitologico del Parco del Beigua è riconosciuto, ormai da molti anni, ed ha portato dapprima all’inclusione dell’area nelle Important Bird Areas italiane (Gariboldi et al. 2000) ed alla successiva designazione a ZPS da parte della Regione Liguria (D.G.R. n. 270 del 25/2/2000); ciò è riconducibile sia all’abbondanza delle specie nidifi canti, che all’entità del fl usso migratorio riguardante molte specie, comprendenti sia rapaci diurni, per i quali è una delle principali zone di migrazione del Mediterraneo occidentale (Zalles & Bildstein 2000; Agostini 2002), che Passeriformi (Spanò et al. 1996; Realini 2002).
L’avifauna include attualmente 204 specie (92 Non Passeriformi e 112 Passeriformi), che rappresentano il 49,8% di quelle note per la Liguria (Baghino et al. 2012) ed il 38,8% di quelle italiane (Fracasso et al. 2009). Tra esse 103 sono nidifi canti (il 64,4% di quelle liguri) e 83 svernanti (delle quali 68 sedentarie), mentre 39 sono presenti solo durante le migrazioni e 44 risultano accidentali.
Nel corso degli anni 2006-2012 l’Ente Parco del Beigua ha poi attuato, nell’area protetta e nella connessa ZPS Beigua – Turchino, un dettagliato piano di monitoraggio dell’avifauna; esso contempla azioni di censimento svolte sia durante il periodo riprodut-tivo che durante le migrazioni (Fasano & Aluigi 2007; Fasano et al. 2009).
L’elenco sistematico delle specie ammesse per l’area a tutto il 2012, che viene ripor-tato nelle pagine successive, comprende l’indicazione della fenologia, sintetizzata utiliz-zando le seguenti sigle:B (Breeding) specie presente nel periodo riproduttivo e nidifi cante.T (Transient) specie presente con popolazioni in transito.
Attribuzione assegnata cercando di rispettare rigorosamente due criteri: specie non residen-ti, che quindi vi giungono regolarmente per nidifi care, per svernare o vi compaiono durante le migrazioni; specie presenti tutto l’anno, ma per le quali sono note ricatture regionali di individui inanellati all’estero o viceversa. Nei limiti del possibile si escludono da questo criterio quei movimenti di emigrazione o immigrazione, prevalentemente attribuibili a fe-nomeni di dispersione giovanile.
W (Wintering) specie indicativamente presente fra il 1 dicembre e il 15 febbraio. La sedentarietà, a livello di specie, è data dall’associazione delle sigle B e W.
V (Vagrant) specie di comparsa accidentale (fi no a 10 segnalazioni; o più di 10, ma in meno di 6 anni, dopo il 1950). Alle categorie B, T e W possono essere affi ancate le seguenti abbreviazioni (in mancanza di queste è sottintesa la regolarità della presenza):
irr. (irregular) presenza constatata più di 10 volte e in almeno 6 anni, ma in meno di 9 degli ultimi 10 anni in riferimento alla categoria associata; specie che ha nidifi cato in 4-8 anni degli ultimi 10 anni.
occ. (occasional) presenza constatata 1-10 volte o più di 10 volte ma in meno di 6 anni in riferimento alla categoria associata; specie che ha nidifi cato solo in 1-3 siti o anni.
int. (introduced) specie o individui immessi in natura volontariamente (generalmente per scopi venatori) o involontariamente (sfuggiti da cattività).
ext. (extinct) si riferisce alla nidifi cazione. Specie non più nidifi cante negli ultimi dieci anni.? indica eventuali situazioni di incertezza e/o carenza di dati certi.
Accanto alle specie incluse nell’allegato I della Direttiva Uccelli è riportato l’emblema dell’Unione europea:
LES OISEAUX
La valeur ornithologique du Parco del Beigua est reconnu, désormais par plusieurs années, et a emmené en premier compte à l’inclusion dans l’aire des Important Bird Areas italiennes (Gariboldi et al. 2000) et à une successive désignation à ZPS par la Regione Liguria (D.G.R. n. 270 du 25/2/2000); ça peut être reconduit soit à l’abondance des espèces nidifiants, que à l’entité du flux migratoire de plusieurs espèces, comprenant soit des oiseaux de proie diurnes, dont une des principales zones de migration est la Méditerranée occidentale (Zalles & Bildstein 2000; Agostini 2002), soit des passeraux (Spanò et al. 1996; Realini 2002). Les oiseaux incluent aujourd’hui 204 espèces (92 non passeraux et 112 passeraux), qui représentent le 49,8% des espèces connues pour la Ligurie (Baghino et al. 2012) et le 38,8% des espèces italiennes (Fracasso et al. 2009). Entre eux, 103 sont nidifiants (64,4% des quelles de la Ligurie) et 83 hivernantes (dont 68 sédentaires), pendant que 39 sont présentes seulement pendant les migrations et 44 sont accidentelles. Au fil des années 2006-2012 l’Ente del Parco del Beigua a activé, dans l’aire protégée et dans l’aire ZPS Beigua – Turchino à lui reliée, un détaillé plan de monitorage des oiseaux ; celui-ci contemple des actions de recension effectuées soit pendant la période de reproduction, soit pendant les migrations (Fasano & Aluigi 2007; Fasano et al. 2009). La liste systématique des espèces admises pour l’aire à tout le 2012, et qui est indiquée dans les pages qui suivent, comprend l’indication de la phénologie, synthétisée avec les sigles qui suivent: B (Breeding) espèce présente dans la période de reproduction et nidifiante. T (Transient) espèce présente avec des populations de passage.
L’attribution est assignée en cherchant de respecter rigoureusement deux critères : espèces non résidentes, qui donc arrivent régulièrement là pour nidifier, pour l’hivernage ou qui apparaît pendant les migrations ; espèces présentes toute l’année, mais dont les captures régionales des individus annulés à l’extérieur ou vice versa sont connues. Dans la mesure du possible, on exclue de ce critère les mouvements d’émigration ou immigration qui soient substantiellement attribuables à des phénomènes de dispersion de jeunesse.
W (Wintering) espèces typiquement présentes entre le 1 décembre et il 15 février . La sedentairété, à niveau d’espèce, est donnée par l’association des sigles B et W.
V (Vagrant) espèces d’apparition accidentelle (jusqu’à 10 signalisations; ou plus de 10, mais en moins de 6 années, après 1950). Aux catégories B, T et W peuvent être associées les abréviations qui suivent (en manque de celles-ci il est sous-entendue la régularité de la présence):
irr. (irregular) présence constatée plus de 10 fois et en au moindre 6 années, mais en moins de 9 des dernières 10 années en référence à la catégorie associée; espèce qui à nidifié en 4-8 années des dernières 10 années.
occ. (occasional) présence constatée 1-10 fois ou plus de 10 fois mais en moins de 6 années en référence à la catégorie associée; espèce qui a nidifié seulement dans 1-3 sites ou années.
int. (introduced) espèces ou individus mises en nature volontairement (en général à fin de chasse) ou involontairement (fuite de la captivité).
ext. (extinct) ça concerne la nidification. Espèce non plus nidifiante dans les dernières dix années.
? montre éventuelles situations d’incertitude et/ou manque de données certaines. Associé aux espèces incluses dans l’Annexe I de la Directive Oiseaux est indiqué l’emblème de l’Union Européenne:

Cigno selvatico Cygnus cygnus V-1 Alzavola Anas crecca V-1
Germano reale Anas platyrhynchos B, T, WMarzaiola Anas querquedula V-2
Pernice rossa Alectoris rufa B, WStarna Perdix perdix B occ. int., W
Quaglia comune Coturnix coturnix B occ., T
Fagiano comune Phasianus colchicus B, WCormorano Phalacrocorax carbo T, W
Nitticora Nycticorax nycticorax T occ.?Airone guardabuoi Bubulcus ibis V-1
Garzetta Egretta garzetta T irr. Airone bianco maggiore Casmerodius albus T, W
Airone cenerino Ardea cinerea B, T, W
Airone rosso Ardea purpurea T irr. Cicogna nera Ciconia nigra T
Cicogna bianca Ciconia ciconia T
Tuffetto Tachybaptus rufi collis B irr., T irr., W irr.Falco pecchiaiolo Pernis apivorus B, T
Nibbio bruno Milvus migrans T Nibbio reale Milvus milvus T
Capovaccaio Neophron percnopterus V-1 Grifone Gyps fulvus T occ.
Biancone Circaetus gallicus B, T, W occ. Falco di palude Circus aeruginosus T
Albanella reale Circus cyaneus T irr., W occ. Albanella pallida Circus macrourus V-3
Albanella minore Circus pygargus T Astore Accipiter gentilis B, W
Sparviere Accipiter nisus B, T, WPoiana Buteo buteo B, T, W
Poiana codabianca Buteo rufi nus V-2 Poiana calzata Buteo lagopus V-1
Aquila anatraia maggiore Aquila clanga V-3Aquila anatraia minore Aquila pomarina V-8
Aquila minore Aquila pennata B occ.?, T Aquila reale Aquila chrysaetos B, WAquila di Bonelli Aquila fasciata V-5Aquila imperiale Aquila heliaca V-1
Falco pescatore Pandion haliaetus T Grillaio Falco naumanni T irr.
21
Cygne sauvage Cygnus cygnus V-1 Sarcelle d’hiver Anas crecca V-1
Canard colvertAnas platyrhynchos B, T, W Arcelle d’été Anas querquedula V-2 Perdrix rouge Alectoris rufa B, W
Perdrix grise Perdix perdix B occ. int., W Caille des blés Coturnix coturnix B occ., T
Faisan de Colchide Phasianus colchicus B, W Grand cormoran Phalacrocorax carbo T, W
Bihoreau gris Nycticorax nycticorax T occ.? Héron garde-bœufs Bubulcus ibis V-1
Aigrette garzette Egretta garzetta T irr. Grande aigrette Casmerodius albus T, W
Héron cendré Ardea cinerea B, T, W Héron pourpré Ardea purpurea T irr.
Cicogne noire Ciconia nigra T Cicogne blanche Ciconia ciconia T
Grèbe castagneux Tachybaptus ruficollis B irr., T irr., W irr. Bondrée apivore Pernis apivorus B, T
Milan noir Milvus migrans T Milan royal Milvus milvus T
Percnoptère Neophron percnopterus V-1 Vautour fauve Gyps fulvus T occ.
Circaète Jean-le-Blanc Circaetus gallicus B, T, W occ. Busard des roseaux e Circus aeruginosus T
Busard Saint-Martin Circus cyaneus T irr., W occ. Busard pâle Circus macrourus V-3 Busard cendré Circus pygargus T
Autour des palombes Accipiter gentilis B, W Epervier d'Europe Accipiter nisus B, T, W
Buse variable Buteo buteo B, T, W Buse féroce Buteo rufinus V-2
Buse pattue Buteo lagopus V-1 Aigle criard Aquila clanga V-3
Aigle pomarine Aquila pomarina V-8 Aigle Aquila pennata B occ.?, T
Aigle royal Aquila chrysaetos B, W Aigle de Bonelli Aquila fasciata V-5 Aigle impériale Aquila heliaca V-1
Balbuzard pêcheur Pandion haliaetus T Faucon crécerellette Falco naumanni T irr.

Gheppio Falco tinnunculus B, T, WFalco cuculo Falco vespertinus T
Smeriglio Falco columbarius T occ.Lodolaio Falco subbuteo B, T
Falco della Regina Falco eleonorae T irr.
Lanario Falco biarmicus V-1 Falco pellegrino Falco peregrinus B, T, W
Porciglione Rallus aquaticus V-1
Gallinella d’acqua Gallinula chloropus B occ., T irr.
Gru Grus grus T Pernice di mare Glareola pratincola V-1
Piviere tortolino Charadrius morinellus T Pavoncella Vanellus vanellus T occ.
Gambecchio comune Calidris minuta V-1
Frullino Lymnocryptes minimus V-1
Beccaccino Gallinago gallinago T occ. Beccaccia Scolopax rusticola T, W
Chiurlo maggiore Numenius arquata V-1Piro piro piccolo Actitis hypoleucos T occ.Piro piro culbianco Tringa ochropus T irr.
Pettegola Tringa totanus V-2Gabbiano comune Chroicocephalus ridibundus T, W
Gabbiano reale Larus michahellis T, WPiccione torraiolo Columba livia var. domestica B, W
Colombella Columba oenas T occ.Colombaccio Columba palumbus B, T, W
Tortora dal collare Streptopelia decaocto B, WTortora selvatica Streptopelia turtur B, T
Cuculo dal ciuffo Clamator glandarius V-1Cuculo Cuculus canorus B, T
Barbagianni Tyto alba B?, T irr., W?Assiolo Otus scops B, T
Gufo reale Bubo bubo B, WCivetta Athene noctua B, W
Allocco Strix aluco B, WGufo comune Asio otus B occ., T
Gufo di palude Asio fl ammeus V-2 Succiacapre Caprimulgus europaeus B, T
Rondone comune Apus apus B, T
Rondone pallido Apus pallidus TRondone maggiore Apus melba T
22
Faucon crécerelle Falco tinnunculus B, T, W Faucon kobez Falco vespertinus T
Faucon émerillon Falco columbarius T occ. Faucon hobereau Falco subbuteo B, T
Faucon d'Éléonore Falco eleonorae T irr. Faucon lanier Falco biarmicus V-1
Faucon pèlerin Falco peregrinus B, T, W Râle d'eau Rallus aquaticus V-1
Gallinule poule d'eau Gallinula chloropus B occ., T irr. Grue cendrée Grus grus T
Glaréole à collier Glareola pratincola V-1 Pluvier guignard Charadrius morinellus T
Vanneau huppé Vanellus vanellus T occ. Bécasseau minute Calidris minuta V-1
Bécassine sourde Lymnocryptes minimus V-1 Bécassine des marais Gallinago gallinago T occ.
Bécasse des bois Scolopax rusticola T, W Courlis cendré Numenius arquata V-1
Chevalier guignette Actitis hypoleucos T occ. Chevalier cul-blanc Tringa ochropus T irr.
Chevalier gambette Tringa totanus V-2 Mouette rieuse Chroicocephalus ridibundus T, W
Goéland leucophée Larus michahellis T, W Pigeon biset Columba livia var. domestica B, W
Pigeon colombin Columba oenas T occ. Pigeon ramier Columba palumbus B, T, W
Tourterelle turque Streptopelia decaocto B, W Tourterelle des bois Streptopelia turtur B, T
Coucou geai Clamator glandarius V-1 Coucou gris Cuculus canorus B, T
Effraie des clochers Tyto alba B?, T irr., W? Petit-duc scops Otus scops B, T
Hibou grand-duc Bubo bubo B, W Chevêche d'Athéna Athene noctua B, W
Chouette hulotte Strix aluco B, W Hibou moyen-duc Asio otus B occ., T
Hibou des marais Asio flammeus V-2 Engoulevent d’Europe Caprimulgus europaeus B, T
Martinet noir Apus apus B, T Martinet pâle Apus pallidus T
Martinet à ventre blanc Apus melba T

Rondone indiano Apus affi nis V-1Martin pescatore Alcedo atthis B occ., T, W
Gruccione Merops apiaster TGhiandaia marina Coracias garrulus T
Upupa Upupa epops B, T C03-05
Torcicollo Jynx torquilla B occ., TPicchio verde Picus viridis B, W
Picchio nero Dryocopus martius B, W
Picchio rosso maggiore Dendrocopos major B, W
Picchio rosso minore Dendrocopos minor B, W
Calandrella Calandrella brachydactyla V-2 (B occ.?) Tottavilla Lullula arborea B, T, W
Allodola Alauda arvensis B, T, W
Topino Riparia riparia TRondine montana Ptyonoprogne rupestris B, T, W
Rondine Hirundo rustica B, T Balestruccio Delichon urbicum B, T
Rondine rossiccia Cecropis daurica TCalandro Anthus campestris B, T Prispolone Anthus trivialis B, TPispola Anthus pratensis T, W
Pispola golarossa Anthus cervinus V-1Spioncello Anthus spinoletta T, W
Cutrettola Motacilla fl ava TBallerina gialla Motacilla cinerea B, T, WBallerina bianca Motacilla alba B, T, WBeccofrusone Bombycilla garrulus V-3Merlo acquaiolo Cinclus cinclus B, W
Scricciolo Troglodytes troglodytes B, T, WPassera scopaiola Prunella modularis T, W
Sordone Prunella collaris T, WPettirosso Erithacus rubecula B, T, WUsignolo Luscinia megarhynchos B, T
Codirosso spazzacamino Phoenicurus ochruros B, T, WCodirosso comune Phoenicurus phoenicurus B, T
Stiaccino Saxicola rubetra TSaltimpalo Saxicola torquatus B, T, W
Culbianco Oenanthe oenanthe B occ., TMonachella nera Oenanthe leucura B ext.
Codirossone Monticola saxatilis B, TPassero solitario Monticola solitarius B, W
23
Martinet des maisons Apus affinis V-1 Martin-pêcheur d'Europe Alcedo atthis B occ., T, W
Guêpier d'Europe Merops apiaster T Rollier d'Europe Coracias garrulus T
Huppe fasciée Upupa epops B, T Torcol fourmilier Jynx torquilla B occ., T
Pic vert Picus viridis B, W Pic noir Dryocopus martius B, W Pic épeiche Dendrocopos major B, W
Pic épeichette Dendrocopos minor B, W Alouette calandrelle Calandrella brachydactyla V-2 (B occ.?)
Alouette lulu Lullula arborea B, T, W Alouette des champs Alauda arvensis B, T, W
Hirondelle de rivage Riparia riparia T Hirondelle de rochers Ptyonoprogne rupestris B, T, W
Hirondelle rustique Hirundo rustica B, T Hirondelle de fenêtre Delichon urbicum B, T
Hirondelle rousseline Cecropis daurica T Pipit rousseline Anthus campestris B, T
Pipit des arbres Anthus trivialis B, T Pipit farlouse Anthus pratensis T, W
Pipit à gorge rousse Anthus cervinus V-1 Pipit spioncelle Anthus spinoletta T, W
Bergeronnette printanière Motacilla flava T Bergeronnette des ruisseaux Motacilla cinerea B, T, W
Bergeronnette grise Motacilla alba B, T, W Jaseur boréal Bombycilla garrulus V-3 Cincle plongeur Cinclus cinclus B, W
Troglodyte mignon Troglodytes troglodytes B, T, W Accenteur mouchet Prunella modularis T, W
Accenteur alpin Prunella collaris T, W Rouge-gorge familier Erithacus rubecula B, T, W Rossignol philomèle Luscinia megarhynchos B, T Rougequeue noir Phoenicurus ochruros B, T, W
Rougequeue à front blanc Phoenicurus phoenicurus B, T Tarier des prés Saxicola rubetra T
Tarier pâtre Saxicola torquatus B, T, W Traquet motteux Oenanthe oenanthe B occ., T
Traquet rieur Oenanthe leucura B ext. Monticole de roche Monticola saxatilis B, T Monticole bleu Monticola solitarius B, W

Merlo dal collare Turdus torquatus V-8Merlo Turdus merula B, T, W
Cesena Turdus pilaris T, WTordo bottaccio Turdus philomelos B, T, W
Tordo sassello Turdus iliacus T, W
Tordela Turdus viscivorus B irr., T, WUsignolo di fi ume Cettia cetti B occ.
Beccamoschino Cisticola juncidis V-1Forapaglie macchiettato Locustella naevia V-1
Forapaglie comune Acrocephalus schoenobaenus V-4 Cannaiola comune Acrocephalus scirpaceus V-2
Canapino maggiore Hippolais icterina T
Canapino comune Hippolais polyglotta B occ., T
Capinera Sylvia atricapilla B, T, W
Beccafi co Sylvia borin TBigiarella Sylvia curruca T occ.
Bigia grossa Sylvia hortensis B occ., T occ.
Sterpazzola Sylvia communis B, TMagnanina comune Sylvia undata B, W
Magnanina sarda Sylvia sarda V-1 Sterpazzolina comune Sylvia cantillans T
Sterpazzolina di Moltoni Sylvia subalpina B, TOcchiocotto Sylvia melanocephala B, W
Luì bianco Phylloscopus bonelli B, TLuì verde Phylloscopus sibilatrix B occ., TLuì piccolo Phylloscopus collybita B, T, W
Luì grosso Phylloscopus trochilus TRegolo Regulus regulus B occ., T, W
Fiorrancino Regulus ignicapilla B, WPigliamosche Muscicapa striata B, T
Balia dal collare Ficedula albicollis V-5 Balia nera Ficedula hypoleuca T
Codibugnolo Aegithalos caudatus B, T, WCinciarella Cyanistes caeruleus B, T, W
Cinciallegra Parus major B, T, WCincia dal ciuffo Lophophanes cristatus B, W
Cincia mora Periparus ater B, T, WCincia bigia Poecile palustris B, T, WPicchio muratore Sitta europaea B, W
Picchio muraiolo Tichodroma muraria W irr.Rampichino comune Certhia brachydactyla B, W
24
Merle à plastron Turdus torquatus V-8 Merle noir Turdus merula B, T, W Grive litorne Turdus pilaris T, W
Grive musicienne Turdus philomelos B, T, W Grive mauvis Turdus iliacus T, W
Grive draine Turdus viscivorus B irr., T, W Bouscarle de Cetti Cettia cetti B occ.
Cisticole des joncs Cisticola juncidis V-1 Locustelle tachetée Locustella naevia V-1
hragmite des joncs Acrocephalus schoenobaenus V-4 Rousserolle effarvatte Acrocephalus scirpaceus V-2
Hypolaïs ictérine Hippolais icterina T Hypolaïs polyglotte Hippolais polyglotta B occ., T
Fauvette à tête noire Sylvia atricapilla B, T, W Fauvette des jardins Sylvia borin T
Fauvette babillarde Sylvia curruca T occ. Fauvette orphée Sylvia hortensis B occ., T occ.
Fauvette grisette Sylvia communis B, T Fauvette pitchou Sylvia undata B, W
Fauvette sarde Sylvia sarda V-1 Fauvette passerinette Sylvia cantillans T
Fauvette de Moltoni Sylvia subalpina B, T Fauvette mélanocéphale Sylvia melanocephala B, W
Pouillot de Bonelli Phylloscopus bonelli B, T Pouillot siffleur Phylloscopus sibilatrix B occ., T Pouillot véloce Phylloscopus collybita B, T, W
Pouillot fitis Phylloscopus trochilus T Roitelet huppé Regulus regulus B occ., T, W
Roitelet à triple bandeau Regulus ignicapilla B, W Gobemouche gris Muscicapa striata B, T
Gobe-mouche à collier Ficedula albicollis V-5 Gobe-mouche noir Ficedula hypoleuca T
Mésange à longue queue Aegithalos caudatus B, T, W Mésange bleue Cyanistes caeruleus B, T, W Mésange charbonnière Parus major B, T, W
Mésange huppée Lophophanes cristatus B, W Mésange noire Periparus ater B, T, W
Mésange nonnette Poecile palustris B, T, W Sittelle torchepot Sitta europaea B, W
Tichodrome échelette Tichodroma muraria W irr. Grimpereau des jardins Certhia brachydactyla B, W

Rigogolo Oriolus oriolus B, TAverla piccola Lanius collurio B, T Averla cenerina Lanius minor V-1
Averla maggiore Lanius excubitor V-1Averla capirossa Lanius senator B occ., T irr.
Ghiandaia Garrulus glandarius B, WGazza Pica pica B, W
Nocciolaia Nucifraga caryocatactes V-1 Gracchio corallino Pyrrhocorax pyrrhocorax V-1
Taccola Corvus monedula B, W
Cornacchia nera Corvus corone B, WCornacchia grigia Corvus cornix B, W
Corvo imperiale Corvus corax B, W
Storno nero Sturnus unicolor V-1
Storno Sturnus vulgaris B, T, W
Storno roseo Pastor roseus V-1Passera d’Italia Passer domesticus italiae B, W
Passera sarda Passer hispaniolensis V-1Passera mattugia Passer montanus T occ.
Fringuello Fringilla coelebs B, T, WPeppola Fringilla montifringilla T, W
Verzellino Serinus serinus B, T, WVerdone Carduelis chloris B, T, W
Cardellino Carduelis carduelis B, T, W Venturone alpino Carduelis citrinella T occ.
Lucherino Carduelis spinus T, WFanello Carduelis cannabina B occ., T
Fanello nordico Carduelis fl avirostris V-1Crociere Loxia curvirostra B occ., T, WCiuffolotto Pyrrhula pyrrhula B, T, W
Frosone Coccothraustes coccothraustes B occ., TZigolo delle nevi Plectrophenax nivalis V-1
Zigolo giallo Emberiza citrinella V-4Zigolo nero Emberiza cirlus B, T, W
Zigolo muciatto Emberiza cia B, T, WOrtolano Emberiza hortulana B occ., T irr.
Ortolano grigio Emberiza caesia V-1 Zigolo minore Emberiza pusilla V-6
Migliarino di palude Emberiza schoeniclus TStrillozzo Emberiza calandra B occ., T irr.
25
Loriot d'Europe Oriolus oriolus B, T Pie-grièche écorcheur Lanius collurio B, T
Pie-grièche à poitrine rose Lanius minor V-1 Pie-grièche grise Lanius excubitor V-1
Pie-grièche à tête rousse Lanius senator B occ., T irr. Geai des chênes Garrulus glandarius B, W
Pie bavarde Pica pica B, W Casse-noix moucheté Nucifraga caryocatactes V-1
Crave à bec rouge Pyrrhocorax pyrrhocorax V-1 Choucas des tours Corvus monedula B, W
Corneille noire Corvus corone B, W Corneille mantelée Corvus cornix B, W
Grand Corbeau Corvus corax B, W Étourneau unicolore Sturnus unicolor V-1
Étourneau sansonnet Sturnus vulgaris B, T, W Étourneau roselin Pastor roseus V-1
Moineau cisalpin Passer domesticus italiae B, W Moineau espagnol Passer hispaniolensis V-1
Moineau friquet Passer montanus T occ. Pinson des arbres Fringilla coelebs B, T, W
Pinson du Nord Fringilla montifringilla T, W Serin cini Serinus serinus B, T, W
Verdier d'Europe Carduelis chloris B, T, W Chardonneret élégant Carduelis carduelis B, T, W Venturon montagnard Carduelis citrinella T occ.
Tarin des aulnes Carduelis spinus T, W Linotte mélodieuse Carduelis cannabina B occ., T
Linotte à bec jaune Carduelis flavirostris V-1 Bec-croisé des sapins Loxia curvirostra B occ., T, W
Bouvreuil pivoine Pyrrhula pyrrhula B, T, W Gros-bec casse-noyaux Coccothraustes coccothraustes B occ, T
Plectrophane des neiges Plectrophenax nivalis V-1 Bruant jaune Emberiza citrinella V-4 Bruant zizi Emberiza cirlus B, T, W Emberiza cia Emberiza cia B, T, W
Bruant ortolan Emberiza hortulana B occ., T irr. Bruant cendrillard Emberiza caesia V-1
Bruant nain Emberiza pusilla V-6 Bruant des roseaux Emberiza schoeniclus T
Bruant proyer Emberiza calandra B occ., T irr.

26 ATTIVITÀ DI RICERCA, CENSIMENTO E MONITORAGGIO
L’AVIFAUNA NIDIFICANTE
Ogni anno nel Parco del Beigua possiamo stimare che nidifi chino tra le 33.100 e le 36.300 coppie di uccelli, un importante patrimonio da conoscere e salvaguardare! Men-tre per numerose specie di importanza comunitaria o locale si sono adottate metodiche particolari (successivamente descritte in dettaglio, ed i cui risultati vengono riportati nel-le schede relative alle singole specie), per la caratterizzazione ed il monitoraggio della comunità ornitica nidifi cante, la tecnica di rilevamento prescelta è stata quella dei punti di ascolto, meglio noti come «Point counts» nella letteratura ornitologica anglosassone. Essi consistono sostanzialmente nel riconoscere e conteggiare tutti gli uccelli visti o sen-titi in un punto scelto casualmente ed in un tempo determinato.
I dati raccolti vengono quindi analizzati calcolando una serie di indici d’abbondanza e trasformati, in sede di elaborazione fi nale, in ‘numero di coppie’, l’unità di studio ritenuta più adeguata per i censimenti effettuati durante il periodo riproduttivo.
Considerando i dati disponibili per il Parco del Beigua e tutti i set-tori ad esso connessi (anni 2000-2012), le specie rilevate durante l’e-secuzione dei 1.468 punti d’ascolto sono state 101; tra queste, per 95 si sono riscontrati indizi di nidifi cazio-ne, rappresentando quindi il 92,2% di quelle attualmente accertate come nidifi canti nell’area di studio. Nel complesso gli indici calcolati per il Beigua evidenziano la presenza di una comunità ricca di specie ed equilibrata nella sua composizio-ne, perfettamente confrontabile con analoghe realtà ornitologiche, con-fermandone l’importanza nell’am-bito della biodiversità dell’avifauna degli ambienti appenninici e, più in generale, mediterranei.
Tra le specie osservate, le più ab-bondanti sono state Pettirosso, Frin-guello, Capinera, Merlo, Cincialle-gra, Luì piccolo, Cuculo, Prispolo-ne, Ghiandaia, Rondone comune e Allodola. Nel grafico viene ripor-tata l’abbondanza relativa (espres-sa come frequenza percentuale del numero di individui contattati) delle principali 35 specie (le barre rosse indicano le specie dominanti, le bar-re gialle quelle sub-dominanti) [a].
l i
Luì bianco
Codibugnolo
Magnanina comune
Cincia bigia
Picchio rosso maggiore
Colombaccio
Sterpazzolina di Moltoni
Codirosso spazzacamino
Fiorrancino
Ciuffolotto
Tortora selvatica
Cincia mora
Saltimpalo
Tordo bottaccio
Picchio verde
Calandro
Scricciolo
Cincia dal ciuffo
Zigolo muciatto
Rampichino comune
Picchio muratore
Cornacchia grigia
Zigolo nero
Cuculo
Prispolone
Ghiandaia
Rondone comune
Allodola
Cinciarella
Pettirosso
Fringuello
Capinera
Merlo
Cinciallegra
Luì piccolo
0 5 10
Abbondanza
[a]
LA COMMUNAUTÉ DES OISEAUX NIDIFIANTS
Chaque année dans le Parco del Beigua on peut estimer que entre 33.100 et 36.300 couples de oiseaux nidifient, un patrimoine important qu’il faut connaître et sauvegarder ! Si par plusieurs espèces d’importance communautaire ou locale on a adopté des méthodiques particulières (ci-dessous décrites en détail, et dont les résultats sont reportés dans les fichiers reliés à chaque espèce), pour la caractérisation et le monitorage de la communauté des oiseaux nidifiant, la technique de détection choisie a été quelle des points d’écoute, aussi connus comme «Point counts» pour la littérature ornithologique anglo saxone. Ils consistent substantiellement dans la détection et comptage des touts les oiseaux vus ou étendus dans un certain point choisit par hasard et en un certain moment. Les données rassemblées sont après analysées en calculant une série d’index d’abondance et étant transformées, au moment de l’élaboration finale, en un “numéro de couples”, l’unité d’étude retenue comme la plus juste pour les recensions effectuées pendant la période de reproduction. En considérant les données disponibles pour le Parco del Beigua et touts les secteurs à lui reliés (années 2000- 2012), les espèces identifiées pendant l’exécution des 1.468 points d’écoute ont été 101; entre eux, pour 95 espèces on à individué des indices de nidification, qui représentent le 92,2% de ces qui sont aujourd’hui vérifiées comme nidifiantes dans l’aire d’étude. Dans le complexe, les indexes calculés pour le Beigua montrent la présence d’une communauté riche d’espèces et au même temps équilibrée en sa composition, parfaitement rapportable avec des réalités ornithologiques analogues, en confirmant l’importance dans le milieu de la biodiversité des oiseaux des environnements des Apennins et, plus en général, de la Méditerranée. Entre les espèces observées, les plus abondantes ont été le Rouge-gorge familier, Pinson des arbres, Fauvette à tête noire, Merle noir, Mésange charbonnière, Pouillot véloce, Coucou gris, Pipit des arbres, Rollier d'Europe, Martinet noir et Alouette des champs. Dans la graphique est indiquée l’abondance relative (exprimée comme fréquence en pourcentage du numéro d’individus contactés) des principales 35 espèces (les barres rouges indiquent les espèces dominantes, les barres jaunes les espèces sub-dominantes) [a].

27
IL MONITORAGGIO DELL’AVIFAUNA NIDIFICANTE IN LIGURIA
Grazie ai dati raccolti attraverso un progetto della Regione Liguria, attuato adottando il modello di monitoraggio avifaunistico già in uso nel Parco del Beigua (che è inoltre l’Ente attuatore del progetto regionale), è possibile sia documentare lo stato attuale, caratterizzando la comunità orni-tica nidifi cante ed evidenziandone i trend, sia monitorare le specie presenti nell’area di studio che siano tutelate dalla Direttiva Uccelli e quelle rare, importanti dal punto di vista scientifi co ovvero di interesse conservazionistico e gestionale locale (Fasano & Aluigi 2007; Fasano et al. 2009; Nicosia et al. 2009a, 2009b; Fasano et al. 2012).I dati raccolti consentono quindi di defi nire i probabili andamenti delle popolazioni nidifi canti di molte specie dell’avifauna ligure, evidenziando, nel periodo 2000-2012, una situazione di relativa stabilità, infl uenzata però negativamente dall’elevato numero di specie in diminuzione; infatti, su 54 specie considerate, il 39% risultano tendenti all’aumento o stabili, il 37% tendono alla diminu-zione, mentre per il 24% la tendenza non è ancora statisticamente defi nita. La tendenza alla dimi-nuzione è più marcata per le specie che nidifi cano in ambienti agrari ed antropizzati e per quelle che svernano nell’Africa sub-sahariana; mentre, le specie che svernano nell’area mediterranea e quelle legate alle praterie presentano andamenti delle loro popolazioni che tendono alla stabilità, e si riscontra inoltre incremento moderato per gli uccelli che preferiscono gli ambienti boscati.Ulteriori analisi confermano l’importanza conservazionistica delle aree liguri appartenenti alla Rete Natura 2000; esse risultano infatti, rispetto alle aree non soggette a tutela, mediamente più importanti per quanto riguarda quantità e qualità delle specie che ospitano. I risultati evidenziano anche come l’andamento delle popolazioni nidifi canti appaia nel complesso simile e coerente sia all’interno che all’esterno dei Siti Natura 2000, ma con una tendenza meno negativa per questi ultimi, mentre è nettamente differenziato per quanto riguarda le specie forestali e delle praterie, con tendenze positive dentro i Siti Natura 2000 e negative all’esterno. Per alcune specie di note-vole importanza conservazionistica vediamo poi come l’andamento degli indici dentro e fuori i Siti della Rete Natura 2000 diverga, con tendenze positive (Biancone e Magnanina comune) o di stabilità (Averla piccola) all’interno dei Siti Natura 2000 e negative all’esterno; la Tottavilla, che complessivamente presenta una diminuzione moderata, mostra andamento simile nei due ambiti, ma con una tendenza decisamente meno negativa all’interno dei Siti Natura 2000.
I dati raccolti nel Parco del Beigua confermano quanto evidenziato a livello regionale, con una situazione di sostanziale stabilità nei probabili andamenti delle popolazioni ni-difi canti. Questo sia per gli uccelli più comuni, che manifestano una tendenza non certa dal punto di vista statistico, ma tendente all’incremento, con leggeri aumenti per diverse delle 35 specie considerate ed una cer-ta tendenza alla diminuzione solo per lo Scricciolo [a]; sia per le numerose specie di maggiore importanza conser-vazionistica che, anzi, in alcuni casi tendono ad un moderato incremento.
I principali ambienti verranno am-piamente trattati di seguito, ma possia-mo anticipare che il maggior numero di specie si è riscontrato nelle aree
0,87 0,861,02
1,10
1,351,15
1
2
ce
0,66
0,87 0,861,02
1,10 1,15
0
1
Indi
ce
02006 2007 2008 2009 2010 2011 2012
Andamento annuo
[a]
LE MONITORAGE DE LA COMMUNAUTÉ DES OISEAUX NIDIFIANTS EN LIGURIE
Grâce aux données rassemblées à travers d’un projet de la Regione Liguria, activé en adoptant le modèle de monitorage des oiseaux déjà utilisé dans le Parco del Beigua (qui est l’organisme d’actuation du projet régional), il est possible soit de documenter l’état actuel, en caractérisant la communauté ornithologique nidifiant et en montrent les tendances, soit de contrôler les espèces présentes dans l’aire d’étude qui soient protégées par la Directive Oiseaux et celles rares, importantes au point de vue scientifique ou d’intérêt de conservation et gestion locale (Fasano & Aluigi 2007; Fasano et al. 2009; Nicosia et al. 2009a, 2009b; Fasano et al. 2012). Les données rassemblées consent donc de définir les probables tendances des populations nidifiants de plusieurs espèces de oiseaux ligures, en montrent, dans la période 2000-2012, une situation de substantielle stabilité, influencée toutefois négativement par l’haut numéro d’espèces en diminution; en effet, sur 54 espèces considérées, le 37% tend à diminuer, par contre pour le 24% d’eux la tendance n’est pas encore définie statistiquement. La tendance à la diminution résulte plus marquée pour les espèces qui nidifient dans des environnements agraires et anthropiques et pour celles qui hivernent dans l’Afrique sub-saharienne ; par contre, les espèces qui hivernent à l’intérieur de l’aire méditerranée et celles reliées aux prairies montrent des tendances des leur populations qui tendent à la stabilité, et dont on retrouve un modéré incrément pour les oiseaux qui préfèrent les environnements boisés. Ultérieures analyses confirment l’importance de conservation des aires ligures faisant partie de la Rete Natura 2000; elles résultent en effet, par rapport à des aires ne pas sujettes à tutelle, en moyenne plus importantes en ce qui concerne la quantité et qualité d’espèces qui hébergent. Les résultats illustrent même que l’évolution des populations nidifiant apparaît dans l’ensemble similaire et cohérente soit à l’intérieur que à l’extérieur des Sites Natura 2000, mais avec une tendance moins négative pour ces derniers, tandis que il est fortement différencié en ce qui concerne les espèces de forêts et des prairies, avec des tendances positives dans les Sites Natura 2000 et négatives hors d’eux. Pour certaines espèces de remarquable importance de conservation, on voit comme la tendance des index dedans et dehors les sites de la Rete Natura 2000 diverge, avec des tendances positives (Circaète Jean-le-Blanc et Fauvette pitchou) ou de stabilité (Pie-grièche écorcheur) à l’intérieur des sites Natura 2000 et négatives à hors d’eux; la Alouette lulu, qui dans l’ensemble présente une tendance modérée, montre une tendance similaire dans les deux milieux, mais avec une tendance fortement moins négative à l’intérieur des Sites Natura 2000.
Les données rassemblées dans le Parco del Beigua confirment ce qui montré à niveau régional, avec une situation de substantielle stabilité dans les probables évolutions des populations nidifiants. Ça est vrai soit pour les oiseaux les plus communs, qui manifestent une tendance ne pas sûre au point de vue statistique, mais qui tend à l’incrémentation, avec des légers augments pour plusieurs des 35 espèces considérées et une certaine tendance à la diminution seulement pour le Troglodyte mignon [a]; soit pour les nombreuses espèces de plus grande importance de conservation qui, au contraire, en certains cases tendent vers une modérée incrémentation.

28 ATTIVITÀ DI RICERCA, CENSIMENTO E MONITORAGGIO
caratterizzate dalla presenza di boschi (in particolare se con pre-senza di latifoglie), brughiere e cespuglieti e praterie [a]; mentre il numero medio di specie contat-tate per punto d’ascolto presenta differenze statisticamente signi-ficative tra i diversi ambienti, ed ha il suo valore massimo nei mo-saici agrari.
Valutando poi il valore ornito-logico e conservazionistico de-gli ambienti presenti, è risultato evidente come, la maggior rile-vanza, sia nettamente da attribu-ire alle praterie ed alla macchia
mediterranea; ma anche gli ambienti con rocce, falesie, rupi e affi oramenti sono con-traddistinti da elevati valori ornitologici. Boschi, brughiere e cespuglieti risultano essere ben diversifi cati, e sono quindi rilevanti anche dal punto di vista qualitativo. Particolare importanza, dovuta alla loro elevata biodiversità, è poi da ascrivere ai mosaici agrari.
Ricchezza specifi ca[b]
N° specie in all. I DU[c]
Aree rocciose
Mosaici agrari
Praterie
Brughiere e cespuglieti
Macchia mediterranea
Boschi di latifoglie
Boschi di conifere
Boschi misti
0 20 40 60 80
Boschi di latifoglie
Numero di Specie
[a]
Altre analisi evidenziano poi come i diversi settori presentino notevoli differenze qualitative e quantitative nell’avifauna che li contraddistingue, sia per ciò che riguarda il numero medio di spe-cie presenti [b], che il numero di specie incluse nell’allegato I del-la direttiva ‘Uccelli’ [c], eviden-ziando come non sempre le aree prioritarie individuate dai due parametri coincidano, ed ai fi ni gestionali sia quindi importante considerarli congiuntamente.
Les principaux environnements seront donc amplement traités ci-dessous, mais on peut anticiper que le plus grand numéro d’espèces a été identifié dans les aires caractérisées par la présence de bois (notamment si en présence de feuillius), de landes, arbustes et prairies [a]; au contraire, pour le numéro moyen d’espèces contactées pour chaque point d’écoute montre différences statistiquement significatives parmi les plusieurs environnements, et a sa valeur maximale dans les mosaïques agraires.
La évaluation de la valeur ornithologique et de conservation des environnements présents, a montré comme la plus grande relevance soit à attribuer aux prairies et à la végétation méditerranée ; toutefois aussi les environnements ayants des roches, falaises, rochers et affleurements sont caractérisés par des valeurs ornithologiques plutôt hautes. Les bois, landes et aires d’arbustes résultent bien diversifiés, et sont donc relevants même en ce qui concerne la qualité. Une particulière importance, due à leur grande biodiversité, est à attribuer aux mosaïques agraires.

29
METODI
La scelta delle metodiche attuate è stata dettata dalla coniugazione di due necessità principali: essere caratterizzate da elevata ripetibilità e confrontabilità dei rilevamenti e, nello stesso tempo, comportare un impegno economico compatibile con la prosecuzione delle attività sul lungo pe-riodo.Per la caratterizzazione ed il monitoraggio dell’ornitocenosi nidifi cante la tecnica di rilevamento prescelta è stata quella dei punti di ascolto senza limiti di distanza (Blondel et al. 1981), adottando una durata del rilevamento di 10 minuti (Dawson 1981; Fornasari et al. 2002; Fuller & Langslow 1984; Gutzwiller 1992), eseguiti una sola volta, indicativamente dal 20 maggio al 10 luglio, nelle fasce orarie comprese dall’alba alle 11.00 e dalle 18.00 al tramonto, e registrando separatamente i contatti entro un raggio prefi ssato (100 metri) da quelli esterni a tale raggio (Hutto et al. 1986). Considerando le dimensioni delle aree oggetto di studio e l’orografi a del territorio indagato si è ritenuto opportuno prevedere che il rilevatore possa distribuire uniformemente i punti d’ascolto, eventualmente lungo percorsi preesistenti quali sentieri e strade, distanziando i singoli punti di almeno 500 metri l’uno dall’altro. Durante le attività di campo, al fi ne di poter mettere in relazione la presenza e l’abbondanza delle specie alle caratteristiche ambientali, viene stimata nel raggio di 100 metri dal punto di rilevamen-to la presenza percentuale delle diverse variabili ambientali secondo le categorie CORINE Land Cover di terzo livello. Nel corso dei rilevamenti effettuati negli anni 2000-2012, in 1.213 punti d’ascolto sono state rilevate 26 variabili ambientali, in fase elaborativa accorpate in 11 categorie, che successive analisi di agglomerazione associano in otto cluster distinti, riconducibili ad am-bienti dominati da: mosaici agrari, boschi di latifoglie, boschi di conifere, boschi misti, praterie, brughiere e cespuglieti, macchia mediterranea ed aree rocciose. Per meglio comprendere le relazioni tra ornitocenosi ed habitat, si è inoltre proceduto alla defi -nizione della vocazione ambientale relativa alle principali specie nidifi canti, adottando una me-todologia analoga a quella applicata a livello nazionale per l’identifi cazione delle specie legate ad agroecosistemi ed ambienti boschivi (Rossi 2011). Il processo logico seguito nel corso delle analisi si può ricondurre ai seguenti punti: selezione preliminare dei taxa da considerare nelle analisi; calcolo del baricentro di distribuzione ambientale per ogni singola specie (Fornasari etal. 2002; Tellini et al. 2005); individuazione di gruppi di specie omogenei dal punto di vista delle preferenze ambientali (mediante analisi di agglomerazione; Gauch 1982; McGarigal et al. 2000); analisi delle caratteristiche ambientali a livello di comunità (analisi delle componenti principali e analisi della corrispondenza; Benzécri 1973; de Leeue & Mair 2007); selezione delle comunità rappresentative dei principali ambienti. Il set di dati considerato è stato relativo a 1.153 punti d’a-scolto eseguiti nel periodo 2006-2012.Gli andamenti annui dell’ornitocenosi nidifi cante nel periodo 2006-2012 sono stati valutati inclu-dendo nell’analisi esclusivamente le aree monitorate in almeno la metà degli anni di indagine. Per la selezione di tali aree si è quindi proceduto considerando le particelle UTM di 1x1 chilometri di lato dell’area di studio che siano state indagate almeno quattro volte su un totale di sette anni. Complessivamente l’area di indagine così selezionata comprende 87 particelle UTM di 1x1 chi-lometri di lato (media annua 78 ± 5,0; 70÷83) nelle quali sono stati effettuati 895 punti d’ascolto (media annua 128 ± 14,2; 115÷155). Per ulteriori confronti si è inoltre considerato il valore ornitologico nazionale delle specie nidifi -canti, che tiene conto di parametri eco-biologici, di vulnerabilità ed antropici (Brichetti & Gari-boldi 1992).
METHODES
La choix des méthodes effectuées à été dirigée par la conjugaison de deux nécessitées principales : être caractérisées par une haute répétabilité et comparabilité des détections et, au même temps, impliquer un engagement économique soutenable avec la prosecution des activités à longue terme. Pour la caractérisation et monitorage de l’ornithocenosis nidifiant, la technique de détection choisie a été celle des points d’écoute sans limitations de distance (Blondel et al. 1981), en adoptant une durée de détection de 10 minutes (Dawson 1981; Fornasari et al. 2002; Fuller & Langslow 1984; Gutzwiller 1992), effectués seulement une fois, substantiellement depuis 20 mai au 10 juillet, dans les tranches horaires comprises de l’aube jusqu’à 11 heures et de 18 heures au coucher de soleil, et enregistrant séparément les contacts entre un rayon préfixé (100 mètres ) des ces hors de dit rayon (Hutto et al. 1986).En considération des dimensions des aires objet d’étude et l’horographie du territoire sous enquête, on a retenu opportun prévoir que le capteur puisse distribuer uniformément les points d’écoute, éventuellement au long des parcours préexistants comme des sentiers ou routes, avec une distance entre chaque point d’écoute de au moins 500 mètres l’un de l’autre. Pendant les activités sur le champ, afin de pouvoir mettre en relation la présence et l’abondance des espèces aux caractéristiques du environnement, on a estimé entre 100 mètres du point de détection la présence en pourcentage des plusieurs variables d’environnement en accord avec les catégories CORINE Land Cover troisième niveau. Pendant le cours des détections effectuées dans les années 2000-2012, en 1.213 points d’écoute, on a identifié 26 variables d’environnement, qui en phase de calcul ont été unifiées en 11 catégories, qui les suivantes analyses d’agglomération ont réduit en 8 cluster différents, chacun reconductible à des environnements contrôlés par : mosaîques agraires, bois de feuillus, bois des conifères, bois mixtes, prairies, landes, aires ayant arbustes, macule méditerranée et aires rocheuses. Afin de mieux comprendre les relations d’ornithocenosis et habitat, on à en outre procédé à la définition de la vocation à l’environnement pour les principales espèces nidifiantes, en adoptant une méthodologie similaire à celle d’application nationale pour l’identification des espèces reliées à écosystèmes agraires et environnements à bois (Rossi 2011). Le processus logique suivi pendant les analyses peut être reconduit aux points suivants: sélection préliminaire des taxa à considérer dans les analyses; calcul du centre de distribution dans l’environnement pour chaque espèce (Fornasari et al. 2002; Tellini et al. 2005); individuation des groupes d’espèces homogènes par rapport aux préférences dans l’environnement (à moyen des analyses d’agglomeration; Gauch 1982; McGarigal et al. 2000); analyse des caractéristiques d’environnement à niveau de communaute (analyse des principales components et analyse de la correspondance; Benzécri 1973; de Leeue & Mair 2007); sélection des communautés répresentatives des principaux environnements. Le set de données considéré à été relié à 1.153 points d’écoute effectués pendant la période 2006-2012.
Les tendances annuelles de l’ornithocenosis nidifiant pendant la période 2006-2012 ont été évaluées en incluant dans l’analyse exclusivement les aires contrôlées et au moindre une moitié des années d’enquête. Pour la sélection de ces aires, on a donc procédé en prenant en considération les particules UTM de 1x1 kilomètres pour chaque coté de l’aire d’étude qui aient été soumises à enquête au moins quatre fois sur un total de sept années. Dans l’ensemble, l’aire d’enquête ici identifiée comprend 87 particules UTM de 1x1 kilomètres pour chaque coté (moyenne annuelle 78 ± 5,0; 70÷83) dans lesquelles ont été effectués 895 point d’écoute (moyenne annuelle 128 ± 14,2; 115÷155).
Pour toutes autres confrontations on a en outre considéré la valeur ornithologique nationale des espèces nidifiantes, qui tient compte des paramètres éco biologiques, de vulnérabilité et anthropiques (Brichetti & Gariboldi 1992).

30
METODI
Il monitoraggio delle specie target è stato effettuato adottando metodiche differenti, idonee alle esi-genze specifi che; di seguito vengono trattate in dettaglio le singole azioni attuate per l’espletamento di questo obiettivo. Il censimento di Falco pecchiaiolo, Biancone, Astore, Aquila reale, Falco pellegrino, Gufo reale e Picchio nero è fi nalizzato alla localizzazione delle coppie nidifi canti ed alla defi nizione dei relativi parametri riproduttivi; mentre per il Succiacapre si è scelto di effettuare il censimento crepuscolare al canto dei maschi in sette aree campione che, complessivamente, interessano una superfi cie di 47 Km2 (Aluigi et al. 2011). Il calcolo della densità riproduttiva, per tutte le specie precedentemente elencate, si è effettuato utilizzato il metodo della Nearest-Neighbour-Distance (Newton 1976). Pernice rossa, Tottavilla, Calandrella, Calandro, Codirossone, Magnanina comune, Cincia dal ciuf-fo, Averla piccola ed Ortolano sono stati censiti adottando il metodo dei transetti lineari con rile-vamento della distanza Distance Sampling (Buckland et al. 2001), le cui assunzioni fondamentali sono ampiamente descritte da Buckland et al. (1993), e la densità delle specie è stata calcolata con il software Distance 5.0 (Buckland et al. 2001; Thomas et al. 2005). Durante le stagioni riproduttive degli anni 2006-2012 sono stati quindi eseguiti 164 transetti per uno sviluppo complessivo di 527,4 chilometri.Il censimento delle coppie nidifi canti di Merlo acquaiolo è stato condotto percorrendo i greti di 25 corsi d’acqua dell’area di studio. Al fi ne di ottenere dati utili al monitoraggio delle popolazioni, i torrenti Erro, Gargassa, Lerone e Sansobbia sono stati monitorati in tutti gli anni indagati, mentre gli altri vengono censiti a rotazione. Ogni percorso è stato ripetuto tre volte da febbraio a maggio; tale frequenza, secondo D’Amico & Hemery (2003) consente di contattare tutte le coppie presenti con probabilità elevatissima. Le preferenze ambientali delle specie sono state valutate mediante l’indice di Jacobs (I = (N1/N - S1/S) / (N1/N + S1/S); dove N1 è il numero di soggetti osservati nell’ambiente 1, N è il numero totale di soggetti osservati nell’area di studio, S1 è la superfi cie dell’ambiente 1 nell’area di studio ed S la superfi cie totale dell’area di studio. Per il Succiacapre la formula utilizzata è invece: I = (X1/Y1 – X2/Y2) / (X1/Y1 + X2/Y2); in cui X1 è la superfi cie usata del tipo 1 nei territori, X2 è la super-fi cie totale del tipo 1 nell’area di studio, Y1 la superfi cie totale dei territori e Y2 la superfi cie totale dell’area di studio). L’indice è compreso tra valori di -1 e +1; assume valori positivi se l’habitat è selezionato, negativi se non viene utilizzato, uguale o vicino a 0 se è usato al pari della disponibilità.Sebbene la serie temporale sia ancora limitata e le risultanze dell’analisi vadano prudenzialmente considerate come semplici variazioni interannuali e non ancora trend delle popolazioni monitorate (che acquisiranno signifi catività proseguendo le indagini sul medio-lungo periodo), si è ritenuto utile testare statisticamente gli andamenti riscontrati. La valutazione di tali andamenti è stata quindi effettuata utilizzando il software TRIM (TRends & Indices for Monitoring Data), liberamente uti-lizzabile e scaricabile dal sito www.ebcc.info. Il software viene normalmente adoperato per stimare indici e andamenti di popolazione, ed è il principale strumento attualmente utilizzato dall’European Bird Census Council per il calcolo degli indici e degli andamenti dell’avifauna nidifi cante in Europa nell’ambito del programma di monitoraggio paneuropeo (Gregory et al. 2005). TRIM stima inoltre quale sia, nell’arco temporale considerato, la variazione percentuale media annua e la tendenza in atto. La classifi cazione della tendenza in atto viene effettuata su base statistica (calcolando l’erro-re standard e verifi cando dove si collocano i valori del limite inferiore e superiore dell’intervallo di confi denza rispetto al valore centrale), distinguendo tra aumento marcato, aumento moderato, stabilità, diminuzione moderata, diminuzione marcata, andamento non certo. Se non diversamente specifi cato, la tendenza delle singole specie è da considerarsi con andamento non certo.
METHODES
Le monitorage des espèces target a été effectué en adoptant des methodiques diverses, qui soient aptes aux exigences spécifiques ; ci-dessous sont décrites en détail les actions effectuées pour l’achèvement de cet objectif.
La recension de la Bondrée apivore, Circaète Jean-le-Blanc , Autour des palombes, Aigle royal, Faucon pélerin, Hibou grand-duc et Pic noir est finalisée à la localisation des couples nidifiantes et à la definition des paramètres de reproduction associés; pour le Engoulevent d’Europe on a préféré effectuer la recension crépusculaire au chant des masculins dans sept aires échantillon qui, dans l’ensemble, concernent une surface de 47 Km2 (Aluigi et al. 2011). Pour le calcul de la densité de reproduction, pour toutes les espèces précédemment listées, on a utilisé le modèle Nearest-Neighbour-Distance (Newton 1976).
Perdrix rouge, Alouette lulu, Calandrella, Pipit rousseline, Monticole de roche, Fauvette pitchou, Mésange huppée, Pie-grièche écorcheur et Bruant ortolan ont été recensés en adoptant la méthode des transepts linéaires avec la détection à distance Distance Sampling (Buckland et al. 2001), dont les hypothèses de base sont largement décrites par Buckland et al. (1993), et la densité des espèces a été calculée avec le logicielDistance 5.0 (Buckland et al. 2001; Thomas et al. 2005). Pendant les saisons de reproduction des années 2006-2012 ont été effectuées 164 transepts pour une développement total de 527,4 kilomètres. La recension de couples nidifiantes de Cincle plongeur a été effectuée en parcourant grèves de 25 courses d’eau dans l’aire d’étude. Afin d’obtenir des données utiles au monitorage des populations, les torrents Erro, Gargassa, Lerone et Sansobbia ont été objet de monitorage pendant toutes les années d’enquête, tandis que les autres sont recensés à rotation. Chaque parcours a été répété trois fois depuis février jusqu’à mai ; cette fréquence, selon D’Amico & Hemery (2003) consent le contact avec toutes les couples présentes avec une très haute probabilité. Les préférences du environnement des espèces ont été évaluées avec l’index de Jacobs (I = (N1/N - S1/S) / (N1/N + S1/S); où N1 est le numéro de sujets observés dans l’environnement 1, N est le numéro total de sujets observés dans l’aire d’étude, S1 est la surface de l’environnement 1 dans l’aire d’étude et S est la surface totale de l’aire d’étude. Pour l’Engoulevent d’Europe la formule utilisée est celle qui suit: I = (X1/ Y1 – X2/Y2) / (X1/Y1 + X2/Y2); dont X1 est la surface utilisée de la typologie 1 dans les territoires, X2 est la surface totale de la typologie 1 dans l’aire d’étude, Y1 est la surface totale des territoires et Y2 est la surface totale de l’aire d’étude. L’index est compris entre les valeurs -1 et +1; il assume des valeurs positives si l’habitat est sélectionné, négatives si n’est pas utilisé, égal ou proche à zero s’il est utilisé tel que la disponibilité.Bien que la série temporale soit encore limitée et les résultantes de l’analyse doivent être pour précaution considérées comme simples variations intra-annuelles et ne pas encore un trend des populations monitorisées (qui acquerront significativité dans le suivi des enquêtes sur une période moyenne ou longue), on a retenu utile tester statistiquemente les tendances trouvées. L’évaluation de ces tendances a été donc effectuée en utilisant le logiciel TRIM (TRends & Indices for Monitoring Data), librement utilisable et téléchargeable du site www.ebcc.info. Le logiciel est typiquement utilisé pour éstimer index et évolutions de population, et il est le principal outil aujourd’hui exploité par l’European Bird Census Council pour le calcul des index et évolutions de la communauté des oiseaux nidifiants en Europe dans le milieu du programme de monitorage paneuropéen (Gregory et al. 2005). TRIM effectue une estimation ce qui soit, dans la période du temps prise en examen, la variation en pourcentage moyenne annuelle et la tendance en cours. La classification de la tendance est effectuée sur une base statistique (avec le calcul de l’erreur standard et en vérifiant où sont placées les valeurs limites inférieure et supérieure de l’intervalle de confiance par rapport à la valeur centrale), en distinguant entre une augmentation marquée, une augmentation modérée, stabilité, diminution modérée, diminution marquée, tendance non certaine. Si non diversement specifié, la tendance de chaque espèce est à considérer avec une évolution non certaine.

31
LA MIGRAZIONE
Fin dall’antichità l’uomo ha provato grande attrazione verso alcuni comportamenti ani-mali, primo fra tutti la migrazione. Infatti, il fenomeno per cui queste creature compiano lunghi spostamenti per poi occupare di nuovo le terre lasciate, resta ancora oggi affasci-nante e, per alcuni aspetti, non del tutto chiarito. L’area del Beigua è ormai nota da molti anni per la sua importanza nella migrazione dei Rapaci diurni, ma non tutti sanno che, estendendosi su un ampio arco stagionale, vi transitano migliaia di individui appartenenti a tante altre specie.
Dal punto di vista ecologico, possiamo distinguere gli uccelli in due categorie principa-li. La prima è rappresentata da specie che lasciano l’Europa prima dell’inverno, raggiun-gendo terre più calde e con maggiori disponibilità trofi che per superare la brutta stagione. La seconda categoria comprende quelli che, spostandosi attraverso l’Europa, raggiungono aree che offrono condizioni di svernamento più favorevoli, quali ad esempio il bacino del Mediterraneo e le coste atlantiche di Spagna, Francia e Gran Bretagna. La differenza tra queste due categorie, che scelgono regioni diverse per lo svernamento, è strettamente infl uenzata dal tipo di alimentazione delle specie. Gli uccelli che si trattengono in Europa e Nord Africa (ad esempio: Pettirosso, Merlo, tordi, Capinera, Luì piccolo, Cincia mora...) possono trovare facilmente cibo di cui nutrirsi, come vegetali (semi, gemme…) oppure invertebrati lungo le spiagge. Al contrario, gli uccelli che si spingono fi no a sud del Sa-hara (dove sverna oltre il 40% delle specie nidifi canti nell’Eurasia) o in Asia sono quasi tutti totalmente o principalmente insettivori. A questa seconda categoria appartengono, ad esempio, alcune delle specie target del Parco del Beigua come il Calandro, svernante nella zona del Sahel ed in Asia sud-occidentale, ed il Codirossone, che trascorre l’inverno nell’Africa a sud del Sahara arrivando a sud-est fi no alla Tanzania. Ad esse si aggiungono poi varie altre specie più o meno comuni, quali Gruccione, Torcicollo, Rondine, Codirosso comune, Sterpazzolina comune e di Moltoni, Beccafi co, Balia nera, etc.
Inoltre il fenomeno della migrazione riguarda anche specie che non nidifi cano in Ligu-ria, in alcuni casi neppure nel territorio italiano, ed in molti casi gran parte delle popola-zioni in transito, pur appartenenti a specie nidifi canti nel nostro territorio, sono costituite da soggetti destinati ad insediarsi al di fuori dei confi ni nazionali; fatti per cui i migratori rappresentano un bene comune a livello, quantomeno, europeo.
Colombacci in migrazione
LA MIGRATION
Depuis ancien temps l'homme a ressenti une grande attraction pour certaines comportements animaux, tout d'abord la migration. En effet, le phénomène par lequel ces créatures parcourent distances longues pour en suite reprendre la terre originaire, reste encore aujourd’hui fascinant, à certains égards, pas tout à fait clair. La région du Beigua est bien connue par de nombreuses années pour sa importance dans la migration des oiseaux de proie diurnes, mais pas tout le monde sait que, en étendant sur un large arc de saison, des milliers d’individus d'autres espèces transitent par là. Du point de vue écologique, on peut distinguer les oiseaux en deux catégories principales. La première est représentée par des espèces qui ont quitté l'Europe avant l'hiver, en atteignant les terres plus chaudes ayant une plus grande disponibilité trophique pour surpasser la mauvaise saison. La deuxième catégorie comprend les oiseaux qui, se déplaçant à travers l'Europe, arrivent aux zones qui offrent des conditions plus favorables d'hivernage, comme par exemple le bassin de la Méditerranée et des côtes de l'Atlantique de le Espagne, la France et la Grande-Bretagne. La différence entre ces deux catégories, qui choisissent régions différentes pour l'hivernage, est étroitement influencée par le type d'alimentation des espèces. Les oiseaux qui restent en Europe et en Afrique du Nord (par exemple, Robin, Merle noir, le Labre merle, Fauvette à tête noire, Pouillot véloce, Mésange noire ...) peuvent facilement trouver de la nourriture qu'ils mangent, comment les plantes (graines, bourgeons ...) ou des invertébrés le long des plages. Au contraire, les oiseaux qui vont au sud du Sahara (où hivernent plus de 40% des espèces nidifiantes d’Eurasie) ou en Asie sont presque tous totalement ou principalement insectivores. A cette catégorie appartiennent les secondes, par exemple, certaines des espèces cible du Parco del Beigua comme le Pipit rousseline, hivernant dans la région du Sahel et dans le sud-ouest de l'Asie, et le Monticole merle de roche, qui passe l'hiver dans l'Afrique au sud du Sahara pour arriver à sud-est jusqu'à atteindre la Tanzanie. S'ajoutent à cela les diverses espèces, puis d'autres plus ous moins fréquentes, telles que le Guêpier d'Europe, Torcol fourmilier, Avale, Rougequeue à front blanc, Fauvette commune et de Moltoni, Fauvette des jardins, Gobemouche noir, etc. En outre, le phénomène de la migration affecte également espèces qui ne nidifient pas en Ligurie, dans certains cases, néanmoins sur le territoire italien, et dans de nombreux cas, la plupart des individus en transit, bien que appartenants à des espèces nidifiantes dans notre région, sont composés des individus destinés à s'installer dans dehors des frontières nationales ; ces faits sont lesquels dont les oiseaux migrants sont un bien commun à niveau au moins européenn.

32 ATTIVITÀ DI RICERCA, CENSIMENTO E MONITORAGGIO
LA MIGRAZIONE DEI RAPACI DIURNI
Da circa quindici anni la migrazione dei ra-paci rappresenta, nel panorama ornitologico nazionale, un ambito tematico di notevole in-teresse ed in costante evoluzione, nel quale i contrafforti appenninici di Arenzano, uno dei siti “storici” sin dall’inizio dei primi controlli sulla migrazione pre-riproduttiva risalenti alla metà degli anni ottanta del secolo scorso (Ba-ghino & Leugio 1989, 1990; Baghino 1996; Zalles & Bildstein 2000), continuano ad es-sere luogo di conteggi sistematici e standar-dizzati tali da offrire una serie temporale di dati relativamente estesa. Attualmente il sito aderisce al Coordinamento Nazionale Rapa-ci Migratori; negli anni si sono inoltre tenuti contatti giornalieri con campi di osservazione nella vicina Francia (regione di Nizza e Costa Azzurra: Bellet e Eze-La Turbie), in Lom-bardia (Lago di Garda, BS) e soprattutto in Toscana (Alpi Apuane, LU) sempre con inte-ressanti e preziosi scambi di informazioni.
In primavera, i rapaci in migrazione sui rilievi montuosi del versante tirrenico si vedo-no arrivare sostanzialmente da sud-ovest, parallelamente alla costa e su un fronte di circa tre- quattro chilometri d’ampiezza, dirigendosi prevalentemente in direzione est (Ponente di Genova), ma anche nord-est; in autunno si osserva uno schema analogo sebbene di direzione opposta, con una primaria direttrice migratoria da est-nord-est verso ovest-sud-ovest. Anche nell’ambito di una stessa specie, la direzione dei vari contingenti, oltre a dipendere della meteorologia locale (forza e provenienza del vento, altezza della coper-tura nuvolosa), può però rifl ettere le diverse destinazioni geografi che delle popolazioni di passaggio: una sorta di invisibile “autostrada nel cielo”, con svincoli e raccordi, uscite e rientri, deviazioni e interruzioni.
Grazie al notevole sforzo di campionamento, sono attualmente 29 le specie di rapaci segnalate per il Parco del Beigua nel sito di Arenzano. Tenendo conto del fatto che il numero di uccelli osservati è in realtà un rapporto tra quelli veramente passati e quelli contati, nel Beigua il volume stagionale della migrazione primaverile dei rapaci può es-sere stimato, considerando tutte le specie, intorno ai 6.000-9.000 individui, con fl uttua-zioni numeriche da un anno all’altro anche considerevoli; la migrazione post-riproduttiva (agosto-novembre) è invece numericamente più limitata.
Le specie di seguito descritte vengono riportate in ordine di abbondanza, dalle più rap-presentate quantitativamente a quelle numericamente più scarse e/o irregolari.
Il Falco pecchiaiolo risulta la specie più abbondante tra quelle censite durante la mi-grazione pre-riproduttiva: attualmente i conteggi permettono di stimare un passaggio an-nuo di circa 4-5.000 soggetti, ed un andamento tendente alla stabilità. Non è nota con esattezza la destinazione, all’interno dell’Europa continentale, di questi contingenti in
Falco pecchiaiolo
LA MIGRATION DES RAPACES DIURNES
Depuis une quinzaine d'années la migration des oiseaux de proie représente, dans le milieu ornithologique national, un domaine thématique d'un intérêt considérable et en constante évolution, où les contreforts des Apennins d’Arenzano, l'un des sites «historiques» depuis le début des premiers contrôles sur la migration pre-reproduction depuis les années quatre-vingt du siècle dernier (Baghino & Leugio 1989, 1990; Baghino 1996; Zalles & Bildstein 2000), continuent à être un lieu de comptages systématiques et standardisés, fournissent une série chronologique de données rélativement élargie. Aujourd'hui le Site adhère à la Coordination Nationale Oiseaux de Proie migratoires ; au fil des années des contacts quotidiens avec les champs d’observation en France (région de Nice Côte d'Azur et Bellet et Eze-La Turbie), Lombardie (Lac Garda BS) et surtout en Toscane (Alpes Apuanes, LU) ont été maintenus, et ont permis toujours des échanges intéressants et précieux de renseignements. Au printemps, les oiseaux de proie en migration sur les montagnes de la côte tyrrhénienne sont vu arriver sensiblement à partir du sud-ouest, parallèle à la côte et sur une front d'environ trois à quatre kilomètres d’amplitude, principalement vers est (Ponente à Gênes), maïs aussi au nord-est ;en automne, on observe une tendance similaire, bien que dans la direction opposée, avec une primaire directrice de migration d'est-nord-est à l'ouest-sud-ouest. Même au sein d’une single espèce, la direction des divers contingents, de plus de la dépendance selon la météorologie locale (force et origine du vent, la hauteur des nuages), peut refléter les différents destinations géographiques des populations de passage, une sorte de invisible "autoroute dans le ciel", avec des jonctions et raccords, sorties et entrées, déviations et interruptions. Grâce à l'effort d'échantillonnage considérable, ils sont aujourd’hui 29 les espèces de oiseaux de proie signalés pour le Parco del Beigua parc sur le site de Arenzano. En tenant compte du fait que le nombre d'oiseaux observés est en fait un rapport entre ceux réellement passés et ceux comptés, dans le Parco del Beigua le volume de la migration saisonnière du printemps des oiseaux de proie peut être estimé, compte tenu de toutes les espèces, en environ 6000-9000 d’individus avec des fluctuations numériques d'année en année considérables ; la migration post-reproductive (Août à Novembre) est numériquement plus limitée. Le espèces ci-dessous décrites sont indiquées selon un ordre d’abondance, à partir des plus représentées quantitativement à ces numériquement inférieures et/ou irrégulières. La Bondrée apivore résulte comme l’espèce la plus abondante entre celles recensées pendant la migration pre-reproductive : aujourd’hui les comptages permettent d’estimer un passage annuel d’environ 4-5.000 individus, et une évolution qui tend à la stabilité. N’est pas connue avec certitude l’exacte destination, dans le milieu de l’Europe continentale, de ces contingents en migration à mai, que vraisemblablement, ou au moins en partie, sont interceptés sur la côte de Brescia du Lac de Garda (Micheli & Leo 2010). De cette espèce, qui fait registrer à niveau qualitatif des concentrations assez plus relevantes que des autres sites-clé pour la migration pre-reproductive (par exemple le Stretto di Messina Giordano et al. 2012), on sait que la migration post-reproductive dans le Parco del Beigua, ayant le pic entre fin d’Août et le début de septembre, est numériquement marginale par rapport au considérable et bien documenté flux qu’on peut observer, dans la même periode, entre l’Italie nord orientale et les Alpes occidentales (Gargioni et al. 2007; Mezzavilla et al. 2003, 2008; Toffoli & Bellone 1996; Belfiore & Giraudo 2012): en Arenzano on observe en proportion plus des exemplaires jeunes (environ 90% en septembre), environ 17-20 jours plus en retard par rapport à les adultes (Baghino 2008; Schmid 2000).

33
migrazione a maggio, che verosimilmente, o almeno in parte, vengono intercettati sulla sponda bresciana del Lago di Garda (Micheli & Leo 2010). Di questa specie, che fa re-gistrare sul piano quantitativo concentrazioni molto più rilevanti in altri siti-chiave per la migrazione pre-riproduttiva (ad es. lo Stretto di Messina; Giordano et al. 2012), si sa che la migrazione post-riproduttiva nel Parco del Beigua, con picco tra fi ne agosto e inizio settembre, è numericamente marginale rispetto al cospicuo e ben documentato fl usso che si osserva, nel medesimo periodo, tra l’Italia nord-orientale e le Alpi occidentali (Gargio-ni et al. 2007; Mezzavilla et al. 2003, 2008; Toffoli & Bellone 1996; Belfi ore & Giraudo 2012): ad Arenzano vengono osservati in proporzione più giovani (circa il 90% in settem-bre), più tardivi di circa 17-20 giorni rispetto agli adulti (Baghino 2008; Schmid 2000).
12141618
zion
e
Migrazione pre-nuziale
02468
101214
1988 1991 1994 1997 2000 2003 2006 2009 2012
Indi
ce d
i mig
razi
one
01988 1991 1994 1997 2000 2003 2006 2009 2012
Anno
Biancone Falco pecchiaiolo
2 5
3,0
3,5
one
Migrazione post-nuziale
0 0
0,5
1,0
1,5
2,0
2,5
3,0
Indi
ce d
i mig
razi
one
0,0
0,5
2000 2001 2002 2003 2004 2005 2006 2007 2008 2009 2010 2011 2012
I
Anno
Biancone Falco pecchiaiolo
Andamento delle popolazioni di Biancone e Falco pecchiaiolo monitorate durante le migrazioni pre- [a] e post-nuziali [b]; in entrambi i periodi il Biancone tende ad un aumento marcato, mentre la tendenza del Falco pecchiaiolo risulta stabile in primavera e statisticamente non certa in autunno.
METODI
Il monitoraggio delle popolazioni in transito di Rapaci diurni e Ciconiformi si è svolto in forma standardizzata mediante la copertura continua dei periodi individuati, conformemente ai program-mi di conteggio dei rapaci in migrazione nei colli di bottiglia indicati da Brambilla et al. (2001).Per questa azione le specie target sono rappresentate da Biancone e Falco pecchiaiolo; negli ultimi anni considerati (dal 2008-2009 ad oggi), durante la migrazione pre-riproduttiva il primo è censito dall’8 al 29 marzo ed il secondo dal 5 al 20 maggio, ed entrambi dall’8 al 29 settembre per ciò che riguarda la migrazione post-riproduttiva.I dati di presenza qui riportati si riferiscono alla globalità dei conteggi effettuati a partire dal 1985; per il numero medio e le estrapolazioni delle stime annuali si sono invece impiegati i risultati dal 2004 al 2012 incluso. Gli andamenti delle popolazioni sono stati valutati utilizzando il softwareTRIM (vedi pag. 30).Le osservazioni sono compiute guardando ad ogni altezza del settore di provenienza dei rapaci e realizzate con l’ausilio di binocoli ad ampio campo visivo (ad es. 7x42) e, per osservazioni di dettaglio o a più forte distanza, di cannocchiali (32x77) e binocoli (15x70) montati su treppiede.Per quanto riguarda le origini delle popolazioni si è fatto riferimento prevalentemente a Spanò e Truffi (1987), Vorišek & Formanek (1996) e Spina & Volponi (2008a).
[a] [b]
METHODES
Le monitorage des populations des oiseaux de proie diurnes en transit et des oiseaux cigogniformes s’est développé en façon standardisée avec une couverture continue des périodes identifiées, conformement aux programmes de comptage des oiseaux de proie en migration dans les cols de bouteille indiqués par Brambilla et al. (2001). Pour cette action les espèces cibles sont représentées par le Circaète Jean-le-Blanc et la Bondrée apivore; dans les dernières années considérées (depuis 2008-2009 à aujourd’hui), pendant la migration pre-reproductive le premier est recensé depuis 8 jusqu’à 20 mars et le deuxième depuis 5 jusqu’à 20 mai, et touts les deux depuis 8 jusqu’à 29 septembre en ce qui concerne la migration post reproductive. Les données de présence ici rapportées se référent à la globalité des comptages effectués à partir de 1985 ; pour le nombre moyen et les extrapolations des estimation annelles, au contraire, on a utilisé les resultats à faire compte de 2004 à 2012 inclus. Les évolutions des populations ont été évaluées en utilisant le logiciel TRIM (voir page. 30). Les observations sont accomplies en observant chaque hauteur du secteur de provenance des oiseaux de proie et sont réalisées au moyen des jumelles à grand champ visuel (par ex. 7x42) et, pour les observations de détail ou à plus grande distance, avec des lunettes (32x77) et jumelles (15x70) installés sur le tripode. En ce qui concerne les origines des populations on a fait référence principalement à Spanò et Truffi (1987), Vorišek & Formanek (1996) et Spina & Volponi (2008a).
Evolution des populations Circaète Jean-le-Blanc et Bondrée apivore controlées pendant la migration pre [a] et post-nuptiales [b]; en toutes les deux les périodes le Circaète Jean-le-Blanc tend à une augmentation marquée, par contre la tendance de la Bondrée apivore se montre stable en printemps et statistiquement non certaine en automne.

34 ATTIVITÀ DI RICERCA, CENSIMENTO E MONITORAGGIO
Il Biancone è in assoluto la specie più intensamente studiata, oggetto del maggior nu-mero di indagini e approfondimenti scientifi ci (Baghino 2009), oltre ad essere quella che, per importanza quali-quantitativa, caratterizza di più il fenomeno migratorio dei Rapaci nel Parco del Beigua. I risultati dei conteggi di marzo indicano un fl usso consistente, con oltre 1.500 soggetti censiti annualmente. Tale ordine di grandezza contribuisce a far rivedere in senso estensivo, attualizzandole, le stime della popolazione nidifi cante italia-na (Baghino & Premuda 2005; Baghino 2009); generalmente i bianconi in transito nel Ponente ligure in marzo sono infatti soggetti adulti destinati ad insediarsi in gran parte, ma non esclusivamente, nella penisola italiana fi no alle regioni centro-meridionali, un dato interessante riguarda infatti un soggetto dotato di trasmettitore GPS, osservato nel marzo 2011 ad Arenzano ed insediatosi poi in Croazia (Baghino 2011). È da sottolineare che la migrazione del Biancone non si esaurisce nel solo mese di marzo con gli adulti riproduttori (data mediana 15/3), ma prosegue ad aprile e anche a maggio con individui immaturi. A settembre il fenomeno è numericamente meno consistente che a marzo, fatto probabilmente legato all’utilizzo di differenti direttrici migratorie di una parte considere-vole delle popolazioni del Nord Italia (Ruggieri et al. 2006; Baghino et al. 2012); in 13 stagioni consecutive, ed in un periodo campione di 12 giorni incentrato sul picco migra-torio della specie, sono stati conteggiati mediamente 732 ind. (circa 80% adulti, 16-17% giovani nati nell’anno in corso e il resto soggetti immaturi). Varie e interessanti sono le evidenze che, a seguito di studi specifi ci sulla migrazione post-riproduttiva, svolti in più siti e in simultanea, sono emerse a riguardo di questa specie:
a) uno schema migratorio denominato “a circuito” (Agostini et al. 2002a) che interessa anche gli immaturi (Premuda et al., 2010): le popolazioni di Biancone in movimento at-traverso la Liguria risalgono la Penisola partendo da molto più a sud di quanto si potesse immaginare (Mellone et al. 2011) per raggiungere i quartieri di svernamento africani passando per lo Stretto di Gibilterra;
b) una strategia migratoria di tipo conservativo (Agostini et al. 2002b); c) una strategia migratoria differenziata tra giovani e adulti (Agostini et al. 2004);d) le interconnessioni tra la topografi a e il ruolo del vento (Panuccio et al. in stampa).
Biancone
Le Circaète Jean-le-Blanc est en absolu l’espèce la plus intensivement étudiée, objet de la plus grande partie des enquêtes et études scientifiques (Baghino 2009), et est aussi quelle que, à la cause de sa importance qualitative et quantitative, caractérise plus le phénomène migratoire des oiseaux de proie dans le Parco del Beigua. Les résultats des comptages de mars indiquent un flux consistent, avec plus de 1500 individus recensés par année. Cet ordre de grandeur contribue à faire corriger en sens extensif, les estimations de la population nidifiante italienne en rendent celles-ci mises à jour (Baghino & Premuda 2005; Baghino 2009); generalement les Circaètes Jean-le-Blanc qui passent sur l’ouest de la Ligurie en mars sont en effet sujets adultes destinés à s’installer pour la plus grande partie, mais ne pas exclusivement, dans la péninsule italienne jusqu’aux régions centrales et méridionales, un renseignement intéressant concerne en effet un sujet ayant un émetteur GPS, observé en mars 2011 en Arenzano et installé en suite en Croatie (Baghino 2011). Il faut sousligner que la migration du Circaète Jean le Blanc ne pas termine seulement entre mars avec les adultes reproducteurs (date moyenne 15/03) mais poursuit en avril et même en mai avec des individus immatures. A septembre le phénomène est numériquement moins consistent qu’en mars, ce fais étant probablement relié à l’utilisation de directrices migratoires différentes par une partie significative des populations du nord de l’Italie (Ruggieri et al. 2006; Baghino et al. 2012); en 13 saisons consecutives, et pendant une période échantillon de 12 jours centrée sur le pic migratoire de l’espèce, on a compté en moyenne 732 ind. (environ 80% adultes, 16-17% jeunes nés dans la dernière année et le reste étant d’individus immatures). Plusieurs et intéressantes sont les évidences qui, après des études spécifiques sur la migration post reproductive, ces études étant effectués en plusieurs sites et en simultanée, sont émergées à l’égard de cette espèce: a) un schéma migratoire « à circuit » (Agostini et al. 2002a) qui concerne aussi les immatures (Premuda et al., 2010): les populations de Circaète Jean-le-Blanc en migration à travers la Ligurie, remontent la péninsule à partir de beaucoup plus à sud de ce qui on pouvait imaginer (Mellone etal. 2011) pour arriver aux quartiers d’hivernage africains en transitant pour le détroit de Gibraltar; b) une stratégie migratoire de type conservatif (Agostini et al. 2002b); c) une stratégie migratoire differenciée entre les jeunes et les adultes (Agostini et al. 2004); d) les interconnexions entre la topographie et le rôle du vent (Panuccio et al. En cours de pubblication). L’évolution des populations de Circaète Jean-le-Blanc apparaît donc clairement positive, avec une tendance à l’augmentation marquée soit en connexion à la migration du printemps que à la migration d’automne.

35
L’andamento delle popolazioni di Bianco-ne appare poi nettamente positivo, con ten-denza all’aumento marcato sia in relazione alla migrazione primaverile da che a quella autunnale.
Classifi cabile come la terza specie in ordi-ne di abbondanza, il Falco di palude mostra un periodo di migrazione primaverile piutto-sto dilatato, con un picco che si colloca nor-malmente tra gli ultimi cinque giorni di mar-zo e la prima decade di aprile, ed un numero medio di oltre 200 ind. a stagione; in autunno il picco è invece intorno al 20 settembre e la migrazione prosegue blandamente nel mese di ottobre, con il massimo conteggio stagio-nale di 78 soggetti nel 2008. L’areale di nidifi cazione dei contingenti che transitano per la Liguria occidentale è riconducibile prevalentemente a Repubblica Ceca e Germania.
Più diffi cile quantifi care la migrazione del Gheppio in considerazione di fattori quali l’esteso periodo di migrazione, le diffi coltà di avvistamento che interessano i rapaci di minori dimensioni e l’elevato numero di individui di incerta attribuzione specifi ca tra Gheppio e Grillaio (86% del totale nel periodo 2004-2012). In primavera la migrazione è compresa tra marzo e maggio, con picco localizzabile ai primi di aprile ed una stima di 3-500 individui annui. In questo caso sono principalmente coinvolti uccelli provenienti da Paesi dell’Europa centrale ed orientale. Scarsamente rappresentativa è invece la mi-grazione autunnale nell’Arenzanese. Sebbene oggettivamente molto più scarso e infre-quente (mediamente 1-2 ind./anno), va qui collegato al Gheppio, per ragioni di affi nità tassonomica, l’assai simile Grillaio, la cui identifi cazione sul campo è stata limitata ai soli maschi adulti rilevati in condizioni ottimali di osservazione.
Il Nibbio bruno, anch’esso regolare e ben rappresentato, mostra, come il Gheppio, un periodo di migrazione primaverile decisamente ampio, tale da abbracciare le due fi nestre temporali adottate per i conteggi del Biancone e del Falco pecchiaiolo, con un fl usso più pronunciato verso aprile e maggio riconducibile a soggetti immaturi, mentre in marzo transitano esclusivamente adulti (Premuda & Baghino 2012). La migrazione post-ripro-duttiva di questa specie è distribuita tra la fi ne di luglio e settembre con un picco noto, di solito scarsamente percepibile in Liguria, che è situato intorno alla prima metà di agosto.
Dal punto di vista quantitativo l’Aquila minore costituisce una peculiarità. Migra-trice regolare in primavera ed autunno ma di fatto molto scarsa fi no al 2004 (media annuale 1985-1994: 1 ind./anno; Premuda & Baghino 2004), ha destato notevole clamore nell’ambiente ornitologico italiano e france-se il particolare caso di migrazione di massa avvenuto nell’Ottobre 2004, con oltre 500 aquile minori arrivate in pochi giorni in Ita-lia dalla Penisola iberica attraverso la Francia mediterranea (Guillosson et al. 2006; Premu-da et al. 2007). Per effetto di quell’inusuale fenomeno (peraltro ripropostosi a distanza di sette anni, nel 2011, con analoghe modalità e abbondanze) e dei movimenti da esso de-
Falco di palude
G. C
arra
ra
Aquila minore
R. P
edem
onte
Etant classifiable comme la troisième espèce comme abondance, le Busard des roseaux montre une période de migration en printemps plutôt agrandie, avec un pic qui se place typiquement entre les dernières cinq journées de mars et la première décade d’avril, et un nombre moyen de plus de 200 ind. à la saison; en automne le pic est environ le 20 du septembre et la migration poursuit légèrement à octobre, avec le comptage maximal de la saison de 78 sujets en 2008. L’aire de nidification des contingents transitent par la Ligurie occidentale, est reconductible substantiellement à la Republique tchèque et Allemagne. Plus dificile resulte quantifier la migration de la Crécerelle, à la cause de facteurs comme l’étendue période de migration, la difficulté de repérage qui intéressent les oiseaux de proie ayant petites dimensions et l’haute nombre d’individus d’incertaine attribution specifique entre le Crecerelle et le Faucon crécerellette (86% du total dans la période 2004-2012). En printemps la migration est comprise entre mars et mai, avec un pic identifiable au début d’avril et une estimation de 3-500 individus par année. En ce cas sont principalement impliqués des oiseaux de l’Europe centrale et orientale. Faiblement representative est au contraire la migration automnale dans la zone d’Arenzano. Bien que objectivement assez plus rare et peu fréquent (typiquement 1-2 ind./année), vaut il relié a la Crécerelle, pour des raisons d’affinité taxonomique, le vraiment similaire Faucon crecerellette, dont l’identification sur le champ a été limitée seulement aux masculins adultes identifiés en conditions d’observation optimales. Le Milan noir, lui aussi régulier et bien représenté, montre, même que la Crécerelle, une période de migration en printemps bien vaste, suffisante à embrasser les deux fenêtres temporales adoptées pour les comptages du Circaète Jean-le-Blanc et de la Bondrée apivore, avec un flux migratoire plus évident vers avril et mai reconductible à sujets immatures, pendant que en mars seulement des adultes transitent tout à fait (Premuda & Baghino 2012). La migration post reproductive est distribuée entre fin de juillet et septembre avec un pic connu, typiquement difficilement identifiable en Ligurie, qui est situé environ à la première moitié d’août. Au point de vue quantitatif, l’Aigle botté constitue une particularité. Migratrice régulière en printemps et automne, mais en effet assez rare jusqu’à 2004 (moyenne annuale 1985-1994: 1 ind./année; Premuda & Baghino 2004), l’Aigle botté a éveillé beaucoup de clameur dans le milieu de l’ornithologie italien et français pour le singulier cas de migration massive arrivé dans l’octobre du 2004, avec plus de 500 aigles bottés qui sont arrivé en quelques journées en Italie à partir de la péninsule ibérique à travers de la France méditerranée (Guillosson et al. 2006; Premuda et al.2007). A la suite de ce phénomène singulier (par ailleurs arrivé une nouvelle fois après sept années, en 2011, avec mêmes modalités et abondance) et des mouvements par lui derivés, le volume de migration en printemps de l’espèces augmentait de plusieurs ordres de grandeur ; ce, surtout dans le mois de mars, lors du transit des sujet adultes (arrivés à partir d’une moyenne de 1 ind./période avant 2005 jusqu’à environ 21 ind. successivement). Aujourd’hui on peut estimer, en printemps, au moins une centaine d’individus qui se bougent des quartiers d’hivernage de la Méditerranée centrale (dans le cas spécifique, l’Italie méridionale; Baghino et al. 2007) vers l’aire de nidification de l’Europe occidentale, en montrent un sens de migration opposé à celui-là des autres espèces identifiées dans le site. Même évolution, avec une direction migratoire cette fois opposée, a été observé dans la deuxième partie de septembre : des 1,6 ind/année individués jusqu’à 2004, on est passés à une moyenne de plus de 48 ind./année depuis 2005 au 2012.

36 ATTIVITÀ DI RICERCA, CENSIMENTO E MONITORAGGIO
rivati, il volume di migrazione primaverile della specie è aumentato di vari ordini di grandezza; questo soprattutto nel mese di marzo, quando a transitare sono soggetti adulti (saliti da una media di 1 ind./periodo prima del 2005 a quasi 21 ind. successivamente). Attualmente si possono stimare, in primavera, almeno un centinaio di individui che si spostano dai quartieri di svernamento del Mediterraneo centrale (nel caso specifi co l’Ita-lia meridionale; Baghino et al. 2007) verso l’areale di nidifi cazione nell’Europa occiden-tale, presentando un senso di migrazione opposto a quello delle altre specie rilevate nel sito. Analogo andamento, con direzione migratoria stavolta opposta, si è osservato nella seconda metà di settembre: da 1,6 ind./anno riscontrati fi no al 2004, si è passati ad una media di oltre 48 ind./anno dal 2005 al 2012.
Il Lodolaio è una specie di comparsa regolare senza però risultare particolarmente abbondante in primavera (in media 19 ind./anno, stima 100-200 ind./anno), ed irrilevante dal punto di vista numerico in autunno.
Tra gli altri migratori regolari ma nel complesso poco abbondanti fi gura lo Sparvie-re, come alcune altre specie (Poiana, Gheppio) di non sempre facile assegnazione alla categoria dei migratori per la contemporanea presenza dei soggetti appartenenti a locali coppie nidifi canti. Mediamente ogni anno vengono conteggiati 14 individui in primavera (stima: ca 150 ind./anno) e 12 in autunno, riconducibili a soggetti le cui aree di nidifi ca-zione si collocano nell’Europa centro-settentrionale e quelle di svernamento nel Medi-terraneo occidentale.
La Poiana presenta parziali analogie con lo Sparviere in fatto di schemi migratori e di numeri complessivi di passo, che risultano in entrambe le specie piuttosto modesti (in media: primavera 15 ind./anno, stima ca 100 ind./anno; autunno 6 ind./anno). In pri-mavera è uno tra i rapaci più precoci (date estreme: 15/2-15/5) ed appare più frequente intorno alla prima metà di marzo. Nella Poiana, la linea di passo che interessa la Liguria è piuttosto marginale e riguarda soggetti di probabile origine centro-europea. Inoltre sono da riportare otto avvistamenti di soggetti in migrazione a fi ne settembre fenotipicamen-te riconducibili alla forma sottospecifi ca vulpinus (Poiana delle steppe; Baghino 2008; Corso 2002).
Presente in modo discontinuo e in quantità fl uttuanti negli anni, il Falco cuculo è un piccolo rapace che compare scarsamente a primavera avanzata (date estreme: 4/4-26/5,
Baghino 1996) e con relativa maggior regolarità tra fi ne aprile-primi maggio, cioé, di norma, prima dei periodi usual-mente indagati per il Falco pecchiaiolo (in media 5,5 ind./anno, stima 50-200 ind./anno). Assai più irregolare, addirit-tura casuale, risulta fi nora la presenza ac-certata in autunno nel Genovese con due sole segnalazioni note, nel settembre del 1984 e del 2009.
L’Albanella minore, seppur regolare, è un migratore poco numeroso, che a metà anni ottanta-novanta del secolo scorso è stato rilevato in stagione primaverile in
un periodo tra il 28 marzo e il 19 maggio, e con un massimo giornaliero, eccezionale, di 12 soggetti il 27/4/1986 (Baghino 1996). In anni più recenti viene osservata ogni primavera
Falco cuculo
M. R
osso
Le Faucon hobereau est une espèce régulière qui, n’est pas particulièrement abondante au printemps (une moyenne de 19 ind. / année, estimée 100-200 ind. / année), et est insignifiant au point de vue numérique en automne. Parmi les autres types de migrateurs réguliers mais généralement pas très abondantes est présent l'Epervier, comme certaines autres espèces (Buzzard, Kestrel) qui ne sont pas toujours facilement attribuables à la catégorie des migrateurs à la cause de la contemporaine présence de sujets appartenant à couples nidifiantes locales. En moyenne, chaque année, on compte 14 individus au printemps (estimation : environ 150 ind. / année) et 12 à l'automne, en raison de sujets dont les zones de nidification sont situées dans l’Europe centrale ou du nord et ces de l’hivernage dans la Méditerranée occidentale. La Buse variable montre des analogies partiales avec l’Epervier en ce qui concerne les schémas migratoires et les nombres totales de pas, qui résultent pour toutes les deux espèces plutôt réduits (en moyenne : 15 ind./année en printemps, estimation 100 ind/année ; automne 6 ind./année). En printemps la buse est l’une des oiseaux de proie les plus précoces (dates externes : 15/2-15/5) et apparaît plus fréquemment autour de la première moitié de mars. Pour la Buse, la ligne de pas qui concerne la Ligurie est plutôt marginale et concerne des sujets de potentielle origine de l’Europe centrale. En outre, sont à signaler huit réperages de sujets en migration vers fin septembre qui sont phénotypiquement reconductibles à la forme sous-spécifique vulpinus (Buse vulpinus; Baghino 2008; Corso 2002). Présent en modalité discontinue et en quantités fluctuantes pendant les années, le Faucon kobez est un petit rapace qui apparaît rarement en printemps avancé (dates extrêmes 4/4-26/5, Baghino 1996) et avec une relative plus haute régularité entre la fin d’avril – début de mai, soit, typiquement, avant les périodes traditionnellement objet d’enquête pour la Bondrée apivore (en moyenne 5,5 ind./année, estimation 50-200 ind./année). Bien plus irrégulière, environ par hasard, est la présence jusqu'à présent confirmée en automne dans l’aire de Genova avec seulement deux détections connues, septembre 1984 et 2009. Le Busard cendré, bien que irrégulièr, est un migrateur peu nombreux, qui à moitié des années 80-90 du dernier siècle a été identifié pendant la saison du printemps dans la période entre 28 mars et 19 mai, et avec un maximal journalier, exceptionnel, équivalent à 12 sujets le 27/4/1986 (Baghino 1996). A fil des années les plus récentes, on a observé chaque printemps une moyenne de 2,9 ind./année et un maximale pour chaque saison de 5 individus en 2004.

37
con una media di 2,9 ind./anno e un massimo stagionale di 5 individui nel 2004. Con una media di meno di 3 individui conteggiati ogni primavera e quasi altrettanti
in autunno, il Falco pescatore è una presenza regolare ma quantitativamente scarsa in Liguria, dove transitano individui di origine prevalentemente scandinava in migrazione tra Africa ed Europa. In primavera mostra un periodo di migrazione abbastanza esteso tra marzo e maggio, in autunno è visibile tra agosto e settembre.
Uno schema fenologicamente simile, ma con volume di migrazione ancor inferiore, riguarda il Nibbio reale, con una media annua di circa due individui a marzo e tre a settembre.
Irregolare (forse anche in relazione ai periodi indagati) e molto scarso nei numeri è il Falco della regina, per il quale tra la fi ne di luglio e i primi giorni di settembre si os-servano in Liguria singoli soggetti non maturi, distanti dalle colonie nidifi canti sparse in svariate piccole isole del Mediterraneo dove nidifi ca.
Il Grifone compare in forma sempre meno occasionale, come nel resto della regio-ne, per effetto dei progetti di reintroduzione, soprattutto francesi, che portano giovani e immaturi a compiere movimenti di dispersione con presenze più frequenti tra aprile e giugno, quantifi cabili con una media di 1-2 individui all’anno (Calvini & Toffoli 2007; Borgo & Galuppo 2007).
Molto scarsa ed irregolare è l’Albanellareale, della quale, tra il 2004 e il 2012, sono stati osservati in migrazione a marzo tre sin-goli soggetti; va comunque considerato che la specie in Liguria compare in numero esi-guo anche in inverno (Baghino 1996, Baghi-no 1998) e la sua migrazione precede o segue i periodi normalmente indagati. Analoga posizione è quella del Falco pellegrino con, dal 2000, nove osservazioni di soggetti migranti nella seconda metà di settembre (Baghino 2008); mentre del tutto casuale è la comparsa dell’Astore, più tardivo.
Tra le altre rarità si possono segnalare l’Aquila anatraia minore, osservata con rela-tiva regolarità negli ultimi anni con un numero ristrettissimo (0-2) di individui giovani provenienti da Paesi dell’Est europeo, e la notevole Aquila anatraia maggiore, che compare ancor più irregolarmente con effettivi altrettanto esigui tra la fi ne di ottobre e metà novembre e nella prima metà di marzo.
Decisamente occasionale e raro nel sito di Arenzano, e per il quale valgono le con-siderazioni fatte a proposito dell’Albanella reale, è lo Smeriglio, con un totale di otto osservazioni dal 1985 al 2012 (Baghino 1996).
Irregolare e molto rara è pure la comparsa dell’Aquila di Bonelli i cui pochissimi dati di presenza si riferiscono a singoli individui giovani o immaturi avvistati in erratismo a fi ne aprile-metà maggio e a settembre (Baghino & Leugio 1990; Baghino 2007; Baghino 2008b, Alessandria & Boano 2011, Baghino ined.).
L’Albanella pallida è un migratore rarissimo di passaggio tra l’Africa e gli ambienti di steppa dell’Europa Orientale: capita irregolarmente tra la fi ne di marzo e i primi d’a-prile e sono note tre segnalazioni distribuite tra il 1985 e il 2012 per un totale di cinque individui, dei quali ben tre nel solo marzo 2011 (Baghino & Leugio 1990; Baghino 2011).
La Poiana codabianca è stata documentata due volte (26/9/2001, Baghino 2008a; 17/10/2011, R. Pedemonte oss.pers.).
Grifone
R. P
edem
onte
Avec une moyenne moindre de 3 individus comptés chaque printemps et environ la même chiffre en automne, le Balbuzard pêcheur est une présence régulière mais quantitativement rare en Ligurie, où transitent individus d’origine substantiellement scandinave en migration entre l’Afrique et l’Europe. En printemps il y a une période de migration assez étendue entre mars et mai, en automne est visible entre août et septembre. Un schéma phénologiquement similaire, mais ayant un volume de migration encore inférieure, concerne le Milan Royal, avec une moyenne annuelle d’environ deux individus en mars et trois en septembre. Irrégulier (peut être en relation aux périodes sous enquête) et assez rare est le Faucon d’Eleonore,dont entre fin de juillet et début de septembre on observe en Ligurie des sujets solitaires non maturés, distant des colonies nidifiantes qui sont présentes en plusieurs petites îles de la Méditerranée, où il nidifie. Le Vautour fauve apparaît toujours en façon moins occasionnelle, comme dans le reste de la région, à la cause des projets de réintroduction, surtout françaises, qui emmènent les jeunes et les immatures à effectuer mouvements de dispersion avec des présences plus fréquentes entre avril et juin, quantifiables en une moyenne d’un ou deux individus par année (Calvini & Toffoli 2007; Borgo & Galuppo 2007). Assez rare et irrégulièr est le Busard Saint Martin, dont entre 2004 et 2012, on a observé en migration en mars seulement trois sujets singles; il faut en tout cas considérer que l’espèce en Ligurie apparaît en nombre réduit en hiver aussi (Baghino 1996, Baghino 1998), et sa migration précède ou suit les périodes traditionnellement sous enquête. La même position est celle du Faucon pélerin avec à partir de l’année 2000, dont ont été effectuées neuf observations de sujets migrant dans la deuxième partie de septembre (Baghino 2008); totalement par hasard arrive l’Autour des palombes, plus tardif. Parmi les autres raretés on peut signaler l’Aigle pomarin, observée avec une relative régularité dans les années dernières avec un nombre très réduit (0-2) d’individus jeunes qui arrivent de l’Est de l’Europe, et la Aigle criard, qui apparaît encore plus irrégulièrement avec des effectifs autant rares entre la fin d’octobre et moitié de novembre et la première moitié de mars. Substantiellement occasionnel et rare dans le site d’Arenzano, et dont valent les mêmes considérations faites à propos de la Busard Saint Martin, est le Faucon émerillon, ayant un totale d’huit observations entre 1985 et 2012 (Baghino 1996). Irrégulière et assez rare est aussi l’apparition de l’Aigle de Bonelli dont on a très peux renseignements de présence qui se referent à des individus singles, jeunes ou immatures, vus en façon erratique à fin april – début mai et en septembre (Baghino & Leugio 1990; Baghino 2007; Baghino 2008b, Alessandria & Boano 2011, Baghino ined.). Le Busard pâle, est un migrateur très rare qui passe entre l’Afrique et les environnements de la steppe de l’Europe orientale : arrive irrégulièrement entre la fin de mars et début d’avril et seulement trois épisodes distribués entre 1985 et 2012 sont connus, pour un total de cinq individus, dont bien trois dans le mois de mars 2011 seulement (Baghino & Leugio 1990; Baghino 2011). La Buse féroce a été documentée deux fois (26/9/2001, Baghino 2008a; 17/10/2011, R. Pedemonte obs. pers.).

38 ATTIVITÀ DI RICERCA, CENSIMENTO E MONITORAGGIO
L’elenco delle specie a vario titolo accidentali, con segnalazioni più o meno recenti nell’Arenzanese, include anche uccelli avvistati una sola volta; tra questi fi gurano Ca-povaccaio (13/5/1990, Baghino 1996), Poiana calzata (14/9/05, Bottero 2006), Aquilaimperiale (9/10/2007, Bottero et al. 2008) e Lanario (29/5/2012, Baghino oss.pers.).
Nella tabella sottostante vengono riportate, per le specie principali, alcune informa-zioni aggiuntive relative alla migrazione primaverile (FF: MP) e/o autunnale (FF: MA).
Capovaccaio
Specie FF N° medio (±ES)
N° Min-Max stagionale
N° Max giornaliero
Datamediana
Dateestreme
Stimastagionale
Falcopecchiaiolo MP 2.941
±468,81.463 (2012)6.072 (2005)
2.751(12/5/2002) 13/5 14/3/2008
27/6/19924.000-5.000
BianconeMP 1.514
±211,4770 (2004)
2.250 (2010)830
(17/3/2010) 15/3 20/2/199019/5/2004 2.000
MA 802±97,1
280 (2003)1.552 (2012)
326(22/9/2008) 21/9 8/8/2001
21/10/2000 1.400
Falco di palude MP 283
±37,582 (2004) 501 (2011)
270(4/4/1990) - 1/3/1992
23/5/1988 400-800
Nibbiobruno MP 74
±12,340 (2006)
135 (2012)34
(13/5/2007) - 22/2/19879/6/1985 250-350
Aquilaminore
MP 14±3,9
2 (2004)47 (2011)
19(16/3/2008) - 10/3/2005
18/5/2005 100
MA 40±12,2
0 (2002)550 (2004)
~600 (2011)
~200(16/10/2004,17/10/2011)
- 15/8/201130/10/2004 100-150
G. C
arra
ra
La liste des espèces à titres diverses accidentelles, ayant des apparitions plus ou moins récentes dans la zone d’Arenzano, comprend aussi des oiseaux vus une seule fois ; entre eux apparaît le Vautour percnoptère (13/5/1990, Baghino 1996), Buse pattue (14/9/05, Bottero 2006), Aigle impérial (9/10/2007, Bottero et al. 2008) et le Faucon lanier (29/5/2012, Baghino obs.pers.).
Dans le tableau ci-dessous sont indiquées, pour les espèces principales, quelques informations en ajoute qui concernent la migration en printemps (FF: MP) et/ou en automne (FF: MA).

39
L’ATTIVITÀ DI INANELLAMENTO A SCOPO SCIENTIFICO
L’inanellamento a scopo scientifi co nac-que in Danimarca nel 1899, ad opera di H. C. C. Mortensen, in seguito ad esperimenti di marcaggio eseguiti su alcuni storni mediante anelli metallici recanti impressi numeri pro-gressivi ed il suo indirizzo al fi ne di ricevere lettere di segnalazione (Jenni 1999). Da allo-ra l’inanellamento è rapidamente evoluto in concreta tecnica di ricerca utilizzata in tutto il mondo e basata sul riconoscimento individua-le degli uccelli mediante anelli metallici di tipo e diametro adatti alle dimensioni ed abitudini delle differenti specie. In Italia, attualmente, tutte le attività vengono condotte, nel rispetto della L. 157/92 e suoi recepimenti regionali, secondo le specifi che dell’Istituto Superiore per la Protezione e la Ricerca Ambientale. Du-rante le attività di campo, su ogni individuo, vengono rilevati molti parametri (specie, ses-so, età, morfometrie...), che permettono di au-mentare le conoscenze su etologia e fi siologia delle specie catturate. I dati raccolti nelle dif-ferenti stazioni consentono inoltre di ottenere informazioni sulla struttura della comunità or-nitica e l’utilizzo dell’habitat. Soggetti marcati e poi ricatturati o trovati morti ci danno inoltre indicazioni su durata della vita e spostamenti defi nendo quindi le rotte migratorie e le aree di sosta, fornendo così indicazioni sulla fenologia spaziale e temporale della migrazione, ma anche dati utili per la creazione e la gestione di aree protette. Tali dati possono poi essere utilizzati per una serie di analisi statistiche basate sulle cosiddette tecniche CMR (cattura-marcatura-ricattura), che con-sentono di stimare dimensioni della popolazione, tassi di sopravvivenza (e mortalità), tassi di emigrazione, reclutamento di nuovi individui nella popolazione, spostamento tra siti e, soprattutto, di mettere in correlazione questi parametri con variabili ambientali. L’inanellamento, stimando i parametri legati a successo riproduttivo, sopravvivenza, im-migrazione ed emigrazione, diviene poi una tecnica essenziale per la comprensione dei cambiamenti nelle popolazioni ornitiche e le loro cause, e di conseguenza per identifi care le problematiche legate alla loro conservazione.
Da tempo questa attività viene svolta con regolarità nell’area di studio nelle stazioni Sorgenti del Cerusa e Passo del Turchino, che effettuano il monitoraggio durante la mi-grazione primaverile ed autunnale in un contesto ornitologico tra i più interessanti della Liguria (Aluigi et al. 2008; Realini 2002). Tra il dicembre 2004 ed il novembre 2005, nell’ambito del progetto “Parco del Beigua: un parco per la biodiversità – Conoscenza, tutela e valorizzazione della ZPS Beigua-Turchino”, in quattro differenti stazioni (Pava-
Assiolo fotografato presso la stazione di inanellamento del “Centro Ornitologico e di Educazione Ambientale” di località Vaccà (Arenzano)
L’ACTIVITÉ DE BAGUEMENT À BUT SCIENTIFIQUE
Le baguement à but scientifique naît en Danemark en 1899, par H. C. C. Mortensen, et à la suite des expérimentations de marquage effectuées sur certains étourneaux au moyen des anneaux métalliques ayant l’impression de chiffres progressives et son adresse afin de recevoir lettres d’identification (Jenni 1999). Depuis ce jour là, le baguement a rapidement évolue en une concrète technique de recherche utilisée partout dans le monde et basée sur l’identification individuelle des oiseaux au moyen d’anneaux métalliques de typologie et diamètre adaptés aux dimensions et habitudes de chaque espèce. En Italie, aujourd’hui, toutes les activités sont conduites, dans le respect de la Loi 157/92 et ses implémentations regionales, selon les spécifiques de l’Istituto Superiore per la Protezione e la Ricerca Ambientale. Pendant les activités sur le champ, pour chaque individu, on identifie plusieurs paramètres (espèces, sexe, âge, morphométrie...), qui permettent d’augmenter les connaissances sur l’éthologie et physiologie des espèces capturées. Les données rassemblées dans les plusieurs stations permettent en outre l’obtention d’informations sur la structure de la communauté des oiseaux et sur l’utilisation de l’habitat. Sujets marqués et en suite nouvellement capturés ou trouvés morts donnent en outre des indications sur la durée de la vie et sur leur mouvements, en définissant des routes migratoires et des aires de repos, en fournissant donc des indications sur la phénologie spatiale et temporale de la migration, mais aussi des données utiles pour la création et la gestion d’aires protégées. Ces données peuvent en outre être utilisées pour une pluralité d’analyses statistiques basées sur les soi-disant techniques CMR (capture – marquage – nouvelle capture), qui permettent l’estimation des dimensions de la population, les taux de survivance (et mortalité), taux d’émigration, recrutement de nouvels individus dans la population, mouvement entre les sites et, surtout, la mise en corrélation de ces paramètres avec les variables du environnement. Le baguement, en estimant les paramètres reliés au succès reproductif, survivance, immigration et émigration, devient une technique essentiale pour la compréhension des changements des populations d’oiseaux et leur causes, et par conséquent pour identifier les problématiques reliées à leur conservation. Ça fait longtemps que cette activité est effectuée régulièrement dans l’aire d’étude des stations Sorgenti del Cerusa et Passo del Turchino, qui effectuent le monitorage pendant la migration du printemps et d’automne en un contexte ornithologique parmi les plus intéressants de la Ligurie (Aluigi et al. 2008; Realini 2002). Entre decembre 2004 et novembre 2005, dans le milieu du projet “Parco del Beigua: un parco per la biodiversità – Conoscenza, tutela e valorizzazione della ZPS Beigua-Turchino”, en quatre différentes stations (Pavaglione, Passo del Faiallo, Costa Cerusa et “Centro Ornitologico e di Educazione Ambientale” en localité Vaccà) a commencé une activité adressée à la caractérisation de la communauté des oiseaux et à l’enquête des liens écologiques, aussi en fonction des plusieurs phases saisonniers, étendant donc le monitorage à toute l’année solaire. A partir de 2006, afin de contrôler les espèces des petits passereaux avec les populations en transit, on a activé la station site auprès le “Centro Ornitologico e di Educazione Ambientale” en localité Vaccà, en activant des enquêtes d’exploration de la migration en printemps et un monitorage plus détaillé de la migration automnal (août-octobre). En outre, en 2012 a été expérimentée une nouvelle station, localisée en proximité de l’aire humide de récent rétablissement du Pian della Badia; les résultats de cette action, qui a impliqué l’entier cycle annuel, permettent l’individuation du site comme le plus diversifié entre ceux à capture passive qui sont activés dans l’aire du Beigua.

40 ATTIVITÀ DI RICERCA, CENSIMENTO E MONITORAGGIO
glione, Passo del Faiallo, Costa Cerusa e “Centro Ornitologico e di Educazione Ambien-tale” di località Vaccà) è stata poi intrapresa un’attività orientata alla caratterizzazione della comunità ornitica ed all’indagine dei legami ecologici, anche in funzione delle di-verse fasi stagionali, estendendo quindi il monitoraggio a tutto l’anno solare. A partire dal 2006, al fi ne di monitorare le specie di piccoli Passeriformi con popolazioni in transito, è stata poi attivata la stazione sita presso il “Centro Ornitologico e di Educazione Ambien-tale” di località Vaccà, attuando indagini esplorative della migrazione primaverile ed un più dettagliato monitoraggio della migrazione autunnale (agosto – ottobre). Inoltre, nel 2012 è stata sperimentata una nuova stazione, localizzata in prossimità dell’area umida di recente ripristino del Pian della Badia; i risultati di questa azione, che ha interessato l’intero ciclo annuale, consentono di individuare il sito come il più diversifi cato tra quelli a cattura passiva attivati nell’area del Beigua.
Al termine di questo nono anno di indagine, iniziamo quindi a delineare la fenologia annuale delle specie catturate, e migliorare, con dati quantitativi, la caratterizzazione dell’ornitocenosi che contraddistingue i periodi di migrazione. Complessivamente sono state effettuate 1.547 catture e ricatture riconducibili a 1.383 individui appartenenti a 54 specie. I periodi di più intenso transito migratorio si collocano tra fi ne marzo e fi ne aprile e da metà settembre a fi ne ottobre, ascrivibili in larga misura al notevole numero di pettirossi. La maggior biodiversità si raggiunge invece durante i periodi di presenza dei migratori trans-sahariani, ed in particolare a fi ne aprile e fi ne agosto quando, nell’area, sostano gli individui che si dirigono ai quartieri di svernamento africani.
L’importanza del Parco del Beigua per la migrazione degli uccelli è confermata anche dall’elevato numero di ricatture note per l’area (Bendini 1981; Spina & Bendini 1983; Bendini & Spina 1990) [a]. Uno dei dati più interessanti è relativo ad un individuo di Balia nera inanellato come pulcino il 24/6/2004 a Siuntio (Finlandia; 60°16’N 24°19’E) e ricatturato in località Vaccà il 24/4/2008. La distanza tra le due località risulta essere di 2.046 chilometri, ma va tenuto conto che, nei 3 anni, 10 mesi e 5 giorni intercorsi tra inanellamento e ricattura, quest’individuo ha percorso per ben otto volte la strada che separa i suoi quartieri di nidifi cazione, localizzabili molto probabilmente in Scandinavia, e le aree di svernamento sub-sahariane della specie (Sahel e Golfo di Guinea).
[a]
Au terme de cette dernière neuvième année d’enquête, on commence donc à délinéer la phénologie annuelle des espèces capturées, en améliorant avec des renseignements quantitatifs la caractérisation de l’ornithocenosis qui caractérise les périodes de migration. Complexivement, ont été effectuées 1.547 captures et récaptures reconductibles à 1.383 individus appartenant à 54 espèces. Les périodes de plus intense transit migratoire se placent entre fin mars et fin avril et à partir de mi-septembre à la fin d’octobre, qui sont imputables substantiellement au significatif nombre de rouges-gorges. La plus grande biodiversité est achevée au contraire pendant les périodes de présence des migratoires trans-sahariens, et notamment à fin avril et fin août lorsque, dans l’aire, restent les individus qui se dirigent aux quartiers d’hivernage africains. L’importance du Parco del Beigua pour la migration des oiseaux est confirmée même par l’haut nombre de recaptures connues pour l’aire (Bendini 1981; Spina & Bendini 1983; Bendini & Spina 1990) [a]. L’une entre les données plus intéressantes concerne un individu de Gobe-mouche noir baguée lors qu’il était petit le 24/6/2004 à Siuntio (Finlandie; 60°16’N 24°19’E) et nouvellement capturé en localité Vaccà le 4/4/2008. La distance entre ces deux localités résulte être 2.046 kilomètres, mais il faut compter que, dans 3 années, 10 moises et 5 jours passés entre le baguement et la nouvelle capture, ce individu a parcouru bien huit fois la route qui sépare ses quartiers de nidification, qui sont localisables très probablement en Scandinave, et les aires d’hivernage sub-sahariennes de l’espèce (Sahel et Golfe de la Guinée).

41
I CHIROTTERI
Con almeno 1.116 specie attualmente descritte e suddivise in 18 Famiglie i Chirotteri, comunemente conosciuti come pipistrelli, costituiscono il secondo più grande Ordine della Classe dei Mammiferi rappresentando circa un quarto di questi, superati solo dai Roditori. Hanno conquistato tutti gli ambienti presenti sulla terra ad esclusione delle calotte polari, delle alte montagne, delle isole più lontane e del centro dei più grandi de-serti, grazie ad adattamenti unici in questa Classe di Vertebrati. Da almeno 55 milioni di anni, infatti, hanno evoluto delle capacità biologiche fuori dal comune nei Mammiferi: il volo, la visione nell’oscurità grazie all’ecolocalizzazione e l’ibernazione nelle regioni temperate.
In Italia sono attualmente presenti 35 specie, delle quali due non più segnalate in tempi recenti (Vespertilio dasicneme Myotis dasycneme e Rinolofo di Blasius Rhinolophusblasii), costituendo così il 30% dei Mammiferi italiani.
Dalle abitudini di vita notturne, i Chirotteri, necessitano di ecosistemi complessi dove trovare rifugi, aree di caccia e corridoi di volo per spostarsi a breve o lungo raggio a se-conda della fase del loro ciclo annuale.
La loro dieta è quasi esclusivamente
come ragni, opilionidi, scolopendre, ma alcune specie europee sono in gra-do di catturare più o meno frequente-mente alcuni vertebrati, quali piccoli passeriformi e pesci, come nei casi di Nottola gigante (Nyctalus lasiopetrus)e Vespertilio di Daubenton.
In tutte le specie europee il periodo degli accoppiamenti coincide con la tarda estate e l’autunno, e in molte situazioni si protrae nei quartieri poi occupati per il letargo. Questi sono spesso gli unici periodi in cui maschi e femmine condividono i ri-fugi; infatti, i parti avvengono in colonie riproduttive, costituite esclusivamente o quasi da femmine. Di norma è partorito un solo piccolo, talvolta due; i parti gemellari sono la regola in poche specie (Pi-pistrello nano, Pipistrello di Savi). In tutte le specie, con una sola eccezione, gli spermatozoi sono conservati nelle vie genitali femminili e la fecondazione è posticipata alla primavera: questi sono adattamenti che permettono di far nascere i piccoli quando la disponibilità di cibo è massi-ma; l’eccezione è il Miniottero (Miniopterus schreibersii),in cui la fecondazione avviene in autunno ed è invece ri-tardato l’annidamento dell’embrione. La gestazione dura in genere 4-6 settimane e altrettanto lo svezzamento, nei casi in cui si prolunga maggiormente, i piccoli sono indi-pendenti all’età di 6-8 settimane. La maturità sessuale è raggiunta all’età di un anno, con diverse eccezioni tra le femmine di molte specie, che possono essere sessualmen-te mature già nel primo autunno, a tre mesi di vita.
Pipistrello nano
Pipistrello di Savi
a base di insetti e di altri artropodi
LES CHIROPTÈRES
Avec au moins 1.116 espèces aujourd’hui décrites et classifiées en 18 Familles de Chiroptères, communément connus comme chauves-souris, constituent le deuxième plus grande Ordre de la Classe des Mammifères en représentant environ un quatrième de ces derniers et étant surpassées seulement par les rongeurs. Il ont conquis toutes les environnements présents sur notre planète en excluant des calottes polaires, des hautes montagnes, des îles plus lointaines et du centre des grands déserts, grâce à des adaptations uniques dans la Classe des Vertébrés. Depuis au moins 55 millions d’années, en effet, il ont évolué leurs capacités biologiques en manière extraordinaire pour les Mammifères: le vol, la vision dans l’obscurité avec l’écholocalisation et l’hibernation dans les régions tempérées. En Italie sont aujourd’hui presentes 35 espèces, dont deux ne sont plus signalées en temps récents (Vespertilio dasicneme Myotis dasycneme et Rinolofo di Blasius Rhinolophus blasii), en constituent donc le 30% des Mammifères italiens. Des habitudes nocturnes, les Chiroptères, nécessitent d’écosystèmes complexes où trouver réfuge, aires de chasse et couloirs de vol pour se déplacer à bref ou long rayon selon la phase du cycle annuel.Leur diète est quasi exclusivement composée par des araignées, opilions, scolopendres, mais certaines espèces européennes sont capables de capturer plus ou moins fréquemment des vertébrés, comme des petits moineaux ou poissons, comme pour la Grande Noptule (Nyctalus lasiopetrus) et le murin de Daubenton. Pour toutes les espèces européennes la période d’accouplement coincide avec la tarde été et l’automne, et en plusieurs situations, arrive aux quartiers plus tard occupés par l’hibernation. Ces derniers sont plusieurs fois les uniques périodes dont les mâles et les femmes partagent les refuges ; en effet les accouchements arrivent dans les colonies reproductives, composées exclusivement ou substantiellement par des femmes. D’habitude, naît seulement un exemplaire, quelque fois deux ; les accouchements gémellaires sont la règle seuelement de quelques espèces (Chauve-souris nain, Chauve-souris de Savi). Pour toutes les espèces, avec une single exception, les spermatozoïdes sont conservés dans les voies génitales féminines, et la fécondation est retardée au printemps : ces adaptations permettent de faire naître les petits lors que la disponibilité de nourriture est maximale ; l’exception est constituée par le Minioptère de Schreibers (Miniopterus schreibersii), dont la fécondation arrive en automne et par contre est retardé l’amorçage de l’embryon. La gestation dure en général 4-6 semaines et le même arrive pour le sevrage. Dans les cas où il se prolonge plus, les petits sont indépendants à l’âge de 6-8 semaines. La maturité sexuelle est achevée à l’âge d’un an, avec plusieurs exceptions entre les femmes de plusieurs espèces, qui peuvent être sexuellement matures déjà dans le premier automne, à trois mois de vie.

42 ATTIVITÀ DI RICERCA, CENSIMENTO E MONITORAGGIO
I maschi vivono isolati o in piccoli gruppi nel periodo primaverile-estivo, mentre in autunno gli individui di alcune specie (Nottole, alcuni Myotis, Pipistrello nano e Pipistrello albolimbato) difendono piccoli harem, costituiti in genere da non più di 10 femmine.
Il periodo invernale è trascorso in letargo, condizione che comporta una diminuzione di tutte le attività vitali, cioè delle frequen-ze respiratoria e cardiaca, della reattività del sistema nervoso, dell’attività musco-lare con conseguente abbassamento del-la temperatura corporea che può scendere fi no a - 5 °C. Con tale cambiamento fi sio-logico delle condizioni corporee, l’anima-le può vivere in stato d’inattività per interi mesi, superando così la stagione fredda consumando esclusivamente le riserve energetiche accumulate nei periodi di vita attiva. La scelta del rifugio per il letargo è importante: il luogo deve essere al sicuro
da eventuali predatori, privo di correnti d’aria, con temperature non eccessivamente bas-se (comprese tre 2-3° e 10 °C) e soprattutto con un buon livello d’umidità per impedire il disseccamento del patagio. Il letargo invernale può subire delle interruzioni per stimoli esterni, se disturbati gli animali possono spostarsi da una parte all’altra di uno stesso rifu-gio o anche cambiarlo, o per stimoli interni, come il raffreddamento eccessivo.
I trasferimenti dalla nursery al rifugio invernale implicano per i Chirotteri degli spo-stamenti, poiché raramente i due rifugi sono nel medesimo luogo; solitamente si muovo-no anche solo di pochi chilometri ma cambiano ambiente: si tratta quindi di movimenti di tipo ecologico e non geografi co. Le specie considerate sedentarie percorrono solitamente distanze che variano tra i 20 e 50 Km tra i quartieri estivi e quelli di svernamento. La maggior parte delle specie, tra cui molti Myotis, effettua migrazioni a breve distanza (da 50 a 300 Km), che non necessariamente hanno un orientamento geografi co preferenziale. Alcune popolazioni di Nottola comune, Nottola di Leisler, Pipistrello nano e Pipistrello di Nathusius intraprendono voli autunnali in direzione sud-ovest dai paesi nordici dove trascorrono l’estate per svernare in Francia, Svizzera e nelle aree mediterranee. In spe-cie che effettuano migrazioni su lunga distanza sono state riscontrate longevità inferiori mentre, in generale, i Chirotteri sono animali considerati molto longevi: parecchie specie, anche di piccole dimensioni, possono superare i 20 anni di vita.
I Chirotteri, essendo animali gregari per la maggior parte dell’anno, utilizzano rifugi comuni, in cui gli individui appaiono aggregati. Aree ricche di habitat diversi, nelle quali compaiono tutte le tipologie di rifugio, presentano le maggiori concentrazioni di pipi-strelli poiché offrono molteplici opportunità di ricovero e d’alimentazione. La maggior parte dei Chirotteri europei, in base alle diverse fasi del ciclo biologico, utilizza differenti tipi di rifugio. Quelli utilizzati nella stagione invernale hanno temperature che oscillano tra i 3° e i 9° C, non soggetti a variazioni di rilievo; tali condizioni favorevoli si trovano nelle grotte ricche d’anfratti, in svariati nascondigli all’interno di edifi ci vicino a camini o tubazioni calde, e nei grossi alberi cavi utilizzati in inverno solo da poche specie (Genere Nyctalus). I rifugi estivi utilizzati dai Chirotteri sono i più svariati: edifi ci, anfratti in muri e rocce, alberi e varie cavità. Questi ambienti devono essere indisturbati e protetti poiché
Nottola di Leisler
Les mâles vivent isolés ou en petits groups dans la période du printemps et de l’été, par contre en automne les individus de certaines espèces (Noctules, certains Myotis, Chauve-souris nain et Chauve-souris kuhlii) défendent leur petits harem, constitués en général par non plus de 10 femmes. La période hivernale est passée en hibernation, condition qui comporte une diminution de toutes les activités vitales, c'est-à-dire de la fréquence respiratoire et du cœur, de la réactivité du système nerveux, de l’activité musculaire avec le conséquent baissement de la température du corps qui peut atteindre jusqu’à - 5 °C. Avec tel changement physiologique des conditions du corps, l’animal peut vivre en état d’inactivité pour plusieurs mois, en pouvant surpasser la saison froide avec la consommation exclusive des ressources accumulées dans les périodes de vie active. Le choix du refuge pour l’hibernation est important : le lieu doit être à l’abri des prédateurs, sans courants d’air, avec des températures ne pas excessivement baisses (entre 2-3° et 10°C) et surtout avec un bon niveau d’humidité pour éviter le sèchement du patagium. L’hibernation hivernale peut subir des interruptions pour les stimulations externes et si dérangés les animaux peuvent se bouger d’une partie à l’autre du refuge, ou même lui changer, ou par des stimulations internes, comme un excessif refroidissement. Les transferts de la nursery vers le refuge hivernal impliquent pour les Chiroptères des déplacements, puisque rarement deux refuges sont placés dans le même lieu ; typiquement ils se déplacent de quelques kilomètres mais ils changent environnement : il s’agit de déplacements de type écologique et non géographique. Les espèces considérées comme sedentaires parcourent typiquement des distances qui varient entre 20 et 50 km entre les quartiers d’été et d’hivernage. La plu partie des espèces, dont plusieurs Myotis, fait des migrations à brève distance (de 50 à 300 Km), qui non nécessairement ont une orientation géographque preferentielle. Certaines populations de Noctule commune, Noctule de Leisler, Chauve-souris nain et Chauve-souris de Nathusius effectuent des vols automnaux en direction sud ouest à partir des pays nordiques où ils passent l’été pour hiverner en France, Suisse et dans les aires de la méditerranée. Les espèces qui effectuent migrations à longue distance sont caractérisées par une moindre longévité tandis que, en général, les chiroptères sont des animaux considérés comme vivants assez longuement : plusieures espèces, même de petite taille, peuvent surpasser les 20 ans de vie. Les Chiroptères, en étant des animaux grégaires pour la plus partie de l’année, utilisent des refuges communes, dont les individus apparaissent comme agrégats. Les aires riches d’habitats différenciés, dans lesquelles apparaissent toutes les typologies de refuge, présentent les plus grandes concentrations de chauves-souris puisque ils offrent plusieurs opportunités d’abri et nourriture. La plus partie des chiroptères européens, en raison des différentes phases du cycle biologique, utilise plusieures typologies de refuge. Celles qui sont utilisées en hiver et qui ont températures qui oscillent entre 3 et 9°C, ne sont pas sujetes à des variations significatives ; ces favorables conditions sont trouvées dans les grottes riches d’anfractuosités, dans les endroits et recès des constructions proche des conduites de la fumée ou conduites chaudes, et dans les grand arbres creux qui sont exploités en hiver seulement par peux espèces (Genre Nyctalus). Les refuges d’été qui sont utilisés par les chiroptères sont les plus différents: édifices, recès dans les murs et rochers, arbres et cavités diverses. Ces lieux doivent être non dérangés et protégés puisque hébergent des sites de réproduction, c'est-à-dire des lieux dont l’ensemble des femmes enceintes accouchent et élèvent ensemble les petits. Les mâles qui ont achevé la maturité sexuelle pendant l’été, vivent souvent seuls ou en petit groups dans des dortoirs séparés, qui sont exploités aussi pour l’accouplement.

43
ospitano i siti di riproduzione, cioè i luoghi in cui l’insieme di femmine gravide parto-riscono e allevano insieme i piccoli. I maschi che hanno raggiunto la maturità sessuale durante l’estate, vivono spesso da soli o in piccoli gruppi all’interno di dormitori separati, sfruttati anche per l’accoppiamento.
I Chirotteri europei, in base alla tipologia di rifugio scelta, possono essere distinti in specie troglofi le, specie antropofi le e specie fi tofi le; le preferenze individuali sono da mettere in relazione anche alle caratteristiche climatiche delle diverse località e alla stagione.
I Chirotteri troglofi li (Generi Rhinolophus, Miniopterus, Plecotus, e buona parte delle specie del genere Myotis) sono più o meno strettamente legate ad ambienti cavernicoli come grotte e miniere ed a fessure di pareti rocciose e scogliere. Tali specie trovano in questi rifugi particolari condizioni microclimatiche come temperature basse e costanti e gradi elevati di umidità dell’aria. In generale nelle grotte con notevole sviluppo, verso il fondo l’aria è più calda e la temperatura rimane più costante. La maggior parte delle specie si trova ad una distanza che varia dai 20 ai 60 metri dall’entrata. I Rinolofi di, che preferiscono temperature più elevate, occupano il fondo della grotta mentre il Barbastello (Barbastella barbastellus) si trova solitamente all’entrata delle cavità.
I Chirotteri antropofi li, come il Pipistrello albolimbato, il Pipistrello nano, il Serotino comune e parzialmente il Pipistrello di Savi, si sono adattati a utilizzare le possibilità offerte dalle costruzioni umane. Alcune specie, come il Vespertilio maggiore, utilizzano le ampie soffi tte con tetti d’ardesia, materiale che si scalda molto bene immagazzinando calore; non vengono invece occupati luoghi con correnti d’aria. Anche i sottotetti delle chiese si adattano molto bene come rifugi in quanto luoghi senza particolari disturbi. Molte specie, grazie alle dimensioni ridotte (Genere Pipistrellus), riescono a insinuarsi in fessure larghe 1,5 cm in muri molto spessi, costruiti in mattoni o pietre combacianti e cavi all’interno.
Rinolofo maggiore
Les Chiroptères européens, en accord à la typologie de refuge choisie, peuvent être distingués en espèces troglodytiques, espèces anthrophyles et espèces phytophyles; les préférences individuelles sont à mettre en rélation aussi avec les caractéristiques climatiques des localités et de la saison. Les Chiroptères troglodytes (Genres Rhinolophus, Miniopterus, Plecotus et une bonne partie des espèces du genre Myotis) sont plus ou moins strictement reliés aux environnements caverneux comme les grottes et les minières, les fissures parois rocheuses et les falaises. Ces espèces trouvent dans ces refuges des particulieres conditions de microclimat comme températures baisses et constantes, et hauts degrés d’humidité de l’air. En général, dans les grottes plutôt grandes, vers la partie plus profonde l’air est plus chaud et la température reste plus constante. La plus partie des espèces se retrouve à une distance qui variait des 20 aux 60 mètres de l’entrée. Les rhinolophes, qui préfèrent températures plus hautes, occupent le fond de la grotte, alors que le Barbastelle commune (Barbastella barbastellus) se retrouve typiquement à l’entrée des cavités. Les chiroptères anthropiens, comme le Chauve-souris de Kuhl, le Chauve-souris nain, la Sérotine commune et partiellement la Vespère de Savi, ont adaptés à utiliser les possibilités offertes par le constructions humaines. Certaines espèces, comme le Grand Murin, utilisent des grands greniers avec toits d’ardoise, un matériau qui chauffe assez bien en emprisonnant la chaleur ; ne sont pas occupés les lieux ayant des courants d’air. Aussi les greniers des églises s’adaptent bien comme refuges, en tant que sans particuliers dérangements. Plusieurs espèces, grâce aux dimensions réduites (Genre Pipistrellus), peuvent s’insinuer dans fissures larges 1,5 cm à l’intérieur de murs assez profonds, construits en briques ou pierres qui coïncidentes et creuses.

44 ATTIVITÀ DI RICERCA, CENSIMENTO E MONITORAGGIO
I Chirotteri fi tofi li, quali il Pi-pistrello di Nathusius ed i Generi Barbastella e Nyctalus, occupano le spaccature naturali degli alberi o quelle prodotte da un fulmine o dallo scavo di un picchio. I rifugi ri-poduttivi dei Chirotteri fi tofi li sono abbastanza piccoli a causa del limi-tato spazio disponibile. Quelli abitati dal genere Nyctalus sono facilmente identifi cabili poiché presentano una lunga e ben visibile striscia d’urina sotto l’ingresso in seguito all’abitu-dine di questi animali di svuotare la vescica prima di entrare nel rifugio.
Attualmente le conoscenze sulla chirotterofauna del Parco del Beigua sono relativamente buone, almeno rispetto ad altre aree protette italia-ne, anche se molti territori restano ancora da indagare e sono poco co-nosciuti gli aspetti ecologici relativi alle aree di caccia. Le specie segna-late sono 15 [a], che costituiscono il 43% di quelle nazionali e il 62% di quelle liguri.
Tutte le specie di Chirotteri con-siderate sono incluse nell’allegato IV della Direttiva Habitat; inoltre tra quelle presenti nel Parco del Beigua Rinolofo maggiore, Vespertilio di
Bechstein e Vespertilio maggiore/minore sono incluse anche nell’allegato II della stessa Direttiva. Dal punto di vista conservazionistico bisogna poi tenere conto che nelle ‘Liste Rosse’ ben 1/3 delle specie presenti sono considerate vulnerabili o in pericolo a livello nazionale ed il Vespertilio di Bechstein è vulnerabile a livello europeo.
I dati a disposizione evidenziano come le specie più comuni e diffuse siano in ordine decrescente, il Pipistrello albolimbato, il Pipistrello nano e il Pipistrello di Savi [a]. Que-sti sono Chirotteri prevalentemente antropofili nella scelta dei siti di rifugio, e piuttosto eclettiche e generaliste nell’utilizzo degli ambienti di caccia che sono spesso costituiti da aree in prossimità di abitazioni, di fonti luminose, parchi e giardini, anche se non disde-gnano ambienti ecotonali posti tra habitat erbacei e forestali.
Le specie più selettive nella scelta degli habitat di foraggiamento e dei siti di rifugio sono meno diffuse, e in alcuni casi piuttosto rare come il Rinolofo maggiore, la cui presen-za è in relazione ad ambienti sotterranei naturali ed artifi ciali, poco rappresentati nell’area protetta. Alcune di queste mostrano una diffusione piuttosto limitata, non tanto per la loro rarità, ma per l’estrema specializzazione ecologica, come ad esempio, il Vespertilio di Daubenton, chirottero tipicamente acquatico che utilizza in maniera quasi esclusiva
Molosso di Cestoni
Notola di Leisler
Tadarida teniotis
Nyctalus leisleri
V tili di
Vespertiliomaggiore /
minore
Nottola comune
Myotis myotis/oxygnatus
Nyctalus noctula
Pipistrello di Nathusius
Rinolofomaggiore
Vespertilio diBechstein
Pipistrellus nathusii
Rhinolophus ferrumequinum
Myotis bechsteinii
Vespertiliomustacchino
Serotinocomune
Nat us us
Myotis mystacinus
Eptesicus serotinus
Pipistrello di Savi
Orecchionecomune e
meridionale
Vespertilio di Daubenton
Hypsugo savii
Plecotus auritus e austriacus
Myotis daubentonii
Pipistrelloambolimbato
Pipistrello nano
Savi
Pipistrellus kuhlii
Pipistrellus pipistrellus
yp g
pambolimbato
0,0% 5,0% 10,0%
Pipistrellus kuhlii
Distribuzione (%)
[a]Pipistrellus kuhlii Chauve-souris de Kuhli Pipistrellus pipistrellus Chauve-souris nain Hypsugo savii Vespértillon de Savi Plecotus auritus et austriacus Oréillard commun et méridionale Myotis daubentonii Vespértillon de Daubenton Myotis mystacinus Vespertillon à moustaches Eptesicus serotinus Serotin communbe Pipistrellus nathusii Chauve-souris dieNathusius Rhinolophus ferrumequinum Grand rhinolophe fer à cheval Myotis bechsteinii Vespertllon de Bechstein Myotis myotis/oxygnatus Grand murin /petit murin Nyctalus noctula NoctuleTadarida teniotis Molosse de Cestoni Nyctalus leisleri Noctule de Leisler
Les chiroptères phytophiles, comme le Chauve-souris de Nathusius et les genres Barbastella etNyctalus, occupent les fentes naturelles des arbres ou quelles qui sont produites par les foudres ou par les pics. Les refuges reproductives des chiroptères phytophiles sont assez petits à la cause du limité espace disponible. Ceux qui sont habités par le genre Nyctalus sont facilement identifiables puisque répresentent une longue et bien visible ligne d’urine sous l’accès, à la suite de l’habitude de ces animaux de vider la vessie avant d’entrer dans le refuge. Aujourd’hui les connaissances sur la faune des chiroptères du Parco del Beigua sont relativement bonnes, au moins par rapport aux autres aires protégées d’Italie, bien que plusieurs territoires restent encore à controler et les aspects écologiques reliés aux aires de chasse sont peu connus. Les espèces signalées sont 15 [a], que constituent le 43% de ces nationales et le 62% de ces de la Ligurie.Toutes les espèces des chiroptères considérées sont inclues dans l’Annexe IV de la Directive Habitat; en outre entre quelles qui sont présentes dans le Parc del Beigua arrivent le Grand rhinolophe fer à cheval, le Vespertillon de Bechstein et le Grand murin /petit murin sont inclues même dans l’Annexe II de la même Directive. Au point de vue de la conservation il faut compter que dans les « listes rouges » bien 1/3 des espèces présentes sont considérées comme vulnérables ou en danger à niveau nationale et le Verspertillon de Bechstein est vulnérable à niveau européen. Les données disponibles montrent comme les espèces plus communes et diffuses soient en ordre décroissant le Chauve-souris de Kuhl, il Chauve-souris nain et il Chauve-souris de Savi [a]. Ces sont des chiroptères en prévalence anthropophyles dans le choix des sites de refuge, et plutôt éclectiques et généralistes dans l’exploitation des environnements de chasse qui sont souvent constitués des aires en proximité des habitations, des sources lumineuses, parcs et jardins, bien que ils ne écartent pas les environnements écotones placés entre habitats herbeux et forestières. Les espèces plus sélectives dans le choix des habitats de nourriture et des sites de refuge sont moins diffuses, et en quelque cas plutôt rares comme le Grand rhinolophe dont la présence est en relation aux environnements souterrains naturels et artificiels, peu représentées dans l’aire protégée. Certaines de ces espèces montrent une diffusion plutôt limitée, ne pas en termes de rareté, mais pour l’extrême spécialisation écologique, comme par exemple, le Vespertillon de Daubenton, chiroptère typiquement aquatique qui utilise en manière quasi exclusive les courses d’eau et les aires humides pour la chasse et les ponts ou les cavités des arbres en proximité des environnements aquatiques comme sites de refuge. Sa présence dans l’aire protégée est donc limitée aux courses d’eau, notamment ces des fonds des vallées avec une présence des parties à faible courant qui sont susceptibles pour la nourriture, constituée substantiellement par des diptères aquatiques comme les chironomidés et quelque petit poisson.

45
corsi d’acqua ed aree umide per la caccia e ponti o cavità arboree in prossimità di ambienti acquatici quali siti di rifugio. La sua presenza nell’area protetta è quin-di limitata ai corsi d’acqua, in particolare quelli di fondovalle con presenza di tratti a corrente debole idonei all’alimentazio-ne, costituita principalmente da ditteri acquatici quali i Chironomidi e, occasio-nalmente, da qualche piccolo pesce.
Apparentemente rare sono le specie strettamente forestali, ad esclusione de-gli orecchioni e del Vespertilio mustac-chino, la cui presenza si limita a poche località, o a singole segnalazioni come il caso della Nottola comune, della Nottola di Leisler e del Vespertilio di Bechstein. Tale rarità non è giustifi cata dalla limita-ta estensione di habitat idonei costituiti da ambienti forestali generalmente maturi, ampiamente diffusi nell’area di studio. Le po-che segnalazioni sono probabilmente imputabili ad un difetto di ricerca e indagini future potranno sicuramente dimostrare una presenza più diffusa.
La diversità ambientale del Beigua, caratterizzata da boschi maturi di differenti tipo-logie, alternati da vaste aree aperte costituite da aree prative di crinale o altri ambienti erbacei, con la presenza di edifi cati ed aree umide, consente quindi la presenza di diverse specie di Chirotteri offrendo molteplici siti di rifugio e ricche aree di alimentazione.
In particolare i boschi offrono siti di rifugio e aree di caccia per alcune specie come le nottole, gli orecchioni, il Vespertilio di Bechstein, già citate in precedenza, ma anche al Vespertilio maggiore, che caccia carabi nella lettiera, o al Pipistrello di Nathusius, le cui popolazioni migratrici provenienti dal nord Europa che utilizzano le aree forestali del Parco come habitat preferenziale nei mesi primaverili ed autunnali. Anche altre specie sfruttano gli ambienti boschivi per l’attività trofi ca, in particolare il loro margine, come il Rinolofo maggiore ed alcune specie gene-raliste spesso associate ad ambienti antropici quali il Pipistrello nano o il Pipistrello albo-limbato.
Gli ambienti erbacei, invece, costituisco habitat elettivi per l’alimentazione del Ve-spertilio minore, che si dedica alla caccia di Ortotteri Tettigonidi in estate, o alcune specie dall’alto volo quali il Pipistrello di Savi o il Molosso di Cestoni che catturano attivamen-te ditteri e, soprattutto, lepidotteri notturni. Orecchione
Molosso di Cestoni
Apparemment rares sont les espèces strictement forestières, avec l’exclusion des Oreillards communes et du Murin à moustaches, dont la présence est limitée à quelques localités, ou à quelque vue comme est le cas de la Noctule commune, de la Noctule de Leisler et du Vespertillon de Bechstein. Cette raréte n’est pas justifiée par une limitée extension de l’habitats apts constitués par les environnements forestières généralement matures, largement diffusés dans l’aire d’étude. Les peux informations sont probablement à imputer à un défaut de recherche et les futures enquêtes pourront bien sûr démontrer une présence plus diffuse. La diversité du environnement du Beigua, caractérisée par les bois maturs des différentes typologies, qui s’alternent entre des vastes zones ouvertes constituées par des aires à prairie sur les lignes de crêtes ou des autres milieux herbés, avec une présence de constructions et aires humides, consent donc la présence de plusieures espèces de Chiroptères en offrant plusieurs sites de refuge et riches aires d’alimentation. Notamment les bois offrent des sites de refuge et des aires de chasse pour certaines espèces comme les Noctules, les Oreillons, le Vespertillon de Bechstein, dèjà cités en précédence mais aussi au Vespertillon majeur, qui chasse des vivanettes dans la litière, ou le Chauve-souris de Nathusius, dont les populations migratoires arrivent du nord de l’Europee et utilisent les aires forestières comme habitat préférentiel des moises de printemps et automne. Aussi des autres espèces exploitent les environnements forestièrs pour l’activité trophique, notamment leur margine, comme le Grand rhinolophe fer à cheval et des autres espèces généralistes souvent associées aux environnements anthropiques comme le Chauve-souris nain ou le Chauve-souris de Kuhl. Les environnements herbeux, par contre, constituent des habitats d’élection pour la nourriture du petit murin, qui se dédie à la chasse des orthoptères tettigoniidae en été, ou certaines à haut vol comme Chauve-souris de Savi ou le Molosse de Cestoni qui capturent activement les diptères et, surtout, lépidoptères nocturnes.

46 ATTIVITÀ DI RICERCA, CENSIMENTO E MONITORAGGIO
I Chirotteri, data la loro particola-re biologia, sono generalmente molto sensibili alle trasformazioni ambien-tali: il basso tasso riproduttivo, i lun-ghi tempi di gestazione e di svezza-mento, la tendenza delle femmine a riunirsi in densi gruppi per il parto e l’allevamento, spiegano come singoli eventi, anche se limitati ad una sola colonia, possano avere effetti sull’in-tera popolazione di una regione.
Negli ultimi cinquant’anni è sta-ta registrata una forte diminuzione dell’abbondanza di questi Mammi-
feri che, come tutti gli animali, hanno due necessità fondamentali: rifugi per il riposo e disponibilità di cibo. Qualsiasi opera o azione che infl uenzi o modifi chi la quantità e la qualità di questi fattori viene risentita dalle comunità dei chirotteri presenti sul territorio.
Le cause della loro rarefazione sono state identifi cate in: alterazioni e distruzioni degli habitat forestali, essenziali per molte specie sia come
rifugio (per le specie fi tofi le) sia come luogo di foraggiamento, con la conseguente riduzione dell’entomofauna a causa dell’eliminazione di alberi morti o deperenti o la riduzione dello strato arbustivo di sottobosco;
alterazione e distruzione degli habitat acquatici, essenziali per le specie che si alimen-tano direttamente sull’acqua o per quelle che utilizzano gli ambienti riparali quali zone di caccia;
“banalizzazione strutturale” delle aree agricole, passando dalle forme di conduzione di tipo tradizionale (ecomosaici con piccoli appezzamenti coltivati, inframmezzati a siepi e relitti forestali) all’agricoltura meccanizzata e intensiva (estese aree unifor-memente coltivate) che ha portato alla riduzione dell’entomofauna da un lato e alla perdita di elementi “lineari” molto importanti per i pipistrelli in quanto utilizzati come linee di riferimento nei loro spostamenti tra i rifugi e le aree di foraggiamento;
chiusura (per motivi di sicurezza o turistici) degli ingressi di grotte, miniere e gallerie ostruendone l’accesso alle specie troglofi -le;
allontanamento di colonie di Chirotteri da vecchie case in via di ristrutturazione;
azioni di disturbo di origine antropica arre-cate alle colonie, sia in edifi ci sia in am-bienti sotterranei;
fenomeni di bioaccumulo da composti or-ganoclorurati, metalli pesanti e policlo-robifenili dovuti all’impiego di pesticidi, insetticidi e di altri trattamenti chimici in agricoltura e per i trattamenti del legno;
oscillazioni climatiche, anche di lieve enti-tà, che possono infl uire sulla disponibilità
Orecchione comune
Vespertilio minore
Les Chiroptères, vu leur particulière biologie, sont généralement très sensibles aux transformations du environnement : le bas taux de reproduction, les longs temps de grossesse et de sevrage, la tendance des femmes à se réunir en groupes denses pour l’accouchement et l’élevage, montrent comme les évents, bien que limités à une colonie single, puissent avoir des effets sur la population entière d’une region. Pendant les dernières cinquante années à été registrée une grande diminution de l’abondance de ces mammifères qui, comme tous les animaux, ont deux nécessitées fondamentales : les refuges pour le repos et disponibilité de nourriture. Toutes le œuvres ou actions qui influencent ou modifient quantité et disponibilité de nourriture sont ressenties par la communauté des chiroptères qui sont présents sur le territoire. Les causes de leur raréfaction ont été identifiées en: altérations et destructions des habitats forestières, qui sont essentiels pour plusieurs espèces soit comme refuge (pour les espèces phytophiles) soit comme lieu d’alimentation, avec la conséquente réduction de l’entomofaune à la cause de l’élimination des arbres morts ou en dépérissement ou la réduction de la couche d’arbustes du sous-bois; altération et destruction des habitats aquatiques, essentiels pour les espèces qui se nourrissent directement sur l’eau ou pour quelles qui utilisent les milieux d’abri comme zones de chasse; “banalisation structurale” des aires agricoles, en passant des formes de conduction de type traditionnel (mosaïques écologiques avec des petites parcelles cultivées et subdivisés par des haies ou zones forestières) à l’agriculture mécanisée et intensive (grandes aires uniformément cultivées) qui a emmené à la réduction de l’entomofaune d’une coté et à la perte d’éléments « lineairs » assez importants pour les chauves-souris étant utilisés comme des lignes de référence pour leur déplacements entre les refuges et les aires de nourriture ; clôture (pour questions d’hygiène ou de tourisme) des entrances des grottes, minières et galeries, en obstruant l’accès aux espèces troglodytes; éloignement des colonies des chiroptères des veilles maisons en phase de restructuration; action de dérangement d’origine anthropique ressenties par les colonies, soit dedans les édifices soit dans les milieux souterrains; phénomènes de bioaccumulation de composées organochlorés, métaux pesants et polychlorobiphényles dus à l’utilisation de pesticides, insecticide et des autres traitements chimiques en agriculture et pour les traitements du bois; variations climatiques, bien que de légère entité, peuvent affecter la disponibilité de nourriture, ou peuvent causer l’abandonnement du site dont on n’a plus les conditions de température et humidité qui sont nécessaires pour la survivance de la colonie;

47
di cibo, oppure possono causare l’abbandono del sito nel quale non si verifi cano più le condizioni di temperatura e umidità necessarie per la sopravvivenza della colonia;
inquinamento luminoso, che riduce le aree di foraggiamento idonee per le specie lu-cifughe, e determina l’abbandono dei rifugi in edifi ci soggetti ad illuminazione not-turna;
eolico industriale, che provoca mortalità diretta per urto contro le pale in movimento degli aerogeneratori o per barotrauma conseguente ai vortici d’aria e determina, in alcuni casi, un peggioramento della qualità ambientale nelle aree di foraggiamento conseguentemente all’apertura di nuove strade specialmente in ambienti forestali.
La conservazione dei Chirotteri ha risentito nel passato di una man-canza di conoscenza delle specie, situazione conseguente alla scar-sa contattabilità di questo gruppo. Oggi è invece chiaro come un’ef-fi cace strategia di conservazione debba tenere in considerazione la complessità dell’ecologia del taxonin questione, che si mostra altamen-te specie specifi ca e, nonostante la limitata mole degli animali, ricca di aspetti ed esigenze differenti nell’arco di un ciclo biologico. Le azioni di tutela devono quindi riguardare i punti qui elencati, che vanno considerati nella loro complessità e stretta dipendenza reciproca: conservazione dei siti di rifugio, la cui valenza conservazionistica aumenta conside-
rando anche l’eventuale presenza contemporanea di più specie e la notevole fi lopatria della componente femminile;
conservazione dei siti di svernamento, cui gli animali tendono ad essere fedeli anno dopo anno;
conservazione degli habitat di caccia, soprattutto per le specie maggiormente specia-liste nella selezione degli stessi e valutate le esigenze trofi che variabili nelle diverse fasi biologiche;
conservazione dei corridoi di volo adottati durante le migrazioni, soprattutto per le specie che compiono migrazioni a lungo raggio nello spostamento tra siti riproduttivi e di svernamento, e conservazione dei corridoi di volo utilizzati durante gli sposta-menti a corto raggio e durante le attività di caccia;
conservazione dei siti di rifugio temporanei, quali i siti di riposo diurno utilizzati du-rante le migrazioni, quelli utilizzati per le attività di aggregazione temporanea (swar-ming), cui è associato un ruolo fondamentale nello scambio genetico tra popolazioni, e quelli utilizzati dai maschi durante il periodo estivo.
È quindi evidente che qualsiasi azione di conservazione dei Chirotteri, affi nché sia ef-fi cace, deve sottintendere una buona conoscenza delle specie e dei popolamenti presenti in un’area.
Riferimenti bibliografi ci: Spagnesi & De Marinis (2002), Calvini (2006) e Toffoli (2011).
Vespertilio di Bechstein
pollution lumineuse, qui réduit les aires de nourritures aptes pour les espèces étant loin de la lumière, et qui cause l’abandonnement des refuges dans les édifices qui sont sujets d’illumination nocturne; éolienne industrielle, qui provoque une mortalité directe par collision contre les pales en mouvement des aérogénérateurs ou à la cause des barotraumatisme en conséquence des vortex d’air et qui cause, en certains cases, une aggravation de la qualité du environnement dans les aires d’alimentation en conséquence de l’ouverture de nouvelles routes notamment dans les milieux forestières. La conservation des chiroptères a ressenti en temps passé d’un manque de connaissance des espèces, en conséquence d’une réduite caractéristique d’identifiabilité de cette groupe. Aujourd’hui est par contre clair comme une efficace stratégie de conservation doit tenir compte de la complexité de l’écologie du taxon sous examen, qui se montre hautement spécifique de l’espèce et, malgré la limitée dimension des animaux, est riche d’aspects et exigences différents à fil d’un cycle biologique. Les actions de protection doivent donc concerner les point ci-dessous indiqués, qu’il faut considérer dans leur complexité et grande dépendance réciproque: conservation des sites de refuge, dont la valence de conservation augmente en considérant l’éventuelle présence contemporaine de plusieurs espèces et la grande philopatrie de la composante féminine; conservation des sites d’hivernage, dont les animaux tendent à être fidèles année après année; conservation des habitats de chasse, surtout pour les espèces plus fortement spécialistes dans la sélection de ces-ci et en considérant les exigences trophiques variables des phases biologiques; conservation des couloirs de vol adoptés pendant le migrations, surtout pour les espèces qui effectuent des migrations à long rayon dans le déplacement entre les sites de reproduction et d’hivernage, et la conservation des couloirs de vol utilisés pendant les déplacements à court rayon et pendant les activités de chasse; conservation des sites de refuge temporaires, comme les sites de repos diurne utilisés pendant les migrations, ces qui sont utilisés pour l’activité d’agrégation temporaire (swarming), dont est associé un rôle fondamental dans le changement génétique entre les populations, et ces qui sont utilisés par les mâles pendant la période de l’été. Il est donc évident que quelconque action de conservation des chiroptères, parce qu’elle puisse être efficace, doit impliquer une bonne connaissance des espèces et des populations qui sont présents dans chaque aire.
Références bibliographiques: Spagnesi & De Marinis (2002), Calvini (2006) et Toffoli (2011).

48 ATTIVITÀ DI RICERCA, CENSIMENTO E MONITORAGGIO
IL TRAPPOLAGGIO FOTOGRAFICO
Il monitoraggio delle specie animali basato seguendo le impronte su polvere, fango, sabbia o neve, è probabilmente il metodo più antico conosciuto per l’individuazione della presenza dei mammiferi in un’area. I conteggi di escrementi, tracce, vocalizzazione e l’osservazione diretta lungo transetti standard vengono ampiamente utilizzati per valuta-re sia la ricchezza che l’abbondanza delle specie animali. Tuttavia negli ultimi anni una nuova tecnica di rilevamento è diventata sempre più popolare: il trappolaggio fotografi co o fototrappolaggio, metodica quantitativa che ha costi relativamente bassi di lavoro, non è invasiva, ed implica un minimo disturbo ambientale.
Questa tecnica consiste nell’ottenere immagini (fotografi che o video) all’insaputa del soggetto e in assenza dell’operatore; si tratta quindi di una trappola che non tocca il soggetto, ma ne “cattura” semplicemente l’immagine. Ciò è possibile utilizzando un’at-trezzatura ottenuta associando ad una fotocamera un dispositivo (sensore di movimento) capace di far scattare automaticamente l’apparecchio al passaggio degli animali che vi passano davanti. Le trappole fotografi che sono effi cienti nel raccogliere dati sia di giorno che di notte e offrono la possibilità di ottenere informazioni addizionali sulla distribuzio-ne e sull’uso dell’habitat delle specie, sulla struttura di popolazione e sul comportamento.
Il metodo è applicabile ad un ampio spettro di situazioni, in quanto è adattabile a variazioni delle condizioni del terreno e del clima; sono quindi molteplici le possibili applicazioni nei campi della ricerca e della gestione faunistica, ed esse riguardano preva-lentemente la documentazione di aspetti eco-etologici di specie presenti a basse densità o particolarmente elusive, e in territori particolarmente diffi cili laddove altre metodologie siano ineffi cienti, acquisendo così dati inediti da integrare con quelli ottenuti mediante altre tecniche.
Nell’ambito delle ricerche previste nel Parco del Beigua, e ad integrazione ed appro-fondimento di quanto si sta già ottenendo con il progetto regionale “Il Lupo in Liguria”, a partire dalla fi ne di novembre del 2011 il trappolaggio fotografi co è stato adottato come tecnica principale per l’acquisizione di informazioni sulla presenza del Lupo.
Faina
“ ”
LES TRAPPES PHOTOGRAPHIQUES
Le monitorage des espèces animales qui se base sur la suite des empreintes sur la poussière, le boue, sable ou neige est probablement la méthode la plus ancienne connue pour l’individuation de la présence des mammifères dans une aire. Les comptages d’excréments, traces, vocalisation et observation directe au long des transepts standard sont amplement utilisés pour évaluer soit la richesse que l’abondance des espèces animales. Toutefois dans les dernières années une nouvelle technique de détection est devenue toujours plus connue: les trappes photographiques ou phototrappage, une méthodique quantitative qui montre coûts relativement baisses de travail, n’est pas invasive, et implique un dérangement minimale d’environnement. Cette technique consiste dans l’obtenir d’images (photographiques ou vidéo) à l’insu du sujet et en absence de l’opérateur; il s’agit donc d’une trappe qui ne touche pas le sujet, mais que capture simplement l’image. Ça est rendu possible en utilisant un équipage obtenu avec l’association d’une camera avec un dispositif (capteur de mouvment), capable d’activer automatiquement l’appareil lors du passage des animaux qui passent là devant. Les trappes photographiques sont efficientes pour le rassemblement de renseignements soit pendant le jour que la nuit et offrent la possibilité d’obtenir informations additionnelles sur la distribution et l’utilisation de l’habitat par les espèces, sur la structure de la population et sur le comportement. La méthode est applicable à un grand milieu de situation, parce que est adaptable aux variations des conditions du sol et du climat ; ils sont donc plusieurs les applications qui peuvent être appliquées dans les champs de la recherche et de la gestion faunistique, et elles concernent préférentiellement la documentation d’aspects écologiques et éthologiques d’espèces présentes en baisses densités ou particulièrement élusives, et dans les territoires qui sont difficiles où de autres méthodologies soient inefficaces, pour apprendre des renseignements neufs à intégrer avec ces qui sont obtenus par des techniques différentes. Dans le milieu des recherches prevues du Parco del Beigua, et comme integration et étude ulterieure de ce qui est déjà en cours d’obtention avec le projet régional “Il Lupo in Liguria”, à partir de la fin de novembre du 2011 les trappes photographiques ont été adoptées comme technique principale pour l’obtention de renseignements sur la présence du loup.

49
Nei settori dell’area d’indagine dove si è registrata una frequentazione continua da parte del predatore, o nei quali si voleva ve-rifi carne la presenza, sono stati individuati una serie di siti dove collocare le trappole fotografi che. La scelta delle stazioni è stata subordinata alle caratteristiche ambientali dell’area, al grado di disturbo antropico e al livello di frequentazione degli ambienti da parte degli animali selvatici, in primisil Lupo. Ogni stazione di rilevamento è stata cartografata e ciascuna fotocamera è stata dotata di un codice identifi cativo. Le trappole fotografi che sono state sottoposte, in relazione alle condizioni climatiche, a frequenti verifi che di funzionamento e di recupero delle immagini memorizzate, per-mettendo di ottenere dati inediti che suc-cessivamente sono stati integrati con quelli ottenuti con le altre tecniche di censimento. Durante il periodo d’indagine sono state conservate tutte le registrazioni foto/video ottenute e le informazioni collezionate da ciascuna apparecchiatura (specie animale monitorata, data e ora di rilevamento, etc.) inserite in uno specifi co archivio informa-tico.
In circa 13 mesi di attività (fi ne novembre 2011 – dicembre 2012) sono state attivate 100 stazioni di rilevamento (84 specifi che per il Lupo e 16 più generiche) che complessivamente hanno operato per 8.614 giornate (in media 662 giornate ogni mese) ottenendo complessivamente oltre 48.000 registrazioni, delle quali 9.348 riconducibili ad animali.
L’analisi di questi materiali ha permesso di identifi care 68 specie: 3 Anfi bi, 1 Rettile,
Ammotrago Capriolo
Trappola fotografi ca
Tasso
Dans les secteurs de l’aire d’enquête, lors qu’on registre une fréquentation continue par les prédateurs, ou dont on cherchait d’en vérifier la présence, ont été individués plusieurs sites où placer les trappes photographiques. Le choix des stations a été subordonné aux caractéristiques de l’aire, au degré de dérangement anthropique et au niveau de fréquentation des environnements par les animaux sauvages, in primis le loup. Chaque station de détection a été mise en cartographie et chaque camera a été identifiée par un code d’identification. Les trappes photographiques ont été soumises, en relation aux conditions climatiques, à des contrôles fréquents tandis que de téléchargement d’images mémorisées, en permettant d’obtenir des données qui plus en avant ont été intégrées avec cettes obtenues par des autres techniques de recension. Pendant la période d’enquête, ont été conservées toutes les registrations photo/vidéo obtenues et les informations collectées par chaque appareil (espèces animales identifiée, date et heure de détection, etc.) ont été introduites dans un archive logiciel spécifique. Environ en 13 mois d’activité (fin novembre 2011 – décembre 2012) ont été activées 100 stations de détection (84 specifiques pour le loup et 16 plus générales) qui dans l’ensemble ont opéré pour 8.614 journées (moyenne 662 journées par mois) en obtenant en complexe plus de 48.000 registrations, dont 9.348 reconductibles à des animaux.

50 ATTIVITÀ DI RICERCA, CENSIMENTO E MONITORAGGIO
43 Uccelli e 21 Mammiferi (16 specie selvatiche [a], 4 domestiche, cane, gatto, bovino e pecora, ed 1 introdotta, l’Ammotrago Ammotragus lervia).
I risultati ottenuti sono particolarmente interessanti per ciò che riguarda i Mammiferi; infatti, escludendo i Chirotteri (per i quali ci sono state comunque 7 registrazioni, purtroppo non determinabili almeno a livello di Genere), le specie contattate rappresentano il 76% di quelle note per il Parco del Beigua; infatti solo per Toporagno d’acqua (Neomys fodiens),Talpa europea (Talpa europaea),Martora (Martes martes) e Mufl one (Ovis orientalis) non ci sono stati contatti, mentre per le altre è possibile quantifi carne le presenze sia complessivamente che nell’arco annuale, calcolandone, ad esempio la ‘frequenza di cattura’ (che equivale al numero di contatti / il numero di giornate di attività delle fototrappole). Vediamo così come, con la tecnica del trappolaggio fotografi co, le specie più abbondanti siano risultate Volpe, Capriolo, Cinghiale e Faina [a]. Il Lupo è la quinta specie in ordine di abbondanza; a parte i Chirotteri è l’unico Mammifero del Beigua contemplato dalla Direttiva Habitat (specie prioritaria, inclusa negli allegati II e IV) e verrà trattato estesamente.
Riferimenti bibliografi ci: Barnes (2001), Bider (1968), Eberhardt & Van Etten (1956), Fashing & Cords (2000), Fragoso (1991), Gannon & Fo-ster (1996), Henschel & Ray (2003), Karanth & Nichols (1998), McNeila-ge et al. (2001), O’Brien et al. (2003), Silveira et al. (2003), Spagnesi & De Marinis (2002), Wegge et al. (2004).
Donnola
Ratto nero
Mustela nivalis
Rattus rattus
Riccio
Daino
Puzzola
Erinaceus europaeus
Dama dama
Mustela putorius
Topo quercino
Ghiro
Eliomys quercinus
Glis glis
Topo selvatico
Leprecomune
Scoiattolo
Apodemus sp.
Lepus europaeus
Sciurus vulgaris
Faina
Lupo
Tasso
Martes foina
Canis lupus
Meles meles
V l
Capriolo
Cinghiale
V l l
Capreolus capreolus
Sus scrofa
Volpe
0,0% 5,0% 10,0% 15,0% 20,0%
Vulpes vulpes
Frequenza di cattura
[a]
L’analyse de ces données a permis l’identification de 68 espèces: 3 amphibiens, 1 reptile, 43 oiseaux et 21 mammifères (16 espèces sauvages [a], 4 domestiques, chien, chat, bœuf et mouton, et 1 introduite, mouflon à manchettes Ammotragus lervia). Les résultats obtenus sont particulièrement intéressants en ce qui concerne les mammifères; en effet, en excluant les chiroptères (dont on a toutefois effectué 7 registrations, malheureusement non attribuables à niveau de Genre), le espèces contactés représentent le 76% de ces qui sont connues dans le Parco del Beigua; en effet seulement le Crossope aquatique d’eau (Neomys fodiens), Taupe européenne (Talpa europaea), Martre (Martes martes) et Muflon (Ovis orientalis) n’a pas eu des contacts, par contre pour les autres espèces il est possible en quantifier les présences soit dans l’ensemble de pendant l’entière année, avec le calcul, par exemple, de la « fréquence de capture » (qui est le nombre de contacts sur le nombre de journées d’activité des trappes photographiques). On peut voir comme, avec la technique des trappes photographiques, les espèces principales sont : le renard, chevreuil, sanglier et la fouine. Le loup est la cinquième espèce en ordre d’abondance; à l’exclusion des Chiroptères, il est l’unique mammifère du Beigua contemplé par la Directive Habitat (espèces prioritaire, inclue dans les annexes II et IV) et sera traité extensivement.
Références bibliographiques: Barnes (2001), Bider (1968), Eberhardt & Van Etten (1956), Fashing & Cords (2000), Fragoso (1991), Gannon & Foster (1996), Henschel & Ray (2003), Karanth & Nichols (1998), McNeilage et al. (2001), OU’Brien et al. (2003), Silveira et al. (2003), Spagnesi & De Marinis (2002), Wegge et al. (2004).
Vulpes vulpes RenardCapreolus capreolus ChevreuilSus scrofa SanglierMartes foina FouineCanis lupus Loup Meles meles BlaireauApodemus sp. Rat sauvage Lepus europaeus Lièvre d'Europe Sciurus vulgaris Ecureuil roux Eliomys quercinus Lérot commun Glis glis Loir gris Erinaceus europaeus Hérisson commun Dama dama Daim européen Mustela putorius PutoisMustela nivalis BeletteRattus rattus Rat noir

51
IL LUPO
Il Lupo (Canis lupus; specie prioritaria, inclusa negli allegati II e IV della Direttiva Habitat) è uno dei Mammiferi selvatici con la distribuzione geografi ca più estesa, origi-nariamente presente in Nord America, in Eurasia e gran parte della Penisola Arabica e della Penisola Indiana. In particolare negli Stati Uniti ed in Europa la specie ha mostrato un marcato declino, che ha causato l’estinzione di diverse popolazioni. In Italia, alla di-minuzione verifi catasi sino agli anni ’70, è seguita un’espansione dell’areale che ha por-tato la specie ad occupare tutta la catena appenninica ed alla ricolonizzazione delle Alpi.
È specie dalla forma slanciata, e dalle dimensioni medio grandi (lunghezza testa – cor-po 110-148 cm, altezza alla spalla 50-70 cm, coda di 30-35 cm), testa massiccia con muso allungato, orecchie relativamente grandi ed erette. Ha colorazione del mantello che va dal grigio pallido al marrone grigiastro. Il peso è molto variabile (fi no a 70 Kg), ma in Italia in media è compreso tra i 25 e i 35 Kg con maschi più pesanti e massicci. Frequenta habi-tat vari, dalla tundra ai deserti, alle foreste di pianura e di montagna; in Italia predilige le aree con densa copertura forestale collinari e montane. Ha alimentazione piuttosto varia che comprende prevalentemente Ungulati selvatici (soprattutto Cinghiale e Capriolo, ma anche Cervo e Mufl one) e secondariamente domestici (in particolare ovini) con presenza di piccoli mammiferi, lepri, frutta, con proporzioni molto variabili secondo la disponibi-lità e la stagione.
Di abitudini prevalentemente notturne, vive in branchi che si spostano, cacciano, si nutrono, riposano insieme, in libera associazione, ma uniti l’un l’altro dai forti legami sociali che si instaurano tra i componenti del gruppo (Mech 1970). Essenzialmente tale unità corrisponde ad un nucleo familiare, che si origina dall’incontro di un maschio e una femmina che si riproducono in un territorio idoneo (Rothman & Mech 1979; Fritts & Mech 1981). Nei carnivori sociali, in passato, la tendenza all’aggregazione è stata interpretata come un adattamento selettivo fi nalizzato alla caccia dei grandi mammiferi erbivori (Bekoff & Well 1980; Zimen 1976). Attualmente l’ipotesi più accreditata per spiegare la tendenza dei lupi a vivere nel branco è quella della kin selection (Schmidt & Mech 1997), secondo la quale gli adulti del nucleo familiare investono energie sulla prole attraverso la condivisione del cibo in esubero e la trasmissione culturale, consentendo così di massimizzare le probabilità di sopravvivenza dei cuccioli.
LE LOUP
Le loup (Canis lupus; espèces prioritaire, incluse dans les annexes II et IV de la Directive Habitat) est l’un des mammifères sauvages avec la distribution geographique la plus étendue, originairement present dans l’Amérique du nord, en Eurasie et dans la plus grande partie de la péninsule arabique et indienne. Notamment dans les Etats-Unis et en Europe, l’espèce a montré un décline marqué, qui a causé l’extintion de plusieurs populations. En Italie, à la diminution arrivée jusqu’aux années 70, est suivie une expansion de l’aire qui a emmené l’espèce à occuper toute la chaîne montagneuse des Apennines et à la nouvelle colonisation des Alpes. Elle est une espèce à forme éloignée, ayant dimensions moyen grandes (longueur de la tête – corps 110-148 cm, hauteur à l’épaule 50-70 cm, queue 30-35 cm), une tête massive avec le museau éloigné, oreilles relativement grandes et érigées. Elle montre une couleur du manteau qui commence du gris pâle au marron-gris. Le poids est plutôt variable (jusqu’à 70 kg) mais en Italie en moyenne il est compris entre 25 et 35 kg avec les mâles plus grands et massifs que les femmes. Elle fréquente les habitats les plus différents, de la toundra aux déserts, aux forêts des plaines et de montagne ; en Italie préfère les airs à dense couverture forestière, les collines et les montagnes. Possède une alimentation plutôt variée, qui comprend typiquement des ongulés sauvages (surtout sanglier et chevreuil, mais aussi cerf et muflon) et secondairement des animaux domestiques (notamment ovins) avec la présence de petits mammifères, lèvres, fruits, avec des proportions assez variables selon la disponibilité et la saison. D’habitude substantiellement nocturne, le loup vive en troupeaux qui se déplacent, chassent, nourrissent et reposent ensemble, en association libre, mais unis l’un l’autre par des forts liens sociaux qui s’instaurent entre les individus du groupe (Mech 1970). Essentiellement, cette unité correspond à un noyau familial, qui trouve origine par un mâle et une femme qui reproduisent dans un endroit adapté (Rothman & Mech 1979; Fritts & Mech 1981). Entre les carnivores sociaux, en passé, la tendance à l’agglomération a été interprétée comme une adaptation sélective finalisée à la chasse des grand mammifères herbivores (Bekoff & Well 1980; Zimen 1976). Aujourd’hui l’hypothèse plus accréditée pour expliquer la tendance des loups à vivre en troupeaux est la thèse de la kin selection (Schmidt & Mech 1997), dont les adultes du noyau familial dépensent des énergies sur les petits avec le partage de la nourriture en surplus et la transmission culturale, en permettant de ce façon la maximisation des probabilités de survivance des petits.

52 ATTIVITÀ DI RICERCA, CENSIMENTO E MONITORAGGIO
Sebbene branchi composti da un elevato numero di individui siano stati osservati in natura, la dimensione media su scala mondiale è di 7 lupi (Mech 1970), da 2 a 7 in Italia. La dimensione del branco varia notevolmente durante l’anno ed è regolata dalla mortali-tà, dalla produttività e dall’età media alla quale gli individui entrano in dispersione. Infat-ti, nei giovani di 2-3 anni si manifesta spesso la tendenza ad abbandonare il branco (Fritts & Mech 1981; Peterson et al. 1984; Hefner & Geffen 1999) e ad andare in dispersione. Durante l’anno si verifi cano due picchi di dispersione: uno tra febbraio-aprile e uno tra ottobre-novembre. Un lupo in fase di dispersione può percorrere da 8 fi no a 354 Km. Solitamente i giovani inesperti si allontanano molto dal territorio natale, conseguendo numerosi fallimenti; gli adulti, invece, tendono a stabilirsi in aree limitrofe al luogo di origine e hanno un successo maggiore (Gese & Mech 1991).
Generalmente all’interno di un branco si accoppiano il maschio e la femmina domi-nanti; i cuccioli (solitamente da 1 a 6) nascono in primavera, vengono partoriti in una tana e svezzati a 7-8 settimane; i giovani rimangono poi con i genitori per almeno un anno e la maturità sessuale viene raggiunta intorno al secondo anno di età (Jordan et al. 1967;Fuller 1989; Boertje & Stephenson 1992; Mech 1970, 1974).
In Italia il Lupo è una specie territoriale e ogni branco tende ad occupare un territorio esclusivo che oscilla intorno a 150-300 Km2 (Boitani & Ciucci 1998; Ciucci et al. 1997;Liccioli 2004), e dal quale eventuali conspefi ci estranei vengono attivamente estromessi (Mech, 1974). Il territorio di un branco comprende aree di caccia, di riproduzione, di spo-stamento e di riposo (Mech 1970) e viene attivamente difeso utilizzando principalmente segnali olfattivo-visivi, come le urine, le raspate e le secrezioni delle ghiandole perianali (Peters & Mech 1975; Rothman & Mech 1979) o acustici, come l’ululato (Harrington & Mech 1979; 1983).
Come ormai noto, il Lupo è riapparso in Liguria per ricolonizzazione spontanea, risa-lendo dall’appennino centro meridionale in seguito a un’inversione di tendenza dovuta a più fattori, tra i quali la protezione legale assicurata alla specie, i radicali mutamen-ti dei territori montani soggetti a spopolamento rurale e l’aumento delle specie preda. Il ritorno di questo superpredatore in Liguria avviene quindi come logica conseguenza dell’incremento delle popolazioni relitte poste molto più a sud (soprattutto in Abruzzo e in Calabria) e ciò si verifi cò dopo vari decenni di estinzione seguita all’uccisione talvolta ben documentata degli ultimi Lupi liguri.
Uno di questi esemplari risale al 1871 ed è l’ultimo Lupo di Liguria che fu conservato in un museo (Mus. Civ. St. Nat. “G. Doria” di Genova) prima del ritorno della specie, e lo uccise il Marchese Marcello De Mari presso Stella Santa Giustina in una località oggi compresa nel Parco del Beigua (Doria in Campora et al. 2009). L’ultima uccisione
Bien que les troupeaux ayant beaucoup d’individus soient été observés en nature, la dimension moyenne sur l’échelle mondiale es 7 loups (Mech 1970), de 2 à 7 en Italie. La dimension du troupeau varie sensiblement pendant l’année et est contrôlée par la mortalité, la reproduction et l’âge moyen dont les individus sont dispersés du troupeau. En effet, pour les jeunes de 2-3 années souvent arrive la tendance à l’abandonnement du troupeau (Fritts & Mech 1981; Peterson et al. 1984; Hefner & Geffen 1999) et à se disperser. A fil de l’année on peut compter deux pics de dispersion : un premier entre février et avril et un deuxième entre octobre et novembre. Un loup éloigné peut parcourir de 8 jusqu’à 354 kilomètres. Traditionnellement les jeunes peu experts s’éloignent beaucoup du territoire natal, et achèvent plusieurs échecs ; les adultes, par contre, montrent la tendance à s’établir dans des aires limitrophes au lieu d’origine et ont un plus grand succès (Gese & Mech 1991). Généralement à l’intérieur d’un troupeau le mâle et la femme s’accouplent entre individus dominants ; les petits (typiquement de 1 à 6) arrivent en printemps, sont accouchés dans une tanière et sevrés à 7-8 semaines; les jeunes restent puis avec les parents pour au moins un an, et la maturité sexuelle est achevé autour de la deuxième année (Jordan et al. 1967; Fuller 1989; Boertje & Stephenson 1992; Mech 1970, 1974). En Italie le loup est une entre les espèces territoriales et chaque troupeau tend à occuper un territoire exclusif qui est d’environ 150-300 Km2 (Boitani & Ciucci 1998; Ciucci et al. 1997; Liccioli 2004), et dont des éventuels individus de la même espèce sont activement exclus (Mech, 1974). Le territoire d’un troupeau comprend des aires de chasse, de reproduction, de déplacement et de repos (Mech 1970) et est activement défendu typiquement avec des signaux olfactifs et visuels, comme les urines, les râpes et les sécretions des glandillons péri-anales (Peters & Mech 1975; Rothman & Mech 1979) ou acoustiques comme l’ululé (Harrington & Mech 1979; 1983). Comme aujourd’hui connu, le loup est nouvellement apparu en Ligurie pour une nouvelle colonisation spontanée, en remontant de l’apennine centrale méridionale à la suite d’une inversion de tendance due à facteurs diverses, entre lesquels on trouve la protection légale assurée à l’espèce, les changements radicaux des territoires montagneux qui sont sujets à dépeuplement rural et l’incrément des espèces proie. Le retour de ce super prédateur en Ligurie arrive donc comme conséquence logique de l’incrément des populations épaves placées beaucoup plus à sud (surtout en Abruzzo et Calabria) et ça se vérifiait après plusieurs décennies d’extinction suivie par l’abattage certaines fois bien documenté des derniers exemplaires de la Ligure. Un entre eux remonte à 1871 et est le dernier Loup de Ligurie qui fut conservé dans une musée (Mus. Civ. St. Nat. “G. Doria” de Genova) avant le retour de l’espèce, et fut abattu par le Marquis Marcello de Mari auprès Stella Santa Giustina dans une localité aujourd’hui comprise dans le Parco del Beigua (Doria in Campora et al. 2009). Le dernier abattage documenté concerne la Val d’Aveto, et notamment en 1946 auprès S. Stefano d’Aveto (Cagnolaro et al. 1974), dont suivent environ quarante années de totale absence de repérages.

53
documentata è invece relativa alla Val d’Aveto, e precisamente nel 1946 presso S. Stefa-no d’Aveto (Cagnolaro et al. 1974), cui seguono circa quarant’anni di totale assenza di segnalazioni.
Già dai primi anni ’80 si iniziò a parlare di un possibile ritorno del Lupo nell’Appenni-no ligure; a quei tempi la presenza era da considerarsi sporadica, e le prime segnalazioni sull’Appennino genovese (Valli Trebbia e Aveto) erano legate prevalentemente a preda-zioni di bestiame (Meriggi & Schenone 1998).
Dopo un’intensifi cazione delle segnalazioni, nell’inverno 1985-86 è dalla vicina Val Borbera, in provincia di Alessandria, che arriva il primo reperto conclamato: un Lupo uc-ciso (Spanò 1997). Da questo momento non esistono più dubbi sul ritorno del Lupo e da lì a poco inizieranno ricerche condotte soprattutto dall’Università di Pavia (Dipartimento di Biologia animale) nelle valli appenniniche delle provincie di Piacenza, Genova, Ales-sandria e Pavia per seguire le fasi di questo evento, cercando di defi nire il territorio dei primi nuclei stabili ed accertarne la riproduzione. Gli andamenti paiono fl uttuanti negli anni, registrando inoltre una marcata ostilità delle popolazioni locali che è talora sfociata in atti di bracconaggio. Tale pratica si è rivelata a volte deleteria, se pur non indirizzata esplicitamente al Lupo, come nel caso dei lacci il cui uso, ancora oggi non del tutto scom-parso, era ancora in gran voga negli anni ’90. E proprio al 1990 risale un evento di rilievo: una coppia di Lupi resta vittima dei lacci di un bracconiere in Val d’Aveto. Questi primi esemplari uccisi e recuperati su suolo ligure dopo gli anni dell’estinzione sono anch’essi conservati al Museo di Genova.
Intanto le valli del Beigua, storicamente tra le ultime ad ospitare il Lupo, in questo periodo sono certamente interessate dal passaggio di esemplari in dispersione, ma nella prima fase di ricolonizzazione sembrano non ospitare nuclei stabili. Il fenomeno è comune a molte al-tre aree appenniniche oggi abitate stabilmente, come le valli del Gottero, tra La Spezia, Parma e Massa Carrara, dove, se pur segnalato sin dal 1988, i primi accertamen-ti di rilievo risalgono al 1992, o il gruppo montuoso del Figne (Parco naturale delle Capanne di Marcarolo e alta val Polcevera), per il quale la formazione di un branco stabile è avvenuta (stando ai dati disponibili) assai dopo il ritorno del Lupo sulle Alpi. Tale clamoroso fatto av-venne inizialmente “passando in sordina” (1987 nelle Alpi Marittime Francesi, e prima osservazione certa per le Alpi italiane in Val Pesio nel 1990; Audino 1997) e poi con grande risonanza dal 1992 nel Parco del Mer-cantour, dove due anni più tardi se ne accertò la ripro-duzione (Poulle et al. 1995).
Più che a una carenza di ricerche specifi che (e conse-guente mancanza di dati) questa espansione a “macchia di leopardo” è forse imputabile alla disponibilità di pre-de nei differenti settori, fattore che ha subito notevoli mutamenti nel corso degli anni (basti pensare al netto incremento numerico osservato per il Cinghiale, ottima
Déjà des premiers années ’80 on commençait parler d’un possible retour du Loup sur l’Apennine Ligure ; dans ce temps-là sa présence était sporadique, et les premiers repérages sur l’Apennine de Genova (Valli Trebbia e Aveto) étaient liés principalement à des prédations de bétail (Meriggi & Schenone 1998). Après une intensification des repérages, dans l’hiver 1985-86 est de la proche Val Borbera, en province d’Alessandria, qui arrive le premier rapport confirmé : un Loup abattu (Spanò 1997). A partir de ce moment n’existent pas plus des doutes sur le retour du Loup et assez rapidement commenceront les recherches par l’Université de Pavia (Département de Biologie Animale) dans les vallées de l’apennine de la province de Piacenza, Genova, Alessandria et Pavia pour suivre les phases du évent, en cherchant de définir le territoire des premiers noyaux stables et en vérifier la reproduction. Les évolutions se ressemblent fluctuantes au cours des années, en registrant en outre une marquée hostilité des populations locales que parfois est arrivée à des actes de braconnage. Cette pratique s’est révélée parfois délétère, bien que non adressée explicitement au Loup, comme dans le cas des laces, dont l’utilisation n’est pas même aujourd’hui encore par tout disparue, était en vogue dans les années ’90. Il est du 1990 l’évent plus important : une couple de Loups est victime des laces d’un braconnier en Val d’Aveto. Ces premiers exemplaires abattus et récupérés sur le sol Ligure après des années d’extinction sont eux-mêmes conservés dans la musée de Genova. Toutefois, dans le vallées du Beigua, historiquement parmi les dernières à héberger le Loup, dans ce période sont certainement intéressées par le passage d’exemplaires en dispersion, mais dans la première phase de recolonisation, il apparaît que n’hébergent pas des noyaux stables. La phénomène est commune pour plusieurs aires de l’Apennine aujourd’hui habitées stablement, comme les vallées du Gottero, entre La Spezia, Parma et Massa Carrara, où, bien que identifié à faire compte du 1988, les premières identifications d’importance remontent au 1992, ou le groupe montagneux du Figne (Parco naturale delle Capanne di Marcarolo et haute vallée Polcevera), dont la formation d’un troupeau stable est arrivée (selon les données disponibles) bien après le retour du Loup dans les Alpes. Ce éclatant fait arrive initialement en passant en sourdine (1987 dans les Alpes Maritimes Françaises, et première observation confirmée pour les Alpes italiennes en Val Pesio en 1990; Audino 1997) et puis avec une grande résonance à partir du 1992 dans le Parco del Mercantour, où deux années plus tard s’en vérifiait la reproduction (Poulle et al. 1995).

54 ATTIVITÀ DI RICERCA, CENSIMENTO E MONITORAGGIO
preda per il Lupo). Nei primi anni ’90, ad esempio, il Capriolo nel Parco del Beigua, se pur già diffuso, non si presentava certamente con le densità attuali; mentre il Daino, specie alquanto apprezzata dal Lupo, era già comune intorno all’Antola e in Val Borbera. Stesse considerazioni valgono per le Alpi, dove i primi branchi di Lupo si stabilirono presso quelle aree connesse a Parchi come Mercantour, Salbertrand e Alpi Marittime, nelle quali si riscontravano elevate densità di Ungulati. Negli anni ‘90 si assiste quindi ad un’espansione del lupo sul territorio alpino italiano, con l’insediamento di branchi dalle Valli Tanaro e Pesio (ed i cui territori interessano anche la Liguria) alle Valli di Lanzo (Torino), ed individui in dispersione che si spingevano verso la Svizzera (Fasoli & Cen-tofanti 1997).
Tornando all’evoluzione della ricolonizzazione del Beigua, i primi indizi risalgono al 1993, con predazioni di agnelli presso Pratorondanino (Masone). Nella seconda metà degli anni ’90 si rinvenivano alcuni segni di presenza, tra i quali una pista su neve di almeno due esemplari e una fatta contenente pelo di pecora. Fino al 2008, nonostante un’assidua frequentazione del territorio, i ritrovamenti di segni di presenza, pur riguar-dando gran parte dell’area, furono alquanto occasionali ed incostanti nel tempo, mentre la loro frequenza si intensifi cava nel vicino Parco delle Capanne di Marcarolo, dove la specie è seguita nell’ambito del “Progetto Lupo Regione Piemonte”. È quindi dal 2009 che, grazie anche ai risultati ottenuti dal progetto regionale “Il Lupo in Liguria”, l’ele-vato numero di dati raccolti permette di individuare l’insediamento di un nucleo stabile nel Parco del Beigua [a].
Le principali tecniche d’indagi-ne impiegate per il monitoraggio del Lupo sono: la raccolta dei segni di presenza lungo una rete di percorsi campione (transetti; metodo princi-pale adottato nell’ambito del proget-to “Il Lupo in Liguria”), l’impiego delle trappole fotografiche, la tecnica dell’ululato indotto (wolf-howling),la tracciatura delle piste d’impronte su substrato nevoso (snow-tracking);inoltre, la raccolta degli escrementi, grazie al recente sviluppo dei metodi della genetica molecolare, permette di ricostruire il profi lo genetico (uni-co per ogni individuo), ottenendo così molte informazioni sulla popolazione in esame. Tutte queste tecniche sono state applicate, anche se in misura va-riabile, nel corso delle ricerche con-dotte nel Parco del Beigua tra l’ottobre 2011 ed il gennaio 2013, ottenendo nel complesso 885 dati riconducibili al Lupo [b].
350
400
450
Numero di dati per anno e tipologia di osservazione(N = 945)
100
150
200
250
300
N°
dati
0
50
2004 2007 2008 2009 2010 2011 2012 2013 (gennaio)Anno
Osservazione Richiami Predazione Tracce Fatta Fototrappola
[a]
[b]
Plus que à une manque de recherches spécifiques (et par conséquent une manque de données), cette expansion à façon de fourrure de léopard peut être attribuée à la disponibilité de proies des différents secteurs, un facteur qui a subi des changements significatifs au cours des années (ça suffit penser au fort incrément numérique observé pour le sanglier, une parfaite proie pour le loup). Au debut des années ’90, par exemple, le chevreuil dans le Parco del Beigua, bien que déjà diffusé, ne se présentait certainement pas avec les densités actuelles, par contre le daim, espèce bien appréciée par le loup, était déjà commune autour de l’Antola et la Val Borbera. Les mêmes considérations sont valides pour les Alpes, dont les premiers troupeaux de loup ont établis auprès quelles aires liées à Parcs comme le parc de la Mercantour, de Salbertrand ou des Alpi Marittime, dont on retrouvait d’hautes densités d’ongulés. Pendant les années ’90 on assiste donc à une expansion du loup dans le territoire alpin italien, avec l’établissement de troupeaux entre les Valli Tanaro et Pesio (dont les territoires intéressent aussi la Ligurie), le Valli di Lanzo (Torino) et des individus en dispersion qui se poussaient vers la Suisse (Fasoli & Centofanti 1997). En retournant à l’évolution et à la recolonisation du Beigua, les premiers indices arrivent au 1993, avec des chasses d’agneaux auprès Pratorondanino (Masone). Dans la deuxième moitié des années ’90, on retrouvait certains signes de leur présence, comme une piste sur neige d’au moins deux exemplaires et une qui contenait seulement poils des mouton. Jusqu’au 2008, malgré une assidue fréquentation du territoire, les découvertes de signes de présence, bien que concernant une grande partie de l’aire, étaient plutôt occasionnels et non constants au cours du temps, tandis que leur fréquence s’intensifiait auprès du proche Parco delle Capanne di Marcarolo, où l’espèce est suivie dans le milieu du projet “Progetto Lupo Regione Piemonte”. Il est à partir du 2009 que, grâce aux résultats obtenus par le projet régional “Il lupo in Liguria”, le grand nombre de données rassemblées permet l’individuation de l’installation d’un noyau stable dans le Parco del Beigua [a].
Les principales techniques d’enquête employées pour le monitorage du Loup sont : le rassemblement de signes de présence au long d’un réseau de parcours échantillons (transepts ; méthode principale adoptée dans le milieu du projet “Il lupo in Liguria”), l’utilisation des trappes photographiques, la technique du hurlement induit (wolf-howling), le traçage de pistes d’empreintes sur le substrat neigeux (snow-tracking); en plus, le rassemblement d’excréments, grâce au récent développement des méthodes de la génétique moléculaire, permet de reconstruire le profil génétique (unique pour chaque individu) en obtenant de ce façon plusieurs informations sur la population en examen. Toutes ces techniques ont été appliquées, bien que en mesure variable, au cours des recherches effectuées dans le Parco del Beigua entre octobre 2011 et janvier 2013, en obtenant dans l’ensemble 885 données reconductibles au Loup [b].

55
Tra i pochi avvistamenti diretti merita menzione quello del 26 febbraio 2012 all’alba, cui è seguito un lungo pedinamento della pista lasciata su neve dal branco al completo, che si è spostato da un’area piuttosto centrale del territorio verso una zona periferica soffermandosi solo su qualche valico; la pista è stata poi persa verso le ore 13 su un pendio ripido privo di neve. A proposito dei richiami, oltre alle numerose testimonianze di persone che abitano nelle zone maggiormente frequentate dai Lupi, durante le nostre attività di campo si sono potuti ascoltare sette episodi d i ululati, con almeno un evento per stagione.
Gli escrementi (‘fatte’) trovati durante le attività sono stati 317, dei quali il 10-15%, utili per le analisi genetiche, sono stati inviati al Laboratorio di Genetica dell’ Istituto Su-periore per la Protezione e Ricerca Ambientale; al momento i risultati preliminari hanno permesso di caratterizzare 10 individui e defi nire le relazioni di parentela per 5 di essi. Per ciò che riguarda la dieta (defi nita in base al contenuto degli escrementi) si conferma una preponderanza del Cinghiale, ma con discrete differenze stagionali; il Cinghiale è stato infatti campionato in modo esclusivo fra il tardo autunno e l’inverno, mentre il Capriolo ha iniziato a comparire da marzo ed è diventato di una certa importanza nei mesi estivi.
Il trappolaggio fotografico ha permesso di ottenere 290 contatti di Lupo [a], con una frequenza di cattura complessiva pari al 4,0% (numero totale di catture diviso per le giornate di attività); nel corso dell’anno tale frequenza è stata però molto variabile, con i valori massimi durante il periodo invernale e frequenze nettamente inferiori nelle altre stagioni [b]. Mediamente si sono osservati 1,6 individui per contatto, con un massimo di 7 in due occasioni.
Si è inoltre stimata la densità secon-do il metodo di Rowcliffe et al. (2008),che si basa su due caratteristiche tec-niche delle fototrappole (raggio d’a-zione del sensore e angolo della zona di rilevamento, ricavati dalla media dei parametri di ogni modello utilizzato, in accordo con lo studio di Anile, 2009) e su un parametro ecologico (la veloci-tà media giornaliera della specie, pari a 27,3 km, mediando quanto riportato da Ciucci et al. 1997 e Jedrzejewski et
60
80
100
120
°da
tiNumero di dati per mese e tipologia di osservazione
ottobre 2011 - gennaio 2013 (N = 885)
0
20
40
O N D G F M A M G L A S O N D G
2011 2012 2013
N°
Anno / Mese
Fototrappola Osservazione Richiami Predazione Tracce Fatta
[a]
8,1% 8,1%
6,6%7,1%
7%
8%
9%
i
Frequenza di cattura
1,7%1 2%
3,1% 3,4%
1,3%
2,5% 2,6% 2,8%
5,6%
,
2%
3%
4%
5%
6%
7%
cont
atti
/ n°
gior
ni
1,2% ,
0%
1%
D G F M A M G L A S O N D
2011 2012
c
Anno / Mese
[b]
Parmi les rares repérages directs est susceptible d’attention celui du 26 fevrier 2012, à l’aube, dont a suivi une longue suivie de la piste laissée sur la neige par le troupeau au complet, qui s’est déplacé d’une aire plutôt centrale du territoire vers une zone périphérique se concentrant uniquement sur quelques cols ; la piste était perdue vers 13 heures vers une pente raide sans neige. A propos des appels, en outre aux plusieurs témoignages des personnes qui habitent entre les aires les plus fréquentées par les Loups, pendant nôs activités de champ on a pu écouter sept épisodes d’hurlement, avec au moins un évent par saison. Les excréments (« faites ») trouvés pendant les activités sont 317, dont 10-15% utiles pour les analyses génétiques, ont été envoyées au Laboratoire de génétique du Istituto Superiore per la Protezione e Ricerca Ambientale; en ce moment les résultats préliminaires ont permis la caractérisation de 10 individus et la définition des degrés de parentèle pour 5 d’eux. En ce qui concerne la diète (définie sur la base du contenu des excréments) on confirme une préponderance du sanglier, mais avec des discrètes différences entre les saisons; le Sanglier a été échantillonné exclusivement entre le tard automne et l’hiver, tandis que le Chevreuil a commencé apparaître au début mars et est devenu d’une certaine importance pendant les mois de l’été. Les trappes photographiques ont permis d’obtenir 290 contacts de Loup [a], avec une fréquence de capture totale du 4,0% (nombre total des captures divisé pour les journées d’activité) ; au cours de l’année cette fréquence a été toutefois plutôt variable, avec les valeurs maximales pendant la période hivernale et fréquences nettement plus basses pendant les autres saisons [b]. En moyenne, on a observé 1,6 individus par contact, avec un maximum de 7 individus en deux occasions. On a aussi estimé la densité selon la méthode Rowcliffe et al. (2008), qui se base sur deux caractéristiques techniques des trappes photographiques (le rayon d’action du capteur et l’angle de la zone de détection, deduits par la moyenne de paramètres de chaque modèle utilisé, en accord avec l’étude d’Anile, 2009) et sur un paramètre écologique (la vitesse moyenne journalière de l’espèce, équivalente à 27,3 km, en effectuant une moyenne de ce qui est reporté par Ciucci et al. 1997 et Jedrzejewski et al. 2001). Avec cette méthodique, on a obtenu une densité de 6,7 ind./100 Km2; une valeur légèrement plus haute de cela individuée dans des autres realités (par exemple dans le Parco Nazionale delle Foreste Casentinesi Monte Falterona e Campigna, estimé en 4,7 ind./100 Km2; Appollonio et al. 2004; Lucchesi et al. 2012).

56 ATTIVITÀ DI RICERCA, CENSIMENTO E MONITORAGGIO
al. 2001). Con questa metodica si è ottenuta una densità pari a 6,7 ind./100 Km2; valore leggermente superiore a quanto riscontrato in altre realtà (ad esempio, nel Parco Nazio-nale delle Foreste Casentinesi Monte Falterona e Campigna, stimata in 4,7 ind./100 Km2;Appollonio et al. 2004; Lucchesi et al. 2012).
Interessante è inoltre la distribuzione oraria degli eventi, che risultano prettamente notturni, con eccezioni solo per orari prossimi all’alba e al tramonto o in altri orari diurni solo per poche giornate caratterizzate da clima piovoso o nubi orografi che, ed un unico evento in pieno sole. La distribuzione giornaliera è stata inoltre analizzata nell’ambito di una Tesi di Laurea a cura di G. Calcagno.
Altre informazioni emerse dal fototrappolaggio riguardano i tragitti che i singoli esem-plari riescono a compiere nell’arco di poche ore (ad esempio, tra due stazioni di rileva-mento poste a circa 5,5 Km l’una dall’altra si sono riscontrati tempi di percorrenza mini-ma di 68 e 75 minuti). Tra questi eventi spicca quello della notte tra il 3 ed il 4 gennaio 2012, durante la quale è stato possibile monitorare il percorso della coppia dominante che, accompagnata dai soggetti più giovani, ha percorso circa 15 Km tra le ore 19 e la mezzanotte.
Risultano poi notevoli le differenze della frequenza di campionamento in relazione alla tipologia dei siti monitorati. In sintesi, su strade e sentieri principali i passaggi di lupo sono piuttosto regolari, e in vari casi hanno frequenza settimanale, mentre lungo le piste di selvatici, spesso utilizzate da elevati numeri di Ungulati, i contatti di Lupo sono stati scarsissimi. In uno di questi siti, solo dopo ben 13 mesi e 18 giorni sono stati ripresi i primi Lupi, e si tratta inoltre dell’unico caso di palese attività di caccia ai danni di un Capriolo femmina e del proprio piccolo, i quali, in precedenza, erano stati costantemen-te rilevati dalla fototrappola (che era posta in uno dei loro punti di riposo); il giorno in questione, dopo mezz’ora dal passaggio in piena corsa dei Lupi, la femmina ricomparve trafelata emettendo suoni di richiamo, raggiunta poi nelle ore successive dal giovane, segno che la caccia dei Lupi non si era conclusa a loro favore.
Même la distribution horaire des évents est intéressante, dont les évents sont pour la plus partie nocturnes, avec des exceptions seulement pour les horaires proches à l’aube et au coucher du soleil ou pendant des autres horaires journaliers seulement pour quelque journée caractérisée par un climat de pluie ou nuageux orographiques, et un single évent en plein soleil. La distribution journalière a été en outre analysée dans le contexte d’une thèse de maîtrise par G. Calcagno. Des autres informations émergées des trappes photographiques concernent les parcours que les exemplaires sont capables d'effectuer au cours de quelques heures (par exemple, entre deux stations de détection placées à environ 5,5 Km l’une de l’autre on a identifié des temps de parcours minimales entre 68 et 75 minutes). Entre eux émerge celui de la nuit entre le 3 et 4 janvier 2012, pendant lequel il était possible de contrôler le parcours de la couple dominante qui, accompagnée par des sujets plus jeunes, a parcouru presque 15 km entre 19 heures et minuit. Les différences de la fréquence d’échantillonnage en relation à la typologie des sites contrôlés sont significatives. En bref, sur les routes et sentiers principaux les passages des loups sont plutôt réguliers, et en plusieurs cas ont une fréquence hebdomadaire, tandis que au long des pistes de sauvages, souvent exploitées par des hautes nombres d’ongulés, les contacts du loup ont été très rares. En un de ces sites, seulement après 13 mois et 18 jours on a identifié les premiers Loups, et il s’agissait en plus de l’unique cas d’évidente activité de chasse au détriment d’une Chevreuil femme et son petit, étaient en précédence déjà identifiés par la trapppe photographique (qui était placée dans l’un de leur points de repos) ; dans ce jour-là, après environ une demi-heure du passage en pleine course des Loups, la femme est apparue essoufflée en émettent des sons de rappel, rejointe dans les heures suivantes par le jeune, une démonstration que la chasse des loups n’avait conclue en faveur d’eux.

57
L’analisi delle immagini raccolte ha inoltre consentito di documentare la riproduzione del Lupo nel Parco del Beigua; nel 2012 questa è avvenuta con due nuclei distinti, che si possono localizzare nel settore centrale ed in quello nord-occidentale dell’area indagata.
Ulteriori approfondimenti porteranno poi a defi nire meglio il rapporto tra i sessi, le classi di età ed eventualmente riconoscere i singoli soggetti presenti nella popolazione in esame, ma è già stato possibile individuare un maschio adulto (probabilmente dominan-te) le cui caratteristiche del mantello lasciano supporre che si tratti di un caso di ibrida-zione tra Lupo e Cane.
La valutazione congiunta di tutte le informazioni disponibili permette ora di defi nire con maggiore precisione il probabile areale occupato dal Lupo nel Parco del Beigua (vi-sualizzato nella mappa con tonalità di rosso crescenti in relazione alla distribuzione delle osservazioni; realizzata mediante Kernel Analysis); possiamo quindi individuare un am-pio territorio che occupa la quasi totalità dell’area centro-occidentale (ed ospitante i due nuclei di riproduttori osservati nel 2012; per i quali, in base anche alle risultanze ottenute con il fototrappolaggio, i confini non appaiono assolutamente netti), ed un secondo nel settore orientale, riconducibile al nucleo da anni insediatosi nel Parco delle Capanne di Marcarolo [a].
Per approfondimenti si rimanda a: Campora et al. (2009), Caniglia et al. (2010), Genovesi (2002), Spagnesi & De Marinis (2002).
[a]
L’analyse des images rassemblées a montré la reproduction du Loup dans le Parco del Beigua ; en 2012 cela est arrivée en deux noyaux distingués, qui se peuvent localiser dans le secteur central et nord occidental de l’aire soumise à enquête. Des ultérieures renseignements emmèneront puis à mieux définir le rapport entre les sexes, les classes d’ancienneté et éventuellement reconnaître les singles sujets qui sont présents dans la population sous enquête, mais il est déjà devenue possible l’individuation d’un mâle adult (probablement dominant) dont les caractéristiques du manteau laissent supposer qu’il s’agit d’un cas d’hybridation entre Loup et Chien. L’évaluation conjointe des toutes les renseignements disponibles permets aujourd’hui de définir avec une meilleure précision la potentielle aire occupée par le Loup dans le Parco del Beigua (visualisé dans la mappe avec une tonalité rouge croissante en relation à la distribution des observations ; effectuée avec Kernel Analysis); on peut donc identifier un grand territoire qui occupe presque la totalité de l’aire centro occidentale (et hébergeant les deux noyaux de reproduction observés en 2012 ; dont en accord aux résultats des trappes photographiques les confins ne semblent pas tout à fait définis), et un deuxième dans le secteur oriental, reconductible au noyau qui depuis plusieurs années s’installait dans le Parco delle Capanne di Marcarolo [a].
Pour renseignements: Campora et al. (2009), Caniglia et al. (2010), Genovesi (2002), Spagnesi & De Marinis (2002).