Embed Size (px)
Citation preview

Wilhelm Roux's Archives 183, 223-231 (1977) Roux's Archives of Developmental Biology �9 by Springer-Verlag 1977
Formation de la membrane de f6condation de roeuf d'Artemia salina
G. De Maeyer-Criel, N. Fautrez-Firlefyn et J. Fautrez
Laboratoire d'Anatomie, Universit~ de Gand, Belgique
Formation of the Fertilization Membrane of the Egg in Artemia salina
Summary. The unfertilized egg of Artemia salina is not covered with any extracellular structure. No special organelles are found in the sub-cortical plasm. From the moment of fertilization, a membrane is progressively secreted by the egg. The membranogenous substance is first seen as large granules in the smooth endoplasmic reticulum, presumably transformed within Golgi elements and extruded in vesicles liberated from the Golgi apparatus. Retained by a glycocoat or by contact with the fluid of the genital tract, it spreads out into a fertilization membrane, soon surrounding a perivitelline space. The process lasts till 1 1/2 h after fertilization.
Key words: Fertilization membrane - Artemia salina.
R~sum~. L'oeuf vierge d'Artemia salina n'est pas entour6 de membranes exocellulaires. Le plasme sous-cortical ne contient pas d'organites sp6ciaux. D6s la f6condation, une membrane est secr6tde par l'oeuf. La substance membranog6ne, contenue dans le reticulum endoplasmique lisse, passe par les 616ments golgiens, oti elle semble modifi6e, et est expuls6e dans des v6sicules qui se d6tachent du Golgi. Retenue par un enduit granuleux, qui couvre le plasmolemme, et qui peut 6tre un glycocoat ou du suc du tractus g~nital, elle s'dtale en une membrane de fdcondation, qui se soul6ve pour constituer l'espace p6rivitellin. Le processus est progressif et dure environ une heure et demi.
Introduction
L'oeuf d'Artemia salina sou16ve une membrane de f6condation environ une heure et demi apr6s la p6n6tration du spermatozo'ide, qui a lieu d6s sa descente

224 G. De Maeyer-Criel et al.
dans l'ovisac. Le ph6nom6ne est visible in vivo. Les oeufs mats et i6g6rement aplatis, deviennent brillants et sph6riques. La membrane, sous tension, refl6te sans doute la lumi6re.
En 1961, deux d'entre nous (Fautrez et Fautrez-Firlefyn) ont &udi~ son soul6vement en collaboration avec Anteunis. Ces recherches, ~ l'aide du micro- scope 61ectronique, avaient conduit/~ la conclusion que la membrane de f6conda- tion se forme simplement par d61amination du plasmolemme.
Comme l 'oeuf mat d'Artemia n'est entour6 ni d 'un chorion, ni d 'une mem- brane vitelline, ni d'une gangue muqueuse, comme c'est les cas pour les germes examin6s ~ ce point de vue, au microscope 61ectronique, il nous a sembl6 int6res- sant de reprendre cette &ude, 5~ un moment o/l la technique ultramicroscopique a fortement 6volu6, et off elle permet un examen plus pr6cis, particuli6rement en ce qui concerne du mat6riel embryologique.
Materiel et technique
Les animaux proviennent d'une culture entretenue au laboratoire /t partir de germes enkyst6s et d'origine californienne. I1 s'agit d'une race amphigonique diplofde. Les oeufs sont pr61ev& dans l'oviducte ou dans l'ovisac. Ils sont fix6s au glutarald6hyde et au t6troxyde d'osmium, tamponn~ au cacodylate 0,2 M A pH 7,2, <<color~s>> in toto ~ l'ac6tate d'uranyle (Farquhar et Palade, 1965) et inclus dans l 'Epon. Les coupes ultrafines sont contrast6es ~ l'ac6tate d'uranyle (Okhura et Takashio, 1966) et au citrate de plomb (Venable et Coggeshall, 1965).
Observations personnelles
1. Oeufs prOlev~s dans l'oviducte (Fig. 1)
Les oeufs descendus dans l'oviducte sont tous au stade de la m~taphase de la premi+re division de maturation. Le fuseau est parall61e/t la surface. Le plasme sous-cortical est relativement ~pais et basophile. On y observe de nombreuses citernes de reticulum endoplasmique lisse, ainsi que des mitochondries. Ce reticulum endoplasmique contient de nombreux gros granules mod6r6ment osmiophiles. L'hyaloplasme est bourr~ de ribosomes, et deci-del/t on observe un microtubule. De minces tubules de reticulum endoplasmique rugueux ne s'observent que sporadiquement; leur lumi+re n'est pas osmiophile.
Le germe est entour~ d 'un plasmolemme peu tendu, qui est une unit-membrane dont la structure trilaminaire est 6vidente aux endroits off l'angle de section est favorable (fl6ches de la Fig. 1). Son 6paisseur totale est de l 'ordre de 100 ~. Sa face externe est couverte d 'un pr~cipit~ granulaire, d'environ 250 ~ d'6paisseur maximum, qui pourrait ~tre le glycocoat, mais certainement pas un chorion. I1 n'est pas exclu cependant, qu'iI s'agisse d 'un artefact de fixation: pr6cipit6 de suc de i'oviducte.
2. Oeufs prdlevds au moment de leur descente clans l'ovisac (Fig. 2)
C'est dbs leur descente dans l'ovisac, que les oeufs sont f6cond6s. On est frapp~ par la presence de nombreux complexes golgiens, que nous n'avons jamais trouvfis dans le plasme sous-cortical des oeufs vierges de l'oviducte. Ces appareils sont habituellement form6s de trois saccules et d'assez rares v6sicules, qui semblent s'en d6tacber. Leur membrane est plus osmiophile que celle du reticnlum endoplasmique: certaines images semblent suggerer une continui't+ entre les deux syst+mes. A c e moment les saccules sont pratiquement vides et tr6s aplatis. Les rares v6sicules, ainsi que les
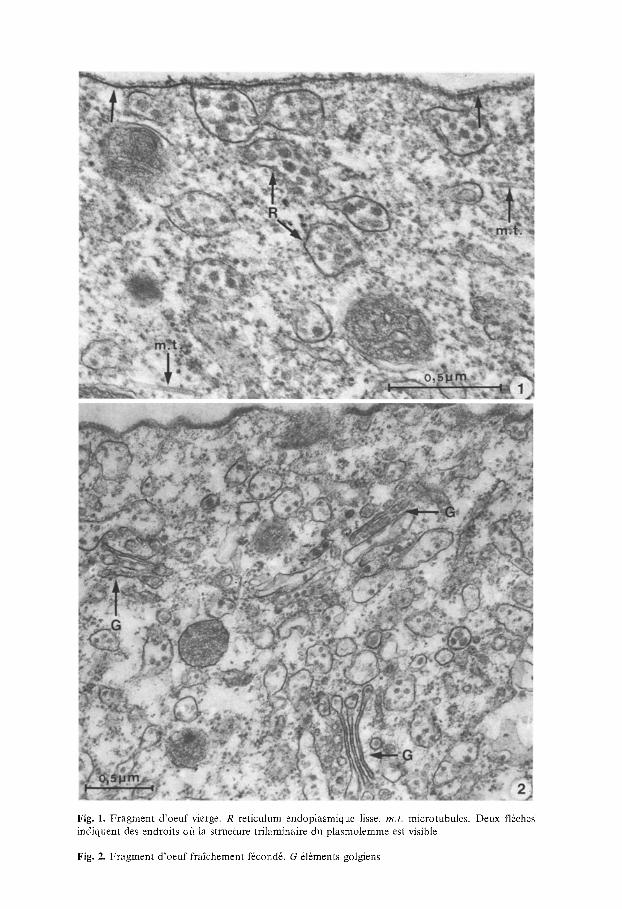
Fig. 1. Fragment d'oeuf vierge. R reticulum endoplasmique lisse, m.t . microtubules. Deux fl6ches indiquent des endroits o/1 la structure trilaminaire du plasmolemme est visible
Fig. 2. Fragment d'oeuf fra]chement f6cond6. G dl~rnents golgiens
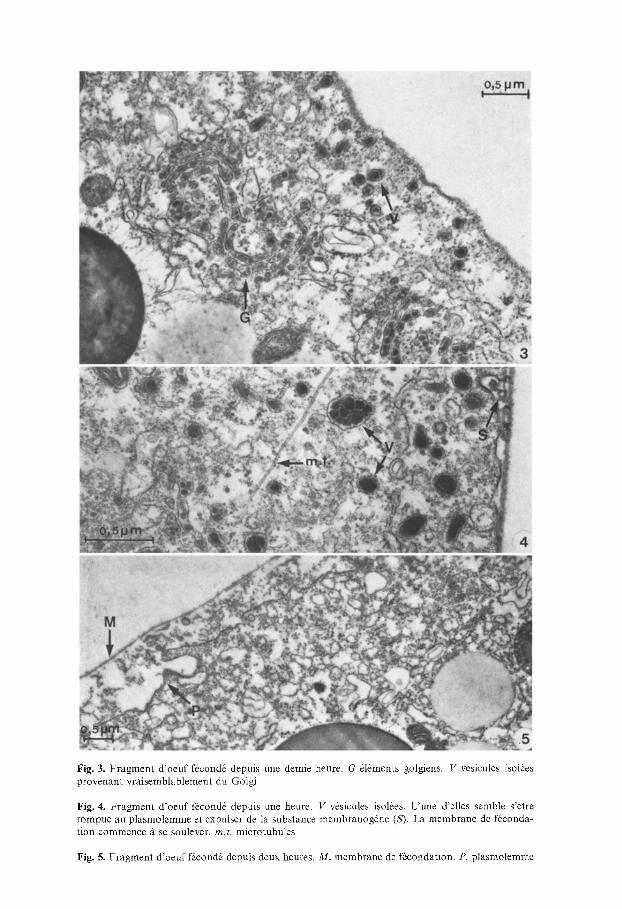
Fig. 3. Fragment d 'oeuf f~cond6 depuis une demie heure. G 616ments golgiens. V v6sicules isol6es provenant vraisemblablement du Golgi
Fig. 4. Fragment d 'oeuf f~cond6 depuis une heure. V v6sicules isol6es. L'une d'elles semble s'6tre rompue au plasmolemme et expulser de la substance membranog+ne (S). La membrane de f6eonda- tion commence fi se soulever, rn.t. microtubules
Fig. 5. Fragment d 'oeuf f~cond~ depuis deux heures. M. membrane de f6condation. P. plasmolemme

Membrane de facondation chez Artemia 227
bords des saccules contiennent des granules, semblables/~ ceux du reticulum endoplasmique, parfois lagarement plus osmiophiles.
Pour le reste la composition de la r~gion cortico-sous-corticale n'a gu~re chang&
3. Oeufs prdlevOs une demie heure aprbs la descente dans l'ovisac (Fig. 3)
A c e moment, on n'est pas seulement frapp6 de la grande abondance d'appareils de Golgi dans le plasme sous-cortical, mais agalement par des signes de grande activitd. On observe de nombreuses vasicules de transition entre reticulum endoplasmique et Golgi. Les saccules sont bourras de gros grains fortement osmiopbiles. Des vasicules s'en d6tachent: elles contiennent habituellement un seul grain osmiophile semblable/l ceux des saccules. Les images suggarent que ces v~sicules migrent, en grand nombre, vers la surface de l'oeuf. Certaines images pourraient ~tre interprataes comme rasultant de l'~clatement en surface de vasicules, rejetant leur contenu ~ la surfac e du plasmolemme.
Le reticulum endoplasmique lisse semble contenir moins de granules qu'aux stades pracadents. A partir de ce moment les minces tubules de reticulum endoplasmique rugueux deviennent
plus nombreux; ils ne contiennent jamais de gros granules osmiophiles.
4. Oeufs prOlevOs une heure aprbs la descente dans l'ovisac (Fig. 4)
L'activita golgienne semble encore plus prononcae. Des vasicules plus grandes s'observent sous le cortex. Leur contenu est parfois forma de nombreux grains fortement osmiophiles, parfois d'une seule masse de substance osmiophile confluae. On retrouve du reticulum endoplasmique lisse; il est beaucoup plus vide qu'aux stades plus jeunes. De rares microtubules sont encore prasents.
En surface on trouve le plasmolemme ondula. De nombreux tubules de reticulum endoplasmique y sont accolas. On y voit des coated pits' avec un contenu osmiophile, qui semble s'incorporer dans une membrane de facondation, qui commence/t se dacoller du plasmolemme. On peut s'imaginer que cette membrane se forme /t partir du produit osmiophile excreta par les 6laments golgiens. Ces produits semblent s'incorporer dans le glycocoat, qui se datache par endroits du plasmolemme.
A c e moment l'oeuf est encore mat, la membrane de facondation ne commen?ant qu'/t se soulever par endroits. Aces endroits, Ie plasmolemme est lisse.
5. Oeufs brillants (descendus clans l'ovisac depuis plus d'une heure el demi) (Fig. 5)
Une membrane de f~condation est soulevee sur ces oeufs brillants: elle est finement striae. La surface ovulaire s'est profondament modifiae. Alors qu'elle n'dtait que tras lagarement ondulae,
aux stades antarieurs, elle prasente de larges digitations repliaes, affaissaes Ies unes sur les autres et constituant ainsi un feutrage tras dense. Le plasmolemme reste mince et lisse. Quelques coated pits persistent sporadiquement. Dans Ie plasme sous-cortical on retrouve des mitochondries, du reticulum endoplasmique lisse et rugueux. Les aliments golgiens sont beaucoup moins nombreux et vidas de leur contenu osmiophile.
Plus tard l'espace parivitellin se remplit de dabris cellnlaires expulsas.
Discussion
La rdg ion c o r t i c o - s o u s - c o r t i c a l e d u g e r m e j o u e un r61e cap i t a l g d ivers m o m e n t s
du d d v e l o p p e m e n t e m b r y o n n a i r e . Elle sub i t n o t a m m e n t des t r a n s f o r m a t i o n s
m o r p h o l o g i q u e s e t p h y s i c o - c h i m i q u e s i m p o r t a n t e s apr~s la f a c o n d a t i o n ou apras
l ' a c t i v a t i o n e x p a r i m e n t a l e de l ' oeuf , c o n n u e s sous te n o r a de r~act ion cort icale .

228 G. De Maeyer-Criel et al.
A c6t6 de la remise en marche des divisions de maturation, qui en est vraisembla- blement une cons6quence (cf. les exp6riences de mise fi l'unisson du noyau spermatique chez l'Oursin, Brachet, 1922), de r6actions profondes et de modifica- tions m6taboliques, elle caract6rise l'activation.
La r6action corticale est complexe et se fait selon des modalit6s tr6s diff6rentes d'une esp6ce g l'autre. Son aspect le plus anciennement connu, mais non unique, est l 'apparition d'un espace p6rivitellin, attribu6 au soul6vement de la membrane de fOcondation. Cette derni6re d6nomination est cependant impropre dans la plupart des cas. L 'oeuf vierge est tr6s souvent entour6 de structures anhistes exocellulaires. On peut observer une membrane plus ou moins 6paisse, la mem- brane vitelline, ou une coque plus structur6e, le chorion, ou bien encore les deux (la membrane vitelline tapissant la face interne du chorion). Ces 616ments peuvent ~tre produits dans l'ovaire soit par l'oocyte, soit par des cellules follicu- leuses, soit encore par les deux. I1 suffit que l 'oeuf se retracte, apr~s f6condation (et m~me parfois avant!) pour qu'apparaisse un espace p6rivitellin. Chez de nombreuses esp6ces, des microvillosit6s ovulaires s'incrustent dans le chorion et seront rompues ~ leur base lors de la formation de l'espace p6rivitellin.
I1 est 6vident que, dans ces conditions, ni ta membrane vitelline ni le chorion, pr6existants, ne m6ritent le nora de membrane de f6condation. Dans certaines esp6ces, cependant, la formation d'une nouvelle membrane a ~t6 d~crite au moment de la f6condation ou imm6diatement apr6s: ce serait une membrane de f6condation r6elle. Elle se place contre les gaines pr6existantes, autour de l'espace p6rivitellin et pourrait aussi se d6velopper autour d'oeufs fi plasmolemme nu, tels ceux d'Artemia salina. C'est ainsi que Grey et ai. (1974) d6crivent, chez Xenopus laevis, une couche de fOcondation venant s'apposer g la face pro- fonde d'une membrane vitelline pr6existante. Cette couche se constitue ~ partir de granules corticaux: ce mat6riel finit par traverser la membrane vitelline et par s'6taler fi sa face externe. Ce ph6nom6ne se passe en 8 fi 9 min. apr6s addition de sperme (f6condation in vitro). La membrane pr6existante serait perm6able au spermatozo'ides, la membrane complexe leur formerait une barri6re (Grey et al., 1976).
La microscopie 61ectronique a permis de pr6ciser la morphologie et la signifi- cation des membranes ovulaires et du plasme sous-cortical, sans que les r6sultats ne se superposent toujours clairement aux donnbes d'autres mbthodes. Le pro- bl6me m6rite donc certes une 6tude comparative plus pouss6e.
Pasteels (1961) a consacr6 une importante revue comparative au probl6me qui nous int~resse ici: le lecteur y trouvera une bibliographie 6tendue jusqu'g cette date.
Depuis lors, les divers aspects de la rbaction corticale ont ~tb r6&udi6s au microscope 61ectronique et ont donn6 lieu /t de nombreuses contributions. En ce qui concerne les Crustac6s, et pour autant que nous soyons inform6s, la note de Anteunis et al. (1961) reste le seul document concernant les remaniements suite ~ la f6condation. C'est ce qui nous a mend ~t reprendre cette ancienne 6tude sur l 'oeuf d'Artemia salina, en esp6rant pouvoir tirer parti des perfectionne- ments de la technique comme de donn6es comparatives r6centes. Ne trouvant rien concernant le ph6nom~ne chez d'autres Crustac6s, force nous sera de compa-

Membrane de f6condation chez Artemia 229
rer nos r6sultats avec des donn6es fournies par les Arthropodes plus ~volu~s, les Insectes, qui connurent plus de succ~s!
L'oocyte de Bombus ferrestris, 6tudi~ par Hopkins et King (1966), est entour6 de cellules folliculaires. Apr~s la vitellog~n~se, une membrane vitelline se consti- tue dans l'interstice entre l 'oocyte et l '@ithelium folliculaire ; l 'un comme l'autre contribuent fi sa formation. Plus tard un chorion est s~cr6t~ contre la face externe de la membrane vitelline par le seul epithelium folliculaire.
Chez la Libellule (Beams et Kessel, 1969) comme chez Ephestia kuhniella (Cummings, 1972)~ l 'oocyte n'interviendrait ni dans la formation de la membrane vitelline, ni ensuite dans celle du chorion. Les deux structures seraient form~es exclusivement par l '@ithelium folliculaire.
Chez Drosophila melanogaster, des cellules folliculaires, qui ne sont pas en contact avec un oocyte, sont capables de s 'entourer d'une membrane semblable /~ la membrane vitelline. I1 semble donc bien qu'elles jouent le r61e principal dans la s6cr&ion de cette derni~re. Plus tard elles forment le chorion (King et Koch, 1963).
La situation est tras diff6rente chez Artemia. En effet il ne se forme, dans l'ovaire, ni follicule, ni chorion, ni membrane vitelline. I1 existe bien des cellules nourrici~res, mais elles sont phagocyt6es en grand nombre et compl~tement incorpor6es dans la partie superficielle de l 'oocyte en vitellog6nase (Fautrez- Firlefyn, 1951) (Anteunis et al., 1966). Lorsque celui-ci quitte l'ovaire vers l'ovi- ducte, son plasmolemme n'est entour~ que d'un mince d@6t finement granulaire, qui pourrait ~tre un glycocoat. C'est donc l 'oocyte lui-mame qui formera la substance de sa membrane de f6condation.
Si l 'on peut assimiler la coque des oeufs durables, / t un chorion, ce dernier sera form6 par des produits de glandes coquilliares, et ne se d@osera qu'aux premiers stades de la segmentation. Plus tard encore, le germe s6cr6terait la couche chitineuse profonde de cette coque (Fautrez et Fautrez-Firlefyn, 1971).
Pour ce qui est de la synth6se de la substance, qui constituera la membrane defOcondation, celle-ci a lieu d6s la descente dans l'ovisac et donc d6s la f6conda- tion, dans le cytoplasme sous-cortical. Nous ne pouvons d6finir ce dernier que de mani6re n6gative. I1 ne contient en effet que des organites cytoplasmiques banaux; on peut donc simplement dire que c'est la mince couche p6riph6rique du cytoplasme, qui, apr~s la vitellogdndse, n'est pas envahie par des grains de vitellus. Dans l 'oeuf vierge, on y trouve un reticulum endoplasmique lisse tr6s d6velopp6 dans un hyaloplasme bourr6 de ribosomes libres, des mitochon- dries et des microtubules. On est frapp6 par l'absence d'61dments golgiens (qu~ l'on trouve par contre de ci-de 1/~ en profondeur, entre les grains de vitellus), et par l'extr6me raret6 du reticulum endoplasmique rugueux.
D6s la f6condation, on voit d6buter un processus de synth6se, qui est tr6s semblable fi celui de la synth6se de la substance destinde/~ la membrane vitelline dans les cellules folliculeuses des Insectes, dont il vient d'6tre question. Les premiers granules de substance membranog6ne s'observent cependant dans un reticulum endoplasmique lisse (et non rugueux, comme c'est le cas chez les Insectes 6tudi6s). Le reticulum endoplasmique rugueux, qui sera plus d6velopp6 vers la fin du processus, ne semble pas y participer. On n'y trouve jamais

230 G. De Maeyer-Criei et al.
de con tenu granuleux. La suite des 6v6nements est semblable fi ce qui a 6t6 d~crit dans les cellules foll iculeuses des Insectes: in tervent ion d'~16ments gol- giens, avec modi f i ca t ion vra i semblab le des granules, l ibdra t ion des v6sicules, don t le con tenu est 61imin6 pa r exocytose. Ce processus c o r r e spond g u n m6ca- nisme g6n6ral de s6cr6tion d6crit dans les cellules somat iques les plus diverses.
La substance s6cr6t6e est comme retenue pa r le d6p6t, qui couvre le p l a smo- lemme, - g lycocoat ou suc ut6rin - et s '6tale en une membrane . Le soul6vement de celle-ci donne lieu /t la f o r m a t i o n de l 'espace p6rivitellin, dans lequel des d6bris ovulai res a p p a r a i t r o n t plus tard. N o t o n s qu 'apr~s le soul+vement, il n 'es t
plus de d6p6t visible sur le p lasmolemme. Le p lasme sous-cor t ica l de l ' oeu f vierge et f ra ichement f6cond6 pr6sente
d 'assez n o m b r e u x micro tubules . On peu t se d e m a n d e r s 'ils in terv iennent dans les d6placements de mat6riel , c o m m e cel~ rut soup~onn6, lors de la vitellog6n6se de l ' A n o d o n t e (Beams et Sekhon, 1966), chez les leucocytes (Golds te in et al., 1970; Zur ie r et al., 1973, 1974; Anteun is et al., 1977).
Les donn6es ici recueill ies nous pe rmet t en t de pr6ciser la note ant6r ieure de An teun i s et al. (1961). L ' o e u f descendu dans l 'u terus ne pr6sente pas de membranes exocellulaires. Si le soul6vement d 'une m e m b r a n e de f6condat ion ne devient visible ~ la loupe que tr6s ta rd ivement , sa s6cr6tion pa r l ' oeu f est un ph6nom6ne tr6s progressif , qui d6bute d6s le m o m e n t de la f6condat ion. Dans le cas par t icu l ie r d ' A r t e m i a sal ina, il nous para~t donc l~gitime de par le r d 'une m e m b r a n e de f6condat ion.
Bibliographic
Anteunis, A., Astesano, A., Robineaux, R. : Ultrastructural characteristics of developing eosinophil leukocytes in human bone marrow during acute leukemia. Evidence for extracellular granular release from human eosinophiles. Inflammation, 2, i7-26 (1977)
Anteunis, A., Fautrez-Firlefyn, N., Fautrez, J. : Ultrastructure du cortex et du plasme sous-cortical de l'oeuf d'Artemia salina. C.R. Soc. Biol. 155, 1393-1394 (1961)
Anteunis, A., Fautrez-Firlefyn, N., Fautrez, J. : L'incorporation de cellules nourrici6res par I'oocyte d'Artemia salina. Etude au microscope 61ectronique. Arch. Biol. 77, 665-676 (1966)
Beams, H.W., Kessel, R.G.: Synthesis and deposition of oocyte enveloppes (vitelline membrane, chorion) and the uptake of yolk in the Dragonfly (Odonata Aeschnidae). J. Cell Sci. 4, 241-264 (1969)
Beams, H.W., Sekhon, S.S.: Electron microscope studies on the oocyte of the fresh-water mussel (Anodonta) with special references to the stalk and mechanism of yolk deposition. J. MorphoI. 114, 477-502 (1966)
Brachet, A.: Sur la f6condation pr6matur6e de l'oeuf d'Oursin. C.R. Soc. Biol. 136, 511 512 (1922) Cummings, M.R.: Formation of the vitelline membrane and chorion in developing oocytes of
Ephestia kuhniella. Z. Zellforsch. 127, 175-188 (1972) Farquhar, M., Palade, G.: Cell junctions on the amphibian skin. J. Cell Biol. 26, 263-291 (1965) Fautrez, J., Fautrez-Firlefyn, N.: Contribution ~ l'6tude des glandes coquilli6res et des coques
de l'oeuf d'Artemia salina. Arch. Biol. 82, 41 --83 (1971) Fautrez-Firlefyn, N.: Etude cytochimique des acides nuclfiiques au cours de la gam6tog~n6se et
des premiers stades du d~veloppement embryonnaire chez Artemia salina. Arch. Biol. 62, 391 438 (1951)
Goldstein, I., Hoffstein, S., Gallin, J., Weissmann, G.: Mechanism of lysosomal assembly and membrane fusion induced by a component of complement. Proc. Nat. Acad. Sci. USA 71), 2916-2920 (1973)

Membrane de f~condation chez Artemia 231
Grey, R.D., Wolf, P., Hedrick, J.L.: Formation and structure of the fertilization envelope in 32enopus laevis. Develop. Biol. 36, 44-61 (1974)
Grey, R.D., Working, P.K., Hedrick, J.L.: Evidence that the fertilization envelope blocks sperm entry in eggs of Xenopus laevis. Interaction of sperm with isolated envelopes. Develop. Biol. 54, 52 -60 (1976)
Hopkins, C.R., King, P.E. : An electron-microscopical and histochemical study of the oocyte peri- phery in Bombus terristris during vitellogenesis. J. Cell Sci. 1, 201-216 (1966)
King, R.C., Koch, E.A.: Studies on the ovarian follicle cells of Drosophila. Quart. J. Microsc. Sci. 104, 297-320 (1963)
Okhura, T., Takashio, M. : BeitrS, ge zur Verbesserung der Elektronenffirbung mit den aus nichtwgtsse- rigen Flftssigkeiten hergestellten Uranylacetatl6sungen. Arch. Histol. Jap. 27, 49 -56 (1966)
Pasteels, J. : La r~action corticale de f6condation ou d'activation (Revue comparative). Bull. Soc. Zool. France 86, 600-629 (1961)
Venabte, J., Coggeshall, R.: A simplified lead citrate stain for use in electron microscopy, J. Cell Biol. 25, 407-408 (1965)
Zurier, R.B., Hoffstein, S., Weismann, G.: Cytochalasin B: Effect on lysosomal enzyme release from human leucocytes. Proc. Nat. Acad. Sci. USA 70, 844-848 (1973)
Zurier, R.B., Weismann, G., Hoffstein, S., Kammermann, S., Hsiung Tai, H.: Mechanism of lysosomal enzyme release from human leucocytes. II. Effects of c-AMP and c-GMP, autonomic antagonists and agents which affect microtubule function. J. Clin. Invest. 53, 297-309 (1974)
Refu le 27 avril 1977 Accept~ le 8 ao~t 1977