Embed Size (px)
Citation preview

Le systkme endophytique d'drceuthobiurn oxycedn'. II. Aspects ultrastructuraux des zones de contact entre les tissus de l'h8te et du parasite
A. SADIK,' L. REY ET S. RENAUDIN Laboratoire de Cytopathologie vLge'tale, Universite' de Nantes, 2, rue de la Houssinikre, 44072 Nantes CEDEX, France
R e ~ u le 21 novembre 1985
SADIK, A,, L. REY et S. RENAUDIN. 1986. Le systbme endophytique d'drceuthobium oxycedri. II. Aspects ultrastructuraux des zones de contact entre les tissus de l'h6te et du parasite. Can. J. Bot. 64 : 2778-2784.
L'Ctude ultrastructumle des zones de contact entre le systbme endophytique d'drceuthobium oxycedri et les tissus de son h6te, le genCvrier oxycbdre, montre qu'autour des plus gros cordons et sur les flancs des su~oirs primaires existe souvent, au niveau du phlobme secondaire de l'hdte, une zone d'kcrasement qui parait peu favorable aux transferts de substances. En revanche, la prksence, au niveau du xylbme secondaire, de demi-ponctuations arColCes ?u contact direct des cellules des su~oirs primaires et surtout secondaires est certainement de nature i faciliter les Cchanges. A la pointe des su~oirs, les cellules du parasite, au cours de leur progression, endommagent les cellules de l'h6te et font parfois irruption dans la lumibre de ses tracheides. Une forte activitC phosphatasique acide et ATP-asique est mise en Cvidence en microscopie Clectronique au niveau des plasmalemmes, des plasmodesmes ainsi que de certaines petites vCsicules d'endocytose ou d'exocytose dans les su~oirs primaires et secondaires. Elle tCmoigne de I'intensitC des phCnombnes de transport actif dans ces parties du systbme endo- phytique. Dans la rigion des zones de contact, une convergence d'aspect apparait entre les cellules de type cellulosique du parasite et de l'h6te.
SADIK, A., L. REY, and S. RENAUDIN. 1986. Le systbme endophytique d'drceuthobium oxycedri. II. Aspects ultrastructuraux des zones de contact entre les tissus de l'h6te et du parasite. Can. J. Bot. 64 : 2778-2784.
Ultrastructural study of the contact areas between the endophytic system of Arceuthobium oxycedri and the tissues of its host, Juniperus oxycedms, shows that a crushed zone often occurs around the large cortical strands and along the primary sinkers in the secondary phloem of the host; such a structure seems rather unfavourable to the transfer of substances. On the other hand, the presence in the secondary xylem of half-bordered pits in direct contact with the cells of the primary and espe- cially secondary sinkers certainly makes it easier for exchanges to take place. At the tip of the sinkers, the cells of the parasite, in the course of their growth, damage the cells of the host and sometimes penetrate into the lumen of its tracheids. Electron microscopy shows important acid phosphatase and ATPase activities at the level of the plasmalemma, plasmodesmata, and also some small endo- or exo-cytotic vesicles in the primary and secondary sinkers. Such activities are a sign of intense active transfer processes in these parts of the endophytic system. Around the contact zones, the parenchymatous cells of the host become very similar in aspect to those of the parasite.
Introduction LIArceuthobium, communCment appelC agui nainn est une
ViscacCe hemiparasite qui pousse sur les rameaux des co- niferes.
Au cours d'un travail prCcCdent (Sadik et al. 1986), nous avons CtudiC les caractbres cytologiques et cytochimiques des diffkrents cordons et su~oirs qui foment le systbme endo- phytique.
Le but du prCsent article est de complCter les rksultats obtenus par une Ctude plus prkcise, au niveau ultrastructural, des zones de contact entre les tissus de l'h6te et ceux du para- site, zones particulibrement importantes puisqu'elles sont le lieu des transferts de substance entre les deux partenaires.
Materiel Les Cchantillons d'drceuthobium oxycedri (DC.) M. Bieb. que
nous avons utilisCs proviennent de la rCgion de Manosque (Alpes de Haute-Provence, France) oh ils ont CtC rCcoltCs sur le genCvrier oxycbdre (Junipems oxycedms L.). Pour notre Ctude cytologique et cytoenzymologique, plusieurs dizaines de blocs sont dCcoupCs i des emplacements divers dans les rameaux parasites.
Methodes Prkparation des e'chantillons pour 1 'ktude en microscopie electronique
Le matCriel a CtC fix6 au glutaraldChyde i 2 % dans un tampon phos- phate ou cacodylate 0,l M (pH 7,3) pendant 2 h i tempkrature ambiante, puis au tCtroxyde d'osmium i 1 % dans le m&me tampon
'Auteur i qui faire parvenir toute correspondence. R i n t d in Canada 1 Imprim6 au Canada
pendant 1 h i tempCrature ambiante et inclus dans 1'Epon selon la technique de Luft (1961). Les coupes ultra-fines ont CtC contrastCes par I'acCtate d'uranyl et le citrate de plomb selon Reynolds (1963).
Localisation ultrastructurale d hctivitgs enzymatiques La mise en evidence de l'activite ATP-asique a CtC effectuCe par la
technique de Wachstein et Meisel (1957) selon le protocole dCcrit par Marx et al. (1982); la localisation des phosphatases acides a CtC CtudiCe par la technique de Gomori (1956) selon le protocole utilisC par Coulomb (1971).
Resultats ~ t u d e de la zone de contact
Les zones de contact entre l'h6te et le parasite n'ont pas le m&me aspect selon les diffkrentes parties du systbme endo- phytique et les differents tissus du rameau de genCvrier qu'elles traversent.
Les cordons de 1'A. oxycedri, qu'ils soient longitudinaux ou circulaires, sont toujours disposCs 2 1'intCrieur du phlobme secondaire du rameau parasit6 (Sadik et al. 1986). Les petits cordons n'entrainent aucun bouleversement notable dans l'organisation de ce tissu et leurs cellules aux parois Cpaisses sont au contact direct des cellules phlobmiennes. En revanche les gros cordons sont entourks, en raison m&me de leur dCveloppement, d'une zone dlCcrasement confuse et irrCgu- libre qui les sCpare plus ou moins du tissu environnant (fig. 1).
Les su~oirs primaires qui s'enfoncent profondkment dans le cylindre central de l'h6te et se prolongent vers 1'extCrieur par une tige aCrienne, traversent de ce fait plusieurs tissus.
La pointe d'un su~oi r primaire est toujours constituCe de cel- lules de type cellulosique caractCrisCes par une paroi t k s
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y G
LA
SGO
W U
NIV
ER
SIT
Y L
IBR
AR
Y o
n 09
/03/
13Fo
r pe
rson
al u
se o
nly.

SADIK ET AL. 2779
tpaisse, de nombreux chloroplastes, d'abondantes inclusions lipidiques; on y observe aussi souve?t des accumulations de profils de rtticulum endoplasmique. A leur contact immtdiat, les trachtides du xylkme secondaire du gentvrier, malgrt l'tpaisseur de leur parqi lignifite, apparaissent frtquemment endommagtes (fig. 2). A la faveur d'une dtchimre, une cellule de la pointe du s u ~ o i r peut parfois faire directement irruption dans la lumikre d'une de ces trachtides.
Sur les flancs du su~oi r , au niveau du xylkme secondaire du gentvrier, les cellules cellulosiques ou les trachtides du para- site sont ttroitement accoltes aux trachtides de l'h6te mais ce contact direct ne donne lieu i aucune particularitt notable. Cependant, des coupes semi-fines tangentielles i un s u ~ o i r montrent qu'autour de celui-ci les trachtides du gentvrier peu- vent former une sorte d'enveloppe concentrique en s'orientant perpendiculairement i leur direction habituelle (fig. 3).
Plus vers l'exttrieur, au niveau du cambium et surtout du phlokme secondaire du gentvrier, on observe souvent autour du s u ~ o i r primaire la prtsence d'une zone d'tcrasement con- fuse, plus ou moins tpaisse, formte de divers dtbris cellulaires parmi lesquels on reconnait ~i et 18 quelques fibres libtriennes (fig. 4). Au deli de cette zone, le tissu phlokmien apparait nettement modifit : alors que dans les rameaux sains, le phlokme secondaire se prtsente sous forme de petites cellules rtgulikrement aligntes, fortement vacuolistes et pauvres en inclusions (fig. 6), dans les rameaux parasitts, ces cellules sont plus grandes, posskdent un cytoplasme plus abondant et surtout des inclusions amyliferes et lipidiques beaucoup plus nombreuses (fig. 5). I1 est i noter que ces modifications s'observent tgalement autour des su~oirs secondaires et i un degrt moindre autour des cordons, si bien que dans une partie de rameau oh su~oirs et cordons sont nombreux, c'est l'ensemble du phlokme secondaire de l'h6te qui apparait ainsi transformt. De mCme que les cellules phlokmiennes, les cellules cambiales s'enrichissent en cytoplasme au contact des su~oirs.
Au niveau du parenchyme cortical de l'hhte, autour de la zone de jonction riche en vaisseaux lignifits qui relient un s u ~ o i r primaire i une tige atrienne, les cellules se chargent souvent de tanins et contiennent des chromoplastes (fig. 7) que nous n'avons pas retrouvts dans les cellules correspondantes du rameau sain.
Les su~oirs secondaires s'enfoncent tgalement profondt- ment dans le xylkme secondaire de l'h6te. Plus minces que les su~oirs primaires, ils sont constituts de cellules de type cellu- losique et de trachtides dispostes sans ordre apparent (Sadik et al. 1986). Cette organisation a deux constquences : il n'apparah jamais de zone d'tcrasement autour des su~oirs secondaires, ni au niveau du xylkme secondaire de lYh6te, ni au niveau du phlokme secondaire; par ailleurs, ce sont soit des cellules cellulosiques, soit des trachtides qui se trouvent au contact des tissus de l'h6te.
A la pointe du su~oi r , l'aspect des cellules cellulosiques avec leur paroi trks tpaisse et leurs abondantes inclusions lipidiques est le m&me que dans le cas d'un s u ~ o i r primaire et l'on peut observer de nombreuses figures de progression de ces cellules a l'inttrieur du rayon mtdullaire envahi (fig. 8).
C'est en effet clairement au niveau des rayons mtdullaires que les su~oirs secondaires s'enfoncent dans les tissus de l'h6te si bien que sur leurs flancs on observe des trachtides i demi-ponctuations artoltes, ponctuations qui i l'origine assu- raient une communication avec les cellules du rayon ligneux. Ces cellules ayant t t t remplackes par des cellules de s u ~ o i r
secondaire d'Arceuthobium, il existe donc de nombreux endroits oh la paroi des tltments de xylkme de lYh6te est amincie au contact du parasite (fig. 9).
Lorsqu'un rayon mtdullaire est envahi par un s u ~ o i r secon- daire et que, sur une coupe, on peut observer c6te i c6te les cellules du parasite et les cellules du rayon mtdullaire qui sub- sistent (fig. lo), on constate que les deux cattgories de cellules se ressemblent beaucoup par la densitt de leur cytoplasme et leur richesse en inclusions lipidiques. Cependant, les plastes, de type chloroplaste dans les cellules du parasite, de type amyloplaste dans les cellules des rayons mtdullaires sont nettement difftrents.
I1 est i noter que dans les rayons mtdullaires non envahis mais situts a proximitt de l'appareil haustorial du parasite, les cellules sont tgalement transformtes : alors que dans le rameau sain, ces cellules sont pauvres en cytoplasme, elles deviennent ici plus grandes avec un cytoplasme plus abondant, un vacuome plus morcelt et des inclusions amyliferes et lipi- diques plus nombreuses . ~ t u d e cytoenzymologique en microscopie e'lectronique
Localisation de 1 'activite' phosphatasique acide Par comparaison avec les coupes ttmoins (fig. 14), les
dtp6ts de phosphate de plomb dans les coupes essais indiquent une activitt des phosphatases acides dans les cellules du systkme endophytique, au niveau du plasmalemme soulignt sur toute sa surface, au niveau des noyaux, au niveau des plasmodesmes et au niveau des petites vacuoles (Fig. 1 1 i 13). Tous ces dtp6ts sont plus abondants dans les su~oirs primaires et secondaires que dans les cordons. Les cellules de l'h6te voisines de systkme endophytique, surtout dans le phlokme secondaire et les rayons mtdullaires, prtsentent tgalement un certain marquage, en particulier au niveau du plasmalemme et du noyau.
Localisation de l'activite' ATP-asique La rtaction qui se traduit par le m&me dtp6t de phosphate de
plomb (comparer la fig. 15 essai i la fig. 16 ttmoin) rtvkle une activitt ATP-asique dans les cellules du systkme endo- phytique, en particulier au niveau du plasmalemme des cellules des su~oirs primaires. Les petites invaginations du plasmalemme et les vtsicules sous-jacentes que nous avons d t j i signaltes dans ces cellules (Sadik et al. 1986) sont marqutes aussi.
Discussion et conclusion L'examen des zones de contact entre le svstkme endo-
phytique d'A. oxycedri et les tissus de gentvrier amkne i se demander comment se fait le passage des substances de l'h6te au parasite. L'existence de zones d'tcrasement autour des gros cordons et des su~oirs primaires, surtout au niveau du phlokme, ne para?t pas Ctre favorable aux tchanges. Cette zone d'tcrasement a t t t tgalement observte par Sallt (1977) autour du s u ~ o i r primaire du gui, au niveau duparenchyme cortical et du phlokme primaire et secondaire; l'tcrasement et l'orienta- tion perturbte des cellules environnantes sont causes par la largeur du c6ne de pknttration du s u ~ o i r primaire qui ne ren- contre pas beaucoup de resistance de la part de ces tissus. Mais au niveau du xylkme secondaire de l'h6te oh les parois cellu- laires lignifites sont plus rtsistantes, les perturbations sont minimes; selon Sallt (1977), cela est dO tgalement 2 la prtsence d'une zone mtristtmatique intercalaire au niveau du cambium qui permet une croissance du s u ~ o i r primaire en har-
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y G
LA
SGO
W U
NIV
ER
SIT
Y L
IBR
AR
Y o
n 09
/03/
13Fo
r pe
rson
al u
se o
nly.

2780 CAN. J . BOT. VOL. 64, 1986
FIG. 1. Gros cordon longitudinal, en sectio? transversale. Il est form6 d'un grand nombre de cellules et comporte en son centre des ClCments conducteurs (EC), trachCides ou vaisseaux. A sa pCriphCrie, des zones comprim6es ou CcrasCes sont visibles dans le phlobme secondaire qui l'entoure (flbches). x220 (Cchelle, 100 pm). FIG. 2. ExtdmitC d'un s u ~ o i r primaire. Au-dessous des cellules de la pointe caractCris6es par leur richesse en inclusions lipidiques (L) et leur paroi cellulaire Cpaisse (PC) on observe une zone d'kcrasement (*) dsultant de la compression des trachCides du xylbme secondaire de 17h6te. x7000 (Cchelle, 2 pm). FIG. 3. Coupe tangentielle d'un s u ~ o i r primaire : les tracheides du genCvrier foment autour de lui une enveloppe concentrique en s'orientant perpendiculairement i leur direction habituelle. x220 (Cchelle, 100 pm). FIG. 4. Contact entre un s u ~ o i r primaire (SP) et le phlobme secondaire de l'h6te. Le long des cellules du su~oi r , on observe une zone irrkgulibre d'Ccrase- ment plus ou moins Cpaisse (*). Dans cette zone confuse, on reconnait ici une fibre libCrienne lignifiCe (F). ~ 3 8 0 0 (Cchelle, 2 pm). FIG. 5. Cel- lule 1ibCrienne de rameau parasit6 de genCvrier riche en cytoplasme et en inclusions lipidiques (L) et amyliferes (A). N, noyau. ~ 3 8 0 0 (Cchelle, 2 pm). FIG. 6 . Cellule 1ibCrienne tCmoin de rameau sain de genCvrier. V, vacuole. ~ 3 9 0 0 (Cchelle, 2 pm).
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y G
LA
SGO
W U
NIV
ER
SIT
Y L
IBR
AR
Y o
n 09
/03/
13Fo
r pe
rson
al u
se o
nly.
SADIK ET AL
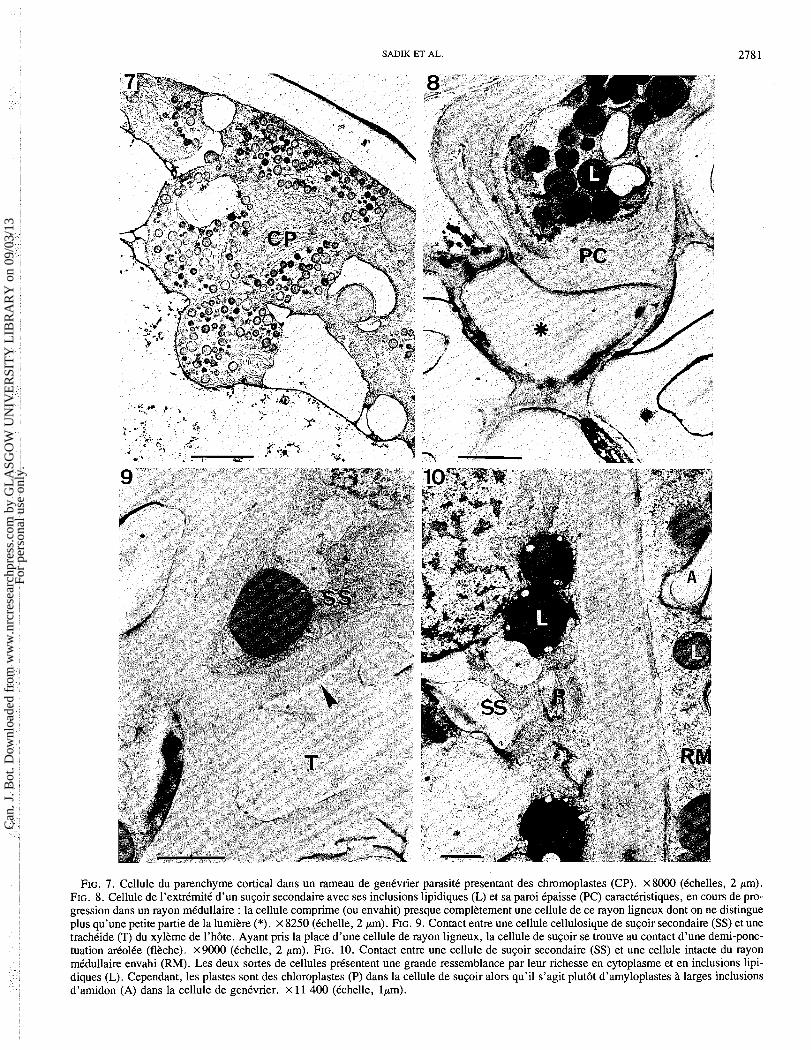
FIG. 7. Cellule du parenchyme cortical dans un rameau de genCvrier parasit6 presentant des chromoplastes (CP). ~ 8 0 0 0 (Cchelles, 2 pm). FIG. 8. Cellule de I'extrCmitC d'un s u ~ o i r secondaire avec ses inclusions lipidiques (L) et sa paroi Cpaisse (PC) caractCristiques, en cours de pro- gression dans un rayon maullaire : la cellule comprime (ou envahit) presque complktement une cellule de ce rayon ligneux dont on ne distingue plus qu'une petite partie de la lumiere (*). x 8250 (Cchelle, 2 pm). FIG. 9. Contact entre une cellule cellulosique de s u ~ o i r secondaire (SS) et une trachCide (T) du xylkme de l'h6te. Ayant pris la place d'une cellule de rayon ligneux, la cellule de s u ~ o i r se trouve au contact d'une demi-ponc- tuation arColCe (flkche). x9000 (Cchelle, 2 pm). FIG. 10. Contact entre une cellule de su~oi r secondaire (SS) et une cellule intacte du rayon maullaire envahi (RM). Les deux sortes de cellules pksentent une grande ressemblance par leur richesse en cytoplasme et en inclusions lipi- diques (L). Cependant, les plastes sont des chloroplastes (P) dans la cellule de s u ~ o i r alors qu'il s'agit plut6t d'amyloplastes i larges inclusions d'amidon (A) dans la cellule de genCvrier. x 11 400 (Cchelle, lpm).
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y G
LA
SGO
W U
NIV
ER
SIT
Y L
IBR
AR
Y o
n 09
/03/
13Fo
r pe
rson
al u
se o
nly.

CAN. I. BOT. VOL. 64, 1986
FIG. 11 i 14. Mise en Cvidence de I'activitC phosphatasique acide dans les cellules d'un s u ~ o i r secondaire. Fig. 11 i 13. Essai. Fig. 14. tCmoin. Fig. 11. Coupe essai. Le pr6cipitC de phosphate de plomb indiquant une activitC enzymatique est present surtout au niveau du plas- malemme. Le noyau (N) est Cgalement un peu marque. On ne note en revanche aucune rCaction dans les trachCides (T). ~ 4 8 0 0 (Cchelle, 2 pm). Fig. 12. Marquage des plasmodesmes indiquant une forte activitC phosphatasique Z i leur niveau. x4200 (Cchelle, 2 pm). Fig. 13. Marquage des petites vacuoles (V) ainsi que de vCsicules dont certaines sont situCes dans le cytoplasme (flbche) et d'autres ouvertes sur l'espace pCriplasmique (double flbche). ~ 9 6 0 0 (Cchelle, 1 pm). Fig. 14. Sur le tCmoin n'apparaissent que de lCgers dCpBts qui correspondent peut-&tre i la presence de phosphates libres. ~ 6 6 0 0 (Cchelle 2 pm). FIG. 15 et 16. Mise en Cvidence de I'activitC ATP-asique dans les cellules d'un s u ~ o i r primaire. Fig. 15. Coupe essai. Le marquage indiquant une activitC enzymatique est surtout important au niveau du plasmalemme et des plasmodesmes. Certaines petites vCsicules, les unes situCes dans le cytoplasme pCriphCrique (flbche), les autres ouvertes sur l'espace periplasmique (double flbche), sont marquCes. X 15 300 (Cchelle, 1 pm). Fig. 16. Coupe tCmoin. x 14 950 (Cchelle, 1 pm).
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y G
LA
SGO
W U
NIV
ER
SIT
Y L
IBR
AR
Y o
n 09
/03/
13Fo
r pe
rson
al u
se o
nly.

SADIK ET AL. 2783
monie avec celle de l'h6te. En ce qui conceme 1'A. oxycedri, nous n'avons pas observt l'existence d'une telle zone mtri- sttmatique.
Quoi qu'il en soit, c'est au niveau du xylkme de l'h6te que s'observent les contacts les plus ttroits entre les tissus de l'h6te qui sont ici des trachtides artoltes et les cellules cellulosiques ou les trachtides des su~oirs. Dans cette zone, les semi-ponc- tuations artoltes qui sont en contact direct avec les cellules des su~oirs secondaires contribuent certainement B favoriser le passage des substances. Un dispositif particulier qui a parfois Ctt observk est la ptnttration d'une cellule a paroi cellulosique de l'extrtmitt d'un s u ~ o i r B l'inttrieur d'une trachtide du xylkme secondaire de l'h6te. Ce phtnomkne rappelle ce que Renaudin (1974) a observt chez Lathraea clandestina L. et ce que Renaudin et Capdepon (1979) ont observC chez Touia alpina L. au niveau des cellules absorbantes. Mais nous n'avons pas constatt ici de rtsorption des parois permettant un passage direct entre la cellule du parasite et la trachtide de l'h6te. Existe-t-il d'autres voies pour le passage des sub- stances? Le phtnomkne de ptnCtration ttant rare, les substances ont certainement a traverser en gtntral la paroi des tltments conducteurs puis celle des cellules du systkme endo- phytique souvent trks tpaisses. Nous n'avons en effet jamais observt de plasmodesmes entre les cellules de l'hbte et celles du parasite, contrairement a ce que Tainter (1971) a avanct au sujet de 1'Arceuthobium pusillum Peck. Les tchanges sont en tous cas certainement trks importants au niveau du plas- malemme des cellules des su~oirs comme en ttmoignent les fortes activitts phosphatasique et ATP-asique que nous y avons enregistrtes. On sait en effet que les phosphatases acides sont impliqukes dans les mkcanismes de transports actifs trans-membranaires (Ziegler 1965; Sauter 1966) et notamment que ces enzymes sont toujours prtsentes dans les cellules oh transitent les glucides (Sauter et Braun 1968; Sauter 1972; Figier 1972; Sauter et Braun 1972; Caussin et al. 1979; Cron- shaw 1980). I1 a t t t tgalement montrt qu'elles interviennent dans les processus de difftrenciation des tltments conducteurs (Mia et Pathak 1968; Hebant 1973). Chez les plantes parasites, Rodriguez et Pannier (1967) avaient dtja tmis l'hypothkse que les phosphatases y jouaient un r61e important dans le transport actif de l'eau et des substances dissoutes en particulier des glucides. Les travaux de Renaudin (1977) sur le s u ~ o i r de L. clandestina vont dans le mCme sens en montrant la prtsence d'une forte activitt phosphatasique acide tout le long du pont- transfert reliant les tltments conducteurs de l'h6te B ceux du parasite. En outre, Tripodi (1970) et Onofeghara (1972) ont avanct que les phosphatases acides intervenaient dans la cyto- lyse des cellules de l'hbte, la difftrenciation et la maturation des cellules du parasite et dans le transfert d'tnergie. Dans le mCme ordre d'idte, Toth et Kuijt (1977) pensent que chez Cornandra les phosphatases, en association avec d'autres enzymes, agissent en ramollissant les parois des cellules de l'h6te favorisant ainsi l'action mtcanique du parasite. Chez Striga hermonthica (Del.) Benth. et Striga gesnerioides (Willd.) Watke., Ba (1983) estime tgalement que ces enzymes, trks actives dans la zone de contact h6te-parasite, sont impliquCes dans les mtcanismes de ptnttration dans l'h6te ainsi que dans le transport des substances de l'h6te vers le parasite.
En ce qui concerne les ATP-ases, leur localisation au niveau des plasmalemmes leur confere un rble central dans le trans- port des ions (Leigh et al. 1975; Leonard et van der Woude 1976) et dans les processus de cotransport des ions et des
glucides (Caussin et al. 1979; Komor et Tanner 1980; Georgieva 1980). Plus prtcisCment, Korenbrot (1977) a montrt que la moltcule d'ATPase (phosphatase neutre) com- prend deux sites fonctionnels, l'un ttant engagt dans la syn- thkse de l'ATP, l'autre dans le transport d'ions B travers 1 'apoplaste.
Chez Arceuthobium oxycedri, nos observations ont montrt que les activitts phosphatasiques acides et ATP-asiques ttaient importantes dans les cellules des su~oirs , au niveau de leur plasmalernme, sans &tre particulikrement intenses B la pointe de ces su~oirs , c'est-2-dire dans la zone de ptnttration. Par constquent, ces enzymes semblent impliqutes ici surtout dans les phtnomknes de transfert. En outre, il faut souligner que nous avons tgalement mis en tvidence une forte activitt phos- phatasique acide dans les cellules de 17h6te entourant le systkme endophytique, toujours au niveau des plasmalemmes; c'est le cas principalement des cellules du phlokme secondaire. On peut donc se demander si le transfert des substances se fait uniquement au niveau du xylkme de l'hbte ou tgalement au niveau du phlokme et si dans ce cas un passage est possible au niveau des cordons. Par ailleurs. les trachtides de l'h6te Ctant parfois au contact immtdiat des trachtides du parasite dans les su~oirs secondaires et la partie profonde des su~oirs primaires, des tchanges de substances organiques ont-ils lieu directement entre ces trachtides ou bien y a-t-il obligatoirement transport actif par l'intermtdiaire de cellules de type cellulosique? Une ttude autoradiographique, en utilisant en particulier des sub- stances migrant sptcifiquement par le phlokme ou le xylkme de 17h6te permettra sans doute d'Cclairer au moins une partie de ces problkmes.
BA, A. T. 1983. Biologie du parasitisme chez deux ScrophulariacCes tropicales, Striga hermonthica (Del.) Benth. et Striga gesnerioides (Willd.) Vatke. Thkse de doctorat 5s sciences naturelles, UniversitC de Dakar, Dakar, SCnCgal.
CAUSSIN, C., J. P. DESPEGHEL, M. FAUCHER, A. LEGER et J. I. BONNEMAIN. 1979. Etude du mkcanisme des Cchanges entre le gamCtophyte et le sporophyte chez les Bryophytes. C. R. Seances Acad. Sci. Ser. D, 289 : 1329-1334.
COULOMB, P. 197 1. Phytolysosomes dans le mCristkme radiculaire de la Courge (Cucurbita pep0 L., CucurbitacCes). ActivitC phospha- tasique acide et activitC peroxydasique. C. R. Seances Acad. Sci. SCr. D, 272 : 48-51.
CRONSHAW, J. 1980. Histochemical localization of enzymes in the phloem. Ber. Dtsch. Bot. Ges. 93 : 141-152.
FIGIER, J. 1972. Localisation infrastmcturale de la phosphatase acide dans les glandes petiolaires d'lmpatiens holstii. RSles possibles de cette enzyme au cours des processus sCcrCtoires. Planta, 108 : 215- 226.
GEORGIEVA, I. D. 1980. Histochemical localization of the adenosine triphosphatase (ATP-ase) and glucose-6-phosphatase during embryogenesis of Lillium regale Wils. Dokl. Akad. Nauk, ISSN 0366-8681, 33 : 117-120.
GOMORI, G. 1956. Histochemical methods for acid phosphatases. J. Histochem. Cytochem. 4 : 453-461.
HEBANT, C. 1973. Acid phosphomonoesterase activities (P-glycero- phosphatase and naphthol AS-MX phosphatase) in conducting tissues of bryophytes. Protoplasma, 77 : 231 -241.
KOMOR, E., et W. TANNER. 1980. Proton-cotransport of sugars in plants. Duns Plant membrane transport: current conceptual issues. Editeurs: R. M. Spanswick, W. T. Lucas et J. Dainty. ElsevierINorth-Holland Biomedical Press, Amsterdam. pp. 247-257.
KORENBROT, J. I. 1977. Ion transport in membranes: incorporation of biological ion-translocating proteins in model membrane systems. Annu. Rev. Physiol. 39 : 19-49.
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y G
LA
SGO
W U
NIV
ER
SIT
Y L
IBR
AR
Y o
n 09
/03/
13Fo
r pe
rson
al u
se o
nly.

2784 CAN. I. BOT. \
LEIGH, R. A., F. A. WILLIAMSON et R. G. WYN JONES. 1975. Pres- ence of two different membrane-bound, KC1-stimulated adenosine triphosphatase activities in maize roots. Plant Physiol. 55 : 678-685.
LEONARD, R. T., et W. J. VAN DER WOUDE. 1976. Isolation of plasma membranes from corn roots by sucrose density gradient centrifugation. An anomalous effect of ficoll. Plant Physiol. 57 : 105 - 114.
LUFT, J. H. 1961. Improvement in epoxy resin embedding methods. J. Biophys. Biochem. Cytol. 9 : 409-414.
MARX, C., J. DEXHEIMER, V. GIANINAZZI-PEARSON et S. GIANIN- AZZI. 1982. Enzymatic studies on the metabolism of vesicu- lar-arbuscular mycorrhizas. IV. Ultracytoenzymological evidence (ATPase) for active transfer processes in the host-arbuscule inter- face. New Phytol. 90 : 37-43.
MIA, A. J., et S. M. PATHAK. 1968. A histochemical study of the shoot apical meristem of Rauwolfia with reference to differentiation of sclereids. Can. J. Bot. 46 : 115 - 120.
ONOFEGHARA, F. A. 1972. Histochemical localization of enzymes in Topinanthus bangwensis: acid phosphatase. Am. J. Bot. 59 : 549-556.
RENAUDIN, S. 1974. Contribution i 1'Ctude de la biologie des phanCrogames parasites : recherches sur Lathraea clandestina L. (ScrophulariacCes). Thbse de doctorat bs sciences Naturelles, Uni- versitC de Nantes, Nantes.
1977. Mise en Cvidence d'activitCs enzymatiques au niveau des suqoirs de Lathraea clandestina L. Bull. Soc. Bot. Fr. 124 : 419-425.
RENAUDIN, S., et M. CAPDEFQN. 1979. Sur quejques aspects de l'ultrastructure des suqoirs de Touia alpina L. I. Etude des cellules absorbantes. Bull. Soc. Bot. Fr. 126 : 165-178.
REYNOLDS, E. S. 1963. The use of lead citrate at high pH as an elec- tron opaque stain in electron microscopy. J. Cell Biol. 17 : 208-212.
RODRIGUEZ, M. P., et P. PANNIER. 1967. ~ t u d e de la distribution de la phosphatase acide dans l'haustorium primaire de Phthirusa pyri-
folia (H.B.K.) Eichl. (Loranthaceae). Rev. Gen. Bot. 74 : 625 - 635.
SADIK, A., L. REY et S. RENAUDIN. 1986. Le systbme endophytique d'drceuthobium oxycedri. I. Organisation, Ctude cytologique et cytochimique, Can. J. Bot. 64 : 1104- 111 1.
SALLB, G. 1977. Etude cytologique, cytochimique et histoautoradio- graphique de Viscum album L. (LoranthacCes). U. Thbse de doctorat bs sciences naturelles, UniversitC de Paris VI, Paris.
SAUTER, J. J. 1966. Untersuchungen zur Physiologie der Pappelholz- strahlen. II. Jahresperiodische Anderungen der Phosphataseakti- vitit im Holzstrahlparenchym und ihre mogliche Bedeutung fiir den Kohlenhydratstoffwechsel und den aktiven Assimilattransport. Z. Pflanzenphysiol. 55 : 349 -362.
1972. Respiratory and phosphatase activities in contact cells of wood rays and their possible role in sugar secretion. Z. Pflanzen- physiol. 67 : 135-145.
SAUTER, J. J., et H. J. BRAUN. 1968. Enzymatic polarity in ray parenchyma cells of conifers in spring. ~ . - ~ f l a n z e n ~ h ~ s i o l . 58 -: 378-381.
1972. Cytochemische Untersuchung der Atmungsaktivitit in den Strasburger-Zellen von Lark und ihre Bedeutung fiir den Assimilattransport. Z. Pflanzenphysiol. 66 : 440-458.
TAINTER, F. H. 1971. The ultrastructure of Arceuthobium pusillum. Can. J . Bot. 49 : 1615 - 1622.
TOTH, R., et J. KUIJT. 1977. Cytochemical localization of acid phos- phatase in endophyte cells of the semiparasitic angiosperm Coman- dra umbellata (Santalaceae). Can. J. Bot. 55 : 470-475.
TRIFQDI, G. 1970. Localization of tryptophan rich proteins and gly- cerophosphatase activity in Cuscuta haustorial cells. Protoplasma, 71 : 191-196.
WACHSTEIN, M., et E. MEISEL. 1957. Histochemistry of the hepatic phosphatases at a physiologic pH. Am. J. Clin. Pathol. 27 : 13-23.
ZIEGLER, H. 1965. Die Physiologie pflanzlicher Driisen. Ber. Dtsch. Bot. Ges. 78 : 466-477.
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y G
LA
SGO
W U
NIV
ER
SIT
Y L
IBR
AR
Y o
n 09
/03/
13Fo
r pe
rson
al u
se o
nly.





























