Embed Size (px)
DESCRIPTION
Â
Citation preview

RECHERCHEAGRONOMIQUESUISSE
J a n v i e r 2 0 1 4 | N u m é r o 1
Ag
rosc
op
e |
OFA
G |
HA
FL |
AG
RID
EA
| E
TH Z
üri
ch
Agroscope
une bonne alimentation, un environnement sain
Production végétale Série ProfiCrops: Cinq idées qui ont changé la recherche en production végétale Page 4
Production végétale La diversité génétique dans l'agriculture Page 12
Eclairage Poulets de chair: la taille des particules de l'aliment influence les accroissements Page 28

ImpressumRecherche Agronomique Suisse / Agrarforschung Schweiz est une publication des stations de recherche agronomique Agroscope et de leurs partenaires. Cette publication paraît en allemand et en français. Elle s’adresse aux scientifiques, spécialistes de la recherche et de l’industrie, enseignants, organisations de conseil et de vulgarisation, offices cantonaux et fédéraux, praticiens, politiciens et autres personnes intéressées.
EditeurAgroscope
Partenairesb Agroscope (Institut des sciences en production végétale IPV;
Institut des sciences en production animale IPA; Institut des sciences en denrées alimentaires IDA; Institut des sciences en durabilité agronomique IDU), www.agroscope.ch
b Office fédéral de l’agriculture OFAG, Berne, www.ofag.chb Haute école des sciences agronomiques forestières et alimentaires HAFL, Zollikofen, www.hafl.chb Centrale de vulgarisation AGRIDEA, Lausanne et Lindau, www.agridea.chb Ecole polytechnique fédérale de Zurich ETH Zürich,
Département des Sciences des Systèmes de l'Environnement, www.usys.ethz.ch
Rédaction Andrea Leuenberger-Minger, Recherche Agronomique Suisse /Agrarforschung Schweiz, Agroscope, Case postale 64, 1725 Posieux, Tél. +41 26 407 72 21, Fax +41 26 407 73 00, e-mail: [email protected]
Judith Auer, Recherche Agronomique Suisse / Agrarforschung Schweiz, Agroscope, Case postale 1012, 1260 Nyon 1 e-mail: [email protected]
Team de rédaction Président: Jean-Philippe Mayor (Responsable Corporate Communication Agroscope), Evelyne Fasnacht, Erika Meili et Sibylle Willi (Agroscope), Karin Bovigny-Ackermann (OFAG), Beat Huber-Eicher (HAFL), Esther Weiss (AGRIDEA), Brigitte Dorn (ETH Zürich).
AbonnementsTarifsRevue: CHF 61.–*, TVA et frais de port compris(étranger + CHF 20.– frais de port), en ligne: CHF 61.–** Tarifs réduits voir: www.rechercheagronomiquesuisse.ch
AdresseNicole Boschung, Recherche Agronomique Suisse/Agrarforschung Schweiz, Agroscope, Case postale 64, 1725 Posieux e-mail: [email protected], Fax +41 26 407 73 00
Changement d'adressee-mail: [email protected], Fax +41 31 325 50 58
Internet www.rechercheagronomiquesuisse.chwww.agrarforschungschweiz.ch
ISSN infosISSN 1663 – 7917 (imprimé)ISSN 1663 – 7925 (en ligne)Titre: Recherche Agronomique SuisseTitre abrégé: Rech. Agron. Suisse
© Copyright Agroscope. Tous droits de reproduction et de traduction réservés. Toute reproduction ou traduction, partielle ou intégrale, doit faire l’objet d’un accord avec la rédaction.
Indexé: Web of Science, CAB Abstracts, AGRIS
SommaireJanvier 2014 | Numéro 1
Depuis le 1er janvier 2014, les trois stations de recherche ACW, ALP-Haras et ART sont réunies sous une même bannière, celle d’Agroscope. Le nouveau mandat de prestations d’Agroscope (2014 -2017) englobe six pôles thématiques qui seront traités en commun par par les quatre instituts d’Agroscope. Vu la pénurie annoncée des ressources naturelles, la mission de la recherche agroalimentaire consistera désormais à trouver des solutions innovatrices. (Photo: Gabriela Brändle, Agroscope)
3 Editorial
Production végétale – Série ProfiCrops
4 Cinq idées qui ont changé la recherche en production végétale Anna Crole-Rees, Vincent Nassar, Arnold Schori,
Willy Kessler et Bernard Jeangros
Production végétale
12 La diversité génétique dans l'agriculture Roland Kölliker, Luisa Last, Felix Herzog et
Franco Widmer
Environnement
20 Effets de mélanges de semences sur la diversité génétique et la performance
Tsipe Aavik, Daniel Bosshard, Peter Edwards,
Rolf Holderegger et Regula Billeter
Eclairage
28 Poulets de chair: la taille des particules de l'aliment influence les accroissements
Danielle Albiker et Ruedi Zweifel
Eclairage
32 Réseau agri benchmark: comparaison dans le contexte international Hildegard Garming et Esther Bravin
36 Portrait
37 Actualités
39 Manifestations

Editorial
3Recherche Agronomique Suisse 5 (1): 3, 2014
Chère lectrice, cher lecteur,
Au cours des prochaines décennies, le système alimentaire mondial devra
relever l’immense défi de fournir à une population toujours plus nombreuse
des aliments sains en suffisance et à un prix abordable, en dépit des ressources
limitées. D’après l’évolution démographique pronostiquée et le bien-être
croissant, l’ONU estime que la demande en denrées alimentaires augmen-
tera de 50 % d’ici à 2030. La demande en produits animaux devrait augmen-
ter encore davantage. La pénurie prévisible des ressources – en particulier
des ressources non renouvelables – poussera la population à changer ses
habitudes de consommation. La recherche agroalimentaire a désormais pour
tâche cruciale de trouver des solutions innovatrices afin de promouvoir une
utilisation parcimonieuse des ressources.
Dans le nouveau mandat de prestations 2014–2017, adopté dans son
intégralité par le Conseil fédéral en décembre 2013, Agroscope met en place
une stratégie pour relever ces défis. L’exécution du mandat de prestations
est du ressort du Conseil Agroscope, organe de conseil pour l’orientation
stratégique de la recherche d’Agroscope, présidé par Bernard Lehmann,
directeur de l’OFAG. Le mandat de prestations tient compte des demandes
émanant à la fois du Conseil fédéral, du Parlement et des principales parties
prenantes d’Agroscope. L’examen du mandat de prestations par les Commis-
sions des finances et les Commissions de l’économie et des redevances de la
Confédération a montré que son orientation générale répond à ces
demandes.
Le mandat de prestations définit les six pôles thématiques qui oriente-
ront à l'avenir les travaux de recherche des quatre nouveaux instituts
d’Agroscope:
•• Intensification écologique
•• Préservation des ressources naturelles
•• Contribution à la protection du climat et adaptation de l’économie
agroalimentaire au changement climatique
•• Qualité et sûreté des denrées alimentaire pour une alimentation saine
•• Amélioration de la compétitivité de l’économie agroalimentaire
•• Vitalité et attractivité de l’espace rural
Les thèmes de chaque pôle seront traités par plusieurs instituts, ce qui ne sera
possible qu’avec une utilisation conséquente des synergies et en se fondant sur
des valeurs communes. Grâce à nos valeurs fondamentales, telles que la
confiance, la responsabilité propre et la fiabilité, nous parviendrons à réaliser
ces objectifs ambitieux, à renforcer notre visibilité et à remplir notre mandat.
Pour mettre en œuvre ce mandat de prestations, le comité de direction
d’Agroscope doit employer de façon optimale les ressources financières et en
personnel. Il s’agit donc non seulement de prendre des décisions stratégiques
adroites en matière de personnel, mais aussi de développer des compétences
de manière ciblée et d’assurer la flexibilité des ressources afin de s’adapter aux
circonstances. En procédant ainsi, Agroscope pourra fournir à la pratique, aux
politiques et aux services chargés de l’exécution des tâches légales de la
Confédération des solutions actuelles et adaptées.
Nous sommes convaincus que nous parviendrons à apporter des réponses
aux défis futurs de l’économie agroalimentaire.
Michael Gysi, chef AgroscopeBernard Lehmann, directeur de l’Office fédéral de l’agriculture OFAG
L’économie agroalimentaire devant des défis de taille

4
Recherche Agronomique Suisse 5 (1): 4–11, 2014
I n t r o d u c t i o n
L’agriculture en général et la production végétale en
particulier font face à de sérieux défis. La libéralisation
des marchés se poursuit et induit une concurrence
accrue de l’étranger. Certains marchés sont saturés. Les
exigences environnementales et sociales quant aux pro-
duits et aux modes de production augmentent. La
vitesse de ces changements a aussi fortement aug-
menté, mue entre autres par les progrès technolo-
giques. La survie des exploitations agricoles dépend
donc de leur capacité à suivre ces changements, à
s’adapter et donc à innover. L’innovation est une idée
qui est réalisée et adoptée. Elle est au centre de tout
processus de changement.
Les travaux du Module Innovation du programme
de recherche ProfiCrops se sont donc centrés sur la
valorisation des produits, des méthodes et des services
développés par Agroscope pour ses clients dans le
domaine de la production végétale. L’objectif de ce
module est de renforcer le processus d’innovation afin
de permettre la production de solutions encore meil-
leures pour la pratique et de renforcer leur taux
d’adoption par les différents utilisateurs.
Les chercheurs se doivent de constamment innover pour faire face aux différents défis et aux nombreux chan-gements des conditions-cadres, économiques, législatives, etc. (Source: http://www.johnthemachine.com/tag/business-innovation)
Anna Crole-Rees1, Vincent Nassar3, Arnold Schori1, Willy Kessler2 et Bernard Jeangros1
1Agroscope, Institut des sciences en production végétale IDP, Suisse,2Agroscope, Institut des sciences en durabilité agronomique IDU, Suisse 3Institut pour l’Entrepreneurship & Management, HES-SO, 3960 Sierre, Suisse
Renseignements: Anna Crole-Rees, [email protected]
Cinq idées qui ont changé la recherche en production végétale
Série ProfiCrops
P r o d u c t i o n v é g é t a l e

Cinq idées qui ont changé la recherche en production végétale | Production végétale
5
Recherche Agronomique Suisse 5 (1): 4–11, 2014
Innover est une condition nécessaire pour les
institutions qui cherchent à maintenir leur
compétitivité dans une économie de plus en
plus libéralisée. Ceci est aussi vrai pour la
recherche agronomique. Un des objectifs de
ProfiCrops, un des programme de recherche
Agroscope, était de promouvoir un processus
d’innovation menant à une plus grande
valeur ajoutée au sein du secteur de produc-
tion végétale. Cet article décrit cinq idées
sélectionnées de manière raisonnée au sein
d’Agroscope. L’idée de base, sa mise en
œuvre et les perspectives de son adoption
sont présentées pour trois innovations de
type processus: l’appareil NIRS (Near-infrared
spectroscopy) portable, le séquençage du
pathogène du feu bactérien et l’utilisation
des marqueurs biochimiques ou moléculaires,
et deux innovations de type produit: l’ana-
lyse du cycle de vie (LCA) et l’agriculture
urbaine. Les résultats montrent que le
processus de l’innovation au sein de la
recherche exige un rayon d’action compre-
nant une mission de recherche claire, des
ressources financières en conséquence, du
temps et une attitude positive face au risque.
Rés
um
é
ProfiCrops
Le programme de recherche Agroscope Profi-
Crops (www.proficrops.ch) a pour objectif de
contribuer à garantir la compétitivité de la
production végétale suisse dans un cadre de
plus en plus libéralisé, et de renforcer la
confiance des consommateurs envers les pro-
duits suisses. Les hypothèses posées en début
de programme stipulaient que l’efficience de
la production devait être améliorée, l’inno-
vation et la valeur ajoutée augmentées, la
confiance des consommateurs renforcée et
les conditions-cadres modifiées. Ces quatre
aspects ont fait l’objet de recherches interdis-
ciplinaires, sous forme de modules: Efficience,
Innovation, Consommateurs et Conditions
cadres, et de projets intégrés et associés: Feu
Bactérien, ProfiVar, ProfiGemüse CH, Coopé-
ration d’assolement, ProfiViti, WIN4 et FUI.
La série d’articles «ProfiCrops» publiée dans
Recherche Agronomique Suisse permet de dif-
fuser une sélection de résultats et de solutions
pour le maintien de la compétitivité de la pro-
duction végétale en Suisse. Ces résultats et
solutions sont exemplaires. Un rapport de syn-
thèse sera disponible début 2014.
L’article «Cinq innovations qui ont changé la
recherche en production végétale», lié au
module Innovation*, présente des innova-
tions, développées ou testées, dans tous les
cas adoptées par les chercheurs d’Agroscope.
Il met en exergue la complexité du processus
de l’innovation.
*( http://www.agroscope.admin.ch/proficrops/05365/index.html?lang=fr)
Les études menées jusqu’à présent visaient surtout à
mieux comprendre l’innovation au niveau des produc-
teurs agricoles en Suisse (Hermier et al. 2006), du sys-
tème de connaissances en agriculture en général (Her-
mans et al. 2010) et de la caractérisation des innovations
(Aouinaït 2013). Plus rares sont les projets qui s’inté-
ressent au processus de l’innovation au sein même des
organismes de recherche en agriculture. Et pourtant, les
chercheurs innovent aussi. Ils font aussi face à des défis
qui les poussent à créer de nouvelles idées adaptées à
leurs besoins (Nassar et Tucci 2012).
Cet article s’attache à décrire le processus d’innova-
tion au travers d’études de cas, afin de mieux identifier
certains facteurs d’innovation au sein de la recherche et
ainsi de renforcer ce processus. Cette étude se concentre
sur les travaux au sein d’Agroscope.
Sélection de cinq études de cas
Les chercheurs impliqués dans le Module Innovation ont
élaboré une liste de «solutions» développées par
Agroscope pour les acteurs de la production végétale.
Plusieurs produits ou méthodes se sont révélés être des

Production végétale | Cinq idées qui ont changé la recherche en production végétale
6 Recherche Agronomique Suisse 5 (1): 4–11, 2014
innovations adoptées par les chercheurs eux-mêmes,
parfois aussi développées par eux. Les nombreuses initia-
tives novatrices permettent aux chercheurs, et donc à
Agroscope:
•• d'offrir de nouveaux produits aux clients (innovation
type produit): colza HOLL, variétés de soja pour le tofu
en Europe;
•• de mieux s’organiser ou s’adapter à certaines exi-
gences (innovation type organisation): système de
management de la qualité, fusion des stations de
recherche Agroscope;
•• d’améliorer sa manière de diffuser les résultats de ses
recherches (innovation type commercialisation):
application Smartphone (p. ex. PhytoPre), spin-off
s’occupant de la commercialisation (MediaPlant,
VariComm) et/ou
•• de renforcer l’efficience de la recherche (innovation
type processus): méthode de floraison accélérée
(sélection de pommiers), phénotypage pour sélection
des céréales, logiciels GenieII et SustainOS, etc.
Cinq études de cas ont été sélectionnées par un échantil-
lonnage raisonné (station à l’origine de la recherche,
type d’innovation), en tenant compte de l’appréciation
des chercheurs impliqués dans le Module Innovation.
Une recherche bibliographique a été menée, puis les
chercheurs ayant adopté un nouveau produit ou une
nouvelle méthode dans le cadre de leur recherche ont
été contactés afin d’identifier les motivations ayant
mené à l’innovation.
1. L’appareil NIRS portable (fig. 1)Agroscope a pour mandat de travailler en pré-récolte sur
les aspects de la qualité en arboriculture et petits fruits.
Les aspects de la qualité, leur définition et leur contrôle
sont devenus indispensables pour le maintien de la com-
pétitivité du secteur. La détermination de la date de
récolte idéale pour une qualité optimale des fruits sur le
marché requiert des analyses chimiques et de prélever des
fruits qui seront détruits par ces analyses. Il n’est donc pas
possible de suivre l’évolution de la maturité des mêmes
fruits depuis l’arbre jusqu’à sa récolte.
Il existe un outil permettant des analyses non-inva-
sives, appelé communément NIRS. Cet appareil utilise la
spectroscopie du rayonnement proche de l’infrarouge
(NIRS: Near-infrared spectroscopy). Il est déjà utilisé à
Agroscope dans le cadre de la recherche à différents
stades de la chaîne de valeur (production, récolte, stoc-
kage), pour différents produits (miel, viande, lait, blé,
fruits, fourrages) et pour de plus en plus de critères
(matière soluble, acidité, matière sèche, teneur en lyco-
pènes, taux de protéines, valeurs de digestibilité, nutri-
ments). Cet appareil s’utilise en laboratoire et est fixe.
L’idée est d’en développer une version qui soit portable
et qui puisse être utilisée dans les parcelles et les serres.
Des contacts ont été pris avec un chercheur à Angers qui
travaillait sur la qualité des pommes. Grâce à des fonds
tiers, ce chercheur a pu être engagé par Agroscope en
2006. Actuellement, les travaux de calibration de cet
appareil portable sont en cours (Camps et Christen 2009).
Figure 1 | Analyse de la qualité, de l’arbre à la récolte.
Innovation Problème-motivation Type innovation
1 Appareil NIRS portable pour analyse de fruitsAnalyse de la qualité de fruits sur l’arbre
jusqu’à leur récolteProcessus
2 Analyse de cycle de vie (LCA)Volonté politique de développer une
agriculture plus écologiqueProduit
3 Séquençage de la bactérie Erwinia amylovoraRecherche d’un moyen de lutte curatif contre le feu
bactérienProcessus
4 Agriculture urbaineDemande externe pour les compétences
agronomiquesProduit
5 Marqueurs biochimiques ou moléculairesAnalyse de caractères génétiques,
indépendamment de l’environnementProcessus
Tableau 1 | Etudes de cas sélectionnées

Cinq idées qui ont changé la recherche en production végétale | Production végétale
7Recherche Agronomique Suisse 5 (1): 4–11, 2014
2009). Cette méthode fournit des données fiables concer-
nant l’impact des produits sur l’environnement, ceci à
l’échelle d’un produit, d’un système de production et/ou
d’une exploitation agricole. Elle permet aussi de compa-
rer des modes de production, des origines de produits
différentes et de calculer des indicateurs agro-environ-
nementaux. Elle est actuellement constituée d’une base
de données des inventaires environnementaux agricoles,
ecoinvent. Elle offre:
•• des modèles de calcul des émissions directes au champ
et à la ferme, comme les nitrates ou les métaux lourds;
•• des méthodes d’évaluation de l’impact environnemen-
tal, sur la biodiversité et la qualité du sol;
•• des outils de calcul pour des différents systèmes
agricoles, notamment pour des exploitations agricoles
et des cultures;
•• un schéma d’interprétation pour les ACV en agriculture.
Les demandes pour ce service proviennent des décideurs
du secteur privé (commerce, industrie agro-alimentaire)
et public (suisses et étrangers), des organisations de pro-
ducteurs, du conseil agricole et d’ONGs.
Les perspectives de ce service pour les prochaines années
vont certainement augmenter avec, notamment, la
demande de solutions pour une utilisation durable des
ressources dans le secteur agro-alimentaire et l’éco-
conception des systèmes de production agro-alimen-
taires. Il est prévu aussi d’appliquer l’ACV dans des nou-
veaux domaines comme les cultures spéciales ou la
production d’aliments.
Les résultats attendus et la portée d’un NIRS portable cali-
bré pour les fruits sont importants. Cet appareil donnera
la possibilité d’améliorer la recherche sur la qualité pré-
récolte, particulièrement pour déterminer la maturité
optimale et pour étudier l’impact des facteurs climatiques.
Il permettra aussi d’augmenter l’efficience de la recherche,
car cette dernière ne sera plus limitée par la taille de
l’échantillon (les fruits ne seront plus détruits), sera plus
rapide (quelques secondes pour une analyse), exigera
moins de main d’œuvre et aucun produit chimique. De
plus, l’utilisation de cet équipement est relativement
simple et permettra une adoption aisée par les chercheurs.
A terme, il pourrait être utilisé par tous les acteurs de la
chaîne de valeur, ce qui permettrait de suivre l’évolution
de la qualité d’un fruit tout au long de la chaîne.
2. L’analyse de cycle de vie (fig. 2)
L’agriculture s’écologise. Cette tendance s’applique à
l’ensemble de la chaîne de valeur et aux produits finis,
jusqu’aux consommateurs. Elle est demandée par la
société, les citoyens, les consommateurs ainsi que par les
autorités qui veulent formuler des stratégies politiques
en adéquation avec les attentes. La question est de savoir
comment mesurer les impacts de la production, voire de
l’ensemble des processus de la chaîne de valeur, sur les
émissions de CO2, la biodiversité, la qualité du sol, etc.?
La direction d’Agroscope a décidé de créer un nouveau
groupe de recherche «Analyse de cycle de vie» (ACV) en
2000. Ce dernier a développé une méthode d’ACV adap-
tée à l’économie agroalimentaire, appelée SALCA (Swiss
Agricultural Life Cycle Assessment) (Gaillard & Nemecek
Figure 2 | Pour une agriculture plus écologique. (Photo: Carole Parodi, Agroscope)

Production végétale | Cinq idées qui ont changé la recherche en production végétale
8 Recherche Agronomique Suisse 5 (1): 4–11, 2014
3. Le séquençage de la bactérie responsable du feu
bactérien (fig. 3)
Le feu bactérien est une maladie très grave pour les
pommiers et les poiriers notamment. Bien que la mala-
die et son agent pathogène, la bactérie Erwinia amylo-
vora, soient connus depuis plus de 100 ans, il n’existe
toujours pas de traitement curatif autre que l’utilisation
d’antibiotiques pendant la floraison. Le centre de com-
pétences sur le feu bactérien à Agroscope travaille sur
les moyens de maîtriser cette maladie. Les recherches
ont essentiellement porté sur les méthodes de lutte pro-
phylactiques, comme la sélection de variétés robustes,
l’analyse de la sensibilité des anciennes variétés, le déve-
loppement d’outils pour la pratique tels que le test de
diagnostic rapide permettant de déterminer la présence
de l’agent pathogène en champs, la prévision de l’infec-
tion florale (www.feubacterien.ch) et le test d’antago-
nistes en laboratoire et en plein champ.
L’un des chercheurs d’Agroscope a formulé l’idée de
changer d’approche et de tenter de mieux connaître
l’agent pathogène pour rechercher son talon d’Achille,
et ainsi de mieux le contrôler. Cette idée a pu être mise
en oeuvre dans le cadre du projet intégré Feu bactérien,
grâce à des moyens financiers supplémentaires impor-
tants apportés entre 2008 et 2013, et sa concrétisation
se poursuit aujourd’hui encore. Une étape marquante a
été le séquençage de la bactérie responsable de cette
maladie en 2010 (Smits et al. 2010). Le génome a été
complètement décrypté.
Depuis le séquençage, plusieurs gènes pouvant être déci-
sifs pour la survie et la virulence de la bactérie ont été
découverts (Smits et al. 2010). Les perspectives de lutte
contre le feu bactérien s’améliorent. Par exemple, l’ana-
lyse des différents isolats a permis de mieux comprendre
la contamination locale. Toutefois, il reste encore beau-
coup de travail. Si les gènes ont été séquencés, il s’agit
maintenant d’identifier leur fonction, opération très
complexe, ainsi que d’analyser les mutations d’ADN selon
l’environnement. Ces connaissances permettront de com-
prendre comment la bactérie fonctionne et où se situe
son talon d’Achille. Ce n’est pas avant plusieurs années
que les effets seront notables sur le terrain. Le séquen-
çage de la bactérie pourrait permettre, à terme, de gérer
le feu bactérien. Une condition est l’utilisation optimale
des moyens de lutte actuels comme la mise en culture
d’arbres tolérants - entre autres solutions.
4. L’agriculture urbaine (fig. 4)
Il existe un intérêt grandissant pour l’agriculture urbaine.
De plus en plus d’initiatives de production alimentaire
en ville sont réalisées. La Suisse n’échappe pas à ce phé-
nomène. Dans le cadre du projet «Qualité urbaine» du
Fonds national pour la recherche suisse (FNRS), un orga-
nisme a souhaité soumettre un projet intégrant l’agricul-
ture urbaine et a contacté différentes institutions, dont
l’Institut des décisions environnementales de l’Ecole
polytechnique fédérale de Zurich et Agroscope. Ce pro-
jet, Food Urbanism Initiative (www.foodurbanism.org),
a été accepté pour une durée de trois ans et la partie
d’Agroscope attribuée à ProfiCrops.
L’origine de l’idée est donc externe. Sa réalisation,
soit la mise à disposition des compétences d’Agroscope
au service des promoteurs de l’agriculture urbaine, a
requis le recrutement de ressources humaines financées
par le FNRS. Elle a aussi exigé un «changement» de point
de vue sur les entités productrices. En effet, les unités de
production en zones urbaines ne sont pas (ou rarement)
des exploitations agricoles «traditionnelles». Il a fallu
ainsi définir, d’une façon reconnue par tous les parte-
naires du projet, une typologie des unités de production
(Crole-Rees et al. 2012).La participation à ce projet a permis d’intégrer les com-
pétences d’Agroscope en production sous serres et en pro-
duction maraîchère. Elle a donné l’occasion de travailler
sur un mandat pour des serres installées sur les toits. Elle a
aussi été une opportunité d’offrir une visibilité auprès
d’un public différent des clients usuels de la recherche en
production végétale. L’agriculture urbaine au sein
d’Agroscope, en tant que service pour les clients, va rester
marginale et son action sporadique. Elle ne fait actuelle-
ment pas l’objet du contrat de prestations de l’OFAG.
Figure 3 | Le séquençage de la bactérie Erwinia amylovora.
Erwinia amylovoraCFBP 1430
chromosome3805573 bp
0 kb
500 kb
1000 kb
1500 kb
2000 kb
2500 kb
3000 kb
3500 kb
pEA2928259 bP

Cinq idées qui ont changé la recherche en production végétale | Production végétale
9Recherche Agronomique Suisse 5 (1): 4–11, 2014
exemple pour évaluer la résistance de variétés de pom-
mier au feu bactérien. Une spécialiste a été engagée
dans le but de soutenir l’amélioration du blé, du triticale
et du soja à Changins.
Actuellement, cette technologie est utilisée à
Agroscope par tous les départements actifs dans la
sélection: la vigne (Agroscope, à paraître), les céréales,
le soja, le pommier, le poirier et l’abricotier. Les applica-
tions concrètes de cette technologie sont, par exemple,
de faciliter la création de variétés de soja destinées à
l’alimentation humaine et de céréales résistantes aux
maladie, le cumul de plusieurs gènes de résistance
contre une seule maladie à l’aide de marqueurs dans
diverses lignées de blé, de permettre l’introgression de
gènes de résistance dans des variétés élites de blé par
rétro-croisements et marqueurs moléculaires, etc.
(Mouillet et al. 2008). Cette technologie améliore l’effi-
cience et ses potentiels, comme par exemple l’utilisation
de plusieurs marqueurs et l’introduction de caractère
quantitatif. Elle permet aussi à Agroscope de rester un
leader reconnu dans le domaine de la sélection.
5. Les marqueurs biochimiques ou moléculaires (fig. 5)
En Suisse, la sélection du blé a pour objectif de créer des
variétés performantes, associant une très haute qualité
boulangère et un bon niveau de résistance aux maladies.
Le résultat des croisements est examiné sur des plantes
«adultes» au champ, c’est-à-dire sur l’expression de leurs
gènes dans un milieu donné. Cela exige du temps et est
parfois trop imprécis, par exemple lorsqu’une maladie
n’est pas présente. Pour connaître néanmoins la consti-
tution génétique du caractère étudié, indépendamment
de son expression dans un milieu donné, des marqueurs
biochimiques et moléculaires ont été développés. Ils per-
mettent de juger de la résistance aux maladies de la
plante, même si la maladie n’est pas déclarée.
Des scientifiques ont cherché à valoriser sur les connais-
sances individuelles des gènes pour les intégrer dans les
schémas de sélection. L’idée était d’appliquer ce saut
technologique au processus de sélection. Les marqueurs
ont l’avantage de ne pas être influencés par l’environne-
ment, d’être observables à n’importe quel stade de
développement de la plante et sur tous les organes, et
de pouvoir être analysés sans détruire la plante. Ils per-
mettent aussi de réduire les tests coûteux en serres, par
Figure 4 | Agriculture urbaine. (Photo: Therese Haller, HAFL)
Figure 5 | Utilisation de marqueurs biochimiques et moléculaires.(Photo: Carole Parodi, Agroscope)

10 Recherche Agronomique Suisse 5 (1): 4–11, 2014
Production végétale | Cinq idées qui ont changé la recherche en production végétale
D i s c u s s i o n s e t c o n c l u s i o n s
La recherche, comme tous les acteurs de la chaîne de
valeur en production végétale, se doit de constamment
innover pour faire face aux différents défis et aux nom-
breux changements des conditions-cadres, économiques,
législatives, etc.
Les études de cas mettent en évidence différents
aspects de l’innovation. La motivation peut être interne
ou externe, comme c’est le cas pour la mise sur le marché
des nouveaux produits ou services comme l'analyse de
cycle de vie et l’agriculture urbaine. Agroscope répond
donc à des demandes externes. Pour les trois procédés,
c’est l’envie de contourner les inconvénients de cer-
taines procédures, comme la lenteur ou l’imprécision de
la sélection traditionnelle pour certains caractères, qui
stimule les chercheurs et les pousse à adopter puis déve-
lopper de nouvelles idées (Nassar et Tucci 2012). La
recherche de solutions se fait rarement en vase clos. En
effet, plusieurs de ces innovations ont un caractère
transversal. Le séquençage de bactéries reprend une
approche qui avait été déjà mise en œuvre dans le sec-
teur de la santé, et l’analyse de cycle de vie provient de
l’industrie. Les échanges avec d’autres chercheurs au
travers de publication, conférences scientifiques, etc.
sont une source d’inspiration et de création. Les études
de cas montrent aussi qu’entre l’idée et l’innovation, il
s’écoule parfois plusieurs années, par exemple pour le
calibrage du NIRS.
Enfin, les idées présentées démontrent la capacité
d’innover au sein de la recherche. Elles ont permis ou
vont permettre de grandes améliorations dans la
recherche et pour les acteurs de la production végétale
en Suisse, particulièrement en matière d’efficience et
d’amélioration de la qualité des produits agricoles au
sens large. Les cinq innovations s’attèlent à la qualité
intrinsèque et extrinsèque (mode de production, impact
sur l’environnement) et à la différenciation des produits.
La capacité d’innover avec succès, c’est-à-dire la
capacité de passer de l’idée à l’innovation, exige du
temps, des ressources, une prise de risque et une vision
à long terme. Cette dernière implique de bonnes
connaissances des tendances dans les domaines spéci-
fiques des chercheurs et dans la politique agricole en
général et une définition claire de la vision et de la mis-
sion d’Agroscope. Le nouveau mandat de prestations en
tient compte. Pour poursuivre les objectifs stratégiques
d’Agroscope (OFAG 2012), «la recherche agricole doit
continuer à pouvoir disposer suffisamment de marge de
manœuvre pour que les bons chercheurs, intuitifs et
créatifs, puissent trouver des solutions pour le futur de
l’agriculture et l’alimentation» (Agroscope 2007). n
ProfiCropsProgrammes de recherche Agroscope

11Recherche Agronomique Suisse 5 (1): 4–11, 2014
Cinq idées qui ont changé la recherche en production végétale | Production végétale
Ria
ssu
nto
Sum
mar
y
▪ Gaillard G. & Nemecek T., 2009. Swiss Agricultural Life Cycle Assessment (SALCA): An integrated environmental assessment concept for agricul-ture. In: Int. Conf. «Integrated Assessment of Agriculture and Sustainable Development, Setting the Agenda for Science and Policy», Egmond aan Zee, The Netherlands. AgSAP Office, Wageningen University, 134–135.
▪ Hermier R., Praz P. & Buser Ch., 2006. Succès des innovations en agricul-ture: le projet InoVagri. Revue suisse d’Agriculture 38 (5), 275–279.
▪ Moullet O., Fossati D., Mascher F., Schori A. & Guadagnuolo R., 2008. Les marqueurs moléculaires comme outils dans la sélection des céréales. Revue suisse d’Agriculture 40 (3), 133–138.
▪ Nassar V. & Tucci Ch., 2012. Creative-Learning Innovation Cycle – CLIC: Work Motivation and Organizational Creativity. Thesis 5502. EPFL: Lausanne
▪ OFAG, 2012. Plan directeur de la recherche agronomique et agroalimen-taire 2013-2016. Berne. Février 2012.
▪ Smits Th., Rezzonico F., Kamber T., Blom J., Goesmann A., Frey J. E & Duffy B., 2010. Complete genome sequence of the fire blight pathogen Erwinia amylovora CFBP 1430 and comparison to other Erwinia spp. Mol Plant Microbe Interact. 2010 Apr 23 (4), 384–93.
Bibliographie ▪ Agroscope, 2007. Agroscope Research Master Plan 2008-2011. ▪ Agroscope, à paraître. Améliorer la compétitivité du secteur de la production végétale suisse. Résultats et expériences du programme de recherche ProfiCrops. Rapport final de programme.
▪ Aouinaït C., 2013. Caractérisation des innovations dans la production végétale suisse. Mémoire de fin d’études présenté pour le diplôme d’Ingénieur de spécialisation Innovations dans les Systèmes Agro- Alimentaires du Monde (ISAM). SupAgro Montpellier.
▪ Camps C. & Christen D., 2009. Non-destructive Assessment of Apricot Fruit Quality by Portable Visible-Near Infrared Spectroscopy (pVNIRs). LWT – Food Science and Technology 42 (6), 1125–1131.
▪ Crole-Rees A., Heitkämper K., Bertschinger L., Dumondel M., Haller Th. & Verzone C., 2012. Urban agriculture: an opportunity for farmers? A Swiss case study. Paper presented at the SHE conference, Angers, July 2012.
▪ Hermans F., Klerkx L. & Roep D., 2010. Comparative analysis and synthe-sis report. SOLINSA. FP7. Deliverables 3.1 A. Accès: http://www.solinsa.org/fileadmin/Files/deliverables/D3_1a_Comparative_analysis_and_syn-thesis_report_final_Nov_2011.pdf [15.9.2013].
Five ideas that have changed research in
the cropping sector
Innovation is now a prerequisite for
institutions aiming to maintain their
competitiveness in a more and more
liberalized economy. This is also true for
agricultural research. One of the objective
of ProfiCrops, the research program
Agroscope, was to promote the innova-
tion process leading to added value in the
cropping sector. This article describes five
ideas, their development into innovation
and the scope for the innovation’s
adoption. The sampling was done purpo-
sively, based on an innovations’ list for the
cropping sector. The sample comprised:
three process innovations: a portable
Near-infrared spectroscopy (NIRS) tool, the
sequence of the fire blight pathogen
genome and the use of molecular markers,
and two service innovations: Life Cycle
Assessment (LCA) in agriculture and urban
agriculture. The results show that the
innovation process within research
requires some scope that includes a clear
research mission, sufficient financial
resources, time and a risk-taking attitude.
Key words: innovation process, agricul-
ture, research, Agroscope.
Cinque idee che hanno cambiato la ricerca
nella produzione vegetale
L’innovazione è una condizione necessaria
per le istituzioni che cercano di mantenere
la loro competitività all’interno di un’eco-
nomia sempre più liberalizzata. Questo
vale anche per la ricerca agronomica. Uno
degli obiettivi di ProfiCrops, uno dei
programmi di ricerca di Agroscope, era di
promuovere un processo d’innovazione
aspirante a dare un valore aggiunto
maggiore al settore della produzione
vegetale. Questo articolo descrive cinque
idee selezionate in modo ragionato
all’interno di Agroscope. L’idea di base, la
sua attuazione e le prospettive d’adozione
sono presentate attraverso tre tipi di
procedimento innovativi: l’apparecchio
NIRS (Near-Infrared spectroscopy) porta-
tile, il sequenziamento del patogeno del
fuoco batterico e l’uso di marcatori
biochimici o molecolari nella selezione.
Inoltre, ci sono altri due prodotti innova-
tivi: l’analisi del ciclo vitale (LCA) e
l’agricoltura urbana. I risultati mostrano
che il successo di questi progetti innovativi
in seno alla ricerca esigono una missione
di ricerca chiara, delle risorse finanziarie
adatte, del tempo e un’attitudine positiva
di fronte al rischio.

12 Recherche Agronomique Suisse 5 (1): 12–19, 2014
tion des espèces et des populations face à des facteurs
externes comme les conditions environnementales ou les
processus de sélection. Bien que l’importance de la diver-
sité des espèces soit généralement reconnue et que l’in-
fluence qu’elle subit par l’agriculture ait été étudiée de
manière approfondie, on dispose de très peu d’informa-
tions sur la diversité génétique à l’intérieur des écosys-
tèmes agricoles. Le but de la présente étude consiste
donc à développer et appliquer des méthodes permet-
tant de cerner la diversité génétique au niveau des
exploitations agricoles. Cette étude a été réalisée dans le
I n t r o d u c t i o n
La fonctionnalité et la productivité des systèmes agri-
coles sont assez fortement influencées par la biodiver-
sité ambiante. Celle-ci peut être subdivisée en trois
niveaux hiérarchiques: diversité de l’écosystème, diver-
sité des espèces, diversité génétique (Vellend et Geber
2005; fig. 2). La diversité génétique comprend la diver-
sité des gènes et des allèles se rapportant à une espèce
et constitue la base de la diversité de tous les organismes
vivants. Elle est aussi le préalable à la capacité d’adapta-
Roland Kölliker, Luisa Last, Felix Herzog et Franco Widmer
Agroscope, Institut des sciences en durabilité agronomique IDU, 8046 Zurich, Suisse
Renseignements: Roland Kölliker, e-mail: [email protected]
La diversité génétique dans l'agriculture
P r o d u c t i o n v é g é t a l e
Figure 1 | La diversité à l'intérieur des espèces et entre les espèces est une composante importante des éco-systèmes agricoles. (Photo: Luisa Last, Agroscope)

La diversité génétique dans l'agriculture | Production végétale
13
Rés
um
é
Recherche Agronomique Suisse 5 (1): 12–19, 2014
La diversité génétique – la variété des gènes
et des allèles à l'intérieur d'une espèce –
définit l'élément de base de la biodiversité.
C'est une condition essentielle pour la
productivité et la durabilité des systèmes de
production agricole. Nous avons développé
et appliqué différentes méthodes pour
apprécier la diversité génétique dans
plusieurs exploitations agricoles d'Europe,
d'Ukraine et d'Ouganda. Une enquête
reposant sur un questionnaire touchant
203 exploitations a montré que la diversité
génétique parmi les plantes cultivées et les
animaux de rente différait fortement d'une
région à l'autre et en fonction des systèmes
de production les plus répandus. Pour mieux
saisir la diversité génétique des prairies,
nous avons examiné 60 populations de
dactyle de Bulgarie, de Norvège et de Suisse
en recourant à des marqueurs génétiques.
Cette espèce se caractérise par une très
grande variabilité à l'intérieur des popula-
tions. Cependant, les différences entre
populations sont plutôt faibles.
cadre du projet de recherche EU BioBio (EU FP7, KKBBE-
227161, www.biobio-indicator.org); il a permis de déve-
lopper et mettre au point des indicateurs fiables pour les
trois niveaux de la biodiversité (Herzog et al. 2012).
La diversité génétique dans l’agriculture
La diversité génétique dans les écosystèmes agricoles
peut être subdivisée en deux catégories principales:
•• La diversité planifiée, qui comprend la diversité
génétique parmi les espèces de plantes cultivées et
parmi les animaux de rente (Vandermeer et al. 1998).
Ces ressources tant végétales qu’animales sont à la
base de la production agricole et elles sont gérées par
les agriculteurs.
•• La diversité associée, qui comprend la diversité parmi
et entre les espèces végétales et animales présentes
dans les écosystèmes agricoles sans être incluses
toutefois dans la gestion de l’exploitation (Biala et al.
2005).
Par rapport à ces définitions, les prairies pluriannuelles et
les pâturages, tels qu’on les rencontre fréquemment en
Europe centrale, occupent une position intermédiaire. Ils
sont bien exploités, au sens agricole du terme, mais leur
diversité génétique n’est influencée par les agriculteurs
qu’au commencement. Par la suite, cette diversité est
Diversité des écosystèmes
Diversité des espèces
Diversité génétique
Figure 2 | Les trois niveaux de la biodiversité en agriculture. (Photos: Luisa Last, Agroscope [en haut à gauche], Salah Garchi, INRGREF [en haut à droite], Markus Zuber [au milieu à droite], Gabriela Brändle, Agroscope [toutes les autres])

Production végétale | La diversité génétique dans l'agriculture
14 Recherche Agronomique Suisse 5 (1): 12–19, 2014
influencée de façon prépondérante par l’environnement
et par le mode d’exploitation. On constate qu’une forte
intensité d’exploitation peut exercer des effets négatifs
sur la diversité génétique (Peter-Schmid et al. 2008).
Détermination de la diversité génétique
Au cours des dernières décennies, une intensification
générale des pratiques agricoles a fortement réduit la
diversité planifiée parmi les espèces végétales et ani-
males. Ainsi, en Allemagne ou en Finlande par exemple,
presque toutes les variétés locales de céréales cultivées
autrefois ont disparu (Hammer et Diederichsen 2009); en
Hollande, le nombre des races locales de bovidés a dimi-
nué de quelque 90 % au cours des 30 dernières années
(Buiteveld et al. 2009). Bien que des études isolées docu-
mentent ces pertes, il manque des méthodes fiables et
simples qui permettent de suivre l’évolution de la diver-
sité génétique dans l’agriculture. Pour cette étude, nous
avons mis au point et utilisé un questionnaire qui permet
une évaluation grossière de la diversité génétique de
variétés de plantes et de races d’animaux. En plus, nous
avons déterminé l’influence du mode d’exploitation et
celle de facteurs environnementaux sur la diversité géné-
tique du dactyle (Dactylis glomerata), une espèce que
l’on rencontre fréquemment dans les prairies et les pâtu-
rages, au moyen de marqueurs génétiques moléculaires.
M a t é r i e l e t m é t h o d e s
Enquête auprès de 203 exploitations
En se basant sur la littérature ainsi que sur une enquête
menée auprès d’experts et de groupes d’intéressés au
projet BioBio; un questionnaire a été développé pour
évaluer la diversité génétique dans 203 exploitations.
Ces questionnaires ont été remplis au cours d’entretiens
personnels avec les agriculteurs. En plus des questions se
rapportant aux systèmes de production agricole, ils pré-
voyaient des questions détaillées sur les noms, la prove-
nance et la quantité des plantes cultivées ainsi que des
animaux de rente. Les relevés ont été répartis en treize
études de cas BioBio (onze en Europe, une en Ouganda
et une en Ukraine; fig. 3). A partir des éléments recueillis,
des indicateurs de diversité ont été calculés, comme la
diversité des espèces, la diversité des variétés, celle des
variétés locales ainsi que le nombre de races de moutons
et de bovins par exploitation. En outre, la part de varié-
tés et de races rares a été analysée.
La diversité génétique du dactyle
Pour l’analyse détaillée de la diversité génétique en éco-
systèmes herbagers, le dactyle a été choisi, une espèce
qui se trouve dans les études conduites en trois régions
herbagères différentes (Rhodopen/Bulgarie, Hedmark/
Norvège et Obwald/Suisse). Chaque étude de cas régio-
nale englobait dix exploitations sur chacune desquelles
deux surfaces ont été échantillonnées. Sur chaque sur-
face, on a prélevé sur 32 plantes des feuilles qui ont été
soumises à l’analyse génétique par marquage molécu-
laire. Pour la partie suisse de l’étude, on a sélectionné
cinq exploitations biologiques (selon les directives de
BioSuisse) et cinq exploitations classiques. Sur chaque
exploitation, on a choisi une surface exploitée intensive-
ment et une surface exploitée de manière peu intensive.
Sur les surfaces suisses, en plus des échantillons de
feuilles, 60 pousses en fleur ont été prélevées, que l’on a
placées en serre, isolées, jusqu’à maturité. La semence en
a été récoltée et les populations F1 semées en serre.
La diversité génétique de 60 populations provenant
des trois études de cas régionales a été déterminée au
moyen de 29 marqueurs SSR (simple sequence repeat)
(Last et al. 2013). Les profils des marqueurs SSR de chaque
plante ont été comparés entre eux et la diversité géné-
tique a été définie à l’intérieur des populations, entre
les populations et entre les régions étudiées. Comme
mesure de la diversité génétique dans les populations,
on a pris en compte l’hétérozygotie moyenne attendue
(HE) et la diversité génotypique selon Shannon (HG).
L’influence du mode d’exploitation a été examinée à
l’aide d’analyses de corrélation et de statistiques multi-
variables.
�
�
�
�
�
�
�
�
�
�
�
�
AT
BG
CHDE
ES1ES2
FR
HUIT
UA
NO
UK
UG
Figure 3 | Les treize analyses de cas régionales BioBio, dans lesquelles on a enregistré les variétés de plantes cultivées et les races d'animaux de rente au moyen de questionnaires (www.biobio-indicator.org).
La diversité génétique dans l'agriculture | Production végétale
15Recherche Agronomique Suisse 5 (1): 12–19, 2014
grandes cultures et herbages, ce qui conduisait naturel-
lement à la collecte d’un nombre d’espèces par exploita-
tion significativement plus élevé.
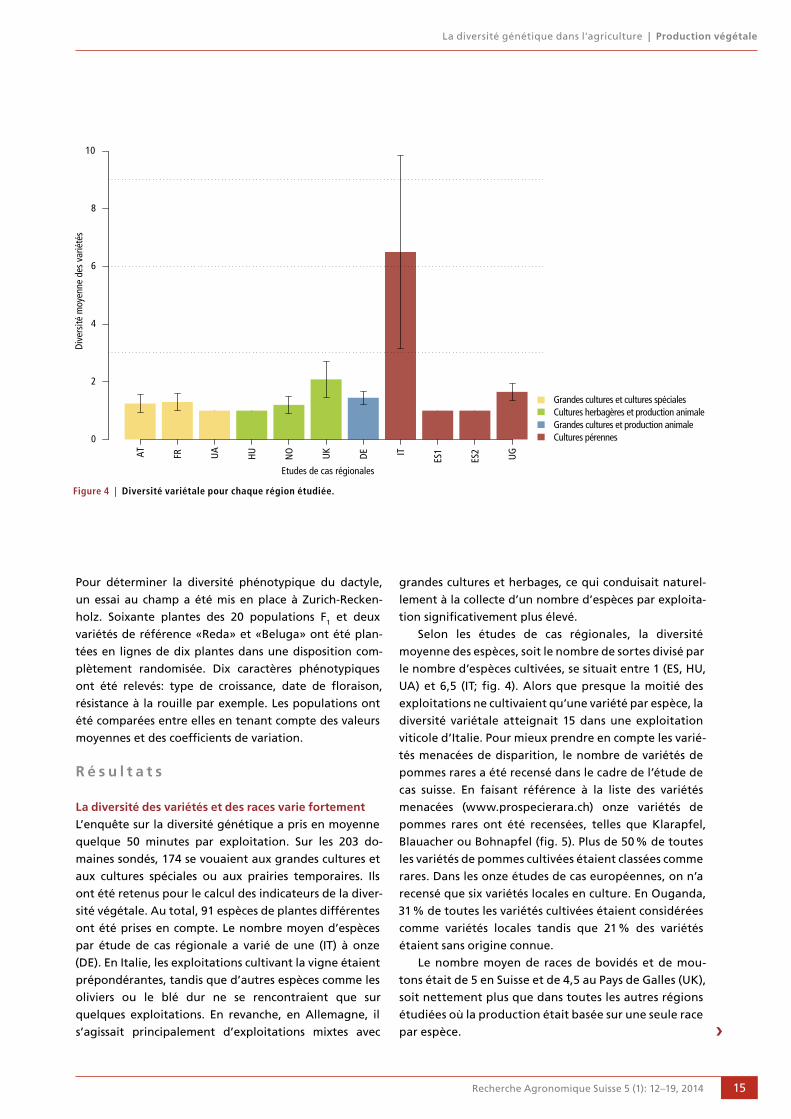
Selon les études de cas régionales, la diversité
moyenne des espèces, soit le nombre de sortes divisé par
le nombre d’espèces cultivées, se situait entre 1 (ES, HU,
UA) et 6,5 (IT; fig. 4). Alors que presque la moitié des
exploitations ne cultivaient qu’une variété par espèce, la
diversité variétale atteignait 15 dans une exploitation
viticole d’Italie. Pour mieux prendre en compte les varié-
tés menacées de disparition, le nombre de variétés de
pommes rares a été recensé dans le cadre de l’étude de
cas suisse. En faisant référence à la liste des variétés
menacées (www.prospecierara.ch) onze variétés de
pommes rares ont été recensées, telles que Klarapfel,
Blauacher ou Bohnapfel (fig. 5). Plus de 50 % de toutes
les variétés de pommes cultivées étaient classées comme
rares. Dans les onze études de cas européennes, on n’a
recensé que six variétés locales en culture. En Ouganda,
31 % de toutes les variétés cultivées étaient considérées
comme variétés locales tandis que 21 % des variétés
étaient sans origine connue.Le nombre moyen de races de bovidés et de mou-
tons était de 5 en Suisse et de 4,5 au Pays de Galles (UK),
soit nettement plus que dans toutes les autres régions
étudiées où la production était basée sur une seule race
par espèce.
Pour déterminer la diversité phénotypique du dactyle,
un essai au champ a été mis en place à Zurich-Recken-
holz. Soixante plantes des 20 populations F1 et deux
variétés de référence «Reda» et «Beluga» ont été plan-
tées en lignes de dix plantes dans une disposition com-
plètement randomisée. Dix caractères phénotypiques
ont été relevés: type de croissance, date de floraison,
résistance à la rouille par exemple. Les populations ont
été comparées entre elles en tenant compte des valeurs
moyennes et des coefficients de variation.
R é s u l t a t s
La diversité des variétés et des races varie fortement
L’enquête sur la diversité génétique a pris en moyenne
quelque 50 minutes par exploitation. Sur les 203 do-
maines sondés, 174 se vouaient aux grandes cultures et
aux cultures spéciales ou aux prairies temporaires. Ils
ont été retenus pour le calcul des indicateurs de la diver-
sité végétale. Au total, 91 espèces de plantes différentes
ont été prises en compte. Le nombre moyen d’espèces
par étude de cas régionale a varié de une (IT) à onze
(DE). En Italie, les exploitations cultivant la vigne étaient
prépondérantes, tandis que d’autres espèces comme les
oliviers ou le blé dur ne se rencontraient que sur
quelques exploitations. En revanche, en Allemagne, il
s’agissait principalement d’exploitations mixtes avec
AT FR UA
HU NO UK
DE IT
ES1
ES2
UG
Etudes de cas régionales
0
2
4
6
8
10
Dive
rsité
moy
enne
des
var
iété
s
Grandes cultures et cultures spécialesCultures herbagères et production animaleGrandes cultures et production animaleCultures pérennes
Figure 4 | Diversité variétale pour chaque région étudiée.

Production végétale | La diversité génétique dans l'agriculture
16 Recherche Agronomique Suisse 5 (1): 12–19, 2014
Grande diversité du dactyle en Europe
Avec les 29 marqueurs SSR 257 allèles différents ont pu
être identifiés sur l’ensemble des plantes de dactyle ana-
lysées. La diversité génétique (hétérozygotie moyenne
attendue HE) à l’intérieur des populations était très éle-
vée dans les trois analyses de cas régionales, variant de
0,44 à 0,59. En Norvège, la HE atteignait 0,54, une valeur
significativement plus élevée que celle de Bulgarie (0,52),
alors qu’en Suisse la HE prenait une position intermé-
diaire avec 0,53. Malgré la grande diversité à l’intérieur
des populations, on pouvait considérer que 62 % des
plantes bulgares et 91 % des plantes suisses étaient spé-
cifiques à la région concernée. Une analyse par clusters
basée sur la fréquence des marqueurs par étude de cas
régionale situait les populations de dactyle suisses dans
le même compartiment que les populations norvé-
giennes, tandis que celles de Bulgarie formaient un
groupe à part (fig. 6A). Ce groupement ne correspondait
cependant pas au groupement par site basé sur la dis-
tance géographique (fig. 6B).
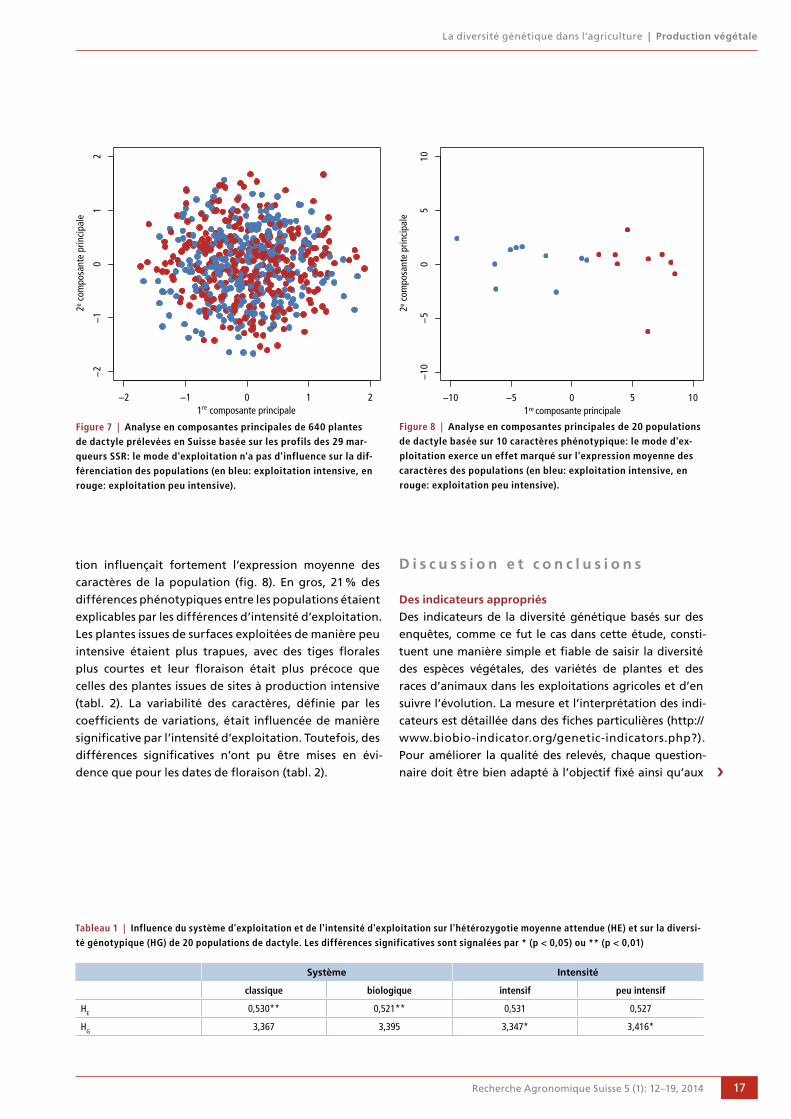
Mode d’exploitation et diversité génétiqueComme mentionné précédemment dans les comparaisons
entre études de cas régionales, la diversité génétique à
l’intérieur des populations est très élevée en Suisse. Seuls
4 % de la diversité observée était liée à des différences
entre populations, en revanche la diversité résiduelle
était attribuable à chaque population. Par conséquent, il
n’était pas possible de subdiviser les populations, ni
d’identifier des différences attribuables au mode d’ex-
ploitation (fig. 7). En revanche, les systèmes d’exploita-
tion (biologique vs. classique) ont eu une influence sur la
diversité génétique (HE), le système intensif correspon-
dant à une fréquence des génotypes (HG) significative-
ment plus faible (tabl. 1). Tous les autres paramètres
décrivant le site (composition botanique, situation et
exposition des surfaces examinées) ne paraissaient pas
influencer la diversité génétique des populations.
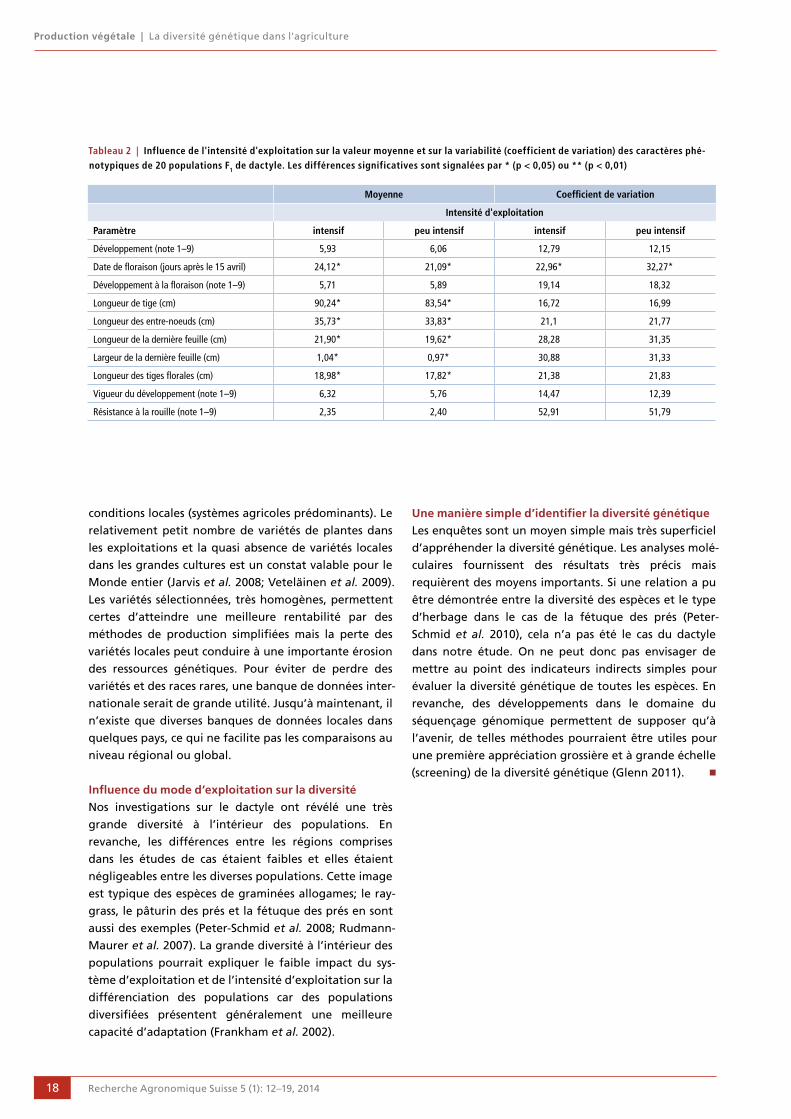
La caractérisation phénotypique des populations F1
a montré que celles-ci se différenciaient nettement par
leurs propriétés agronomiques; l’intensité d’exploita-
Suis
se
Bulg
arie
Nor
vège
(A) Marqueurs SSR
Dist
ance
euc
lidie
nne
0,0
0,5
1,0
1,5
2,0
Nor
vège
Suis
se
Bulg
arie
(B) Position géographique
Dist
ance
(km
)
0
750
1500
2250
Figure 6 | Analyse par clusters dans les trois études de cas régionales, en Bulgarie, Norvège et en Suisse, appliquées sur les analyses SSR des populations de dactyle (A) ainsi que sur les distances géographiques entre les régions de collecte (B).
Figure 5 | Des variétés de pommes rares: Klarapfel, Blauacher, Bohnapfel (de gauche à droite). (Photos: Markus Zuber)
La diversité génétique dans l'agriculture | Production végétale
17Recherche Agronomique Suisse 5 (1): 12–19, 2014
D i s c u s s i o n e t c o n c l u s i o n s
Des indicateurs appropriés
Des indicateurs de la diversité génétique basés sur des
enquêtes, comme ce fut le cas dans cette étude, consti-
tuent une manière simple et fiable de saisir la diversité
des espèces végétales, des variétés de plantes et des
races d’animaux dans les exploitations agricoles et d’en
suivre l’évolution. La mesure et l’interprétation des indi-
cateurs est détaillée dans des fiches particulières (http://
www.biobio-indicator.org/genetic-indicators.php?).
Pour améliorer la qualité des relevés, chaque question-
naire doit être bien adapté à l’objectif fixé ainsi qu’aux
−2 −1 0 1 2
−2
−1
01
2
1re composante principale
2e com
posa
nte
prin
cipa
le
Figure 7 | Analyse en composantes principales de 640 plantes de dactyle prélevées en Suisse basée sur les profils des 29 mar-queurs SSR: le mode d'exploitation n'a pas d'influence sur la dif-férenciation des populations (en bleu: exploitation intensive, en rouge: exploitation peu intensive).
Système Intensité
classique biologique intensif peu intensif
HE 0,530** 0,521** 0,531 0,527
HG 3,367 3,395 3,347* 3,416*
Tableau 1 | Influence du système d'exploitation et de l'intensité d'exploitation sur l'hétérozygotie moyenne attendue (HE) et sur la diversi-té génotypique (HG) de 20 populations de dactyle. Les différences significatives sont signalées par * (p < 0,05) ou ** (p < 0,01)
tion influençait fortement l’expression moyenne des
caractères de la population (fig. 8). En gros, 21 % des
différences phénotypiques entre les populations étaient
explicables par les différences d’intensité d’exploitation.
Les plantes issues de surfaces exploitées de manière peu
intensive étaient plus trapues, avec des tiges florales
plus courtes et leur floraison était plus précoce que
celles des plantes issues de sites à production intensive
(tabl. 2). La variabilité des caractères, définie par les
coefficients de variations, était influencée de manière
significative par l’intensité d’exploitation. Toutefois, des
différences significatives n’ont pu être mises en évi-
dence que pour les dates de floraison (tabl. 2).
−10 −5 0 5 10
−10
−5
05
101re composante principale
2e com
posa
nte
prin
cipa
le
Figure 8 | Analyse en composantes principales de 20 populations de dactyle basée sur 10 caractères phénotypique: le mode d'ex-ploitation exerce un effet marqué sur l'expression moyenne des caractères des populations (en bleu: exploitation intensive, en rouge: exploitation peu intensive).
18
Production végétale | La diversité génétique dans l'agriculture
Recherche Agronomique Suisse 5 (1): 12–19, 2014
conditions locales (systèmes agricoles prédominants). Le
relativement petit nombre de variétés de plantes dans
les exploitations et la quasi absence de variétés locales
dans les grandes cultures est un constat valable pour le
Monde entier (Jarvis et al. 2008; Veteläinen et al. 2009).
Les variétés sélectionnées, très homogènes, permettent
certes d’atteindre une meilleure rentabilité par des
méthodes de production simplifiées mais la perte des
variétés locales peut conduire à une importante érosion
des ressources génétiques. Pour éviter de perdre des
variétés et des races rares, une banque de données inter-
nationale serait de grande utilité. Jusqu’à maintenant, il
n’existe que diverses banques de données locales dans
quelques pays, ce qui ne facilite pas les comparaisons au
niveau régional ou global.
Influence du mode d’exploitation sur la diversité
Nos investigations sur le dactyle ont révélé une très
grande diversité à l’intérieur des populations. En
revanche, les différences entre les régions comprises
dans les études de cas étaient faibles et elles étaient
négligeables entre les diverses populations. Cette image
est typique des espèces de graminées allogames; le ray-
grass, le pâturin des prés et la fétuque des prés en sont
aussi des exemples (Peter-Schmid et al. 2008; Rudmann-
Maurer et al. 2007). La grande diversité à l’intérieur des
populations pourrait expliquer le faible impact du sys-
tème d’exploitation et de l’intensité d’exploitation sur la
différenciation des populations car des populations
diversifiées présentent généralement une meilleure
capacité d’adaptation (Frankham et al. 2002).
Une manière simple d’identifier la diversité génétique
Les enquêtes sont un moyen simple mais très superficiel
d’appréhender la diversité génétique. Les analyses molé-
culaires fournissent des résultats très précis mais
requièrent des moyens importants. Si une relation a pu
être démontrée entre la diversité des espèces et le type
d’herbage dans le cas de la fétuque des prés (Peter-
Schmid et al. 2010), cela n’a pas été le cas du dactyle
dans notre étude. On ne peut donc pas envisager de
mettre au point des indicateurs indirects simples pour
évaluer la diversité génétique de toutes les espèces. En
revanche, des développements dans le domaine du
séquençage génomique permettent de supposer qu’à
l’avenir, de telles méthodes pourraient être utiles pour
une première appréciation grossière et à grande échelle
(screening) de la diversité génétique (Glenn 2011). n
Moyenne Coefficient de variation
Intensité d'exploitation
Paramètre intensif peu intensif intensif peu intensif
Développement (note 1–9) 5,93 6,06 12,79 12,15
Date de floraison (jours après le 15 avril) 24,12* 21,09* 22,96* 32,27*
Développement à la floraison (note 1–9) 5,71 5,89 19,14 18,32
Longueur de tige (cm) 90,24* 83,54* 16,72 16,99
Longueur des entre-noeuds (cm) 35,73* 33,83* 21,1 21,77
Longueur de la dernière feuille (cm) 21,90* 19,62* 28,28 31,35
Largeur de la dernière feuille (cm) 1,04* 0,97* 30,88 31,33
Longueur des tiges florales (cm) 18,98* 17,82* 21,38 21,83
Vigueur du développement (note 1–9) 6,32 5,76 14,47 12,39
Résistance à la rouille (note 1–9) 2,35 2,40 52,91 51,79
Tableau 2 | Influence de l'intensité d'exploitation sur la valeur moyenne et sur la variabilité (coefficient de variation) des caractères phé-notypiques de 20 populations F1 de dactyle. Les différences significatives sont signalées par * (p < 0,05) ou ** (p < 0,01)

19
La diversité génétique dans l'agriculture | Production végétale
Ria
ssu
nto
Sum
mar
y
Genetic diversity in agriculture
Genetic diversity – the variety of genes
and alleles within a species – constitutes
the most basic level of biodiversity, and is
an important prerequisite for productivity
and sustainability in agricultural produc-
tion systems. We have developed and
applied various methods to determine
genetic diversity on farms in Europe,
Ukraine and Uganda. A questionnaire-
based survey conducted on a total of 203
farms has shown that the genetic diversity
of crops and livestock varies greatly
between different regions, and depends
upon the prevailing production systems. In
order to obtain a more accurate insight
into the genetic diversity of grassland, we
studied 60 populations of orchard grass
from Bulgaria, Norway and Switzerland
with molecular genetic markers. In doing
so, we learned that although this species
exhibits a very large variability within the
populations, the differences between the
populations tend to be minor.
Key words: genetic diversity, indicators,
grassland.
Diversità genetica nell'agricoltura
La diversità genetica, ossia la varietà dei
geni o degli alleli in una specie, è alla
base della biodiversità e costituisce un
importante presupposto per la produtti-
vità e la sostenibilità dei sistemi produt-
tivi agricoli. Abbiamo sviluppato e
applicato diversi metodi per determi-
nare la diversità genetica presente nelle
aziende agricole in Europa, Ucraina e
Uganda. Un sondaggio basato su di un
questionario, effettuato su un totale di
203 aziende, ha mostrato che la diver-
sità genetica di piante coltivate e
animali allevati varia notevolmente tra
le diverse regioni ed è strettamente correlata al sistema di produzione
predominante. Per avere una visione più
precisa della diversità genetica della
superficie erbosa, abbiamo analizzato
60 popolazioni di dattile provenienti
dalla Bulgaria, dalla Norvegia e dalla
Svizzera con marcatori genetico-moleco-
lari. Quest’analisi ha mostrato che questa
specie presenta una grande variabilità
all’interno delle popolazioni, tuttavia le
differenze tra le varie popolazioni si sono
dimostrate piuttosto esigue.
Recherche Agronomique Suisse 5 (1): 12–19, 2014
Bibliographie ▪ Biala K., Peeters A., Muys B., Hermy M., Brouckaert V. et al., 2005. Bio-diversitiy indicators as a tool to assess sustainability levels of agro- ecosystems, with a special consideration of grassland areas. Options Méditerranéennes. Série A: Séminaires Méditerranéens 67, 439–443.
▪ Buiteveld J., van Veller M.G.P., Hiemstra S.J., ten Brink B. & Tekelenburg T., 2009. An exploration of monitoring and modelling agrobiodiversity: From indicator development towards modelling biodiversity in agricultu-ral systems on the sub-specific level. CGN/DLO Foundation, Wageningen, The Netherlands. Accès: http://edepot.wur.nl/51240 [22.10.2013]
▪ Frankham R., Ballou J.D. & Briscoe D.A., 2002. Introduction to conserva-tion genetics. Cambridge University Press, Cambridge, UK, 617 p.
▪ Glenn T.C., 2011. Field guide to next-generation DNA sequencers. Molecular Ecology Resources 11 (5), 759–769.
▪ Hammer K. & Diederichsen A., 2009. Evolution, status and perspectives for landraces in Europe. In: European landraces: On-farm conservation managment and use (Eds. M. Veteläinen, V. Negri & N. Maxted). Bio-diversity International, Italy, Rome, 23–44.
▪ Herzog F., Balázs K., Dennis P., Friedel J., Geijzendorffer I. et al., 2012. Biodiversity indicators for European farming systems. A guidebook. ART-Schriftenreihe 17, 1–99.
▪ Jarvis D.I., Brown A.H.D., Cuong P.H., Collado-Panduro L., Latournerie-Moreno L. et al., 2008. A global perspective of the richness and evenness of traditional crop-variety diversity maintained by farming communities. Proceedings of the National Academy of Sciences 105 (14), 5326–5331.
▪ Last L., Widmer F., Fjellstad W., Stoyanova S. & Kölliker R., 2013. Genetic diversity of natural orchardgrass (Dactylis glomerata L.) populations in three regions in Europe. BMC Genetics 14, 102.
▪ Peter-Schmid M., Boller B. & Kölliker R., 2008. Habitat and management affect genetic structure of Festuca pratensis but not Lolium multiflorum ecotype populations. Plant Breeding 127 (5), 510–517.
▪ Peter-Schmid M., Kölliker R. & Boller B., 2010. Genetic diversity of Festuca pratensis Huds. and Lolium multiflorum Lam. ecotype populati-ons in relation to species diversity and grassland type. In: Grassland Bio-diversity: Habitat Types, Ecological Processes and Environmental Impacts (Eds. J. Runas & T. Dahlgren). Nova Science Publishers, New York, 377 p.
▪ Rudmann-Maurer K., Weyand A., Fischer M. & Stocklin J., 2007. Micro-satellite diversity of the agriculturally important alpine grass Poa alpina in relation to land use and natural environment. Annals of Botany 100 (6), 1249–1258.
▪ Vandermeer J., van Noordwijk M., Anderson J., Ong C. & Perfecto I., 1998. Global change and multi-species agroecosystems: Concepts and issues. Agriculture, Ecosystems and Environment 67 (1), 1–22.
▪ Vellend M. & Geber M.A., 2005. Connection between species diversity and genetic diversity. Ecology Letters 8, 767–781.
▪ Veteläinen M., Negri V. & Maxted N., 2009. European landraces: On-farm conservation managment and use. Bioversity International, Rome, Italy, 358 p.

20 Recherche Agronomique Suisse 5 (1): 20–27, 2014
une colonisation naturelle d’espèces typiques à partir
des environs, ces surfaces sont souvent ensemencées à
l’aide d’un mélange de graines. Cette démarche pose
cependant certains problèmes (Vander Mijnsbrugge et
al. 2010). D’une part, les semences des mélanges peuvent
ne pas provenir de la même région. Il est alors possible
qu’elles soient peu ou pas du tout adaptées aux surfaces
renaturées (Bischoff et al. 2006). C’est pourquoi Info-
Flora a proposé aux producteurs de semences de n’utili-
ser que des semences de la même région biogéogra-
phique pour leurs mélanges (http://www.infoflora.ch/fr/
flore/semences-plantes-sauvages/). Mais même si les
graines proviennent de la même région, leur multiplica-
tion dans le jardin du producteur peut entraîner des
pertes d’adaptation aux conditions naturelles (Schoen &
Brown 2001). En outre, les mélanges ne sont en général
accompagnés d’aucune information sur la qualité géné-
tique des semences. Pourtant, la diversité biologique
peut exercer un effet significatif sur la performance à
court terme des plantes et ainsi influencer de façon
déterminante le succès des ensemencements. Une diver-
sité génétique élevée aide également à conserver la
capacité d’adaptation des plantes. Il importe donc de
connaître la composition génétique et la performance
des mélanges de semences afin d’éviter des consé-
quences négatives dues à des semences mal adaptées et/
ou génétiquement appauvries. Cette étude visait à étu-
dier la diversité génétique et la performance de popula-
tions naturelles et de populations semées de Lychnis flos-
cuculi dans le paysage agricole de Haute-Argovie. En
outre, des essais ont été menés en plein champ et en
jardin d’essai, afin d’étudier l’adaptabilité à l’environne-
ment local des plantes issues de populations naturelles
et de populations issues d’ensemencements, ainsi que de
plantes élevées à partir de graines obtenues directement
chez les producteurs. Enfin, une expérience en chambre
climatique, en conditions contrôlées, a permis d’étudier
la réaction des mêmes catégories de plantes à la disponi-
bilité en eau et en nutriments.
I n t r o d u c t i o n
La perte de grandes surfaces d’habitats naturels à la
suite de l’exploitation intensive des terres a entraîné une
diminution significative de la biodiversité dans les pay-
sages agricoles (Billeter et al. 2008). Diverses mesures ont
été prises pour stopper ces pertes et développer les
réseaux entre les habitats restants. Les surfaces de com-
pensation agricoles vont dans ce sens. Comme le réser-
voir local d’espèces est trop appauvri pour permettre
Effets de mélanges de semences sur la diversité génétique et la performanceTsipe Aavik1,2, Daniel Bosshard2, Peter Edwards2, Rolf Holderegger2,3 et Regula Billeter2,4
1Institut d’écologie et des sciences de la terre, Université de Tartu, 51005 Tartu, Estonie2Institut de biologie intégrée, EPF Zurich, 8092 Zurich, Suisse3Institut fédéral de recherches sur la forêt, la neige et le paysage WSL, 8903 Birmensdorf, Suisse4School of Life Sciences and Facility Management ZHAW, 8820 Wädenswil, Suisse
Renseignements: Regula Billeter, e-mail: [email protected]
E n v i r o n n e m e n t
Figure 1 | Diverses mesures de valorisation ont été prises en Haute-Argovie, par exemple la renaturation de prairies exploitées extensivement. La bande visible sur la photo fait partie d’une sur-face de compensation écologique ensemencée à l’aide d’un mé-lange de semences sauvages contenant aussi des graines de la silène fleur de coucou (Lychnis flos-cuculi).

Effets de mélanges de semences sur la diversité génétique et la performance | Environnement
21
Rés
um
é
Recherche Agronomique Suisse 5 (1): 20–27, 2014
Des mélanges de semences de plantes
sauvages sont souvent utilisés pour ense-
mencer des surfaces renaturées, notamment
pour augmenter la diversité des espèces.
Mais les conséquences de tels mélanges sur
la diversité génétique et sur la performance
sont souvent peu claires. La diversité
génétique de populations naturelles et de
populations issues de semis de la silène fleur
de coucou (Lychnis flos-cuculi) a été étudiée
en Haute-Argovie, sur le Plateau suisse. En
outre, les effets de la diversité génétique, de
l’origine et des conditions environnemen-
tales sur la croissance et la capacité de survie
des plantes ont été mesurées en plein champ,
en jardin d’essai et en chambres climatiques.
La diversité génétique de ces deux types de
population ne se distingue presque pas, mais
le taux de consanguinité est significative-
ment plus élevé dans les populations semées
que dans les populations naturelles. La
composition des deux types de populations
est également très différente. Les plantes
issues de graines obtenues directement chez
les producteurs ont fleuri moins souvent et
tendanciellement plus tard que les popula-
tions naturelles. Mais sinon, l’origine ou la
diversité génétique n’influencent pas la
performance des plantes. En conclusion,
l’ensemencement avec des mélanges de
semences est une mesure adéquate, surtout
lorsque la production de semences s’effectue
sur une base génétique suffisamment large.
Dans ce cas, pratiquement aucun effet
négatif sur la performance n’est attendu.
Mais lorsque c’est possible, la colonisation
naturelle à partir de l’espace environnant est
préférable.
M a t é r i e l e t m é t h o d e s
Populations étudiées, relevés de terrain et analyses
génétiques
L’étude a été menée dans une région du réseau Eme-
raude, localisée en Haute-Argovie et sur le canton de
Berne. Il s’agit d’une région agricole typique de cultures
et de pâturages. A la suite de la construction de Rail
2000, de nombreuses surfaces de compensation ont été
créées entre 2001 et 2003 le long de fossés et de ruis-
seaux. Ces bandes ont été ensemencées à l’aide d’un
mélange de semences issues de plantes sauvages (fig. 1)
contenant la silène fleur de coucou étudiée (Lychnis flos-
cuculi; fig. 2). En 2005 et 2006, de nouvelles mesures de
valorisation ont été réalisées et de nouvelles surfaces
ensemencées avec le même mélange. En 2009 et 2010,
toutes les populations de L. flos-cuculi ont été cartogra-
phiées dans le périmètre d’étude. Lychnis flos-cuculi a
été semée à huit endroits et quinze populations natu-
relles de cette espèce ont été trouvées. Le mélange de
semences a en outre été répandu dans des prairies
humides à trois emplacements, si bien qu’il n’était pas
possible de savoir avec certitude si les populations ren-
contrées étaient naturelles ou semées.
Figure 2 | La silène fleur de coucou (Lychnis flos-cuculi) est une espèce des prairies calcicoles et humides. Cette plante est rare dans les espaces agricoles exploités intensivement; elle est en général localisée sur le bord des fossés et des ruisseaux ou en lisière.
Environnement | Effets de mélanges de semences sur la diversité génétique et la performance
22 Recherche Agronomique Suisse 5 (1): 20–27, 2014
Nous avons déterminé la taille de chaque population et
récolté des feuilles de 30 individus par population. Ces
feuilles ont été séchées et préparées en vue de l’analyse
ADN, pour laquelle nous avons utilisé six marqueurs
microsatellites (Galeuchet et al. 2002; Moccia et al. 2009).
Le déroulement de ces analyses est décrit en détail dans
Aavik et al. (2012). Les critères de performance suivants
ont également été décrits, pour 30 individus choisis au
hasard dans 20 populations: nombre de fleurs, nombre
de tiges par plante et hauteur de la plante. En outre,
100 graines par plante ont été récoltées et pesées. Nous
avons déterminé le taux de germination de 50 graines
prélevées chaque fois sur 15 individus par population et
semées dans des boîtes de Petri. Le nombre de graines
ayant germé a été compté après 30 jours. Enfin, des
échantillons de sol ont été prélevés pour chaque popula-
tion, afin de déterminer les teneurs en nitrate et en
phosphore et l’humidité du sol a été mesurée (détails sur
la méthode: voir Aavik et al. en impression).
Expérimentations
Afin de comparer la performance de plantes issues de
populations naturelles, d’un ensemencement ou de
semences obtenues directement chez les producteurs,
nous les avons semées en plein champ et dans le jardin
d’essai de l’EPF à Hönggerberg (Zurich). Nous avons dans
ce but utilisé des semences de deux populations de
Haute-Argovie issues de semis ainsi que des semences de
deux producteurs différents. L’expérience a débuté en
septembre 2010 sur deux sites en Haute- Argovie et au
jardin d’essai. Sur le terrain, quatre platebandes ont été
installées à chaque endroit, avec six plots chacune. Dans
le jardin d’essai, 24 pots ont été remplis d’un mélange de
terre et de sable, puis recouverts d’une fine couche d’hu-
mus. Nous avons semé 200 graines dans chaque plot,
que ce soit sur le terrain ou au jardin. Les semences de
chaque origine ont été semées une fois dans chaque pla-
tebande avec une origine par plot (deux populations
naturelles, deux populations semées et les semences de
deux producteurs). Dans le jardin d’essai, chaque origine
a été semée dans quatre pots différents. En mai 2011, le
nombre de plantules a été réduit à dix individus qui ont
pu poursuivre leur croissance. En septembre 2011, le
taux de survie a été mesuré, ainsi que divers critères de
performance (diamètre de la rosette, feuille la plus
longue, hauteur de la plante, nombre de pousses,
nombre de fleurs et biomasse des individus séchés). Dans
une deuxième expérience, nous avons étudié l’effet des
nutriments et de l’humidité du sol sur la performance
des plantes, avec les mêmes populations utilisées dans
l’essai en chambre climatique mentionnée précédem-
ment. Nous avons choisi au hasard 36 plantules de
chaque population et les avons repiquées en pot. Les
216 pots obtenus au total ont été répartis de façon aléa-
toire dans les trois chambres climatiques. Quatre traite-
ments différents ont été réalisés, qui consistaient à com-
biner de faibles et de fortes quantités d’eau avec des
doses d’engrais faibles et fortes. Le début de la floraison
de chaque plante a été notifié. Après 70 jours, nous
avons mesuré la hauteur des plantes en fleur, le dia-
Figure 3 | (a) Diversité génétique (moyenne ± erreur-standard) et (b) coefficient de consanguinité de populations semées et naturelles de la silène fleur de coucou (Lychnis flos-cuculi) en Haute-Argovie (Aavik et al. 2012). Les populations semées présentent une consanguinité signi-ficativement plus élevée que les populations naturelles (pseudo-F1,20=11,634, P=0,003).
Semé
0,60
0,63
0,66
0,69
Dive
rsité
gén
étiq
ue
0,00
0,04
0,08
0,12
0,16
Croi
sem
ent
Naturel Semé Naturel
(a) (b)
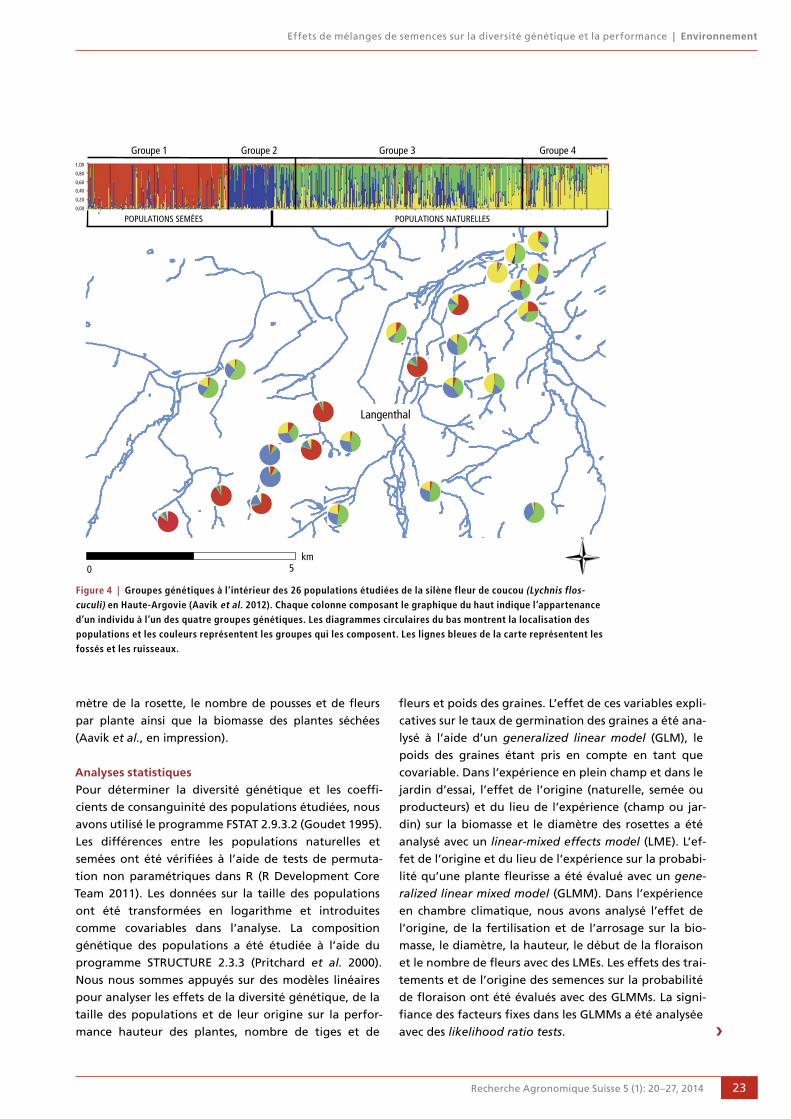
Effets de mélanges de semences sur la diversité génétique et la performance | Environnement
23Recherche Agronomique Suisse 5 (1): 20–27, 2014
fleurs et poids des graines. L’effet de ces variables expli-
catives sur le taux de germination des graines a été ana-
lysé à l’aide d’un generalized linear model (GLM), le
poids des graines étant pris en compte en tant que
covariable. Dans l’expérience en plein champ et dans le
jardin d’essai, l’effet de l’origine (naturelle, semée ou
producteurs) et du lieu de l’expérience (champ ou jar-
din) sur la biomasse et le diamètre des rosettes a été
analysé avec un linear-mixed effects model (LME). L’ef-
fet de l’origine et du lieu de l’expérience sur la probabi-
lité qu’une plante fleurisse a été évalué avec un gene-
ralized linear mixed model (GLMM). Dans l’expérience
en chambre climatique, nous avons analysé l’effet de
l’origine, de la fertilisation et de l’arrosage sur la bio-
masse, le diamètre, la hauteur, le début de la floraison
et le nombre de fleurs avec des LMEs. Les effets des trai-
tements et de l’origine des semences sur la probabilité
de floraison ont été évalués avec des GLMMs. La signi-
fiance des facteurs fixes dans les GLMMs a été analysée
avec des likelihood ratio tests.
mètre de la rosette, le nombre de pousses et de fleurs
par plante ainsi que la biomasse des plantes séchées
(Aavik et al., en impression).
Analyses statistiques
Pour déterminer la diversité génétique et les coeffi-
cients de consanguinité des populations étudiées, nous
avons utilisé le programme FSTAT 2.9.3.2 (Goudet 1995).
Les différences entre les populations naturelles et
semées ont été vérifiées à l’aide de tests de permuta-
tion non paramétriques dans R (R Development Core
Team 2011). Les données sur la taille des populations
ont été transformées en logarithme et introduites
comme covariables dans l’analyse. La composition
génétique des populations a été étudiée à l’aide du
programme STRUCTURE 2.3.3 (Pritchard et al. 2000).
Nous nous sommes appuyés sur des modèles linéaires
pour analyser les effets de la diversité génétique, de la
taille des populations et de leur origine sur la perfor-
mance hauteur des plantes, nombre de tiges et de
Figure 4 | Groupes génétiques à l’intérieur des 26 populations étudiées de la silène fleur de coucou (Lychnis flos- cuculi) en Haute-Argovie (Aavik et al. 2012). Chaque colonne composant le graphique du haut indique l’appartenance d’un individu à l’un des quatre groupes génétiques. Les diagrammes circulaires du bas montrent la localisation des populations et les couleurs représentent les groupes qui les composent. Les lignes bleues de la carte représentent les fossés et les ruisseaux.
Groupe 1 Groupe 2 Groupe 3 Groupe 4
POPULATIONS SEMÉES POPULATIONS NATURELLES
Langenthal
0 5 km
1,00
0,80
0,60
0,40
0,20
0,00
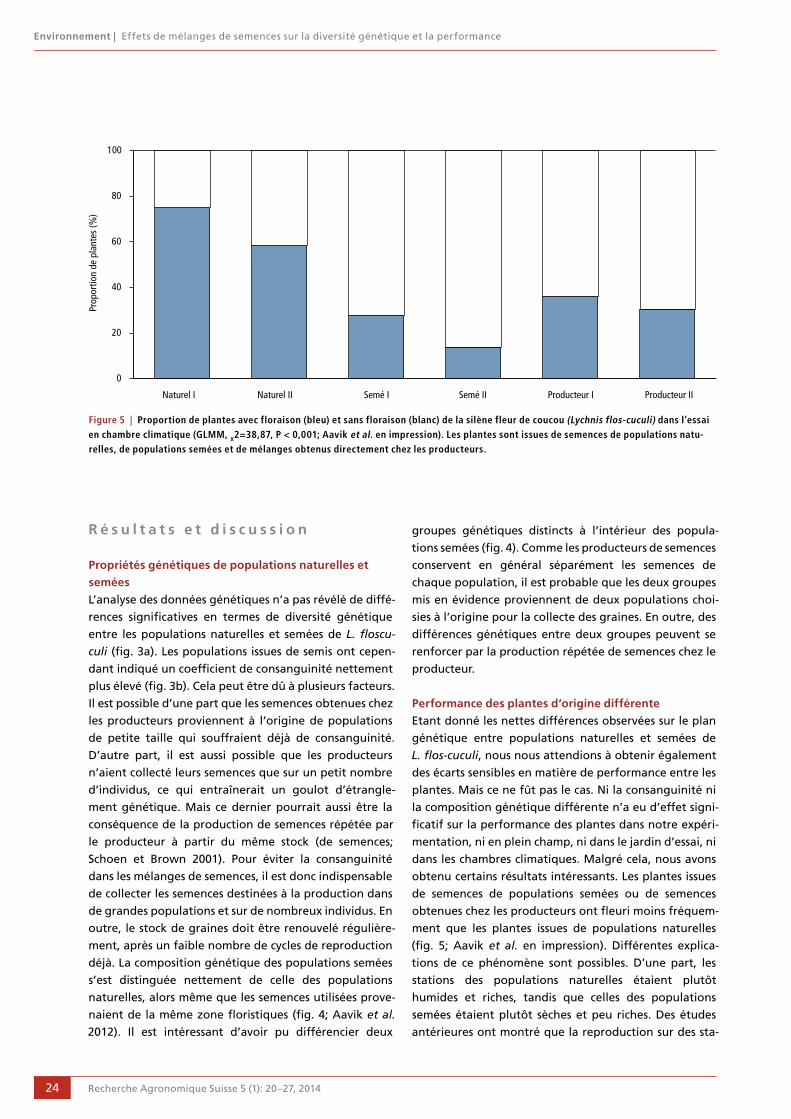
Environnement | Effets de mélanges de semences sur la diversité génétique et la performance
24 Recherche Agronomique Suisse 5 (1): 20–27, 2014
R é s u l t a t s e t d i s c u s s i o n
Propriétés génétiques de populations naturelles et
semées
L’analyse des données génétiques n’a pas révélé de diffé-
rences significatives en termes de diversité génétique
entre les populations naturelles et semées de L. floscu-
culi (fig. 3a). Les populations issues de semis ont cepen-
dant indiqué un coefficient de consanguinité nettement
plus élevé (fig. 3b). Cela peut être dû à plusieurs facteurs.
Il est possible d’une part que les semences obtenues chez
les producteurs proviennent à l’origine de populations
de petite taille qui souffraient déjà de consanguinité.
D’autre part, il est aussi possible que les producteurs
n’aient collecté leurs semences que sur un petit nombre
d’individus, ce qui entraînerait un goulot d’étrangle-
ment génétique. Mais ce dernier pourrait aussi être la
conséquence de la production de semences répétée par
le producteur à partir du même stock (de semences;
Schoen et Brown 2001). Pour éviter la consanguinité
dans les mélanges de semences, il est donc indispensable
de collecter les semences destinées à la production dans
de grandes populations et sur de nombreux individus. En
outre, le stock de graines doit être renouvelé régulière-
ment, après un faible nombre de cycles de reproduction
déjà. La composition génétique des populations semées
s’est distinguée nettement de celle des populations
naturelles, alors même que les semences utilisées prove-
naient de la même zone floristiques (fig. 4; Aavik et al.
2012). Il est intéressant d’avoir pu différencier deux
groupes génétiques distincts à l’intérieur des popula-
tions semées (fig. 4). Comme les producteurs de semences
conservent en général séparément les semences de
chaque population, il est probable que les deux groupes
mis en évidence proviennent de deux populations choi-
sies à l’origine pour la collecte des graines. En outre, des
différences génétiques entre deux groupes peuvent se
renforcer par la production répétée de semences chez le
producteur.
Performance des plantes d’origine différente
Etant donné les nettes différences observées sur le plan
génétique entre populations naturelles et semées de
L. flos-cuculi, nous nous attendions à obtenir également
des écarts sensibles en matière de performance entre les
plantes. Mais ce ne fût pas le cas. Ni la consanguinité ni
la composition génétique différente n’a eu d’effet signi-
ficatif sur la performance des plantes dans notre expéri-
mentation, ni en plein champ, ni dans le jardin d’essai, ni
dans les chambres climatiques. Malgré cela, nous avons
obtenu certains résultats intéressants. Les plantes issues
de semences de populations semées ou de semences
obtenues chez les producteurs ont fleuri moins fréquem-
ment que les plantes issues de populations naturelles
(fig. 5; Aavik et al. en impression). Différentes explica-
tions de ce phénomène sont possibles. D’une part, les
stations des populations naturelles étaient plutôt
humides et riches, tandis que celles des populations
semées étaient plutôt sèches et peu riches. Des études
antérieures ont montré que la reproduction sur des sta-
Figure 5 | Proportion de plantes avec floraison (bleu) et sans floraison (blanc) de la silène fleur de coucou (Lychnis flos-cuculi) dans l’essai en chambre climatique (GLMM, X2=38,87, P < 0,001; Aavik et al. en impression). Les plantes sont issues de semences de populations natu-relles, de populations semées et de mélanges obtenus directement chez les producteurs.
0
20
40
60
80
100
Naturel I Naturel II Semé I Semé II Producteur I Producteur II
Prop
ortio
n de
pla
ntes
(%)
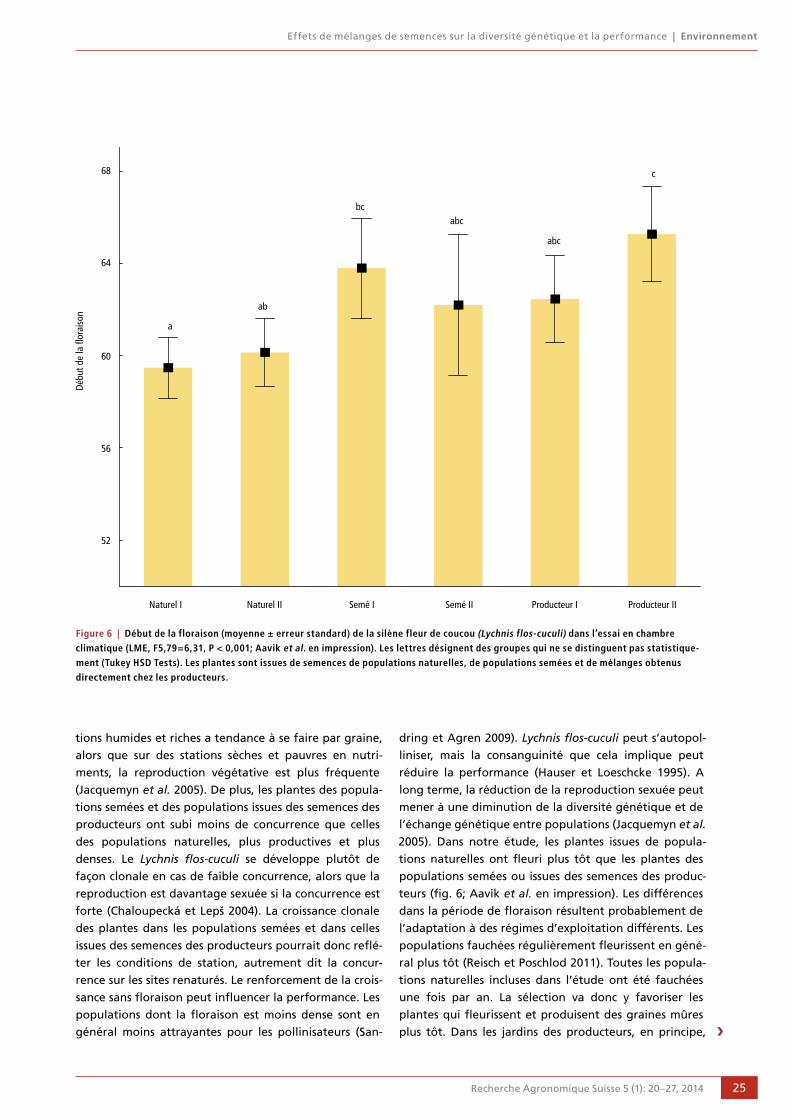
Effets de mélanges de semences sur la diversité génétique et la performance | Environnement
25Recherche Agronomique Suisse 5 (1): 20–27, 2014
dring et Agren 2009). Lychnis flos-cuculi peut s’autopol-
liniser, mais la consanguinité que cela implique peut
réduire la performance (Hauser et Loeschcke 1995). A
long terme, la réduction de la reproduction sexuée peut
mener à une diminution de la diversité génétique et de
l’échange génétique entre populations (Jacquemyn et al.
2005). Dans notre étude, les plantes issues de popula-
tions naturelles ont fleuri plus tôt que les plantes des
populations semées ou issues des semences des produc-
teurs (fig. 6; Aavik et al. en impression). Les différences
dans la période de floraison résultent probablement de
l’adaptation à des régimes d’exploitation différents. Les
populations fauchées régulièrement fleurissent en géné-
ral plus tôt (Reisch et Poschlod 2011). Toutes les popula-
tions naturelles incluses dans l’étude ont été fauchées
une fois par an. La sélection va donc y favoriser les
plantes qui fleurissent et produisent des graines mûres
plus tôt. Dans les jardins des producteurs, en principe,
tions humides et riches a tendance à se faire par graine,
alors que sur des stations sèches et pauvres en nutri-
ments, la reproduction végétative est plus fréquente
(Jacquemyn et al. 2005). De plus, les plantes des popula-
tions semées et des populations issues des semences des
producteurs ont subi moins de concurrence que celles
des populations naturelles, plus productives et plus
denses. Le Lychnis flos-cuculi se développe plutôt de
façon clonale en cas de faible concurrence, alors que la
reproduction est davantage sexuée si la concurrence est
forte (Chaloupecká et Lepš 2004). La croissance clonale
des plantes dans les populations semées et dans celles
issues des semences des producteurs pourrait donc reflé-
ter les conditions de station, autrement dit la concur-
rence sur les sites renaturés. Le renforcement de la crois-
sance sans floraison peut influencer la performance. Les
populations dont la floraison est moins dense sont en
général moins attrayantes pour les pollinisateurs (San-
Figure 6 | Début de la floraison (moyenne ± erreur standard) de la silène fleur de coucou (Lychnis flos-cuculi) dans l’essai en chambre climatique (LME, F5,79=6,31, P < 0,001; Aavik et al. en impression). Les lettres désignent des groupes qui ne se distinguent pas statistique-ment (Tukey HSD Tests). Les plantes sont issues de semences de populations naturelles, de populations semées et de mélanges obtenus directement chez les producteurs.
Naturel I Naturel II Semé I Semé II Producteur I Producteur II
52
56
60
64
68
Débu
t de
la fl
orai
son
a
ab
bcabc
abc
c

26
Environnement | Effets de mélanges de semences sur la diversité génétique et la performance
Recherche Agronomique Suisse 5 (1): 20–27, 2014
Bibliographie ▪ Aavik T., Edwards P., Holderegger R., Graf R. & Billeter R., 2012. Genetic consequences of using seed mixtures in restoration: a case study of a wetland plant Lychnis flos-cuculi. Biological Conservation 145, 195–204.
▪ Aavik T., Holderegger R., Edwards P. J. & Billeter R., 2013. Patterns of contemporary gene flow suggest low functional connectivity of grass-lands in a fragmented agricultural landscape. Journal of Applied Ecology 50, 395–403.
▪ Aavik T., Bosshard D., Edwards P., Holderegger R. & Billeter R., en impression. Fitness in naturally occurring and restored populations of a grassland plant Lychnis flos-cuculi in a Swiss agricultural landscape. Restoration Ecology.
▪ Billeter R., Liira J., Bailey D., Bugter R., Arens P., Augenstein I., Aviron S., Baudry J., Bukacek R., Burel F., Cerny M., De Blust G., De Cock R., Dieköt-ter T., Dietz H., Dirksen J., Dormann C. F., Durka W., Frenzel M., Hamer-sky R., Hendrickx F., Herzog F., Klotz S., Koolstra B., Lausch A., Le Coeur D., Maelfait J. P., Opdam P., Roubalova M., Schermann A., Schermann N., Schmidt T., Schweiger O., Smulders M. J. M., Speelmans M., Simova P., Verboom J., van Wingerden W., Zobel M. & Edwards P. J., 2008. Indica-tors for biodiversity in agricultural landscapes: a pan-European study. Journal of Applied Ecology 45, 141–150.
▪ Bischoff A., Vonlanthen B., Steinger T. & Müller-Schärer H., 2006. Seed provenance matters – effects on germination of four plant species used for ecological restoration. Basic and Applied Ecology 7, 347–359.
▪ Chaloupecká E. & Lepš J., 2004. Equivalence of competitor effects and tra-deoff between vegetative multiplication and generative reproduction: case study with Lychnis flos-cuculi and Myosotis nemorosa. Flora 199, 157–167.
▪ Goudet J., 1995. FSTAT (version 1.2): a computer program to calculate F-statistics. Journal of Heredity 86, 485–486.
▪ Hauser T. P. & Loeschcke V., 1995. Inbreeding depression in Lychnis flos-cuculi (Caryophyllaceae): effects of different levels of inbreeding. Journal of Evolutionary Biology 8, 589–600.
▪ Jacquemyn H., Brys R., Honnay O., Hermy M. & Roldan-Ruiz I., 2005. Local forest environment largely affects below-ground growth, clonal diversity and fine-scale spatial genetic structure in the temperate deci-duous forest herb Paris quadrifolia. Molecular Ecology 14, 4479–4488.
▪ Leng X., Musters C. J. M. & de Snoo G. R., 2011. Effects of mowing date on the opportunities of seed dispersal of ditch bank plant species under diffe-rent management regimes. Journal for Nature Conservation 19, 166–174.
▪ Pritchard J. K., Stephens M. & Donnelly P., 2000. Inference of Population Structure Using Multilocus Genotype Data. Genetics 155, 945–959.
▪ R Development Core Team (2011) R: a language and environment for statistical computing. http://www.r-project.org.
▪ Reisch C. & Poschlod P., 2011. Morphology and phenology of Scabiosa columbaria from mown and grazed habitats – results of a simulation experiment. Flora 206, 887–891.
▪ Sandring S. & Agren J., 2009. Pollinator-mediated selection on floral display and flowering time in the perennial herb Arabidopsis lyrata. Evolution 63, 1292–1300.
▪ Schoen D. J. & Brown A. H. D., 2001. The conservation of wild plant spe-cies in seed banks. BioScience 51, 960–966.
▪ Vander Mijnsbrugge K., Bischoff A. & Smith B., 2010. A question of origin: where and how to collect seed for ecological restoration. Basic and Applied Ecology 11, 300–311.
cette sélection n’existe pas. La floraison plus tardive des
populations semées peut cependant influencer le succès
des valorisations écologiques. Les prairies extensives des
surfaces de compensation peuvent être fauchées à par-
tir du 15 juin. Cette date ne correspond pas à la période
de production maximale des graines de L. flos-cuculi
(Leng et al. 2011). En Haute-Argovie, on fauche souvent
les bords de prairies jouxtant des fossés encore plus tôt,
afin de créer des conditions favorables à une espèce de
libellule menacée. Cela peut gêner la reproduction de
plantes semées, dont les graines mûrissent plus tard que
celles des populations naturelles. Des périodes de florai-
son variables rendent les échanges génétiques par le
pollen plus difficiles entre populations naturelles et
semées. Une analyse en Haute-Argovie a en effet révélé
que ce taux d’échange pour L. flos-cuculi était faible
(Aavik et al. 2013).
C o n c l u s i o n s
Cette étude montre que les populations issues de
mélanges de semences commercialisées sont génétique-
ment différentes des populations naturelles. Cela pour-
rait être évité si les graines d’origine utilisées pour pro-
duire ces mélanges de semences étaient issues de
grandes populations et prélevées sur de nombreux indi-
vidus de la même région. Le stock de graines utilisé pour
la reproduction doit aussi être renouvelé régulièrement.
Par exemple, les différences de périodes de floraison
entre des populations semées et naturelles indiquent
que le matériel des producteurs de semences provient à
l’origine de populations soumises à d’autres conditions
de station et de concurrence ainsi qu’à un autre régime
d’exploitation. Cependant, les différences mentionnées
n’ont exercé jusqu’à présent aucun effet sur la perfor-
mance des plantes. Ceci montre que l’ensemencement à
l’aide de mélanges de semences est une méthode parti-
culièrement adéquate lorsque la production des
semences repose sur une base génétique suffisamment
large. Dans ces conditions, on ne peut guère s’attendre à
des effets négatifs sur la performance. Lorsque c’est pos-
sible, il faut cependant préférer la colonisation sponta-
née à partir de l’environnement naturel. n
Remerciements
Ce travail a été soutenu financièrement par le «Competence Centre Environment and Sustainability» (CCES; projets ENHANCE et GENEREACH) et par l’Union euro-péenne (T. A. Mobilitas subvention MJD113).

27
Effets de mélanges de semences sur la diversité génétique et la performance | Environnement
Ria
ssu
nto
Sum
mar
y
Recherche Agronomique Suisse 5 (1): 20–27, 2014
Genetic and fitness consequences of using
wildflower seed mixtures in ecological
restoration
Wildflower seed mixtures are widely used
for restoration in areas with impoverished
species pools. However, the genetic and
fitness consequences of using seed
mixtures are often not considered in
practical restoration. We studied the
genetic characteristics of sown and
naturally occurring populations of the
grassland plant Lychnis flos-cuculi in an
agricultural landscape in the Oberaargau
region in Switzerland. Furthermore, we
examined various fitness parameters of
these populations, and carried out
experiments in the study area, in an
experimental garden and in climate
chambers, in order to study the effect of
genetic diversity, origin and environmen-
tal conditions on the viability of plants.
Sown and natural populations were
characterized by similar genetic diversity.
Inbreeding coefficients, by contrast, were
significantly higher in sown populations.
Sown populations were genetically
different from natural populations.
Experiments revealed that plants originat-
ing from sown populations and from seed
companies were less likely to produce
flowers and tended to flower later than
plants from natural populations. We
conclude that there was no substantial
influence of origin and genetic diversity
on plant fitness. However, seed mixtures
used for restoration should originate from
genetically diverse sources to avoid
potential negative consequences for
fitness. Wherever possible, natural
recolonization should be favoured.
Key words: ecological compensation areas,
genetic diversity, grasslands, inbreeding,
seed mixtures.
Diversità genetica nelle miscele di semente
di piante selvatiche
Miscele di semente di piante selvatiche
sono spesso usate per la rinaturalizzazione
di superfici, in particolare allo scopo di
aumentare la biodiversità in aree povere
di specie. Tuttavia, le conseguenze di simili
miscele per la diversità genetica e la
«fitness» sono spesso poco chiare.
Attraverso questo studio abbiamo
analizzato la diversità genetica delle
popolazioni naturali e di quelle seminate
del fior di cuculo (Lychnis flos-cuculi) nella
regione dell’Argovia superiore nell’Alti-
piano svizzero. Mediante prove in campo,
vivaio e in camere climatiche abbiamo
misurato gli effetti che diversità genetica,
provenienza e condizioni climatiche
esercitano sulla crescita e la capacità di
sopravvivenza delle piante. La diversità
genetica delle popolazioni naturali e
seminate era molto simile, ma le popola-
zioni seminate presentavano una consan-
guineità significativamente più elevata.
Inoltre, anche nella loro composizione
genetica, le popolazioni naturali si
differenziavano notevolmente da quelle
seminate. Le nostre prove hanno mostrato
che piante seminate, così come piante
ottenute da semente acquistata presso
produttori, fioriscono meno frequente-
mente e, tendono a fiorire più tardi
rispetto alle piante di popolazioni naturali.
La provenienza e la diversità genetica non
hanno, percontro, praticamente nessun
influsso sulla «fitness» delle piante. Ne
deduciamo che la semina con miscele di
sememente rappresenta una misura
sensata soprattutto quando la produzione
dei semi avviene su una base genetica
sufficientemente ampia. In quel caso non
ci si deve aspettare effetti negativi sulla
«fitness». Quando possibile dovrebbe
essere favorito l’insediamento naturale
proveniente dalle zone limitrofe.

28 Recherche Agronomique Suisse 5 (1): 28–31, 2014
graissement de l‘entreprise UFA SA (Sursee) ont été
moulus avec un broyeur à marteaux (fin) ou un moulin à
cylindres (grossier). La moitié des animaux a reçu à
chaque fois l’aliment avec des CB fins ou grossiers. Les
deux groupes ont été transformés sans répétition dans le
processus normal d’abattage et de découpe de Bell SA à
Zell. Le 28e et le 36e jour d’essai, la qualité de la litière a
été évaluée visuellement sur la base de la part de croû-
tage en pourcent et du degré d’humidité. Le 28e et le 35e
jour, la santé de la plante des pieds et des talons de dix
animaux par compartiment a été évaluée d’après la
méthode d‘Ekstrand et al. (1997). Le jour précédant
l’abattage, le gésier et l’intestin de 20 animaux par pro-
cédé d’alimentation ont été pesés par section à Avifo-
rum (tabl. 3).
Analyse par tamisage
Une tamiseuse électromagnétique à mouvement tridi-
mensionnel et mailles de 2 mm, 1 mm et 0,5 mm de
Haver & Boecker a été utilisée pour analyser la taille des
particules dans l’aliment (fig. 2). Le réglage du temps de
tamisage optimal (une minute) et de l’amplitude d’oscil-
lation (0,7) a été calculé d’après Retsch® (2004). Le dia-
mètre géométrique moyen (DGM) des particules des
deux aliments a été calculé sur la base de la part d’ali-
ment par taille de maille du tamis (racine énième du pro-
duit des valeurs n) pour pouvoir comparer les résultats
des essais avec la littérature (fig. 3).
La taille des particules dans les granulés de l'aliment
influence les performances d’engraissement des poulets
de chair. La taille doit donc être choisie de manière à
avoir une bonne qualité de granulés sans prétériter les
performances. Un essai a été mené à Aviforum pour étu-
dier l’impact de la taille des composants bruts (CB) maïs,
blé et tourteau d’extraction de soja d’un aliment sous
forme de granulés de 3,5 mm sur les performances d’en-
graissement de poulets de chair à croissance rapide.
M a t é r i e l e t m é t h o d e s
Au cours de deux essais menés chaque fois avec 5600
poussins Ross PM3 et Ross 308 de sexes mélangés (as hat-
ched), les animaux ont été répartis au hasard dans
vingt compartiments (fig. 1). La litière était composée de
granulés de farine de paille, à raison de 1,2 kg par m2. La
garde s’est faite dans les conditions habituelles prati-
quées en Suisse. La durée d’engraissement était de
36 jours. Les CB maïs, blé et tourteau d’extraction de soja
utilisés dans l’aliment de démarrage et l’aliment d’en-
Poulets de chair: la taille des particules de l'aliment influence les accroissementsDanielle Albiker et Ruedi Zweifel
Aviforum, 3052 Zollikofen, Suisse
Renseignements: Danielle Albiker, e-mail: [email protected]
E c l a i r a g e
Figure 1 | Installation d’essai dans le poulailler d’engraissement avec vingt compartiments de 280 animaux chacun. (Photo: Aviforum)
Figure 2 | Tamiseuse de Haver & Boecker. (Photo: Aviforum)
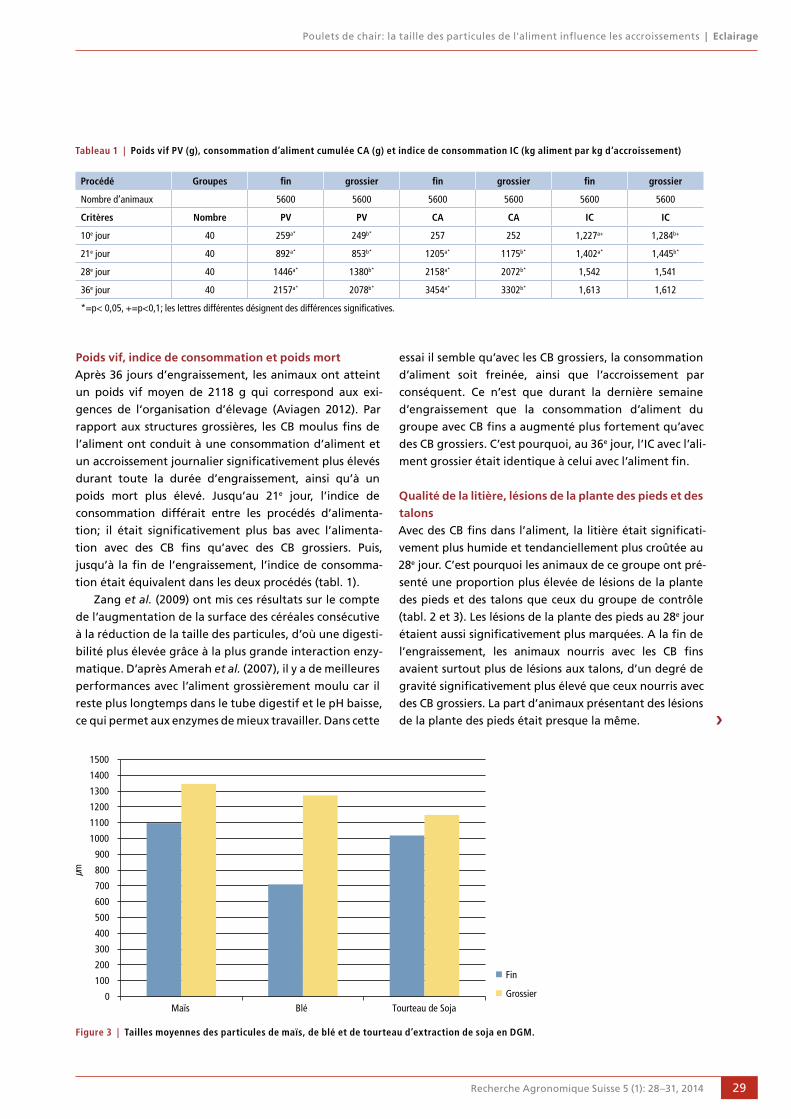
Poulets de chair: la taille des particules de l'aliment influence les accroissements | Eclairage
29Recherche Agronomique Suisse 5 (1): 28–31, 2014
Poids vif, indice de consommation et poids mort
Après 36 jours d’engraissement, les animaux ont atteint
un poids vif moyen de 2118 g qui correspond aux exi-
gences de l‘organisation d’élevage (Aviagen 2012). Par
rapport aux structures grossières, les CB moulus fins de
l’aliment ont conduit à une consommation d’aliment et
un accroissement journalier significativement plus élevés
durant toute la durée d’engraissement, ainsi qu’à un
poids mort plus élevé. Jusqu’au 21e jour, l’indice de
consommation différait entre les procédés d’alimenta-
tion; il était significativement plus bas avec l’alimenta-
tion avec des CB fins qu’avec des CB grossiers. Puis,
jusqu’à la fin de l’engraissement, l’indice de consomma-
tion était équivalent dans les deux procédés (tabl. 1).
Zang et al. (2009) ont mis ces résultats sur le compte
de l’augmentation de la surface des céréales consécutive
à la réduction de la taille des particules, d’où une digesti-
bilité plus élevée grâce à la plus grande interaction enzy-
matique. D’après Amerah et al. (2007), il y a de meilleures
performances avec l’aliment grossièrement moulu car il
reste plus longtemps dans le tube digestif et le pH baisse,
ce qui permet aux enzymes de mieux travailler. Dans cette
essai il semble qu’avec les CB grossiers, la consommation
d’aliment soit freinée, ainsi que l’accroissement par
conséquent. Ce n’est que durant la dernière semaine
d’engraissement que la consommation d’aliment du
groupe avec CB fins a augmenté plus fortement qu’avec
des CB grossiers. C’est pourquoi, au 36e jour, l’IC avec l’ali-
ment grossier était identique à celui avec l’aliment fin.
Qualité de la litière, lésions de la plante des pieds et des
talons
Avec des CB fins dans l’aliment, la litière était significati-
vement plus humide et tendanciellement plus croûtée au
28e jour. C’est pourquoi les animaux de ce groupe ont pré-
senté une proportion plus élevée de lésions de la plante
des pieds et des talons que ceux du groupe de contrôle
(tabl. 2 et 3). Les lésions de la plante des pieds au 28e jour
étaient aussi significativement plus marquées. A la fin de
l’engraissement, les animaux nourris avec les CB fins
avaient surtout plus de lésions aux talons, d’un degré de
gravité significativement plus élevé que ceux nourris avec
des CB grossiers. La part d’animaux présentant des lésions
de la plante des pieds était presque la même.
Procédé Groupes fin grossier fin grossier fin grossier
Nombre d‘animaux 5600 5600 5600 5600 5600 5600
Critères Nombre PV PV CA CA IC IC
10e jour 40 259a* 249b* 257 252 1,227a+ 1,284b+
21e jour 40 892a* 853b* 1205a* 1175b* 1,402a* 1,445b*
28e jour 40 1446a* 1380b* 2158a* 2072b* 1,542 1,541
36e jour 40 2157a* 2078b* 3454a* 3302b* 1,613 1,612
*=p< 0,05, +=p<0,1; les lettres différentes désignent des différences significatives.
Tableau 1 | Poids vif PV (g), consommation d’aliment cumulée CA (g) et indice de consommation IC (kg aliment par kg d‘accroissement)
0
100
200
300
400
500
600
700
800
900
1000
1100
1200
1300
1400
1500
Maïs Blé Tourteau de Soja
µm
Fin
Grossier
Figure 3 | Tailles moyennes des particules de maïs, de blé et de tourteau d’extraction de soja en DGM.
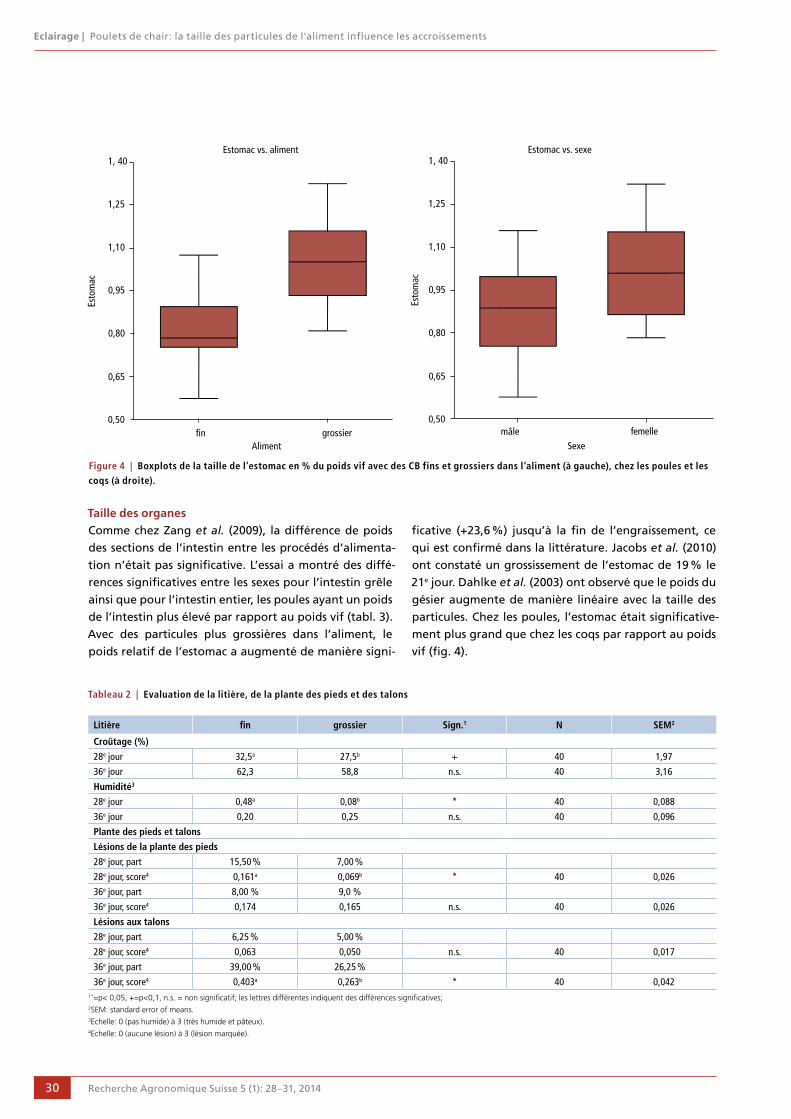
Eclairage | Poulets de chair: la taille des particules de l'aliment influence les accroissements
30 Recherche Agronomique Suisse 5 (1): 28–31, 2014
Taille des organes
Comme chez Zang et al. (2009), la différence de poids
des sections de l’intestin entre les procédés d‘alimenta-
tion n’était pas significative. L’essai a montré des diffé-
rences significatives entre les sexes pour l’intestin grêle
ainsi que pour l’intestin entier, les poules ayant un poids
de l’intestin plus élevé par rapport au poids vif (tabl. 3).
Avec des particules plus grossières dans l’aliment, le
poids relatif de l’estomac a augmenté de manière signi-
ficative (+23,6 %) jusqu’à la fin de l’engraissement, ce
qui est confirmé dans la littérature. Jacobs et al. (2010)
ont constaté un grossissement de l’estomac de 19 % le
21e jour. Dahlke et al. (2003) ont observé que le poids du
gésier augmente de manière linéaire avec la taille des
particules. Chez les poules, l’estomac était significative-
ment plus grand que chez les coqs par rapport au poids
vif (fig. 4).
fin grossier0,50
1, 40
1,25
1,10
0,95
0,80
0,65
Aliment
Estomac vs. aliment
Esto
mac
mâle femelleSexe
Estomac vs. sexe
Esto
mac
0,50
1, 40
1,25
1,10
0,95
0,80
0,65
Figure 4 | Boxplots de la taille de l’estomac en % du poids vif avec des CB fins et grossiers dans l‘aliment (à gauche), chez les poules et les coqs (à droite).
Litière fin grossier Sign.1 N SEM2
Croûtage (%)
28e jour 32,5a 27,5b + 40 1,97
36e jour 62,3 58,8 n.s. 40 3,16
Humidité3
28e jour 0,48a 0,08b * 40 0,088
36e jour 0,20 0,25 n.s. 40 0,096
Plante des pieds et talons
Lésions de la plante des pieds
28e jour, part 15,50 % 7,00 %
28e jour, score4 0,161a 0,069b * 40 0,026
36e jour, part 8,00 % 9,0 %
36e jour, score4 0,174 0,165 n.s. 40 0,026
Lésions aux talons
28e jour, part 6,25 % 5,00 %
28e jour, score4 0,063 0,050 n.s. 40 0,017
36e jour, part 39,00 % 26,25 %
36e jour, score4 0,403a 0,263b * 40 0,0421*=p< 0,05, +=p<0,1, n.s. = non significatif; les lettres différentes indiquent des différences significatives; 2SEM: standard error of means.3Echelle: 0 (pas humide) à 3 (très humide et pâteux).4Echelle: 0 (aucune lésion) à 3 (lésion marquée).
Tableau 2 | Evaluation de la litière, de la plante des pieds et des talons
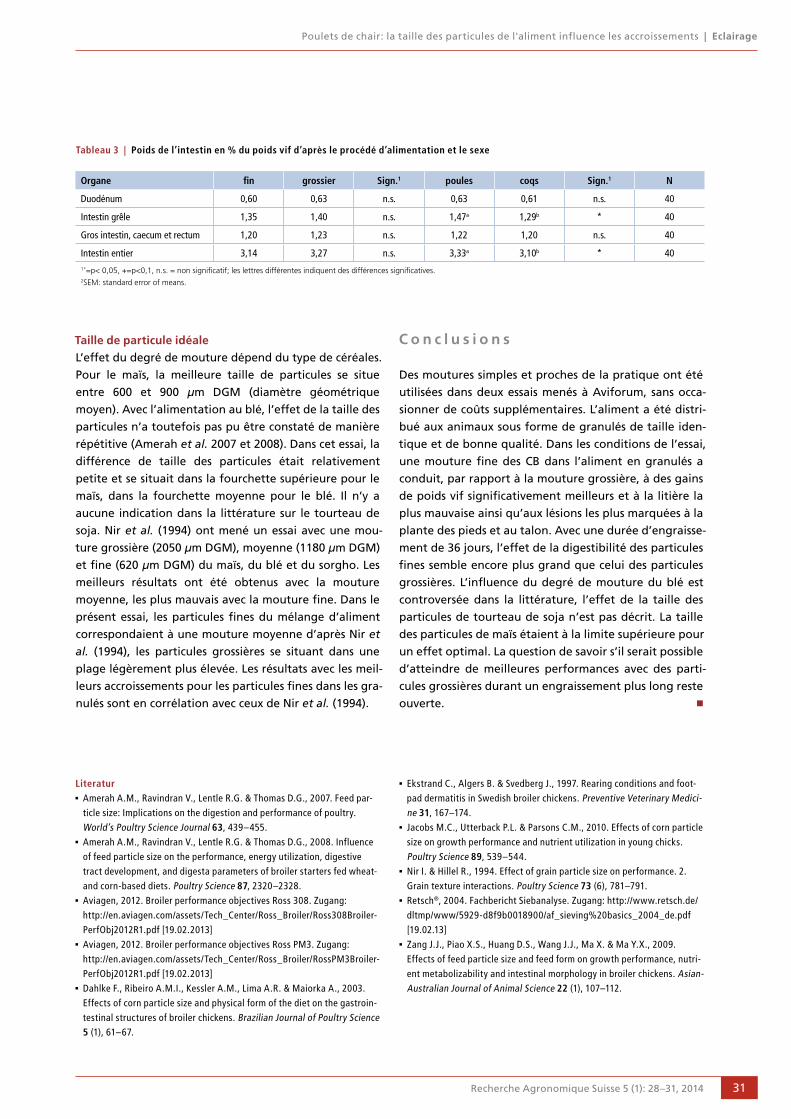
Poulets de chair: la taille des particules de l'aliment influence les accroissements | Eclairage
31Recherche Agronomique Suisse 5 (1): 28–31, 2014
C o n c l u s i o n s
Des moutures simples et proches de la pratique ont été
utilisées dans deux essais menés à Aviforum, sans occa-
sionner de coûts supplémentaires. L’aliment a été distri-
bué aux animaux sous forme de granulés de taille iden-
tique et de bonne qualité. Dans les conditions de l’essai,
une mouture fine des CB dans l’aliment en granulés a
conduit, par rapport à la mouture grossière, à des gains
de poids vif significativement meilleurs et à la litière la
plus mauvaise ainsi qu’aux lésions les plus marquées à la
plante des pieds et au talon. Avec une durée d’engraisse-
ment de 36 jours, l’effet de la digestibilité des particules
fines semble encore plus grand que celui des particules
grossières. L’influence du degré de mouture du blé est
controversée dans la littérature, l’effet de la taille des
particules de tourteau de soja n’est pas décrit. La taille
des particules de maïs étaient à la limite supérieure pour
un effet optimal. La question de savoir s’il serait possible
d’atteindre de meilleures performances avec des parti-
cules grossières durant un engraissement plus long reste
ouverte. n
Taille de particule idéaleL’effet du degré de mouture dépend du type de céréales.
Pour le maïs, la meilleure taille de particules se situe
entre 600 et 900 µm DGM (diamètre géométrique
moyen). Avec l’alimentation au blé, l’effet de la taille des
particules n’a toutefois pas pu être constaté de manière
répétitive (Amerah et al. 2007 et 2008). Dans cet essai, la
différence de taille des particules était relativement
petite et se situait dans la fourchette supérieure pour le
maïs, dans la fourchette moyenne pour le blé. Il n’y a
aucune indication dans la littérature sur le tourteau de
soja. Nir et al. (1994) ont mené un essai avec une mou-
ture grossière (2050 µm DGM), moyenne (1180 µm DGM)
et fine (620 µm DGM) du maïs, du blé et du sorgho. Les
meilleurs résultats ont été obtenus avec la mouture
moyenne, les plus mauvais avec la mouture fine. Dans le
présent essai, les particules fines du mélange d’aliment
correspondaient à une mouture moyenne d’après Nir et
al. (1994), les particules grossières se situant dans une
plage légèrement plus élevée. Les résultats avec les meil-
leurs accroissements pour les particules fines dans les gra-
nulés sont en corrélation avec ceux de Nir et al. (1994).
Literatur ▪ Amerah A.M., Ravindran V., Lentle R.G. & Thomas D.G., 2007. Feed par-ticle size: Implications on the digestion and performance of poultry. World’s Poultry Science Journal 63, 439–455.
▪ Amerah A.M., Ravindran V., Lentle R.G. & Thomas D.G., 2008. Influence of feed particle size on the performance, energy utilization, digestive tract development, and digesta parameters of broiler starters fed wheat- and corn-based diets. Poultry Science 87, 2320–2328.
▪ Aviagen, 2012. Broiler performance objectives Ross 308. Zugang: http://en.aviagen.com/assets/Tech_Center/Ross_Broiler/Ross308Broiler-PerfObj2012R1.pdf [19.02.2013]
▪ Aviagen, 2012. Broiler performance objectives Ross PM3. Zugang: http://en.aviagen.com/assets/Tech_Center/Ross_Broiler/RossPM3Broiler-PerfObj2012R1.pdf [19.02.2013]
▪ Dahlke F., Ribeiro A.M.I., Kessler A.M., Lima A.R. & Maiorka A., 2003. Effects of corn particle size and physical form of the diet on the gastroin-testinal structures of broiler chickens. Brazilian Journal of Poultry Science 5 (1), 61–67.
▪ Ekstrand C., Algers B. & Svedberg J., 1997. Rearing conditions and foot-pad dermatitis in Swedish broiler chickens. Preventive Veterinary Medici-ne 31, 167–174.
▪ Jacobs M.C., Utterback P.L. & Parsons C.M., 2010. Effects of corn particle size on growth performance and nutrient utilization in young chicks. Poultry Science 89, 539–544.
▪ Nir I. & Hillel R., 1994. Effect of grain particle size on performance. 2. Grain texture interactions. Poultry Science 73 (6), 781–791.
▪ Retsch®, 2004. Fachbericht Siebanalyse. Zugang: http://www.retsch.de/dltmp/www/5929-d8f9b0018900/af_sieving%20basics_2004_de.pdf [19.02.13]
▪ Zang J.J., Piao X.S., Huang D.S., Wang J.J., Ma X. & Ma Y.X., 2009. Effects of feed particle size and feed form on growth performance, nutri-ent metabolizability and intestinal morphology in broiler chickens. Asian-Australian Journal of Animal Science 22 (1), 107–112.
Organe fin grossier Sign.1 poules coqs Sign.1 N
Duodénum 0,60 0,63 n.s. 0,63 0,61 n.s. 40
Intestin grêle 1,35 1,40 n.s. 1,47a 1,29b * 40
Gros intestin, caecum et rectum 1,20 1,23 n.s. 1,22 1,20 n.s. 40
Intestin entier 3,14 3,27 n.s. 3,33a 3,10b * 401*=p< 0,05, +=p<0,1, n.s. = non significatif; les lettres différentes indiquent des différences significatives. 2SEM: standard error of means.
Tableau 3 | Poids de l’intestin en % du poids vif d’après le procédé d’alimentation et le sexe

32 Recherche Agronomique Suisse 5 (1): 32–35, 2014
et porc), des grandes cultures et du lait; depuis 2012, il
s’étend à l’arboriculture fruitière et à la vigne. Agroscope,
par son groupe Extension fruits, s’est associée pour la
première fois en 2013 au réseau en lui fournissant des
données provenant de la production de pommes.
Le réseau agri benchmark compare la production agricole
de certains pays sous l’angle économique, en particulier
quant aux structures des coûts, aux rendements ou à la
rentabilité des systèmes de production. Fondé en 1997, le
réseau couvre les branches de la viande (bœuf, mouton
Réseau agri benchmark: comparaison dans le contexte internationalHildegard Garming1 et Esther Bravin2 1Thünen-Institut für Betriebswirtschaft, 38116 Braunschweig, Allemagne2Agroscope, Institut des sciences en production végétale IPV, 8020 Wädenswil, Suisse
Renseignements: Hildegard Garming, e-mail: [email protected]
Production de pommes en Thurgovie. (Photo: Adeline Kilchenmann, Agroscope)
E c l a i r a g e

33
Réseau agri benchmark: comparaison dans le contexte international | Eclairage
Recherche Agronomique Suisse 5 (1): 32–35, 2014
La production suisse de pommes
La Suisse produit annuellement quelque 120 000 à
165 000 tonnes de pommes sur 3900 ha de vergers (OFAG
2013). Les importations de pommes sont restées à un
niveau constant (env. 10 000 tonnes) au cours des vingt
dernières années (AFD 2011). La modicité relative des
importations (moins de 8 % de la production indigène)
tient vraisemblablement plus au système douanier qu’à
la qualité de la production locale ou à quelque avantage
de coûts du côté suisse. Le système des deux phases
régissant les importations de fruits frais a été introduit
en 1995; durant la phase administrée (15 juillet au
14 juin), les pommes de table sont protégées par un taux
de droit de douane hors contingent de CHF 153.– /100 kg
(Bitzer et al. 2012). Durant la période du 15 juin au
14 juillet, le taux de droit de douane applicable est de
CHF 2.– /100 kg et l’on trouve alors davantage de pommes
d’origine étrangère dans les magasins. La comparaison
des données fournies par la FAO montre que les prix à la
production en Suisse sont plus élevés d’au moins 25 %
que ceux pratiqués dans les principaux pays producteurs
de fruits tels l’Allemagne, l’Italie, la France et la Pologne
(Bravin et al. 2010).
La production mondiale de pommes s’est considéra-
blement développée au cours des cinq dernières années.
Certaines nations, telles la Chine et la Pologne, prennent
toujours plus de place parmi les «acteurs» et entrent en
concurrence avec les producteurs traditionnels de
pommes que sont par exemple la France ou l’Italie
(O’Rourke 2012). Dans notre voisinage immédiat, l’Italie
(Tyrol du sud) ou l’Allemagne doivent améliorer leur
capacité concurrentielle pour assurer à l’avenir la vente
de leur production à l’intérieur comme à l’extérieur de
leurs frontières. La consommation de pommes suit une
tendance fléchissante. Les consommateurs européens
ont découvert depuis longtemps les fruits en prove-
nance du Sud ainsi que d’autres manières de consommer
des fruits (O’Rourke 2012). En cas de libéralisation du
marché des pommes, ces tendances du marché global
des pommes exposeraient les producteurs suisses de
pommes à des difficultés supplémentaires.
Pourquoi le réseau agri benchmark?
Quelle est la situation de la production indigène dans le
contexte international? Cette question intéresse la pro-
duction aussi bien que la politique. L’analyse de la com-
pétitivité, basée sur les coûts de production ainsi que sur
les structures des exploitations et sur les systèmes de pro-
duction qui les sous-tendent, fournit d’importantes
informations pour discerner le potentiel d’améliorations.
En effet, les statistiques agricoles ne contiennent que
peu de données sur les systèmes et structures de produc-
tion. Les études consacrées dans différents pays aux
coûts de production sont rarement comparables entre
elles, par exemple en raison de différences méthodolo-
giques. D’autre part, les études ne proposent souvent
que des instantanés, plus rarement une évaluation sur le
long terme incluant les tendances.
Agri benchmark poursuit l’objectif de comparer des
systèmes de production agricole dans le monde entier
quant à leur rentabilité, aux forces agissant lors de leur
adaptation ainsi qu’à leurs perspectives. Divers instituts
scientifiques, organisations de conseil et partenaires
industriels de différents pays, coordonnés par l’Institut
d’économie d’entreprise allemand Thünen, ont consti-
tué des réseaux spécialisés par branche. Chacun de ces
réseaux s’occupe d’un secteur de la production agricole.
Ceux qui ont vu le jour jusqu’ici sont par exemple agri
benchmark Cash Crop (depuis 2003, 26 pays), agri bench-
mark Beef and Sheep (2001, 25 pays) et agri benchmark
Horticulture (depuis 2012, 8 pays).
Des exploitations typiques comme sources de donnéesLes données servant aux analyses sont celles d’exploita-
tions ou de modèles typiques d’exploitations comprenant
des paramètres techniques, physiques et économiques
représentatifs d’une région donnée de production à l’in-
térieur d’un pays. Ces modèles d’exploitations typiques
sont développés selon une méthode standardisée: sur la
base de données statistiques, on établit pour chaque
pays les principales régions de production, et à l’intérieur
de celles-ci les structures et grandeurs les plus courantes
parmi les entreprises. Les conseillers experts de la branche
concrétisent alors l’exploitation modèle avec des indica-
tions sur les surfaces, la main-d’oeuvre, les équipements
techniques et le système de production. Ce modèle est
alors adapté et validé dans des groupes de discussion
incluant des chefs d’exploitation.
Les données étant actualisées chaque année, il en
résulte des séries périodiques permettant une analyse
exhaustive de la situation économique des exploitations.
Ces relevés permettent aussi de repérer, plus rapide-
ment que dans les statistiques officielles, les nouveaux
développements intervenus dans la technologie de pro-
duction ou dans les marchés d’écoulement des produits.
Résultats récents d’agri benchmark Beef & Sheep et
Cash Crop
Exemple: Beef
La production de viande de bœuf est un exemple d’une
tendance globale que les statistiques officielles ne per-
mettent pas de distinguer. La tendance actuelle est aux
«feed lots», de grandes installations d’engraissement où
les bœufs en stabulation sont alimentés de fourrage
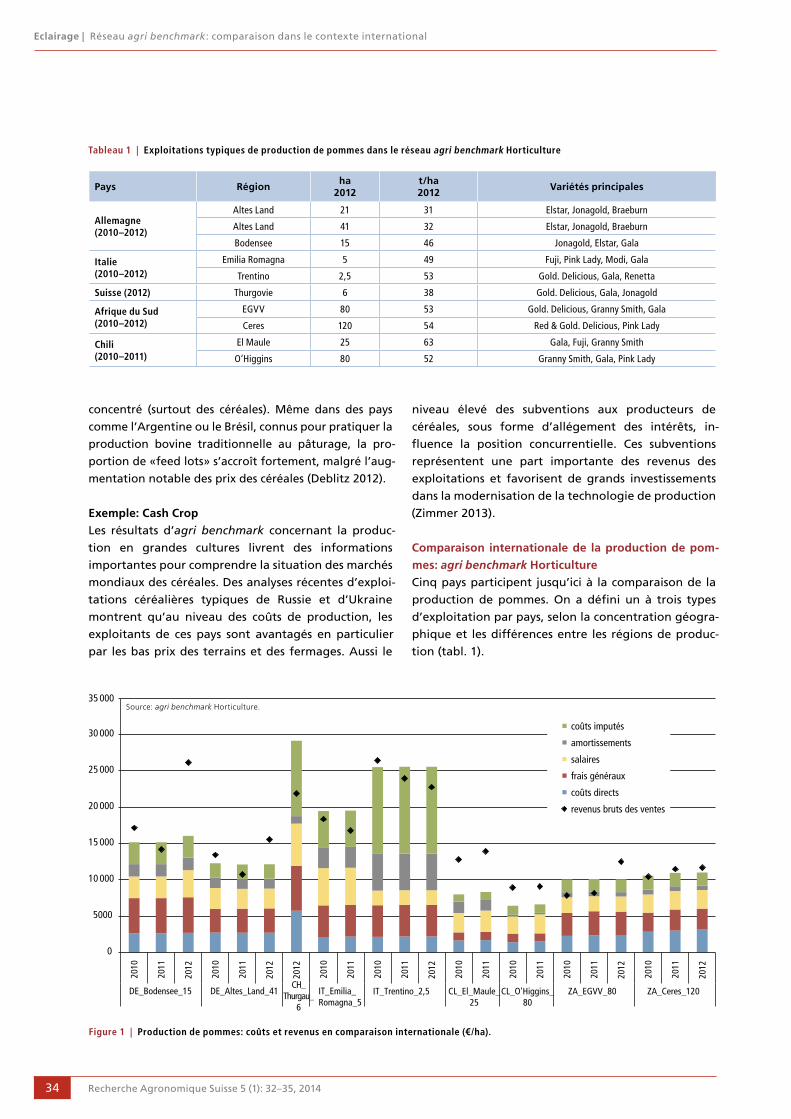
34
Eclairage | Réseau agri benchmark: comparaison dans le contexte international
Recherche Agronomique Suisse 5 (1): 32–35, 2014
concentré (surtout des céréales). Même dans des pays
comme l’Argentine ou le Brésil, connus pour pratiquer la
production bovine traditionnelle au pâturage, la pro-
portion de «feed lots» s’accroît fortement, malgré l’aug-
mentation notable des prix des céréales (Deblitz 2012).
Exemple: Cash Crop
Les résultats d’agri benchmark concernant la produc-
tion en grandes cultures livrent des informations
importantes pour comprendre la situation des marchés
mondiaux des céréales. Des analyses récentes d’exploi-
tations céréalières typiques de Russie et d’Ukraine
montrent qu’au niveau des coûts de production, les
exploitants de ces pays sont avantagés en particulier
par les bas prix des terrains et des fermages. Aussi le
niveau élevé des subventions aux producteurs de
céréales, sous forme d’allégement des intérêts, in-
fluence la position concurrentielle. Ces subventions
représentent une part importante des revenus des
exploitations et favorisent de grands investissements
dans la modernisation de la technologie de production
(Zimmer 2013).
Comparaison internationale de la production de pom-
mes: agri benchmark Horticulture
Cinq pays participent jusqu’ici à la comparaison de la
production de pommes. On a défini un à trois types
d’exploitation par pays, selon la concentration géogra-
phique et les différences entre les régions de produc-
tion (tabl. 1).
Pays Régionha
2012t/ha2012
Variétés principales
Allemagne(2010–2012)
Altes Land 21 31 Elstar, Jonagold, Braeburn
Altes Land 41 32 Elstar, Jonagold, Braeburn
Bodensee 15 46 Jonagold, Elstar, Gala
Italie (2010–2012)
Emilia Romagna 5 49 Fuji, Pink Lady, Modi, Gala
Trentino 2,5 53 Gold. Delicious, Gala, Renetta
Suisse (2012) Thurgovie 6 38 Gold. Delicious, Gala, Jonagold
Afrique du Sud (2010–2012)
EGVV 80 53 Gold. Delicious, Granny Smith, Gala
Ceres 120 54 Red & Gold. Delicious, Pink Lady
Chili (2010–2011)
El Maule 25 63 Gala, Fuji, Granny Smith
O‘Higgins 80 52 Granny Smith, Gala, Pink Lady
Tableau 1 | Exploitations typiques de production de pommes dans le réseau agri benchmark Horticulture
Figure 1 | Production de pommes: coûts et revenus en comparaison internationale (€/ha).
0
5000
10 000
15 000
20 000
25 000
30 000
35 000
2010
2011
2012
2010
2011
2012
2012
2010
2011
2010
2011
2012
2010
2011
2010
2011
2010
2011
2012
2010
2011
2012
DE_Bodensee_15 DE_Altes_Land_41CH_
Thurgau_6
IT_Emilia_Romagna_5
IT_Trentino_2,5 CL_El_Maule_25
CL_O'Higgins_80
ZA_EGVV_80 ZA_Ceres_120
coûts imputés
amortissements
salaires
frais généraux
coûts directs
revenus bruts des ventes
Source: agri benchmark Horticulture.

35
Réseau agri benchmark: comparaison dans le contexte international | Eclairage
Recherche Agronomique Suisse 5 (1): 32–35, 2014
par la fumure, la protection des plantes, l’irrigation et la
pépinière sont plutôt bas en comparaison, comme dans
la plupart des pays.
C o n c l u s i o n s
Le réseau agri benchmark offre à la branche suisse de la
production fruitière une belle opportunité d’analyser
objectivement sa situation et de discerner à temps les
actions à entreprendre. Pour d’autres produits agricoles
aussi, la comparaison de la capacité concurrentielle
pourrait être intéressante à l’avenir, surtout dans la pers-
pective de possibles modifications des conditions-cadres
de la politique agricole.
Pour d’autres informations concernant les différents
réseaux agri benchmark: www.agribenchmark.org n
Les exploitations typiques ne correspondent pas néces-
sairement à la moyenne statistique: il s’agit en effet de
représenter la plus grande partie possible de la produc-
tion de pommes du pays concerné; on considère à ce
titre des entreprises exploitées à temps plein, typiques
quant à la surface et aux équipements techniques.
Premiers résultats
La mise en valeur des résultats fournis par l’analyse des
exploitations typiques montre des différences notables
de coûts de production entre les pays européens d’une
part, le Chili et l’Afrique du Sud d’autre part. Si les coûts
de production sont bas dans ces derniers, les rendements
financiers bruts sont aussi relativement bas; ainsi, en
Afrique du Sud, les coûts totaux sont à peine couverts
par les revenus. Le Chili atteint la meilleure rentabilité
de tous les pays.
En Europe, les coûts de production sont les plus bas
en Allemagne (fig. 1). Cela tient principalement à la
taille des exploitations, plus grande que celle des exploi-
tations italiennes ou suisses. Les coûts des machines
(amortissements) et de la main-d’œuvre familiale (inclue
dans le calcul des coûts) sont divisés par une surface plus
grande. Cependant, comme les revenus bruts des ventes
sont très variables, la couverture des coûts n’est pas réa-
lisée chaque année.
C’est en Italie et en Suisse que les revenus à l’hectare
sont les plus élevés, dans les années ayant fait jusqu’ici
l’objet de relevés. En effet, les rendements en quantités
récoltées sont dans l’ensemble plus élevés qu’en Alle-
magne. Toutefois, les coûts imputés, incluant les coûts
de location des terrains en propriété de l’exploitant,
ceux du capital propre et des salaires de la main-d’œuvre
familiale, sont très élevés. Les coûts directs occasionnés
Bibliographie ▪ Bitzer A., Bregy G., Schuler R, 2012. Perspektiven für den Schweizer Apfel, Hochschule Luzern HSLU, Luzern.
▪ Office fédéral de l'agriculture (OFAG) 2013. Observation du marché / fruits et légumes, URL: http://www.blw.admin.ch/ [7.11.13], Berne.
▪ Bravin E., Kilchenmann A., 2010. Comparaison internationale de la production de pomme. Recherche Agronomie Suisse 1 (2), 52–59.
▪ Deblitz C., 2012. Feed lots: a new tendency in global beef production? Working Paper 2/2011, updated July 2012, agri benchmark Beef and Sheep, Braunschweig.
▪ Administration fédérale des douanes (AFD), 2011. Statistique suisse du commerce extérieur, Berne.
▪ O’Rourke D., 2012. The apple in the World. Vortrag an der Interpoma 2012, Bozen.
▪ Zimmer Y., 2013. Economics of Russian Grain Production – driven by low competition for land and strong political support. Pressemitteilung 28.07.2013, agri benchmark Cash Crop, Braunschweig.

36
P o r t r a i t
Recherche Agronomique Suisse 5 (1): 36, 2014
«Les compétences chez Agroscope sont impressionnantes»
Pour le chef Agroscope Michael Gysi, le plus grand défi
d’Agroscope consiste à augmenter encore la qualité de
la recherche et à renforcer le lien avec la pratique.
Quatre instituts de recherche sous une même bannière:
depuis le début de l’année, les trois anciennes stations
de recherche agronomique collaborent plus étroite-
ment. «Augmenter la reconnaissance auprès des par-
ties prenantes et des clients», tel est le souhait de
Michael Gysi. Et à la question «Quels sont selon vous les
défis les plus importants qu’Agroscope doit relever?»,
la réponse de Michael Gysi est précise: «Traiter les pôles
thématiques communs dans le cadre de la nouvelle
structure d’Agroscope avec quatre instituts, tout en
réalisant les objectifs du mandat de prestations; main-
tenir et développer le lien avec la pratique, augmenter
encore la qualité scientifique.» Un autre grand défi
consiste à créer une culture d’entreprise Agroscope. Et
Michael Gysi d’ajouter: «L’objectif est que tous les colla-
borateurs et collaboratrices d’Agroscope travaillent
avec encore davantage de fierté et d’enthousiasme
pour Agroscope».
En contact avec la recherche agronomique dès l’enfance
Né en 1968, Michael Gysi est pour ainsi dire tombé dans
la marmite de la recherche agronomique quand il était
petit: enfant, il allait rendre visite à son père sur son lieu
de travail, à la station de recherche de Wädenswil ZH.
Pendant sa thèse de doctorat à l’EPFZ, il découvre la FAT
de Tänikon TG, où il deviendra plus tard sous-directeur.
En 2006, il déménage avec femme et enfants à Berne
pour diriger la station de recherche ALP Liebefeld-
Posieux, qui deviendra plus tard ALP-Haras lors de l’inté-
gration du Haras national dans ALP en 2008. Aujourd’hui,
Michael Gysi est le chef Agroscope et dirige à ce titre la
totalité de l’infrastructure de la recherche agronomique
de la Confédération. «La diversité et les compétences de
l’ensemble des collaborateurs d’Agroscope sont impres-
sionnantes!», s’émerveille Michael Gysi. Il voit cepen-
dant encore un potentiel d’amélioration: «Les solutions
d’Agroscope pour résoudre les problèmes actuels,
comme le feu bactérien et ceux liés aux antibiotiques,
accroître la compétitivité de l’agriculture ou mettre un
frein à la mortalité des abeilles, doivent être à l’avenir
diffusées dans la pratique avec encore davantage d’effi-
cacité.»
Chef Agroscope, père de famille et alpiniste chevronné
Au service de la recherche agronomique, père de trois
enfants et passionné de montagne, autant dire que
l’agenda du chef Agroscope est bien rempli. Michael
Gysi souhaite cependant préserver un équilibre entre sa
vie professionnelle et sa vie privée: «Grâce à la souplesse
de la Confédération comme employeur et à des sup-
pléances adéquates, j’y parviens – et j’espère que, comme
moi, les collaborateurs d’Agroscope arrivent à concilier
vie de famille et vie professionnelle.» Et bien qu’il ne
désire pas s’épancher sur sa vie privée, il trouve qu’«aider
mes enfants à faire leurs devoirs est un gros challenge.»
Sans le «feu sacré» des collaborateurs, rien n’est possible
Ancien chercheur devenu manager, Michael Gysi
regrette-t-il l’époque où il faisait de la recherche? Le
chef Agroscope répond avec un petit sourire et deux
expressions françaises: «Non, je ne regrette rien» et le
«feu sacré». L’une des activités les plus agréables dans
son quotidien professionnel sont les rencontres avec les
collaborateurs sur leur lieu de travail. Il perçoit alors le
«feu sacré» qui les anime. Dans ces moments, il lui arrive
d’éprouver le désir de se remettre à faire quelques mani-
pulations en laboratoire ou, comme à l’époque de sa
thèse sur l’amélioration des sols, de la recherche.
Andrea Leuenberger-Minger, Recherche Agronomique Suisse
Christine Caron-Wickli, Agroscope
Michael Gysi, Chef Agroscope. (Photo: Carole Parodi)

37
A c t u a l i t é s
Recherche Agronomique Suisse 5 (1): 37–39, 2014
N o u v e l l e s p u b l i c a t i o n s
Rapport ART 765
Dans la détention des chèvres en stabulation libre, il
est courant d’intégrer des animaux étrangers dans des
groupes existants pour compléter ou agrandir l’effectif.
De telles arrivées conduisent à une augmentation des
affrontements agressifs et représentent un stress pour
les chèvres, qui se traduit par des perturbations de leur
comportement au repos et lors des repas. D’un autre
côté, le fait d’être séparé du troupeau est une source
de stress pour les chèvres qui sont des animaux sociables.
Cependant, il est parfois nécessaire de séparer provisoi-
rement certaines bêtes du reste du troupeau à l’occasion
de la mise bas ou pour cause de blessures. Par consé-
quent, on peut se demander comment réduire au maxi-
mum le stress causé par l’intégration d’une chèvre dans
un troupeau étranger, de même que par la séparation
d’un animal du reste du troupeau. Des essais réalisés à
la station de recherche Agroscope Reckenholz-Tänikon
ART ont étudié ces mesures de management sur des
chèvres avec et sans cornes. Comme les conflits sont plus
fréquents dans les petits groupes et, qu’en Suisse, les
chèvres sont essentiellement détenues en petits effec-
tifs, les études ont été effectuées sur de petits groupes.
Les résultats montrent que l’intégration d’une seule
chèvre dans un troupeau déjà formé représente une
source de stress importante et durable pour le nouvel
animal, indépendamment de la présence de cornes, et
qu’il est recommandé de l’éviter si possible. Il est donc
préférable d’intégrer simultanément dans un troupeau
plusieurs chèvres qui se connaissent, et de leur laisser
suffisamment de place à cette occasion, par exemple au
pâturage. Les chèvres ne devraient être séparées de leur
troupeau que dans des cas justifiés. Dans de tels cas, il est
recommandé de permettre aux chèvres d’avoir le plus de
contact possible avec leur troupeau. Le stress est réduit
lorsque les chèvres peuvent continuer à voir leur trou-
peau et avoir un contact avec leurs congénères à travers
la séparation des boxes.
Nina M. Keil et Antonia Patt, Office vétérinaire fédéral OVF, Centre
spécialisé dans la détention convenable des ruminants et des porcs,
ART
Impressum
Edition:Station de recherche AgroscopeReckenholz-Tänikon ART,Tänikon, CH-8356 Ettenhausen,Traduction:ART
Les Rapports ART paraissentenviron 20 fois par an.Abonnement annuel: Fr. 60.–.Commandes d‘abonnementset de numéros particuliers:ART,Bibliothèque, 8356 EttenhausenT +41 (0)52 368 31 31F +41 (0)52 365 11 [email protected]: www.agroscope.ch
ISSN 1661-7576
Rapport ART 765
Intégration et séparation des chèvres
Comment réduire le stress?
Autrices
Nina M. Keil et Antonia Patt,Office vétérinaire fédéralOVF, Centre spécialisé dansla détention convenable desruminants et des porcs,[email protected]
Juillet 2013
Dans la détention des chèvres en stabula-tion libre, il est courant d’intégrer des ani-maux étrangers dans des groupes exis-tants pour compléter ou agrandir l’effec-tif. De telles arrivées conduisent à uneaugmentation des affrontements agres-sifs et représentent un stress pour leschèvres qui se traduit par des perturba-tions de leur comportement au repos etlors des repas. D’un autre côté, le faitd’être séparé du troupeau est une sourcede stress pour les chèvres qui sont des ani-maux sociables. Cependant, il peut parfoisêtre nécessaire de séparer provisoirementcertaines bêtes du reste du troupeau àl’occasion de la mise bas ou pour cause deblessures. Par conséquent, on peut sedemander comment réduire au maximumle stress causé par l’intégration d’unechèvre dans un troupeau étranger, demême que par la séparation d’un animaldu reste du troupeau.Des essais réalisés à la station de rechercheAgroscope Reckenholz-Tänikon ART ontétudié ces mesures de management surdes chèvres avec et sans cornes. Comme
les conflits sont plus fréquents dans lespetits groupes et qu’en Suisse, les chèvressont essentiellement détenues en petitseffectifs, les études ont été effectuées surde petits groupes. Les résultats montrentque l’intégration d’une seule chèvre dansun troupeau déjà formé représente unesource de stress importante et durablepour le nouvel animal, indépendammentde la présence de cornes, et qu’il estrecommandé de l’éviter si possible. Parconséquent, il est préférable d’intégrersimultanément dans un troupeau plu-sieurs chèvres qui se connaissent, et deleur laisser suffisamment de place à cetteoccasion, par exemple au pâturage.
Les chèvres ne devraient être séparées deleur troupeau que dans des cas justifiés.Dans de tels cas, il est recommandé de per-mettre aux chèvres d’avoir le plus decontact possible avec leur troupeau. Lestress est réduit lorsque les chèvrespeuvent continuer à voir leur troupeau etavoir un contact avec leurs congénères àtravers la séparation des boxes.
Fig.1: Dans un troupeau stable, les chèvres se tolèrent les unes les autres et entretiennent descontacts positifs. Les animaux étrangers ne sont pas facilement acceptés dans le troupeau.
Intégration et séparation des chèvres

38
www.agroscope.admin.ch/medienmitteilungen
Actualités
Recherche Agronomique Suisse 5 (1): 37–39, 2014
N o u v e l l e s p u b l i c a t i o n s
C o m m u n i q u é s d e p r e s s e
9.12.2013 Un front international contre la tavelure du pommier La tavelure est la principale maladie fongique des
pommiers. Dans toutes les zones de production à prin-
temps humide, les traitements anti tavelure sont incon-
tournables, en production intégrée comme en culture
biologique. Les variétés de pommes pourvues de gènes
de résistance dépendent moins de la protection phyto-
sanitaire. Cependant, dans bien des régions, le cham-
pignon a contourné la résistance à la tavelure. Pour
faire face à ce problème, Agroscope a lancé l'initiative
internationale VINQUEST. Les spécialistes identifient
actuellement les gènes de résistance encore efficaces
et mettent leurs connaissances au service de la sélec-
tion variétale classique. L'objectif est de créer des varié-
tés de pommes munies d’une combinaison de facteurs
de résistance à la tavelure, qui possèdent ainsi une
résistance plus durable.
3.12.2013 Qualité du lait de chèvre et de brebis
Aujourd’hui, il n’existe pas de normes et de valeurs-
limites reconnues d’une manière générale au niveau
international pour contrôler et définir la qualité du lait
de chèvre et de brebis. Il est ainsi très difficile de définir
un paiement des producteurs selon des critères de qua-
lité. Agroscope a conçu un guide pour soutenir les spé-
cialistes lors de l’évaluation de la qualité.
18.11.2013 Qualité organoleptique et boulangère des varié-tés de blé suisses en culture biologique La sélection du blé tendre par Agroscope et Delley Semence
et plantes (DSP) est orientée vers une excellente qualité
boulangère. Depuis une dizaine d’années, la qualité bou-
langère et organoleptique des variétés, cultivées dans des
essais en petites parcelles selon les normes de production
biologique, est analysée au laboratoire d’Agroscope à
Changins. Ces analyses ont pour objectif de vérifier si les
variétés cultivées en mode bio, sans apport d’engrais azoté
de synthèse, gardent leur bonne qualité boulangère.
www.agroscope.admin.ch/communiques
«Avenir de l’économie
alpestre suisse. Faits, ana-
lyses et éléments de
réflexion issus du pro-
gramme de recherche Alp-
FUTUR» – Parution le 1er
octobre en allemand, en
été 2014 pour le français.
Année après année, des
milliers d’alpagistes con-
duisent leur bétail à l’al-
page. Depuis des siècles,
ils exploitent les pâturages alpestres d’altitude pour esti-
ver 100 000 vaches laitières, 35 000 vaches-mères, 180 000
génisses et 90 000 veaux ainsi que des moutons et des
chèvres et pour fabriquer des produits de qualité. Tradi-
tion et modernité se rejoignent, l’économie alpestre est
en constante évolution. Dans le cadre du programme de
recherche interdisciplinaire AlpFUTUR et de ses 22 pro-
jets, des chercheurs et des experts ont étudié l’avenir de
l’économie alpestre suisse sous toutes ses facettes. Alp-
FUTUR est coordonné par Agroscope et l’Institut fédéral
de recherches sur la forêt, la neige et le paysage WSL.
Le livre du programme de recherche fait la synthèse
d’AlpFUTUR. Il réunit les connaissances acquises en les
organisant par thème, de manière aisément compréhen-
sible et émet des recommandations clairement formu-
lées. Il s’adresse à tous ceux et celles qui s’intéressent à
l’économie alpestre et à son avenir, sur le plan profes-
sionnel ou privé.
Le livre est accompagné de films d’application Alp-
FUTUR – «D’alpagistes à alpagistes» et du documentaire
«Sommerzeit» en DVD. La publication des versions fran-
çaise et italienne est prévue pour le début de l’été 2014.
Le livre peut être commandé auprès de l’Institut fédéral
de recherches WSL pour Fr. 30.– (frais de port non com-
pris): www.alpfutur.ch/livre

39
Informationen: www.agroscope.admin.ch/veranstaltungen
Actualités
Recherche Agronomique Suisse 5 (1): 37–39, 2014
M a n i f e s t a t i o n s
Informations: www.agroscope.admin.ch/manifestations
L i e n s I n t e r n e t
2014 - Année internationale de l'agriculture familiale
www.familyfarming.ch
L’ONU a officiellement proclamé 2014 Année internatio-
nale de l’agriculture familiale (AIAF).
Différentes organisations qui vouent une partie
déterminante de leurs activités à la défense des revendi-
cations des exploitations paysannes familiales en Suisse
ou dans le monde se sont retrouvées en Suisse pour pré-
parer l'AIAF 2014.
Janvier 2014
18.01.2014Journée d’information HAFLHaute école des sciences agronomiques, forestières et alimentaires HAFLZollikofenInformations: www.hafl.bfh.ch
21. – 24.01.2014Agroscope à AgrovinaMartigny
23.01.2014Nachhaltigkeitstagung 2014«Wasser in der Landwirtschaft – heute und in Zukunft» AgroscopeAgroscope, 8046 Zürich
31.01.2014Pflanzenschutztagung Feldbau 2014Agroscope, 8046 Zürich
Février 2014
06. 02.20141ère Journée Nationale Grandes culturesAgroscope, le Forum Ackerbau, swissgranum, Agridea et PAG-CHInforama Rütti, 3052 Zollikofen
Mai 2014
06. – 07.05.2014Landtechnik im AlpenraumAgroscope et BLT WieselburgFeldkirch, Österreich
21.05.2014AgriMontana – Zukünftige Perspektiven der BerglandwirtschaftAgriMontana / AgroscopeLandquart
Février 2014 / Numéro 2
D a n s l e p r o c h a i n n u m é r o
La santé des abeilles est au coeur des préoccupations des chercheurs du Centre de recherche apicole (CRA). Un groupe d’experts, dont fait partie Peter Gallmann, ancien responsable du CRA, est en train d’élaborer un catalogue de mesures destinées à protéger la santé des abeilles dans notre pays. (Photo: BGD/SSA)
•• Femmes et hommes dans l’agriculture,
Esther Thalmann et al., Agridea,
•• Effets à long-terme d’une conversion à l’agriculture
biologique, Adrian Honegger et al., Agroscope
•• Estimation de la valeur nutritive d’ensilage de
mélanges protéagineux et céréales immatures,
Yves Arrigo, Agroscope
•• Série ProfiCrops: Agriculture urbaine: le projet FUI,
Katja Heitkämper et al., Agroscope et HAFL
•• Listes recommandées des variétés de soja et maïs
pour la récolte 2014

‣
Informations et inscription : hafl.bfh.ch
Journée d’information 18 janvier 2014
Etudes de bachelor :
Etudes de master :
– Agronomie – Foresterie – Technologie alimentaire
– Sciences appliquées agronomiques et forestières. Nouveau avec Major « Master alpin »loin
Voir
Jeudi 6 février 2014 (8h45 – 17h00)1ère Journée Nationale Grandes culturesOrganisée par Agroscope (Institut des sciences en production végétale),le Forum Ackerbau, swiss granum, AGRIDEA et la PAG-CH
Concilier économie et écologiedans les grandes culturesInforama Rütti, 3052 Zollikofen
ForumAckerbau
Objectif de la journéePar des exposés, un marché de l’information (posters) et une tableronde, échanger et discuter sur les possibilités actuelles et les pers-pectives pour concilier économie et écologie dans les grandescultures.
Le directeur de l’Office fédéral de l’agriculture, Bernard Lehmann,lancera les débats par un 1er exposé intitulé « Intensification durable– complexité et préconditions ». Des représentants de tous lesmilieux concernés prendront ensuite la parole.
PublicCette journée s’adresse à un large public: conseiller(ère)s etenseignant(e)s, chercheur(euse)s, représentant(e)s du commerceagricole, agriculteur(trice)s, membres de la PAG-CH, membres d’or-ganisations agricoles et services cantonaux, entrepreneurs(euses)agricoles et toutes les personnes intéressées par les grandescultures.Les présentations se feront soit en français, soit en allemand(ppt dans les 2 langues).
Programmewww.agridea-lausanne.ch ou www.agroscope.admin.ch/manifestations
Inscription (jusqu’au 30 janvier 2014)par e-mail [email protected] ou sur le site www.agridea-lausanne.ch





























