Embed Size (px)
Citation preview

MISE AU POINT DU DOSAGE
DE LA THYMULINE
‘PAR IMMUNONEPHELEMiZTRIE A
SUPPORTS MICROPARTICULAIRES
Christian CARLES *, Agnès GARTNER *, Paul MONTAGNE * * . 1
.
* Laboratoire de Nutrition Tropicale (L.N.T.) ORSTOM Montpellier
* * Laboratoire d’immunologie de la Faculté A de Médecioe, Nancy

Editions de I’ORSTOM / KK3T1;TUT FRANÇAIS DE RECHERCHE SCIEN.TlFIQUE
POUR LE DÉVELOPPEMENT EN COOPÉMTION
Favaux et documents microédités . . -
ISfjN 2-7099-0967-7
0 C ORSTOM .:
u La. loi du 11 mars 1957 n’autorisant, aux termes des alinéas 2 et 3 de l’anicI<
Y 41, d’une part, * que les utopies ou reproductions strictement réservées E
u 4. ‘.,. Pusqge priv6 du copiste et non destinées i une utilisarion collective, et, d’autrt
u part, que les analystes et les courtes citaSons dans un but d’exemple et d’illus ., (Y tration, utoute repr&sentation ou reproduclion intégrale, ou partielle, faite San!
p le ‘consentement de l’auteur ou de ses ayants droit ou ayants cause, est illicite>
a (alinéa”ler de l’article 40).
R Cette représemation ou reproduction, par quelque prochdè que ce soil.
a constituerait donc une contrefaçon sanctionnGe par les articles 425 et suivant!
a du Code Pbnala.


La mise au point du dosage de la thymuline s'inscrit dans le cadre du grand programme "Atteinte et restauration de la fonction immunitaire" au sein de 1' UR 6 "Maladies de dénutrition" du Département SANTE de 1'ORSTOM.
Ce sujet a été établi puis mené en collaboration avec le Laboratoire d'immunologie du Professeur J. DUHEILLE à la Faculté de Médecine de Nancy. Initiés par B. JAMBON les travaux ont été poursuivis à Nancy, jusqu'en janvier 1986, puis à Montpellier au Laboratoire de Nutrition ,- . Tropicale de ~IORSTOM (L.N.T.), par C. CARLES et A. GARTNER. Cette étude a toujours bénéficié de la collaboration scientifique de P. MONTAGNE et de toute l'équipe de Nancy.
La mise au point du dosage de la thymuline par une technique nouvelle et performante,, l'immunonéphélémétrie à supports microparticulaires, a bénéficié d'un financement du Ministère de la Recherche et de l'Industrie (d écisions numéros 82.L.1189 et 84.M.1121), de la Commission des Communautés Européennes (contrat numéro TSD.M.384.F) et de 1'ORSTOM.
3

SOMMAIRE
PRESENTATION DU PROJET
I- INTRODUCTION . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 9
II- LA THYMULINR . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 10
a'- Une hormone thymique ................................................ 11 a-l . Structure du thymus ...................................... 11 a-2 . Fonction du thymus ....................................... 12 a-3 . Les hormones thymiques ................................... -13
b- Activité biologique de la thymuline ; le test des rosettes .......... 14 c - Purification et caractérisation ..................................... 17 d- Essais de dosage de la thymuline .................................... 19 e - Etude de conformation en RMN-2D ..................................... 21
MISE AU POINT DU DOSAGE PAR IMMUNONEPHELEMETRIE A SUPPORTS MICROPARTICULAIRES
1 - PRINCIPES . . . . . . ..~............~..~..................~........... 25
a - Mesure néphélémétrigue .............................................. 25 b- Principe de la technique utilisée ................................... 28 c - Possibilités de couplage ............................................ 29
c-l . La thymuline ............................................. 29 c-2 . Les microsphères natives ................................. 32
d- Principe du dosage par inhibition ................................... 33 e - Caractéristiques et interprétation des réactions .................... 36 f- Protocole général de mise en réaction ............................... 38
II- MISE AU POINT DU DOSAGE EN MILIEU SYNTHETIQUE . . . . . . . . . . . . . . . . . . . 38
1 . PROTOCOLES . . . . . . . . . . . . . . . . . . . . . . ..~............................. 38
a - Obtention d'immunsérums . . . . . . . . . . . . . . ..I............................ 38
5

b - Immunofluorescence indirecte sur coupes de thymus ................... 42 c - Obtention des microsphères couplées à la thymuline .................. 42 d - Milieux réactionnels ................................................ 44
2. RESULTATS . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 45
a - Les immunsérums . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 45 b - Premier couplage sur les microsphères . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 47 c - Microsphères-SAB-thymuline . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 48 d - Test du stock d'immunsérums avec les microsphères-SAB-thymuline . . . . . SS e - Essais de nouvelles protéines intermédiaires . . . . . . . . . . . . . . . . . . . . . . . . 58
e-l . L'acide polyaspartigue . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 58 e-2 . La'métallothionéine . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 65
f - Le milieu réactionnel . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 68 g - Conclusions sur les microsphères . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 76
g-l . Influence du porteur protéique . . . . . . . . . . . . . . . . . . . . . . . . . . . 76 g-2 . Réactivité comparée selon la qualité des immunsérums
et des microsphères . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 77 g-3 . Agglutination comparée selon la gualité des immunsérums
et des microsphères dans un milieu avec ou sans zinc . . . . . 79 h- Diamètre des microsphères et appareillage . . . . . . . . . . . . . . . . . . . . . . . . . . . 80
h-l . Comparaison entre le néphélémètre BEHRING et le prototype de NANCY . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 81
h-2 . Comparaison entre les néphélémètres BEHRING et HYLAND pour les diamètres de 78 et 105 nm . . . . . . . . . . . . . . . . . . . . . . . 84
1 - Suivi de la production d'immunsérums D............................... 87
CONCLUSION 95 . ..............................................................
ANNEXES ................................................................ 101
REFERENCES BIBLIOGRAPHIQUES . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 107

PRESENTATION DU PROJET


1 - INTRODUCTION
La malnutrition protéino-énergétique (MPE) représente une importante endémie mondiale, touchant gravement les populations des pays en voie de développement et notamment les enfants.
Les formes graves de MPE sont les mieux caractérisées cliniquement mais les formes modérées et marginales sont de loin les plus fréquentes et peuvent échapper au diagnostic. Les divers degrés de malnutrition forment un continuum et sont difficiles à détecter et à différencier par les seules analyses biochimiques de laboratoire ou les mesures anthropométriques. Une étude plus précise des épisodes infectieux et des conséquences biologiques de la MPE sur l'immunité permet une autre approche des risques liés à la malnutrition. En effet la MPE est la première cause de déficits immunitaires acquis et elle est à l'origine d'une importante mortalité infantile, du fait principalement d'une susceptibilité accrue aux infections.
En càs de MPE les réponses métaboliques spécifiques au jeûne, au stress et aux infections sont associées à des degrés divers.
Nous voulons en évaluer les conséquences sur l'immunité par la mesure du taux sérique d'une hormone produite par l'épithélium du thymus : la thymuline.
Le thymus est très sensible à la malnutrition. Des observations faites chez l'enfant et de nombreuses études de MPE expérimentale chez l'animal ont montré que le thymus présente l'atrophie la plus sévère parmi l'ensemble des organes lymphoïdes (47, 82). Une involution importante du thymus a été constatée chez l'enfant mort de MPE (1, 46, 82, 83). Chez de jeunes enfants Sénégalais une corrélation à été montrée entre l'état nutritionnel apprécié par anthropométrie et l'épaisseur du thymus mesurée par échographie (64, 69). L'involution thymique serait une des conséquences les plus précoces de la MPE (46, 81). D'après ZIEGLER l'effet le plus direct du déficit nutritionnel serait une destruction du thymus d'abord fonctionnelle. puis anatomique (83). De même chez l'animal, des études suggèrent qu'un effet premier de la MPE sur le système immunitaire doit se situer au niveau de l'épithélium thymique chez des souris sévèrement carencées (58, 59).
Lors d'une étude post mortem d'enfants dénutris, JAMBON a pu aisément mettre en évidence des altérations liées au déficit thymique sur la
9

structure et le peuplement cellulaire des zones thymodépendantes des organes lymphoïdes (45, 82, 83).
L'ensemble de ces conséquences est probablement à la base du déficit de l'immunité à médiation cellulaire observé en cas de MPE (32, 40, 56, 76). Une altération numérique et fonctionnelle des lymphocytes thymodépendants, une modification des proportions de leurs sous-populations (18, 19) et une augmentation corrélative des lymphocytes non-T non-B (16, 47) suggèrent une atteinte de la fonction lymphodifférenciatrice du thymus, notamment au niveau de la production de thymuline. En effet notre équipe a trouvé une diminution de la thymuline associée à l'épithélium thymique en cas de MPE (44, 46, 81, 83). Cela est confirmé par l'observation d'une activité biologique de la thymuline sérique diminuée chez l'enfant (15, 16, 78) ou chez l'animal malnutri (17, 41, 60).
L'ampleur et les conséquences de l'altération des défenses immunitaires et de la susceptibilité aux maladies en cas de MPE, sont une sérieuse motivation pour déterminer la nature de ces effets, notamment par l'évaluation de la fonction thymique. Selon CBANDRA la baisse du taux de thymuline circulante est un facteur essentiel de la dépression immunitaire (16) -
Les taux de thymuline thymique et sérique abaissés lors de la malnutrition renforcent l'intérêt de doser cette hormone au niveau de la circulation, d'autant plus qu'il existe peu de méthodes non invasives d'étude de l'immunité chez l'enfant. Cette approche pourrait apporter une aide au diagnostic et aux indications thérapeutiques.
Le seul dosage de la thymuline sérigue existant actuellement est basé sur son activité biologique in vitro (4). Il est complexe, non spécifique, semi quantitatif et perturbable en cas d'infection (22, 78). Il serait donc intéressant de disposer d'une méthode quantitative et facile à mettre en oeuvre.
Nous avons envisagé l'application pour la mise au point du dosage de la thymuline, d'une nouvelle technique innnunochimique de haute sensibilité et non radioisotopique : l'immunonéphélémétrie à supports microparticulaires (10, 50, 70).
Cet outil permettrait d'évaluer les conséquences de la MPE à un niveau précoce et critique. Cette méthode sans contraintes techniques pourrait être mise à la disposition des équipes de terrain.
Après la présentation de la thymuline et de l'état des connaissances sur le sujet, nous décrirons en détail la mise au point des différents réactifs nécessaires au dosage par immunonéphélémétrie à supports microparticulaires en milieu synthétique.
II -IATHYMULI~
. :
10

a - Une hormone thymique
a-l . Structure du thymus
Le thymus est le premier organe lymphoïde a apparaitre chez e l'homme. Lors de l'ontogénèse le thymus est d'abord constitué par une petite masse dense de cellules épithéliales. Des cellules souches;véhiculées dans le sang, y pénètrent et se différencient en lymphocytes sous l'influence du microenvironnement épithélial. Une séparation cortico-médullaire apparait. L'organogénése du thymus est termin6e au cours même de 'la vie foetale.
Le thymus, situé 3 la base du cou, est constitué de deux lobes reliés entre eux sur la ligne médiane par du tissu conjonctif. Chaque lobe est cloisonné en lobules comprenant une partie périphérique très riche en lymphocytes, le cortex, et une médullaire moins riche en lymphocytes.
La trame du thymus est essentiellement constituée par des cellules épithéliales, plus nombreuses et de plus grande taille dans la médullaire, Oii elles se regroupent en structures arrondies appelées corpuscules de Hassal, dont le rôle est encore obscur. Dans la corticale les cellules épithéliales délimitent des espaces libres où se logent des groupes de lymphocytes.
Aspect du thymus humain Schématisation d'un lobule thymique
(d'après 7)
11

Les cellules épithéliales, notamment dans la mGdullaire, sont b l'origine des sdcrétions d'hormones thymiques.
Une involution physiologique commence au moment de la puberté, avec une r6duction du rapport cortico-médullaire,
' complètement. mais le thymus ne disparait pas
Le parenchyme s'infiltre de tissu adipeux. Cette involution s'accompagne d'une réduction progressive de la sécrétion d'hormones thymiques.
Klms
Involution du thymus avec l'âge chez l'homme _
a-2 - Fonction du thymus
Le thymus est un organe lymphoïde central ayant un rôle essentiel dans le développement du système lymphoïde et dans le maintien de l'équilibre immunitaire. C'est l'organe clé de l'immunité à médiation cellulaire. Chez l'enfant le thymus est particulièrement important car il assure le développement de l'immunocomp&ence nécessaire ensuite pour la résistance aux infections induites par de nombreux agents pathogènes.
Le thymus est le seul organe lymphoïde connu dont le développement se fait indépendamment de toute stimulation antigénigue. Son poids et l'activité des cellules ne sont pas modifiés par les inununisations, contrairement à ce gui est observé dans la rate et les ganglions.
Le thymus contrôle la production des lymphocytes T. Les cellules . souches lymphoïdes, provenant de la moelle osseuse, colonisent le thymus,
pour 90% dans le cortex, et s'y multiplient. Leur migration fait intervenir des facteurs chimiotactigues sécrétés par l'épithélium. La majorité des thymocytes corticaux meurent sur place. Très peu de thymocytes (5%) sont matures et sont localisés dans la médullaire. L'origine précise et le niveau
12

de maturité des cellules T gui migrent hors du thymus ne sont pas encore clairement établis. Les lymphocytes T se retrouvent au niveau de la circulation et colonisent les zones thymodépendantes ou paracorticales des organes lymphoïdes périphériques.
La fonction lymphodifférenciatrice et régulatrice du thymus sur les lymphocytes T est due à des facteurs sécrétés par l'épithélium et retrouvés au niveau de la circulation sanguine.
a-3 - Les hormones thymiques
Une partie essentielle du rôle du thymus se fait par l'intermédiaire d'un mécanisme hormonal. De nombreuses études ont analysé les effets d'une thymectomie ou d'une restauration par du tissu thymique. Depuis une quinzaine d'années, les équipes s'intéressent à isoler et identifier les hormones responsables des fonctions du thymus. Il existe encore trop d'inconnues pour apporter une conclusion définitive sur les hormones thymiques. Pour l'instant on imagine qu'il existe plusieurs hormones agissant éventuellement à différents niveaux de la différenciation des lymphocytes T. Il semble que le contact avec le microenvironnement thymique soit nécessaire pour les premières phases de cette différenciation. Une ou plusieurs hormones pourraient ensuite intervenir dans le thymus et à distance. SAVINO a montré que plusieurs hormones thymiques étaient produites par les mêmes cellules épithéliales (72).
Des taux abaissés ou une utilisation déficiente des hormones thymiques entrainent un déséquilibre immunitaire pouvant contribuer à l'étiologie de nombreuses maladies. Des extraits thymiques plus ou moins grossiers ainsi que des facteurs synthétiques ont été testés in vitro (33) ou administrés à des malades atteints de déficits immunitaires, avec des résultats de restauration prometteurs (11, 77).
Nous présentons sommairement les facteurs d'origine thymique ayant des propriétés de stimulation des lymphocytes T les mieux caractérisés actuellement.
* La thymosine fraction V, obtenue par A.L. GOLDSTEIN, est un extrait thymique de veau partiellement .purifié pouvant restaurer les fonctions immunitaires d'animaux thymectomisés ou immunodéprimés (37). Deux . peptides biologiguement actifs ont d'abord été isolés de cette fraction : les thymosines a1 (28 acides aminés (AA)) et J34 (43 AA) (54). D'autres thymosines plus récemment décrites présentent une homologie importante avec la thymosine al (a11 de 112 AA et pro-a de 35 AA) et de même activité biologique, ou avec la thymosine R4 (~38 de 39 AA, 09 de 41 AA, l310 de 42 AA, 1311 de 41 AA) et sans activité biologique rapportée (55).
* La thymopoïétine, isolée du thymus par G. GOLDSTEIN, induit la transformation des précurseurs de la moelle osseuse en thymocytes (38). C'est un polypeptide de 49 AA, de séquence connue (74), dont peuvent être isolés deux fragments actifs.. de 11 et 5 AA.. Ce dernier est appelé thymopentine ou TP5 (39). ,, ~.

* Un facteur humoral thymigue a été isolé par TRAININ à partir du thymus de veau (48). C'est un polypeptide de 30 résidus, de poids moléculaire d'environ 3 000 D et de séquence non connue, gui a la propriété d,'augmenter la réponse des splénocytes aux mitogènes et de restaurer une réaction de greffon contre l'hôte pour des splénocytes de souris thymectomisée.
* La thymuline, anciennement dénommée facteur thymique sérigue @TSI, a été découverte par BACH (4) gui a eu le mérite de l'isoler du sang ( c.omme son nom l'indique) et non d'extraits thymiques comme les autres hormones, ce gui a permis de lsenvisager accessible à un dosage au niveau de la circulation. Son mode d'action est encore en partie inconnu, mais semble faire intervenir la fixation sur des récepteurs spécifiques (8, 35), puis la synthèse d'AMP cyclique et peut-être de prostaglandine PGE2.
La thymuline est la seule hormone thymique de structure chimiquement définie dont la présence dans la circulation ait été démontrée.
L'ensemble des travaux ayant mené à la caractérisation de la th>ymuline est basé sur la mesure de son activité biologique in vitro.
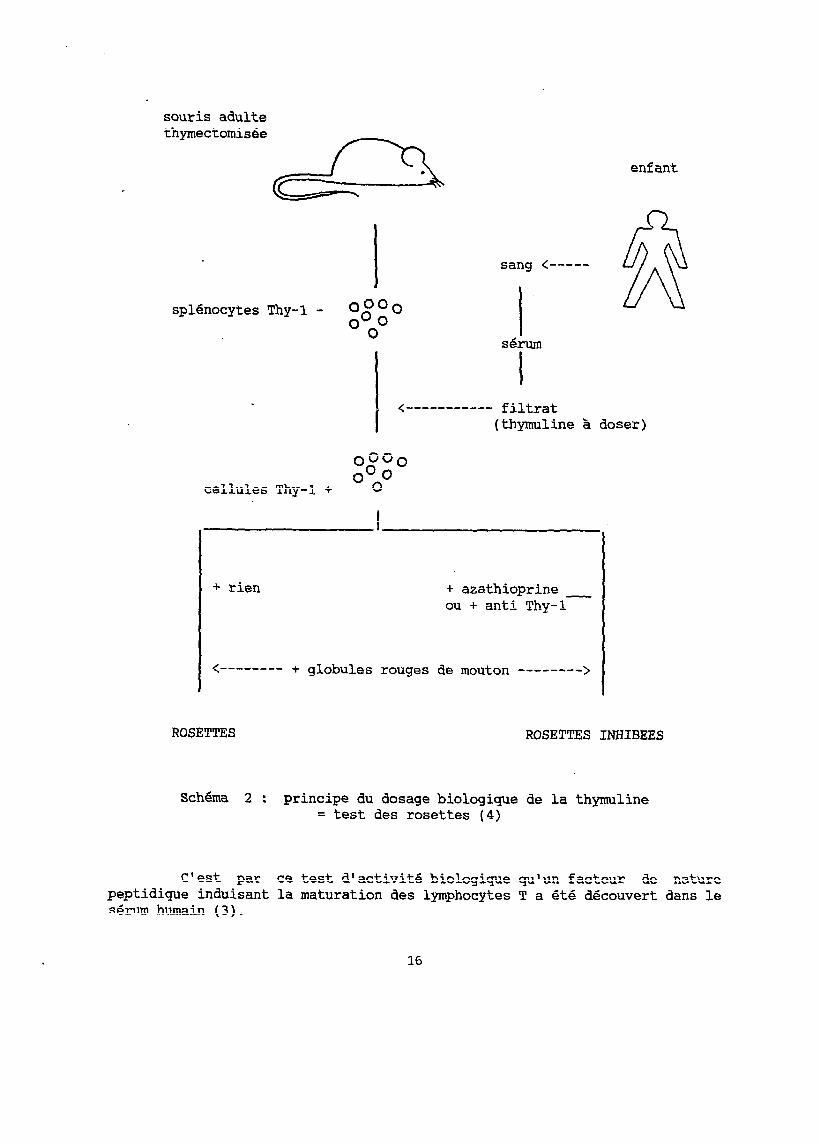
b- Activité biologique de la thmline; sa mesure par le test des rosettes
Les lymphocytes T ont la propriété de réagir in vitro avec les globules rouges de mouton et de former des "rosettes". La réaction peut être facilement visualisée et permet la mesure de cette population lymphocytaire ou l'étude de certaines de ses caractéristiques membranaires intervenant dans la réaction.
L'action biologique de la thymuline permet la maturation des lymphocytes T, provoquant ainsi l'apparition de marqueurs membranaires spécifiques dont celui dénommé Thy-1, gui intervient dans la formation des "'rosettes".
Les lymphocytes T immatures Thy-l- (absence du marqueur) ainsi que les lymphocytes T rendus matures par la thymuline, Thy-1+ (présence du marqueur), peuvent former des "rosettes".
Mais en présence d'azathioprine ou d'anticorps se fixant sur le marqueur Thy-1, les lymphocytes matures Thy-l+ ne peuvent plus former de "rosettes" alors que les lymphocytes immatures Thy-l- ne sont pas influencés par ces inhibiteurs dans leur réaction avec les globules rouges de mouton (voir schéma 1).
Cette inhibition de la formation des "rosettes" est donc le reflet des lymphocytes Thy-1+ et par là de la thymuline gui a provoqué leur maturation.
Cela est à la base du dosage biologique de la thymuline.
Pour doser la thymuline, des splénocytes de souris adulte thymectomisée, donc immatures Thy-1-, dont la formation en "rosettes" ne peut être inhibée, sont incubés pendant 30 minutes à 37Oc avec les échantillons biologiques dont on veut doser l'hormone. La thymuline entraine la maturation des lymphocytes T, l'apparition du marqueur Thy-1, et restaure la possibilité d'inhiber, par l'azathioprine ou des anticorps anti Thy-1, la
14

formation des "rosettes" par ces cellules.
La dernière dilution d'échantillon permettant que la formation des rosettes soit encore inhibée à 70% dans les conditions du dosage, donne le titre en thymuline (voir schéma 2).
0 thymuline T -----------_----------- > 0 T
Thy-1 - Thy-1 +
lymphocyte T immature
lymphocyte T rendu mature par la thymuline apparition du marqueur membranaire Thy-1
+ azathioprine ou + anti-Thy-1 + rien
t azathioprine ou + anti Thy-1 + rien
+ -- globules rouges-
de mouton
+ -- globules rouges-
de mouton
ROSETTES ROSETTES NON ROSETTES ROSETTES INHIBEES INHIBEES
Schéma 1 : formation de "rosettes" par les lymphocytes T
15
souris adulte thymectomis&e
enfant
n
I sang <-----
splénocytes Thy-1 - o*oo OO I-
sérum
I
1 < ----------- filtrat (thymuline à doser)
0000
cellules Thy-1 + OOo”
+ rien + azathioprine ou + anti Thy-l-
< -------- + globules rouges de mouton -------- >
ROSETTES ROSETTES INHIBEES
Schgma 2 : principe du dosage biologique de la thymuline = test des rosettes (4)
C'est par ce test d'activité biologique qu'un facteur de nature peptidigue induisant la maturation des lymphocytes T a été découvert dans le sérum humain (3).
16

La présence de ce facteur a été mise en évidence dans le sérum de diverses espèces (souris, rat, porc, poulet, homme). Son niveau sérique dépend de l'âge. Il est stable jusqu'à 6 mois chez la souris et 20 ans chez l'homme puis diminue ensuite parallèlement au poids du thymus (4).
La thymodépendance de ce facteur isolé du milieu sérique, a été initialement suggérée par sa disparition de la circulation après thymectomie et sa réapparition après greffe de thymus chez la souris (3). Ensuite de la thymuline a été retrouvée dans des extraits thymiques (24). COHEN a montré qu'une sous-population de cellules épithéliales humaines en culture sécrète de la thymuline, sécrétion modulée par la thymuline elle même (21). Des extraits de rate, de muscle ou de cerveau n'ont pas cette même activité biologique (4), confirmant la spécificité thymique de l'hormone.
c - Purification et caractérisation
L'équipe de BACH a purifié la thymuline à partir de sang de porc défibriné, ultrafiltré, concentré et soumis à une filtration sur gel, une chromatographie sur carboxyméthylcellulose, une lyophilisation et une dernière filtration sur gel (4).
La forte augmentation de l'activité biologique au cours de la purification est en partie due à l'élimination d'une fraction protéique gui est inhibitrice dans le test des rosettes (6) et gui est responsable de la baisse du taux de thymuline constatée dans certains cas (25, 28, 31).
La thymuline ne peut donc pas être détectée par son activité biologique dans le sérum total. Les échantillons doivent être débarrassés des inhibiteurs de hauts poids moléculaires par filtration (4, 55).
La thymuline n'est pas fixée de façon covalente ni détruite par cet inhibiteur. En effet après filtration un retour au niveau normal d'inhibiteur est observé si le rétentat de la filtration et la thymuline dosée dans le filtrat sont préincubés ensemble (6).
Il existe aussi des inhibiteurs de faibles poids moléculaires (4.000 à 20.000 D) gui nécessitent des filtrations plus fines pour être éliminés (6, 25). Ceci tendrait à montrer gue pour être correctement détectée par le dosage biologique, la thymuline doit être en grande partie purifiée.
La thymuline circule dans le sang en partie fixée à des transporteurs. Son activité biologique est retrouvée, après fractionnement du sérum, dans deux fractions dont les poids moléculaires correspondent à ceux de la thymuline libre et de la préalbumine, mais en des proportions rapportées très différentes selon les auteurs. D'aprk D-ARDENNE environ 20 % de l'activité thymuline du sérumest associée à la préalbumine (23) alors gue cette fraction serait de 95 % selon WHITE (79).ou BURTON (14). Ces différences de résultat soulignent la difficulté d'évaluer l'activité biologique de la thymuline non seulement dans le sérum, mais aussi dans des fractions sérigues ou d'évaluer l'association de la thymuline à une protéine sérigue selon le traitement auquel est soumis le sérum.
Dans du sérum de souris thymectomisée ces deux fractions ne
17

présentent pas l'activité thymuline, gui réapparait si l'hormone pure est injectée aux animaux (23).
La préalbumine' est un transporteur connu pour les peptides et certains auteurs considèrent qu'elle se comporte comme une hormone thymique quant à ses effets in vitro, notamment dans le test des rosettes (14). Ce serait une hormone circulante régulant la réponse immunitaire de populations de lymphocytes T (52, 53).
Ayant observé que l'activité de la thymuline peut être prolongée par une préincubation avec du sérum de souris thymectomisée, DAPDENNE isole un porteur protéique de la thymuline et postule qu'il s'agit de la préalbumine (23). Mais certaines fonctions immunologiques associées à la préalbumine seraient indépendantes de la thymuline (52).
SAVINO a mis en évidence quatre molécules de poids moléculaire compris entre 48.000 et 59.000 D, reconnues dans des cellules épithéliales thymiques humaines en culture par des anticorps monoclonaux anti-thymuline (73), gui pourraient être des transporteurs ou des précurseurs.
La thymuline ainsi Caractéris&e et purifiée est un nonapeptide d'un poids moléculaire de 890 D. Sa séquence en acides aminés a été déterminée : (5, 65)
<Glu - Ala - Lys - Ser - Gln - Gly - Gly - Ser - Asn
Il n'existe apparemment pas de spécificité d'espèce puisque la séquence en acides aminés est la même pour la thymuline de porc et de veau (24) ou d'homme (49, 67).
De la thymuline synthétique a alors été fabriquée. La thymuline naturelle et la thymuline synthétique sont identiques dans lës approches biologiques, biochimiques ou radioimmunologigues. Elles agissent à la même concentration dans le test des rosettes (6).
Le peptide synthétique présente donc une activité biologique mesurable. Plusieurs laboratoires ont synthétisé et confirmé sa haute activité biologique. Cependant la préparation fortuite de lots instables ou inactifs a suggéré que la thymuline peut exister sous deux formes, actives ou inactives biologîguement. Finalement il s'est avéré que l'activité biologique du peptide nécessite du zinc fixé à la molécule (26, 27). La présence de zinc dans la thymulîne synthétique a été confirmée par absorption atomique. Elle acquiert vraisemblablement l'activité biologique par des traces de zinc contaminant les réactifs (27).
Le peptide synthétique ou naturel perd son activité biologique après traitement par un agent chélatant les cations divalents. L'addition de chlorure de zinc à de la thymuline désactivée restaure son activité (27, 36).
D'autre part chez des souris déficientes en zinc, le taux sérigue de l'hormone est bas mais l'addition de chlorure de zinc au sérum permet de retrouver l'activité biologique normale (29). Donc le peptide est présent dans le sérum même si le déficit en zinc entraine une baisse de l'activité mesurée. Ces résultats semblent montrer que le test des rosettes ne détecte
18

que la forme associée au zinc, ce gui pourrait expliquer des possibilités d'erreur ou de contradiction selon le milieu, plus ou moins chélatant, utilisé.
Avec la thymuline synthétique, associée au zinc ou non, différentes équipes ont fabriqué des anticorps (polyclonaux chez le lapin et monoclonaux chez la souris) dont l'efficacité est testée par leur effet inhibiteur de l'action biologique de la thymuline dans le test des rosettes. Ces anticorps ont été utilisés pour poursuivre la caractérisation et l'étude de la thymuline, notamment sa localisation spécifique dans le thymus, l'étude de la relation structure-activité avec divers analogues synthétiques, l'étude comparative de la thymuline avec ou sans zinc et la mise au point de nouveaux dosages non biologiques
La présence de thymuline dans les cellules épithéliales du thymus a été directement démontrée par immunofluorescence (43, 61, 71), ou par obtention d'anticorps anti-thymuline chez des animaux immunisés avec des cellules épithéliales thymiques (9). Les résultats ont été confirmés par microscopie électronique (2, 75).
Divers analogues, avec un ou plusieurs acides aminés changés, ont été synthétisés pour étudier l'importance de la séquence dans certaines propriétés (13, 55). Ainsi Glu 1 et Ala 2 ne sont pas nécessaires à l'activité biologique, mais leur présence (surtout Ala 2) augmente beaucoup l'activité de l'heptapeptide C-terminal. Glu 1 et Ala 2 ne sont pas nécessaires à l'antigénicité (42) ni la chaine latérale de la Lys 3 (34). Par contre selon YOSHIDA de faibles changements en C-terminal entrainent la disparition des activités biologiques et antigénigues et l'octapeptide C- terminal est la partie minimum essentielle de la molécule de thymuline pour l'expression de son activité biologique (80).
Ces différences de résultats sur la relation stucture-activité peuvent s'expliquer par le fait que le dosage biologique utilisé n'est pas toujours équivalent d'un laboratoire à l'autre.
L'antigénicité du peptide est modifiée selon qu'il est associé ou non au zinc (30, 68).
a- Essais de dosages de la thymuline
Différentes équipes ont produi t des anticorps dans le but de mettre au point un dosage non biologique de l'hormone, par des méthodes radioimmunologigues (RIA) ou enzymoimmunologigues (ELISA, EIA).
Dans les dosages radioimmunologigues en milieu synthétique, un immunsérum de lapin permet d'obtenir une bien meilleure sensibilité gu'un anticorps monoclonal de souris (34, 63).
PLFAU a mis au point un RIA avec le peptide synthétique associé ou non au zinc et radiomarqué, permettant de différencier les deux formes en
19

milieu synthétique. Avec ce dosage seule de la thymuline ayant fixé du zinc était détectée dans le sérum de souris (68). Plus récemment METREAU a mis au point un EIA gui en milieu synthétique détecte de la même façon les deux formes du peptide (57).'
Les différentes équipes ayant mis au point un dosage non biologique de la thymuline n'ont pas pu l'appliquer au milieu sérique entier à cause de protéines interférentes. Tous préconisent une filtration, une extraction ou une purification de la thymuline pour pouvoir la doser dans les échantillons Biologiques. Certains auteurs ont essayé de doser avec leur méthode des sérums traités, mais les traitements nécessaires s'avèrent très complexes (comprenant par exemple successivement une précipitation au TCA, une extraction à l'éther, deux lyophilisations et une chromatographie de gel filtration (42)). De plus les faibles taux sériques obligent à concentrer les échantillons.
Leurs résultats sont présentés dans le tableau suivant :
concentration limite de taux sérigues déplaçant de détection de thymuline méthode référence 50% le signal mesurés
de base --------w--w --------- --------------------- ------- ---------
200 pg/ml 20 w/ml porc : 100 - 500 pg/ml RIA (66) ----------------------------------------------------------------------
400 pg/ml 20 w/ml enfant 5 ans : 44 pg/ml homme 63 ans : < 20 pg/ml RIA (63)
---------------------------------------------------------------------- 20 w/ml 2 pdml porc 7 jours : 22 pg/ml
souris Balb/c : 1.3 pg/ml souris nude : non detecté RIA (42)
-----------------------------------------------------------~---------- non citée 50 pdml homme : 26 pg /ml
porc : 32 Pdml souris : 45 pg/ml; souris thymectomisée : 2 pg/ml RIA (6)
---------------------------------------------------------------------- 32.5 pg/ml 5 pdml impossible EIA (57)
---------------------------------------------------------------------- 10 Pg non citée impossible RIA (68)
---------------------------------------------------------------------- 15 Pg 5 Pg impossible RIA (67)
---------------------------------------------------------------------- non citée 20 pdml RIA (34)
---------------------------------------------------------------------- non citée non citée suivi de la production ELISA (62)
d'anticorps ---------=,-,-,,,,,--,,-----,-----,---,-------------------------------
20

Aucune des méthodes ainsi mises au point et publiées n'a été ensuite exploitée pour des études plus approfondies et plus variées des taux de thymuline. Cela est dù à leur mangue de sensibilité et à la présence d'interférences sérigues ce gui impose un traitement très lourd à l'échantillon biologique, équivalent à une purification de la thymuline.
En conséquence, malgré la facilité de doser la thymuline synthétique, il n'existe toujours pas aujourd'hui d'autres moyens de doser le thymuline naturelle, au niveau du sérum traité, que par la mesure de son activité biologique dans le test des rosettes.
Par l'activité biologique ce n'est pas la molécule de thymuline qui est dosée mais une fonction associée à sa présence. La maturation des lymphocytes T peut être induite par d'autres hormones thymiques ou d'autres molécules que la thymuline. Par exemple un "facteur allogénigue", imitant l'action de la thymuline mais n'étant pas reconnu par des anticorps anti- thymuline, a été mis en évidence dans le sérum en cas d'infection (22, 78). Ce dosage n'est donc pas spécifique.
Comme pour le dosage par l'activité biologique, les méthodes immunochimigues gui dosent pourtant directement la molécule de thymuline synthétique, sont perturbées par des composants du sérum et nécessitent d'isoler la thymuline naturelle par ultrafiltration essentiellement. Un dosage plus simple et plus sensible permettrait de diluer les khantillons biologiques pour éviter les effets aspécifigues de milieu et d'aborder plus aisément le problème des interférences sérigues.
Il est donc intéressant d'essayer de mettre au point un dosage de la thymuline par une méthode différente, comme nous l'avons envisagé. La méthode choisie permet d'espérer atteindre une sensibilité inférieure au pdml (10). De plus le complexe résultant de la réaction de l'antigéne avec l'anticorps est directement détecté en néphélémétrie. Cette méthode est spécifique.
Contrairement aux méthodes radio- ou enzymoimmunologigues, la néphélémétrie à supports microparticulaires est d'une grande simplicité d'utilisation. Ses avantages sont notamment qu'elle n'utilise pas de radioactivité et que le résultat est obtenu en une ou deux heures après une seule étape de mise en réaction de trois produits.
e - Etude de conformation en RMN-2D
DARDENNE a montré gue la thymuline peut exister sous deux formes, l'une biologiguement active associée au zinc, l'autre biologiquement inactive dépourvue de zinc (26, 27).
L'antigénicité d'une molécule est liée de façon étroite à sa conformation, influant sur la reconnaissance par un anticorps, et peut conditionner les performances d'un dosage immunochimigue.
C'est pourquoi une étude des conformères de la thymuline avec ou sans zinc par résonnance magnétique nucléaire à deux dimensions (PMN-2D) a été initiée par le Laboratoire d'immunologie du Professeur DUHEILLE à la Faculté de Médecine de Nancy en association avec deux équipes de
21

physicochimistes : le Laboratoire de Biophysique de l'université de Nancy 1 et le Laboratoire de Chimie Physique Macromoléculaire de 1'Ecole Nationale Supérieure des Industries Chimiques de Nancy (51)
Une première approche a été menée en milieu organique (diméthylsulfoxide, DMSO). Une conformation thermodynamiquement probable par repliement de la chaine du peptide libre a pu être décrite. Ensuite en prkience de zinc, l'existence de deux complexes métallo-peptidigues différents a été montrée, alors qu'un seul était décrit jusqu'alors. Un atome de zinc pourrait complexer soit une soit deux molécules de thymuline. Les constantes dPassociation calculées n'indiquent pas d'affinité forte pour le.métal.
Ces premiers résultats en milieu organique ont montré la sensibilité ,de la molécule de thymuline aux paramètres de son environnement physicochimique.
Une étude de la conformation de l'hormone en milieu aqueux a ensuite été menée, dans un mélange H20/D20 95/5 v/v, puis avec de la thymuline marquée par le 13C. Des mesures de dichroïsme circulaire dans différents environnements ont été aussi prises en compte pour la modélisation conformationnelle.
Les résultats concordent pour montrer que la thymuline n'adopte probablement pas une conformation unique et repliée dans l'eau, contrairement à ce gui avait été observé en milieu organique (DMSO). La proportion des conformères repliés thermodynamiquement plus stables est faible et ils s'échangent rapidement avec des formes non,repliées. En milieu aqueux il n'est plus possible de caractériser un complexe stable du peptide avec le zinc. Les molécules liées au métal sséchangeraient rapidement et en permanence avec le pool de peptide libre.
Ces résultats de RMN montrent que la thymuline peut exister sous différents conformères en équilibre entre eux.
Des équilibres dépendants de la concentration en zinc s'instaurent entre les conformères avec et sans zinc. De même des équilibres s'établissent entre les différents conformères sans zinc, et entre les différents conformères avec zinc.
Le dosage électif d'un conformère par fixation sur un anticorps le soustrait du milieu et induit un déplacement de l'équilibre entre les conformères et la réapparition de celui gui est reconnu et fixé par l'anticorps. Un tel dosage est donc dépendant du milieu et notamment du zinc. C'est pourquoi nous avons mis au point le dosage en milieu séquestrant les cations divalents pour s'affranchir de cette incertitude liée au degré de complexation de la thymuline avec le zinc, et ne doser que les formes du peptide sans ion métallique.
22

MISE AU POINT DU DOSAGE
PAR IMMUNONEPHELEMETRIE A
SUPPORTS MICROPARTICULAIRES
23


1. PRINCIPES
Le dosage de la thymuline par immunonéphélémétrie nécessite :
Des microsphères greffées par la thymuline
Des anticorps dirigés contre la thymuline
Un milieu réactionnel adapté
Un néphélémètre
La mise au point commence lorsque des microsphères greffées sont reconnues et agrégées par les anticorps. L'agrégation, gui ne peut se faire que dans un milieu adapté, est détectée par un néphélémètre. On voit donc que les quatre acteurs du dosage interagissent et gue l'on doit figer trois des composants pour étudier le quatrième.
Précisons qu'en l'absence d'agrégation la mise au point ne peut commencer et gu'aucun des quatre constituants ne peut être validé, d'où la difficulté de la phase primaire de la mise au point.
Seule la spécificité des knmunsérums peut être appréciée mais par une autre méthode ce gui ne les valide pas forcément pour l'immunonéphélémétrie .
Pour une meilleure compréhension de la méthode, il est important d'en préciser quelques principes en détail.
a - Mesure néphélémétrique
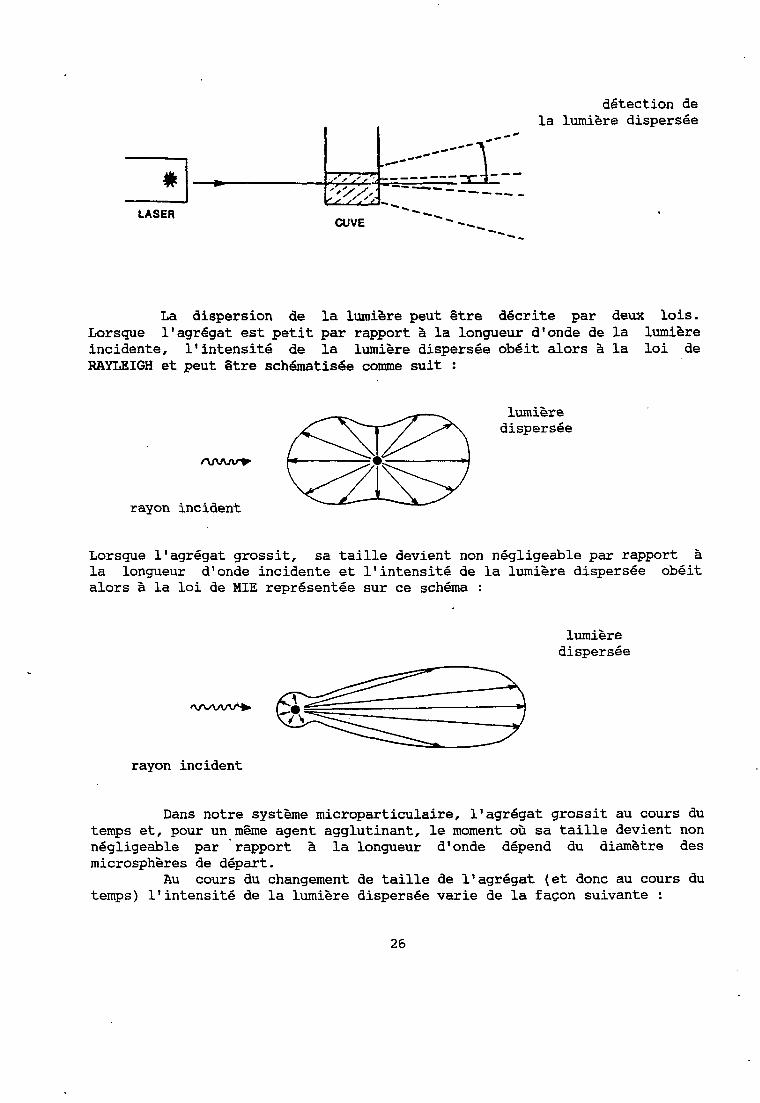
La néphélémétrie est la mesure de la lumière diffusée par des particules en suspension dans un liquide ou un gaz.
Les néphélémètres récents utilisent un laser Hélium-Néon gui fournit une source monochromatique, de longueur d'onde 631.8 nm, puissante, dirigée vers la cuve échantillon.
La différence entre les appareils vient surtout de l'angle choisi par rapport à la lumière incidente pour placer le système de détection de la lumière diffusée.
25
dgtection de
La dispersion de la lumière peut être décrite par deux lois. Lorsque l'agrégat est petit par rapport à la longueur d'onde de la lumière incidente, l'intensit6 de la lumière dispersée obéit alors à la loi de RAYLEIGH et peut être schématisée comme suit :
lumière dispersée
rayon incident
Lorsque l'agrégat grossit, sa taille devient non négligeable par rapport à la longueur d'onde incidente et l'intensité de la lumière dispersée obéit alors à la loi de MIE représentée sur ce schéma :
lumière dispersée
rayon incident
Dans notre syst'ème microparticulaire, l'agrégat grossit au cours du temps et, pour un même agent agglutinant, le moment où sa taille devient non négligeable par 'rapport à la longueur d'onde dépend du diamètre des microsphères de départ.
Au cours du changement de taille de l'agrdgat (et donc au cours du temps) l'intensité de la lumière dispersee varie de la façon suivante :
26

-
3 --@sgg
4 -*
agrégat au début de sa formation
sa taille est encore très inférieure à la longueur d'onde
sa taille est devenue non négligeable par rapport a la longueur d'onde
la taille continue à augmenter
Nous constatons donc que l'augmentation de la lumière dispersée au cours du temps n'est pas équivalente si l'angle de mesure est petit (environ 10 ") ou grand (environ 30 ").
A un petit angle de détection l'intensité varie peu au début lorsqu'elle suit la loi de RAYLEIGH et varie de façon plus importante ensuite suivant la loi de MIE.
intensité de la
lumière dispersée
la détection est peu significative au début de la réaction mais le devient ensuite
temps
A cet angle les agrégats sont d'autant plus vite détectés qu'ils sont gros. L'emploi de microparticules assez grosses au départ permettra de suivre la croissance de l'agrégat pendant longtemps, mais il faut savoir gue le début de leur formation ne sera pas détecté.
A un plus grand angle de détection l'intensité de la lumière dispersée varie plus au début lorsqu'elle suit la loi de RAYLEIGH'qu'ensuite lorsque les agrégats sont devenus gros. Au delà d'une certaine taille leur augmentation n'est plus détectée. La détection dans ce cas est intéressante à des temps faibles de réaction et avec des agrégats et donc des microsphères de départ de petite taille.
27

intensité de la
lumière dispersee
la détection est plus significative au début de la réaction qu'ensuite
temps
A un grand angle de mesure la détection de la lumière dispersée, PhS faible, nécessite une plus grande amplification qu'à un petit angle.
Au cours des mises au point que nous décrivons nous avons utilisé trois néphélémètres:
- WLAND : angle de mesure 30 O, photomultiplicateur - BEBEING : angle de mesure 10 O, photodiode - Prototype NANCY : angle de mesure 10 O, photodiode
(appareil mis au point par le Laboratoire d'lknunologie du Pr. DUBEILLE de la Faculté de Médecine de Nancy)
Le photomultiplicateur donne une plus forte amplification que la photodiode.
Ajoutons gu'un grand angle de lecture permet l'emploi de réactifs plus dilués, par contre il nécessite de tenir compte des blancs réactifs car la mesure est plus sensible aux petites particules contaminantes.
b- Principe de la technique utilisée : llimnunonéphélémétrie
En milieu liquide la réaction entre des anticorps divalents et les antigènes correspondants, pourvu qu'ils soient multivalents, conduit à la formation d'un réseau entrainant un trouble du milieu.
L'inununonéphélémétrie classique est la quantification de cette réaction par la mesure de la lumière dispersée par le trouble formé. Cette technique est rapide et simple à mettre en oeuvre mais présente des inconvénients importants, notamment :
- l'impossibilité de doser des haptènes (comme la thymuline), petites molécules monovalentes pour leur réaction avec les anticorps.
- le seuil de détection est médiocre car les complexes antigènes-anticorps doivent être assez gros pour être détectés. La sensibilité n'est qie de
-l'ordre du pg/ml, ce gui est bien moins performant que celle des techniques plus complexes comme les dosages radio- ou immunoenzyrnologiques.
Le Laboratoire d'immunologie du Pr. DUBFILLE à la Faculté de Médecine de Nancy a mis au point un réactif particulaire (microsphère)
28

stable sur lequel des molécules protéiques peuvent être couplées chimiquement. Cela a ouvert la voie à une nouvelle technique : l'immunonéphélémétrie à supports microparticulaires (70).
Le grossissement de l'antigène ou de l'anticorps par sa fixation sur les microsphères permet une amplification de la taille du complexe antigène-anticorps pour une même réaction et donc une augmentation de la lumière dispersée. Il en résulte une meilleure détection de la réaction par l'appareil, ce gui accroit la sensibilité de la technique, l'amenant au niveau des méthodes radio- ou enzymoimmunologigues.
Le support particulaire constitue donc un amplificateur passif du signal néphélémétrigue. De plus la fixation de plusieurs molécules d'antigènes sur une microsphère rend possible le dosage des haptènes.
Cette technique est simple et d'excellente sensibilité et nous avons décidé de l'appliquer au dosage de la thymuline, en profitant de l'expérience et des succès des chercheurs de Nancy pour d'autres dosages de protéines sérigues et d'hormones thyroïdiennes (10, 50, 70).
c - Possibilités de couplage
La réaction d'immunonéphélémétrie à supports microparticulaires nécessite le couplage de la thymuline sur une microsphère, soit directement, soit par l'intermédiaire d'un porteur protéique. En effet la thymuline est une petite molécule et son couplage direct n'a pas permis de réaliser une "couverture protéique" importante autour de la microsphère.
La thymuline fixée sur les microsphères doit être reconnue par les anticorps pour permettre leur agrégation. Il est donc souhaitable que la thymuline soit fixée par le même coté sur les microsphères et dans le polymère utilisé pour l'immunisation (cf II-l-a). C'est contre la partie libre de la thymuline que sont produits les anticorps et c'est cette même partie libre qu'ils reconnaissent sur la surface des microsphères.
Enfin la liaison chimique entre la microsphère et la thymuline d'une part, et la protéine porteuse de l'irmnunogène et la thymuline d'autre part, doit être différente sinon des anticorps "anti-liaison", produits en même temps gue les anti-thymuline, agglutineraient non spécifiquement les microsphères.
La partie C-terminale de la thymuline correspond aux propriétés biologiques et antigénigues et c'est donc cette partie gui doit rester libre, la fixation se faisant du coté N-terminal.
La configuration doit aussi permettre une réaction immunologigue croisée avec la thymuline native à doser.
c-l I La thymuline
Plusieurs possibilités de réaction sont envisageables entre la thymuline et un support selon les fonctions mises en jeu. Nous allons les évoquer au vu de la structure primaire de la thymuline :
29

NH2 I NH2 c=o NH2 I I 1
y 0 I’; iil” fi (cr; y 1 y i f; c; jN\ II O=C C-C-N-C-C-N-C-C-N-C-C-N-C-C-N-C-C-N-C-C-N-C-C-N-C-COOH I I II II II II II II II II
/\-î\ HH HH HH HH HH HH HH HH
H HH H
<Glu Ala LYS Ser Gln Gly GUY Ser Asn
1 2 3 4 5 ‘6 7 a 9
* L'amine primaire en epsilon de la lysine 3 est la fonction la plus réactive de la molécule et peut conduire à une réaction de condensation sur un aldéhyde ou une cétone :
k thymuline - NH2 -+ O=C - ligand --------> thymuline - N = C - ligand
* La fonction carboxyle de l'asparagine 9 est aussi très réactive. Les carbodiimides peuvent l'activer et la faire réagir avec une amine primaire :
thymuline -C-OH + R'-N=C=N- R -----> thymuline - C - 0 - C = N - R II
.i II ’ *’ I 0 0 NH - R'
-------------- > thymuline - C - N - ligand + H- N - ligand
I II I 0 H
H
* Les groupements amides (asparagine 9 et glutamine 5) 'sont activables par l'hydrazine ou le glutaraldéhyde et dans un deuxième temps peuvent réagir avec une amine primaire :
,, '.
30 .

glutaraldéhyde :
CH0 I
thymuline C=CH-(CH2)3-CH0 CH0 1 1
thymuline CH0 I 1 l
CONHZ + (CH2)2 ------__ > CONH-CH-CH-(CH2)2-CH=C-(CH2)2-CH0 1 1 CH (CH2)3 1 I C-(CH2)2-CH0 CH0 I CH0
glutaraldéhyde thymuline activée
CH0 CH0 + H2N - ligand I I ----------------> ,thymuline-CONH-CH-CH-(CH2)2-CH-CH-(CH2)2-CH0
I I (CH2)3 I Y CH0 ligand
Hydrazine :
ii thymuline-C-NH2 + H2N-NH2
hydrazine
+ H2N-ligand
.NaN02 -w---m- > thymline-C-N3
thymuline activée
------------- > thymuline-C-NH-ligand
Signalons que la fixation par l'asparagine se conçoit plus facilement par activation directe de son carboxyle que par activation indirecte de son amide.
31

Par ailleurs l'activation de la glutamine conduirait à fixer la thymuline par le centre de la molécule, ce gui est peu souhaitable pour favoriser une reconnaissance par des anticorps gui devront réagir aussi avec la thymuline native.
L'activation de ces amides implique aussi la fixation sur une amine et est donc impossible par réaction directe sur une microsphère mais uniquement sur les groupements aminés d'une protéine intermédiaire. '
* Les dernières fonctions envisageables sont les hydroxyles des sérines 4 et a. Le résultat conduit lui aussi à une fixation sur une amine primaire mais les conditions de réaction sont difficiles et instables avec un pH élevé et des Aactions secondaires.
c-2 0 Les microsphères natives
Les fonctions aldéhydes réactives des microsphères peuvent se condenser aisément avec des amines primaires de toute molécule qui peut ainsi se fixer sur les microparticules sans aucune activation préalable.
* Des possibilités réactionnelles des microsphères et de la thymuline, on peut concevoir une fixation directe sur la microsphère :
- C=O + H - N - thymuline ------- > -C=N- thymuline
* Un autre schéma de couplage peut être envisagé en dégageant la thymuline ( quelques dizaines d'Angstroms ) de la microsphère gui a un diamètre de 1000 Angstroms environ : la microsphère est liée à une molécule protéique intermédiaire elle même liée à la thymuline.
Les molécules protéiques réagissent directement avec les microsphères par leurs fonctions amines primaires. Elles peuvent être des protéines, des polyaminoacides ou des "bras espaceurs" avec des fonctions amines ou acides.
Pour fixer la thymuline sur le porteur il faut :
- soit activer les carboxyles en 9 de la thymuline pour qu'ils réagissent avec les amines du porteur - soit activer les carboxyles du porteur pour qu'ils réagissant avec les amines en epsilon des lysines en position 3 de la thymuline.
Cette dernière solution semble la meilleure envisageable puisque le
32

fait de toucher à la partie C-terminale dans le premier cas modifierait les propriétés antigéniques de la thymuline.
d- Principe du dosage envisagé : par inhibition
Les microsphères recouvertes de thymuline, gui donnent seules un signal néphélémétrigue négligeable, sont agglutinées par les anticorps des immunsérums anti-thymuline, ce gui donne le signal maximal de référence (ou zéro d'inhibition). Pour le dosage en une étape par inhibition, l'ajout de l'antigène libre (thymuline synthétique pour l'étalonnage ou échantillon sérigue pour le dosage) consomme le réactif agglutinant et entraine une baisse du signal en relation avec la quantité de molécules à doser.
La mise en oeuvre d'un tel dosage demande donc que l'on dispose de particules sensibilisées avec l'antigène, d'anticorps spécifiques, d'un milieu réactionnel approprié et d'un appareillage adapté.
Les principes et les étapes du dosage, résumés ci-dessous, sont représentés dans les schémas des deux pages suivantes.
obtention des réactifs
microsphères-thymuline anticorps anti-thymuline
étalonnage en milieu synthétique
agglutination I
inhibition par la thymuline
I signal de référence
l dosage en une étape
Le but des différentes mises au point des paramètres de la réaction est d'obtenir la meilleure sensibilité possible pour la réaction d'inhibition c'est à dire de doser la plus faible concentration possible de thymuline synthétique.
. . . ,i
33 1 I

* Réactif antigène : couplage chimique cavalent de la thymuline sur le support microparticulaire
Microsphères + Thymuline ------------> MICROSPHERES-THYMULINE couplage
* Réactif anticorps : production d'immunskum chez le lapin par injection de thymuline fixée à l'ovalbumine
glutaraldéhyde Thymuline + Ovalbumine -------------- > polymère
injection au lapin
prélèvements d'immunsérum
ANTICORPS ANTI-THYMULINE
I B - 3’ 1 .
immunogène ;.
u
Y YY Y Y Y
34

PRINCIPE DE IA REAC!l!ION
l- DBtermination des conditions opkatoires pour l'obtention du signal de référence
Microsphères-Thymuline + anticorps ---------- > AGGLUTINATION anti-thymuline
2- Dosage en une étape par inhibition
Microsphères-Thymuline + Anticorps anti-thymuline
+
+
OIJ OO
thymuline
---------------- > INHIBITION
agglutination inhibée
révélée par une diminution du signal de référence 9
35

e - Caractéristiques et interprétatian des r&actions
Lors de la mise au point de ce dosage deux tests principaux sont utilisés pour évaluer la qualité des rgactifs mis en jeu :
- l'agglutination des microsphkes-thymuline par l'immunsérum spkifigue - l'inhibition de cette réaction par de la thymuline synthétique
Le milieu synthétique est un mélange complexe choisi dans lequel sont faites ces réactions de mise au point, c'est à dire sans ajout d'aucun sérum autre que l'immunskum de lapin anti-thymuline.
Une courbe d'agglutination, pour une concentration de microsphères donnée et à un te-s donné, représente le signal néphélémétrigue obtenu en fonction des dilutions de 1' immunsérum. Cette courbe a en général une allure "en cloche" similaire à celle de la précipitation classique antigène- anticorps en milieu liquide :
trouble du milieu dù à la
précipitation
h excès H--N excès
d'anticorps ,,#' '\x, d'antigène 3-x /
\ \ \ II \ \ +
/I \ \ /' \ \
1' \ \ /' \ \
I/ \ \ ,' \
dilution d'immunsérum
L'allure de la courbe est due au fait que l'excès d'anticorps ou l'excès d'antigene défavorise respectivement la réaction bivalente d'un anticorps avec deux antigènes et d'un antigène avec deux anticorps, ce gui diminue les possibilités de formation d'un réseau.
x x excès d'anticorps
précipitation maximale
4---w 1 1
&y+
-b-
e--l+ + +
-b--P +-9 excès d'antigène
36

Les signaux évoluent au cours du temps. Pour une concentration de microsphères donnée un ensemble de courbes d'agglutination est obtenu pour les differents temps de lecture.
signal néphél&nétrigue
dilutions de l'immunsérum
Cet ensemble de courbes est ensuite obtenu pour différentes dilutions de microsphères et permet de mieux cerner la dilution souhaitable d'utilisation.
La qualité de la réaction est &Valuée par le taux de multiplication du signal de base ("blanc microsphères", correspondant aux microsphères- thymuline seules sans anticorps) à un temps donné, le temps nécessaire pour que ce taux soit égal à 2, la cinétique de la réaction et l'allure de la courbe.
La qualité réactionnelle de l'immunsérum est estimge par la dernière dilution donnant un signal détectable et par la dilution donnant le signal maximal.
La qualité des microsphères est appréciée par leur dilution d'emploi pour obtenir un signal choisi, à une dilution d'immunsérum et à un temps de lecture donnés.
Ces ensembles de résultats permettent de sélectionner la courbe d'agglutination la plus adaptée pour avoir un signal satisfaisant, B partir de laquelle sera déterminée la dilution de travail de l'immunsérum pour l'inhibition, choisie dans le haut de la partie descendante de la courbe, pour ne pas être en excès d'anticorps et avoir un signal de référence maximal.
Les conditions -opératoires sont ainsi établies pour doser par inhibition.
La courbe d'inhibition représente la diminution du signal de référence en fonction des quantités croissantes d'antigène libre à doser.
37

thymuline
L'inhibition de l'agglutination par l'antigène est une preuve de la spécificité de la réaction établie.
La sensibilité de la réaction est évaluée par la plus faible dose de thymuline synthétique entrainant une baisse significative du signal de référence. Le résultat de l'inhibition est aussi exprimé par les doses inhibant à 50% et 100% le signal.
f.- Protocole général de mise en réaction
En pratique les mises en réaction sont faites à l'aide d'un diluteur, directement dans la cuve de mesure et pour un volume final de 300 pl.
Les cuves sont lues au néphélémètre à des temps successifs de réaction.
Pour l'agglutination, l'immunsérum est dilué avec du tampon dans les cuves en une série de raison 2 depuis la dilution 1/50 en général. Les "blancs cuves" sont lus avant l'ajout des microsphères-thymuline. Une cuve sans immunsérum permettra de mesurer le "blanc microsphères" à prendre en compte dans le taux de multiplication du signal de base. Les microsphères- thymuline prédiluées extemporanément sont ensuite ajoutées avec du tampon.
Pour l'inhibition la thymuline synthétique est diluée par du tampon dans les cuves en une série de raison 2, 3 ou 10 en fonction de la gamme d'étalonnage souhaitée. L'immunsérum prédilué est ajout6 et incubé 30 minutes à température ambiante avec la thymuline. Puis les "blancs cuves" correspondant sont lus et notés. Enfin les microsphères prédiluées extemporanément sont ajoutées avec du tampon.
II - HISE AU POINT DU DOSAGE EN HILIXU SYNTElETIQUE
l- PROTOCOLES
a - Obtention d'imnmsérums
Afin de rendre immunogène la thymuline, gui est un haptène, elle doit être fixée sur un porteur.
38

La thymuline est couplée à l'ovalbumine par l'intermédiaire du glutaraldéhyde. Ce procédé est préconisé par BACH et donne selon lui de meilleurs résultats que l'emploi du carbodiimide comme agent de couplage (61. Ce dernier a été utilisé pour le couplage sur les microsphères afin d'éviter toute réaction immunologigue croisée avec la liaison de la thymuline dans l'immunogène.
Le glutaraldéhyde est un réactif bifonctionnel gui établit un pontage cavalent entre les groupements aminés du porteur et de l'haptène. La réaction fait intervenir la formation de deux bases de Schiff :
Ovalbumine - NH2 + O=C - (CH2)3 -C=O I I
+ H2N - thymuline
glutaraldéhyde
-------> ovalbumine - N = CH - (CH2)3 - CH = N - thymuline
Dans la formation d'un tel complexe le NH2 latéral de la lysine 3 est préférentiellement réactif par rapport au NH2 de l'asparagine 9 de par leur différence de point isoélectrigue (pHi), ce gui est plus satisfaisant et permet de ne pas modifier la molécule de thymuline du coté C-terminal.
La réaction de polymérisation est provoquée par addition lente de glutaraldéhyde à un mélange de thymuline et d'ovalbumine. Après quelques heures la réaction est arrétée par du métabisulfite de sodium. L'immunogène est débarassé des réactifs en excès par dialyse.
Le protocole est détaillé dans l'annexe 1.
L'immunogène ainsi obtenu est mélangé à volume égal avec de l'adjuvant incomplet de Freund (complet pour la première injection). L'ensemble est agité fortement au vortex jusqu'à obtenir une émulsion, puis injecté à des lapins.
Pour des prélèvements en petites quantités, juste nécessaires aux tests d'agglutination et d'inhibition, quelques microlitres de sang sont prélevés par pigure d'une veine de l'oreille. Un petit appareil en forme de stylo (AUTOLET Owen Mumford, Woodstock, Angleterre) permet, en pressant sur son capuchon, d'actionner un ressort gui entraine une pointe faisant des pigures franches et nettes comme celles effectuées chez l'homme au bout du doigt. Les gouttes de sang sont recueillies dans de fins capillaires (tubes à microhématocrites). Après coagulation du sang les tubes sont centrifugés, les globules rouges se tassent au fond du tube et le sérum est récupéré en cassant le tube.
Pour l'obtention de grandes quantités de sang les prélèvements sont
39

faits à l'oreille, préalablement chauffée pour faire grossir les veines et les rendre plus visibles. L'oreille est incisée au niveau d'une veine puis introduite dans le col d'une fiole à vide. Le sang est aspiré et 25 à 35 ml peuvent être recueillis.
Après coagulation une nuit à +4OC , une centrifugation permet de récupérer le sérum. De l'azide de sodium est ajouté pour prévenir le développement de contaminations, et le sérum est aliquoté sous 1 ml et stocké à -2O“C.
Trois séries d'immunisation ont été effectuées.
* Première série :
Pour cette série de 8 lapins nommés 001 à 008, le protocole retenu est celui d'immunisation lente utilisé par MONIER (61). 4 lapins (001, 002, 083, 004) ont reçu l'immunogène ovalbumine-thymuline et 4 lapins (005, 006, 007, 008) l'immunogène fait avec le peptide synthétique mélangé à du chlorure de zinc (ZnC12). Les injections sont toutes faites par voie intradermique en de multiples points du dos; seuls les lapins 001, 003, 005 et 007 ont reçu en plus une injection intrasplénigue lors de la première innnunisation.
Le calendrier a été le suivant :
jour
0 A (avant injection) 0 PREMIERE 3 B 7 C
14 D 20 E 35 F 57 G
102 H 116 RAPPEL no 1 119 1 123 J 130 K 148 L 158 M 187 RAPPEL no 2 190 N
' 199 0 310 .P 313 Q
injection prélèvement
40

* Deuxième série :
Un essai de protocole d'.immunisation plus rapide et plus intense a été mené chez 3 lapins nommés 050 à 07b, avec l'immunogène ovalbumine- thymuline. Toutes les injections ont été faites par voie intradermique en plusieurs points du dos, selon le calendrier suivant:
jour injection prélèvement
0 1 8
15 20
22-24 26-29 31-33 36-39
40 43 46
182 183 187
A (avant injection) PREMIERE RAPPEL no 1 RAPPEL no 2
B
RAPPELS no 3 à 11 injections consécutives
C D E
RAPPEL no 12 F
* Troisième série :
Une dernière série de 3 lapins, nommés 100, 200 et 300, a été. immunisée suivant le protocole lent avec le complexe ovalbumine-thymuline, par voie intradermique en plusieurs points du dos. Le calendrier a été le suivant :
jour injection prélèvement
0 PREMIERE 94 RAPPEL no 1 -
219 RAPPEL no 2 - 225 A 226 B 296 RAPPELnO - 302 C 303 .P 305 E 359 RAPPEL no 4 - 362 F 364 G 366 H 371 1 376 J
41

fJ- Immunofluorescence indirecte SKW coupes de thymus
Une technique d'immunofluorescence standard a été utilisée pour tester les immunsérums pendant la phase initiale de mise au point du dosage car nous ne disposions pas encore de réaction d'agglutination.
Des coupes sont effectuées sur des thymus de porcelet congelés, et déposées sur des lames porte-objets. Elles sont réhydratées puis incubées avec les dilutions dlimmunsérum anti-thymuline. Les lames sont lavées pour éliminer l'excès de ce premier anticorps n'ayant pas réagi. Un anticorps fabriqué contre les immunoglobulines G de lapin et conjugué à la fluorescéine est ensuite incubé sur les coupes. Après lavage les préparations sont observées au microscope à fluorescence.
Afin d'éliminer les risques de marquage non spécifique, les immunsérums sont adsorbés sur une poudre acétonique de foie et de rate de porc I préparée au laboratoire, avant d'être déposés sur coupe de thymus, selon une méthode employée au laboratoire du Pr. DUHEILLE à Nancy (12, 20).
Le titre d'un prélèvement d'immunsérum est défini comme étant la dernière dilution donnant un marquage fluorescent de l'épithélium thymique et notamment des corpuscules de Hassal .
Le protocole détaillé est donné dans l'annexe II.
c - Obtention de microsphères-thymnline
La fabrication du réactif microparticulaire sphérique a été mise au point par le Laboratoire d'immunologie du Pr DUHEILLE de la Faculté de Médecine de Nancy et reste confidentielle. Le procédé est protégé par un brevet appartenant au groupe SANOFI-RECHERCHE (MONTPELLIER-PARIS).
Les microsphères sont obtenues par polymérisation sous rayonnement gamma de quatre réactifs. Le NN'méthylène bisacrylamide
CH2 = CH - C - NH - CH2 - NH - C - CH = CH2 Il II 0 0
sert d'agent réticulant pour les trois autres gui apportent chacun un atout aux microsphères :
- stabilité de par la fonction acide apportée par l'acide méthacrylique
CH2 - CH - COOH I CH3
- hydrophilie grâce à la fonction hydroxyle de l'hydroxyéthylméthacrylate
42

CH2 = CH - C - CH2 - CH2 - OH 1 Il CH3 0
- réactivité avec la fonction aldéhyde de l'acroléine
CH2 = CH - C=O 1 H
Il est possible d'obtenir diffirents diamètres de sphère (50 B 300 ml et leur taille peut être mesurée, après fabrication, au microscope électronique à balayage.
Les microsphères sont stables en suspension, hydrophiles et facilement couplées à l'antigène voulu.
Leurs fonctions alddhydes permettent un couplage cavalent de molécules biologiques possédant des fonctions amines primaires, par formation de liaisons imines :
C=O + NH2-R ---------- > =N- R + H20
La présentation de l'antigène sur les microparticules gouverne l'accessibilité de ses déterminants antigdniques aux sites anticorps. Ce paramètre a fait l'objet d'études systématiques par le Laboratoire de Nancy quant aux quantités de réactifs a mettre au couplage, et nous avons pu au départ suivre un protocole déjà prêt que nous avons adapté au cas de la thymuline.
Les microsphères natives, consenrées en milieu antioxydant, sont dialysées dans du tampon puis simplement incubées avec l'antigène pendant plusieurs heures. Ensuite les fonctions aldéhydes des microsphères n'ayant pas réagi sont saturées par de l'éthanolamine ( NH2 - CH2 - CH2 - OH ) par l'intermédiaire de sa fonction amine primaire. Cela permet de ne pas faire varier la charge des microsphères et d'en renforcer l'hydrophilie grâce à la fonction hydroxyle de l'éthanolamine.
Les ligands non couplés sont éliminés par dialyse si leur poids moléculaire est assez petit ( < 10 000 D ) ou par ultracentrifugation sur gradient discontinu de saccharose si leur poids moléculaire est élevé.
Les microsphères sont enfin dialysées dans du tampon dans lequel elles sont conservées à +4V.
A chaque étape le signal néphélémétrigue des microsphères est contrôlé sur l'appareil HYLAND gui détecte de façon très sensible l'agglutination des microsphères de diamètres autour de 100 nm. Cela permet
43

de vérifier la stabilité des microsphères au cours du couplage. Une augmentation du signal des microsphères seules indique une agglutination spontanée et nécessite une modification de l'étape impliquée.
Une autre possibilité de couplage consiste à intercaler une protéine entre la microsphère et la thymuline.
La protéine intermédiaire est fixée sur les microsphères natives par simple incubation puis élimination de Ifexcès comme décrit ci dessus. Ensuite les microsphères sont dialysées contre une solution de NaCl 0.9%, car le phosphate gène la réaction suivante. Les fonctions carboxyles de la protéine sont activées par l'ajout de carbodiimide sur les microsphères couplées. La thymuline est alors introduite et se fixe sur les fonctions activées par l'intermédiaire d'un groupe amine (NH2).
carbodiimide NH-R' ii 1
Protéine-C-OH + R-N=C=N-R1 ----------> protéine-C-O-C=N-R
+ thymuline - NH2
Au pH de la réaction c'est préférentiellement le NH2 en epsilon de la lysine 3 de la thymuline gui réagit ce gui évite de toucher à la partie C-terminale de la molécule.
Après plusieurs heures d'incubation de l'urée est ajouté pour éventuellement saturer les fonctions activées nsayant pas réagi. Les * microsphères sont ensuite dialysées contre du tampon phosphate dans lequel elles sont conservées à +4OC.
En fonction du porteur choisi, certaines adaptations ont été apportées au protocole général; elles sont mentionnées en annexe.
Le protocole détaillé de couplage est donné dans l'annexe III.
d - Milieux réactionnels
Trois types de milieux ont été utilisés pour la mise au point de la réaction en milieu synthétique.
- le tampon PBS : phosphate O.OlM, NaCl 0.15M, pH 7.2
- Na2-HP04 12H20 2.575 g/l - Na-H2P04.H20 0.396 g/l - NaCl a.2 g/l
- le tampon NEPH : phosphate 0.05 M, NaCl 0.33M, pH 7.2
44

- Na2-HP04 12H20 12.9 g/l - Na-H2P04.H20 1.929 gjl - NaCl 19.28 g/l - PEG 6000 30 dl (polyéthylèneglycol) - EDTA 0.56 gjl (éthylènediaminetétraacétigue) - Triton X100 2 dl - NaN3 2 dl (azide de sodium)
- le tampon HEPES (acide(hydroxy-2-éthyl)-4-pipérazinyl-1,2- éthanesulfonique) 0.01 M, NaCl 0.2 M, pH 7.5 et contenant du PEG, de l'EDTA, du Triton X100 et du NaN3 comme le tampon NEPH, est utilisé lorsqu'on veut travailler en milieu tamponné ne chélatant pas les cations métalliques.
Nous reviendrons sur le choix et les caractéristiques de ces tampons par la suite.
La mise au point des réactifs et des conditions de la réaction d'immunonéphélémétrie à supports microparticulaires représente un ensemble d'étapes à franchir pour obtenir dans un premier temps une réaction d'étalonnage en milieu synthétique.
L'absence de méthode de référence disponible rend difficile l'appréciation des réactifs fabriqués tant que l'on n'a pas obtenu une réaction, et donc mis au point plusieurs réactifs performants.
C'est pourquoi le plan de la présentation qui va suivre n'est pas. ordonné de façon à présenter successivement les résultats de chacun des paramètres séparés.
2 - RESULTATS
a - Les ilmmlnsérums
Chronologiquement la plupart des immunsérums ( sauf ceux de la troisième série) ont été fabriqués avant l'obtention du modèle de .microsphères-thymuline gui a permis d'obtenir les premiers, résultats spécifiques.
Pour le suivi de la production d'anticorps anti-thymuline par les lapins il n'y avait donc pas de réaction néphélémétrique envisageable. C'est pourquoi ce suivi a été fait pour les première et deuxième séries par la seule méthode disponible de détection d'anticorps anti-thymuline, l'immunofluorescence indirecte sur coupes de thymus (20).
Mais cette technique est bien différente de celle pour laquelle les anticorps sont fabriqués et ne rend pas compte des propriétés attendues pour l'immunonéphélémétrie, notamment la capacité d'agglutination.
45
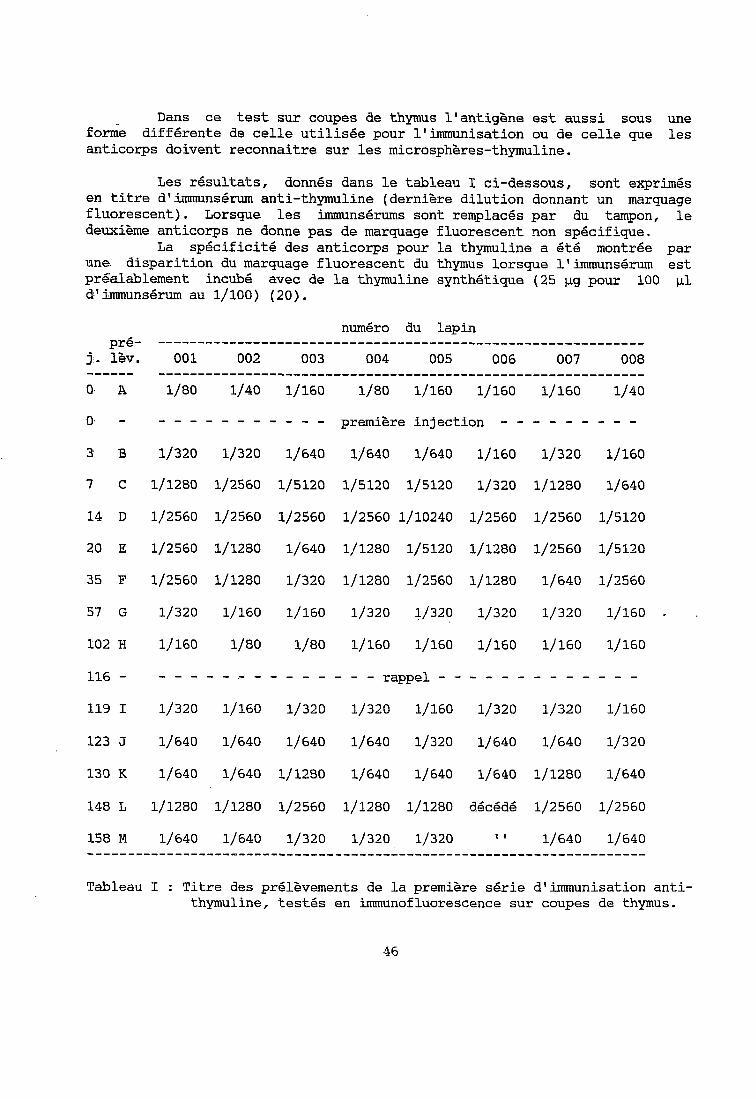
Dans ce test sur coupes de thymus l'antigène est aussi sous une forme différente de celle utilisée pour l'immunisation ou de celle que les anticorps doivent reconnaitre sur les microsphères-thymuline.
Les résultats, donnés dans le tableau I ci-dessous, sont exprimés en titre d'immunsérum anti-thymuline (dernière dilution donnant un marquage fluorescent). Lorsque les immunsérums sont remplacés par du tampon, le deuxième anticorps ne donne pas de marquage fluorescent non spécifique.
La spécificité des anticorps pour la thymuline a été montrée par une disparition du marquage fluorescent du thymus lorsque l'immunsérum est préalablement incubé avec de la thymuline synthétique (25 pg pour 100 CL1 d'immunsérum au l/lOO) (20).
pré- j:. lèv. ------ 0. A
0, -
3 B
7' c
14 D
20 E
35 F
57 G
numéro du lapin -------------------------------------------------------------
001 002 003 004 005 006 007 008 -------------------------------------------------------------
l/ao 1/40 1/160 l/ao lj160 lj160 1/160 1/40
----------- première injection - - - - - - - - -
1/320 lj320 1/640 1/640 lj640 lj160 lj320 lj160
1/1280 1/2560 1/5120 1/5120 1/5120 1/320 1/1280 1/640
1/2560 112560 1/2560 1/2560 1/10240 1/2560 1/2560 1/5120
1/2560 1/1280 1/640 1/1280 1/5120 1/1280 1/2560 1/5120
1/2560 1/1280 1/320 1/1280 1/2560 1/1280 1/640 1/2560
1/320 1/160 1/160 1/320 1/320 1/320 1/320 1/160
102 H
116 -
119 1
123 J
lj160 l/ao l/ao 1/160 1/160 1/160 1/160 1/160
- - - - - .- - - - - - - - - rappel - - - - - - - - - - - - -
1/320 lj160 1/320 1/320 1/160 lj320 lj320 lj160
1/640 1/640 1/640 1/640 lj320 1/640 1/640 1/320
130 K 1/640 lj640 1/12BO 1/640 1/640 1/640 1/1280 1/640
148 L
158 M
1/1280 lj1280 1/2560 1/1280 1/1280 dkédé 1/2560 1/2560
lj640 lj640 1/320 1/320 lj320 1 1 1/640 1/640 ----------------------------------------------------------------------
Tableau 1 : Titre des prélèvements de la première série d'immunisation anti- thymuline, testés en immunofluorescence sur coupes de thymus.
46

D'après ce test les lapins de la Premiere série semblent avoir produit des anticorps anti-thymuline, notamment aux jours 7, 14, 20 et 35 après la première injection et au jour 148, 32 jours après le rappel. Il est à noter que les huit lapins ont répondu de la même façon. La réponse après le premier rappel est moins importante qu'après la première injection. Les derniers prélèvements, N, 0, P et Q, n'ont pas été testés sur coupes de thymus.
La deuxième série ne concerne que trois lapins. Les résultats sont donnés dans le tableau II ci-dessous. La réponse est très faible après le deuxième rappel et commence à augmenter après la série des rappels 3 à 11. Les titres des prélèvements de cette série d'immunisation sont inférieurs à ceux de la première série. Le prélèvement 050 C présente le titre en anticorps le plus élevé.
numéro du lapin prélè- -------------------------------
jour vement 050 060 070 ------ ------ -------------------------------
0 A l/lOO l/lOO 1/40
1-8-15 - - lère injection et rappels 1 et 2
20 B 1/200 lj200 lj200
22 à 40 - - - - - rappels 3 a Il - - - - -
43 C 1/3200 l./800 1/800 -.
46 D 1/800 ------------------------------------------------
Tableau II : Titre des prélèvements d'immunsérums anti-thymuline de la deuxième série d'immunisation, testés en immunofluoresence
indirecte sur coupes de thymus.
b- Premier couplage sur les microsphères
Sauf mention différente, toutes les réactions présentées ici ont été obtenues avec des microsphères de 105 nm de diamètre et lues sur le néphélémètre BEHRING.
Lors des premiers essais de microsphères sensibilisées par la thymuline, nous avons fixé la,molécule directement sur la microsphère native. La microsphère-thymuline ainsi obtenue était stable en milieu synthétique mais présentait une grande instabilité et une importante agglutination en présence de sérum (immunsérum ou non). Cette agrégation
47

n'était pas inhibée par la thymuline synthétique libre. Ce type de réactif n'était donc pas exploitable dans ces
conditions. Cela était vraisemblablement dû à une couverture protéique
insuffisante de la microsphère, à cause de la petite taille de la thymuline. En effet la microsphère doit présenter deux propriétés contradictoires : être suffisamment stable pour rester en suspension et suffisamment instable pour s'agréger fortement en présence de quantité minime d'agent agglutinant. Dans le cas des microsphères-thymuline la déstabilisation était produite non spécifiquement par un composant du sérum, faisant intervenir vraisemblablement des forces électrostatiques et des charges d'ions puisque cette agglutination non spécifique était, par exemple, inhibée par l'ajout de chlorure de zinc (ZnC12) dans le milieu.
Ces premiers résultats nous ont conduits à envisager de sensibïliser les microsphères en les recouvrant d'abord d'une protéine intermédiaire enrobant complètement la microsphère, et sur laquelle la thymuline sera ensuite greffée chimiquement.
L'accessibilité et le microenvironnement de la thymuline dépendent de la protéine choisie. Nous présentons les résultats pour trois porteurs.
c - Microsphères-SAB-thymAine
La sérum-albumine bovine (SAB) a d'abord été choisie cormne protéine intermédiaire car elle avait déjà été utilisée dans d'autres systèmes par le Laboratoire d'immunologie du Pr. DUHEILLE de la Faculté de Médecine de Nancy. La SAB est apportée au couplage à raison de 2.10-8 moles pour 10 mg de microsphères.
Le premier couplage de la thymuline a été fait avec 10-7 moles apportées pour 10 mg de microsphères-SAB. Ces microsphères-SAB-thymuline (10-7) se sont avérées stables car elles n'étaient pas agglutinées en présence d.'immunsérums non spécifiques. Elles ont été alors testées en tampon PBS, avec les prélèvements disponibles les plus tardifs de la première série d'immunisation, correspondant à priori au meilleur moment de production des anticorps anti-thymuline. La réaction est lue sur le néphélém‘étre HYLAND.
Les microsphères-SAB-thymuline (10-7) ont été les premières à donner, avec le prélèvement 003L, une agglutination spécifiquement inhibée par la thymuline synthétique.
Les courbes de la figure 1 présentent les résultats d'agglutination et d'inhibition. Le maximum d'agglutination est obtenu pour l'immunsérum au lj800. La gamme d'inhibition s 'Qtend de 1 à 100 ng/ml environ.
Par l'obtention d'une agglutination spécifique des microsphères- SAB-thymuline nous savions gue le type de microsphère choisi, avec porteur protéique, convenait et que nous possédions des immunsérums spécifiques de la thymuline et agglutinants.
48

4 T AGGLUTINATION
0-l , I I 1 1 I I 4 50 100 200 400 800 1600
inverses des dilutions de I’immunserum
INHIBITION
0- 1/250
2 -- oy3L5 lpgo
=
I/l0
0 10-310-210-1 1 10 10+210+310+410+5
Thymuline en nanogrammes par millilitre
Figure 1 : Agglutination et inhibition avec les microsphères-SAB-thyline 10-7 et l'immunsérum 003L, en tampon PBS. Mesures faites sur
un néphélémètre HYLAND.
49

Cette première réaction a été effectuée en tampon PBS et mesurée avec un néphélémètre HYLAND utilisé jusque là pour tous nos essais. La cinétique est lente, mais il est possible de tenter de l'améliorer en faisant varier la concentration de thymuline apportée au couplage.
Nous avons donc cerné la concentration précédenunent utilisée, et fait des couplages sur les microsphères-SAB avec de la thymuline apportée à raison de 10-6, 5.10-7 et 10-8 moles pour 10 mg de microsphères-SAB.
A partir de maintenant, nous travaillons en tampon NEPH dont nous étudierons les qualités dans un chapitre ci-après. Toutes les microsphères- SAB-thymuline seront testées avec le prélèvement d'immunsérum 003L.
La concentration de 10-8 moles de thymuline au couplage est trop faible puisque les microsphères-thymuline ne donnent aucune agglutination.
Les courbes suivantes présentent les résultats d'agglutination et d'inhibition pour les microsphères fabriquées avec les concentrations de 10-6 et 5.10-7 moles de thymuline.
Pour 10-6, les résultats sont donnés pour les réactions mesurées sur les appareils HYLAND et BEHRING (figures 2 et 3). Ce dernier est l'appareil utilisé ensuite pour la lecture de toutes les réactions.
A l'agglutination l'allure des 2 courbes est semblable l/;OO
sans "cloche" et avec une pente descendante rapide entre les-dilutions et 1/400 et entre l/lOO et 1/400 de 003L respectivement pour les réactions lues sur les néphélimètres HYLAND ou BEHRING.
La gamme dtinhibition se situe entre 5 et 625 ng/ml avec 50 % d'inhibition pour 78 ng/ml de thymuline pour la réaction lue sur le HYLAND (figure 2). Ces valeurs sont respectivement aux environs de 10, 300 et 40 ng/ml de thymuline pour la réaction lue au BEHRING (figure 3).
Avec moins de thymuline apportie au couplage (figure 4), c'est à dire 5.10-7 moles pour 10 mg de microsphères-SAB, le signal à l'agglutination est moins important que précédemment pour les microsphères utilisées à la même dilution (figure 3).. Par contre la courbe a une allure "en cloche" et la pente descendante se situe entre les dilutions, l/lOO et k/800 de 003L.
L'inhibition couvre une gamme allant de 5 à 100 ngjml avec 50 % d'inhibition entre 12 et 37 ng/ml de thymuline.
La cinétique est améliorée puisqu'on a une réaction correcte en 1 heure pour les microsphères-SAB-thymuline (10-6) et en 3 heures pour les microsphères-SAB-thymuline (5.10-7), dont la sensibilité à l'inhibition est légèrement améliorée.
Pour la suite nous avons retenu ces deux types de microsphères-SAB- thymuline pour différents tests. <. . . ,. :
50 .

I (XRLS)
50 100 200 400 800 1800 3200 6400
Inverses des dilutions de I’immunserum
1 (XRLS)
1504 INHIBITION
o- 1/200
OOT :‘ioo =
0-l 1 I I i 0 0.6 1.2 2,3 4.9 9,8 19 39 78 156 312 625
lhymuline en nanogra~mes par millilitre
Figure 2 : Agglutination et inhibition avec les microsphères-SAB-thyline 10-6 et l'immunsérum 003L, en tampon NEPH. Mesures faites sur
un néphélémètre HYLAND.
51

AGGLUTINATION
n I * t I * 4 46 96 192 384 768 1636 3072 6144
inverse8 do8 diluffons de i’immunaerum
v0lts/100
INHIBITION 100..
50.-
o- 1/200
**T X0" =
04 I I s I I t , . . I 0 0,s 1.5 4,5 13,7 41 123 370 1110 3333
Thymuhe en nanogmmmes paf milliRr0
Figure 3 : Agglutination et inhibition avec les microsphères-SAB-thyline 10-6 et l'immunsérum 003L,en tampon NEPH. Mesures faites sur
un néphélémètre BEHRING.
52

volts/1 00
AGGLUTINATION
04 * I I t 48 96 192 304 766 1536 3072 6144
inverses des dilutions de I’immunserum
i.
“8 o,i5 0,k i,i7 4:1 ii,3 ;ilr iii 3b Goo
Thymuline en nanogmmmes par millilitre
Figure 4 : Agglutination et inhibition avec les microsphères-SAB-thymuline 5.10-Y et l'irmnunsérum 003L, en tampon NEPH. Mesures faites sur
un néphélémètre BEHRINC. / ._ : '. 1
.53 I ,

Dans le but d'amBliorer les qualit& de la réaction, le couplage d'un homologue Lys-thymuline de 10 acides aminés a été effectué sur les microsphères-SAB. La fixation par la partie N-terminale, rendue possible par l'ajout d'une lysine, pouvait modifier la prgsentation de la thymuline sur la microsphère, et donc sa réaction avec les anticorps.
Cet homologue a été apporté à raison de 10-6 moles par 10 mg de microsphères-SAH au couplage.
Avec l'immunsérum 003L (figure S), l'agglutination n'est pas très différente de celle obtenue avec les microsphères-SAB-thymuline (10-6) (figure 3). Par contre l'inhibition par la thymuline synthétique est moins sensible puisque la gamme de concentrations couvertes est comprise entre 120 ng/ml et 3 pg/ml environ avec 50 % d'inhibition vers 1 pg/ml de thymuline.
volts/1 00
200 T AGGLUTINATION
04 1 I t 48 96 192 384 768 1536 3072 6144
inverses das dilutions de I’immunserum
volts/1 00
200 T INHIBITION
150.-
100~-
0. t t , i 0 0,5 1,5 4,5 13.7 41 123 370 11103333
Thymuline en nanogrammss par millilitre
Figure 5 : Agglutination et inhibition avec les microsphères-SAB-homologue 10-6 et l'immunsérum 003L, en tampon NEPH. Mesures faites sur
un néphélémètre BEHRING.
54

L'emploi de, cet homologue de la thymuline modifie bien les résultats mais nous ne l'avons pas retenu car ceux-ci sont moins bons qu'avec la thymuline.
L'inhibition par l'homologue Lys-thymuline utilisé au couplage n'est pas intéressante puisque ce n'est pas cette molécule, différente de la thymuline naturelle, que nous voulons atteindre avec le dosage dans les échantillons biologiques.
Les microsphères-SAB-thymuline retenues ont permis dans un premier temps de sélectionner les immunsérums réactifs et exploitables en néphélémétrie. Ces prélèvements ont servi à évaluer la qualité des différents types de microsphères ou des conditions réactionnelles au cours des mises au point.
d - Test du stock d' inununsérums avec les microsphères-!3AE+thymuline
Tous les prélèvements d'immunsérum des séries d'immunisation 1 et 2 ont été testés avec les microsphères-SAB-thymuline. Ce travail a été fait à Nancy dans le cadre de la collaboration avec le Laboratoire d'immunologie du Pr. DUHEILLE à la Faculté de Médecine. La qualité des prélèvements a été évaluée selon le titre à l'agglutination, la dilution à employer pour l'inhibition et la sensibilité de l'inhibition par la thymuline synthétique.
Les résultats sont regroupés dans le tableau III ci-après. Le code (-) correspond à une absence totale de réaction et le nombre de signes (+) permet de comparer qualitativement les prélèvements.
Les constatations suivantes peuvent être faites pour la première série : aucune réponse anti-thymuline n'est détectée après la première injection pour les huit lapins. Deux semaines après le premier rappel (J 130) un seul lapin, le 003, commence à donner une réponse anti-thymuline positive. Parmi les six lapins répondants 32 jours après le premier rappel, le meilleur à l'agglutination est le 003, le moins bon est le 007. Le second rappel (J 187) permet à trois lapins seulement (002, 003 et 008) de continuer à présenter une réponse agglutinante anti-thymuline. Ces trois mêmes lapins répondent après le troisième rappel (J 306). Le titre à l'agglutination est amélioré seulement pour le lapin 002.
Les tests d'inhibition réalisés pour les prélèvements agglutinants ont montré que tous, sauf les 002, sont inhibés par la thymuline synthétique à des concentrations inférieures à 500 ng/ml.
Sur huit lapins tous les types de réponse ont été observés. Un lapin n'a jamais produit d'anticorps (le n"5) et un autre est mort (le n06). Le lapin donnant les meilleurs prélèvements est celui gui a répondu le plus tôt (le n03). Pour trois lapins (les no 1, 4 et 7) la réponse obtenue après le premier rappel a disparu ensuite. Pour 2 lapins (les no 2 et 8) la réponse s'est améliorée au cours des rappels.
55
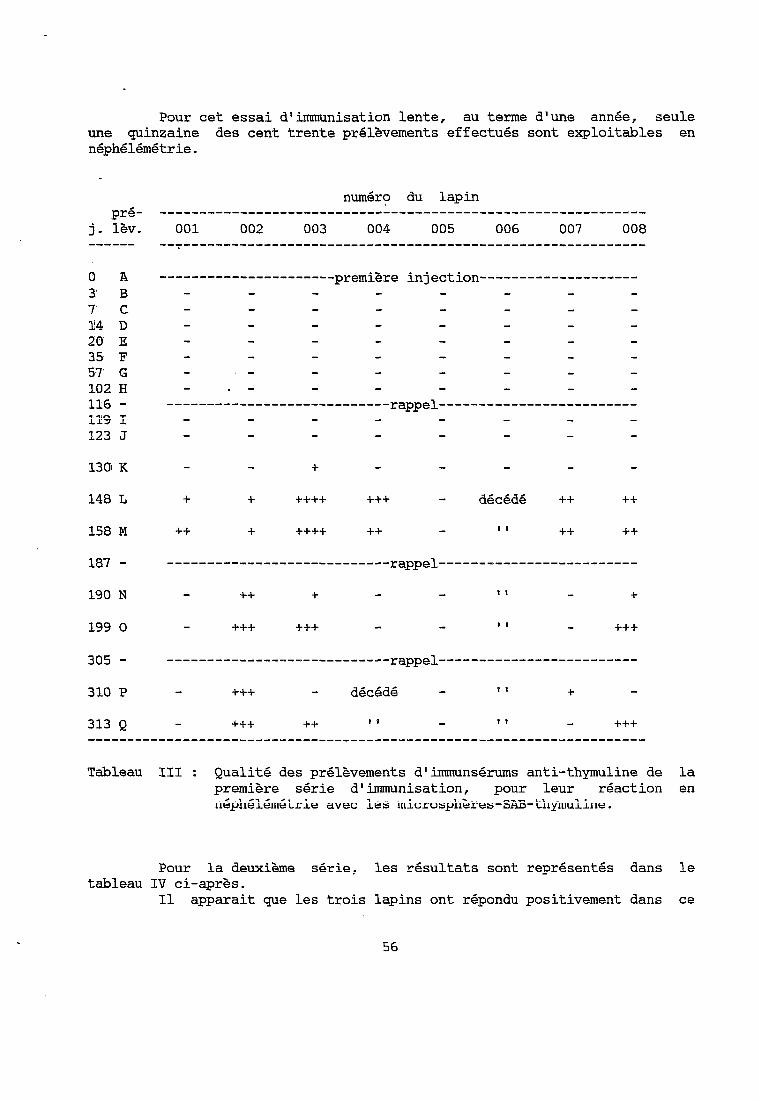
Pour cet essai d'immunisation lente, au terme d'une année, seule une quinzaine des cent trente prélèvements effectués sont exploitables en néphélémétrie.
pré- j. lèv. ------
0, A 3' B 7' c 14 D 20 E 35, F 57 G 102 H 116 - 119 1 123 J
130 K
148 L
158 M
la7 -
190 N
199 0
305 -
310 P
313 Q
numéro du lapin -------------------------------------------------------------
001 002 003 004 005 006 007 008 -------------------------------------------------------------
----------------------première injection--------------------
-
. -
----------------------------rappel-------------------------
+
+ + ++++ +++ décédé f+ ++
++ + ++++ ++ I 1 ++ ++
----------------------------rappel----------------------------
++ + I I +
+++ +++ 1 1 +++
----------------------------rappel-------------------------
+++ décédé - 1 1 +
+++ ++ I 9 1 1 +++ ----------------------------------------------------------------------
Tableau III : Qualité des prélèvements d'immunsérums anti-thymuline de la première série d'immunisation, pour leur réaction en néphélémétrie avec les microsphères-SAB-thymuline.
Pour la deuxième série, les résultats sont représentés dans le tableau IV ci-après.
Il apparait que les trois lapins ont répondu positivement dans ce
56

test mais seulement pour les prélèvements réalisés en fin du protocole d'immunisation. Ces réactions sont difficilement inhibées et nécessitent environ 10 ug/ml de thymuline, soit beaucoup plus que pour les réactions obtenues avec la première série. Ces immunsérums apparaissent spécifiques de la thymuline mais la réaction d'inhibition est très peu sensible. Sur trois lapins le protocole rapide d'immunisation a donné de moins bons résultats.
numéro du lapin -----------------------------
050 060 070 jour prélèvement -----------------------------
1 ----------première injection------
a et 15 ----------rappels no 1 et 2-------
20 B
22 à 40 ----------rappels n0 3 à 11-------
43 C + + +
46 D + t t 182 E t t t
183 ------------ rappel n0 12 --------
187 F + + + ------------------------------------------------------
Tableau IV : Qualité des prélèvements d'immunsérums anti-thymuline. de la deuxième série d'immunisation, pour leur réaction en néphélémétrie avec les microsphères-SAB-thymuline.
Ces résultats montrent la différence d'appréciation des immunsérums selon la méthode employée. Par immunofluorescence sur coupes de thymus, des anticorps anti-thymuline avaient été détectés, à des titres importants, après la première injection, alors qu'aucun de ces pr81èvements n'agglutinent les microsphères-SAB-thymuline. Après le premier rappel les résultats sont déjà plus concordants mais il n'y a pas de corrélation correcte permettant de valider le test sur coupes de thymus pour le suivi de la production d'anticorps destinés à la néphélémétrie.
Les résultats de la comparaison des performances des immunsérums en immunonéphélémétrie avec les microsphères-SAB-thymuline ont amené à choisir les meilleurs d'entre eux pour tester ensuite les différentes microsphères au cours de leur mise au point.
Ainsi ont principalement été utilisés les immunsérums :
57

----em
003L
003M
0030
003Q
002P
osoc
bonne agglutination ---------------------
++
-!-+
+-l-+
+
++
+
inhibition sensible ------------------^--
++
++
++-t+
++
---------------------------------------------------
Les prélèvements 002P, OSOC et même 0035) ont été utilisés pour les seules comparaisons de réactions d'agglutination.
Tous ces résultats ont été obtenus avec les microsphères-SAS- thymuline. Mais le choix définitif de la SAB comme protéine intermédiaire dans un système de dosage n'est pas recommandé car il peut exister des anticorps anti-SAB dans les échantillons sériques à doser, ce gui interfèrerait avec la mesure de la thymuline.
D'autre part au cours de la mise au point du dosage nous avons cherché à améliorer la sensibilité de la réaction d'inhibition. Ceci peut être envisagé par l'essai de nouveaux porteurs protéiques entre la microsphère et la thymuline.
Nous avons donc testé de nouvelles protéines intermédiaires.
e - Essais de nouvelles protéines intermédiaires de couplage
La molécule protéique doit présenter des propriétés réactionnelles c'est à dire posséder des groupements carboxyles ou des amines primaires par exemple.
Après greffage, l'excès de protéine est éliminé par simple passage au travers d'un sac de dialyse pour les poids moléculaires inférieurs à 10.000 D, mais nécessite une ultracentrifugation en gradient de saccharose, suivie d'une dialyse pour les poids moléculaires supérieurs.
Le protocole est donc plus rapide et plus simple lorsque la protéine intermédiaire a un poids moléculaire inférieur à 10 000 D.
e-l . L'acide polyaspartique
Un polymère de l'acide polyaspartique, possédant une centaine de fonctions carboxyles, semblait répondre aux critères de choix.
Son poids moléculaire de 15 000 D ne permettait pas de l'éliminer
58

par simple dialyse. Pour éviter l'ultracentrifugation nous avons choisi d'utiliser dans
un premier temps des microsphères d'un diamètre de 300 nm gui peuvent décanter sur une centrifugeuse classique à 12 000 tours/mn et être débarrassées de l'excès d'acide polyaspartique par centrifugations et lavages successifs.
Pour le premier couplage 2.10-8 moles d'acide polyaspartigue et 2.10-7 moles de thymuline ont été apportées pour 10 mg de microsphères.
Les résultats obtenus avec l'immunsérum 003L (figure 6) montrent une importante agglutination encore présente à une dilution au l/lOOO du 003L. L'inhibition couvre une gamme allant de 1 à 100 ng/ml environ avec 50 % d'inhibition vers 20 ng/ml de thymuline.
Ces résultats encourageants, un peu meilleurs qu'avec les microsphères-SAB-thymuline, nous ont incités à développer le modèle.
Dans le but d'améliorer la sensibilité de 'la réaction à l'inhibition, les couplages ont été poursuivis avec des microsphères de 105 nm de diamètre. Après couplage l'élimination de l'acide polyaspartique n'ayant pas réagi a été faite par une ultracentrifugation sur gradient discontinu de saccharose.
Dans le cas d'un homopolymère comme l'acide polyaspartigue, il était souhaitable d'essayer aussi différentes doses & couplage. Des protocoles ont été prévus avec deux concentrations d'acide polyaspartigue et- pour chacune trois concentrations de thymuline. Soit au total six types de microsphères-polyaspartigue-thymuline dénommées avec les abréviations données dans le tableau suivant :
-----------__---_--_------------------------------------------ THYMULINE
(moles/10 mg de microsphères) -------------------------------
----------
POLYASPARTIQUE
(moles/10 mg de microsphères)
10-6 10+ 10-=
------------------------------
R7F6 R7F7 R7F8
R8F6 R8F7 R8F8
---------------------------------------------------------------
59
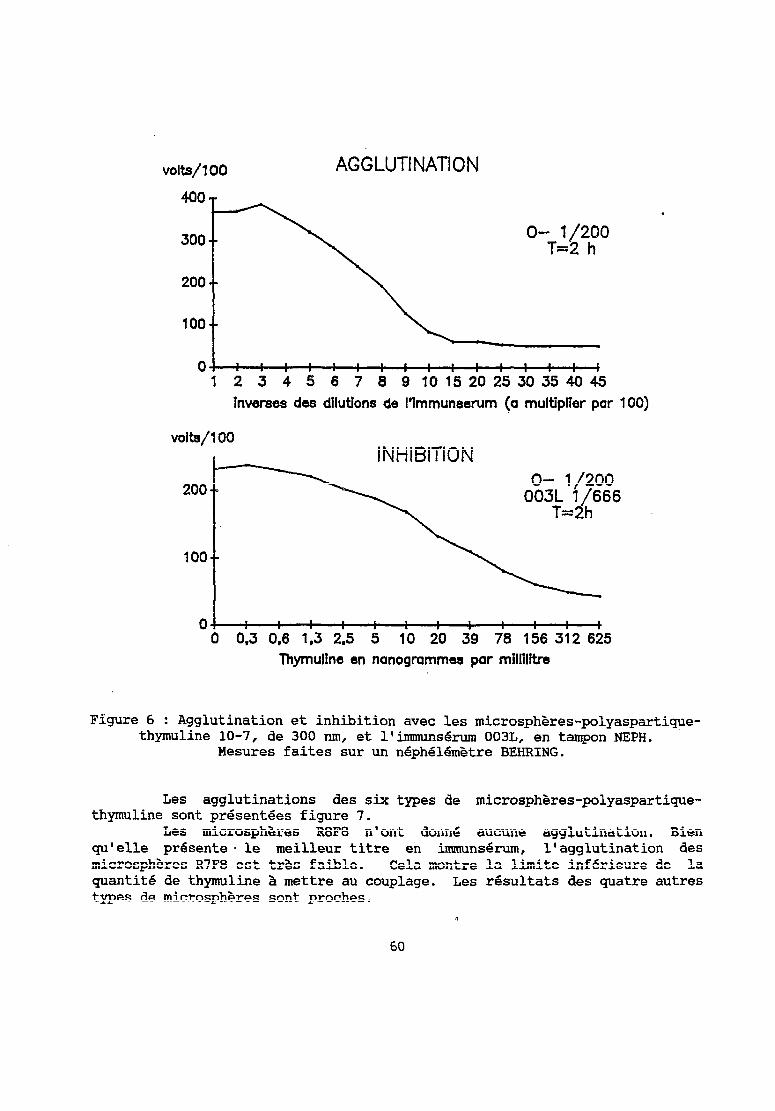
volts/1 00 AGGLUTINATION
o!:::::::::::::::t 1 2 3 4 5 6 7 8 9 10 15 20 25 30 35 40 45
invwees des dilutions de I’immuntwum (a multiplier par
volt8/lOO
I - INHIBITION
100)
04 1 0 0.3 0,6 1,3 2.5 5 10 20 39 78 156 312 625
Thymullns en nanogrammea par millîlitm
Fïgure 6 : Agglutination et inhibition avec les microsphères-polyaspartique- thymuline 10-7, de 300 nm, et l'immunsérum 003L, en tampon NEPH.
Mesures faites sur un néphél&mètre BEHRING.
Les agglutinations des six types de microsphères-polyaspartigue- thymuline sont présentées figure 7.
Les microsphères R8F8 n'ont donné aucune agglutination. Bien qu'elle prkente. le meilleur titre en immunsérum, l'agglutination des microsphères R7F8 est très faible. Cela montre la limite inférieure de la quantitk de thymuline à mettre au couplage. Les résultats des quatre autres types de microsphères sont proches.
60

60
20
60
invenma des diluüons
R7F7 lnveraas des dilutions
inverses des diluffons
I Y0lts/100 R7F8
inverses dea dilutiona
inversea des dilutions
R8F8
PAS DE REACTION
Figure 7 : Agglutination des cinq types de microsphères- polyaspartique- thymuline au l/lOO avec l'immunsérum 003 L, en tampon NEPH. Mesures
faites sur un néphélémètre BEHRING.
61

La figure 8 montre que pour obtenir un signal équivalent, avec les microsphères couplées avec des concentrations plus faibles de thymuline et de polyaspartigue, il faut augmenter la concentration des microsphères dans la cuve et le temps de lecture de la réaction.
invm dea dilutions
R7F6 1/2OO,T=90mn
R7F7 1/200,T=90mn
R7F8 1/5O,T4 20mn
R8F6 1/200,T=300mn
R8F7 1 /l OO,T=300mn
Figure 8 : Comparaison des conditions opératoires à l'agglutination des cinq types de microsphères-polyaspartigue-thymuline avec l'immunsérum 003L en
tampon NEPH. Mesures faites sur un néphélémètre BEBRING.
Les microsphères les moins sensibles à l'inhibition (figures 9 et 10) sont greffées avec 10-6 moles de thymuline par 10 mg de microsphères (R7F6 et R8F6), ce gui est donc une concentration trop élevée.
Les performances des 3 autres lots, R7F7, R7F8 et R8F7, sont assez comparables et permettent une gamme d'inhibition de 1 à 100 ng/ml de thymuline avec 50 % d'inhibition pour environ 15 ngjml Pour une dilution de travail correcte, seules les microsphères-polyaspartigue greffées avec 10-7 moles de thymuline (R7F7 et R8F7) conviennent.
62

I I I I . I . abe lb I%f46 137 43 1&3 87 141 * ldlm
Thymuline (ng/ml)
1 200 l d
200 5 mn
_ _ _
20 “I
Thyniuliné
volts/1 00 hiw
R7FB
o- 1/po
5( Thymulim (ng/ml) ’ ’ ’
15 vdts/~oo R8F7
R8F8
Figure 9 : Inhibition avec les cinq types de microsphères-polyaspartique- thymuline et l'immunsérum 003L, en tampon NJZPH. Mesures faites sur
un nGphélémètre BEHRING. .,
63

vottfJ,‘l 00 - R7F6
00x 11200 11200
T=40mn
+ R7F7 1,‘200 00x 1/200 T=l50mn
- R7F8 003L
1140 11320
T=GOmn
8 R8F6 0031
11300 11400
T=l50mn
* R8F-7 003L
11100 11200
T=l50mn
01 I , 1 I < I I 8 l I t 6 9 I I 1 8 I 0 0.05
1 1.5 0.45 1,37 4.1 12,3 37 111 333 1000
Thymuflm h/mO
Figure 10 : Comparaison des conditions opkatoires des cinq types de microsphères-polyaspartique-thymuline à l'inhibition avec l'immunsérum 003L
en tampon NEPH. Mesures faites sur un néphélémètre BEHRING.
La comparaison des résultats obtenus avec les microsphères- polyaspartique-thymuline de 300 nm et de 100 nm de diamètre est concordante avec la théorie : pour un plus petit diamètre, l'agglutination est moins bonne mais l'inhibition est améliorée (de 50 à 15 ng/ml de thymuline pour 50% d'inhibition).
Les microsphères-polyaspartique-thymuline donnent donc des résultats un peu meilleurs que les microsphères-SAB-thymuline.
64

Après l'étude des doses au couplage dans ce système, leur influence sur le résultat et les performances des meilleures réactions, un autre porteur protéique a été testé.
e-2 . La métallothionéine
La métallothionéine est une protéine d'un poids moléculaire de 6.400 D, fixant les minéraux.
Son faible poids moliculaire permet une dialyse au lieu d'une ultracentrifugation pour l'éliminer après couplage.
Sa capacité de fixer le zinc pourrait modifier la conformation de la thymuline, et peut-être la réactivité des microsphères.
Des essais de couplage ont été faits en apportant la métallothionéine aux doses de 8.10-8, 2.10-8 et 5.10-q moles et.la thymuline à 10-7 moles pour i0 mg de microsphères de 105 nm.
Les microsphères-métallothionéine-thymuline ont été testées avec l'immunsérum 003M et non 003L comme les deux types de microsphères précédents.
Les microsphères greffées avec 8.10-8 moles de métallothionéine n'ont pas donné d'agglutination.
La figure 11 présente les courbes d'agglutination et d'inhibition des microsphères fabriquées avec les deux autres doses de métallothionéine. L'agglutination obtenue avec les microsphères-métallothionéine(2.10-8)- thymuline est forte et présente un maximum pour l'immunsérum au l/lOO. Avec les microsphères-mGtallothionéine(S.lO-9)-thymuline la réaction est plus faible et le maximum se situe au 1/200.
Ces deux types de microsphères permettent d'obtenir des sensibilités identiques à l'inhibition, soit une gamme située entre 0.15 et - 40 ng/ml avec 50 % d'inhibition pour 4 ng/ml de thymuline.
Ce résultat est meilleur que ceux obtenus jusqu'à présent avec les microsphères greffées par la SAB ou l'acide polyaspartique.
La plus forte réactivité des microsphères faites avec la concentration 2.10-8 moles de métallothionéine permet d'employer moins d'immunsérum et de microsphères à l'inhibition.
La dose de métallothionéine au couplage de 2.10-8 moles pour 10 mg de microsphères est donc retenue.
Les microsphères-métallothionéine(2.10-8)-thymuline ont été testées avec le prélèvement suivant d'immunsérum du même lapin obtenu après un rappel : le 0030.
La figure 12 montre une inhibition entre 5 et 10.000 pg/ml de thymuline avec 50 % d'inhibition autour de 200 pg/ml apportant un gain d'un facteur de 10 par rapport au résultat précédent.
65

1. t
O& 1 I I I t
9’s lé2 3$ 4 766 lti630726lb
hvemss des diltions
1 7 volts/1 00 inhibition
.I I t I I I a I b O,b5 0,15 0,45 1.57 4,l 12.3 1000
nanogmmmea do Thymullne/ml
Métallothionéine 2.10-8
Figure 11 : Agglutination et métallothionéine-thymuline 2.10-8 et
inhibition avec les microsphères- 5.10-9 et l'immunsérum 003L en tampon
NEPH, au temps 2h. Mesures faites sur un néphélémètre BEHFZNG.
volte/1 00
agglutination
o-l/lOO
Inverses des dilutions
volts/1 00 in hi bition
nanogtammaa de ThymuRne/ml
HétallothionBine 5.10-g
66

60
15 45 137 4il 1233 picogmmmes de Thymuline/ml
Figure 12 : Inhibition avec les microsphères-métallothionéine-thymuline et l'immunsérum 0030 en tampon NEPH.
Mesures faites sur un néphélémètre BEHRING.
Cette courbe était la première exprimge en picogrammes de thymuline par millilitre et montre le gain de sensibilité apporté aussi bien par le changement de protéine intermédiaire sur les microsphères que par le changement d'immunsérum.
Avec les deux prélèvements d'immunsérums 003M et 0030, les microsphères-métallothionéine-thymuline donnent les inhibitions les plus performantes. Ces microsphères ont donc été utilisées pour la suite de la mise au poinf des paramètres du dosage.
67

f,- Le milieu réactionnel
L'optimisation du dosage passe par le choix d'un milieu réactionnel de composition bien définie, car il influence la stabilité des microsphères, la conformation de la thymuline et sa liaison avec les anticorps.
Les immunoglobulines de l'immunsérum doivent, pour agglutiner les microsphères, se rapprocher puis se fixer. Cette agglutination s'accompagne de phénomènes de solvatation et impliquent des forces électrostatiques dues aux charges des microsphères et des anticorps. Ces charges et répulsions peuvent être modulées selon la composition du milieu.
Un milieu séquestrant le zinc a été choisi pour doser la totalité du peptide sans se préoccuper des diverses formes pouvant exister en fonction de la fixation d'un atome de zinc sur la thymuline. Le pouvoir chélatant est assuré par 1'EDTA bien que les phosphates du tampon séquestrent également le zinc.
Deux tampons usuels de la néphélémétrie sont le PBS et le tampon NEPH dont les compositions sont rappelées ci dessous :
- le tampon PBS : phosphate O.OlM, NaCl O.lSM, pH 7.2
- Na2-HP04 12H20 2.576 g/l - Na-H2P04.H20 0.396 g/l - NaCl 8.2 g/l
- le tampon de néphélémétrie (tampon NEPH) : phosphate 0.05 M, NaCl 0.33M, pH 7.2
- Na2-HP04 12H20 12.9 g/l - Na-H2P04.H20 1.929 g/l - NaCl 19.28 g,'l - PEG 6000 30 411 (polyéthylèneglycol) - EDTA 0.56 g/l (éthylènediaminetétraacétique) - Triton X100 2 41 - NaN3 2 dl (azide de sodium)
Des phosphates mono- et disodigues sont la base du tampon PBS et permettent d'avoir un pH de 7.2. Du chlorure de sodium est ajouté à une concentration identique à celle des milieux biologiques.
Le tampon NEPH comporte les mêmes phosphates pour assurer le pH de 7.2. Le pH agit aussi sur la charge des microsphères et des anticorps et peut influer sur l'agglutination. Le chlorure de sodium ajouté en plus grande quantité, augmente la force ionique, ce gui diminue la mobilité des moîécules et stabilise aussi le complexe antigène-anticorps. Ce tampon comporte des éléments supplémentaires. Le polyéthylèneglycol 6000 gui se substitue aux molécules d'eau entourant les anticorps, les rend moins solubles et favorise leur fixation sur l'antigène des microsphères. Une fois les complexes formés, le PEG 6000 les rend plus stables dans le temps et
68

limite leur croissance 6vitant ainsi la 4dimentation de trop gros agrhgats. L'EDTA dissocie vraisemblablement le zinc de la thymuline. Le Triton X100 est un agent mouillant gui facilite un mélange homogène dans la cuve et évite les adsorptions sur les parois. L'azide de sodium permet de prévenir le développement de contaminations.
Les courbes de la figure 13 permettent de comparer les agglutinations obtenues dans chacun des deux tampons. La réaction en PBS est terminée au bout d'une heure alors qu'elle continue au moins jusqu'à trois heures et même au delà en tampon NEPH pour atteindre un signal bien plus élevé.
1000
800
600
VOltqA 00
- Tampon NEPH
+ Tampon PBS
inverses des dilutions
Figure 13 : Influence du milieu réactionnel sur l'agglutination en tampon PBS ou NEPH des microsphères-métallothionéine-thymuline au 1/50 avec
l'immunsérum 003M, au temps 3h. Néphélémètre BEHRING.
69
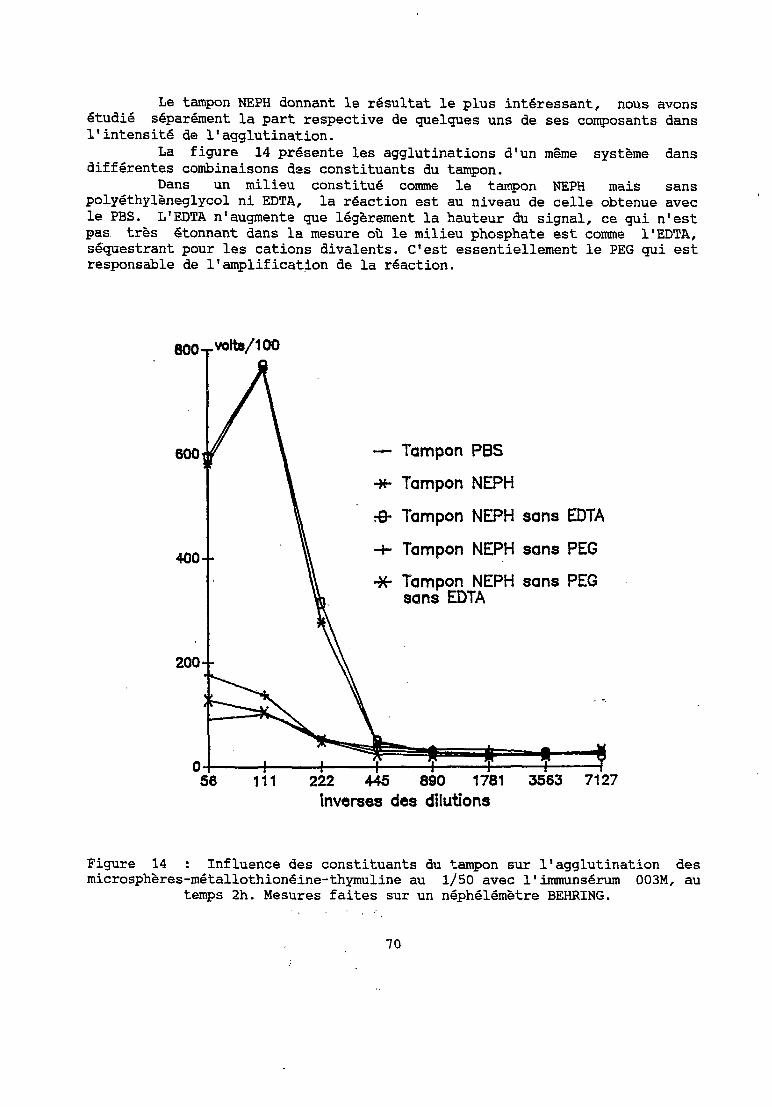
Le tampon NEPH donnant le r6sultat le plus intéressant, nous avons étudié séparément la part respective de quelques uns de ses composants dans l'intensité de l'agglutination.
La figure 14 présente les agglutinations d'un même système dans différentes combinaisons des constituants du tampon.
Dans un milieu constitué comme le tampon NEPH mais sans polyéthylèneglycol ni EDTA, la réaction est au niveau de celle obtenue avec le PBS. L'EDTA n'augmente gue légèrement la hauteur du signal, ce gui n'est pas très étonnant dans la mesure où le milieu phosphate est comme lIEDTA, s&questrant pour les cations divalents. C'est essentiellement le PEG gui est responsable de l'amplification de la réaction.
56 111 222 445 890 1781 3563 7127
- Tampon PBS
+ Tampon NEPH
-8- Tampon NEPH sans EDTA
+ Tampon NEPH sans PEG
+? Tampon NEPH sans PEG sans EDTA
inverses des dilutions
Figure 14 : Influence des constituants du tampon sur l'agglutination des microsphères-métallothionéine-thymuline au 1/50 avec l'immunsérum 003M, au
temps 2h. Mesures faites sur un néphélémètre BEHRING. ., .,:
70
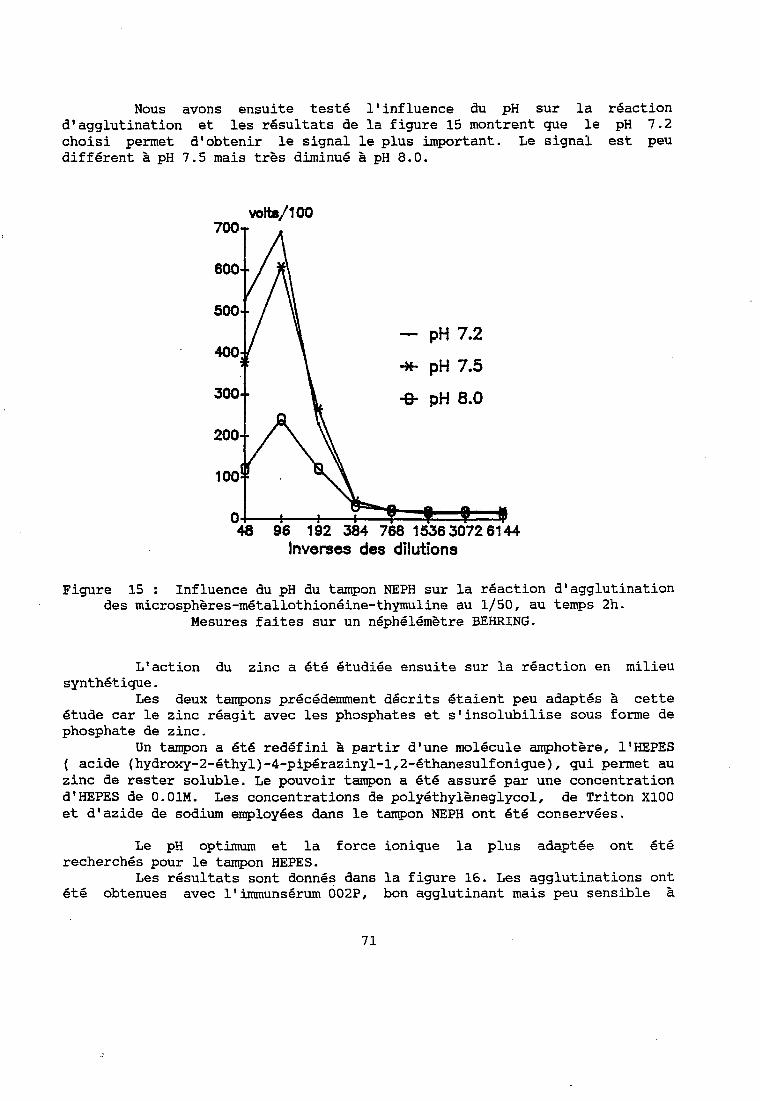
Nous avons ensuite testé l'influence du pH sur la réaction d'agglutination et les résultats de la figure 15 montrent que le pH 7.2 choisi permet d'obtenir le signal le plus important. Le signal est peu différent a pH 7.5 mais très diminué à pH 8.0.
Yotb~100
700T A
- pH 7.2
sç pH 7.5
8 pH 8.0
46 96 192 384 768 153630726144 Inverses des dilutions
Figure 15 : Influence du pH du tampon NEPH sur la réaction d'agglutination des microsphères-métallothionéine-thymuline au 1/50, au temps 2h.
Mesures faites sur un nGphélémètre BEHRING.
L'action du zinc a Bté étudiée ensuite sur la réaction en milieu synthétique.
Les deux tampons prkédemment décrits étaient peu adaptés à cette étude car le zinc réagit avec les phosphates et s'insolubilise sous forme de phosphate de zinc.
Un tampon a ét& redéfini à partir d'une molécule amphotère, 1'HEPES ( acide (hydroxy-2-éthyl)-4-pip~razinyl-l,2-éthanesulfoni~e), gui permet au zinc de rester soluble. Le pouvoir tampon a été assuré par une concentration d'HEPES de O.OlM. Les concentrations de polyéthylèneglycol, de Triton X100 et d'azide de sodium employ6es dans le tampon NEPH ont été conservées.
Le pH optimum et la force ionique la plus adaptée ont été recherchés pour le tampon HEPES.
Les résultats sont donnés dans la figure 16. Les agglutinations ont été obtenues avec l'immunsérum 002P, bon agglutinant mais peu sensible à
71

l'inhibition.
- 0,o M
+ 0,l M
- 0,2 M
* 0,3 M
4 0,4 M
Jt 0,5 M
courbe (b)
- pH 7.0
SC- pH 7.5
0 pH 8.0
Figure 16 : Influence de la concentration de NaCl (a) et du pH (b) du tampon HEPES sur l'agglutination des microsphères-métallothionéine-thymuline.
Mesures faites sur un néphélémètre BEHRING.
72

La force ionique, testée entre 0 et 0.5M en chlorure de sodium, nous permet de retenir la concentration de 0.2M en raison de la hauteur du signal et aussi de l'effet de zone accentué aux fortes concentrations d'immunsérum. Le même raisonnement a permis de retenir le pH 7.5 parmi les 3 pH testés. C'est le pHi de 1'HEPES auquel son pouvoir tampon est le mieux assuré.
La mise au point de ce tampon avait pour but de pouvoir comparer dans le même milieu la réaction soit en présence de zinc soit en milieu séquestrant.
Le tampon HEPES a été additionné soit de 0.56 g/l d'SDTA soit de 13 mg/1 de zinc (apporté sous forme de chlorure de zinc).
L'agglutination a été testée avec l'immunsérum 002P comme précédemment, mais aussi avec un bon immunsérum (le meilleur à l'inhibition testé jusqu'ici), le 0030 (figure 17).
Avec le 002P les résultats sont quasiment identiques dans les deux tampons, seul l'effet de zone disparait en présence d'EDTA. Par contre avec le 0030 le signal est fortement diminué par l'addition de zinc.
La figure 18 présente les performances obtenues à l'inhibition avec l'immunsérum 0030, en présence d'EDTA ou de zinc.
Les réactifs sont employés plus concentrés dans le deuxième cas pour compenser la baisse de signal.
La gamme d'inhibition se situe entre 10 pg/ml et 10 ng/ml avec 50 % d'inhibition autour de 1 ng/ml de thymuline dans le milieu avec EDTA, et entre 10 pg/ml et 100 ng/ml avec 50 % d'inhibition vers 10 ng/ml dans le milieu contenant du zinc.
En présence d'EDTA les conditions réactionnelles, déjà meilleures pour l'agglutination, sont plus favorables à une bonne sensibilité vue la moindre concentration des réactifs. La comparaison des sensibilités à l'inhibition est donc difficile ici.
_.. . . Pour un bon immunsérum il apparait qu'en milieu HEPES la présence
d'EDTA permet l'obtention de signaux plus élevés.
Dans le cas du tampon NEPH l'influence de la concentration d'EDTA a été étudiée et le même phénomène a été retrouvé.
Pour la dilution d'immunsérum la plus réactive, ici 0030 au 1/192, le signal d'agglutination est multiplié par 2 lorsque la concentration d'EDTA passe de 0.56 à 5 g/l (figure 19).
Le tampon de néphélémétrie utilisé (NEPH) est bien adapté à la réaction. L'intensAté de l'agglutination est due au PEG et au pouvoir séquestrant du milieu.
.a, :.
73 .
. .

OOZP
- HEPES + EDTA
+ HEPES + ZINC
0030
4 HEPES + EDTA
-w HEPES + ZINC
Figure 17 : Agglutination des microsphères-inétallothionéine-thymuline p= les immunsérums OOZP et 0030, en tampon HEPES avec zinc ou EDTA, à 2h.
Mesures faites sur un néphélémètre BEHRING.
74

-n HEPES EDTA
ZINC
picogrammes de Thymuline par millilitre
Figure 18 : Comparaison de l'inhibition avec les microsphères- métallothionéine-thymuline au 1/250 et l'immunsénnn 0030, en tampon HEPES + EDTA (0030 1/400, t=75mn) et HEPES + Zinc (0030 lj200, t=240mn).
Mesures faites sur un néphélémètre BEHRING.
0 A . . t * I I 96 la2 384 tbs 1336 3072 6144
inverses des dilutions
Figure 19 : Effet de la concentration d'EDTA du tampon NEPH sur l'agglutination des microsphères-métallothioneine-thymuline au lj250 avec
l'immunsérum 0030, au temps 6h. Néphélémètre BEHRING.
75

II - Conclusion sur les microsphères
g-l . Influence du porteur protéique
La figure 20 présente la comparaison des meilleures courbes d'inhibition obtenues avec chacun des 3 porteurs protéiques employés dans la fabrication des microsphères :
- microsphères-SAB-thymuline - microsphères-polyaspartique-thymuline - microsphères-métallothionéine-thymuline
testées avec le même prélèvement d'immunsérum 003L.
- 0-SAB-lh 003L l/lcr
uhe au t/t 20
t=QOmn
au
o! < , t I I I I I I 4 I I I 8 I I 1 I I
0 0,050,150,45 1.37 4.1 12,3 37 111 333 1000
nanogrammes de Thymuline/ml
Figure 20 : Comparaison, pour les trois types de microsphères, de l'inhibition avec l'immunsérum 003L en tampon NEPH. Mesures faites sur
un néphélémètre BEHRING.
76

comme nous l'avons vu dans le chapitre des microsphères- métallothionéine-thymuline, un changement d'immunsérum peut amener à des sensibilités bien meilleures.
Les inhibitions sont donc plus intéressantes à comparer entre type de microsphères comme présenté ici avec un même immunsérum, qu'en valeur absolue.
La gamme d'inhibition se trouve entre 1 et 100 ng/ml pour les microsphères-SAB-thymuline et entre 0.1 et 100 ng/ml de thymuline pour les 2 autres types de microsphères.
Les concentrations de thymuline donnant 50 % d'inhibition sont :
- microsphères-SAB-thymuline : 30 ng/ml
- microsphères-polyaspartigue-thymuline : 10 ng/ml
- microsphères métallothionéine-thymuline : 1 ng/ml
Les microsphères-métallothionéine-thymuline sont les plus performantes.
g-2 .Réactivité comparée selon la qualité des immunsérums et des microsphères.
Il est intéressant de comparer les différences des agglutinations et des .inhibitions pour plusieurs prélèvements d'immunsérum avec chacun des deux types de microsphères fabriquées avec l'acide polyaspartigue ou la métallothionéine.
Les résultats sont présentés à la figure 21.
Avec les microsphères-métallothionéine-thymuline une discrimination selon la qualité de l'immunsérum peut être faite dès l'agglutination. L'immunsérum le plus agglutinant, le 003M, donne bien le meilleur résultat à l'inhibition et c'est aussi celui gui montre un "effet de zone" aux fortes concentrations de 003M à l'agglutination.
Par contre avec les microsphères-polyaspartigue-thymuline la différence de qualité des immunsérums n'est révélée gu'à l'inhibition puisqu'à l'agglutination leurs résultats sont tous plutôt faibles et encore équivalents.
C'est toujours le prélèvement 003M gui est le meilleur.
La discrimination plus fine des meilleurs immunsérums est un atout supplémentaire pour les microsphères-métallothionéine-thymuline gui sont aussi les meilleures essayées jusqu'ici.
77

Inwmea dea dhrfJon~
nanogmmmea de lhymuh/ml nanogmmme8 de ThymuIine/ml
microsph&res-m&allothion6ine- thymuline
microsphères-polyaspartique- thymuline R7F7
Figure 21 : Comparaison d'immunsérums et de microsphères à l'agglutination et à l'inhibition en tampon NEPH.
Mesures faites sur un néphélémètre BEHRING.
78

g-3 Agglutination comparée selon la qualité des imnunsérums et des microsphères dans un milieu avec ou sans zinc
Les agglutinations présentées figure 22 sont faites dans des conditions plus ou moins bonnes selon que l'on combine des réactifs qualifiés de "bons" ou "mauvais" sur la base de leurs performances Compar&es pour la sensibilité à l'inhibition essentiellement ou dès l'agglutination.
I 1 a
O-R7F8,050C - *
lb) vdb~loo HEPES O-R7F8.0030
HEPES+ZlNC
HEPES+EDTA
Imrsrws dei8 dilulionr
- HEPES
* HEPES+ZINC
- HEPES
SC HEPES+ZlNC
1 =@ HEPES+EDTA
Invmm dea dilutions
- HEPES
-n HEPES+ZlNC
0 HEPES+EDI’A
Figure 22 : Comparaison de l'agglutination en tampon BEPES avec ou sans zinc et EDTA, des microsphères-métallothionéine- et -polyaspartigue-thyline au
1/200, au temps 2h, avec les'immunsérums 05OC et 0030. Mesures faites sur un néphélémètre BEHRING.
79

Les microsphères-polyaspartigue-thymuline ("mauvaises") ou les microsphères-métallothionéine-thymuline ("bonnes") sont agglutin6es par les immunsérums OSOC ("mauvais" car peu sensible) ou 0030 (le meilleur a l'inhibition) et ce dans trois milieux possibles : HBPES seul contenant 77 ng/ml de zinc (défavorable donc "mauvais", dù à une contamination r&iduelle, mesurée par absorption atomique), additionné de zinc à 780 ng/ml (,également "mauvais") ou additionné d'EDTA et donc séquestrant le zinc ("bon"). .
Le système le moins performant (courbe a) mettant en jeu les microsphères-polyaspartigue-thymuline et l'immunsérum 05OC, discrimine à peine la présence ou non d'EDTA et le titre est décalé d'une dilution seulement quand du zinc est ajouté.
Le système le plus performant (courbe d) mettant en jeu les microsphères-métallothionéine-thymuline et l'immunsérum 003 0, est bloqué par l'ajout de zinc, presque entièrement bloqué si le milieu n'est pas séquestrant et par contre très rdactif en milieu séquestrant.
Pour chaque type de microsphère la sensibilité au milieu séquestrant ou non est augmentée avec le bon immunsérum (0030) par rapport au mauvais (05OC). De plus cette différence de sensibilité est plus accentuée avec les bonnes microsphères (-métallothionéine-thymuline).
Nous sommes amenés à conclure que les réactifs les plus performants à l'agglutination et à l'inhibition par de la thymuline sont les plus sensibles aux conditions réactionnelles, notamment en ce gui concerne la disponibilité ou non du zinc.
Dans le but constant d'augmenter la sensibilité à l'inhibition, nous avons étudié les réactifs et conditions de milieu nécessaires B la réaction. Nous avons pu définir la meilleure combinaison pour l'obtention d'une agglutination importante et de l'inhibition la plus sensible.
Un autre moyen d'améliorer la sensibilité. repose sur le choix des paramètres physiques de détection gue sont le diamètre des microsphères et l'angle de lecture selon le néphélémètre.
h- Diamètre des microsphères et appareillage
La mesure de l'agglutination et la sensibilité à l'inhibition dépendent du diamètre des microsphères et du type de néphélémètre.
* le néphélémètre HYLAND détecte sensiblement les petits agrégats et donc leur formation au début de la réaction immunologigue. Des microsphères de départ de petit diamètre et faiblement concentrées sont donc plus favorables dans ces conditions de mesure.
* le néphélémètre BEHRING et le.prototype de NANCY détectent de gros agrégats et donc l'évolution, mais pas le début, de .leur formation. La . . . . . .
80 '

détection sera d'autant plus précoce qu e les microsphères de départ auront une plus grande taille. Si les microsphères sont petites il faut en employer une plus grande quantité.
Ainsi les protocoles de dosage sont différents selon l'appareil utilisé, mais peuvent conduire à une même limite de détection dans le milieu à doser (10).
L'expérience du Laboratoire d'immunologie du Pr. DUHEILLE de la Faculté de Médecine de Nancy, a montré gue; pour un dosage d'antigène par inhibition, les deux néphélémètres HYLAND et BEHRING donnent des résultats satisfaisants avec des microsphères d'un diamètre moyen de 100 nm (10, 70).
Pour cette raison nous avons dès le début utilisé des microsphères de 105 nm de diamètre.
?our essayer d'abaisser la limite de détection des réactions à l'inhibition, nous avons comparé les résultats obtenus avec des microsphères-métallothionéine-thymuline de différents diamètres (78 nm, 105 nm et 295 nm), en tampon NEPH et sur les trois néphélkriètres.
Lorsque l'on compare les résultats obtenus sur deux appareils différents, les échelles de mesure de l'intensité lumineuse ne sont pas les mêmes et ne peuvent donc pas être comparées en valeur absolue. Il faut .prendre en compte le facteur de multiplication du signal de base pour évaluer si l'amplitude des signaux est suffisante. Les concentrations des réactifs- peuvent différer aussi selon l'appareil pour une détection correcte. Les critères d'évaluation d'une réaction restent les mêmes avec essentiellement la sensibilité à l'inhibition.
Les résultats présentés sont obtenus avec les immunsérums 003Q ou 003 0.
Avec les microsphères-polyaspartigue-thymuline les résultats déjà évoqués au chapitre II-2-el ont montré que l'agglutination, lue au BEHRING, est moins importante avec des microsphères de 105 nm qu'avec des microsphères de 295 nm de diamètre. Le résultat à l'inhibition est un peu amélioré avec les microsphères de 105 nm.
, '.
h-i, . Comparaison entre le néphélémètre BEHRING et le prototype de NANCY :
* pour les microsphères de 105 nm de diamètre testées avec l'immunsérum 003Q : (voir figure 23)
les agglutinations mesurées sur les néphélémètres BEHRING ou NANCY donnent un résultat semblable, à savoir une courbe en cloche avec un sommet pour l'immunsérum dilué au l/lOO et une dernière dilution agglutinante vers le 1/400. La sensibilité à l'inhibition est la même, pour une gamme couvrant
., l'ensemble des concentrations testées, avec 50 % d'inhibition vers 1.3 ngjml de thymuline.
81

Inverses des dilutions Inverses de8 dilutions
150
0-l I I 1 0. 0,OS 0.15 0,4S 1.37 4,l 12.3 37 111
nanogmmmes ds lhyrnoline par millilitre
Figure 23 : Agglutination et
signal néohélémètre
50 T signal
0' 6
i 0,05 0,lS O,hS 1.37 4,l 12,3 37 111
nanogmmmes de Thymulhe par milliiitru
inhibition avec les microsphères- métallothionéine-thymuline de 105 nm et l'immunsérum 0034, mesurées
sur les néphélémètres BEHRING et NANCY.
82
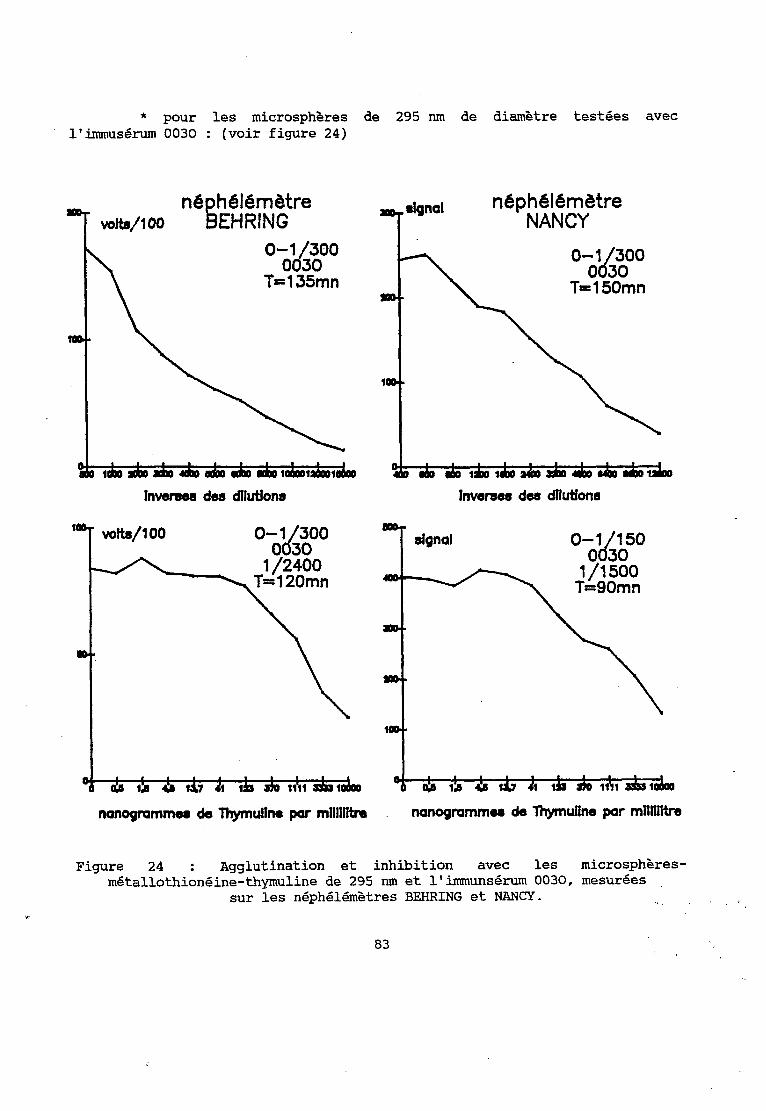
c
* pour les microsphères l'immusérum 0030 : (voir figure 24)
DR né hélémètre volt6/100 I!i EHRING
T=135mn
nanogmmmem de lhymullno par mlllilitm
de 295 nm de diamètre testées avec
rlgnal
7.
nephélemètre NANCY
O-? 3F0 CI T= 150mn
immruea dea diluths
nanogmmmea*l3ymdinepar millUth
Figure 24 : Agglutination et inhibition avec les microsphères- métallothionéine-thyline de 295 nm et l'immunsérum 0030, mesurées
sur les néphélémètres BEHRING et NANCY. .., ,:
83

Le signal de base est assez élevé, et l'agglutination, plus importante, persiste lorsque l'immusérum est encore dilué au 1/10 000 environ pour les 2 appareils. De nouveau les inhibitions sont identiques mesurées sur les néphélémètres BEHRING ou NANCY, avec 50 % d'inhibition autour de 3 ug/ml dans une gamme allant de 41 ng/ml à 10 pg/ml de thymuline.
Nous avons vu jusqu'à maintenant gue le principal facteur dont dgpend la qualité de la réaction reste le changement d'immunsérum. Nous ne pouvons donc pas comparer les résultats obtenus avec les immunsérums 003 0 et. 003 Q, mais pour chacun des deux diamètres il s'avère que les résultats sont quasiment identiques sur ces deux appareils dont l'angle de détection est le même.
h-2 _ Comparaison entre les néphélémètres BEXRING et TAYLAND pou les deux diamètres 78 et 105 nm :
Lors de la mesure sur le néphélémètre HYWND, les réactifs sont employés plus dilués pour détecter le début de réaction de façon sensible. De même les microsphères de 295 nm de diamètre n'ont pas été employées pour rester dans le domaine d'utilisation de cet appareil.
Les résultats sont présentés figures 25 et 26.
Avec les deux néphélémètres BEHRING et HYLAND, l'augmentation du diamètre des microsphères entraine une agglutination plus importante, persistant pour. des dilutions plus grandes d'immunsérum. L'allure des courbes est à peu près la même pour chaque diamètre sur les deux appareils.
Quelle que soit la taille des microsphères, l'inhibition est plus sensible pour la réaction lue sur le néphélémètre BEHRING.
Quel gue soit l'appareil, le seuil de détection à l'inhibition est le même pour les deux diamètres de microsphère. Par contre les concentrations de thymuline inhibant totalement ou à 50 % la réaction sont légèrement plus faibles pour les plus petites microsphères.
En conclusion :
Le néphélémètre BEHRING et le prototype de NANCY permettent l'obtention de résultats identiques, avec des dilutions de travail un Peu différentes pour les réactifs.
Quel que soit le diamètre des microsphères, la réaction est bien meilleure dans les conditions de travail et de lecture du néphélémètre BEHRING que du néphélémètre HYUND.
L'emploi du néphélémètre BEHRING et de microsphères de 105 nm de diamètre est donc adapté à notre mise au point.
La limite de détection n'a pas été améliorée, par rapport aux conditions opératoires initiales, en modifiant les deux paramètres étudiés dans ce chapitre.
84

n Microsphéres 78 nm
a
lnwuea dea dilutions %
. I 1 I
162 708 ’ 30’72 i
immmer dea dilutlona
dte/l 00
I I I I s . . , t
11’11 lob00 m . I . I I I *
. I I 1 . I
13 13,7 123 13 13.7 123 1111 10000
nanogmmmem de lIymulin0 par mIllllRre nanogmmmea de Thymuhe par miliilitm
Figure 25 : Agglutination et inhibition avec les microsphères- métallothionéine-thymuline de 78 et 105 nm et l'immunsérum 0030,
mesurées sur le néphélémètre BEHRING.
85

1& Mo lb- I/l0
MicrosphZres 78 nm MicrosphGres 105 nm
O--l 3ooo d
lnvsnwm de8 dl!utIons
I * . . * . l fi l I . I
13,7 ’ 123 11’11 0
. . . *‘* a e l l ‘ n
13 10600~ l 1;s 13.7 123 il11 lObo
nanogmmmea de llynulh par mll!Mtru nanogmmmea de lhymulhs par milMtre
Figure 26 : Agglutination et inhibition avec les microsphères- métallothionéine-thymuline de 78 et 105 nm et l'immunsérum 0030,
mesurées sur le néphélémètre HYLAND.
86

i - Suivi de la production d'immmsérums
Le type de microsphères, les concentrations de protéine et de thymuline au couplage et le tampon une fois définis, c'est seulement en changeant d'immunsérum que la sensibilité de la réaction peut être améliorée.
Au cours de l'évaluation du protocole d'immunisation, les immunsérums ne peuvent être prélevés en grande quantité sans assurance de leur qualité, évaluée à posteriori.
Ce mode de prélèvement ne peut être effectué fréquemment sans risquer l'anémie ou même la mort du lapin.
Pour obtenir le meilleur immunsérum, un suivi quotidien de la réponse est nécessaire. Dans ces conditions seuls les microprélèvements sont possibles.
Un suivi précis de la production d'anticorps a été mené dans la troisième série d'immunisation, testée dans les conditions de réaction néphélémètrique préalablement mises au point.
Compte tenu des résultats des première et deuxième séries, aucun prélèvement n'a été fait après la première injection ni après le premier rappel. Un prélèvement important a été fait 6 et 7 jours après le deuxième rwwl, puis 6 jours après le troisième rappel. 1 et 3 jours plus tard deux microprélèvements ont été effectués pour commencer à cerner le meilleur jour de prélèvement. C'est après le quatrième rappel qu'un suivi quotidien systématique a été mené avec cinq microprélèvements.
Dans un suivi précis la qualité des prélèvements doit être déterminée rapidement, pour décider d'un éventuel prélèvement important le même jour. Pour cela les microsphères-métallothionéine-thymuline ont été employées plus concentrées qu'habituellement, soit au l/lOO, ce qui donne des signaux importants en 1 heure.
Les prélèvements peuvent être comparés entre eux mais l'emploi de microsphères plus concentrées est fait au détriment de la sensibilité à l'inhibition, comme le montre le résultat avec l'immunsérum 0030 pour lequel des résultats meilleurs avaient été obtenus antérieurement.
Le deuxième rappel a eu lieu le jour 219. Les prélèvements importants A etB ont été faits les jours 225 et 226.
Le troisième rappel, fait au jour 296, a été suivi du prélèvement important C le jour 302. Les microprélèvements D et E ont eu lieu aux jours 303 et 305.
Les résultats d'agglutination des prélèvements A, B, C, D et E sont présentés figure 27. Après les deuxième et troisième rappels, l'agglutination est bien meilleure (la dilution d'immunsérum est plus élevée pour l'obtention du signal maximal, qui est plus important) avec les prélèvements du lapin 300 qu'avec ceux des deux autres lapins. Le maximum d'agglutination est obtenu pour la dilution 1/1536 pour les prélèvements 300A et 300B après le deuxième rappel, et pour 1/768 pour les prélèvements 3OOC et 300D et 1/1536 pour le prélèvement 300E après le troisième rappel.
87

1 Itg 00
FWbvsment A
Pr6bvement B
LAPINS
- 100
* 200
8 300
lb/100 &blWmrnt C
PrWvemont D
Figure 27 : Agglutination des microsphères-métallothionéine-thymuline au l/lOO par les prélèvements des lapins 100, 200 et 300 après les l‘ème et
3ème rappels, en tampon NEPH, mesurée sur un néphélémètre BEHFSNG. 4,
88

après le La figure 28 montre que pour chaque lapin les prélèvements A et B
deuxième rappel donnent de bien-, meilleures agglutinations que les trois prélèvements C, D et E après le troisième rappel. Le signal maximum est obtenu au 1/768 pour les meilleurs prdlèvements des lapins 100 et 200 et au 1/1536 chez le lapin 300..,
1ooA
1008
looc
1OOD
+ 2008
100 - 200E
0 48 88 182 38+ 780 1538 3072 a*
In- ch88 dilution8
‘zii VOlt8/100 A
182 3B4 1838 3072 6144
i-
*
0
3008
300E
Figure 28 : Agglutination des microsph$res-métallothionéine-thymuline au l/lOO par les immunsérums des cinq premiers prélèvements des lapins 100, 200
et 300, en tampon NEPH, au temps 2h, mesurée sur un néphélémètre BEHRING.
89

Les inhibitions sont pr6sent6es figure 29, comparées avec 1"immunsérum 0030 de la première série, le plus performant obtenu jusgu'ici. Les- prélèvements 100B et 200B donnent des résultats proches du 0030. Les performances à l'inhibition sont en faveur du 300B. Les autres prélkements des lapins 100 et 200 n'ont pas été testés à l'inhibition a cause de la faiblesse des signaux à l'agglutination.
a
- pou l/lOOO, œ * ia---~loo
+ PCJJ 1/1000, O~Y)lOO
picogmmme8 de Ihymullne par mIllMtre
Figure 29 : Inhibition avec les microsphères-métallothionéine-thymuline et les immunsérums du prélèvement B des lapins 100, 200 et 300 comparés à
l'immunsérum 0030. Tampon NBPH, néphélémètre BEHRING.
Pour le lapin 300, les résultats au cours des prélèvements sont donnés à la figure 30. Ils sont comparables et ne sont pas influencés par le jour de prélèvement.
90

100
0 I
l I I I I I I I I 8 n 8 I I I I t ’ 18s 43 1&7 41 123 370 lY11 3333 moo
picogmmmea de lhymuh par mTllllitm
Figure 30 : Inhibition avec les microsphères-métallothionéine-thymuline et les prélèvements A, B, C et E du lapin 300, en tampon NEPH.
Mesures faites sur un néphélémètre BEBEING.
C'est apparemment le critère d'agglutination maximale qui peut être retenu pour sélectionner le meilleur jour de prélèvement.
Le quatrième rappel a eu lieu le jour 359, cette fois ci avec de l'adjuvant complet. Les cinq microprélèvements F, G, H, 1 et J, ont eu lieu aux jours 362, 364, 366, 371, 376 soit les 3ème, Sème, .7ème, 12ème et 17ème jours après ce rappel.
La figure 31 présente leur résultat d'agglutination.
91

0 . . I . . l I , i
48 88 la2 364 78a 1538 5472 6144
lm doa dllutlons
182 364 7m 9538 3072 8144 . In-. de8 dllutlons
o+ * I . , I . I I I t 40 00 102 304 780 t53a 3072 0144
In- dea dilutions
Figure 31 : Agglutination des microsphères-métallothionéine-thymuline par les prélèvements F, G, H, 1 et J après le 4ème rappel des lapins
100, 200 et 300, en tampon NEPH, mesurée sur un néphélémètre BEHRING.
92

De façon étonnante, le maximum d'agglutination des prélèvements du lapin 200 est faible mais obtenu à une dilution élevée d'immunsérum (1/1536).
Pour le lapin 300 le meilleur résultat a été obtenu 7 jours après le rappel et un prélèvement important a été effectué.
Pour les lapins 100 et 200 jamais l'agglutination n'a été assez bonne pour effectuer un gros prélèvement.
Les séries de courbes des trois lapins ont la même allure. La dilution donnant le maximum d'agglutination varie en fonction du jour de prélèvement : elle augmente, passe par un maximum au septi'ème jour (pour les lapins 100 et 300) et au douzième jour (pour le lapin 200), puis diminue. La période utile de microprélèvements pour suivre la production d'anticorps semble donc comprise entre le 7ème et le 12ème jour.
Au terme de cette série nous possédons la technique pour suivre la production d'immunsérums par les lapins et intervenir pour prélever au meilleur moment.
93


CONCLUSION
95


Parmi les connaissances sur la thymuline les faits clairement établis sont peu nombreux et la mise au point d'un dosage nouveau non biologique est une entreprise difficile où il existe encore beaucoup d'inconnues. En effet l'absence de dosage non biologique ne permet pas de disposer d'une méthode de référence gui aiderait à évaluer les nouveaux systèmes tout au long de leur mise au point.
L'immunonéphélémétrie est basée sur la mesure de la dispersion de la lumière due à l'agglutination d'antigènes par les anticorps correspondants. L'emploi de supports microparticulaires sur lesquels est présenté l'antigène, ainsi artificiellement grossi, permet une amplification passive du signal néphélémétrigue. La fixation de plusieurs molécules sur une microsphère rend possible le dosage des haptènes comme la thymuline.
.Les microsphères recouvertes de thymuline sont agglutinées par les anticorps anti-thymuline, ce gui donne le signal maximal de référence. Pour le dosage en une étape par inhibition, l'ajout de l'antigène libre à .doser consomme les anticorps agglutinants et entraine une baisse du signal de référence en relation avec la quantité de molécules à doser.
Le but des différentes mises au point des paramètres de la réaction est d'obtenir la meilleure sensibilité possible pour la réaction d'inhibition.
Pour être utilisables dans notre dosage, les microsphires doivent être recouvertes d'une protéine sur laquelle est ensuite fixée la thymuline. Les différents essais ont permis de retenir la métallothionéine comme intermédiaire..
Pour le couplage, 2.10-8 moles de métallothionéine et 10-7 moles de thymuline sont apportées pour 10 mg de microsphères.
Un diamètre des microparticules de 100 nm est satisfaisant et préférable à des diamètres plus importants.
Etant donné que la technique de dosage fait appel à une compétition entre antigène libre à doser et antigène couplé à la microsphère, le seuil de détection est gouverné par le coefficient d'association des anticorps vis à vis de l'antigène. Les coefficients d'affinité les plus forts sont en général obtenus avec des anticorps polyclonaux plutôt qu'avec des anticorps monoclonaux.
L'immunisation avec des injections espacées est la plus favorable à l'obtention d'anticorps anti-thymuline chez les lapins et un rappel au moins est nécessaire avant le début de la réponse.
La production d'immunsérums nécessite un assez grand nombre de lapins car la réponse à l'immunisation est rarement bonne, souvent faible voire absente. Cet aspect imprévisible est certainement dû à la faible immunogénicité de l'haptène thymuline.
97

Les microprélèvements ont permis de constater gue la qualité des immunsérums semble maximale une semaine après un rappel au cours de l'immunisation lente. Ce résultat permet de rationaliser le suivi de la production d'anticorps.
La composition du milieu de réaction module l'énergie de liaison des complexes antigènes-anticorps et influe sur la cinétique des réactions, que l'apport de certains agents, comme le polyéthylèneglycol (PEG), permet d'accéltier.
L'étude des conformations de la thymuline par RMN-2D a montré gue la molécule se présente sous de multiples conformations globalement étirées en équilibre rapide en l!absence de zinc, et que le peptide lié au métal n'adopte pas de conformation figée, mais s'échange aussi rapidement avec l'es chaines libres.
La forme thymuline-zinc, connue comme biologiquement active dans le test des rosettes, est donc minoritaire dans la totalité du peptide libre en milieu aqueux.
De façon à doser la totalité de la thymuline libre et pour éviter les incertitudes liées à la complexation du peptide au zinc, le dosage est fait dans un milieu gui séquestre les cations, propriété assurée par l'ethylènediaminetétraacétique (EDTA) et par les phosphates du tampon NEPH.
Le néphélémètre BEHRING permet une détection, à un angle de 10°, plus favorable à une bonne sensibilité de la réaction d'inhibition gue le néphélémètre HYLAND gui mesure la lumière dispersée à 30°.
En conclusion le dosage de la thymuline en milieu synthétique par inhibition de l'agglutination, permet d'atteindre des concentrations comprises entre quelques picogrsmmes et le nanogramme par millilitre, en deux heures, dans un tampon phosphate à pH 7.2 contenant du PEG et de ~'EDTA, avec des microsphères-métallothion~ine(2.lO-8)-th~line(lO-7), en likant la réaction sur un néphélémètre BEHRING.
La sensibilité du dosage est de l'ordre de 15 pg/ml.
Ce dosage est plus simple et plus performant que ceux relatés dans la littérature. r-
A l'agglutination la conformation reconnue par l'anticorps est une de. celle de la thymuline fixée sur la microsphère par l'intermédiaire de la métallothionéine. Pour la molécule libre une conformation se trouve stabilisée par la liaison au site anticorps. Cette interaction sélectionne parmi les multiples conformations en constant échange de l'hormone en solution, celle correspondant à la structure du site de liaison.
Le dosage prend donc en compte le pool de peptide accessible aux sites de liaison de l'anticorps, tout en déplaçant l'équilibre vers l'apparition de ce conformère, sans permettre d'accéder à la connaissance de la conformation préexistante du peptide puisqu'elle n'est pas unique.
. 98

C'est certainement pour ces raisons que la présence de zinc dans le milieu a un effet variable d'un immunsérum à l'autre sur la capacité à agglutiner les microsphères sensibilisées. Une catégorie d'anticorps, gui se sont révélés les moins favorables à une bonne sensibilité de l'inhibition, agglutinent de façon équivalente avec ou sans zinc dans le milieu. Un autre groupe d'immunsérums, ceux donnant les meilleurs résultats à l'inhibition, agglutinent bien mieux en l'absence de zinc ou en présence de chélateurs que lorsque du zinc est présent dans le milieu.
Le fait de changer de prélèvement d'immunsérum reste le principal facteur qui influe sur le résultat de l'inhibition.
Ce phénomène est d'autant plus sensible que les autres paramètres (protéine intermédiaire et milieu réactionnel) sont les plus favorables à une inhibition sensible.
Pour un même immunsérum, la métallothionéine est le porteur gui permet l'obtention des réactions les plus performantes à l'agglutination et les plus sensibles à l'inhibition.
De plus les microsphères-métallothionéine-thymuline discriminent bien mieux dès l'agglutination la qualité des immunsérums, alors que les microsphères-polyaspartique-thymuline donnent des agglutinations faibles et identiques avec ces mêmes prélèvements.
D'autre part les meilleures microsphères agglutinées par le meilleur immunsérum forment le système le plus sensible à la présence de zinc ou d'EDTA dans le milieu.
De légers gains en sensibilité peuvent encore être espérés en milieu synthétique par l'emploi de microsphères de diamètre légèrement plus faible et un appareil plus performant (comme le prototype de Nancy) gui . permet l'emploi de réactifs plus dilués.
La sensibilité déjà atteinte parait suffisante pour aborder le dosage de la thymuline naturelle en milieu sérique. Les rares données de la littérature mentionnent des concentrations de l'ordre de 25 à 50' pg/ml de thymuline dans le sérum humain; 11 est donc envisageable de la doser, SUP des échantillons biologiques peu dilués, avec la sensibilité de la réaction en immunonéphélémétrie à supports microparticulaires obtenue pour la thymuline synthétique dans un milieu de composition définie.
99


ANNEXES
101


ANNEXE 1 : Fabrication de 1' ïmmunogène ovalbumine-thymuline
Pour 20 doses d'immunogène :
40 mg d'ovalbumine (SIGMA) et 10 mg de thymuline synthétique (FTS BACHEM ou SIGMA) sont dissouts dans 4 ml de tampon phosphate 0.1 M, NaCl 0.15 M, pH 7.4 et mélangés sous agitation magnétique.
2 ml de glutaraldéhyde 0.5 % (p/v) préparé en tampon phosphate 0.1 M, pH 7.4, sont ajoutés et le mélange est agité pendant 3 heures à température ambiante.
La réaction est arrétée par l'ajout de 0.4 ml d'une solution de métabisulfite de sodium 5 %
L'ensemble est di$gedans l'eau). a contre du tampon phosphate 0.1 M, NaCl 0-15 M, pH 7.4 à +4='C. Son volume est ensuite ajusta à 10 ml avec le même tampon.
L'immunogène ainsi obtenu est aliquoté par fractions de 0.5 ml correspondant à 0.5 mg de thymuline, et conservé à -2OOC.
ANNEXE II : Irmrmnofluorescence sur coupes de thymus
Le thymus de porc, congelé dans l'azote .liguide dès son- obtention, est conservé à -8OT.
Des coupes de 4 w d'épaisseur sont préparées au microtome à congélation et récupérées sur des lames porte-objets. Les coupes sont entourées de graisse siliconnée. Ces préparations sont réhydratées en tampon phosphate 0.01 M, NaCl 0.15 M, pH 7.2 (PBS) et placées en chambre humide.
50 à 100 ~1 de la dilution d'immunsérum sont déposés sur la coupe. Après 1 heure d'incubation à température ambiante en chambre
humide, les coupes sont lavées 3 fois 5 min en PBS. 50 ~1 d'anticorps anti-immunoglobulines G de lapin marqués à la
fluorescéine sont déposés sur la coupe à une dilution, en PBS, préalablement établie.
L'incubation se fait durant 30 min à +4OC en chambre humide et à - l'obscurité.
Les coupes sont lavées 3 fois 5 min en PBS, recouvertes d'une goutte de glycérine tamponnée et d'une lamelle, puis sont observées au microscope à fluorescence.
ANNEXE III : Couplage sur les microsphères
Nous présentons le protocole général puis les..adaptations propres à
103

chacun des 3 porteurs protéiques utilisés.
1 ) Protocole général :
* couplage du porteur :
1 volume de microsphères natives est additionné de 0.1 volume de tampon de couplage préalablement dégazé et saturé en argon.
Les microsphères sont dialysées contre 3 fois 1 litre de tampon de couplage à +4V.
Les microsphères, le porteur protéique et du tampon de couplage sont mélangés de façon à avoir en final :
- les microsphères à 10 mg/ml - la protéine à la concentration choisie
Ce mélange est incubg à +4OC pendant 1 nuit et sans agitation.
* saturation des microsphères-protéine : Une solution composée de 1 volume de tampon de couplage, 0.2
volumes d'éthanolamine et 0.184 volume d'acide acétique est ajouté de façon à être au 1/20 dans le mélange final, pour saturer les fonctions aldéhydes des microsphères n'ayant pas réagi. L'incubation dure 1 heure à température ambiante sans agitation.
*. séparation de la protéine libre :
Les microsphères-protéine sont séparées de la protéine libre par ultracentrifugation en gradient discontinu de saccharose à 80 % et 20 % (p/v en tampon de couplage). Les microspères sont récupérées à l'interface, reajustées au volume initial avec le tampon de couplage, et dialysées contre 5 fois 1 litre de NaCl 0.9 %, pH 6.5 à +4Y.
* couplage de la thymuline sur la protéine :
De l'hyhdrochlorure de N(3 diméthylaminopropyl)- N'éthyl- carbodiimide ou ZDC (MERJX ref 800907) .est pesé au fond d'un tube en verre (5 mg pour 10 mg de microsphères). Dans l'ordre sont ajoutées les mïcrosphères-protéine puis la thymuline apportée à 2.10-6 moles /ml en NaCl 0.9 %, pH 6.5, 'sous un volume adapté pour obtenir la concentration voulue.
L'ensemble est mélangé au vortex et incubé durant 4 heures à température ambiante sans agitation.
Les fonctions activées de la protéine n'ayant pas réagi sont saturées par de l'urée 4 M en tampon phosphate 0.05 M, NaCl 0.3 M (250 ~1 pour 10 mg de microsphères).
Les microsphères sont immédiatement mises en dialyse contre 3 fois 1 litre de tampon phosphate 0.01 M, NaCl 0.15 M, pH 7.2 à +4OC.
2 ) Cas particuliers :
104

Pour la sérum-albumine bovine, 1Jacide polyaspartigue et la métallothionéine gui ont été employés comme intermédiaire entre la microsphère et la thymuline, sont précisées ci-après les étapes gui diffèrent du protocole général.
a ) La sérum-albumine bovine (SAB) :
- Le tampon de couplage est du borate 0.1 M, NaCl 0.3 M, pH 8.2.
- Le porteur est la SAB, apportée à 14 mg/ml soit 2.10-7 moles /ml sous un volume gui dépend de la concentration voulue.
b ) L'acide polyaspartique :
- Le tampon de couplage est du phosphate 0.01 M, NaCl 0.15 M, pH 7.2.
- Le porteur est l'acide polyaspartique (SIGMA ref 5387) apporté à 3 mg/ml soit 2.10-7 moles /ml sous un volume gui dépend de la concentration voulue.
- La saturation des microsphères-polyaspartique est faite avec de la glycine préparée à 27 mg/ml en tampon phosphate 0.1 M, NaCl 0.3 M, pH 7.2, apportée à raison de 500 ~1 pour 50 mg de microsphères-polyaspartique, et incubée 2 heures à température ambiante. Cela remplace la saturation par l'éthanolamine.
- Après le gradient de saccharose les microsphgres-polyaspartigue sont dialysées contre 3 fois 1 litre de tampon phosphate 0.1 M, pH 8. La liaison imine entre la microsphère et le polyaspartique est ensuite réduite au borohydrure de sodium (NaBH4). Le NaBH4 est pesé au fond d'un bécher (25 mg pour 50 mg de microsphères). Les microsphères-polyaspartique sont ajoutées et le mélange est agité pendant 2 heures à température ambiante, en maintenant constamment le pH entre 8 et 9 maximun avec de l'acide acétique 1 M. Le pH est ensuite amené à 6 et l'agitation est maintenue jusqu'à la fin du dégagement gazeux.
Les microsphères-polyaspartique sont dialysées contre 3 fois 1 litre de NaCl 0.9 0, 0, pH 6.5 et le couplage de la thymuline se poursuit selon le protocole général.
c ) La métallothionéine :
- Le tampon de couplage est le phosphate O..Ol M, NaCl 0.15 M, pH 7.2.
- Le porteur est la métallothionéine (SIGMA ref 7641) apportée à 1.5 mg/ml soit 2.10-7 moles/ml sous un volume gui dépend de la concentration voulue.
105

- Les microsphères-métallothionéine ne sont pas saturées.
- La métallothionéina non couplée est éliminée par dialyse contre 5 fois 1 litre de NaCl 0.9 "a,,. pH 6.5 supprimant le gradient de saccharose. Le couplage de la thymuline est poursuivi selon le protocole général.
RECAPITULATION:
sérum-albumine bovine acide polyaspartique
dialyse pH 8.2 dialyse pH 7.2
+ SAB + polyaspartique
saturation saturation éthanolamine glycine
gradient saccharose gradient saccharose
dialyse pH 8
réduction NaBH4
dialyse pH 6.5 dialyse pH 6.5
activation au activation au carbodiimide carbodiimide et couplage de et couplage de la thymuline la thymuline
métallothionéine
dialyse pH 7.2
+ métallothionéine
dialyse pH 6.5
activation au carbodiimide et couplage de la thymuline
106

REFERENCES BIBLIOGRAPHIQUES
107


1 -
2 -
3 -
4 -
5 -
6 -
7 -
8 -
9 -
10 -
11 -
12 -
13 -
AREF G.H., ABDEL-AZIZ A., ELARABY I-F., ABDEL-MONEIM M.A., HEBEISHY N.A. and RAHMY AI (1982). A post mortem study of the thymolymphatic system in protein energy malnutrition. J. Trop. Med. Hyg. 85 : 109- 114.
AUGER C., MONIER J.C., DARDENNE M., PLEAU J.M. and BACH J.F. (1982). Identification of FTS (facteur thymique sérigue) on thymus ultrathin sections using monoclonal antibodies. Immunol. Letters 5 : 213-216.
BACH J.F., PAPIERNIK M., LEVASSEUR P., DARDENNE M., BAROIS A. and LEBRIGAND H. (1972). Evidence for a serum-factor secreted by the human thymus. The Lancet 2 : 1056-1058.
BACH J.F., DARDENNE M., PLEAU J.M. and BACH M.A. (1975). Isolation, biochemical characteristics and biological activity of a circulating thymie hormone in the mouse and in the human. Ann. N.Y. Acad. Sci. 249 : 186-210.
BACH J.F., DARDENNE M., PLEAU J.M. and ROSA J. (1977). Biochemical characterisation of a serum thymie factor. Nature 266,5597 : 55-57.
BACH J.F., BACH M.A., BLANOT D., BRICAS E., CHARREIRE J., DARDENNE M FOURNIER C. and PLEAU J.M. J3kketin de l'Institut Pasteur 76
(1978). Thymie serum factor (FTS). : 325-398.
BACH J.F. (1986). Immunologie. FLAMMARION Ed. Paris.
BENE M.C.,, FAURE G., MONTAGNE P:, JAMBON B., REMBAUM A. and DUHEILLE J. (1982). S.E.M. visualization of human lymphocytes membrane receptors for facteur thymique serique (FTS). S.E.M. Congress may 1982. Anheim, USA.
BERRIH S., SAVINO W., AZOULAY M., DARDENNE M. and BACH J.F. (1984). Production of .anti-thymulin (FTS) monoclonal antibodies by immunization against human thymie epithelial cells. J. Histochem. Cytochem. 32,4 : 432-438:
BESSOU T. (1985). Immunonéphélémétrie à supports microparticulaires. Application aux dosages biologiques. Thèse de Docteur Ingénieur, 106 p., Nancy.
BORDIGONI P., FAURE G., BENE M.C., DARDENNE M., BACH J.F., DUHEILLE J. and OLIVE D. (1982). Improvement of cellular immunity and IgA production in immunodeficient children after treatment with synthetic serum thymie factor (FTS). The Lancet 2 : 293-297.
BRAYER M.P. (1981). Contribution à l'étude des procédés de dosage du facteur thymique sérigue (FTS). Thèse de Pharmacie, 107 p, Nancy.
BRICAS E. (1980). Etude des relations structure-activité de
109

14 -
151 -
16 -
17, -
38’ -
19 -
20 -
21 -
22' -
23: -
2.4 -
25 -
l'hormone thymique circulante (Facteur Thymique Sérique : FTS). Compte rendu de fin d'étude d'une recherche financée par la Délégation Générale S la Recherche Scientifique et Technique.
BURTON P., IDEN S., MITCHELL K. and WHITE A. (1978). Thymie hormone- like restoration by human prealbumin of azathioprine sensitivity of spleen cells from thymectomized mice. h-oc. Natl. Acad. Sci. USA 75,2 : 823-827.
CHANDRA R.K. (1979). Serum thymie hormone activity in protein- energy malnutrition. Clin. exp. Immunol. 38 : 228-230.
CHANDRA R.K. (1980). Immunology of nutritional disorders.,ARNOLD Ed. London.
CHANDRA R-K., HERES1 G. and AU B. (1980). Serum thymie factor activity in deficiencies of calories, zinc, vitamin A and pyridoxine. Clin. exp. Immunol. 42 : 332-335.
CHANDRA R.K., GUPTA S. and SINGH H. (1982). Inducer and suppressor T ce11 subsets in protein-energy malnutrition : analysis by monoclonal antibodies. Nut. Res. 2 : 21-26.
CHANDRA R.K. (1983). Numerical and functional deficiency in T helper cells in protein energy malnutrition. Clin. exp. Immunol. 51 : 126- 132.
CHENDID TLEMCANI R. (1985). Contribution au dosage de S'hormone thymique. DEA de Biochimie Pharmacologie, Nancy.
COHEN S., BERRIH S., DARDENNE M. and BACH J.F. (1983). Régulation in vitro de la sécrétion de thymuline (FTS) par les cellules épithéliales thymiques humaines. C. R. Acad. Sci. (Paris) 297, série III : 63-65.
DARDENNE M. and BACH J.F. (1977). Demonstration and characterization of a serum factor produced by activated T cells. Immunology 33 : 643-351.
DARDENNE M., PLEAU J.M. and BACH J.F.' (1980). Evidence of the presence in normal serum of a carrier of the serum thymie factor (FTS). EU~. J. Immunol. 10 : 83-86.
DARDENNE M., PLEAU J.M., BLOUQUIT J.Y. and BACH J.F. (1980). Characterization of facteur thymique sérique (FTS) in the thymus. II. Direct demonstration of the presence of FTS in thymosin fraction V. Clin. exp. Immunol. 42 :.477-482.
DARDENNE M. and BACH J.F. (1981). Thymie hormones. In : The thymus gland, MD KENDALL Ed. Academic Press, London : 113-131.
110

26 -
27 -
28 -
29 -
30 -
31 -
32 -
33 -
34 -
35 -
36 -
37 -
DARDENNE M., PLEAU J.M., LEFRANCIER P. and BACH J.F. (1981). Rôle du zinc et d'autres métaux dans l'activité biologique du FTS (thymuline). C. R. Acad. Sai. (Paris) 292, Série III : 793-795.
DARDENNE M., PLEAU J.M., NABARRA B., LEFRANCTER P., DERRIEN M., CHOAY J. and BACH J.F. (1982). Contribution of zinc and other metals to the biological activity of the serum thymie factor. Proc. Natl. Acad. Sci. USA 79 : 5370-5373.
DARDENNE M., SAVINO W., GASTINEL L.N., NABARRA B. and BACH J.F. (1983). Thymie dysfunction in the mutant diabetic (db/db) mouse. J. nzlmunol. 130,3 : 1195-1199.
DARDENNE M., SAVINO W., WADE S., BENSMAN A., NABARRA B. and BACH J.F. (1984). In vivo and in vitro studies of thymulin in marginally zinc-deficient mice. Eur. J. Immunol. 14 : 454-458.
DARDENNE M., SAVINO W., BERRIH S. and BACH J.F. (1985). A zinc dependent epitope on the molecule of thymulin, a thymie hormone. Proc. Natl. Acad. Sci. USA 82 : 7035-7038.
DARDENNE M., SAVINO W., DUVAL D., KAISERLIAN D., HASSID J. and BACH J.F. (1986). Thymie hormone-containing cells. VII.Adrenals and gonads control the in vivo secretion of thymulin and its plasmatic inhibitor. J. Inmmnol. 136,4 : 1303-1308.
EDELMAN R. (1977). Cell-mediated immune response in protein-calorie malnutrition - A.Review. In : Malnutrition and the Immune Response, RM SUSKIND Ed, Raven Press, New,York : 47-75.
FAURE G., BENE M.C., TAMISIER J.N., GAUCHER A. and DUHEILLE J. ., (1984). Thymulin (FTS-Zn) induced in vitro modulation of T ce11 subset markers on lymphocytes from rheumatoid arthritis and.systemic lupus erythematosus patients. Int. J. Izmunophakacol. 6 : 382.
FOK K.F., OHGA K., INCEFY G.S. and ERICKSON B.W. (1982). Antigenic specificity of two antibodies directed against the thymie hormone serum thymie factor (FTS). Molec. Immunol. 19,12 : 1667-1673.
GASTINEL L.N., PLEAU J.M., DARDENNE M., SASAKI A., BRICAS E., MORGAT J.L. and BACH J.F. (1982). High affinity binding sites on plasma membrane obtained from the lymphoblastoid cultured 1301 ce11 line for highly radioactive serum thymie factor. Biochim. Biophys. Acta 684 : 117-126.
GASTINEL L.N., DARDENNE M., PLEAU J.M. and BACH J.F. (1984). Studies on the zinc binding site to the serum thymie factor. Biochim. Biophys. Acta 797 : 147-155.
GOLDSTEIN A.L., SLATER F.D. and WHITE A. (1966). Preparation, assay
111

30
39,
40'
4%
42
43
45
46
47
and partial purification of a thymie lymphocytopoietic factor (thymosin). Proc. Natl. Acad. Sci. USA 56 : lOIO-1017.
GOLDSTEIN G. (1974). Isolation of a bovin thymin. A polypeptide hormone of the thymus. Nature 247 : 11-14.
GOLDSTEIN G., SCEID M.P., BOYSE E.A., SCHLESINGER D.H. and VANWAURE J. (1979). A synthetic pentapeptide with biological activity characteristic of the thymie hormone thymopoietin. Science 204 : 1309.
GROSS R.L. and NEWBERNE P.M. (1980). Role of nutrition in inununological function. Physiol. Rev. 60,l : 188-302.
HERES1 G. and CHANDRA R.K. (1980). Effects of severe calorie restriction on thymie factor activity and lymphocyte stimulation response in rats. J. Nutr. 110 : 1888-1893.
IKEYAMA S., KATO K., TAKAOKI M., TAKEUCHI M. and KAKINUMA A. (1984). Radioimmunoassay of serum thymie factor (FTS). J. Irmnunoassay 5 : 87-97.
JAMBON B., MONTAGNE P., BENE M.C., BRAYER M.P., FAURE G. and DUHEILLE J. (1981). Immunohistologic localization of "facteur thymique sérique" (FTS) in human thymie epithelium. J. Imunol. 127,s : 2055-2059.
JAMBON B., ZIEGLER O., MAIRE B., BENEM.C., FAURE G., PATRIS A. and DUHEILLE J. (1984). Serum thymie factor (FTS) contents of the thymus in undernourished senegalese children. 'PA
' immunohistological study". post-mortem
In Marker Proteins in Inflammation, Vol 2, Walter de Gruyter, Berlin-New York : 453-455.
JAMBON B., ZIEGLER O., AKIMWOLERE A., MAIRE B. and DUHEILLE J. (1985). Relations entre l'atteinte fonctionnelle du thymus et le peuplement lymphocytaire des ganglions lymphatiques mésentérigues chez l'enfant décédé en état de malnutrition. Cahiers de Nutrition et de Diététique 20. : 185.
JAMBON B., ZIEGLER 0. and DUHEILLE J. (1987). Fonction hormonale lymphodifférenciatrice du thymus et malnutrition protéino- énergétique. In : Les malnutritions dans les pays du Tiers-Monde, Colloque INSERM.136 : 333-339. D. LEMONNIER, Y.INGENBLEEK Eds. Paris.
KEUSH G.T., WILSON C.S. and WAKSAL S.D. (1983). Nutrition, host defenses and the lymphoid system. In : Advance in host defense mechanisms vol 2 : 275-359. JI GALLIN and AS FAUCI Ed. Raven Press, New York.
112

48, - KOOK A-L., YAKIR Y. and TRAININ N. (1975). Isolation and partial chemical characterization of THF, a thymus hormone involved in immune maturation of lymphoid cells. Cell. ~rnmunol. 19 : i51-157.
49 - LACOVERA J. and UTERMOHLEN V. (1983). Isolation and assay of a human plasma factor affecting human thymus derived lymphocytes. Clin. Imunol. Inmmnopathol. 27,3 : 428-432.
50 - LAROCHE P. (1987). Immunonéphélémétrie sur supports microparticulaires. Mise au point de dosages biologiques. Thèse de Doctorat, 225 p, Nancy.
51 - LAUSSAC J.P., CUNG M.T., PASDELOUP M., HARAN R., MARRAUD M., LEFRANCIER P., DARDENNE M. and BACH J.F. (1986). NMR study of thymulin, a lymphocyte differentiating thymie nonapeptide. Conformational states of free peptide in solution. 3. Biol. Chem. 261,17 : 7784-7790.
52 -' LE J., IKEHARA S., PAHWA R., BURTON P.,, WHITE A. and GOOD R.A. (1981). In vitro effects of prealbumin on T-cell-dependent immune responses of the mouse. Cell. Innnunol. 65 : l-12.
53 - LEUNG K.H., EHRKE M.J., BERCSENYI K. and MIHICH E. (1982). Human prealbumin fraction : effects on cell-mediated immunity and tumor rejection. Immunopharkcology 4 : 55-67.
54 -' LOW L.T.K., THRUMAN G.B., McADOO.M., McCLURE J., ROSSI0 J.L., NAYLOR P.H. and GOLDSTEIN A.L. (1979). The chemistry and biology of thymosin. I.Isolation, characterization and biological activities of thymosin a1 and polypeptide Bl from calf thymus. J. Biol. Chem. 254 : 981-986.
55 - LOW T.L.K. and GOLDSTEIN A.L. (1986). Thymie hormones : an overview. Methods in Enzymology 116 : 213-279.
56 - McMURRAY D.N. (1984). Cell-mediated immunity in nutritional deficiency. Progress in Food and Nutrition Science 8 : 193-228.
57 - METREAU E., PLEAU J.M., DARDENNE M., BACH J.F. and PRADELLES P. (1987). An enzyme immunoassay for synthetic thymulin. J. Immunol. Methods 102 : 233-242.
58 - MITTAL A. and WOODWARD' B. (1985). Thymie epithelial cells of severly undernourished mice : accumulation of cholesteryl esters and absence of cytoplasmic vacuoles. Proc. Soc. Exp. Biol. Med. 178 : 385-391.
59 - MITTAL A. and WOODWARD B. (1986). Ultrastructural and morphometric analysis of thymie epithelial secretory vacuoles in severly protein- energy malnourished weanling mice. Nutr. Res. 6 : 663-671.
113

60 -
a! -
62 -
63 -.
64 -
65, -
66 -
67 -
68 -
69 -
70 -
MITTAL A., WOODWARD B. and CHANDRA R.K. (1988). Involution of thymie epithelium and low serum thymulin bioactivity in weanling mice subjected to severe food intake restriction or severe protein deficiency. Exp. Molec. Pathol. 48 : 226-235.
MONIER J.C., DARDENNE M., PLEAU J.M., SCHMITT D., DESCHAUX P. and BACH J.F. (1980). Characterization of facteur thymique sérigue (FTS) in the thymus. I.Fixation of anti-FTS antibodies on thymie reticulo- epithelial cells. Clin. exp. Immunol. 42 : 470-476.
OHGA K., INCEFY G.S., WANG C.Y. and GOOD R.A. (1982). Generation of a monoclonal antibody against facteur thymique sérique (FTS). Clin. exp. Immunol. 47 : 725-734.
OHGA K., INCEFY G.S., FOK K.F., ERICKSON B.W. and GOOD R.A. (1983). Radioimmunoassays for the thymie hormone serum thymie factor (FTS). J. Immunol. Methods 57 : 171-184.
PARENT G., JAMBON B., MAIRE B. and CHEVALIER P. (1987). Echographie du thymus. Interêt diagnostique et pronostique dans la malnutrition. 3èmes Journées Scientifiques Internationales du Groupe d'Etude et de Recherche sur la Malnutrition, juillet 1987, Dakar.
PLEAU J.M., DARDENNE M., BLOUQUIT J.Y. and BACH .J.F. (1977). Structural study of circulating thymie factor : a peptide isolated from pig serum. 1I:Amino acid sequence. J. Biol. Chem. 252 : 8045- 8047.
PLEAU J.M., PASQUES D., BACH J.F., GROS C. and DRAY F.. (1977). Dosage radioimmunologigue du facteur thymique sérigue (FTS). Radioinmnznoassay and Related Procedures in Medicine 2 : 505-510 International Atomic Energy Agency, Vienna.
PLEAU J.M. and DARDENNE M. (1981). Le FTS : Dosage radioimmunologique. Hormone humaine. Analogues structuraux. Compte rendu de fin d'étude d'une recherche financée par la Délégation Générale à la Recherche Scientifique et Technique.
PLEAU J.M., DARDENNE M. and BACH J.F. (1981). Modification de l'antigénicité .du FTS (thymuline) par le zinc. C. R. Acad. Sci. (Paris) 292, Série III : 991-993.
RICARD D. (1985). Exploration échographique du thymus et nutrition chez l'enfant en milieu tropical. Thèse de Doctorat en Médecine, 140 p; Tours.
RIOCHET D. (1984). Immunonéphélémétrie à supports microparticulaires. Synthèse et sensibilisation du réactif. Thèse de Docteur Ingénieur, 134 p, Nancy.
114

71 -
72 -
73 -
74 -
75 -
76 -
77 -
78 -
79 -
80 -
81 -
82 -
SAVINO W., DARDENNE M., PAPIERNIK M. and BACH J.F. (1982). Thymie hormone-containing cells. Characterization and localization of serum thymie factor in Young mouse thymus studied by monoclonal antibodies. J. Exp. Med. 156 : 628-633.
SAVINO W. and DARDENNE M. (1984). Thymie hormone containing cells. IV Immunohistologic evidence for the simultaneous presence of thymulin, thymopoietin and thymosin a1 in normal and pathological human thymus. Europ. J. Iznmunol. 14 : 987-991.
SAVINO W., GASTINEL L.N., WOLFF A., DARDENNE M. and BACH J.F. (1985). Caractérisation immunochimigue des molécules reconnues dans l'épithélium thymique humain par des anticorps monoclonaux anti- thymuline. C. R. Acad. Sci. (Paris) 301, série III,11 : 577-579.
SCHLESINGER D.H. and GOLDSTEIN G. (1975). The amino acid seguence of thymopoietin II. Ce11 5 : 361-365.
SCHMITT D., MONIER J.C., DARD&E M., PLEATJ J.M., DESCHAUX P. and BACH J.F. (1980). Cytoplasmic localization of FTS (facteur thymique sérigue) in thymie epithelial cells. An immunoelectron microscopical study. Thymus 2 : 177.
SUSKIND R.M. (1977). Malnutrition and the Immune Response. Raven Press, New York.
SZTEIN M.B. and GOLDSTEIN A.L. (1986). Thymie hormones. A clinical update. Springer Seminars in Immunopathology 9 : 1-18. .
WADE S., PARENT G., BLEIBERG-DANIEL F., MAIRE B., FALL M., SCHNEIDER D LEMOULLAC B. in' protein-energy
and DARDENNE M. (1988). Thymulin (Zn-FTS) activity malnutrition : new evidence for interaction
between malnutrition and infection on thymie function. Am. J.--Clin Nutr. 47 : 305-311.
WHITE A. and BURTON P. (1979). Isolation from human plasma of a protein fraction with thymie hormone-like activity. Ann. fi.Y. Acad. Sci. 332 : l-8.
YOSHIDA T., TAKAOKI M., KATO K., TAKEUSHI M., NISHIMURA O., FUJINO M. and KAKINUMA A. (1984). The activity of synthetic analogs of serum thymie factor (FTS) to convert mouse pre-T cells into Thy-1 positive cells. Int. J. Immunopharmac. 6,2 : 141-146.
ZIEGLER 0. (1983). Les conséquences de la malnutrition protéino- énergétique sur la structure et le contenu du thymus en facteur thymique sérigue (FTS). Etude histo-immunologigue chez 58 enfants sénégalais. Thèse de doctorat en Médecine, 283 p, Nancy.
ZIEGLER O., JAMBON B., MAIRE B., DARDELIN R., FALL M. and DUHEILLE
115

' J. (1984). The clinico-pathological consequences of severe protein- energy malnutrition in deceased senegalese children. In : Marker Proteins in Inflammation, Vol 2, Walter de Gruyter, Berlin-New York : 457-460.
83' - ZIEGLER O., JAMBON B. and DUHEILLE J. (1987). Les déterminants de l'involution thymique chez l'enfant malnutri. In : Les malnutritions dans les pays du Tiers-Monde. Colloque INSERM 136 : 341-347. D. LEMONNIER, Y.INGENBLEEK Eds. Paris.
116





![Le dosage de l’inuline : mise au point dosage...entre dosage de fructose et de glucose si la réaction a lieu à 75 et non pas à 100 C [26]. Résorcinol Ce dosage se base sur la](https://img.pdfslide.fr/doc/110x75/6082d106c887c069e27bd4cf/le-dosage-de-lainuline-mise-au-point-dosage-entre-dosage-de-fructose-et-de.jpg)
















![Dosage acide-base [Mode de compatibilit ]](https://img.pdfslide.fr/doc/110x75/62b37c5bb878fe2f74541c96/dosage-acide-base-mode-de-compatibilit-.jpg)





